
Submitted:
23 August 2024
Posted:
26 August 2024
You are already at the latest version
Abstract
Medicago sativa L. (Alfalfa) is a globally recognized forage legume that has recently gained attention for its high protein content, making it suitable for both human and animal consumption. However, due to its perennial nature and autotetraploid genetics, conventional plant breeding requires a longer timeframe compared to other crops. Therefore, genetic engineering offers a faster route for trait modification and improvement. Here, we describe a protocol for achieving efficient transient gene expression in alfalfa through genetic transformation with the Agrobacterium tumefaciens pCAMBIA1304 vector. This vector contains the reporter genes β-glucuronidase (GUS) and green fluorescent protein (GFP), along with a selectable hygromycin B phosphotransferase gene, all driven by the CaMV 35s promoter. Various transformation parameters—such as different explant types, leaf ages, leaf sizes, wounding types, bacterial concentrations (OD600nm), tissue preculture periods, incubation periods, co-cultivation periods, and different concentrations of acetosyringone, silver nitrate, and calcium chloride—were optimized using 3-week-old plantlets raised in vitro. Results were attained from data based on the semi-quantitative observation of the percentage and number of GUS spots on different days of agro-infection in alfalfa explants. The highest percentage (76.2%) of GUS-positive alfalfa leaf explants was observed after 3 days of agroinfection at a bacterial concentration of 0.6, 2 days of preculture, 30 min of infection time, 150 µM acetosyringone, 4 mM calcium chloride, and 75 µM silver nitrate. The transient expression of genes of interest was confirmed via histochemical GUS and GFP assays. The results based on transient reporter gene expression suggest that various factors influence T-DNA delivery in the Agrobacterium-mediated transformation of alfalfa.
Keywords:
Medicago sativa L. (Alfalfa)
; Agrobacterium tumefaciens
; β-glucuronidase (GUS)
; Green fluorescent protein (GFP)
; Transient gene
1. Introduction
Medicago sativa L. (Alfalfa) is a popular forage legume crop for the production of high-quality biomass, which is utilized for hay, silage, cover crops, and green manure [1]. The cultivation of alfalfa is valued for its role in soil protection, bioremediation, the nitrogen cycle, and wildlife habitat preservation. Being a perennial crop with nitrogen-fixing capabilities, it is extensively cultivated as a primary source of proteins, enzymes (amylase, coagulase, peroxidase, pepsin, lipase, invertase, and pectinase), antioxidants, minerals, and vitamins A, C, K, and E, as well as valuable phytopharmaceutical components [2]. Moreover, it can be used as a cover crop in grasslands for improved weed control, in the production of recombinant pharmaceutical proteins [3], and in phytoremediation [4]. Alfalfa is an autotetraploid with a chromosome count of 32 (2n=4x=32) and a genome size ranging from 800 to 900 Mbp in length [5]. Researchers have undergone more than 50 years of efforts aimed at enhancing the productivity and quality traits of alfalfa. However, success has been limited [6], due to the intricacy of this species and the challenges involved in its genetic enhancement using traditional breeding methods. The utilization of biotechnological tools and techniques is essential to facilitate the large-scale production of alfalfa for industrial use.
Genetic transformation introduces novel traits or manipulates gene expression, leading to valuable phenotypic variations that are crucial for crop enhancement and to improve the understanding of gene function. This process entails integrating exogenous genes and regulating endogenous gene expression through transformation [7]. Various techniques are available for transferring desired genes into the plant genome, via direct and indirect techniques such as electroporation, micro-projectile bombardment, and Agrobacterium-mediated transformation [8]. Among these, the Agrobacterium-mediated technique has become a common practice in numerous laboratories [9], involving the transfer of DNA from the Agrobacterium plasmid into the cells of numerous dicotyledonous and some monocotyledonous plants [10]. Since the initial report by Deak et al. (1986) [11], multiple efforts have been made to introduce beneficial genes into the alfalfa genome via Agrobacterium-mediated transformation, such as insect resistance against H. postica larvae [12], increased salt tolerance [13,14], improved nutritional quality [15], higher yields via delayed leaf senescence [16], and higher rates of phosphate uptake into cells [17].
Reporter genes utilized as suitable markers allow for the visualization of gene expression and protein localization in vivo for a wide range of prokaryote and eukaryote genomes. For example, genes encoding chloramphenicol acetyltransferase (CAT), green fluorescent protein (GFP), luciferase (LUC), β-glucuronidase (GUS), and red fluorescent protein (DsRed2) are commonly employed as reporters [18,19]. The Escherichia coli gusA (uidA) gene encoding for the enzyme β-glucuronidase (GUS) offers a clear indication that transferred genes are expressed in transgenic plant cells in either a transient or stable manner [20,21]. The GUS gene cleaves a β-glucuronidase substrate variant, which is used in spectrophotometric, fluorimetric, and histochemical assays, resulting in the formation of an indigo–blue precipitate within cells. Green fluorescent protein (GFP), isolated from jellyfish (Aequorea victoria), is a powerful tool in molecular biology and cell biology research. It serves as a widely utilized reporter gene across numerous living organisms [22,23], frequently aiding in the identification of transformed cells, tissues, and organs in both plants and animals, often along with genes conferring antibiotic or herbicide resistance. GFP’s strong fluorescent intensity makes it an excellent visual marker for selecting positive transformants and has the potential to track the expression of transgenes in genetically engineered crops. In addition, it does not require any exogenous substrates. This also allows for the monitoring of transgenic expression from early events of transformation through the regeneration of transgenic plants. Another advantage is the relatively small (26.9 kD) genome size, which assists with both N- and C-terminal protein fusions, lending to the study of localization and intracellular protein trafficking [23].
Genetic engineering technology has been used in alfalfa breeding programs to target the improvement of interesting agronomic traits such as herbicide tolerance and forage quality. Roundup Ready alfalfa [24] and low-lignin alfalfa have been deregulated and commercialized in the US. Various methods have been attempted for the genetic transformation of alfalfa [11,25,26,27]. Since the identification of a highly regenerable genotype, Regen SY4D [28,29], Agrobacterium-mediated transformation of alfalfa has become easier and more efficient. To date, the transformation method that uses infection of the leaf as an explant by Agrobacterium tumefaciens, followed by the production of somatic embryos for transgenic plant regeneration, has been widely used in many laboratories [26]. Improving the efficiency of Agrobacterium-mediated transformation is crucial for enhancing virulence and increasing transformation frequency. Despite scattered studies on limited genotypes of alfalfa for transformation, the optimization and establishment of these protocols have primarily focused on limited parameters such as the use of different explants, genotypes, or cultivars; hormone components and ratios in plant regeneration media; in planta transformation; selection schemes; and Agrobacterium strains and transformation vectors [9,30,31,32,33,34,35]. However, many of these methods are either complex and costly or yield lower transformation frequency. A review of the existing literature has made it apparent that replicating the findings of prior studies presents significant challenges for the large-scale production of alfalfa for the pharmaceutical industry. These challenges occur within various genotypes where a large amount of variation is present. This requires the optimization of efficient transformation protocols.
Transient expression is a critical step for successful transformation, which is widely utilized across various areas such as gene expression, functional studies, plant biological material production [8], promoter characterization [36], gene silencing [37,38], elicitor identification [39], vaccine production [40], and CRISPR/Cas9-based genome editing [38,41]. Despite its widespread utility, a comprehensive review of the literature indicates that the optimization of transient gene expression in alfalfa remains underexplored and has not been directly addressed in extensive transformation studies. Previous research in this area has predominantly used single reporter genes and relied on kanamycin as a selectable marker [26,27], overlooking the superior efficacy of hygromycin in distinguishing transformed from non-transformed cells. To address this gap, the present study aims to establish a robust, versatile, and systematic protocol for optimizing Agrobacterium-mediated transient gene expression in alfalfa. Our research incorporates dual reporter genes, GUS and GFP, with hygromycin as a selectable marker, thereby enhancing the efficiency and effectiveness of transformation protocols in alfalfa. Therefore, the main objective of this study is to evaluate and improve various parameters in alfalfa transformation that influence transient gene expression.
2. Results
Transient assays are a valuable resource for examining gene function in plants. The Agrobacterium-mediated transformation approach is a rapid and exceptionally efficient technique that does not require costly equipment. Multiple methods have been documented to enhance transformation effectiveness for different plant species, as demonstrated in [42]. In this study, the effectiveness of the Agrobacterium-mediated transformation system in M. sativa was assessed by observing the influence of various factors. The success of the transformation was determined by the percentage of observed blue spots, indicating transient expression of GUS-positive transformants. GFP gene expression was identified by the presence of green fluorescent cells.
2.1. Effects of explant types, leaf age, and sizes of explants on transient GUS expression
The type of explant used influences the ability of the Agrobacterium to transfer T-DNA into the genome of the host plant cell [43,44,45,46]. Five explants—leaf, stem, petiole, root, and leaf-derived embryogenic callus initiated at 3 weeks in the dark—were selected as the target explants for this transient gene expression study. Approximately 2 mm leaf segments from well-grown, 3-week-old alfalfa plants were cultured in a callus induction medium. Callus initiation started with the curling of leaf layers, and after one week a greenish or yellowish callus was observed. The different phases of callus formation are presented in Figure 1A-C. To optimize the transient expression protocol, the transformation efficiencies of different explants were evaluated (Figure 2A-H). Our study recorded variations in transformation frequencies, with the highest transient GUS expression being observed in 76.2% of the leaf explants, followed by 38.7% for stems, 20.3% for petioles, 14.5% for roots, and 42.9% for calli in alfalfa (Figure 2A-H and Figure 3A).
The age of the leaf explant is a critical determinant of transient GUS expression [47]. The transient GUS expression decreased with the increasing age of the leaf explants. Young leaves, specifically those that were 3 weeks old, exhibited a markedly higher transient GUS expression (74.2%) compared to older leaves of 5 and 7 weeks old, which only exhibited 56.6% and 48.4% expression, respectively (Figure 3B). Leaves that were less than one week old yielded a transient GUS expression of only 30.1%. Conversely, leaf explants aged 9 weeks exhibited higher resistance to agro-infection, with only 28.6% GUS expression observed. One possible explanation could be that the older leaves possess a more robust defense mechanism, potentially diminishing the ability of Agrobacterium to grow and transfer T-DNA. However, leaves aged more than 5 weeks are too brittle for agro-infection. The sizes of leaf explants are also a crucial factor affecting transient GUS expression. Transformation efficiency was significantly decreased in whole or intact explants compared to excised ones. The excised explants demonstrated the highest GUS expression, at 91.1%, whereas only 56.2% expression was observed in the whole explants (Figure 2A,D and Figure 3C).
2.2. Effect of wounding types on transient GUS expression
In nature, Agrobacterium infects plants through natural openings, such as wounds. Therefore, the wounding or pricking of the plant cells is essential to establish a pathway from Agrobacterium infection, enabling the delivery of T-DNA to explants [48]. This is required to overcome physical barriers such as waxy cuticles on the plant’s epidermis, which could impede T-DNA transfer [47]. Additionally, a plant’s wound response stimulates the release of phenolic compounds for the activation of vir genes. Furthermore, cell division is stimulated, and the cell walls become less rigid, making it easier for the bacterium to breach. At this stage, active DNA replication can assist in T-DNA integration [48]. In this study, the effects of different wounding types of leaf explants were examined. There was a significant increase (over 70%) in T-DNA transfer observed in wounded plants as compared to unwounded explants infected with Agrobacterium. The highest transient GUS expression was found in wounding with a scalpel, at 86.3%, whereas that of wounding with a needle, wounding with mash, wounding with a cell strainer pestle, and control, unwounded, or intact were 53.5, 49.6, 35.9, and 12.9%, respectively (Figure 4).
2.3. Effects of Agrobacterium concentration or optimal density (OD600nm) on transient GUS expression
The efficiency of transformation depends upon the concentration of Agrobacterium, as adequate attachment of Agrobacterium to plant cells is crucial for successful gene transfer to take place [49]. Different plants exhibit varying transformation efficiencies with Agrobacterium concentrations ranging from 0.1 to 2.0 [50]. In this study, we tested the effect of Agrobacterium concentration on transformation efficiencies and found that the specific density (0.1–1.2 OD600nm) of Agrobacterium resulted in distinct levels of transient GUS expression in alfalfa explants. After 3 days of co-cultivation, the transformation rate of alfalfa explants significantly increased, with the Agrobacterium concentration ranging from 0.1 to 0.6 OD600nm. In this study, the greatest percentage of GUS-stained alfalfa explants (63.6%) was observed in a suspension culture of the Agrobacterium strain GV3101, at 0.6 OD600nm, followed by 0.4 and 0.8 with 55.9% and 45.3% GUS expression, respectively (Figure 5). However, the results obtained at 0.6 OD600nm were not significantly different from those at 0.4 OD600nm. High concentrations of Agrobacterium suspension cultures (1.0 and 1.2 of OD600nm) significantly decreased the number of transformed cells, by 42.5%, and 30.5%, respectively. High Agrobacterium concentrations increased the number of infected plant cells, but they also disrupted the physiological processes in the explants, leading to the termination of the transformation process.
2.4. Effects of preculture periods on transient GUS expression
A preculture period involves the culturing of the explants in preculture medium for different durations before Agrobacterium infection and co-cultivation [51]. During this time, the plant tissues undergo physiological and developmental changes that significantly enhance their ability to regenerate, thereby promoting transformation efficiency [45]. As the preculture medium supplemented with growth regulators generally promotes cell division, actively dividing cells are more suitable for the T-DNA delivery and integration of transgenes [52]. The impacts of varying preculture periods (ranging from 1 to 7 days) on the efficiency of alfalfa’s transient expression were evaluated. The results showed that alfalfa precultured for 2 days yielded the highest positive GUS results, at 72.7%, followed by preculture periods of 1 and 3 days, at 51.9% and 44.5%, respectively (Figure 6). With longer preculture periods (4, 5, 6, and 7 days) the transformation efficiency decreased to 43.8%, 38.7%, 29.6%, and 19.1%, respectively. The extended preculture period allowed the alfalfa explants to increase their nutrient uptake from the medium, stimulating cells to become competent for Agrobacterium attachment.
2.5. Effects of incubation periods on transient GUS expression
To achieve efficient Agrobacterium attachment and subsequent transformation of plant cells, a suitable incubation period is required. Precultured alfalfa leaf explants were immersed in an Agrobacterium suspension culture with an OD600nm of 0.6 for varying periods of 5 to 45 mins before co-cultivation. The duration of immersion significantly affected GUS expression, with the highest transient expression (78.5%) observed after 30 mins, followed by 25 and 35 mins, which displayed GUS expression levels of 72.3 and 62.9%, respectively (Figure 7). Incubation for less than 25 mins resulted in lower percentages of 30.9, 39.5, 50.3, and 56.3% GUS expression for 5, 10, 15, and 20 mins, respectively, due to insufficient time for the Agrobacterium to infect the leaf explants. However, extending the incubation period beyond 35 mins (data not shown) decreased the transient GUS expression due to Agrobacterium overgrowth, leading to stress and subsequent browning or blackening of the explants. Such stress could trigger necrotic and hypersensitive symptoms and negatively affect transformation rates.
2.6. Effects of the co-cultivation or post-incubation period on transient GUS expression
Co-cultivation is the process whereby explants are incubated with Agrobacterium. However, in different studies, it is also meant for the post-incubation period, which refers to the time between the removal of unattached Agrobacterium after agro-infection and the observation or selection of transgenic lines from the explants [51,53,54,55,56]. During this stage, alfalfa leaf explants are cultured with attached Agrobacterium to facilitate further infection on a medium lacking selection pressure and anti-Agrobacterium agents. The transient expression of GUS has been observed in alfalfa leaf explants co-cultivated with Agrobacterium for different periods (1 to 7 days). The highest frequency of 83.5% was achieved after 3 days of co-cultivation, while the transformation frequencies were much lower for 1- and 2-day co-cultivation periods, at 35.2% and 45.7%, respectively (Figure 8). Co-cultivation for 2–3 days has been used in Spinacia oleracea L. [45] and Moringa oleifera Lam. [57]. Co-cultivation periods of less than 3 days produced few transformed lines, due to insufficient time for complete or maximized transfer of Agrobacterium T-DNA into the alfalfa genome. Liu et al. (2008) [58] reported that no transient GUS-positive explants and spots in soybeans were found within 0 days of a co-cultivation period. After 4 days of co-cultivation in alfalfa leaf explants, around 62.5% transient expression was observed, and for a co-cultivation period of 5, 6, and 7 days, the frequency of blue spots on the alfalfa leaf explants was 42.9, 28.9, and 16.7%, respectively.
2.7. Effects of acetosyringone concentrations on transient GUS expression
Acetosyringone (AC), a phenolic inducer that affects the virulence genes of Agrobacterium, is essential for infection. AC is also necessary for the infection and co-cultivation medium in order to facilitate the transfer and incorporation of T-DNA into the recalcitrant host plant—particularly monocotyledons, which cannot naturally synthesize this compound and, therefore, are not suitable hosts for Agrobacterium. Alfalfa leaf explants were immersed in different concentrations of AC, ranging from 50-350 µM, in order to assess its effect on transformation. The results showed that the transient transformation efficiency significantly improved up to a concentration of 150 µM, indicating the critical role of AC in alfalfa transformation (Figure 9). In species where low levels of AC were present, such as Oncidium and Cymbidium orchids, Carrizo, and Hamlin lime, the vir genes were induced synergistically by sugars, e.g., glucose and galactose, leading to an increase in transformation efficiency [59]. In the case of alfalfa, the maximum percentage (71.4%) of GUS-positive leaf explants was observed with the addition of 150 µM AC, followed by 46.8% in 100 µM and 39.1% in 200 µM AC. However, at concentrations higher than 150 µM (250, 300, and 350 µM produced GUS expression of 33.2, 25, and 11.1%, respectively), the transient transformation efficiency decreased, and in some cases it was almost completely inhibited. A similar result was obtained in the transformation of Dendrobium orchids, where higher concentrations of AC resulted in the inhibition of transformation. The inhibition at higher concentrations could be attributed to the toxicity of AC to explants, which produced hypersensitive symptoms due to the harmful effect of supra-optimal AC concentration on Agrobacterium inoculation, thus affecting the transformation rate [20].
2.8. Effects of calcium chloride (CaCl2) concentrations on transient GUS expression
It is known that calcium is an essential macronutrient for plants, playing a crucial role in the cell wall structure; it acts as an ionic cross-linkage, binding carboxyl groups of linear macromolecules in the plant cell wall. Typically, the plant cell wall contains a concentration of 1–10 mM calcium chloride (CaCl2), which can make up more than 60% of the total calcium in the cell [20,60,61]. Research has shown that an increase in CaCl2 concentration can stimulate cell dissociation, leading to hyper-secretion of polysaccharide compounds [62]. Bacteria were made competent with CaCl2 and transformed with plasmids by heat shock [63]. In this study on alfalfa leaf explants, different treatments with varying CaCl2 strengths (ranging from 1 to 10 mM) were evaluated, and the highest percentage of GUS expression (68.4%), a marker for transient gene expression, was observed at a CaCl2 concentration of 4 mM, followed by 58.2% and 47.3% for CaCl2 concentrations of 3 and 5 mM, respectively (Figure 10). However, transient GUS activity decreased by 23.8% when the CaCl2 concentration exceeded 6 mM.
2.9. Effects of silver nitrate (AgNO3) concentrations on transient GUS expression
Ethylene, a colorless hydrocarbon-based gas, is produced during agro-infection, which can reduce the efficiency of gene transfer mechanisms in Agrobacterium-mediated transformation. However, silver nitrate (AgNO3) has been found to inhibit ethylene production, which affects cell division mechanisms [64]. Ethylene production is enhanced by wounding during explant preparation, which can lead to browning or tissue damage. Nevertheless, adding AgNO3 to the co-cultivation medium at an optimal level can significantly suppress Agrobacterium growth, without compromising T-DNA delivery and subsequent T-DNA integration, by reducing the capacity of Ag2+ to bind to the ethylene receptor produced through ethylene biosynthesis [65].
Tissue browning is a significant factor that can reduce transformation efficiency in plants [66], including alfalfa. To assess transformation efficiency and prevent tissue browning based on transient GUS expression, the impacts of varying concentrations of AgNO3 (10–150 µM) on alfalfa leaf explants were investigated. The results showed that the highest percentage of transient GUS gene expression (69.1%) was observed at 75 µM, followed by 38.3 and 46.5% for 50 and 100 µM, respectively. Conversely, 33.6% expression was seen at 150 µM AgNO3 (Figure 11). We postulate that this could be attributed to the phytotoxic effects of high AgNO3 concentrations during co-cultivation on alfalfa leaf explants.
2.10. Green Fluorescence Protein (GFP) gene transient expression
Successful gene delivery into alfalfa explants was confirmed through GFP gene expression, with transformed cells displaying GFP production and becoming distinguishable from non-transformed cells. Transient green fluorescence was detected in alfalfa leaf explants (Figure 12A-F) and leaf-derived calli (Figure 12G-L) compared with non-transformed plants, which emitted chlorophyll-induced red autofluorescence. The leaves of alfalfa explants produced GFP after 3 days of co-cultivation with Agrobacterium, while the GFP-positive callus was developed 3–4 weeks after co-cultivation in an optimized medium, and then gradually decreased in intensity thereafter. Approximately 58% of the alfalfa explants exhibited transient GFP gene expression. Notably, severe tissue necrosis, characterized by browning of the culture, was observed following co-cultivation. GFP-positive callus formations were detected from co-cultivated explants after 28 days of culture (data not shown).
3. Discussion
Tissue culture and transformation are fundamental techniques in functional plant genomics. Although Agrobacterium-mediated transformation has been widely employed in various plant species for decades, some legume crops, such as alfalfa, continue to pose challenges for achieving efficient genetic transformation and high-level expression of target genes [9]. Since the first report of Agrobacterium tumefaciens-mediated transformation in alfalfa [11], numerous studies have investigated various methods for the regeneration and genetic transformation of alfalfa [9,32,33], although these methods often prove less effective for this species. Identifying all factors that affect transformation efficiency is crucial for developing effective expression systems in recalcitrant legume crops such as alfalfa. Agrobacterium-mediated transformation for transient gene expression is particularly useful for this purpose, as it facilitates the rapid testing of a variety of parameters. In comparison to existing transformation procedures, our newly established method offers several advantages. Our research on transient expression in alfalfa has shown that different explants can successfully express GUS and GFP genes. This study highlights that our transient expression protocol effectively optimizes key parameters, leading to high levels of GFP and GUS expression.
The types of explants, age of leaves, and tissue sizes are crucial factors in Agrobacterium-mediated transient GUS expression. In this study, we found that leaf explants excised from 3-week-old plants exhibited the highest GUS expression levels compared to stem-, petiole-, root-, and leaf-derived embryogenic calli. One possible explanation for this could be that intact or whole leaves older than 3 weeks have a more developed defense mechanism, which might reduce the ability of Agrobacterium to grow and transfer T-DNA. In addition, the highest transient GUS expression was found in wounding with a scalpel compared to other approaches. Several methods of wounding are employed in plant transformation, including small incisions made with blades, micro-wounding using particle guns or sonication, and injection via a syringe [49]. However, mechanical wounding has significantly improved transformation efficiency in some plant species, including recalcitrant grain legumes such as green gram [52], black gram [67], soybeans [48], and chickpeas [51].
The optimal density of the Agrobacterium culture significantly influences both stable and transient transformation efficiency. Our finding of higher GUS expression with the Agrobacterium concentration at 0.6 OD600nm is consistent with those of previous studies conducted in chickpeas [51,56], rice [53], ramie [68], peanuts [69,70], and soybeans [48]. The results highlight that both variable Agrobacterium concentrations and plant species are crucial factors in the efficacy of plant transformation mediated by Agrobacterium. Our findings also highlight the importance of optimizing the preculture period for successful plant transformation experiments, as the transient gene expression in our study had high transformation efficiency at 2 days of preculture. While successful applications of explant preculture before Agrobacterium infection have been observed in various leguminous plants, such as soybeans [71], peanuts [69], green gram [72], and chickpeas [51], it is nevertheless crucial to note that the ideal preculture period may differ for other plant species [57] and can be influenced by numerous factors, such as the type of tissue and the transformation method employed.
Notably, the optimal incubation time is variable among plant species and explant tissue types. For instance, a 30-min incubation was sufficient for transient expression in cotyledonary nodes of soybeans [44] and chickpeas [46], while a 60-min incubation was needed for in vitro Melastomataceae spp. explants [73]. In our study, the highest transient expression (78.5%) was observed after 30 mins. During the co-cultivation process, explants were incubated with Agrobacterium. The transient GUS expression was low after 1 day of co-cultivation, and the maximum result was at 3 days in our study, which is consistent with previous findings reported in Dierama erectum Hilliard [49], broccoli [74], bananas [66], chickpeas [46], and Phalaenopsis violacea orchids [20]. However, longer co-cultivation periods of 6–7 days also significantly reduced the transient GUS expression efficiency due to the overgrowth of other bacteria, with a contaminant resulting in suffocation of the alfalfa leaf explants and turning necrotic. Comparable results have been reported in chickpeas [55], Vigna radiata [72], Vigna mungo [75], and Glycine max [48].
Li et al. (2022) [76] demonstrated the involvement of various molecules in the transfer of T-DNA from bacterium to host, stimulated by proteins encoded by the virulence (vir) genes. Acetosyringone (AC) is one of these phenolic inducer molecules that stimulate the genes of Agrobacterium. However, in some plant species—such as poplar, Mexican lime, and tobacco—AC did not improve the transformation efficiency in the Agrobacterium-mediated system, possibly due to the production of high amounts of phenolics secreted by the plant tissues, rendering AC unable to induce virulence and infection [59]. Nevertheless, the addition of AC during infection and co-cultivation has been shown to significantly improve the Agrobacterium-mediated system in some plants, including Dierama erectum Hilliard [49], Phalaenopsis orchids [20], and Spinacia oleracea L. [45]. In our study, the transient transformation efficiency significantly improved at a concentration of 150 µM, which is comparable to previous findings where the application of 100–500 µM AC to the co-culture medium was effective for Agrobacterium-mediated transformation in peanuts [70], black gram [75], cowpeas [77], green gram [72], soybeans [48], and chickpeas [51].
Calcium is a vital macronutrient that is essential for cell wall repair. Reduced calcium concentrations can cause cell wall deterioration. The study demonstrated that supplementation with 4 mM calcium chloride enhanced transient transformation efficiency. Interestingly, a lack of calcium in plants may increase the transformation rate, as it can alter the structure of the cell wall, reducing the matrix and making it less conducive to Agrobacterium cell attachment, as supported by the results of transient GUS gene expression in the calcium-free medium [78,79]. On the other hand, higher calcium concentrations can reinforce the plant cell wall by being absorbed into it. Calcium also acts as an inducer, influencing gene expression and changing the composition of the cell wall. Strengthening the cell wall may result in lower rates of T-DNA gene segment transfer from Agrobacterium to plant cells.
Silver nitrate is an anti-necrotic agent that can minimize oxidative bursts during the interaction between plant tissue and A. tumefaciens [66,80,81]. The addition of AgNO3 to the co-culture medium has been known to exhibit anti-ethylene activity, which is a common occurrence during in vitro plant cultures. This subsequently slows down Agrobacterium growth on the target explants, ultimately facilitating plant cell recovery and increasing transformation efficiency in various plant species, such as soybeans [82], cassava [83], Prunus avium (L.) cv Stella [80], maize [84], apple [85], Phalaenopsis violacea, and Dendrobium orchids [20]. The use of AgNO3 has been found to have significant effects on plant tissue culture, including enhancement of somatic embryogenesis, organogenesis, and micropropagation in many species [86]. In the present study, the maximum transient transformation efficiency improved at a concentration of 75 µM. Previous studies have shown that the addition of varying concentrations of AgNO3 increased the rate of shoot elongation in the 88-1 soybean variety. Moreover, the transformation efficiency of the 88-1 soybean variety also showed an improvement, rising from 3.2 to 5.5% upon the addition of 15 mg/L AgNO3 [44]. In addition, AgNO3 has been shown to stimulate direct shoot regeneration in in vitro cultures and, when co-cultivated with A. tumefaciens for genetic transformation studies, it inhibits bacterial growth after co-cultivation. For instance, the addition of 100 mg/L AgNO3 to the plant regeneration medium after co-cultivation can completely halt bacterial growth for at least 3 weeks. As a result, AgNO3 can serve as a substitute for commonly used antibiotics such as cefotaxime and carbenicillin, which are frequently employed during the initial passage after co-cultivation with Agrobacterium strains in wheat [61,87].
4. Materials and Methods
4.1. Establishment of In vitro cultures
Seeds of the alfalfa cultivar Regen-SY germplasm (PI 537440) were collected from Western Regional PI Station through the U.S. National Plant Germplasm System. Alfalfa seeds were surface sterilized with 70% ethyl alcohol for 30 s followed by 20% bleach (Clorox®) treatment for 10 min [88]. The treated seeds were rinsed with sterile distilled water three-to-five times and then placed in 100 mm × 15 mm Petri dishes on Murashige and Skoog (MS) basal medium (PhytoTechnology Laboratories, KS, USA) containing 3% sucrose and 0.8% w/v agar at pH 5.7.
The Petri dishes were kept in the dark for 3 weeks at 24 ◦C. After 3 weeks, nine germinated seeds per Petri dish were inoculated in the magenta GA7 boxes and incubated at a temperature of 24 °C for 7 days in a growth chamber supplied with a 16/8 h photoperiod using cool, white, fluorescent light (75 lmol s-1 m-2). Emerged seedlings with well-developed shoots and roots were maintained in a growth chamber, and only healthy plantlets were chosen for further propagation. Sterile scalpels were used to excise 3-week-old subcultured plants with four different explants, namely, the leaves, stems, petioles, and roots. Callus was obtained from alfalfa leaf explants following the modified protocol [88,89]. Fully expanded leaves from 3-week-old alfalfa plants, were carefully removed, and cut into small segments of uniform size (≈2 mm). Leaf explants were placed on a callus induction medium using Gamborg’s B5 basal (B5H) medium, supplemented with 3% maltose, 4.5 μM 2,4-dichlorophenoxyacetic acid (2,4-D), 0.9 μM kinetin, 6.65 g of glutamine, 0.8 g of serine, 0.004 g of adenine, and 0.08 g of L-glutathione. The cultures were incubated in the dark at 24 ± 2 ℃ for 3 weeks [90,91].
4.2. Agrobacterium Strain and Plasmid Vector
A. tumefaciens strain GV3101 harboring the pCAMBIA1304 plasmid expressing both GUS and GFP gene fusion was used for transformation. Both GFP-GUS gene fusions were under the control of a Cauliflower mosaic virus (CaMV) 35s promoter and NOS terminator. The T-DNA region of the plasmid also contained the hygromycin B phosphotransferase gene, driven by the CaMV 35s promoter, as a selection marker; pCAMBIA1304 has been routinely used to demonstrate or establish the transient expression of several plant species, such as Arabidopsis [31], several medicinal plants [92], rice [93], tobacco [94], and Dendrobium Sonia [95]. The vector pCAMBIA1300—not containing reporter genes—was used as a control. A single Agrobacterium colony derived from stock culture as described by Dutt and Grosser (2009)[59] was cultured in liquid Luria–Bertani (LB) medium [92] containing 100 mg L-1 kanamycin and was grown on a shaker at 120 rpm at a temperature of 28 °C for 16 hrs.
4.3. Inoculation and co-cultivation of alfalfa with A. tumefaciens
Glycerol stocks containing 12% glycerol were utilized to maintain the Agrobacterium culture at -20 ˚C, with initiation 3 days prior to weekly experiments. The Agrobacterium strain GV3101 with construct pCAMBIA1304 was grown on Luria–Bertani (LB) agar containing 100 mg/L kanamycin and 100 mg/L gentamicin at 28 ˚C for 2 days. A single colony of Agrobacterium from a fresh subculture plate was collected with a sterile loop inoculated into 2 mL of LB broth liquid medium containing 100 mg/L kanamycin and 100 mg/L gentamicin for 8 hours at 28 ˚C, 200 rpm. The following day, 200 µl of bacterial suspension was cultured from the stock into 15 ml of liquid LB medium and grown overnight (24 hrs.) at 28 °C in a shaker with 200 rpm agitation until the desired density at OD600nm was obtained. The selected explants were then inoculated with the prepared Agrobacterium. To eliminate Agrobacterium, the explants were washed twice with MS basal liquid medium, dried using filter paper, and then placed on a selection medium containing 350 mg/L cefotaxime and 12.5 mg/L hygromycin (Figure 13).
4.4. Optimization of transient gene expression parameters
Several factors affecting Agrobacterium-mediated transformation frequency in alfalfa were evaluated. Factors such as explant type (leaf-, stem-, petiole-, root-, and leaf-derived 3-week-old calli), the age of the explant (1, 3, 5, 7, and 9 weeks old), the size of leaf explants (excised and whole), wounding technique (control, unwounded, or intact; wounding with cell strainer pestle; wounding with 80 µm mash; wounding with a scalpel; wounding with a 0.1 mm needle), Agrobacterium concentration (0, 0.1, 0.2, 0.4, 0.6, 0.8, 1.0, and 1.2 at OD600nm), preculture period (0, 1, 2, 3, 4, 5, 6, and 7 days), co-cultivation period (0, 1, 2, 3, 4, 5, 6, and 7 days), incubation period (0, 5, 10, 15, 20, 25, 30, 35, and 45 min), acetosyringone concentration in incubation solution (0, 50, 100, 150, 200, 250, 300, and 350 µM), calcium strength (0, 1, 2, 3, 4, 5, 6, and 10 mM), and silver nitrate (0, 10, 25, 50, 75, 100, 125, and 150 µM) were investigated.
To determine the optimal conditions for transformation, several parameters were used, and their effects on the percentage of GFP and transient GUS gene expression were evaluated. Explants co-cultured without agro-infection were used as controls. At the end of the co-cultivation period, the treated leaf explants were assessed and optimized based on their GFP gene expression (visible under a microscope with fluorescent green light) and GUS gene expression (visible as blue spots). The results were determined based on the percentage of explants exhibiting GFP and GUS spots out of the total number of inoculated explants, which were observed 3 days after transformation. A sample was considered to be GUS-positive if at least 25% of the leaf explant displayed blue spots. This experiment was conducted three times, and equivalent results were obtained. The experiments were performed using 16 individual explants and repeated four times.
4.5. GUS histochemical activity assay and GFP assay
Confirmation of transient transformed events for GUS activity was performed in different tissues according to the method described by Jefferson in 1987 [21]. After 3 days of co-cultivation, the alfalfa explants were immersed in a solution of 2 mM X-Gluc (5-bromo-4-chloro-3-indolyl—β-D-glucuronide) containing 0.1 M NaH2PO4 (pH 7.0), 0.1 M potassium ferricyanide, 0.1 M ferrocyanide, and 0.1% (v/v) Triton X-100. Before the observation of transient gene expression in different explants, they were incubated at 37 °C for 16–24 hrs. For removing chlorophyll, the X-Glue solution was replaced with double distilled water followed by washing with 75% ethanol. The mixture of acetone/methanol (1:3) was added and incubated at 4 oC for 1 h. The explants were washed with double distilled water 4–5 times after obtaining clear tissue and then stored in 50% glycerol. The transient GUS activity of the alfalfa explants was examined under a Leica microsystems EMSPIRA 3 digital microscope with a magnification of 1.2x (Life Sciences companies @ Danaher corporation, USA) and scored as blue spots, irrespective of size.
GFP gene expression in alfalfa tissues was observed using an Olympus SZX12 Stereo fluorescence-equipped microscope with an HBO 100 W mercury bulb light mounted with a long-pass GFP filter supplied with a DP72 camera. The magnification level used for the observation was 16x. The GFP filter with excitation wavelengths of 460–480 nm, emission wavelengths of 495–540 nm, along with a GFPA filter for separation of GFP and blue-excited fluorescence with excitation wavelengths of 460–490 nm and emission wavelengths of 510–550 nm, was used to identity the leaves, stems, petioles, roots, and embryogenic calli expressing GFP events. Photographs were taken with an Olympus SZX12 automatic exposure photomicrographic system. To determine the percentage of transient GFP expression, the number of explants producing a GFP positive was divided by the total number of explants co-cultivated with Agrobacterium. An explant with ten or more cells that expressed GFP was considered to be positive for transient expression.
4.6. Determination of Agrobacterium Infection Efficiency and Statistical Analysis
The efficiency of Agrobacterium infection was calculated by measuring the percentage of transient GUS expression in alfalfa explants. The rate of transient GUS expression (%) was calculated using the following formula: rate of transient GUS expression (%) = (number of explants exhibiting blue coloration/total number of explants stained) × 100%. Statistical analysis was performed using one-way analysis of variance (ANOVA). The differences in means between the two groups were assessed using Student’s t-test, while differences among multiple treatments were analyzed using Duncan’s multiple range test [96]. Graphs were generated using Prism v. 5.0 (GraphPad Software, La Jolla, CA) and Microsoft Excel.
5. Conclusions
Genetic transformation is a promising technique for improving crops through incorporating desired traits into their genomes. However, a significant challenge in this process is to increase the responsiveness of cells to Agrobacterium-mediated transformation. Hence, it is essential to identify and optimize critical parameters that positively influence Agrobacterium-mediated transformation efficiency during the infection and co-cultivation stages. This study aimed to improve existing protocols through systematically evaluating various parameters to achieve higher levels of transient gene expression in alfalfa. Additionally, this study extends the benefits of the Agrobacterium-mediated transformation system by employing a dual reporter system—GUS and GFP—along with hygromycin as a selectable marker. The GUS histochemical assay developed for alfalfa in this study could offer a rapid method for visualizing spatial gene expression and functionally testing gene promoters and other cis-regulatory elements in alfalfa. Furthermore, the GFP reporter gene is used as a form of non-invasive and real-time visualization, enabling the tracking of gene expression without damaging the cells and tissues. The dual reporter, combining GUS and GFP, enhances the precision of monitoring and visualizing gene expression and provides cross-validation of results through reducing the likelihood of false positives or negatives and validating the efficiency of gene expression.
Standardizing these parameters will facilitate the efficient transfer of agronomically important traits for the improvement of alfalfa. Moreover, optimizing transient gene expression in tissues may require the use of various additives to manage Agrobacterium overgrowth. Our research highlights key factors that influence T-DNA transfer efficiency and gene expression in alfalfa. Analysis of transient gene expression confirmed successful transformation in alfalfa using the Agrobacterium strain. The presence of blue spots in alfalfa explants indicates GUS reporter gene expression throughout the plants, driven by the constitutive CaMV 35s promoter. This successful Agrobacterium-mediated transformation of alfalfa could be further optimized for generating stable transformants using genome editing techniques such as CRISPR/Cas9, facilitating the functional testing of both genes and cis-regulatory regions such as promoters and enhancers. Our findings not only enhance the existing alfalfa transformation protocol, but also emphasize the importance of a standardized transformation protocol to maximize the yield of transgenic alfalfa plants.
Author Contributions
Conceptualization, S.B. and S.K.D.; methodology, S.B.; software, S.B.; validation, S.B. and S.K.D.; formal analysis, S.B.; investigation, S.B. and S.K.D.; resources, S.K.D.; data curation, S.B.; writing—original draft preparation, S.B.; writing—review and editing, S.B., D.P., S.D., A.S., and S.K.D.; visualization, S.B.; supervision, S.K.D.; project administration, S.K.D.; funding acquisition, S.K.D. All authors have read and agreed to the published version of the manuscript.
Funding
This project was partially supported by the National Institute of Food and Agriculture (NIFA) Agricultural Research at 1890 Land-Grant Institutions, Evans-Allen capacity grant [No. GEOX-5223]. This research was funded by grants from the Department of Education, Minority Science, and Engineering Improvement Program (MSEIP: grant number P120A2000016) and the National Science Foundation Historically Black Colleges and Universities (HBCU)-Undergraduate Program (HBCU-UP: grant number HRD-2011903).
Data Availability Statement
All data are available within the manuscript, and further information can be obtained upon request from the corresponding authors
Acknowledgments
The authors would like to express their sincere thanks and gratitude to Dr. Deborah Samac at the University of Minnesota (USDA/ARS) for supplying the plant material, as wel as thanks to the graduate students for their support during the research work.
We would like to extend our sincere gratitude to Dr. Amit Dhingraat theDepartment of Horticultural Sciences, Texas A&M University, College Station, TX 77843, USA, for providing us with the pCAMBIA1304 vector.
Conflicts of Interest
The authors declare no potential conflicts of interest.
References
- Yazicilar, B.; Chang, Y.L. Embryogenic Callus Differentiation in Short-Term Callus Derived from Leaf Explants of Alfalfa Cultivars. Natural Products and Biotechnology 2022, 2, 99–104. [Google Scholar] [CrossRef]
- Bora, K.S.; Sharma, A. Phytochemical and pharmacological potential of Medicago sativa: A review. Pharmaceutical Biology 2011, 49, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Fu, G.; Grbic, V.; Ma, S.; Tian, L. Evaluation of somatic embryos of alfalfa for recombinant protein expression. Plant Cell Reports 2015, 34, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Nirola, R.; Megharaj, M.; Beecham, S.; Aryal, R.; Thavamani, P.; Vankateswarlu, K.; Saint, C. Remediation of metalliferous mines, revegetation challenges and emerging prospects in semi-arid and arid conditions. Environmental Science and Pollution Research 2016, 23, 20131–20150. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S. Biotechnological advancements in alfalfa improvement. Journal of Applied Genetics 2011, 52, 111–124. [Google Scholar] [CrossRef]
- Tichá, M.; Illésová, P.; Hrbáčková, M.; Basheer, J.; Novák, D.; Hlaváčková, K.; Šamajová, O.; Niehaus, K.; Ovečka, M.; Šamaj, J. Tissue culture, genetic transformation, interaction with beneficial microbes, and modern bio-imaging techniques in alfalfa research. Critical Reviews in Biotechnology 2020, 40, 1265–1280. [Google Scholar] [CrossRef]
- Vignesh, M.; Shanmugavadivel, P.S.; Prabha, M.; Kokiladevi, E. Transformation Studies in Pea–A Review. Agricultural Reviews 2010, 31, 68–72. [Google Scholar]
- Liu, K.; Yang, Q.; Yang, T.; Wu, Y.; Wang, G.; Yang, F.; Wang, R.; Lin, X.; Li, G. Development of Agrobacterium-mediated transient expression system in Caragana intermedia and characterization of CiDREB1C in stress response. BMC Plant Biology 2019, 19, 1–13. [Google Scholar] [CrossRef]
- Liu, W.; Liang, Z.; Shan, C.; Marsolais, F.; Tian, L. Genetic transformation and full recovery of alfalfa plants via secondary somatic embryogenesis. In Vitro Cellular Developmental Biology-Plant 2013, 49, 17–23. [Google Scholar] [CrossRef]
- Azizi-Dargahlou, S.; Pouresmaeil, M. (2023). Agrobacterium tumefaciens-mediated Plant Transformation: A Review. Molecular Biotechnology, 2023; 1–18. [Google Scholar]
- Deak, M.; Kiss, G.B.; Koncz, C.; Dudits, D. Transformation of Medicago by Agrobacterium-mediated gene transfer. Plant Cell Reports 1986, 5, 97–100. [Google Scholar] [CrossRef]
- Tohidfar, M.; Zare, N.; Jouzani, G.S.; Eftekhari, S.M. Agrobacterium-mediated transformation of alfalfa (Medicago sativa) using a synthetic cry3a gene to enhance resistance against alfalfa weevil. Plant Cell, Tissue and Organ Culture (PCTOC) 2013, 113, 227–235. [Google Scholar] [CrossRef]
- Jin, T.; Chang, Q.; Li, W.; Yin, D.; Li, Z.; Wang, D.; Liu, B.; Liu, L. Stress-inducible expression of GmDREB1 conferred salt tolerance in transgenic alfalfa. Plant Cell, Tissue and Organ Culture (PCTOC) 2010, 100, 219–227. [Google Scholar] [CrossRef]
- Zhang, W.J.; Wang, T. Enhanced salt tolerance of alfalfa (Medicago sativa) by rstB gene transformation. Plant Science 2015, 234, 110–118. [Google Scholar] [CrossRef]
- Avraham, T.; Badani, H.; Galili, S.; Amir, R. (2005). Enhanced levels of methionine and cysteine in transgenic alfalfa (Medicago sativa L.) plants over-expressing the Arabidopsis cystathionine γ-synthase gene. Plant Biotechnology Journal 2005, 3, 71–79. [Google Scholar] [CrossRef]
- Calderini, O.; Bovone, T.; Scotti, C.; Pupilli, F.; Piano, E.; Arcioni, S. Delay of leaf senescence in Medicago sativa transformed with the ipt gene controlled by the senescence-specific promoter SAG12. Plant Cell Reports 2007, 26, 611–615. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Gao, Y.; Wu, X.; Ma, H.; Zheng, C.; Wang, X.; Zhang, H.; Yang, H. The relative contributions of pH, organic anions, and phosphatase to rhizosphere soil phosphorus mobilization and crop phosphorus uptake in maize/alfalfa polyculture. Plant and Soil 2020, 447, 117–133. [Google Scholar] [CrossRef]
- Komel, T.; Bosnjak, M.; Sersa, G.; Cemazar, M. Expression of GFP and DsRed fluorescent proteins after gene electrotransfer of tumor cells in vitro. Bioelectrochemistry 2023, 108490. [Google Scholar] [CrossRef] [PubMed]
- Udayabhanu, J.; Huang, T.; Xin, S.; Cheng, J.; Hua, Y.; Huang, H. Optimization of the transformation protocol for increased efficiency of genetic transformation in Hevea brasiliensis. Plants 2022, 11, 1067. [Google Scholar] [CrossRef]
- Sreeramanan, S.; Maziah, M.; Abdullah, M.P.; Sariah, M.; Xavier, R. Transient expression of gusA and gfp gene in Agrobacterium-mediated banana transformation using single tiny meristematic bud. Asian Journal of Plant Science 2006, 5, 468–480. [Google Scholar]
- Jefferson, R.A. Assaying chimeric genes in plants: the GUS gene fusion system. Plant Molecular Biology Reporter 1987, 5, 387–405. [Google Scholar] [CrossRef]
- Shimomura, O.; Johnson, F.H.; Saiga, Y. Extraction, purification and properties of aequorin, a bioluminescent protein from the luminous hydromedusan, Aequorea. Journal of Cellular and Comparative Physiology 1962, 59, 223–239. [Google Scholar] [CrossRef]
- Qin, Y.; Wang, D.; Fu, J.; Zhang, Z.; Qin, Y.; Hu, G.; Zhao, J. Agrobacterium rhizogenes-mediated hairy root transformation as an efficient system for gene function analysis in Litchi chinensis. Plant Methods 2021, 17, 1–9. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Brummer, E.C. Is genetic engineering ever going to take off in forage, turf and bioenergy crop breeding? Annals of Botany 2012, 110, 1317–1325. [Google Scholar] [CrossRef] [PubMed]
- Austin, S.; Bingham, E.T.; Mathews, D.E.; Shahan, M.N.; Will, J.; Burgess, R.R. Production and field performance of transgenic alfalfa (Medicago sativa L.) expressing alpha-amylase and manganese-dependent lignin peroxidase. In The Methodology of Plant Genetic Manipulation: Criteria for Decision Making: Proceedings of the Eucarpia Plant Genetic Manipulation Section Meeting held at Cork, Ireland. Springer Netherlands, 1995, September 11 to September 14, pp. 381–393.
- Samac, D.A.; Austin-Phillips, S. Alfalfa (Medicago sativa L.). Agrobacterium Protocols 2006, 301–312. [Google Scholar]
- Shahin, E.A.; Spielmann, A.; Sukhapinda, K.; Simpson, R.B.; Yashar, M. Transformation of Cultivated Alfalfa Using Disarmed Agrobacterium tumefaciens. Crop Science 1986, 26, 1235–1239. [Google Scholar] [CrossRef]
- Bingham, E.T. Registration of alfalfa hybrid Regen-SY germplasm for tissue culture and transformation research. Crop Science 1991, 31, 1098–1118. [Google Scholar]
- Chen, F.; Srinivasa Reddy, M.S.; Temple, S.; Jackson, L.; Shadle, G.; Dixon, R.A. Multi-site genetic modulation of monolignol biosynthesis suggests new routes for formation of syringyl lignin and wall-bound ferulic acid in alfalfa (Medicago sativa L.). The Plant Journal 2006, 48, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Zhang, Y.; Wu, H.Z.; Min, X.; Zhang, B.; Kim, D.S.; Yan, X.; Zhang, C.J. Optimization and establishment of Agrobacterium-mediated transformation in alfalfa (Medicago sativa L.) using eGFP as a visual reporter. Plant Cell, Tissue and Organ Culture (PCTOC) 2024, 156, 7. [Google Scholar] [CrossRef]
- Jiang, Q.; Fu, C.; Wang, Z.Y. A unified Agrobacterium-mediated transformation protocol for alfalfa (Medicago sativa L.) and Medicago truncatula. Transgenic Plants: Methods and Protocols 2019, 153–163. [Google Scholar]
- Ninkovié, S. ; J. Miljus-Djukic, J.; Vinterhalter, B.; Neskovic, M. Improved transformation of alfalfa somatic embryos using a superbinary vector. Acta Biologica Cracoviensia Series Botanica, 2004; 46, 139–143. [Google Scholar]
- Rosellini, D. , Stefano, C.; Nicoletta, F.; Maria, L.S.S.; Alessandro, N.; Fabio, V. Non-antibiotic, efficient selection for alfalfa genetic engineering. Plant cell reports 2007, 26, 1035–1044. [Google Scholar]
- Weeks, J.T.; Ye, J.; Rommens, C.M. Development of an in-planta method for transformation of alfalfa (Medicago sativa). Transgenic Research 2008, 17, 587–597. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Huang, Q.M.; Jin, S.U. Development of alfalfa (Medicago sativa L.) regeneration system and Agrobacterium-mediated genetic transformation. Agricultural Sciences in China 2010, 9, 170–178. [Google Scholar] [CrossRef]
- Xu, H.; Chang, Q.; Huang, L.; Wei, P.; Song, Y.; Guo, Z.; Peng, Y.; Fan, J. An Agrobacterium-mediated Transient Expression Method for Functional Assay of Genes Promoting Disease in Monocots. International Journal of Molecular Sciences 2023, 24, 7636. [Google Scholar] [CrossRef] [PubMed]
- Dubey, N.K.; Eizenberg, H.; Edelstein, M.; Gal-On, A.; Aly, R. Enhanced host-parasite resistance based on down-regulation of Phelipanche aegyptiaca target genes is likely by mobile small RNA. Frontiers in Plant Science 2017, 8, 251119. [Google Scholar] [CrossRef]
- Kaur, M.; Manchanda, P.; Kalia, A.; Ahmed, F.K.; Nepovimova, E.; Kuca, K.; Abd-Elsalam, K.A. Agroinfiltration mediated scalable transient gene expression in genome-edited crop plants. International Journal of Molecular Sciences 2021, 22, 10882. [Google Scholar] [CrossRef]
- Wu, H.Y.; Liu, K.H.; Wang, Y.C.; Wu, J.F.; Chiu, W.L.; Chen, C.Y.; Wu, S.H.; Sheen, J.; Lai, E.M. AGROBEST: an efficient Agrobacterium-mediated transient expression method for versatile gene function analyses in Arabidopsis seedlings. Plant Methods 2014, 10, 1–16. [Google Scholar] [CrossRef]
- Spiegel, H.; Schillberg, S.; Nölke, G. Production of Recombinant Proteins by Agrobacterium-mediated Transient Expression. In Recombinant Proteins in Plants: Methods and Protocols. New York, NY: Springer US, 2022, pp. 89–102.
- Laforest, L.C.; Nadakuduti, S.S. Advances in delivery mechanisms of CRISPR gene-editing reagents in plants. Frontiers in Genome Editing 2022, 4, 830178. [Google Scholar] [CrossRef]
- Bhatt, R.; Asopa, P.P.; Jain, R.; Kothari-Chajer, A.; Kothari, S.L.; Kachhwaha, S. Optimization of Agrobacterium mediated genetic transformation in Paspalum scrobiculatum L. (Kodo Millet). Agronomy 2021, 11, 1104. [Google Scholar] [CrossRef]
- Atif, R.M.; Patat-Ochatt, E.M.; Svabova, L.; Ondrej, V.; Klenoticova, H.; Jacas, L.; Griga, M.; Ochatt, S.J. Gene transfer in legumes. In Progress in Botany: Berlin, Heidelberg: Springer Berlin Heidelberg, 2012, Vol. 74, pp. 37–100.
- Li, S.; Cong, Y.; Liu, Y.; Wang, T.; Shuai, Q.; Chen, N.; Gai, J.; Li, Y. Optimization of Agrobacterium-mediated transformation in soybean. Frontiers in Plant Science 2017, 8, 246. [Google Scholar] [CrossRef]
- Naderi, D.; Zohrabi, Z.; Shakib, A.M.; Mahmoudi, E.; Khasmakhi-Sabet, S.A.; Olfati, J.A. Optimization of micropropagation and Agrobacterium-mediated gene transformation to spinach (Spinacia oleracea L.). Advances in Bioscience and Biotechnology 2012, 3, 876–880. [Google Scholar] [CrossRef]
- Sadhu, S.K.; Jogam, P.; Gande, K.; Banoth, R.; Penna, S.; Peddaboina, V. Optimization of different factors for an Agrobacterium-mediated genetic transformation system using embryo axis explants of chickpea (Cicer arietinum L.). Journal of Plant Biotechnology 2022, 49, 61–73. [Google Scholar] [CrossRef]
- Salazar-González, J.; Castro-Medina, M.; Martínez-Terrazas, E.; Casson, S.A.; Urrea-Lopez, R. Leaf wounding and jasmonic acid act synergistically to enable efficient Agrobacterium-mediated transient transformation of Persea americana. Research Square 2022, 1–18. [Google Scholar]
- Hada, A.; Krishnan, V.; Mohamed Jaabir, M.S.; Kumari, A.; Jolly, M.; Praveen, S.; Sachdev, A. Improved Agrobacterium tumefaciens-mediated transformation of soybean [Glycine max (L.) Merr.] following optimization of culture conditions and mechanical techniques. In Vitro Cellular Developmental Biology-Plant 2018, 54, 672–688. [Google Scholar] [CrossRef]
- Koetle, M.J.; Baskaran, P.; Finnie, J.F.; Soos, V.; Balázs, E.; Van Staden, J. Optimization of transient GUS expression of Agrobacterium-mediated transformation in Dierama erectum Hilliard using sonication and Agrobacterium. South African Journal of Botany 2017, 111, 307–312. [Google Scholar] [CrossRef]
- Wang, Y.L.; Xu, M.X.; Yin, G.X.; Tao, L.L.; Wang, D.W.; Ye, X.G. Transgenic wheat plants derived from Agrobacterium-mediated transformation of mature embryo tissues. Cereal Research Communications 2009, 37, 1–12. [Google Scholar] [CrossRef]
- Srivastava, J.; Datta, S.; Mishra, S.P. Development of an efficient Agrobacterium- mediated transformation system for chickpeas (Cicer arietinum). Biologia 2017, 72, 153–160. [Google Scholar] [CrossRef]
- Yadav, S.K.; Katikala, S.; Yellisetty, V.; Kannepalle, A.; Narayana, J.L.; Maddi, V.; Mandapaka, M.; Shanker, A.K.; Bandi, V.; Bharadwaja, K.P. Optimization of Agrobacterium-mediated genetic transformation of cotyledonary node explants of Vigna radiata. Springer Plus 2012, 1, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rahman, Z.A.; Seman, Z.A.; Basirun, N.; Julkifle, A.L.; Zainal, Z.; Subramaniam, S. Preliminary investigations of Agrobacterium-mediated transformation in indica rice MR219 embryogenic callus using gusA gene. African Journal of Biotechnology 2011, 10, 7805–7813. [Google Scholar]
- Tripathi, L.; Singh, A.K.; Singh, S.; Singh, R.; Chaudhary, S.; Sanyal, I.; Amla, D.V. Optimization of regeneration and Agrobacterium-mediated transformation of immature cotyledons of chickpea (Cicer arietinum L.). Plant Cell, Tissue and Organ Culture (PCTOC) 2013, 113, 513–527. [Google Scholar] [CrossRef]
- Das, A.; Datta, S.; Thakur, S.; Shukla, A.; Ansari, J.; Sujayanand, G.K.; Chaturvedi, S.K.; Kumar, P.A.; Singh, N.P. Expression of a chimeric gene encoding insecticidal crystal protein Cry1Aabc of Bacillus thuringiensis in chickpea (Cicer arietinum L.) confers resistance to gram pod borer (Helicoverpa armigera Hubner.). Frontiers in Plant Science 2017, 8, 1423. [Google Scholar] [CrossRef]
- Das Bhowmik, S.S.; Cheng, A.Y.; Long, H.; Tan, G.Z.H.; Hoang, T.M.L.; Karbaschi, M.R.; Williams, B.; Higgins, T.J.V.; Mundree, S.G. Robust genetic transformation system to obtain non-chimeric transgenic chickpeas. Frontiers in Plant Science 2019, 10, 524. [Google Scholar] [CrossRef]
- Zhang, J.; Lin, M.; Chen, H.; Chen, X. Agrobacterium tumefaciens-mediated transformation of drumstick (Moringa oleifera Lam.). Biotechnology Biotechnological Equipment 2017, 31, 1126–1131. [Google Scholar] [CrossRef]
- Liu, S.J.; Wei, Z.M.; Huang, J.Q. The effect of co-cultivation and selection parameters on Agrobacterium-mediated transformation of Chinese soybean varieties. Plant Cell Reports 2008, 27, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Dutt, M.; Grosser, J.W. Evaluation of parameters affecting Agrobacterium-mediated transformation of citrus. Plant Cell, Tissue and Organ Culture (PCTOC) 2009, 98, 331–340. [Google Scholar] [CrossRef]
- Molina-Risco, M.; Ibarra, O.; Faion-Molina, M.; Kim, B.; Septiningsih, E.M.; Thomson, M.J. Optimizing Agrobacterium-mediated transformation and CRISPR-Cas9 gene editing in the tropical japonica rice variety presidio. International Journal of Molecular Sciences 2021, 22, 10909. [Google Scholar] [CrossRef]
- Benny, A.; Alex, S.; Soni, K.B.; Anith, K.N.; Kiran, A.G.; Viji, M.M. (2022). Improved transformation of Agrobacterium assisted by silver nanoparticles. BioTechnologia 2022, 103, 311. [Google Scholar] [CrossRef]
- Bruce, M.A.; Shoup Rupp, J.L. Agrobacterium-mediated transformation of Solanum tuberosum L.; potato. Transgenic Plants: Methods and Protocols 2019, 203–223. [Google Scholar]
- Gill, A.R.; Siwach, P. Agrobacterium rhizogenes mediated genetic transformation in Ficus religiosa L. and optimization of acetylcholinesterase inhibitory activity in hairy roots. South African Journal of Botany 2022, 151, 349–356. [Google Scholar] [CrossRef]
- Prem Kumar, G.; Sivakumar, S.; Siva, G.; Vigneswaran, M.; Senthil Kumar, T.; Jayabalan, N. Silver nitrate promotes high-frequency multiple-shoot regeneration in cotton (Gossypium hirsutum L.) by inhibiting ethylene production and phenolic secretion. In Vitro Cellular Developmental Biology-Plant 2016, 52, 408–418. [Google Scholar] [CrossRef]
- Arraes, F.B.M.; Beneventi, M.A.; Lisei de Sa, M.E.; Paixao, J.F.R.; Albuquerque, E.V.S.; Marin, S.R.R.; Purgatto, E.; Nepomuceno, A.L.; Grossi-de-Sa, M.F. Implications of ethylene biosynthesis and signaling in soybean drought stress tolerance. BMC Plant Biology 2015, 15, 1–20. [Google Scholar] [CrossRef]
- Namukwaya, B.; Magambo, B.; Arinaitwe, G.; Oweitu, C.; Erima, R.; Tarengera, D.; Bossa, D.L.; Karamura, G.; Muwonge, A.; Kubiriba, J.; Tushemereirwe, W. Antioxidants enhance banana embryogenic cell competence to Agrobacterium-mediated transformation. African Journal of Biotechnology 2019, 18, 735–743. [Google Scholar]
- Sainger, M.; Chaudhary, D.; Dahiya, S.; Jaiwal, R.; Jaiwal, P.K. Development of an efficient in vitro plant regeneration system amenable to Agrobacterium-mediated transformation of a recalcitrant grain legume black gram (Vigna mungo L. Hepper). Physiology and Molecular Biology of Plants 2015, 21, 505–517. [Google Scholar] [CrossRef] [PubMed]
- An, X.; Wang, B.; Liu, L.; Jiang, H.; Chen, J.; Ye, S.; Chen, L.; Guo, P.; Huang, X.; Peng, D. Agrobacterium-mediated genetic transformation and regeneration of transgenic plants using leaf midribs as explants in ramie [Boehmeria nivea (L. ) Gaud]. Molecular Biology Reports 2014, 41, 3257–3269. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, V.; Chaturvedi, A.K.; Mishra, A.; Jha, B. An efficient method of Agrobacterium-mediated genetic transformation and regeneration in local Indian cultivar of groundnut (Arachis hypogaea) using grafting. Applied Biochemistry and Biotechnology 2015, 175, 436–453. [Google Scholar] [CrossRef]
- Karthik, S.; Pavan, G.; Sathish, S.; Siva, R.; Kumar, P.S.; Manickavasagam, M. Genotype-independent and enhanced in planta Agrobacterium tumefaciens-mediated genetic transformation of peanut [Arachis hypogaea (L.)]. 3 Biotech 2018, 8, 1–15. [Google Scholar] [CrossRef]
- Arun, M.; Subramanyam, K.; Mariashibu, T.S.; Theboral, J.; Shivanandhan, G.; Manickavasagam, M.; Ganapathi, A. Application of sonication in combination with vacuum infiltration enhances the Agrobacterium-mediated genetic transformation in Indian soybean cultivars. Applied Biochemistry and Biotechnology 2015, 175, 2266–2287. [Google Scholar] [CrossRef]
- Mekala, G.K.; Juturu, V.N.; Mallikarjuna, G.; Kirti, P.B.; Yadav, S.K. Optimization of Agrobacterium-mediated genetic transformation of shoot tip explants of green gram (Vigna radiata (L. ) Wilczek). Plant Cell, Tissue and Organ Culture (PCTOC) 2016, 127, 651–663. [Google Scholar] [CrossRef]
- Yong, W.T.L.; Abdullah, J.O.; Mahmood, M. Optimization of Agrobacterium-mediated transformation parameters for Melastomataceae spp. using green fluorescent protein (GFP) as a reporter. Scientia Horticulturae 2006, 109, 78–85. [Google Scholar] [CrossRef]
- Zhao, Y.; Yang, D.; Liu, Y.; Han, F.; Li, Z. A highly efficient genetic transformation system for broccoli and subcellular localization. Frontiers in Plant Science 2023, 14, 1091588. [Google Scholar] [CrossRef]
- Kapildev, G.; Chinnathambi, A.; Sivanandhan, G.; Rajesh, M.; Vasudevan, V.; Mayavan, S.; Arun, M.; Jeyaraj, M.; Alharbi, S.A.; Selvaraj, N.; Ganapathi, A. High-efficient Agrobacterium-mediated in planta transformation in black gram (Vigna mungo (L. ) Hepper). Acta Physiologiae Plantarum 2016, 38, 1–13. [Google Scholar] [CrossRef]
- Li, Y.; Tang, D.; Liu, Z.; Chen, J.; Cheng, B.; Kumar, R.; Yer, H.; Li, Y. An improved procedure for Agrobacterium-mediated transformation of ‘Carrizo’Citrange. Plants 2022, 11, 1457. [Google Scholar] [CrossRef]
- Bett, B.; Gollasch, S.; Moore, A.; Harding, R.; Higgins, T.J. An improved transformation system for cowpea (Vigna unguiculata L. Walp) via sonication and a kanamycin-geneticin selection regime. Frontiers in Plant Science 2019, 10, 219. [Google Scholar] [CrossRef] [PubMed]
- Sabir, M.; Anwar, Y.; Khan, A.; Ali, M.; Yousuf, P.Y.; Al-Ghamdi, K. ChiC Gene Enhances Fungal Resistance in Indigenous Potato Variety (Diamant) via Agrobacterium-mediated Transformation. Biosciences Biotechnology Research Asia 2019, 16, 343–350. [Google Scholar] [CrossRef]
- Cody, J.P.; Maher, M.F.; Nasti, R.A.; Starker, C.G.; Chamness, J.C.; Voytas, D.F. Direct delivery and fast-treated Agrobacterium co-culture (Fast-TrACC) plant transformation methods for Nicotiana benthamiana. Nature Protocols 2023, 18, 81–107. [Google Scholar] [CrossRef]
- Sgamma, T.; Thomas, B.; Muleo, R. Ethylene inhibitor silver nitrate enhances regeneration and genetic transformation of Prunus avium (L. ) cv Stella. Plant Cell, Tissue and Organ Culture (PCTOC) 2015, 120, 79–88. [Google Scholar] [CrossRef]
- Sivanandhan, G.; Moon, J.; Sung, C.; Bae, S.; Yang, Z.H.; Jeong, S.Y.; Choi, S.R.; Kim, S.G.; Lim, Y.P. L-Cysteine increases the transformation efficiency of Chinese cabbage (Brassica rapa ssp. pekinensis). Frontiers in Plant Science 2021, 12, 767140. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.F.; Yu, X.Q.; Zhou, Z.; Ma, W.J.; Tang, G.X. A high-efficiency Agrobacterium tumefaciens mediated transformation system using cotyledonary nodes as explants in soybean (Glycine max L. ). Acta Physiologiae Plantarum 2016, 38, 1–10. [Google Scholar] [CrossRef]
- Guo, Y.; Yin, Q.; Kong, H.; Huang, Q.; Zuo, J.; He, L.; Guo, A. Effect of silver nitrate on Agrobacterium-mediated genetic transformation of cassava. Biotechnology Bulletin 2013, 166–170. [Google Scholar]
- Lee, H.; Zhang, Z.J. Agrobacterium-mediated transformation of maize (Zea mays) immature embryos. Cereal Genomics: Methods and Protocols 2014, 273–280. [Google Scholar]
- Lee, J.; Naing, A.H.; Park, K.I.; Kim, C.K. Silver nitrate reduces hyperhydricity in shoots regenerated from the hypocotyl of snapdragon cv. Maryland Apple Blossom. Scientia Horticulturae 2023, 308, 111593. [Google Scholar] [CrossRef]
- Kala, R.G.; Abraham, V.; Sobha, S.; Jayasree, P.K.; Suni, A.M.; Thulaseedharan, A. Agrobacterium-mediated genetic transformation and somatic embryogenesis from leaf callus of Hevea brasiliensis: effect of silver nitrate. In Prospects in Bioscience: Addressing the Issues. Springer India2013, 303–315.
- Hassan, M.F.; Islam, S.S. Effect of silver nitrate and growth regulators to enhance anther culture response in wheat (Triticum aestivum L.). Heliyon 2021, 7.
- Sangra, A.; Shahin, L.; Dhir, S.K. Long-term maintainable somatic embryogenesis system in alfalfa (Medicago sativa) using leaf explants: embryogenic sustainability approach. Plants 2019, 8, 278. [Google Scholar] [CrossRef]
- Gupta, S.; Gupta, S.; Bhat, V.; Gupta, M.G. Somatic embryogenesis and Agrobacterium-mediated genetic transformation in Indian accessions of lucerne (Medicago sativa L.). Indian Journal of Biotechnology 2006, 5, 269–275. [Google Scholar]
- Basak, S.; Parajulee, D.; Brearley, T.; Dhir, S. High-efficiency Somatic Embryogenesis and Plant Regeneration in Alfalfa (Medicago sativa L.). Society for In Vitro Biology (SIVB), Virginia, USA, 10-14 June 2023, P-2034.
- Parajulee, D.; Basak, S.; Dhir, S. (2023) Somatic Embryogenesis and Plant Regeneration from in vitro grown leaf explants from Alfalfa (Medicago sativa L.). American Society of Plant Biologist, Savannah, Georgia, USA, 5-9 August 2023, P-80039.
- Xia, P.; Hu, W.; Liang, T.; Yang, D.; Liang, Z. An attempt to establish an Agrobacterium-mediated transient expression system in medicinal plants. Protoplasma 2020, 257, 1497–1505. [Google Scholar] [CrossRef] [PubMed]
- Ratanasut, K.; Rod-In, W.; Sujipuli, K. In planta Agrobacterium-mediated transformation of rice. Rice Science 2017, 24, 181–186. [Google Scholar] [CrossRef]
- Heidari Japelaghi, R.; Haddad, R.; Valizadeh, M.; Dorani Uliaie, E.; Jalali Javaran, M. High-efficiency Agrobacterium-mediated transformation of tobacco (Nicotiana tabacum). Journal of Plant Molecular Breeding 2018, 6, 38–50. [Google Scholar]
- Pinthong, R.; Sujipuli, K.; Ratanasut, K. Agroinfiltration for transient gene expression in floral tissues of Dendrobium sonia ‘Earsakul’. Journal of Agricultural Technology 2014, 10, 459–465. [Google Scholar]
- Duncan, D.B. Multiple range and multiple F tests. Biometrics 1995, 11, 1–42. [Google Scholar] [CrossRef]
Figure 1.
Different developmental stages of callus induction: (A) Approximately 2 mm excised leaf segments were cultured on callus induction medium; (B) 2-week-old leaf-derived callus; (C) 3-week-old leaf-derived callus.
Figure 1.
Different developmental stages of callus induction: (A) Approximately 2 mm excised leaf segments were cultured on callus induction medium; (B) 2-week-old leaf-derived callus; (C) 3-week-old leaf-derived callus.
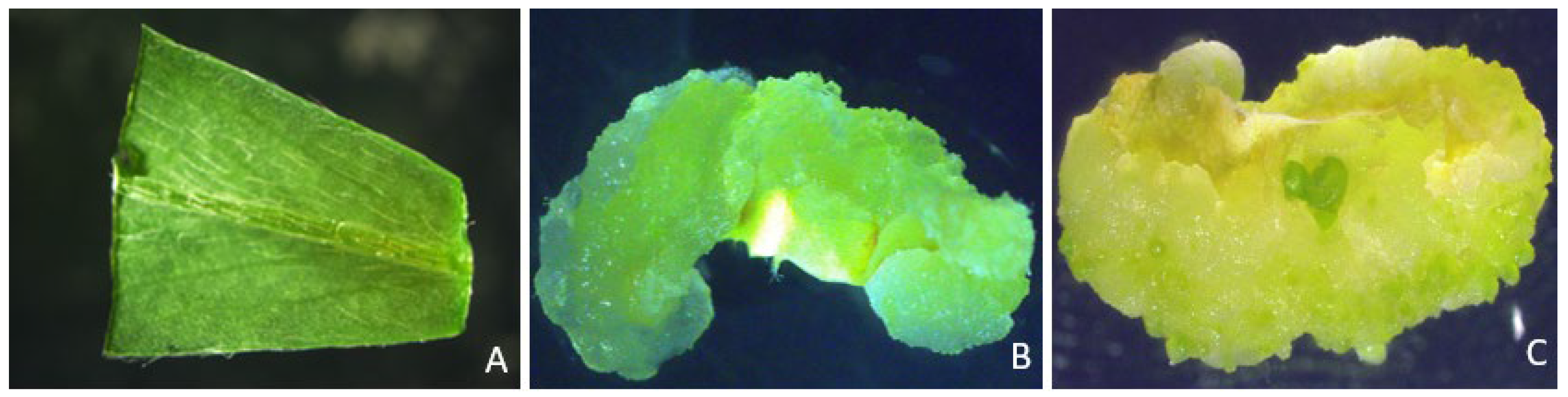
Figure 2.
Transient GUS gene expression in different explants of alfalfa: (A) GUS gene expression in segmented leaf (2 mm), as indicated by dark blue spots; (B) high GUS gene expression observed in the midrib region of the larger section of a 4 mm leaf explant (data not shown); (C) magnified view (6x) of a single blue spot of GUS expression with multiple cells in the midrib area of the explant, as shown in panel B; (D) pattern of dark-to-light blue within single or multiple cells, indicating the distribution of GUS-positive cells in the whole leaf; (E) concentrated GUS expression in the axillary bud and nodal region in the stem, showing dark spots with high GUS expression; (F) high GUS gene expression observed in petioles; (G) high GUS gene expression observed in roots; (H) scattered GUS gene expression in 3-week-old leaf-derived callus. Arrows indicate specific areas of GUS gene expression in the various explants and leaf-derived calli.
Figure 2.
Transient GUS gene expression in different explants of alfalfa: (A) GUS gene expression in segmented leaf (2 mm), as indicated by dark blue spots; (B) high GUS gene expression observed in the midrib region of the larger section of a 4 mm leaf explant (data not shown); (C) magnified view (6x) of a single blue spot of GUS expression with multiple cells in the midrib area of the explant, as shown in panel B; (D) pattern of dark-to-light blue within single or multiple cells, indicating the distribution of GUS-positive cells in the whole leaf; (E) concentrated GUS expression in the axillary bud and nodal region in the stem, showing dark spots with high GUS expression; (F) high GUS gene expression observed in petioles; (G) high GUS gene expression observed in roots; (H) scattered GUS gene expression in 3-week-old leaf-derived callus. Arrows indicate specific areas of GUS gene expression in the various explants and leaf-derived calli.
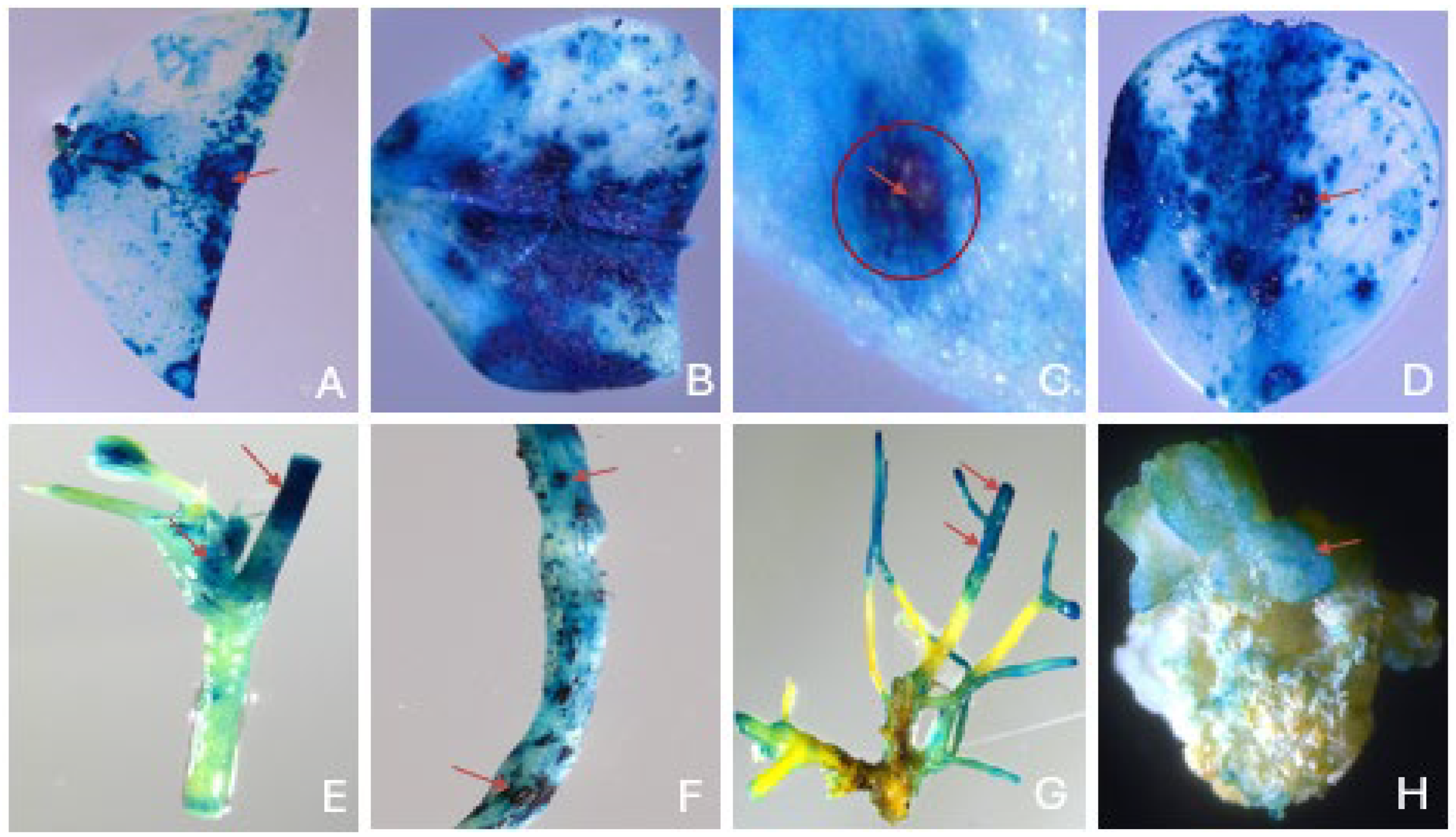
Figure 3.
Optimization of transient GUS gene expression in alfalfa: Percent transient GUS gene expression response relative to the non-treated control alfalfa under varying rates of different parameters. (A) Effects of different explants on transient GUS gene expression. (B) Effect of age of leaf on transient GUS gene expression. (C) Effects of different sizes of leaf explants on transient GUS gene expression. All experiments were repeated four times. Results were pooled over experimental runs. Vertical bars represent the standard errors ±5 (P < 0.0001) of the means (n = 64).
Figure 3.
Optimization of transient GUS gene expression in alfalfa: Percent transient GUS gene expression response relative to the non-treated control alfalfa under varying rates of different parameters. (A) Effects of different explants on transient GUS gene expression. (B) Effect of age of leaf on transient GUS gene expression. (C) Effects of different sizes of leaf explants on transient GUS gene expression. All experiments were repeated four times. Results were pooled over experimental runs. Vertical bars represent the standard errors ±5 (P < 0.0001) of the means (n = 64).
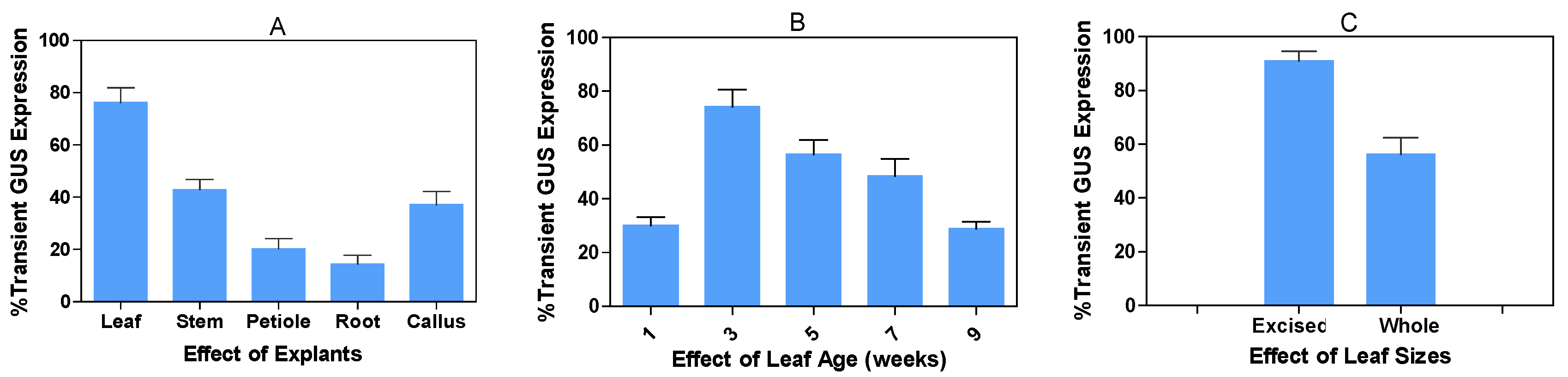
Figure 4.
Optimization of transient GUS gene expression in alfalfa: Percent transient GUS gene expression response relative to the non-treated control alfalfa under the varying effects of leaf tissue wounding on transient GUS gene expression. All experiments were repeated four times. Results were pooled over experimental runs. Vertical bars represent the standard errors ±5 (P < 0.0001) of the means (n = 64). .
Figure 4.
Optimization of transient GUS gene expression in alfalfa: Percent transient GUS gene expression response relative to the non-treated control alfalfa under the varying effects of leaf tissue wounding on transient GUS gene expression. All experiments were repeated four times. Results were pooled over experimental runs. Vertical bars represent the standard errors ±5 (P < 0.0001) of the means (n = 64). .
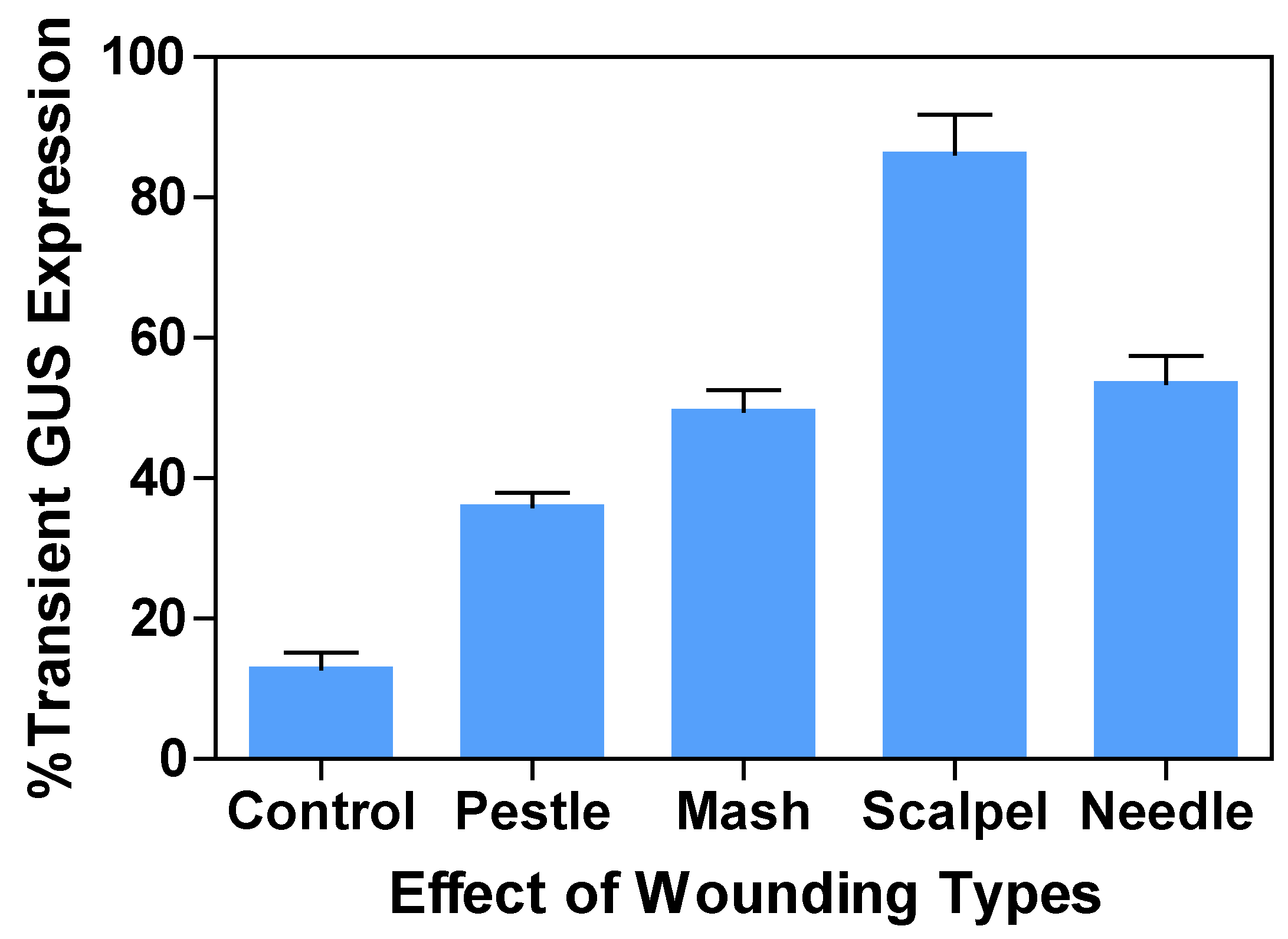
Figure 5.
Optimization of transient GUS gene expression in alfalfa: Percent transient GUS gene expression response relative to the non-treated control alfalfa under varying effects of Agrobacterium concentration on transient GUS gene expression. All experiments were repeated four times. Results were pooled over experimental runs. Vertical bars represent the standard errors ±5 (P < 0.0001) of the means (n = 64).
Figure 5.
Optimization of transient GUS gene expression in alfalfa: Percent transient GUS gene expression response relative to the non-treated control alfalfa under varying effects of Agrobacterium concentration on transient GUS gene expression. All experiments were repeated four times. Results were pooled over experimental runs. Vertical bars represent the standard errors ±5 (P < 0.0001) of the means (n = 64).
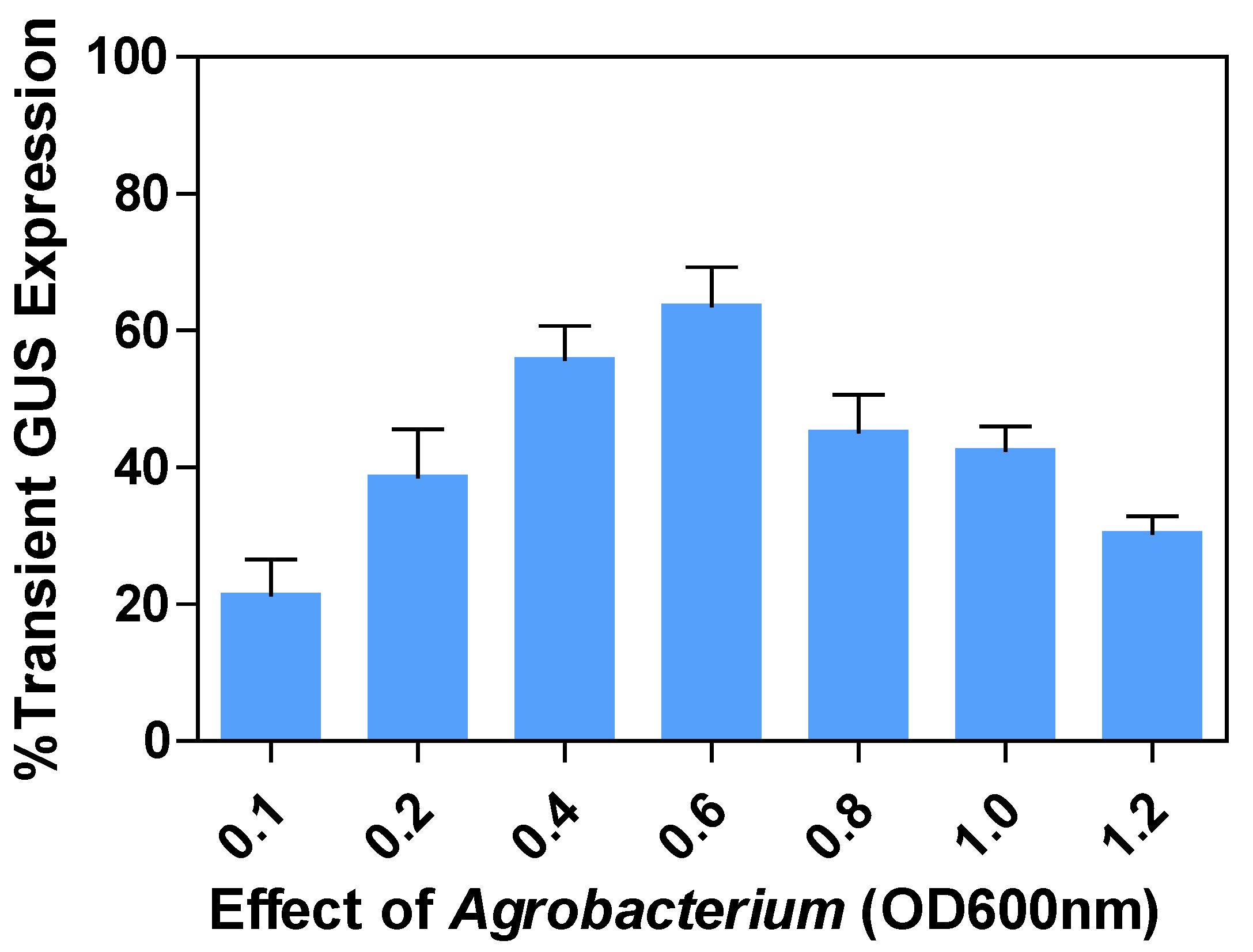
Figure 6.
Optimization of transient GUS gene expression in alfalfa: Percent transient GUS gene expression response is relative to the non-treated control alfalfa under the varying effects of preculture of leaf tissues on transient GUS gene expression. All experiments were repeated four times. Results were pooled over experimental runs. Vertical bars represent the standard errors ±5 (P < 0.0001) of the means (n = 64).
Figure 6.
Optimization of transient GUS gene expression in alfalfa: Percent transient GUS gene expression response is relative to the non-treated control alfalfa under the varying effects of preculture of leaf tissues on transient GUS gene expression. All experiments were repeated four times. Results were pooled over experimental runs. Vertical bars represent the standard errors ±5 (P < 0.0001) of the means (n = 64).
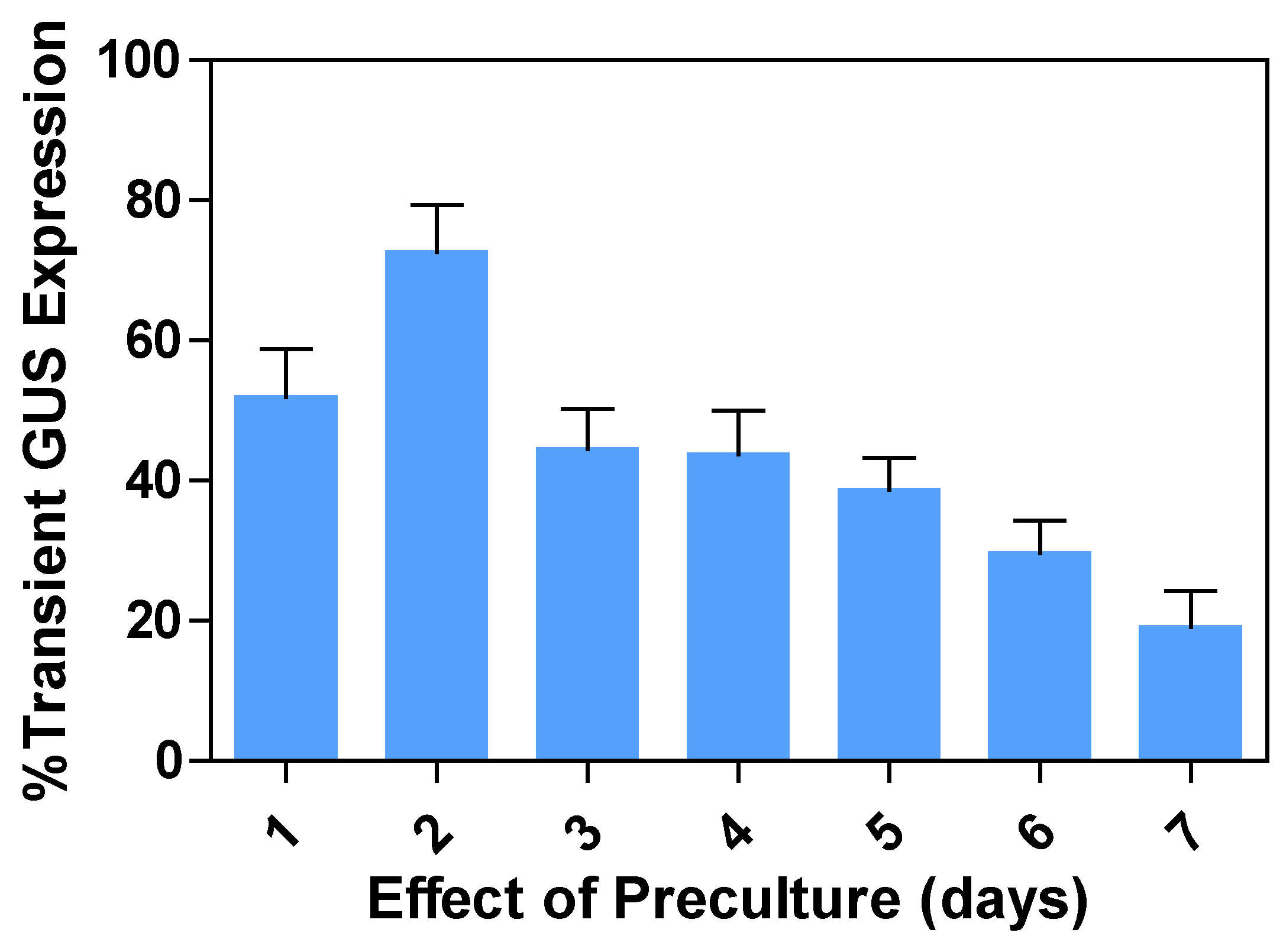
Figure 7.
Optimization of transient GUS gene expression in alfalfa: Percent transient GUS gene expression response relative to the non-treated control alfalfa under varying effects of incubation period on transient GUS gene expression. All experiments were repeated four times. Results were pooled over experimental runs. Vertical bars represent the standard errors ±5 (P < 0.0001) of the means (n = 64).
Figure 7.
Optimization of transient GUS gene expression in alfalfa: Percent transient GUS gene expression response relative to the non-treated control alfalfa under varying effects of incubation period on transient GUS gene expression. All experiments were repeated four times. Results were pooled over experimental runs. Vertical bars represent the standard errors ±5 (P < 0.0001) of the means (n = 64).
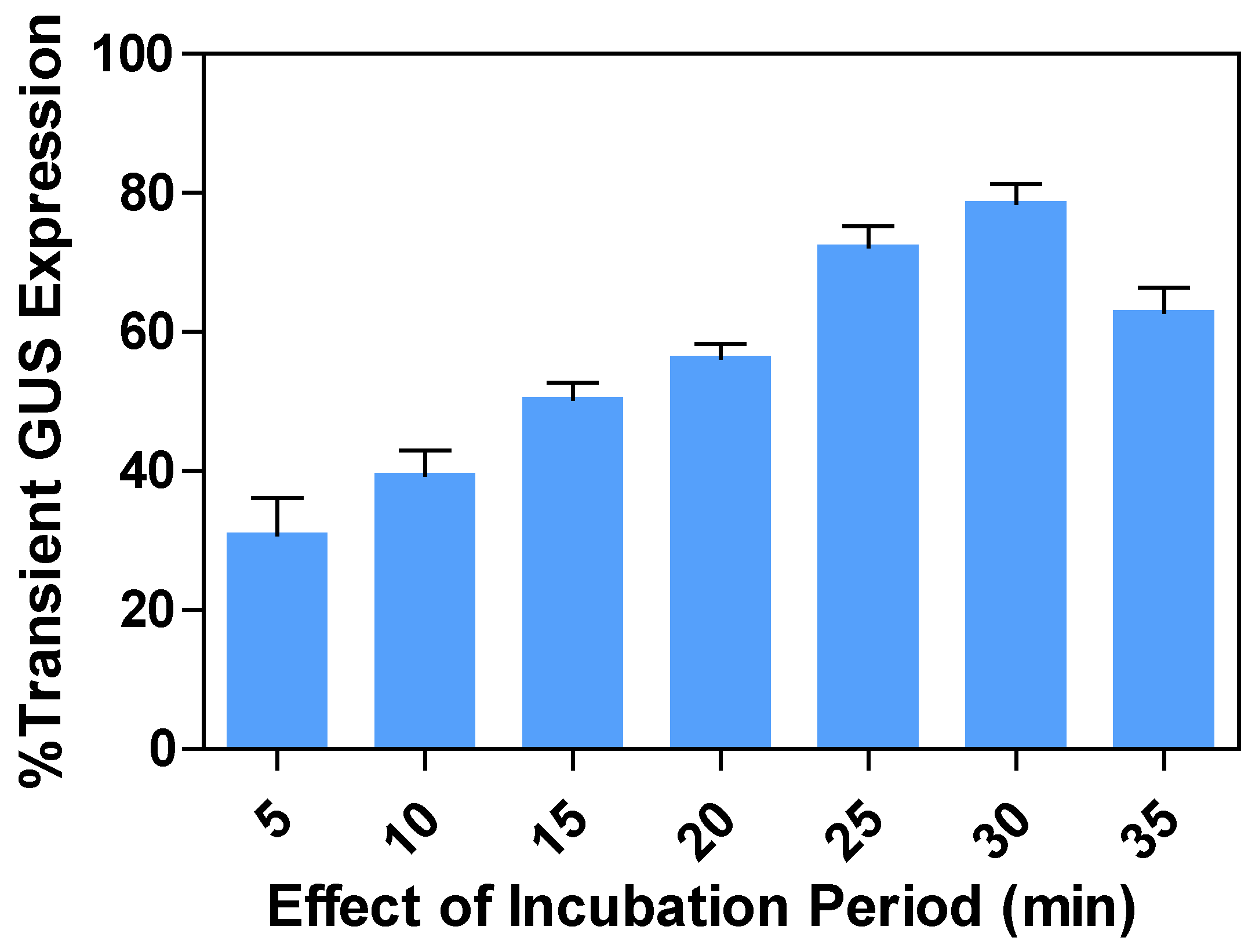
Figure 8.
Optimization of transient GUS gene expression in alfalfa: Percent transient GUS gene expression response is relative to the non-treated control alfalfa under the varying effects of the co-cultivation period on transient GUS gene expression. All experiments were repeated four times. Results were pooled over experimental runs. Vertical bars represent the standard errors ±5 (P < 0.0001) of the means (n = 64).
Figure 8.
Optimization of transient GUS gene expression in alfalfa: Percent transient GUS gene expression response is relative to the non-treated control alfalfa under the varying effects of the co-cultivation period on transient GUS gene expression. All experiments were repeated four times. Results were pooled over experimental runs. Vertical bars represent the standard errors ±5 (P < 0.0001) of the means (n = 64).
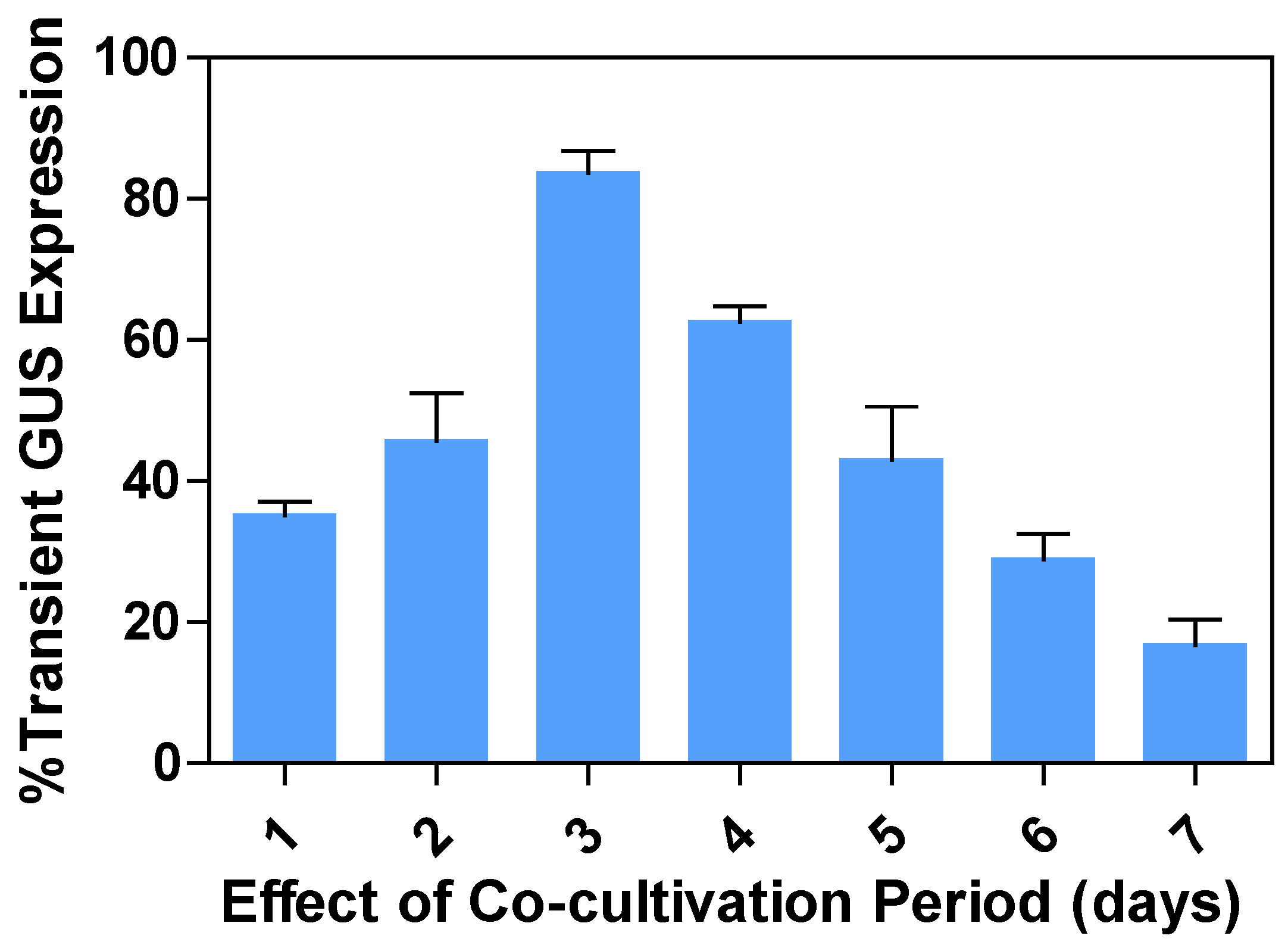
Figure 9.
Optimization of transient GUS gene expression in alfalfa: Percent transient GUS gene expression response is relative to the non-treated control alfalfa under the varying effects of acetosyringone on transient GUS gene expression. All experiments were repeated four times. Results were pooled over experimental runs. Vertical bars represent the standard errors ±5 (P < 0.0001) of the means (n = 64).
Figure 9.
Optimization of transient GUS gene expression in alfalfa: Percent transient GUS gene expression response is relative to the non-treated control alfalfa under the varying effects of acetosyringone on transient GUS gene expression. All experiments were repeated four times. Results were pooled over experimental runs. Vertical bars represent the standard errors ±5 (P < 0.0001) of the means (n = 64).
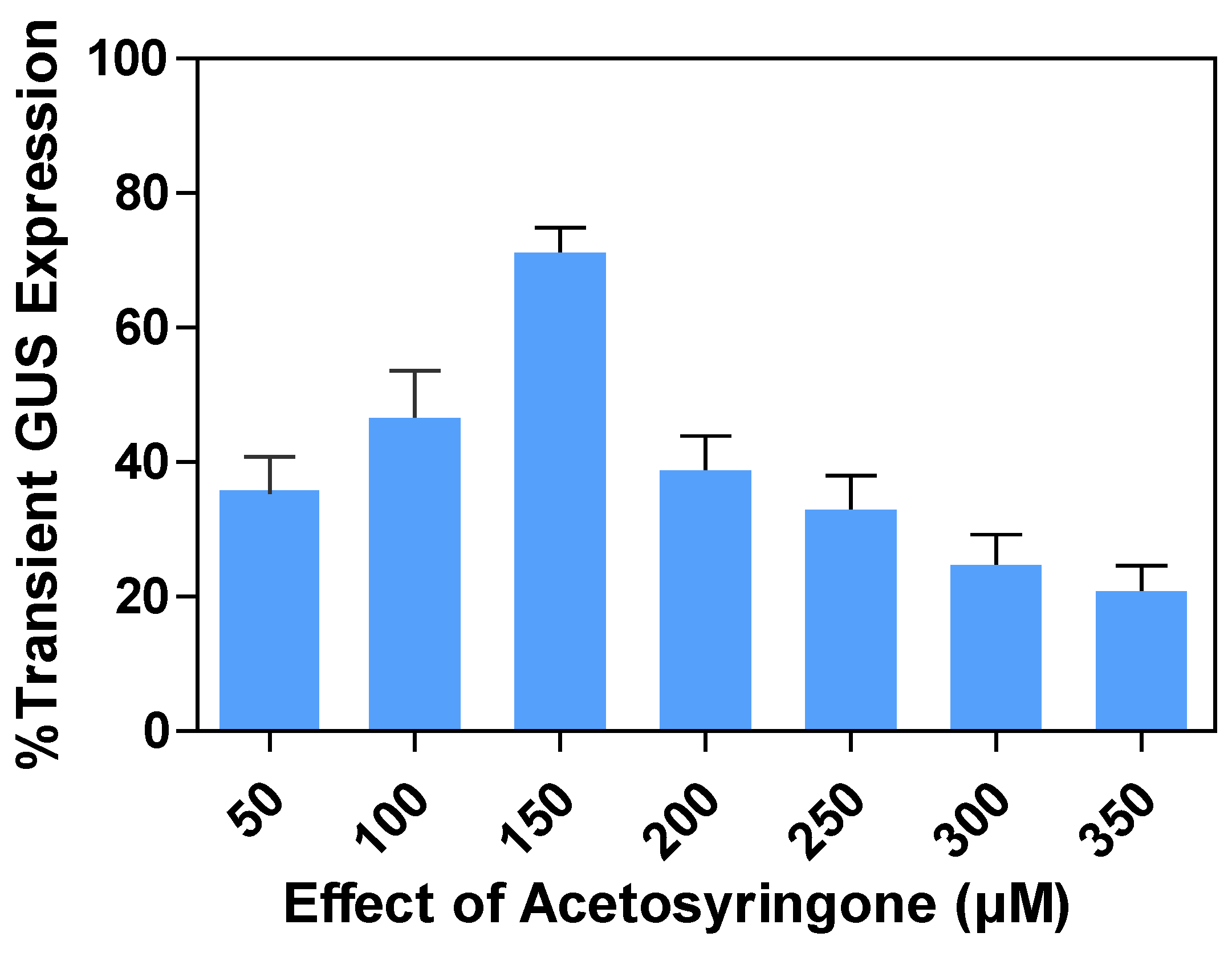
Figure 10.
Optimization of transient GUS gene expression in alfalfa: Percent transient GUS gene expression response relative to the non-treated control alfalfa under the varying effects of calcium chloride on transient GUS gene expression. All experiments were repeated four times. Results were pooled over experimental runs. Vertical bars represent the standard errors ±5 (P < 0.0001) of the means (n = 64).
Figure 10.
Optimization of transient GUS gene expression in alfalfa: Percent transient GUS gene expression response relative to the non-treated control alfalfa under the varying effects of calcium chloride on transient GUS gene expression. All experiments were repeated four times. Results were pooled over experimental runs. Vertical bars represent the standard errors ±5 (P < 0.0001) of the means (n = 64).
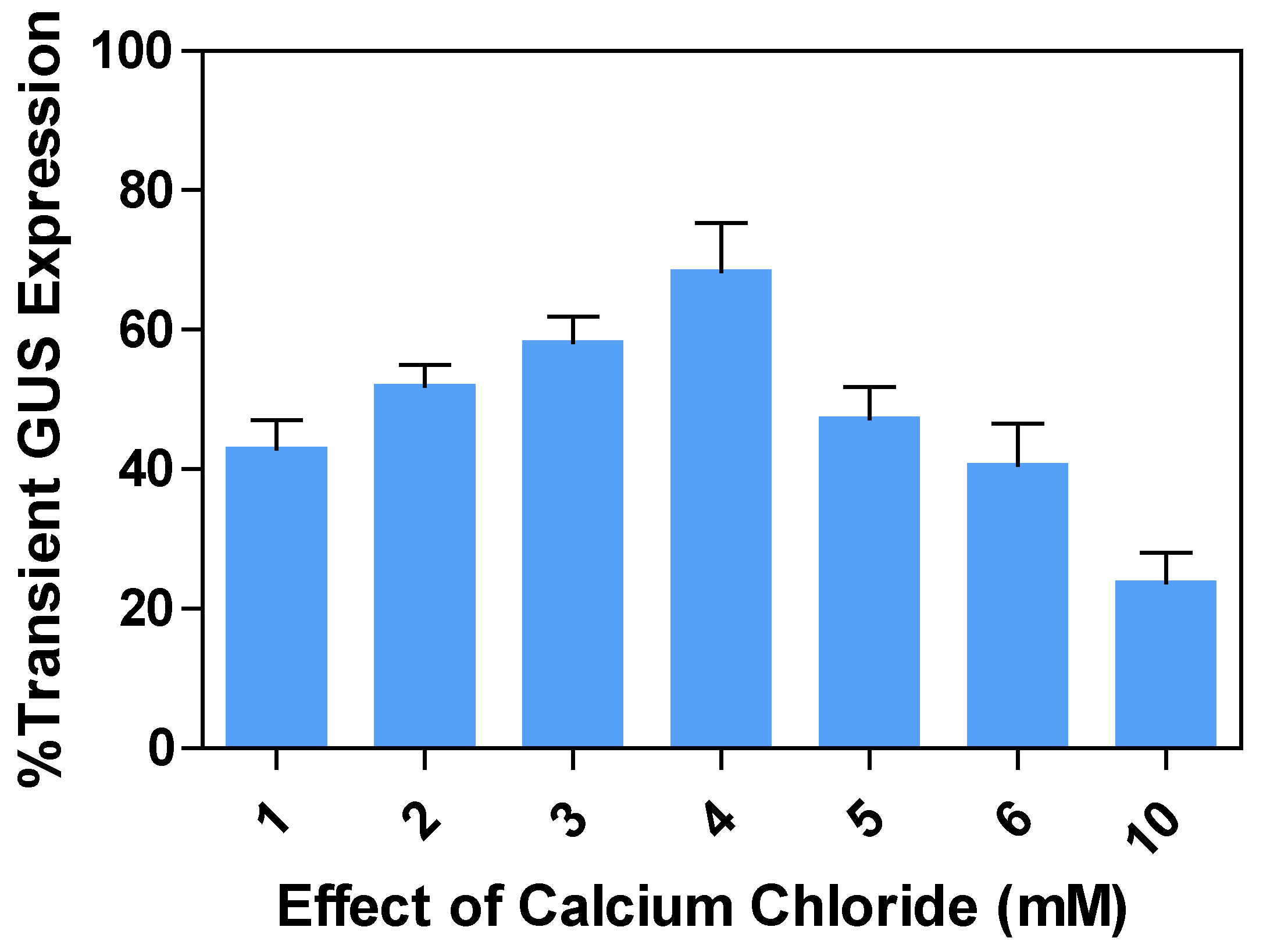
Figure 11.
Optimization of transient GUS gene expression in alfalfa: Percent transient GUS gene expression response relative to the non-treated control alfalfa under the varying effects of silver nitrate on transient GUS gene expression. All experiments were repeated four times. Results were pooled over experimental runs. Vertical bars represent the standard errors ±5 (P < 0.0001) of the means (n = 64).
Figure 11.
Optimization of transient GUS gene expression in alfalfa: Percent transient GUS gene expression response relative to the non-treated control alfalfa under the varying effects of silver nitrate on transient GUS gene expression. All experiments were repeated four times. Results were pooled over experimental runs. Vertical bars represent the standard errors ±5 (P < 0.0001) of the means (n = 64).
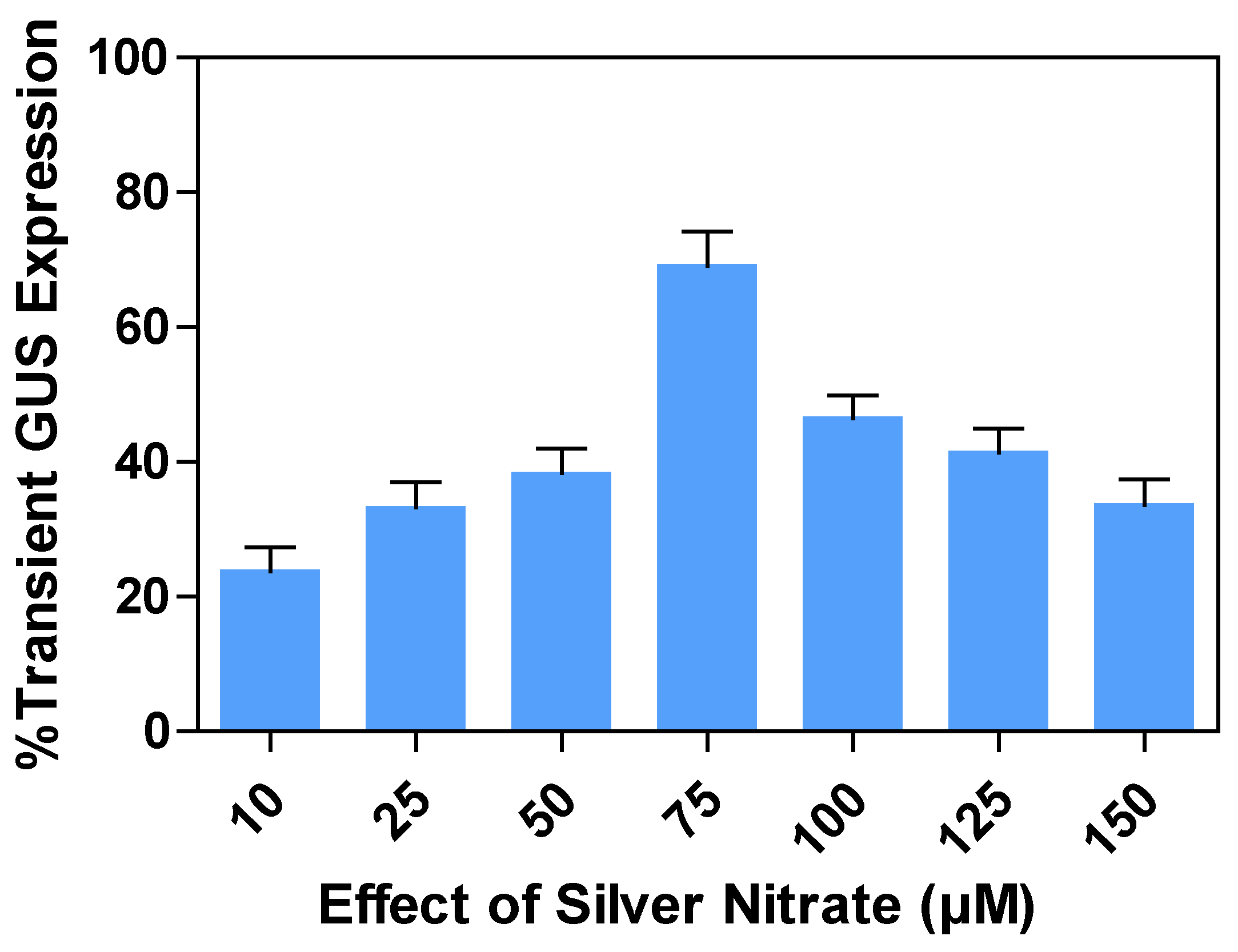
Figure 12.
GFP fluorescence was observed in alfalfa leaf explants (A-F) and embryonic calli (G-L) after co-cultivation for 3 days with Agrobacterium at an OD600nm of 0.6, followed by two washes with MS washing media. Images were captured using an Olympus SZX12 Stereo Fluorescence Microscope with a GFP filter (Olympus America Inc., Melville, NY, USA). Arrows indicate GFP gene expression. [A (chloroplast autofluorescence), B (GFP fluorescence)]: Fluorescence image of clustered multiple cells in 3-week-old wounded leaves; [C (chloroplast autofluorescence), D (GFP fluorescence)]: Isolated spots of GFP on excised leaf explants; [E (chloroplast autofluorescence), F (GFP fluorescence)]: GFP gene expression in the wounded area of the leaf explants; [G (chloroplast autofluorescence), H (GFP fluorescence)]: GFP transient gene expression in 1-week-old pro-embryonic callus derived from leaves; [I (chloroplast autofluorescence), J (GFP fluorescence)]: GFP transient gene expression in 2-week-old pro-embryonic callus derived from leaves; [K (chloroplast autofluorescence), L (GFP fluorescence)]: GFP transient gene expression in 3-week-old pro-embryonic callus derived from leaves. GFP expression was particularly notable in the globular embryos from a 3-week-old leaf-derived callus grown under non-selection conditions.
Figure 12.
GFP fluorescence was observed in alfalfa leaf explants (A-F) and embryonic calli (G-L) after co-cultivation for 3 days with Agrobacterium at an OD600nm of 0.6, followed by two washes with MS washing media. Images were captured using an Olympus SZX12 Stereo Fluorescence Microscope with a GFP filter (Olympus America Inc., Melville, NY, USA). Arrows indicate GFP gene expression. [A (chloroplast autofluorescence), B (GFP fluorescence)]: Fluorescence image of clustered multiple cells in 3-week-old wounded leaves; [C (chloroplast autofluorescence), D (GFP fluorescence)]: Isolated spots of GFP on excised leaf explants; [E (chloroplast autofluorescence), F (GFP fluorescence)]: GFP gene expression in the wounded area of the leaf explants; [G (chloroplast autofluorescence), H (GFP fluorescence)]: GFP transient gene expression in 1-week-old pro-embryonic callus derived from leaves; [I (chloroplast autofluorescence), J (GFP fluorescence)]: GFP transient gene expression in 2-week-old pro-embryonic callus derived from leaves; [K (chloroplast autofluorescence), L (GFP fluorescence)]: GFP transient gene expression in 3-week-old pro-embryonic callus derived from leaves. GFP expression was particularly notable in the globular embryos from a 3-week-old leaf-derived callus grown under non-selection conditions.
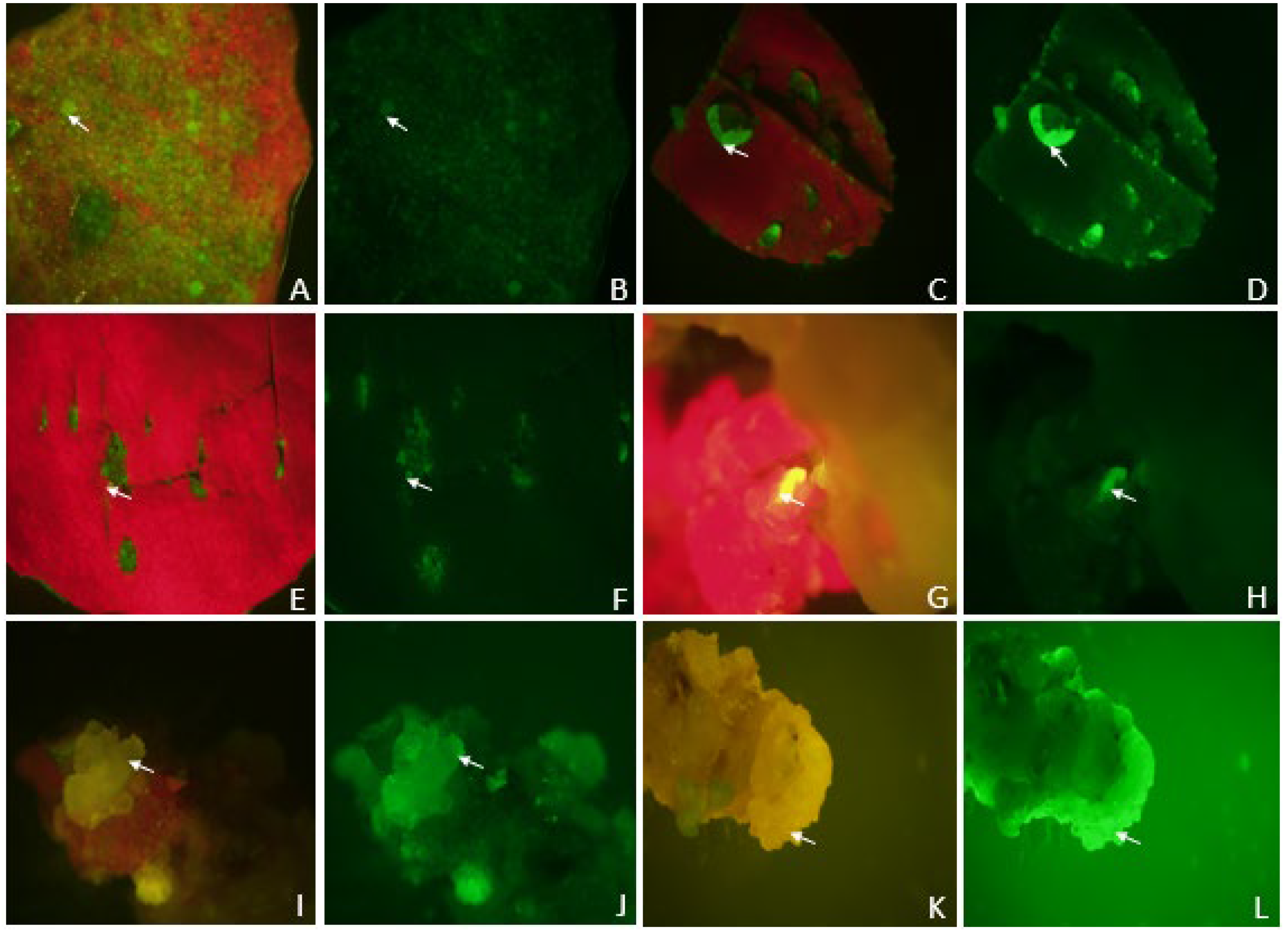
Figure 13.
Step-by-step protocol for transient expression in Medicago sativa (alfalfa).
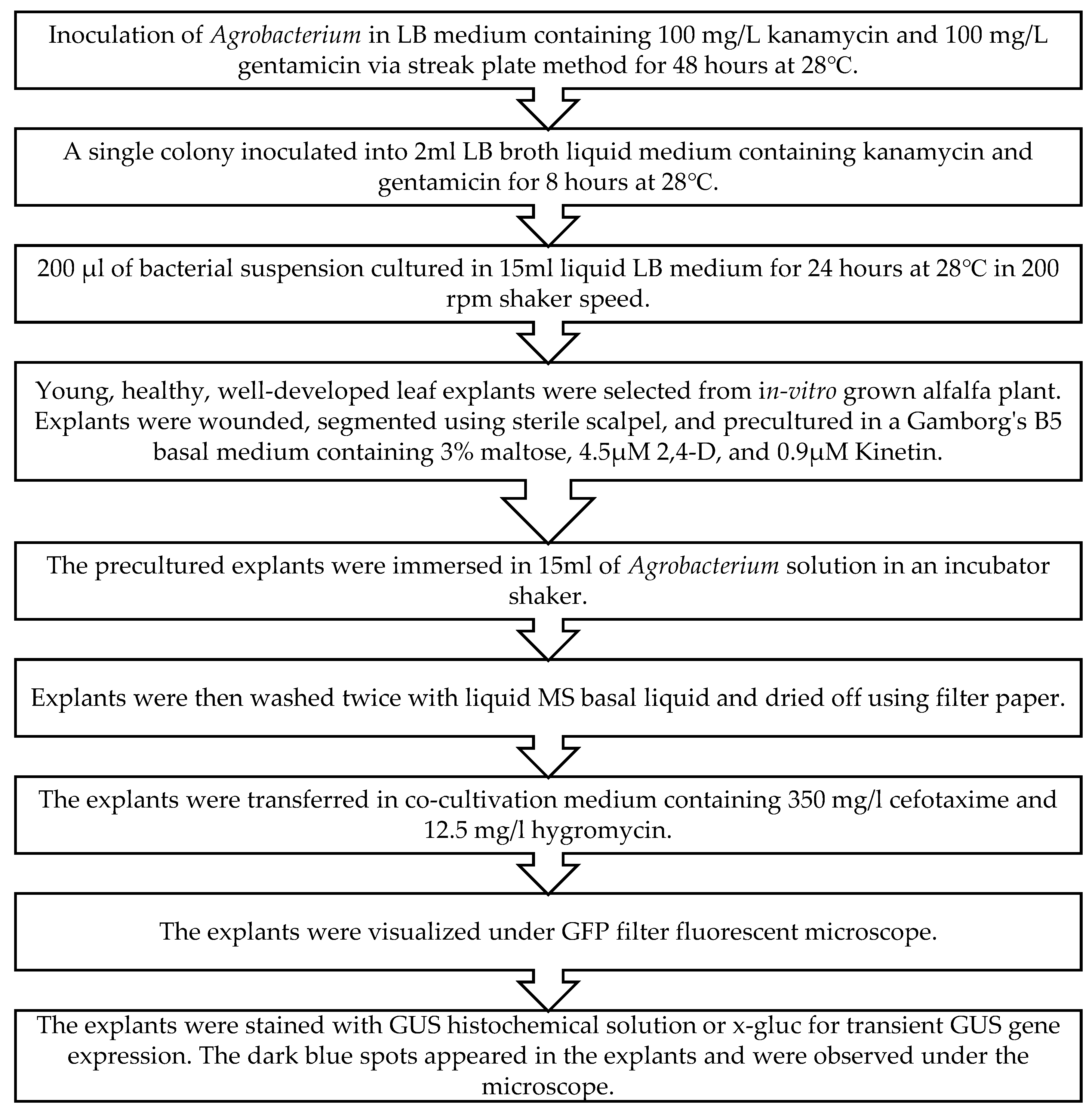
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



