
Submitted:
29 August 2024
Posted:
29 August 2024
You are already at the latest version
Abstract
Genetic transformation is a critical tool for gene manipulation and functional analyses in plants, enabling the exploration of key phenotypes and agronomic traits at the genetic level. While dicotyledonous plants offer various tissues for in vitro culture and transformation, monocotyledonous plants, such as rice, have limited options. This study presents an efficient method for genetically transforming rice (Oryza sativa L.) using seed-derived embryogenic calli as explants. Two target genes were utilized to assess regeneration efficiency: green fluorescent protein (eGFP) and the apple FLOWERING LOCUS T (FT)-like gene (MdFT1). Antisense MdFT1 was cloned into a vector controlled by the rice α-amylase 3D (Ramy3D) promoter, while eGFP was fused to Cas9 under the Ubi promoter. These vectors were introduced separately into rice embryogenic calli from two Korean cultivars using Agrobacterium-mediated transformation. Transgenic seedlings were successfully regenerated via hygromycin selection using an in vitro cultivation system. PCR confirmed stable transgene integration in the transgenic calli and their progeny. Fluorescence microscopy revealed eGFP expression, and antisense MdFT1-expressing lines exhibited notable phenotypic changes, including variations in plant height and grain quality. This study demonstrated the effective use of seed-derived embryogenic calli for rice transformation, offering a promising approach for developing transgenic plants in monocot species.
Keywords:
genetic transformation
; Agrobacterium-mediated transformation
; embryogenic callus
; transgenic rice
; eGFP reporter gene
; MdFT1 gene
; heterologous expression
1. Introduction
Rice (Oryza sativa L.) is one of the most important crops worldwide, serving as a staple food for over half of the world’s population, especially in Asia, where it has great cultural significance. Its rich nutritional content, particularly as a key source of carbohydrates, makes it essential in the daily diet of billions [1,2,3]. Economically, rice farming supports millions of people’s livelihoods and plays a crucial role in both local and global markets. Its adaptability to diverse environments, from floodplains to terraced hills, makes it a resilient crop vital for food security, especially in regions prone to climate variability. Advances in rice cultivation, including high-yield and disease-resistant varieties, have further enhanced its importance by helping to meet global food demands and alleviate poverty in rural communities.
Genetic transformation is a fundamental process in biotechnology and genetic engineering, in which the genetic material of an organism is altered by introducing foreign DNA into its genome. This process is facilitated by various methods, such as bacterial transformation, electroporation, microinjection, polyethylene glycol (PEG)-mediated transformation, biolistics (e.g., particle bombardment, gene gun), and Agrobacterium-mediated transformation, each suited to different types of cells and organisms. Moreover, vectors such as plasmids are commonly used to transport foreign DNA into host cells [4,5,6,7]. This technique is crucial for a wide range of applications, from studying gene function to developing genetically modified crops for agriculture and gene therapies in medicine. Genetic transformation is a powerful tool that enables the precise manipulation of genetic material, driving advances in understanding life at the molecular level and offering innovative solutions in various fields.
Agrobacterium-mediated transformation is a process in which Agrobacterium tumefaciens (or Agrobacterium rhizogenes) naturally introduces foreign DNA into plant cells using Ti (tumor-inducing) or Ri (root-inducing) plasmids [8,9,10]. When the bacterium attaches to a wound site in a plant, it activates virulence genes that process and transfer a specific DNA segment, known as T-DNA, from the plasmid to the plant cell. The T-DNA is then integrated into the plant genome, where it expresses foreign genes and confers new traits to the plant [4,11,12,13,14]. This method, which is widely used in genetic engineering, allows the stable incorporation of desired genes into plant genomes, making it a powerful tool for creating genetically modified plants [15]. Agrobacterium-mediated transformation is a highly efficient and widely used method for introducing foreign DNA into plant cells. It offers advantages such as targeted gene integration, low transgene copy number, stable and heritable transformations, and minimal tissue damage compared to other methods such as gene guns or electroporation [16,17,18].
Agrobacterium-mediated transformation has been successfully employed in various plant species, including tobacco, rice, maize, tomato, and soybean [12,20,21,22]. Numerous studies have demonstrated the effectiveness of this method for generating transgenic plants of these species. Tobacco is one of the species most frequently transformed using this technique, leading to various genetic modifications and trait improvements [19,20,21]. Additionally, Agrobacterium has been used for rice transformation [22,23,24,25,26], providing a platform for the development of genetically modified varieties with enhanced agronomic traits [27,28]. Similar successes have been reported in maize, where Agrobacterium-mediated methods have facilitated the introduction of new traits [29,30]. Tomatoes, soybeans, and potatoes are significant examples of species transformed using this approach, highlighting the versatility and wide applicability of Agrobacterium-mediated transformation in plant biotechnology [29,31,32,33,34].
Rice, as well as other monocot plants, face challenges associated with genetic transformation, where the use of explants is more limited compared to dicots. In addition, previous methods for transforming rice relied on complex culture media, resulting in time-consuming preparation and increased costs. Therefore, by presenting a simplified procedure using embryogenic calli as explants, this study aimed to streamline the process of obtaining transgenic rice, making it more efficient and cost-effective. We developed an efficient genetic transformation method using Agrobacterium to introduce the foreign genes eGFP and MdFT1 into the rice genome. Finally, transgenic rice plants were established via an in vitro culture system using embryogenic calli as explants. Furthermore, we evaluated the gene expression on the phenotype (agronomic trait) in subsequent generations of transgenic rice plants heterologous expressed apple MdFT1 gene. This genetic transformation system offers crucial technical tools for functional gene analysis, uncovering molecular mechanisms in key rice traits, and extends the potential for application to other crops.
2. Results
2.1. Binary Expression Vector for Rice Transformation
To obtain genetic material for rice transformation, two genes of interest (GOI), eGFP and MdFT1, were cloned into two binary expression vectors driven by different promoter systems. The eGFP gene was fused to Cas9 under the control of the rice ubiquitin promoter (Ubi) with the NOS as the terminator, resulting in the recombinant expression vector Ubi::Cas9–eGFP (Figure 1a). The MdFT1 gene was cloned into the binary vector in an antisense orientation through homologous recombination, driven by the rice alpha-amylase (Ramy3D) promoter with the 3’-UTR as the terminator creating the Ramy3D::MdFT1 expression vector (Figure 1b). Both binary expression vectors used the same selection marker gene, HygR, which confers hygromycin B resistance. These binary expression vectors were separately transformed into A. tumefaciens EHA105, which was then used for embryogenic callus transformation.
2.2. Establishment of Transgenic Rice using Agrobacterium‒Mediated Transformation via In Vitro Culture System
Using an in vitro tissue culture system, numerous transgenic lines expressing Ramy3D::MdFT1 and Ubi::Cas9–eGFP were obtained (Figure 2). After in vitro culture in callus induction media (N6CI), seed‒derived embryogenic calli were used as explants for genetic transformation using Agrobacterium‒mediated transformation (Figure 2a‒c). Following Agrobacterium infection and co-culture, the infected rice calli were grown on a selective medium containing hygromycin B (N6SE). Hygromycin-resistant calli began to differentiate with the appearance of new callus after 3–4 weeks. In contrast, false or non-transformed calli did not grow on N6SE, turned brown, and gradually died (Figure 2d). Putative transgenic hygromycin-resistant lines were screened and continuously propagated on selection medium (N6SE) (Figure 2e, f). Subcultures were conducted every 3 weeks. For shoot differentiation, transgenic rice calli were transferred to the shooting medium (MSS). Meristem shoots were initiated from the calli after 4–6 weeks (Figure 2g, h) and quickly developed into mature shoots with leaves (Figure 2 i, j). Roots began to appear within 1 week of culture and elongated rapidly during root differentiation. Whole seedlings with roots were obtained on the rooting medium (MSR) (Figure 2k). After 4–6 weeks of cultured on rooting medium, whole seedlings, including leaves and full roots, were transplanted to the soil and grown in a glass house (Figure 2l). Subsequently, we obtained transgenic rice plants via an in vitro culture system from seed-derived embryogenic calli as the initial materials to whole seedlings with roots.
Putative transgenic hygromycin-resistant lines were continuously propagated on N6SE via subculturing. The calli of the putative transgenic hygromycin-resistant lines were sampled for screening using gDNA PCR (Figure 3). To screen transgenic callus lines with the introduced Ram3D::MdFT1 vector, a specific primer set for MdFT1 amplification (set 3, Table 1), which amplified a 525-bp fragment (Figure 3a), was used. To screen transgenic callus lines expressing Ubi::Cas9‒eGFP, a specific primer set for eGFP amplification (set 2, Table 1) was used to amplify a 717-bp fragment (Figure 3b). Additionally, the presence of HygR selection marker gene in both Ram3D::MdFT1 and Ubi::Cas9‒eGFP was confirmed using a pair of primers (set 1, Table 1), which amplified the 563-bp fragment (Figure 3a, b). Moreover, RT-PCR analysis of MdFT1 mRNA in transgenic calli showed that MdFT1 was transcribed in rice cells (Figure 3c).
2.3. Stable and Inherited Expression of Transgenes in Transgenic Plants
Stable and inherited expression of transgenes in transgenic plants is crucial for the successful establishment of genetically modified crops. To determine whether the target genes introduced into the rice genome could be stably transmitted to the offspring, gDNA PCR amplification was performed. T1 generation plants were generated from seeds harvested from T0 transgenic plants, which were germinated in soil and cultivated in a glasshouse (Figure 4a, b). Leaves of T1 transgenic plants were sampled for gDNA PCR. For molecular confirmation, 20 lines were sampled from the progeny of Ram3D::MdFT1 and Ubi::Cas9‒eGFP plants. Successful integration of the T-DNA region into the rice genome was assessed based on the presence of target genes (MdTFL1 and eGFP) and a selection marker gene (HygR). Following the same procedure as that used for the PCR screening of transgenic callus lines, transgenes in the T1 progeny were detected in Ram3D::MdFT1 (Figure 4c) and Ubi::Cas9‒eGFP (Figure 4d) plants. This indicated that the transgene was consistently inherited and remained stable across generations. The transgenic plants were grown for seed harvesting and further analysis.
2.4. Transgenic Rice Expressed Ubi::Cas9–eGFP
To evaluate whether the target gene eGFP was functionally expressed in transgenic rice, the green fluorescence of the eGFP protein was assessed using microscopic imaging. Fluorescence was observed in both transgenic calli and T1 plants, including in different organs (leaves, stems, and roots) (Figure 5). Six callus lines of transgenic Ubi::Cas9‒eGFP and wild type ‘Samkwang’ were selected for eGFP fluorescence observation. Green fluorescence was observed under the excitation spectra in the calli of the transgenic rice using an in vivo imaging system (Figure 5a, Figure S1b). GFP fluorescence was not observed in WT callus lines (Figure 5a, Figure S1a). As expected, a non-fluorescent transition signal was observed in the WT seedlings (Figure 5b–d, Figure S2). Similar to the callus lines, eGFP fluorescence was visualized in different organs, including the roots, stems, and leaves of the transgenic seedlings. Green fluorescence of the eGFP protein was observed in all tissues of the Ubi::Cas9‒eGFP plants (Figure 5b–d, Figure S3). Moreover, eGFP protein expression was visualized at the cellular scale in transgenic calli (Figure 5a, right panel) and in different organs of T1 seedlings (Figure 5b-d, right panel) using a confocal microscope. Supporting the gDNA PCR data of the transgenic calli (Figure 3) and transgenic T1 plants (Figure 4), this confirms that the eGFP gene, after integration into the rice genome, was functionally expressed and inherited by the progeny.
2.5. Heterologous Expression of the MdFT1 Gene Resulted in Overall Changes in Agronomic Traits of Transgenic Rice
Upon cultivation, transgenic Ramy3D::MdFT1 rice exhibited distinct phenotypic changes, including branching, leaf angle, internode length, plant height, and grain productivity. Phenotyping of agronomic traits was conducted to assess morphological changes in comparison with the WT ‘Dongjin’ cultivar. Introduction of the antisense MdFT1 gene was aimed at delaying flowering time (heading date, HD) in transgenic rice by suppressing its function. The HD of the transgenic Ramy3D::MdFT1 and WT plants were counted from the sowing date until the emergence of the first panicle (Figure 6a). Transgenic Ramy3D::MdFT1 rice showed a slight delay in HD compared to the WT, with an average HD of 65.6 d in WT and 67.1 d in Ramy3D::MdFT1, although there were no significant differences (Figure 6b).
Furthermore, transgenic Ramy3D::MdFT1 rice exhibited greater total height and internode length than the WT rice (Figure 7a‒c). This may be because the introduction of MdFT1 affects cell development. To investigate this, cells in the elongation zone of the internode stem were examined under a light microscope, and their sizes were measured to identify differences (Figure 7d‒g). Cell size differed between the transgenic and WT plants. Although the cell widths were similar, the cell length of the transgenic plants was significantly greater than that of the WT.
In addition to the overall phenotypic description above, another important agronomic trait that determines yield potential is seed productive ability. Seed production is affected by the number of grains in every panicle and the weight and size of the grains. In this study, transgenic rice introducing the apple MdFT1 gene showed a lower-setting seed with a significantly smaller number of grains per panicle (Figure 8a, b). However, the grain size of the transgenic Ramy3D::MdFT1 rice was larger than that of the WT. Although grain size did not show significant differences in width between transgenic and WT rice, the grain length of the transgenic rice was significantly greater (Figure 8c–f). This led to a significant increase in the length-to-width ratio and weight of the rice grains (Figure 8g, h). In addition to assessing the appearance of rice grains, we quantified their nutrient quality by analyzing their free amino acid content. Analyzing the free amino acid content of rice grains provides a deeper understanding of their nutritional quality, as amino acids are the building blocks of proteins and play vital roles in various biological processes. It allows for a more accurate understanding of the rice’s health benefits, flavor profile, and suitability for various culinary applications. High free amino acid content, especially in essential amino acids, enhances the overall nutritional quality, making the rice more valuable from both a health and economic perspective. Rice grains from the WT and transgenic lines were analyzed using an automatic amino acid analyzer, with two replicates for each line (Table S1‒4). The grain rice of the Ramy3D::MdFT1 transgenic line was significantly higher than that of the WT for all the analyzed free amino acids (Table 2).
3. Discussion
The basic classical method of Agrobacterium-mediated transformation to obtain transgenic rice has been described previously [22,24]. However, it requires an expensive medium with many different ingredients and takes a long time to prepare. Specifically, this process requires the preparation of separate and complex media for plant tissue transformation. This study demonstrated that liquid N6CI medium could be utilized throughout the entire Agrobacterium-mediated plant tissue transformation process. In addition, Agrobacterium culture is processed by spreading it on solid LB media, making it easy to collect and directly use for plant tissue infection. In contrast, various studies have grown Agrobacterium in YEP liquid media, which require a centrifugation step to remove the culture medium before it can be used for plant transformation. Thus, we simplified the transformation method for embryonic rice calli, reducing time-consuming preparation and costs.
Transgenic rice plants were generated using seed-derived embryogenic calli in conjunction with Agrobacterium-mediated transformation. This approach has proven to be a robust and reliable method for introducing and achieving stable expression of foreign genes in rice and has traditionally been a challenging task because of the monocotyledonous nature of the plant. The use of seed-derived embryogenic calli as starting explant material is particularly advantageous. Embryogenic calli are known for their high regenerative capacity, which makes them ideal candidates for transformation [35,36,37]. Unlike other explant sources that may be less responsive or more prone to somaclonal variation, seed-derived embryogenic calli can provide a more uniform and consistent response to transformation and regeneration protocols, which is crucial for achieving high transformation efficiency and the subsequent regeneration of healthy, viable, transgenic plants [38]. As a potential explant, transgenic maize has been obtained from embryogenic calli using particle and microprojectile bombardment transformation methods [39,40].
The Agrobacterium-mediated transformation method used in the present study further enhanced the efficiency of gene transfer. A. tumefaciens is widely recognized for its ability to transfer foreign DNA into plant cells. This method has several advantages over other transformation techniques such as biolistics (gene gun), including lower cost, and higher transformation efficiency [24]. Herein, the use of Agrobacterium allowed for the successful introduction of binary expression vectors containing the eGFP and MdFT1 genes (Figure 1) into the rice genome, resulting in the generation of hygromycin-resistant calli (Figure 2) that could be further regenerated into transgenic plants (Figure 3 and Figure 4). Two Korean rice cultivars, ‘Dongjin’ and ‘Samkwang,’ were utilized as explant materials for Agrobacterium-mediated transformation of two target genes under the control of two different promoter systems. The eGFP gene, driven by the rice ubiquitin promoter (Ubi) in the CRISPR/Cas9 vector, is effective for gene-editing in transgenic rice [41,42,43,44]. For genetic transformation in rice, the rice Ubi promoter was more effective than the widely used constitutive promoter CaMV 35S. Using Agrobacterium-mediated transformation of two Australian rice cultivars ‘Jarrah’ and ‘Amaroo’, Narayana et al. (2000) found that the Ubi promoter produced 30-fold higher GUS activity than the CaMV35S promoter in transgenic plants [45]. The apple MdFT1 gene was expressed under the control of the rice α-amylase 3D promoter (Ramy3D), a strong promoter commonly used for producing recombinant proteins in transgenic rice callus suspension cultures [46,47,48,49].
Next, the integration and stable expression of the target genes eGFP and MdFT1 in the transgenic plants were investigated. The eGFP gene served as a reporter, allowing easy visualization of transgene expression through fluorescence microscopy. The clear detection of GFP fluorescence in the transformed calli and various tissues of the transgenic plants provided strong evidence of successful gene integration and expression (Figure 5). This not only validated the effectiveness of the transformation protocol but also demonstrated the functionality of the introduced genes within the rice genome. Moreover, the MdFT1 gene, introduced in an antisense orientation, was specifically designed to modulate key agronomic traits, such as flowering time and plant height. The successful expression of this gene, as confirmed by RT-PCR (Figure 3c) and the observed phenotypic changes (Figure 6, Figure 7 and Figure 8), indicated that the genetic modifications were not only stably inherited but also biologically active in influencing plant development [20,28]. The ability to manipulate such traits through targeted genetic engineering holds significant promise for crop improvement as it allows for the fine-tuning of characteristics that are critical for yield and adaptability.
In this study, we present a clear step-by-step guide, that not only provides a protocol on plant tissues in vitro culture systems but also on how to establish transgenic plants for the functional study of GOI in the field of genetic engineering. We meticulously documented the entire plant transformation process, starting from the cultivation of seed-derived embryogenic calli to the cloning of target genes into binary expression vectors and the generation of fully transformed plants. This was followed by comprehensive genetic and phenotypic analyses of the transgenic plants to confirm the success of the transformation.
Overall, we developed an efficient Agrobacterium-mediated genetic transformation method to obtain transgenic rice plants. The expression of target genes in transgenic rice has been shown to be stable and heritable. The development of genetic transformation systems offers critical tools for functional analysis of GOI, exploring molecular mechanisms behind key traits, and creating genetically modified crops with enhanced traits.
4. Materials and Methods
4.1. Gene Cloning and Construction of Plant Transformation Binary Vectors
First-strand cDNA of apple MdFT1 was synthesized through RT-PCR using mRNA isolated from the follower buds of ‘Fuji’ apple cultivar. To facilitate cloning, a pair of specific PCR primers (primer set 3, Table 1) was designed to amplify the MdFT1 gene, incorporating KpnI and XbaI restriction enzyme sites at the 5′ end of the forward and reverse primers, respectively. PCR products were analyzed on 1.0% agarose gels, and the target band of these genes was recovered from gels and purified using a MEGAquick-spin™ plus fragment DNA purification kit according to the manufacturer’s instructions (Cat. No: 17290; iNtRON Biotechonology, Seongnam, Korea). The fragment was then ligated into a pGEM®-T Easy vector (Cat. No: A1360; Promega, Madison, WI, USA) using T4 DNA ligase (Cat. No: 2011B; Takara, Kusatsu, Japan) and transformed into Escherichia coli DH5α competent cells (Cat. No: 9057; Takara, Kusatsu, Japan). Subsequently, the plasmid DNA was sequenced (Macrogen, Seoul, Korea). After sequencing, the cloned sequence of MdFT1 libraries was blasted/aligned to the original sequence obtained from the NCBI database (GenBank Accession No. AB161112.1) using CLC Genomics Workbench 12 (Qiagen, Hilden, Germany). MdFT1 fragment was released from the pGEM®-T Easy vector plasmid by digesting with KpnI–XbaI restriction enzymes. The MdFT1 gene was then ligated into the binary vector pMYD317 [50], which was digested with the same restriction enzymes (XbaI–KpnI). XbaI was located downstream of the Ramy3D promoter, and KpnI upstream of the 3’ UTR. The MdFT1 gene was cloned into the binary vector in an antisense orientation by homologous recombination, resulting in the recombinant expression vector Ramy3D::MdFT1.
For vector construction of the eGFP reporter gene, the eGFP gene was cloned into pRGEB32 (Plasmid #63142; Addgene, Watertown, MA, USA) by GenScript Biotech (Piscataway, NJ, USA). eGFP was fused downstream of Cas9, driven by the rice ubiquitin promoter (Ubi), with NOS as the terminator, and designated as the Ubi::Cas9–eGFP expression vector.
These binary expression vectors (Ramy3D::MdFT1 and Ubi::Cas9–eGFP) were transformed into E. coli DH5α competent cells to produce a large amount of plasmid DNA vectors. Plasmid DNA vectors were isolated from transformed E. coli DH5α using a fast DNA-spin™ plasmid DNA purification kit (Cat.No: 17013; iNtRON Biotechonology, Seongnam, Korea). The presence of the ligated target genes MdFT1 and eGFP in the plasmid DNA of the binary expression vectors was confirmed by enzyme digestion using KpnI–BamHI and XbaI–KpnI, respectively.
4.2. Agrobacterium Transformation
Each constructed expression vector, Ramy3D::MdFT1 and Ubi::Cas9–eGFP were transformed into A. tumefaciens EHA105 competent cells using the freeze-thaw method [51]. After transformation, A. tumefaciens EHA105 cells were thoroughly spread on LB agar plates (BD Difco, Ref 244520, NJ, USA) containing 50 μg/mL kanamycin and 100 μg/mL rifampicin. They were cultured in a sharking incubator at 180 rpm, 28°C in the dark for 2 d. Single colonies were then picked from the selective plates, inoculated in 5 mL liquid LB broth medium (BD Difco, Ref 244620, NJ, USA) supplemented with antibiotics, and incubated at 28°C, 180 rpm, overnight in the dark. Plasmid DNA vectors in transformed A. tumefaciens EHA105 cells were isolated and confirmed by PCR and enzymatic digestion. The transformed Agrobacteria were then mixed with 30% glycerol and stored in a freezer as stock at –80°C.
4.3. Culture Media and Plant Materials
Culture media were prepared using the assigned components in specific amounts and concentrations, as presented in Table 3. The pH was adjusted to the desired value prior to autoclaving. Media were autoclaved at 121°C for 10 min, then cooled down to approximately 60–65°C, followed by adding hormones, antibiotics, and acetosyringone. Note that some components were added at this temperature to prevent the loss of activity due to autoclaving. The media were mixed well, poured onto culture dishes and bottles, and allowed to solidify and dry on a clean bench.
Mature seeds of rice O. sativa L. from two Korean rice cultivars, ‘Dongjin’ and ‘Samkwang,’ were utilized for in vitro culture. Rice seeds were dehusked to remove the seed cover, and 10 mL of the dehusked seeds were collected in a 50 mL falcon tube for surface sterilization. Briefly, rice seeds were prewashed in 40 mL of sterile water by soaking for 2 min, washed with 70% ethanol for 5 min, and rinsed with a 2.5% sodium hypochlorite (NaOCl) solution containing 0.01% Tween20 (P9416; Sigma Aldrich, Munich, Germany) for 10 min. They were finally rinsed 3–5 times with sterile water to remove any residual disinfectants. During the sterilization process, 40 mL of the disinfectant was used, and the falcon tube was shaken frequently to increase the sterilization efficiency. Sterilized seeds were blotted on sterile 3M Whatman filter paper before being transferred to the culture medium. Rice seeds were cultivated in vitro in various media with different components at specific cultivation stages, as listed in Table 3. Cultivation was conducted in a culture room under controlled conditions: 25°C, 100 μM.m−2.s−1 light intensity, and a 16-h light/8-h dark photoperiod.
4.4. Rice Embryogenic Callus Induction and Transformation
To prepare the plant explants for genetic transformation, sterilized rice seeds, which were obtained as described in Section 4.3, were cultivated on callus induction medium (N6CI) with 10–15 seeds per N6CI plate. After 15 d of culture on N6CI medium, the rice scutellum-derived calli were cut from the in vitro germinated seeds using a sterile scalpel and tweezers. The detached embryogenic calli were placed in fresh N6CI and cultured for 5 d before being used for genetic transformation.
The Agrobacterium-mediated transformation method was utilized to deliver the binary vectors Ramy3D::MdFT1 and Ubi::Cas9–eGFP into embryogenic rice calli, modified from a previous study [52]. Briefly, 10 µL of transformed Agrobacterium stock, prepared as described in Section 4.2, was inoculated in 5 mL liquid LB broth medium with 50 μg/mL kanamycin and 100 μg/mL rifampicin and incubated overnight. The next day, 100 µL of overnight cultured agrobacterial cells were thoroughly spread on the LB agar-supplemented antibiotics plate using a cell spreader and incubated overnight at 28°C in the dark. Subsequently, 2–3 mL of N6CI liquid medium was added to the plate, agrobacterial cells were harvested using a cell scraper, and the cell suspension was collected in a 50 mL falcon tube using a pipette. Harvested cells were diluted in N6CI liquid medium, adjusted to a final OD 600 of 0.5, and then supplemented with 200 µM of acetosyringone.
For genetic transformation, embryogenic calli induced from rice O. sativa L. ‘Dongjin’ and ‘Samkwang’ cultivars were used as explants for the transformation of the binary expression vectors Ramy3D::MdFT1 and Ubi::Cas9–eGFP, respectively. Rice embryogenic calli (40–60 calli from four plates) were prepared as described above and immersed in 40 mL of cell suspension for Agrobacterium infection. This transformation was conducted in a 50 mL falcon tube for 10 min with gentle shaking to improve agrobacterial cell infection. After Agrobacterium infection, the infected rice embryogenic calli were blotted on multiple layers of sterile 3M Whatman filter paper before being transferred to N6CO medium (Table 3) and co-cultured in the dark for 3 d. After co-cultivation, rice calli were prewashed several times with sterile water to remove calli overcovered with Agrobacterium and then rinsed once with 500 mg/L cefotaxime in sterile water. Rice calli were then blotted on sterile 3M Whatman filter paper for drying, then placed on selection medium (N6SE). Transgenic hygromycin-resistant calli developed after 3 weeks under antibiotic selection conditions. The cultivation conditions were the same as those described in Section 4.3.
4.5. Transgenic Plants In Vitro Cultivation
Putative transgenic hygromycin-resistant lines were propagated on N6SE via subculturing. To obtain transgenic plants, calli were transferred to MSS (Table 3) for shoot induction. Mature shoots were then separated and transferred to MSR for rooting. Cultivation was conducted in vitro in a culture room as described in Section 4.3.
Whole seedlings with roots were established and prepared for ex vitro culture (i.e., sown in soil). Before transplantation, culture bottles were gradually opened for 1 d until the cap of the culture bottle was completely opened. Rice seedlings were removed from the culture bottles, rinsed under running tap water to eliminate residual medium, and the upper leaves were trimmed. The roots were wrapped in Kimwipes and placed in 50 mL Falcon tubes filled with water for 2 d of acclimatization. Seedlings were transplanted into the commercially purchased garden soil in growth pots (depth: 24 cm × diameter: 12 cm) and grown in a glass house with the following cultivation conditions: temperature of 32/25°C (day/night), 60% humidity, and natural light. Rice plants were cultivated in 2021 (for the T0 generation) and 2022 (for the T1 generation) during warm periods from late April to early October, representing the rice-producing season in South Korea.
4.6. PCR Analysis
To select transgenic lines, PCR analysis was employed to identify the transgene in the genomic DNA of the transgenic plants. We conducted two rounds of PCR at two different stages of plant generation: the callus and seedling stages. In the first round, putative transgenic lines obtained from hygromycin-resistant selection media were sampled for genomic DNA (gDNA) isolation. Leaves of T1 plants in the vegetative stage (before panicle initiation) were sampled for the second round of gDNA PCR.
The calli and leaves of the transgenic lines were collected in 2.0 mL Eppendorf tubes containing two stainless steel beads (5 mm), pre-frozen in liquid nitrogen, and pulverized using a TissueLyser II (Qiagen, Hilden, Germany). Homogenized tissues were processed for gDNA extraction using a DNeasy® Plant Mini Kit (Cat. No: 69204; Qiagen, Hilden, Germany). PCR was conducted at a total reaction volume of 20 μL, including 1 µL of gDNA, 1 µL of each forward and reverse primer, and 17 µL of DNase-free sterile water using the Maxime™ PCR PreMix (i-StarTaq) (Cat. No: 25167; Intron, Seongnam, Republic of Korea). Integration of the T-DNA region (harboring the target genes) into the rice genome was investigated using specific primer sets (Table 1). To detect the selective marker gene HypR, as well as reporter genes eGFP, and MdFT1, PCR was conducted with primer sets 1, 2, and 3, respectively. PCR was processed as follows: initial denaturation step of 95°C for 5 min; followed by 32 cycles at 95°C for 45 s, 58°C for 45 s, and 72°C for 1 min; and a final extension step at 72°C for 5 min. The PCR products of each target gene were separated on 1.0% agarose gels, and the amplicon length was visualized using the Image LabTM program (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
4.7. RNA Isolation, cDNA Synthesis, and Quantification of Transgene Expression
To quantify the transcript expression of the MdFT1 gene in transgenic rice, callus tissues were used for RNA isolation. Total RNA extraction and cDNA synthesis were performed as previously described [20]. A real-time reverse transcription–polymerase chain reaction (RT‒PCR) assay was used to quantify MdFT1 mRNA expression level. The transcript expression of MdFT1 in transgenic callus tissues was normalized to that of the rice reference gene OsAct1 (Accession No. KC140129.1). RT‒PCR was carried out in 20 μL of mixture reaction containing 1 µL of cDNA, 1 µL of each forward and reverse primer, and distilled water using the Maxime™ PCR PreMix using running conditions as described in a previous study [28]. The primers used for RT-PCR of MdFT1 and OsAct1 are listed in Table 1 as primer sets 3 and 4, respectively. Amplicons were separated on 1.0% agarose gels and visualized using the Image LabTM program.
4.8. Plant Phenotyping
The T1 generation of Ramy3D::MdFT1 rice cultivated in 2022 was used for plant phenotyping, as previously described [28]. The heading date was recorded when the first panicle emerged. Leaf angle was measured during the vegetative phase for the third and fourth leaves using a protractor. Branching (tiler number) was evaluated during the vegetative phase. Plant height and internode length were measured during the ripening phase, and plant height was determined from the soil base to the panicles. Number of grains per panicle, grain weight, and grain size were assessed at maturity.
4.9. Microscopic Imaging Analysis
A light microscope (Axioskop 2; Carl Zeiss, Oberkochen, Germany) was used to analyze stem elongation at the cellular level. Sample preparation and light microscopy were performed as described previously [20]. Cell size was measured using Olympus cellSens Standard software (Olympus, Shinjuku, Tokyo, Japan).
To observe GFP fluorescence in calli and whole plants, including in different organs (leaves, stems, and roots), an In Vivo Imaging System (IVIS Lumina III; PerkinElmer, Massachusetts, USA) was employed. The specimens for observation were pre-adapted to complete darkness in a dark chamber for 1.5 h prior to evaluation. GFP imaging data were processed using a program sequence setup consisting of an automatic unmixing method with all wavelengths and a manual unmixing method with excitation at 488 nm and emission at 504–770 nm in 20 nm increments for 20 min. The optical fluorescence imaging data were displayed in pseudocolor, representing intensity. Measurements were repeated thrice with different specimens, and the signal intensity of each optical image was calculated within the region of interest.
eGFP fluorescence was visualized at the cellular level using a Zeiss LSM 900 Airyscan 2/ GaAsP Confocal Multiplex Microscope (Zeiss, Oberkochen, Germany). Transgenic tissues of Ubi::Cas9–eGFP plants were visualized using a blue filter, which induced green light transmission from the GFP protein.
4.10. Measurements of Free Amino Acid Contents
T1 rice seeds of Ramy3D::MdFT1 and ‘Dongjin’ WT plants were ground to a fine powder in liquid nitrogen using a mortar and pestle for free amino acid content analyses. Free amino acid content in the rice seeds was analyzed using a Hitachi LA8080 automatic amino acid analyzer (Hitachi High-Tech Group, Tokyo, Japan). Free amino acid content in rice seeds was measured using the post-column ninhydrin method [53]. Briefly, 50 mg of powdered grain was homogenized with 1 ml of 4% sulfosalicylic acid by vortexing, followed by centrifugation at 1,200 rpm for 15 min at 4°C. Subsequently, the collected supernatant was mixed with an equal volume of 200 mM HCl, then filtrated through a 0.22-µm filter to remove debris. A 20 μL aliquot of filtered sample was injected to measure free amino acid content. Owing to the ninhydrin reaction, a blue-violet substance was produced, and its absorbance was measured at 570 nm.
4.11. Data Analysis
Statistical analyses were performed using Microsoft Excel and IBM SPSS (IBM Corp., Armonk, NY, USA). Significant differences between pairwise groups were analyzed using one-way ANOVA. All data are presented as the means ± standard deviations (SD) from multiple replicates of independent experiments, with p < 0.05 indicating significant differences.
5. Conclusions
In this study, we developed an efficient and cost-effective Agrobacterium-mediated transformation method for producing transgenic rice plants using seed-derived embryogenic calli in an in vitro tissue culture system. By eliminating the need for complex culture media, we reduced both the time and resources required while maintaining high transformation efficiency. The use of seed-derived embryogenic calli is crucial to ensure consistent and robust regeneration. This method enabled the stable integration and expression of the eGFP and MdFT1 genes in the rice genome. The eGFP gene allowed for clear visualization of transgene expression, while introducing the antisense MdFT1 gene effectively modified key agronomic traits such as plant height and yield potential, highlighting the biological impact of genetic modifications. Additionally, this study underscores the effectiveness of the Agrobacterium-mediated transformation method in gene transfer, particularly when used with strong promoter systems such as the Ubi and Ramy3D promoters. The introduced traits were stably inherited across generations, thus confirming the reliability of this approach. The process described here is a simplified, scalable method for the genetic transformation of rice, paving the way for enhanced functional gene analysis and the development of genetically modified crops with improved traits.
Supplementary Materials
The following supporting information can be downloaded from: preprints.org. Figure S1. Observation of green fluorescent protein (eGFP) in rice calli of wildtype (a) and transgenic Ubi::Cas9‒eGFP lines (b); Figure S2. Observation of green fluorescent protein (eGFP) in different organs, including leaves (a), stems and roots (b) in wildtype rice plants (T1 seedlings); Figure S3. Observation of green fluorescent protein (eGFP) in rice plants (T1 seedlings) of transgenic Ubi::Cas9‒eGFP lines in different organs including leave (a), stem and root (b). Table S1‒4. Free amino acid content.
Author Contributions
Conceptualization, V.G.D.; methodology, V.G.D., Y.L. and G.D.; validation, J.P.; H.K. and S.K.; formal analysis, V.G.D.; Y.L. and N.M.W.; investigation, S.K.; V.G.D. and Y.L.; resources, S.-I.K. and S.Y.; data curation, J.P., G.D. and S.K.; writing—original draft preparation, V.G.D.; writing—review and editing, V.G.D.; Y.L.; S.K. and N.M.W.; visualization, Y.L., V.G.D. and G.D.; supervision, S.-I.K.; H.K. and S.Y.; project administration, Y.L.; funding acquisition, Y.L. and S.-I.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the International Technology Cooperation Center Research Program (grant number PJ0173002024), Rural Development Administration, Republic of Korea.
Data Availability Statement
Data are presented within the article and Supplementary Materials.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mohidem, N.A.; Hashim, N.; Shamsudin, R.; Che Man, H. Rice for food security: Revisiting its production, diversity, rice milling process and nutrient content. Agriculture 2022, 12, 741. [Google Scholar] [CrossRef]
- Zhao, M.; Lin, Y.; Chen, H. Improving nutritional quality of rice for human health. Theor. Appl. Genet. 2020, 133, 1397–1413. [Google Scholar] [CrossRef] [PubMed]
- Fukagawa, N.K.; Ziska, L.H. Rice: Importance for global nutrition. J. Nutr. Sci. Vitaminol. 2019, 65, S2–S3. [Google Scholar] [CrossRef]
- Gelvin, S.B. Traversing the cell: Agrobacterium T-DNA’s journey to the host genome. Front. Plant Sci. 2012, 3, 52. [Google Scholar] [CrossRef]
- Binns, A.N.; Thomashow, M.F. Cell biology of Agrobacterium infection and transformation of plants. Annu. Rev. Microbiol. 1988, 42, 575–606. [Google Scholar] [CrossRef]
- Ayres, N.M.; Park, W.D. Genetic transformation of rice. Crit. Rev. Plant Sci. 1994, 13, 219–239. [Google Scholar] [CrossRef]
- Lv, Z.; Jiang, R.; Chen, J.; Chen, W. Nanoparticle-mediated gene transformation strategies for plant genetic engineering. Plant J. 2020, 104, 880–891. [Google Scholar] [CrossRef] [PubMed]
- Azizi-Dargahlou, S.; Pouresmaeil, M. Agrobacterium tumefaciens-mediated plant Transformation: a review. Mol. Biotechnol. 2024, 66, 1563–1580. [Google Scholar] [CrossRef]
- Gelvin, S.B. Agrobacterium-mediated plant transformation: the biology behind the “gene-jockeying” tool. Microbiol. Mol. Biol. 2003, 67, 16–37. [Google Scholar] [CrossRef]
- Ziemienowicz, A.; Tzfira, T.; Hohn, B. Mechanisms of T-DNA integration. In In: Tzfira, T., Citovsky, V. (eds) Agrobacterium: from biology to biotechnology; Springer: New York, USA. 2008; p. 395440. [Google Scholar] [CrossRef]
- Christie, P.J. Type IV secretion: the Agrobacterium VirB/D4 and related conjugation systems. Biochim. Biophys. Acta - Mol. Cell Res. 2004, 1694, 219–234. [Google Scholar] [CrossRef]
- Anand, A.; Mysore, K. Agrobacterium biology and crown gall disease. In: Gnanamanickam, S.S. (eds) Plant-Associated Bacteria. Springer: Dordrecht, Netherlands. 2006; pp. 359‒384. [CrossRef]
- Pitzschke, A.; Hirt, H. New insights into an old story: Agrobacterium-induced tumour formation in plants by plant transformation. EMBO J. 2010, 29, 1021–1032. [Google Scholar] [CrossRef]
- Gelvin, S.B. Integration of Agrobacterium T-DNA into the plant genome. Annu. Rev. Genet. 2017, 51, 195–217. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.-H.; Yu, M.; Lai, E.-M. Agrobacterium-mediated plant transformation: biology and applications. The arabidopsis book 2017, 15. [Google Scholar] [CrossRef] [PubMed]
- Dai, S.; Zheng, P.; Marmey, P.; Zhang, S.; Tian, W.; Chen, S.; Beachy, R.N.; Fauquet, C. Comparative analysis of transgenic rice plants obtained by Agrobacterium-mediated transformation and particle bombardment. Mol. Breed. 2001, 7, 25–33. [Google Scholar] [CrossRef]
- Hobbs, S.L.; Warkentin, T.D.; DeLong, C.M. Transgene copy number can be positively or negatively associated with transgene expression. Plant Mol. Biol. 1993, 21, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Rivera, A.L.; Gómez-Lim, M.; Fernández, F.; Loske, A.M. Physical methods for genetic plant transformation. Phys. Life Rev. 2012, 9, 308–345. [Google Scholar] [CrossRef]
- Heidari Japelaghi, R.; Haddad, R.; Valizadeh, M.; Dorani Uliaie, E.; Jalali Javaran, M. High-efficiency Agrobacterium-mediated transformation of tobacco (Nicotiana tabacum). J. Plant Mol. Breed. 2018, 6, 38–50. [Google Scholar] [CrossRef]
- Do, V.G.; Lee, Y.; Kim, S.; Kweon, H.; Do, G. Antisense expression of apple TFL1-like gene (MdTFL1) promotes early flowering and causes phenotypic changes in tobacco. Int. J. Mol. Sci. 2022, 23, 6006. [Google Scholar] [CrossRef]
- Niedbała, G.; Niazian, M.; Sabbatini, P. Modeling Agrobacterium-mediated gene transformation of tobacco (Nicotiana tabacum)—a model plant for gene transformation studies. Front. Plant Sci. 2021, 12, 695110. [Google Scholar] [CrossRef]
- Chan, M.-T.; Chang, H.-H.; Ho, S.-L.; Tong, W.-F.; Yu, S.-M. Agrobacterium-mediated production of transgenic rice plants expressing a chimeric α-amylase promoter/β-glucuronidase gene. Plant Mol. Biol. 1993, 22, 491–506. [Google Scholar] [CrossRef]
- Dong, J.; Teng, W.; Buchholz, W.G.; Hall, T.C. Agrobacterium-mediated transformation of Javanica rice. Mol. Breed. 1996, 2, 267–276. [Google Scholar] [CrossRef]
- Hiei, Y.; Ohta, S.; Komari, T.; Kumashiro, T. Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J. 1994, 6, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Hiei, Y.; Komari, T. Improved protocols for transformation of indica rice mediated by Agrobacterium tumefaciens. Plant Cell Tissue Organ Cult. 2006, 85, 271–283. [Google Scholar] [CrossRef]
- Nishimura, A.; Aichi, I.; Matsuoka, M. A protocol for Agrobacterium-mediated transformation in rice. Nat. Protoc. 2006, 1, 2796–2802. [Google Scholar] [CrossRef]
- Rahman, S.U.; Khan, M.O.; Ullah, R.; Ahmad, F.; Raza, G. Agrobacterium-mediated transformation for the development of transgenic crops; present and future prospects. Mol. Biotechnol. 2023, 1–17. [Google Scholar] [CrossRef]
- Do, V.G.; Lee, Y.; Kim, S.; Yang, S.; Park, J.; Do, G. Introducing MdTFL1 promotes heading date and produces semi-draft phenotype in rice. Int. J. Mol. Sci. 2023, 24, 10365. [Google Scholar] [CrossRef]
- Olhoft, P.M.; Bernal, L.M.; Grist, L.B.; Hill, D.S.; Mankin, S.L.; Shen, Y.; Kalogerakis, M.; Wiley, H.; Toren, E.; Song, H.-S. A novel Agrobacterium rhizogenes-mediated transformation method of soybean [Glycine max (L.) Merrill] using primary-node explants from seedlings. In Vitro Cell. Dev. Biol. Plant. 2007, 43, 536–549. [Google Scholar] [CrossRef]
- Ishida, Y.; Hiei, Y.; Komari, T. Agrobacterium-mediated transformation of maize. Nat. Protoc. 2007, 2, 1614–1621. [Google Scholar] [CrossRef]
- Hashmi, M.H.; Saeed, F.; Demirel, U.; Bakhsh, A. Establishment of highly efficient and reproducible Agrobacterium-mediated transformation system for tomato (Solanum lycopersicum L.). In Vitro Cell. Dev. Biol. Plant. 2022, 58, 1066–1076. [Google Scholar] [CrossRef]
- Kong, J.; Martin-Ortigosa, S.; Finer, J.; Orchard, N.; Gunadi, A.; Batts, L.A.; Thakare, D.; Rush, B.; Schmitz, O.; Stuiver, M. Overexpression of the transcription factor GROWTH-REGULATING FACTOR5 improves transformation of dicot and monocot species. Front. Plant Sci. 2020, 11, 572319. [Google Scholar] [CrossRef]
- Noureen, A.; Zuhaib Khan, M.; Amin, I.; Zainab, T.; Ahmad, N.; Haider, S.; Mansoor, S. Broad-spectrum resistance against multiple PVY-strains by CRSIPR/Cas13 system in Solanum tuberosum crop. GM crops & food. 2022, 13, 97–111. [Google Scholar] [CrossRef]
- Bakhsh, A. Development of efficient, reproducible and stable Agrobacterium-mediated genetic transformation of five potato cultivars. Food Technol. Biotechnol. 2020, 58, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Howe, A.; Sato, S.; Dweikat, I.; Fromm, M.; Clemente, T. Rapid and reproducible Agrobacterium-mediated transformation of sorghum. Plant Cell Rep. 2006, 25, 784–791. [Google Scholar] [CrossRef]
- Hiei, Y.; Komari, T. Agrobacterium-mediated transformation of rice using immature embryos or calli induced from mature seed. Nat. Protoc. 2008, 3, 824–834. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, K.; Kawahigashi, H.; Kayano, T.; Ohkawa, Y. Enhancement of regeneration of rice (Oryza sativa L.) calli by integration of the gene involved in regeneration ability of the callus. Plant Sci. 2003, 165, 395–402. [Google Scholar] [CrossRef]
- Toki, S. Rapid and efficient Agrobacterium-mediated transformation. Plant Mol. Biol. Rep. 1997, 15, 16–21. [Google Scholar] [CrossRef]
- Frame, B.R.; Zhang, H.; Cocciolone, S.M.; Sidorenko, L.V.; Dietrich, C.R.; Pegg, S.E.; Zhen, S.; Schnable, P.S.; Wang, K. Production of transgenic maize from bombarded type II callus: effect of gold particle size and callus morphology on transformation efficiency. In Vitro Cell. Dev. Biol. Plant. 2000, 36, 21–29. [Google Scholar] [CrossRef]
- Gordon-Kamm, W.J.; Spencer, T.M.; Mangano, M.L.; Adams, T.R.; Daines, R.J.; Start, W.G.; O’Brien, J.V.; Chambers, S.A.; Adams, W.R.; Willetts, N.G. Transformation of maize cells and regeneration of fertile transgenic plants. The Plant Cell. 1990, 2, 603–618. [Google Scholar] [CrossRef]
- Xie, K.; Minkenberg, B.; Yang, Y. Boosting CRISPR/Cas9 multiplex editing capability with the endogenous tRNA-processing system. PNAS. 2015, 112, 3570–3575. [Google Scholar] [CrossRef]
- Zafar, K.; Khan, M.Z.; Amin, I.; Mukhtar, Z.; Yasmin, S.; Arif, M.; Ejaz, K.; Mansoor, S. Precise CRISPR-Cas9 mediated genome editing in super basmati rice for resistance against bacterial blight by targeting the major susceptibility gene. Front. Plant Sci. 2020, 11, 575. [Google Scholar] [CrossRef]
- Molina-Risco, M.; Ibarra, O.; Faion-Molina, M.; Kim, B.; Septiningsih, E.M.; Thomson, M.J. Optimizing Agrobacterium-mediated transformation and CRISPR-Cas9 gene editing in the tropical japonica rice variety presidio. Int. J. Mol. Sci. 2021, 22, 10909. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Tian, J.; Li, R.; Chen, X.; Luo, Z.; Chen, M.; Zhao, X.; Zhang, D.; Persson, S.; Yuan, Z. Investigation of CRISPR/Cas9-induced SD1 rice mutants highlights the importance of molecular characterization in plant molecular breeding. J Genet Genome. 2020, 47, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Upadhyaya, N.M.; Surin, B.; Ramm, K.; Gaudron, J.; Schünmann, P.H.; Taylor, W.; Waterhouse, P.M.; Wang, M.-B. Agrobacterium-mediated transformation of Australian rice cultivars Jarrah and Amaroo using modified promoters and selectable markers. Funct. Plant Biol. 2000, 27, 201–210. [Google Scholar] [CrossRef]
- Shin, Y.J.; Hong, S.Y.; Kwon, T.H.; Jang, Y.S.; Yang, M.S. High level of expression of recombinant human granulocyte-macrophage colony stimulating factor in transgenic rice cell suspension culture. Biotechnol. Bioeng. 2003, 82, 778–783. [Google Scholar] [CrossRef]
- Van Giap, D.; Jung, J.-W.; Kim, N.-S. Production of functional recombinant cyclic citrullinated peptide monoclonal antibody in transgenic rice cell suspension culture. Transgenic Res. 2019, 28, 177–188. [Google Scholar] [CrossRef]
- Chung, N.-D.; Kim, N.-S.; Jang, S.-H.; Oh, S.-M.; Jang, S.-H.; Kim, T.-G.; Jang, Y.-S.; Yang, M.-S. Production of functional human vascular endothelial growth factor165 in transgenic rice cell suspension cultures. Enzyme Microb. Technol. 2014, 63, 58–63. [Google Scholar] [CrossRef]
- Jung, J.-W.; Huy, N.-X.; Kim, H.-B.; Kim, N.-S.; Yang, M.-S. Production of recombinant human acid α-glucosidase with high-mannose glycans in gnt1 rice for the treatment of Pompe disease. J. Biotech. 2017, 249, 42–50. [Google Scholar] [CrossRef]
- Do, V.G.; Yang, M.-S. Production of mature recombinant human Activin A in transgenic rice cell suspension culture. Curr. Issues Mol. Biol. 2024, 46, 1164–1176. [Google Scholar] [CrossRef]
- Weigel, D.; Glazebrook, J. Transformation of Agrobacterium using the freeze-thaw method. Cold Spring Harb. Protoc. 2006, 7, pdb–prot4666. [Google Scholar] [CrossRef]
- Do, V.G.; Lee, Y.; Kim, J.-H.; Kwon, Y.-S.; Park, J.-T.; Yang, S.; Park, J.; Win, N.M.; Kim, S. The synergistic effects of environmental and genetic factors on the regulation of anthocyanin accumulation in plant tissues. Int. J. Mol. Sci. 2023, 24, 12946. [Google Scholar] [CrossRef]
- Liu, H.; Gong, X.; Deng, H.; Tan, J.; Sun, Y.; Wang, F.; Wu, W.; Zhou, Z.; Xu, R.; He, H. The rice aspartyl-tRNA synthetase YLC3 regulates amino acid homeostasis and chloroplast development under low temperature. Front. Plant Sci. 2022, 13, 847364. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Construction of Ubi::Cas9–eGFP (a) and Ramy3D::MdFT1 (b) binary vectors for the expression of GOI in rice embryonic calli. The eGFP gene was cloned into the binary vector pRGEB32, whereas the MdFT1 gene was cloned into the pMYD317 expression vector between the XbaI‒KpnI sites. LB, T-DNA left border; poly(A), cauliflower mosaic virus (CaMV) polyadenylation signal; HygR, hygromycin B phosphotransferase; 35S.p, CaMV 35S promoter; Ubi.p, rice (Oryza sativa) polyubiquitin promoter; Cas9, Cas9 (Csn1) endonuclease from the Streptococcus pyogenes Type II CRISPR/Cas system; NOS, nopaline synthase terminator; eGFP, enhanced green fluorescent protein; Ramy3D.p, rice alpha-amylase promoter; 3′-UTR, 3′-untranslated region of the rice α-amylase 3D gene; RB, T-DNA right border.
Figure 1.
Construction of Ubi::Cas9–eGFP (a) and Ramy3D::MdFT1 (b) binary vectors for the expression of GOI in rice embryonic calli. The eGFP gene was cloned into the binary vector pRGEB32, whereas the MdFT1 gene was cloned into the pMYD317 expression vector between the XbaI‒KpnI sites. LB, T-DNA left border; poly(A), cauliflower mosaic virus (CaMV) polyadenylation signal; HygR, hygromycin B phosphotransferase; 35S.p, CaMV 35S promoter; Ubi.p, rice (Oryza sativa) polyubiquitin promoter; Cas9, Cas9 (Csn1) endonuclease from the Streptococcus pyogenes Type II CRISPR/Cas system; NOS, nopaline synthase terminator; eGFP, enhanced green fluorescent protein; Ramy3D.p, rice alpha-amylase promoter; 3′-UTR, 3′-untranslated region of the rice α-amylase 3D gene; RB, T-DNA right border.
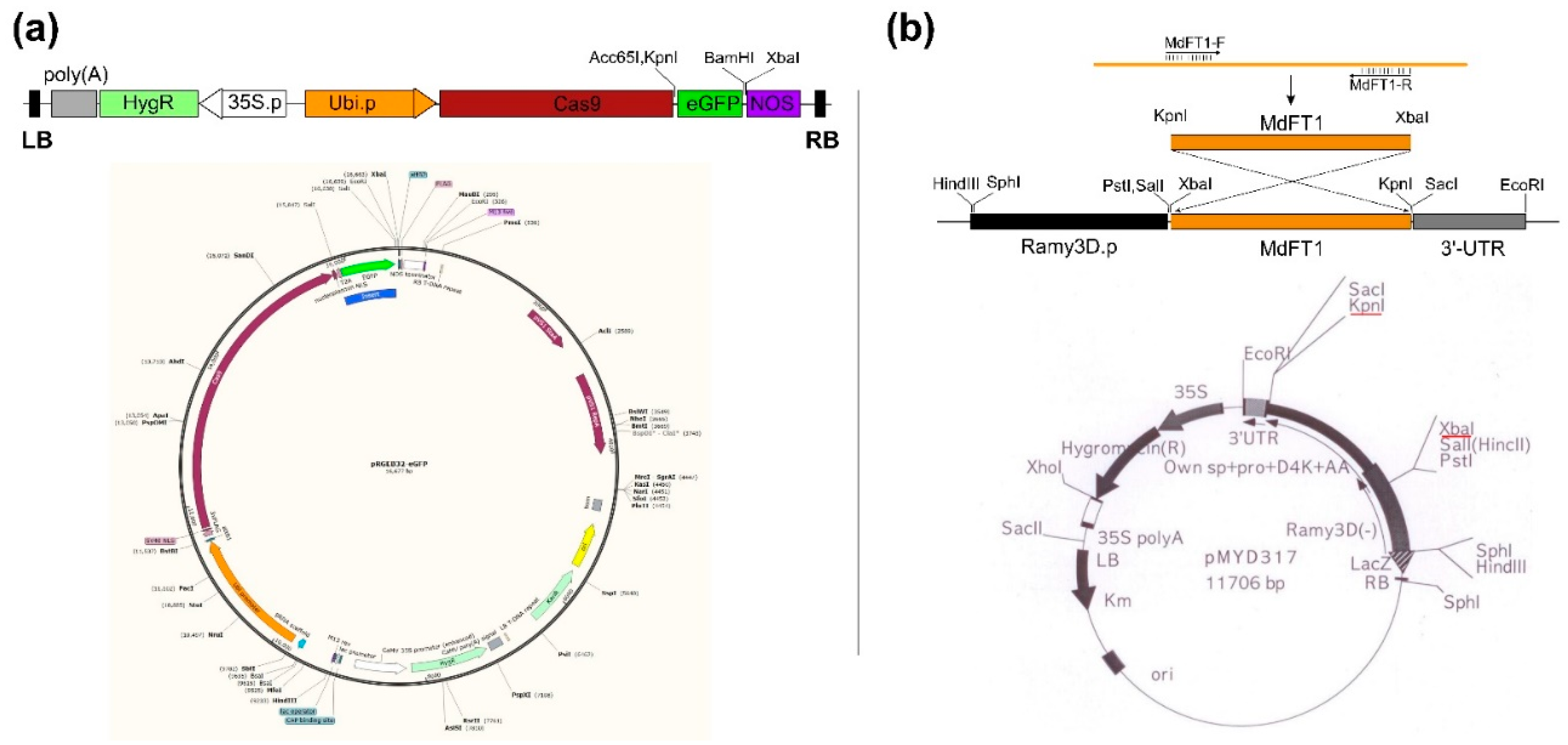
Figure 2.
Scheme establishment of transgenic rice using Agrobacterium-mediated transformation via in vitro tissue culture. (a) Scutellum-derived calli were induced from rice seeds on callus induction medium N6CI. (b) Embryonic calli were used as explant for genetic transformation. (c) Co-culture of Agrobacterium and rice embryonic calli after Agrobacterium-mediated transformation. (d) Transgenic hygromycin-resistant calli growth on selection medium N6SE. (e) Putative transgenic rice calli propagation on N6SE. (f) Magnified image of transgenic rice callus line. (g, h) Growth of transgenic lines on shooting medium MSS with a magnified image. (i, j) Bud differentiation on MSS after 4 and 6 weeks, respectively. (k) Seedling growth on rooting medium MSR. (l) Transplantation of fully rooted seedlings for ex vitro culture (sowing into the soil). The red circle in (d) indicates the non-transformed callus cannot grow on N6SE. The red arrow in (g, h) indicates meristem shoots were initiated from the calli.
Figure 2.
Scheme establishment of transgenic rice using Agrobacterium-mediated transformation via in vitro tissue culture. (a) Scutellum-derived calli were induced from rice seeds on callus induction medium N6CI. (b) Embryonic calli were used as explant for genetic transformation. (c) Co-culture of Agrobacterium and rice embryonic calli after Agrobacterium-mediated transformation. (d) Transgenic hygromycin-resistant calli growth on selection medium N6SE. (e) Putative transgenic rice calli propagation on N6SE. (f) Magnified image of transgenic rice callus line. (g, h) Growth of transgenic lines on shooting medium MSS with a magnified image. (i, j) Bud differentiation on MSS after 4 and 6 weeks, respectively. (k) Seedling growth on rooting medium MSR. (l) Transplantation of fully rooted seedlings for ex vitro culture (sowing into the soil). The red circle in (d) indicates the non-transformed callus cannot grow on N6SE. The red arrow in (g, h) indicates meristem shoots were initiated from the calli.

Figure 3.
PCR-based screening of transgenic rice calli expressing Ram3D::MdFT1. (a) Detection of the selection marker gene HygR and MdFT1 using specific primers. (b) Detection of HygR and eGFP in Ubi::Cas9‒eGFP transgenic lines. (c) RT-PCR analysis to evaluate the expression of MdFT1 mRNA in transgenic rice calli, normalized to the rice reference gene OsAct1. (d) 50 bp and 1 Kb plus DNA ladders with size standards (Dyne Bio Inc. Seongnam-si, Republic of Korea). M, 1 Kb plus DNA ladder; PC, plasmid DNA of 3D::MdTFL1 and Ubi::Cas9‒eGFP (isolated from E. coli) as positive control; NC, genomic DNA of WT callus lines as negative control.
Figure 3.
PCR-based screening of transgenic rice calli expressing Ram3D::MdFT1. (a) Detection of the selection marker gene HygR and MdFT1 using specific primers. (b) Detection of HygR and eGFP in Ubi::Cas9‒eGFP transgenic lines. (c) RT-PCR analysis to evaluate the expression of MdFT1 mRNA in transgenic rice calli, normalized to the rice reference gene OsAct1. (d) 50 bp and 1 Kb plus DNA ladders with size standards (Dyne Bio Inc. Seongnam-si, Republic of Korea). M, 1 Kb plus DNA ladder; PC, plasmid DNA of 3D::MdTFL1 and Ubi::Cas9‒eGFP (isolated from E. coli) as positive control; NC, genomic DNA of WT callus lines as negative control.
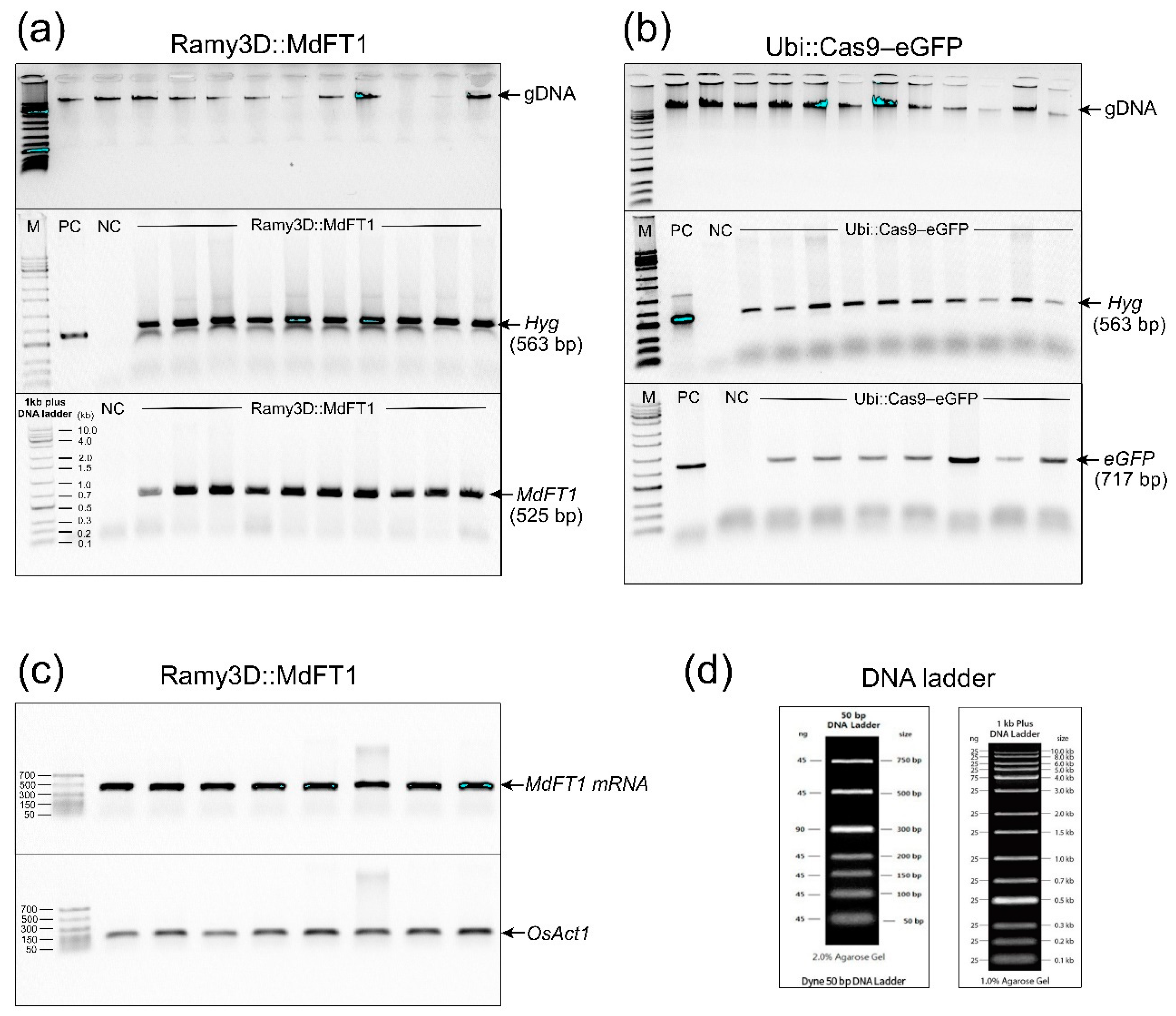
Figure 4.
gDNA PCR confirmation of transgenes in T1 transgenic rice plants. Seedlings of transgenic rice Ram3D::MdFT1 (a) and Ubi::Cas9‒eGFP (b) were grown in a greenhouse. Leaves from T1 transgenic plants at the vegetative stage were collected for gDNA PCR. PCR was conducted to detect the MdFT1 gene (c), reporter gene eGFP (d), and selective marker gene HypR (c, d) using specific primer sets. M, 1 Kb plus DNA ladder; PC, positive control used plasmid DNA of 3D::MdTFL1 and Ubi::Cas9‒eGFP (isolated from E. coli) as DNA templates; NC, negative control used genomic DNA of WT lines as DNA templates.
Figure 4.
gDNA PCR confirmation of transgenes in T1 transgenic rice plants. Seedlings of transgenic rice Ram3D::MdFT1 (a) and Ubi::Cas9‒eGFP (b) were grown in a greenhouse. Leaves from T1 transgenic plants at the vegetative stage were collected for gDNA PCR. PCR was conducted to detect the MdFT1 gene (c), reporter gene eGFP (d), and selective marker gene HypR (c, d) using specific primer sets. M, 1 Kb plus DNA ladder; PC, positive control used plasmid DNA of 3D::MdTFL1 and Ubi::Cas9‒eGFP (isolated from E. coli) as DNA templates; NC, negative control used genomic DNA of WT lines as DNA templates.

Figure 5.
Observation of green fluorescent protein (eGFP) in rice calli (a) and various organs of T1 seedlings expressing Ubi::Cas9‒eGFP, including leaves (b), stems (c), and roots (d), compared to WT plants.
Figure 5.
Observation of green fluorescent protein (eGFP) in rice calli (a) and various organs of T1 seedlings expressing Ubi::Cas9‒eGFP, including leaves (b), stems (c), and roots (d), compared to WT plants.
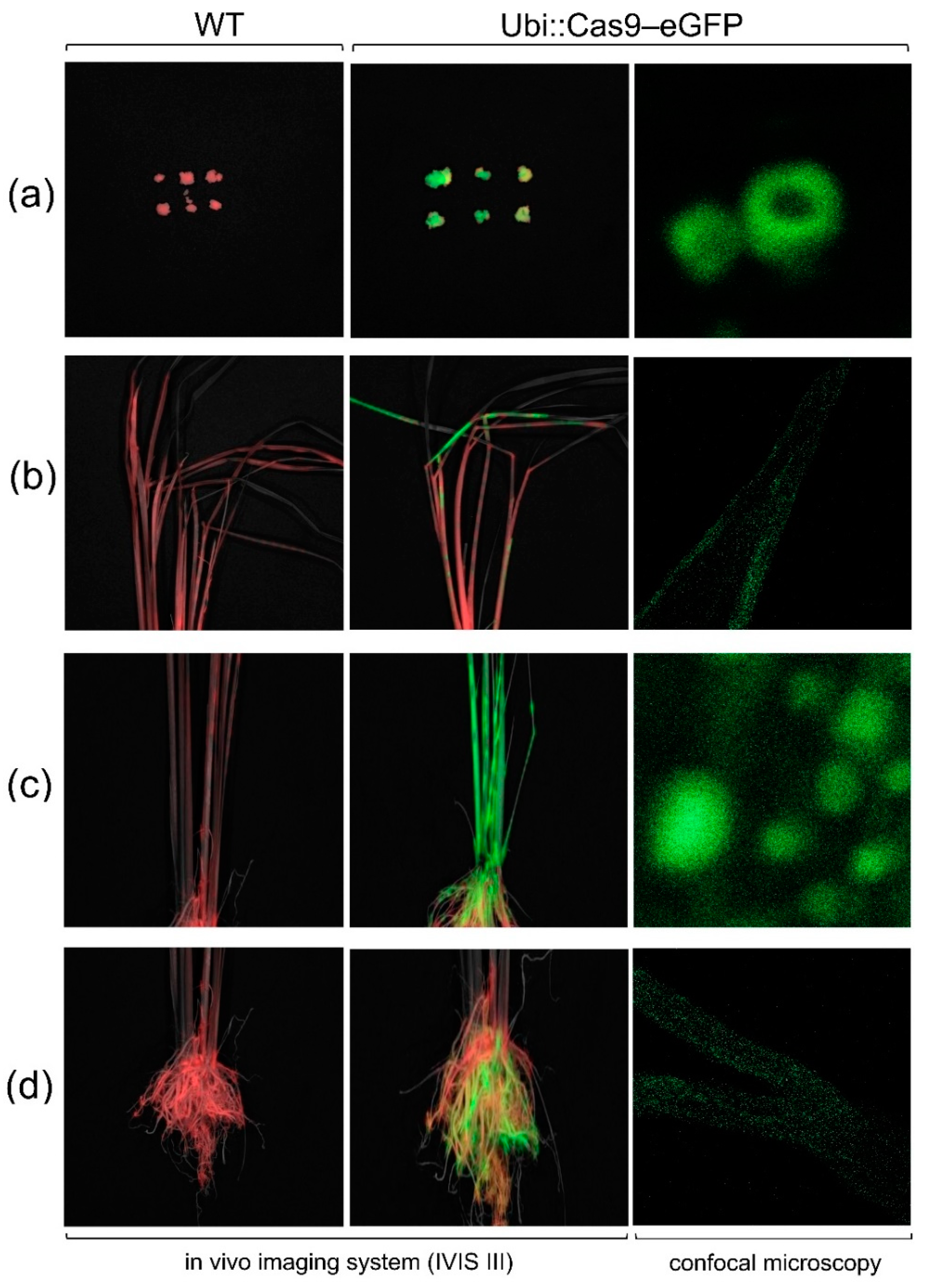
Figure 6.
Phenotyping of transgenic rice expressing Ramy3D::MdFT1 compared to WT ‘Dongjin’ cultivar. The analysis includes evaluation of heading dates (a, b), leaf angle (c, d), branching at vegetative developmental stage (e, f), branching at harvest stage (g), and the number of tillers during different developmental stages (h). Data are presented as means ± standard deviations (SD) (n ≥ 20). Asterisks indicate significant differences (p < 0.05).
Figure 6.
Phenotyping of transgenic rice expressing Ramy3D::MdFT1 compared to WT ‘Dongjin’ cultivar. The analysis includes evaluation of heading dates (a, b), leaf angle (c, d), branching at vegetative developmental stage (e, f), branching at harvest stage (g), and the number of tillers during different developmental stages (h). Data are presented as means ± standard deviations (SD) (n ≥ 20). Asterisks indicate significant differences (p < 0.05).
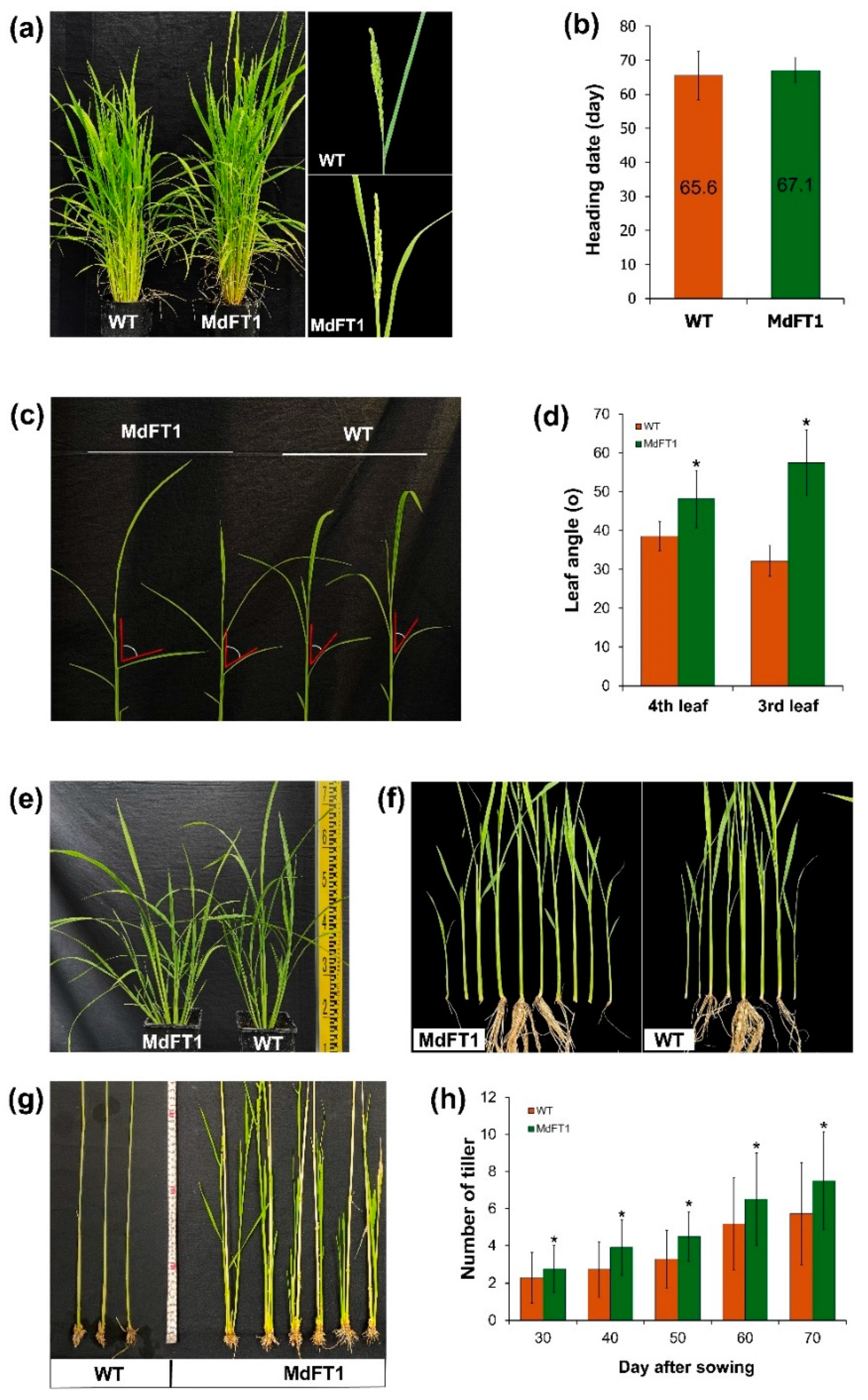
Figure 7.
Phenotyping of transgenic rice Ramy3D::MdFT1 in comparison to wild type ‘Dongjin’ cultivar. Transgenic rice developed with taller in overall heights (a) and internode (IN2) lengths (b, c). Microscopy imaging of elongation zone (IN2) (d), and cellular size in width (e), length (f), and length/width ratio (g). Data are presented as means ± SD (n ≥ 20). Asterisks indicate significant differences (p < 0.05).
Figure 7.
Phenotyping of transgenic rice Ramy3D::MdFT1 in comparison to wild type ‘Dongjin’ cultivar. Transgenic rice developed with taller in overall heights (a) and internode (IN2) lengths (b, c). Microscopy imaging of elongation zone (IN2) (d), and cellular size in width (e), length (f), and length/width ratio (g). Data are presented as means ± SD (n ≥ 20). Asterisks indicate significant differences (p < 0.05).
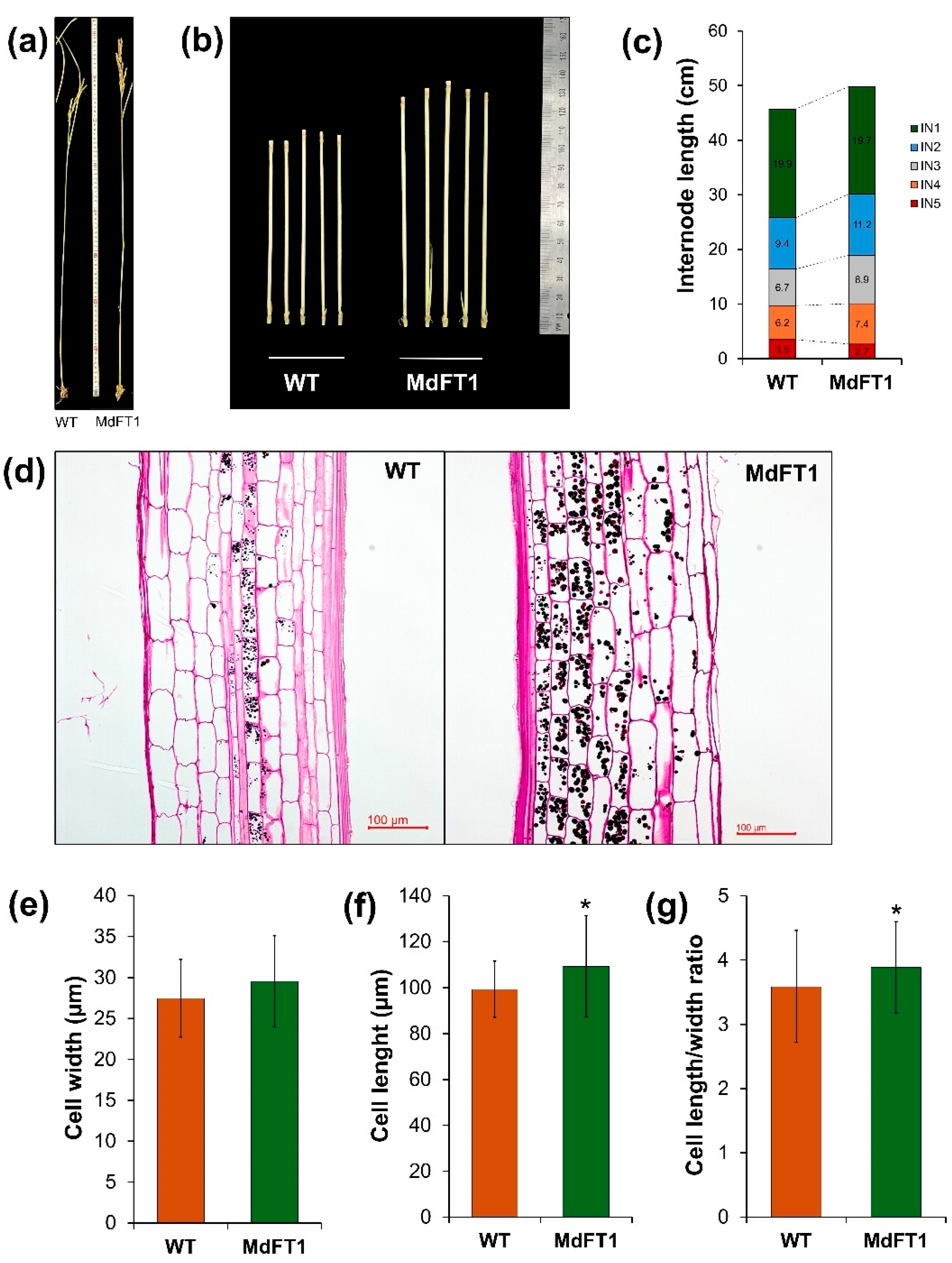
Figure 8.
Evaluation of yield potential in transgenic rice expressing MdFT1. Rice panicles at ripening stage (a) with yield potential (number of grains per panicle) (b). Seed production was measured with 100 grains (c). Grain size (d‒g) and the average weight of 100 grains (h) were also assessed. Data are presented as means ± SD. Asterisks indicate significant differences (p < 0.05).
Figure 8.
Evaluation of yield potential in transgenic rice expressing MdFT1. Rice panicles at ripening stage (a) with yield potential (number of grains per panicle) (b). Seed production was measured with 100 grains (c). Grain size (d‒g) and the average weight of 100 grains (h) were also assessed. Data are presented as means ± SD. Asterisks indicate significant differences (p < 0.05).
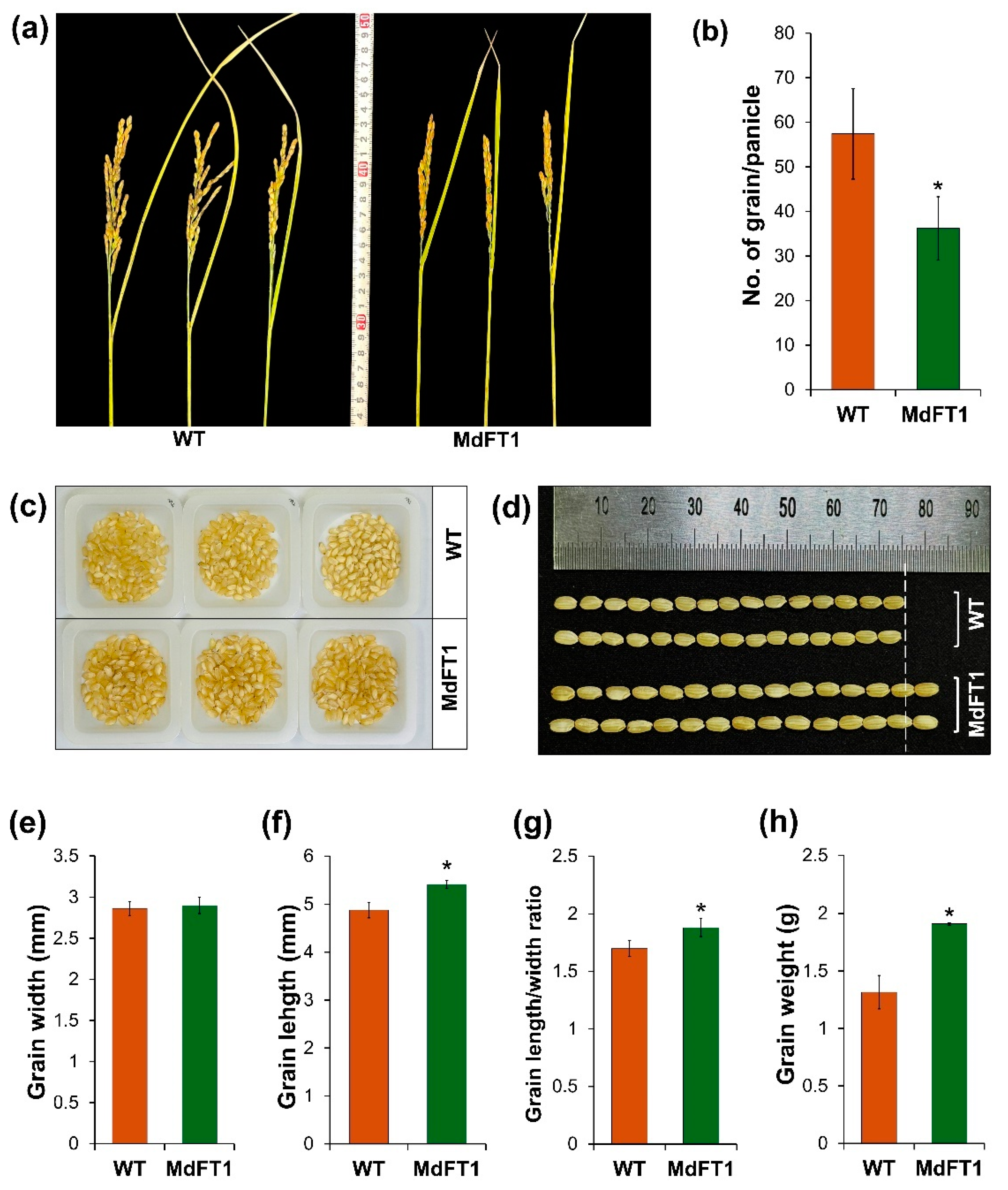

Table 1.
Primer sequences for gene cloning, genomic DNA PCR, and qRT-PCR.
| Primer Set | Primer Name | Sequence (5′→3′) | Amplicon length (bp) | Note | |
|---|---|---|---|---|---|
| 1 | HygR | F | CTCGGAGGGCGAAGAATCTC | 563 | Used for gDNA PCR |
| R | CAATGACCGCTGTTATGCGG | ||||
| 2 | eGFP | F | GTGAGCAAGGGCGAGGAGCT | 717 | Used for gDNA PCR |
| R | TTACTTGTACAGCTCGTCCATGCCGAG | ||||
| 3 | MdFT1 | F | GGTACCATGCCTAGGGATAGGGAC | 525 | Used for gene cloning and gDNA PCR and RT-PCR |
| R | TCTAGATTATCTTCTCCTTCCACCG | ||||
| 4 | OsAct1 | F | GCGTCTGGATTGGTGGTTCT | 142 | Used for RT-PCR |
| R | ACCGCTCTACAAACTTGGCA | ||||
The underlined sequences indicate the restriction enzyme sites. The bold sequences indicate the start and stop codons. F and R denote the forward and reverse primers, respectively.
Table 2.
Comparison of free amino acid in grains of WT and Ramy3D::MdFT1 transgenic rice.
| Amino acid (a.a) | Concentration (mg/mL) * | ||
|---|---|---|---|
| Full name | Abbreviation | WT | MdFT1 |
| Aspartic acid | Asp | 9.04 ± 0.17 | 10.72 ± 0.02 |
| Threonine | Thr | 3.42 ± 0.08 | 4.13 ± 0.01 |
| Serine | Ser | 4.03 ± 0.09 | 4.68 ± 0.01 |
| Glutamic acid | Glu | 15.27 ± 0.32 | 17.38 ± 0.10 |
| Glycine | Gly | 3.68 ± 0.11 | 4.91 ± 0.03 |
| Alanine | Ala | 7.16 ± 0.26 | 7.90 ± 0.04 |
| Cysteine | Cys | 0.88 ± 0.03 | 1.36 ± 0.01 |
| Valine | Val | 52.68 ± 1.53 | 59.69 ± 0.34 |
| Methionine | Met | 1.94 ± 0.06 | 2.27 ± 0.01 |
| Isoleucine | Ile | 3.92 ± 0.12 | 4.48 ± 0.02 |
| Leucine | Leu | 8.32 ± 0.22 | 9.49 ± 0.02 |
| Tyrosine | Tyr | 2.19 ± 0.04 | 2.69 ± 0.02 |
| Phenylalanine | Phe | 5.17 ± 0.11 | 5.97 ± 0.01 |
| Lysine | Lys | 4.30 ± 0.09 | 5.52 ± 0.01 |
| NH3 | 1.91 ± 0.10 | 2.02 ± 0.04 | |
| Histidine | His | 2.31 ± 0.05 | 2.82 ± 0.00 |
| Arginine | Arg | 7.08 ± 0.15 | 7.80 ± 0.01 |
| Proline | Pro | 3.63 ± 0.72 | 4.66 ± 0.93 |
Data are the means ± SD (n = 2). Means followed by a double asterisk (*) within columns of concentration are significantly different according to one-way ANOVA (p < 0.05).
Table 3.
Composition of the media used for plant tissue in vitro culture.
| Component | Producer /Product No. |
Callus induction (N6CI) |
Co-culture (N6CO) |
Selection (N6SE) |
Shooting (MSS) |
Rooting (MSR) |
|---|---|---|---|---|---|---|
| CHU (N6) plus vitamins |
Duchefa Biochemie C0204.0050 |
4.0 | 4.0 | 4.0 | ||
| MS plus vitamins |
Duchefa Biochemie M0222.0050 |
4.3 | 4.3 | |||
| Sucrose | Duchefa Biochemie S0809.100 |
30 | 30 | 30 | 30 | 30 |
| Glucose | Sigma Aldrich G5767-500G |
10 | ||||
| Sorbitol | Sigma Aldrich S3889-1KG |
30 | ||||
| Phytagel | Sigma Aldrich P8169-500G |
2.3 | 2.3 | 2.3 | ||
| Gerlite | Duchefa Biochemie G1101.0500 |
4.0 | 4.0 | |||
| 2,4-D | Duchefa Biochemie D0911.0250 |
2 mg/L | 2 mg/L | 2 mg/L | ||
| Kinetin | Duchefa Biochemie K0905.005 |
2 mg/L | 2 mg/L | 2 mg/L | 5 mg/L | |
| NAA | Duchefa Biochemie N0903.0025 |
1 mg/L | ||||
| IBA | Sigma Aldrich I5386-5G |
0.5 mg/L | ||||
| Acetosyringone | Sigma Aldrich D134406-5G |
100 µM | ||||
| Cefotaxime | Duchefa Biochemie C0111.0005 |
250 mg/L | 250 mg/L (OP)* | 250 mg/L (OP) | ||
| Hygromycin B | Duchefa Biochemie H0192.0001 |
50 mg/L | 50 mg/L (OP) |
50 mg/L (OP) |
||
| pH | 5.8 | 5.2 | 5.8 | 5.8 | 5.8 |
All hormones (2,4-D, Kinetin, NAA, IBA), antibiotics (Cefotaxime, Hygromycin B), and acetosyringone were of analytical grade. They were prepared in stock solutions (1.0 mg/mL for all hormones); 100 mM for acetosyringone; 100 mg/mL for Cefotaxime; and 50 mg/mL for Hygromycin B, and then sterilized using a 0.2-µm filter. * OP indicates optional, because transgenic lines had already been confirmed by gDNA PCR at the selection stage of in vitro cultivation.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



