
Submitted:
25 July 2024
Posted:
25 July 2024
You are already at the latest version
Abstract
Strawberry fruit is highly appreciated worldwide for its organoleptic and healthy properties. However, this plant is attacked by many pathogenic fungi, which significantly affect fruit production and quality at pre- and post-harvest stages, making chemical applications the most effective but undesirable strategy to control diseases, so far. Alternatively, genetic manipulation employing plant key genes involved in defense, such as members of the NPR-like gene family, has been successful in many crops to improve resistance. The identification and use of the endogenous counterpart genes in the plant of interest (as it is the case of strawberry) is desirable as it would increase the favorable outcome and requires prior knowledge of their defense related function. Using RNAi technology in strawberry, we show that transient silencing of Fragaria ananassa NPR3 members in fruit significantly reduces tissue damage after Colletotrichum acutatum infection whereas the ectopic expression of either FaNPR3.1 or FaNPR3.2 does not have an apparent effect. Furthermore, the ectopic expression of FaNPR3.2 in Arabidopsis thaliana double mutant npr3npr4 reverts the disease resistance phenotype to Pseudomonas syringe to wild type levels. Therefore, our results reveal that members of the strawberry FaNPR3 clade negatively regulate the defense response to pathogens, as do their Arabidopsis AtNPR3/AtNPR4 orthologs. Also, we provide evidence that FaNPR3 members act in strawberry (F. ananassa) as positive regulators of WRKY genes, FaWRKY19 and FaWRKY24, and that in Arabidopsis, FaNPR3.2 negatively regulates its orthologous genes AtNPR3/AtNPR4. We report for the first time the functional characterization of FaNPR3 members in F. ananassa which provides a relevant molecular basis for the improvement of resistance in this species through new breeding technologies.
Keywords:
Fragaria ananassa
; strawberry defense
; strawberry resistance
; Colletotrichum acutatum
; FaNPR3
; NPR-like genes
; AtNPR3
; AtNPR4
; FaWRKY19
; FaWRKY24
1. Introduction
Strawberry (Fragaria spp.) represents a valuable and important food crop, being its fruit highly appreciated by consumers worldwide [1]. Beyond its nutritional interest and sensory attributes, such as texture, color, flavor and aroma, strawberry fruit provides substantial health benefits. This is due to the presence of bioactive compounds with a range of functions like control of blood glucose level, high antioxidant capacity and potential cancer prevention effects [2-5].
Strawberry is vulnerable to a wide diversity of pathogens, including fungi, which significantly affect fruit yield and quality, and Colletotrichum acutatum, is considered a serious fungal pathogen of this crop, causing severe losses in production [6-10]. The lack of pathogen-resistant cultivars is a major challenge for strawberry cultivation, making chemical applications the more widely used strategy to control diseases, endangering ecological and food security [11]. Indeed, classical breeding for resistance is an arduous task in strawberry due to the polygenic and quantitative inherited nature of this trait and to the complexity of the octoploid genome of commercial varieties (Fragaria x ananassa) [12-16]. Therefore, innovative solutions for a more sustainable strawberry production are highly in demand.
Although many environmentally friendly strategies have been applied in strawberry to overcome susceptibility to pathogens [17-22], the genetic modification approaches using key genes that control main defense pathways in plants has led to improve resistance in many crops [23-27] and the heterologous expression of defense genes from other species, including those encoding chitinases, beta-1,3-glucanases and thaumatin II, has been reported in strawberry to successfully enhance resistance [28-35]. More recently, new breeding strategies have emerged as important tools to accelerate crop improvement based on powerful biotechnological techniques such as CRISPR/Cas and novel concepts of cisgenesis and intragenesis-assisted by synthetic biology [36-40], which have provided a valuable and more ecological framework for strawberry disease management than the chemical agents used until now [41]. However, it requires the prior identification and characterization of the appropriate endogenous candidate genes [14,42].
Over the last two decades, studies on the genome structure and the strawberry-pathogen interactions have provided new molecular information on putative components of main defensive pathways in this crop [43-52]. Nevertheless, accurate defense pathways remain yet elusive in strawberry and key defense network elements are still largely unknown and need to be functionally validated. For this reason, our research focuses on delving into endogenous strawberry genes homologous to crucial components of the defense pathways known in other plants, with the aim to characterize and functionally validate valuable genetic elements to accelerate improvement of resistance in this crop.
In this sense, members of the nonexpressor of pathogenesis-related gene family (NPR gene family) have been described as important molecular elements in the immune responses in plants. Indeed, Arabidopsis thaliana (Arabidopsis) AtNPR1 has been identified as a master regulator of salicylic acid (SA)-mediated defense responses [53,54]. AtNPR family differentiates into clade I (NPR1 and NPR2), clade II (NPR3 and NPR4) and clade III (NPR5/BOP2 and NPR6/BOP1) where only members of clade I and II have been associated with defense. Thus, single mutants npr1 and npr2 show increased susceptibility to Pseudomonas syringe (Pst) pathogen infection whereas double mutant npr3npr4 displays enhanced resistance [55]. AtNPR1 protein acts as “sensor” of SA and in response to SA-induced redox changes is released from an inactive oligomeric cytoplasmic state and translocated as a monomeric form to the nucleus where, in association with transcription factors like TGAs, activate defense-related genes [53,56-58]. On the other hand, AtNPR3 and AtNPR4 also interact with TGAs, but opposite to AtNPR1, after binding SA, negatively regulate the immune response. Thus, it has been proposed that AtNPR3/4 may act directly as transcriptional corepressors of defense-related genes [59] and also mediate the degradation of AtNPR1 and defense-related JAZs (Jasmonate-zim represor proteins), acting as adaptors for the ubiquitin ligase (CRL3) complex to enhance defense against pathogens [53,60,61].
The wide relevance of NPR family members in plant defense is evidenced by the fact that orthologs of all three AtNPR clades are conserved in most angiosperm species. Furthermore, the ectopic expression of AtNPR1 in many crop species, including strawberry, enhances broad-spectrum disease resistance, suggesting the existence of closely related defense response mechanisms among these species [62-68]. However, the molecular functions of AtNPR1 and its paralogs are still not fully understood and increased resistance using Arabidopsis AtNPR members often correlates with undesirable fitness costs [53,69-71], which highlights the importance of using functionally well-characterized endogenous NPR orthologs to engineer improved resistance in crops.
Recently, a comprehensive identification and phylogenic analysis of the strawberry NPR-like family has been published [49]. Accordingly, 6 NPR-like members are present in the diploid species (Fragaria vesca) and 23 members in cultivated species (F. ananassa), with each F. vesca member matching 3 to 5 homoalleles in the cultivated species and each clade headed by two Arabidopsis orthologs. Thus, the unique clade I member in F. vesca, FveNPR1, and its 5 homoalleles in F. ananassa, FaNPR1a-e, display high degree of identity among them and the Arabidopsis clade I members, AtNPR1 and AtNPR2. Similarly, the F. vesca clade II members, FveNPR31, FveNPR32, and FveNPR33, and their corresponding F. ananassa homoalleles FaNPR31a-c, FaNPR32a-g, and FaNPR33a-d, show high identity to Arabidopsis clade II members, AtNPR3 and AtNPR4. Finally, clade III FveNPR5 members in the diploid species and its F. ananassa 4 homoalleles, FaNPR5a-d, share high identity with Arabidopsis clade III members, AtNPR5 and AtNPR6. Based on these comparative molecular studies and on the transcriptomic profile of these genes in response to Colletotrichum fructicola, the potential orthologues of AtNPR1 and AtNPR3/4 were identified in strawberry.
The spite these studies, the molecular functions of strawberry NPR-like members in promoting plant survival against pathogens remains uncovered. Just a first approach to functional characterization of the F. vesca gene FvNPR31 by its ectopic expression in a heterologous system such as wild-type Arabidopsis has been published [72]. In the present work, we describe for the first time the functional characterization in the cultivated species F. ananassa of members of clade II NPR-like gene family using RNAi technology and gene complementation studies in Arabidopsis double mutant npr3npr4. Our results show that members of the FaNPR3 clade negatively regulate the defense response to pathogens, as do their Arabidopsis AtNPR3/AtNPR4 orthologs.
2. Results
2.1. Expression Pattern of FaNPR Gene Family after Transient Silencing and Overexpression of the FaNPR3 Members in Strawberry Fruit
To understand the biological role that the FaNPR3 family plays in strawberry defense, we use a transient expression-induced gene silencing and overexpressing approach in fruit. Thus, for silencing, one of the two halves of a fruit was infiltrated with Agrobacterium tumefaciens (agroinfiltrated) carrying the silencing construct (either pFRN::FaNPR3all.RNAi or pB7GWIWG2::FaNPR32.RNAi) and the opposite half of the same fruit with the control construct (either pFRN or pB7GWIWG2). Two days after agroinfitration (dai), the fruit was inoculated in both halves with C. acutatum, (see experimental design in Material and methods). Changes in the expression pattern of all FaNPR genes for every experimental condition were analysed by qRT-PCR.
Results in Figure 1 show the silencing effect of pFRN::FaNPR3all.RNAi agroinfiltration on the expression of all FaNPR3 paralogous genes. Two days after agroinfiltration and throughout the time analyzed, a significant decrease on FaNPR3.1 transcript accumulation in pFRN::FaNPR3all.RNAi samples was detected compared to that of their corresponding pFRN control ones (Figure 1A). The expression of FaNPR3.1 decreased to a lower significant level at 5 dai, which corresponds to 3 days post inoculation with C. acutatum (3 dpi). Figures 1B,C, show that the expression patterns of genes FaNPR3.2 and FaNPR3.3, respectively, were similar to that of FaNPR3.1 and a significant reduction in transcript accumulation was detected for both genes after 2 dai, in fruit half agroinfiltrated with the silencing construct compared with that of its corresponding fruit half control. As for FaNPR3.1, a remarkable and significant decrease in transcript accumulation was observed around 5 dai in FaNPR3.2 and FaNPR3.3 genes. On the contrary, no significant differences in the expression level of other members of the FaNPR gene family such as FaNPR1 and FaNPR5 genes, were detected between fruit control and fruit silenced samples (data not shown). On the other hand, in fruit control samples, a relevant induction in the expression of FaNPR3.2 was detected after C. acutatum inoculation which remained significantly high even after 5 dpi (Figure 1B), compared to that of genes FaNPR3.1 and FaNPR3.3 (Figures 1A,C), which indicates that FaNPR3.2 gene members are preferentially activated in response to pathogen infection. Under these experimental conditions, similar patterns of gene silencing were detected for all the FaNPR3 genes in fruit agroinfiltrated with the silencing construct regardless of whether or not it had been inoculated with C. acutatum (data not shown). All in all, these results reveal that, after fruit agroinfiltration, the pFRN::FaNPR3all.RNAi construct is successfully silencing specifically the three FaNPR3 variants of the NPR-like gene family.
Figure 2 shows the effect of the pB7GWIWG2::FaNPR32.RNAi construct on the expression of FaNPR3 members after transient agroinfiltration of fruit. Thus, a significant reduction in transcript accumulation of FaNPR3.2 was detected after 2 dai in the fruit half sample agroinfiltrated with this construct as compared to its corresponding opposite fruit half agroinfiltrated with the control vector pB7GWIWG2 (Figure 2B). This significant reduction was evident for up to 7 dai (5 dpi). However, no significant changes in transcript accumulation were detected for FaNPR3.1 (Figures 2A) and FaNPR3.3 (Figures 2C) genes at any time sampled when comparing the fruit half silenced with its corresponding fruit half control. Similarly to the results shown in Figure 1B, after C. acutatum inoculation, a relevant induction in the expression of FaNPR3.2 was detected in fruit control samples (Figure 2B) whereas no significant changes in gene expression were detected for FaNPR3.1 and FaNPR3.3 (Figures 2A and C, respectively), which strongly support that FaNPR3.2 is preferentially activated in response to pathogen infection. As aforementioned, under these experimental conditions, no relevant changes in transcript accumulation were found for any of the other gene members of the NPR-like family, FaNPR1 and FaNPR5, and similar patterns of gene silencing were detected for all the FaNPR3 genes when fruit agroinfiltrated was not inoculated with C. acutatum (data not shown). These results evidence that silencing of FaNPR3.2 is mostly achieved in strawberry fruit after agroinfiltration with the pB7GWIWG2::FaNPR32.RNAi construct.
The effect of ectopic overexpression of FaNPR3.1 and FaNPR3.2 genes in strawberry fruit on the transcriptional pattern of all strawberry NPR-like gene family members was also analyzed using the pK7WG2::FaNPR31.OE and pB7WG2::FaNPR32.OE constructs for transient fruit expression, respectively, which carry the inserted genes under the control of the strong promoter CaMV35S. In our experimental conditions, the transient overexpression of either FaNPR3.1 or FaNPR3.2 variants (see Supplementary Figure 1) in strawberry fruit did not significantly alter the transcript accumulation level of any other endogenous member of the FaNPR gene family (data not shown).
2.2. The Silencing of FaNPR3 Genes in Strawberry Fruit Reduced Fruit Tissue Damage after C. acutatum Inoculation
According to the above results, the evaluation of fruit tissue damage, and the comparative study of susceptibility to C. acutatum between the two opposite halves of the same fruit (one agroinfiltrated with either the silencing or the overexpression construct and the other with the corresponding empty vector as control) was accomplished after 6 dai. Overall, no relevant visual differences were observed in external tissue damage of opposite halves of the same fruit, in fruit samples silenced with the pFRN::FaNPR3all.RNAi or pB7GWIWG2::FaNPR32.RNAi silencing construct (upper panels, Figures 3A and 4A, respectively) and fruit samples where FaNPR3.1 or FaNPR3.2 was overexpressed (upper panels, Figures 3B and 4B, respectively). However, a relevant reduction in internal tissue damage was clear within fruit half agroinfiltrated with the silencing construct versus fruit half agroinfiltrated with control vector in both pFRN::FaNPR3all.RNAi (Figure 3A, lower panel) and pB7GWIWG2::FaNPR32.RNAi (Figure 4A, lower panel) silenced fruit samples. When genes FaNPR3.1 and FaNPR3.2 were ectopically overexpressed in strawberry fruit (lower panels of Figures 3B and 4B, respectively), no relevant visual difference in internal tissue damage was observed within fruit half agroinfiltrated with the overexpression construct compared to opposite half agroinfiltrated with the control vector.
A statistical analysis of the internal fruit tissue damage was conducted either in silenced and overexpressed fruit halves and compared to their corresponding control halves. The tissue damage value obtained by normalizing fruit halves transformed with the silencing construct with respect to the corresponding opposite fruit halves transformed with the empty vector were significantly reduced in pFRN::FaNPR3all.RNAi and pB7GWIWG2::FaNPR32.RNAi silenced samples (mean value of 0.6603, and 0.8242, respectively) compared to those obtained when both fruit halves were agroinfiltrated with control constructs (mean value of 1.0556, and 0.9958, respectively) (Figures 3C and 4C, left). However, no significant differences were found in FaNPR3.1 and FaNPR3.2 overexpressed samples (mean value of 1.0737, and 1.0097, respectively) compared to their corresponding control samples (mean of 1.0152, and 1.0327, respectively) (Figures 3C and 4C, right). For external fruit tissue damage, all the ratio values show no significant differences either for silenced or overexpressed samples (data not shown). These results unravel a positive correlation between the silencing of members of the FaNPR3 gene family and an increase in fruit resistance to C. acutatum infection.
2.3. Analysis of Defense Related Genes in Strawberry Fruit Silenced in FaNPR3 Genes
To gain insights into the strawberry defense network associated with members of the FaNPR3 clade, we analyzed the expression profile of several strawberry genes already known to respond to C. acutatum infection such as FaPR1-1(AtPR1 ortholog), FaPR2-1 (AtPR2 ortholog), FaPR5.2 (AT4G11650 ortholog), FaWRKY19 (previously reported as FaWRKY33-2; AtWRKY25/33/26 ortholog), FaWRKY24 (previously reported as FaWRKY1; AtWRKY75 ortholog), FaWRKY41 (previously reported as FaWRKY70-1; AtWRKY54/70 ortholog), and FaWRKY60 (previously reported as FaWRKY70-2; AtWRKY54/70 ortholog). The expression level of FaWRKY19 and FaWRKY24 genes increased in fruit control sample two days after agroinfiltration with either pFRN or pB7GWIWG2) and remained significantly high upon C. acutatum inoculation (Figure 5A and B). However, a significantly reduction in transcript accumulation was detected for both genes, FaWRKY19 and FaWRKY24, in the corresponding opposite fruit half agroinfiltrated with the silencing constructs pFRN::FaNPR3all.RNAi (Figure 5A) and pB7GWIWG2::FaNPR32.RNAi (Figure 5B). Similar patterns of down-regulation were detected for FaWRKY19 and FaWRKY24 genes when the agroinfiltrated fruit was not inoculated with C. acutatum (data not shown). For FaPR1-1, FaPR2-1, FaPR5-2, FaWRKY41, and FaWRKY60 genes, no significant difference was found in their transcript accumulation patterns when comparing fruit agroinfiltrated with the silencing construct (either pFRN::FaNPR3all.RNAi or pB7GWIWG2::FaNPR32.RNAi) vs its corresponding control construct (data not shown).
These results strongly evidence that the expression of genes FaWRKY19 and FaWRKY24 is positively modulated in fruit by FaNPR3 members of the strawberry NPR3-like gene family.
2.4. The Strawberry FaNPR3.2 Gene Complements the Arabidopsis Atnpr3npr4 Double Mutant Disease Resistance Phenotype
To get a deeper insight into the function of the strawberry FaNPR3 family, in parallel to the silencing of the FaNPR3.1 and FaNPR3.2 in strawberry, a heterologous complementation analysis has been conducted in Arabidopsis plants with FaNPR3.2.
Therefore, Arabidopsis transgenic lines were generated by transforming WT and npr3npr4 double mutant plants with pB7WG2::FaNPR32-OE, the DNA cassette driving the expression of the strawberry FaNPR3.2 under the control of a strong promoter. Homozygous plants from each line were selected to perform functional complementation tests by characterizing the expression of the transgene and their disease resistance phenotype upon inoculation with virulent Pseudomonas syringae pathovar tomato strain DC3000 (Pst).
The FaNPR3.2 transcript accumulates abundantly in the leaves of the overexpressing lines npr3npr4::FaNPR3.2 and even at higher levels in WT::FaNPR3.2 (Figure 6A). Interestingly, as shown in Figure 6B, the disease symptoms development at 3, 5 and 7 dpi on the leaves of npr3npr4::FaNPR3.2 and WT::FaNPR3.2 overexpressing transgenic plants was similar to that of WT plants, showing chlorotic symptoms and necrotic lesions, in contrast to the asymptomatic npr3npr4 double mutant plants.
Next, we analyzed the Pst-induced cell death phenotype by trypan blue staining and found that plants overexpressing FaNPR3.2 in the WT and npr3npr4 mutant genetic background showed noticeably enhanced cell death compared to WT and double mutant npr3npr4, respectively (Figure 6C). Indeed, our results show that the presence of endogenous AtNPR3 and AtNPR4 in Arabidopsis together with the strawberry FaNPR3.2 results in cell death spreading across the entire infected leaves of WT::FaNPR3.2 plants, whereas it was more restricted in npr3npr4::FaNPR3.2 leaves. In addition, bacterial growth of virulent Pst was significantly restrained in the absence of AtNPR3 and AtNPR4 (double mutant npr3npr4) at all times tested (3, 5 and 7 dpi), while overexpression of FaNPR3.2 reverted the WT disease phenotype in npr3npr4 plants, displaying similar bacterial density than WT plants (Figure 6D).
Thus, Arabidopsis npr3npr4::FaNPR3.2 plants reverts the Pst-increased resistance phenotype of the double mutant npr3npr4 to wild type levels, in terms of disease symptoms development and bacterial growth.
2.5. Changes in the expression profile of defence-related genes in Arabidopsis
To shed light on the molecular mechanisms underlying FaNPR3.2 complementation of Arabidopsis npr3npr4 double mutant, the expression profile of the typical SA defense pathway marker genes PR1, PR2, and PR5, was analyzed both at basal levels and after infection with Pst and compared among all the Arabidopsis lines. The results show that transcript accumulation of this set of genes was barely detectable in naïve non infected WT plants (Figure 7A), and it increased following infection (Figure 7B), indicative of the activation of the defense response. However, in enhanced resistant npr3npr4 mutant plants the basal transcript level for these genes far exceeded that of WT, in uninfected conditions, and was further strongly induced after Pst infection. Curiously, Arabidopsis plants overexpressing FaNPR3.2, which reverts the WT disease phenotype, displayed remarkably high basal levels of PRs transcript and these marker genes were even further induced upon Pst challenge (Figure 7B).
Ours results reflect a clear uncoupling between the resistance phenotype and the induction of these defense-related markers in Arabidopsis plants overexpressing the strawberry FaNPR3.2 gene.
We next explored whether the enhanced basal level of these classic defense markers observed in lines overexpressing FaNPR3.2 was related with an impaired expression profile of any of the endogenous members of the NPR gene family in Arabidopsis, AtNPR1, AtNPR3 and AtNPR4. Thus, the NPR1 basal transcript level in uninfected plants was similar in WT, npr3npr4 mutant and their corresponding overexpressing FaNPR3.2 lines (Figure 8A), and also upon Pst infection, the AtNPR1 gene expression remained unaltered in all lines tested (Figure 8B). As expected, no AtNPR3 and AtNPR4 transcript was detected in npr3npr4 and npr3npr4::FaNPR3.2 lines. Interestingly, the basal transcript level of both endogenous AtNPR3 and AtNPR4 was significantly reduced in WT::FaNPR3.2 compared to that of WT plants. (Figure 8A). In addition, a significantly stronger induction of AtNPR3 and AtNPR4 genes was observed in WT::FaNPR3.2 overexpressing lines compared to that of WT lines (Figure 8B).
3. Discussion
3.1. Members of the FaNPR3 Clade Negatively Modulate Strawberry Fruit Resistance Against Colletotrichum acutatum
This report primarily focuses on understanding the molecular role that FaNPR3 members of the strawberry NPR-like gene family play in the defense response against pathogens. Thus, we have been quite successful in transiently silencing all members of FaNPR3 clade II in strawberry fruit using A. tumefaciens carrying the silencing construct pFRN::FaNPR3all-RNAi (Figure 1). This construct contains a DNA sequence that was previously predicted to promote highly effective siRNAs matching all FaNPR3 genes in strawberry (F. ananassa) [40]. Furthermore, we have also transiently silenced predominantly FaNPR3.2 homeologs in strawberry fruit (Figure 2) using the pB7GWIWG2::FaNPR32-RNAi silencing construct, highly predicted to produce siRNAs targeting essentially FaNPR3.2 alleles. Transient reduction of either all FaNPR3.1, FaNPR3.2, and FaNPR3.3 transcripts or mainly FaNPR3.2 transcripts, led to a clearly noticeable decrease in disease symptoms and significantly reduced internal strawberry fruit tissue damage after C. acutatum inoculation (Figures 3C and 4C, left). On the contrary, transient overexpression of either FaNPR3.1 or FaNPR3.2 genes did not significantly affect fruit susceptibility to C. acutatum (Figures 3C and 4C, right). According to previously published results and consequently with the agroinfiltration methodology used, no clear difference in the external surface damage was detected between silenced (or overexpressed) and control fruit samples [48].
Interestingly, the reduction of the internal fruit tissue damage was more relevant after the silencing of all members of the FaNPR3 gene family than only FaNPR3.2, evidencing that the FaNPR3 paralogs negatively modulate strawberry fruit defense against C. acutatum. Supporting this idea, it has also been recently shown that ectopic expression of the FvNPRL-1 (the homolog of FaNPR3.1 in F. vesca) in wild-type Arabidopsis led to plants suppressing resistance to Pst, suggesting that FvNPRL-1 could probably function as a negative regulator of the SA-mediated defense in this heterologous system [72]. All in all, these results are consistent with a role of the strawberry FaNPR3 gene family similar to that of their Arabidopsis orthologs, AtNPR3/4, as negative regulators of plant defense [59].
3.2. FaNPR3.2 Negatively Modulates Resistance in Arabidopsis
Analysis of gene knockout mutants constitutes the priority and direct approach in revealing and clarifying gene function. However, so far there are no known strawberry varieties with FaNPR3 knockout genes. Thus, to deepen the knowledge of FaNPR3 members have in the defense response, a heterologous expression system approach was used. We have previously used this procedure to successfully carry out functional studies of strawberry defense-related genes in Arabidopsis [44]. Accordingly, Arabidopsis npr3npr4 double mutant plants, lacking FaNPR3.2 orthologues, that display enhanced resistance against virulent Pst [55] were complemented with its strawberry counterpart. Convincingly, the enhanced disease resistance of Arabidopsis double mutant npr3npr4 to virulent Pst is fully complemented by overexpression of FaNPR3.2, as the resistance phenotype in the Arabidopsis npr3npr4::FaNPR3.2 plants, in terms of disease symptoms development and bacterial growth, resembles that of the wild type plants (Figure 6 A, B and D). Similarly, ectopic expression of this strawberry gene FaNPR3.2 in the WT background resulted in plants (WT::FaNPR3.2) that exhibit no appreciable differences in the resistance phenotype compared with WT plants. These results strongly support the aforementioned role of the strawberry FaNPR3.2 protein, similar to that proposed in Arabidopsis for NPR3/NPR4 in negatively regulating resistance [59]. This is not surprising given the high amino acid sequence similarity and the existence of well conserved motifs between the FaNPR3.2 protein and its Arabidopsis orthologs [49,72,73].
Curiously, overexpression of FaNPR3.2 in the Arabidopsis WT and npr3npr4 background results in increased cell death in response to Pst infection compared to WT and the double mutant npr3npr4 plants, especially when coexisting with Arabidopsis proteins AtNPR3 and AtNPR4 (Figure 6C). However, it does not appear to have an appreciable effect on restricting bacterial growth (Figure 6D). In this sense, our results agree with an additional role for FaNPR3.2 as positive regulator of cell death in Arabidopsis throughout the defense response to Pst, as it has been reported for AtNPR3/4. Consequently, the double mutant npr3npr4 suppresses cell death in response to avirulent pathogen infections [74].
3.3. Silencing of FaNPR3 Members in Strawberry Fruit Downregulates FaWRKY19 and FaWRKY24 Gene Expression
In an attempt to expand our understanding of potential molecular players within the strawberry regulatory defense network downstream FaNPR3, we have monitored the molecular signature of several strawberry SA- and JA- responsive genes known to be up-regulated after C. acutatum infection [42]. Only transcript accumulation of the WRKY genes FaWRKY19 and FaWRKY24 [50] was found to be significantly reduced in fruit after agroinfiltration with both FaNPR3.1 and FaNPR3.2 silencing constructs (Figure 4), and no relevant change was detected in the expression level for the remaining SA- and JA-responsive strawberry genes analyzed (data not shown).
Unexpectedly, and in contrast to the transcriptional co-repressor role described for its Arabidopsis NPR3/4 orthologs [59], our results reveal that in strawberry fruit members of the FaNPR3 clade positively regulate FaWRKY19 and FaWRKY24 genes. Very intriguingly, the concomitant decrease in transcript accumulation of both genes following silencing of FaNPR3 members correlates with the decrease in tissue damage observed in fruit after C. acutatum inoculation. This result is consistent with our previous report that silencing of the FaWRKY24 gene (previously reported as FaWRKY1) in strawberry fruit increases the resistance of this tissue to C. acutatum infection [44]. In fact, WRKY genes are well known to modulate defense responses either positively or negatively [75] and in F. ananassa, down-regulation of other members of the WRKY family such as FaWRKY29 or FaWRKY64 has also been correlated with enhanced resistance against pathogens [52]. Interestingly, the strawberry FaWRKY19 protein shows high degree of amino acid similarity with Arabidopsis AtWRKY25/33/26 proteins [50] and WRKY25/33 orthologs have also been correlated with plant increased resistance. AtWRKY33 protein has been described as a key component of the defense related pathway in plants which exhibits a complex and contradictory functional role. Indeed, silencing of AtWRKY33 gene led to plants with increased susceptibility to necrotrophic fungal pathogens [76] but also, the ectopic expression of this gene caused enhanced susceptibility of plants to the bacterial pathogen Pst [77], suggesting that this AtWRKY33 protein can act either positively or negatively on plant defense depending on the pathogen lifestyle [78]. Consistent with our results in strawberry, down regulation of WRKY33 orthologs in rice led to enhanced resistance to Xanthomonas oryzae [79]. Similarly, AtWRKY25-overexpressing plants display increased bacterial growth and enhanced disease symptoms, while silencing of AtWRKY25 reduced disease symptoms after Pst infection [80]. These reports highlight that different plant species can assemble slightly different molecular mechanisms in response to pathogens, and that components of the plant defense network can exhibit a versatile way of acting depending on the lifestyle of the attacker encountered [78,81].
In strawberry, it remains to be further studied whether FaNPR3 members control downstream defense-response components differently than their orthologs in Arabidopsis and/or whether this protein acts as a molecular switch when the type of pathogen differs. Curiously, based on the structural differences between strawberry NPR-like proteins and their orthologs in Arabidopsis, it appears that functional divergence may occur in strawberry NPRs orthologs [49].
3.4. Resistance to Pseudomonas syringae in Arabidopsis Plants Overexpressing FaNPR3.2 is Uncoupled from PRs Gene Expression
Our results showing the recovery of npr3npr4 mutant plants resistance phenotype to wild type levels, in terms of disease development, cell death and bacterial growth, by ectopic expression of FaNPR3.2 (Figure 6), strongly support that FaNPR3.2 and AtNPR3/AtNPR4 proteins have similar functions in plant defense. To further extend our knowledge about the role of FaNPR3.2, we monitored the expression pattern of the classical PR markers (a hallmark of the SA signaling pathway) [55] in Arabidopsis plants overexpressing FaNPR3.2 and compared them with those in WT and npr3npr4 double mutant plants.
Paradoxically, our data reflect the uncoupling between PRs transcript accumulation and the resistance phenotype. Indeed, overexpression of FaNPR3.2 in both npr3npr4 and WT genetic backgrounds, results in higher constitutive expression levels for this set of PR genes, that are further induced upon Pst infection (Figure 7). Interestingly, the uncoupling between SA-dependent markers expression profiles and the resistance phenotype has been reported previously by our group when complementing the Arabidopsis mutant wrky75 by overexpressing strawberry FaWRKY1 [44]. Furthermore, uncoupling has been previously reported by Zeier et al., 2004 [82], who evidenced that systemic acquire resistance can be executed independently from PRs markers under specific external conditions.
At this point, explaining the uncoupling mechanism driven by FaNPR3.2 would be speculative. However, our results show that AtNPR1 transcript level in absence of infection is similar in WT, npr3npr4 mutant and overexpressing FaNPR3.2 plants (Figure 8A, upper panel) and the level of AtNPR1 induction in Pst-challenged plants does not differ significantly among all the Arabidopsis lines (Figure 8B, upper panel). In contrast, as expected, AtNPR3 and AtNPR4 transcripts are absent in npr3npr4::FaNPR3.2 lines (Figure 8A, middle and lower panels), but interestingly, the transcript basal level of these two Arabidopsis genes is reduced in WT::FaNPR3.2 compared to that of wild-type plants (Figure 8B, middle and lower panels). This last data is very revealing since the FaNPR3.2 overexpression in Arabidopsis correlates with a decrease in the levels of the two well-known negative coregulators of PR expression [59] which could explain the high constitutive level of PRs observed in these plants (Figure 7). Also, AtNPR3/4 have been described as Cullin 3 RING ubiquitin ligases adaptors mediating NPR1 degradation [60]. Thus, the reduced basal level of NPR3/4 in the WT::FaNPR32 overexpressing background is consistent with the reduced degradation of the positive defense regulator AtNPR1, which also could contribute to the higher PRs basal level detected in those plants.
Interestingly, in response to Pst infection, AtNPR3 and AtNPR4 transcripts levels are significantly induced and restored to wild-type levels in FaNPR3.2 overexpressing plants (Figure 8B). This result evidence that the repression that FaNPR3.2 exerts on AtNPR3 and AtNPR4 genes is abolished in response to the pathogen, which suggests that FaNPR3.2 may be sensitive to redox changes and/or to variations in SA levels related to the plant response to infection, like its Arabidopsis orthologs. Besides, it is known that under those conditions, the master regulator NPR1 is not the only substrate for NPR3/4- mediated degradation [53]. NPR3/4 proteins can as well bind and act as adaptors to other key regulatory proteins that mediate immune responses, such as the JA transcriptional repressor JAZ1, target them towards the Cullin 3 RING ubiquitin ligase mediated degradation pathway [60]. In fact, AtNPR3/4- mediated SA-dependent degradation of JAZs makes it possible to tailor an efficient specific defense response against biotrophic pathogens without compromising resistance to necrotrophic pathogens due to activation of the JA-signaling pathway [61]. Our data on the role of FaNPR3.2 in Arabidopsis are reminiscent of that of AtNPR3/4 in the crosstalk between the SA and JA signaling pathways to prioritize one over the other and thus adjust the response against necrotrophic or biotrophic pathogens as necessary [83]. Although more research is needed to clarify this aspect, the results presented here support a dual role for FaNPR3.2 in Arabidopsis as negative regulators of AtNPR3 and AtNPR4 genes and the defense response, and modulators of the extent of the immune response after infection.
On the other hand, our results are not in concordance with those described by Shu et al. [72], reporting that the ectopic expression of FvNPRL-1 in Arabidopsis wild type seedlings suppressed SA mediated PR1 expression. To further investigate this discrepancy, we grew Arabidopsis WT, npr3npr4 double mutant, and their corresponding FaNPR3.2 overexpressing lines on MS plates with or without SA and monitored the PRs expression pattern in those seedlings. As shown in Supplementary Figure 1, all seedling lines analyzed, displayed a similar PR1, PR2 and PR5 expression pattern in response to SA treatment profile to that observed previously in plants upon Pst infection. These results strengthen the positive correlation observed in plants between FaNPR3.2 overexpression and the higher constitutive basal and Pst induced levels of PRs and differ notably from those reported by Shu et al. 2018. Although intriguing, this inconsistency can be attributed to the specific NPR3 alleles overexpressed, FvNPR3.1, from F. vesca, vs FaNPR3.2, from F. ananassa. Indeed, the putative hinge region (LENRV) in the SA- binding core is conserved in FvNPR3.1 protein and in all members in strawberry FaNPR3.1 and FaNPR3.3 clade but not in FaNPR3.2 (FENRV) (Supplementary Figure 3) [49,72]. Also, this region is conserved in AtNPR1 (LENRV) but differs in AtNPR3 and AtNPR4 (LEKRV). It has been described that punctual amino acid differences in this region conduct to slightly differences in SA-binding capacity of AtNPR4 and its interaction with AtNPR1 [73,84].
The plant immune system is a complex mechanism of highly connected molecular network whose components can exhibit diverse functions to refine a successful plant defense against different pathogens. Unraveling the interaction of FaNPR3 members and downstream components of the different defense related pathways in strawberry represents a challenge and an exciting task that will help to engineer broad-spectrum disease resistance in this important crop.
4. Materials and Methods
4.1. Biological Material, Growing Conditions, and Pathogen and Elicitor Treatments
Strawberry fruit (Fragaria × ananassa cv. Primoris) was harvested and grown under field conditions at the Experimental Farm "El Cebollar", IFAPA (Huelva, Spain). Fruits were collected with pedicel (about ten centimeters long) at an early red stage with a degree of pigmentation of about 25% as described [85], sterilized with commercial bleach (1:60 v/v), and kept individually with the pedicel immersed in sterile MS medium (0.25% Murashige Skoog) supplemented with 0,4% (w/v) sucrose, throughout the whole assay period (7 days). The MS medium was replaced every two days to minimize the effect of water stress. Fruits were kept at 25 ⁰C, with a photoperiod of 16h light/8h dark.
Colletotrichum acutatum strain CECT 20240 was used for fruit inoculation experiments. C. acutatum was grown at 20°C with 16/8 photoperiod, on strawberry agar (500 g/L liquefied strawberry berries and 1.5 % bacteriological agar) to improve the infectivity of the pathogen. For pathogen inoculations, starting conidia suspensions (105 conidia/mL) were prepared by diluting a stock of conidia previously obtained by scraping the surface of mycelia grown for 4 weeks, in sterile distilled water (0.03% Tween-80), filtering it through glass wool, and counting cells in a Neubauer Chamber.
The Arabidopsis thaliana plants used in this study belong to in the Columbia (Col-0) ecotype. Wild-type control plants (WT) (Nottingham Arabidopsis Stock Centre, N1093), npr3npr4 double knockout mutant plants (provided by X. Dong, Duke University, Durham, USA), and the transgenic lines generated in this work were germinated and grown under controlled conditions (22ºC, 50% humidity, 9/15 h photoperiod at a light intensity of 125 mol m-2 s-1) as previously described [44].
The Arabidopsis infection experiments were performed with 3- to 4-week-old plants using Pseudomonas syringae pv. tomato virulent strain DC3000 (Pst) provided by Dr. Antonio Molina (CBGP-UPM, Spain).
For the transient ectopic expression and silencing experiments in strawberry fruit, Agrobacterium tumefaciens strain AGL0 was used. For the stable transformation of Arabidopsis plants, the A. tumefaciens strain GV3101 was used.
For the salicylic acid (SA) treatment, Arabidopsis seeds were sterilized for 15 minutes (70% ethanol, 0,05% Triton), and then grown on solid Murashige-Skoog (MS) supplemented with 1% sucrose and 0,4% phytagel, under the controlled conditions mentioned above. Twelve-day-old Arabidopsis seedlings were transferred to equivalent medium containing 200 μM SA or a MS medium without SA (control) and incubated for 24h as described by Shu et al., 2018 [72].
4.2. Plasmid Construction for Silencing and Overexpressing FaNPR3 Genes
For silencing, binary plasmids pFRN (courtesy of Dr. Marten Denekamp, Department of Molecular Cell Biology, University of Utrecht (Netherlands) and pB7GWIWG2.0 vector (VIGS-Plant Systems Biology, Belgium) were used. Standard Invitrogen protocols were used for the cloning steps using gateway technology. For the silencing of all members of FaNPR3.1, -3.2, and -3.3 the 407 bp DNA candidate sequence described in Súnico et al., 2022 [40] was cloned into pCR8/GW/TOPO (Invitrogen) and subsequently transferred to pFRN destination vector to obtain the RNAi silencing construct pFRN::FaNPR3all.RNAi. For the silencing of FaNPR3.2 members, a 535 bp DNA candidate sequence (Supplementary Figure 2) from the 5'UTR region of the FvNPR3.2 gene was amplified by PCR and cloned into pDONR221 (InvitrogenTM), and subsequently transferred to pB7GWIWG2.0 destination vector to obtain the RNAi silencing construct pB7GWIWG2::FaNPR32.RNAi. Candidate sequences were selected and amplified from Fragaria x ananassa genome using www.invivogen.com/sirnawizard/design.php and http://sirna.wi.mit.edu as previously described [40]. The presence of the sense and antisense orientation of the candidate DNA fragments spaced by the CHS intron was confirmed by sequencing. For the ectopic expression of FaNPR3.1, a 1.764 kb cDNA fragment (Supplementary Figure 3) carrying the complete FaNPR3.1 ORF sequence was cloned into pENTR/D/TOPO (InvitrogenTM) and then transferred to pK7WG2.0 destination vector [86]. For the ectopic expression of FaNPR3.2, a 1.785 kb cDNA fragment (Supplementary Figure 4) carrying a complete FaNPR3.2 ORF sequence was cloned into pDONR221 (InvitrogenTM) and then transferred to pB7WG2 destination vector [86]. Both final constructs, pK7WG2::FaNPR31.OE and pB7WG2::FaNPR32.OE, respectively, carry the inserted genes under the control of the CaMV35S promoter. All cDNA fragments were obtained from Fragaria ananassa using total RNA as template. Primer information is provided in Supplementary Table 1. All DNA inserts were sequenced prior further manipulations.
All constructs were introduced into the A. tumefaciens strains indicated above. For the transient ectopic expression and silencing experiments in strawberry fruit, A. tumefaciens strain AGL0 carrying the pK7WG2::FaNPR31.OE, pB7WG2::FaNPR32.OE, pFRN::FaNPR31all.RNAi, and pB7GWIWG2::FaNPR32.RNAi construct, respectively, and their corresponding empty vectors was used. For the stable transformation of Arabidopsis plants, the A. tumefaciens strain GV3101 carrying the pB7WG2::FaNPR32.OE construct was used [87].
4.3. Agroinfiltration of Strawberry Fruit and Experimental Design
To analyze the effect of silencing of FaNPR3 genes on the defense response to C. acutatum inoculations, we confronted a query silencing situation to its non-silenced control within the same fruit, thus avoiding the existing variability among the strawberry fruits used in the assay. Thus, one half of the fruit was infiltrated with A. tumefaciens (agroinfiltration) carrying the silencing/overexpression construct and the opposite half with A. tumefaciens carrying the corresponding empty vector, as a control. Then, we compared the defence response to C. acutatum inoculations between halves of the same fruit. The complete and detailed protocol of this agroinfiltration procedure has been previously described in Higuera et al, 2019 [48]. Briefly, for each condition and gene, a set of 144 strawberry fruits were agroinfiltrated and two days later, 120 of them were inoculated with C. acutatum in both halves, leaving the remaining 24 fruits to analyze the silencing of each gene under no infection conditions. From the 120 inoculated fruits, 24 of them were used for gene expression studies and the 96 remaining fruits were used for tissue damage evaluation and statistical purposes. According to Higuera et al, 2019 [48], the whole assay was repeated twice during strawberry fruiting season for two years. Samples were collected from each of the two halves of three fruits at different times for 7 days after agroinfiltration (7 dai), were quickly frozen with liquid nitrogen and kept at -80ºC, to be used later for the gene expression analysis by RT-qPCR. Likewise, every year, fruit tissue damage was evaluated at 6 dai, which corresponds to 4 days post inoculation with C. acutatum (4 dpi), both in 48 infected fruits, previously agroinfiltrated with the silencing construct in one half of the fruit and the empty vector in the opposite half, and in 48 infected fruits where both halves were agroinfiltrated with the empty vector, as a control for statistical purposes [48]. The effect of ectopic expression of FaNPR3.1 and FaNPR3.2 genes in the response of strawberry fruits to C. acutatum inoculation was evaluated following an identical protocol and experimental design but monitoring tissue damage evaluation after fruit agroinfiltration with A. tumefaciens strains bearing the constructs pK7WG2::FaNPR31.OE and pB7WG2::FaNPR32.OE, respectively, and their corresponding empty vectors.
4.4. Stable Transformation of Arabidopsis Plants
Arabidopsis wild-type and npr3npr4 mutant plants were transformed with the A. tumefaciens GV301 strain containing both, the construct pB7WG2::FaNPR32.OE and its corresponding empty vector using the floral dip procedure [88]. Seeds from the transformed plants (T1), were harvested, sown, and selected based on their resistance to BASTA herbicide (ammonium glufosinate, Bayer CropScience, Ireland) by spraying 2-week-old plants 3 times every 48 hours with a solution containing 1.2% BASTA, 0.05% L-77 silwet (Phytotechnology Laboratory, USA). The resulting progeny (T2) segregating 3:1 (BASTA resistant: susceptible) was selected as carrying a single insertion and homozygous lines (T3) were obtained from at least 10 individuals for each construct and confirmed for homozygosis in the offspring for expression of the transgene by RT-PCR using the specific primers (Supplementary Table S1) and selected for further analyses.
4.5. Arabidopsis Infection Assay with Pseudomonas syringae
Pst growth, plant inoculation and in planta bacterial growth analyses were performed as described previously with 106 CFU mL-1 in 10 mM MgCl2 and were pressure infiltrated into leaves [89]. For monitoring cell death, 12 leaves per genotype and treatment were collected 24 h after inoculation in control and Pst infected leaves and stained using Tripan Blue as previously described previously [90]. All the experiments were repeated at least three times using ten plants per experiment and treatment.
4.6. RNA Extraction and Real-Time qPCR
Total RNA from frozen independent halves of agroinfiltrated strawberry fruits was extracted with the Maxwell ® 16 LEV Plant RNA kit (Promega, Madison, WI, USA), according to the instructions provided by the manufacturer. Arabidopsis total RNA was extracted using 50 mg leaf sample from 12 days old seedlings (4 biological replicates each consisting of seedlings grown in the same plate) or 2-3 weeks old soil grown plants (4 biological replicates each consisting of leaves from 3 plants), using the Invitrap Spin Plant RNA MiniKit (Invitek Molecular, Germany) and contaminating genomic DNA was removed by DNase I (Invitrogen) treatment. Purified RNA was quantified spectrophotometrically on NanoDrop 1000 (Thermo Scientific) and RNA integrity (RIN) was verified using the Agilent 2100 Bioanalyser (Agilent Technologies, Germany). Reverse transcription (RT) was carried out using 250 ng and 2 µg of purified total RNA as template from strawberry and Arabidopsis samples respectively with an RIN value ≥ 8 following manufacturer instructions [iScript cDNA synthesis kit (Bio-Rad)], then 50 ng cDNA were used for RT-qPCRs using SsoAdvancedTM SYBR ® Green Supermix, MyIQ v1.004 and iCycler v3.1 real-time PCR systems (Bio-Rad) and specific primers for each of the gene tested with similar PCR efficiencies. Three technical replicates in the same run and three to four biological replicates in different runs were performed, as described in Encinas-Villarejo et al. 2009 [44]. All primers for qRT-PCR used in this study are listed in Supplementary Table S1.
Relative expression values were determined by the 2−ΔΔCt method [91], using as internal standards the housekeeping genes actine 1 (FaACT1) and elongation factor 1α (FaEF1a) for strawberry [92], and ACTIN2 for Arabidopsis [44]. One-way ANOVA followed by HSD Tukey’s testing was computed using R functions “lm”, “anova” and "TukeyHSD".
4.7. Assessment and Statistical Analysis after Pathogen Infection
For both the silencing and the ectopic expression experiments in strawberry, a phenotypic evaluation was performed to determine fruit tissue damage after 4 days post inoculation (4 dpi) with C. acutatum, corresponding to 6 days after agroinfiltration (6 dai). For statistical purposes, fruits with both halves agroinfiltrated with the empty vector were used as control. Per year, a sample of 48 half fruits for silencing/overexpression, and their corresponding 48 control half fruits, were evaluated for internal damage, according to a scale of 1 to 5 (1 , 0% damage, asymptomatic tissues; 2 , up to 10% damage, weak injury; 3, between 10-25% damage, moderate injury; 4, between 25 and 50% damage, severe injury; 5, more than 50% damage, severely affected fruit), as described in Higuera et al. (2019) [48]. Therefore, the internal damage ratio was calculated by dividing the tissue damage value of the fruit half where the gene was silenced/overexpressed by the tissue damage value corresponding to the opposite half of the same fruit agroinfiltrated with the empty vector. Means and SE were calculated by Fisher's LSD (α = 0.05) and Statistix software (v9.0). A ratio value of 1 indicates no difference between the two halves of the same fruit.
For Arabidopsis, the statistical analysis of the bacterial growth data was performed as in Encinas-Villarejo et al. 2009 [44].
5. Conclusions
We have shown that FaNPR3 members in strawberry act as negative regulators of resistance to C. acutatum and as positive regulators of the FaWRKY19 and FaWRKY24 defense genes. Furthermore, our results in Arabidopsis agree with this role of FaNPR3.2 as negative regulator of defense responses and, in addition, evidence a positive effect of this strawberry gene on the induction of the classic PR resistant marker genes in this heterologous system. The novel results presented in this work highlight strawberry FaNPR3 members as promising candidates for new environmentally friendly breeding technology strategies to accelerate strawberry resistance improvement while minimizing fitness costs.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1; Figure S2, Figure S3, Figure S4; Table S1.
Author Contributions
V.S and J.J.H have contributed equally to this work, carrying out the experimental work, analyzed data, interpretation of results, and writing the draft; F.A.R, has contributed to the strawberry experimental work and statistical analysis of gene expression; I.A.G. and C.J.L.H., have contributed to strawberry susceptibility assays and statistical analysis of fruit tissue damage against C. acutatum; J.M.B. contributed to supervise experimental design, discussion and draft writing; J.L.C and A.M.M are both corresponding authors, have designed methods and experiments, interpreted the results, supervised the work, wrote original draft, and reviewed and edited the final version; J.L.C, was responsible for funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Spanish Government (Ministerio de Ciencia y Tecnología), grants DGICYT AGL2006-13401-C02-02/AGR and AGL2009-13359-C02-02, and the Spanish Regional Government (Junta de Andalucía), grant PO7-FQM-02689.
Acknowledgments
We are grateful to Luis Miranda-Enamorado and the staff of IFAPA (Finca Experimental “El Cebollar”, Huelva, Spain) for the kind supply of strawberry plants and their maintenance in field conditions. Likewise, we thank Dr. José Garrido Gala for his help in collecting strawberry samples, and Fabiola Valverde Bella for helping in the FaNPR3.2 cloning.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Mezzetti, B.; Giampieri, F.; Zhang, Y.-t.; Zhong, C.-f. Status of strawberry breeding programs and cultivation systems in Europe and the rest of the world. Journal of Berry Research 2018, 8, 205. [Google Scholar] [CrossRef]
- Battino, M.; Beekwilder, J.; Denoyes-Rothan, B.; Laimer, M.; McDougall, G.J.; Mezzetti, B. Bioactive compounds in berries relevant to human health. Nutrition Reviews 2009, 67, S145. [Google Scholar] [CrossRef] [PubMed]
- Giampieri, F.; Alvarez-Suarez, J.M.; Cordero, M.D.; Gasparrini, M.; Forbes-Hernandez, T.Y.; Afrin, S.; Santos-Buelga, C.; González-Paramás, A.M.; Astolfi, P.; Rubini, C.; et al. Strawberry consumption improves aging-associated impairments, mitochondrial biogenesis and functionality through the AMP-activated protein kinase signaling cascade. Food Chemistry 2017, 234, 464. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Shi, N.; Afzali, A. Chemopreventive Effects of Strawberry and Black Raspberry on Colorectal Cancer in Inflammatory Bowel Disease. Nutrients 2019, 11, 1261. [Google Scholar] [CrossRef] [PubMed]
- Lucioli, S.; Pastorino, F.; Nota, P.; Ballan, G.; Frattarelli, A.; Fabbri, A.; Forni, C.; Caboni, E. Extracts from Cell Suspension Cultures of Strawberry (Fragaria x ananassa Duch): Cytotoxic Effects on Human Cancer Cells. Molecules 2019, 24, 1738. [Google Scholar] [CrossRef] [PubMed]
- Maas, J.L. Compendium of Strawberry Diseases, 2nd ed.; Maas, J.L., Ed.; APS Press, The American phytopathological Society: St. Paul, 1998. [Google Scholar]
- Prusky, D.; Freeman, S.; Dickman, M.B. Colletotrichum: Host Specificity, Pathology, and Host-pathogen Interaction; APS Press, 2000. [Google Scholar]
- Freeman, S.; Horowitz, S.; Sharon, A. Pathogenic and Nonpathogenic Lifestyles in Colletotrichum acutatum from Strawberry and Other Plants. Phytopathology® 2001, 91, 986. [Google Scholar] [CrossRef] [PubMed]
- Garrido, C.; Carbú, M.; Fernández-Acero, F.; Gonzalez Rodriguez, V.E.; Cantoral, J. New insight in the study of strawberry fungal pathogens. G3-Genes Genomes Genetics 2011, 5, 24–39. [Google Scholar]
- Ji, Y.; Li, X.; Gao, Q.-H.; Geng, C.; Duan, K. Colletotrichum species pathogenic to strawberry: discovery history, global diversity, prevalence in China, and the host range of top two species. Phytopathology Research 2022, 4, 42. [Google Scholar] [CrossRef]
- López-Aranda, J.M.; Domínguez, P.; Miranda, L.; de los Santos, B.; Talavera, M.; Daugovish, O.; Soria, C.; Chamorro, M.; Medina, J.J. Fumigant Use for Strawberry Production in Europe: The Current Landscape and Solutions. International Journal of Fruit Science 2016, 16, 1. [Google Scholar] [CrossRef]
- Shulaev, V.; Sargent, D.J.; Crowhurst, R.N.; Mockler, T.C.; Folkerts, O.; Delcher, A.L.; Jaiswal, P.; Mockaitis, K.; Liston, A.; Mane, S.P.; et al. The genome of woodland strawberry (Fragaria vesca). Nature Genetics 2011, 43, 109. [Google Scholar] [CrossRef]
- Edger, P.P.; Poorten, T.J.; VanBuren, R.; Hardigan, M.A.; Colle, M.; McKain, M.R.; Smith, R.D.; Teresi, S.J.; Nelson, A.D.L.; Wai, C.M.; et al. Origin and evolution of the octoploid strawberry genome. Nature Genetics 2019, 51, 541. [Google Scholar] [CrossRef] [PubMed]
- Folta, K.M.; Barbey, C.R. The strawberry genome: a complicated past and promising future. Horticulture Research 2019, 6. [Google Scholar] [CrossRef] [PubMed]
- Cockerton, H.M.; Karlström, A.; Johnson, A.W.; Li, B.; Stavridou, E.; Hopson, K.J.; Whitehouse, A.B.; Harrison, R.J. Genomic Informed Breeding Strategies for Strawberry Yield and Fruit Quality Traits. Frontiers in Plant Science 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Q.; Edger, P.P.; Xue, L.; Qiong, L.; Lu, J.; Zhang, Y.; Cao, Q.; Yocca, A.E.; Platts, A.E.; Knapp, S.J.; et al. Evolutionary history and pan-genome dynamics of strawberry (Fragaria spp.). Proceedings of the National Academy of Sciences 2021, 118, e2105431118. [Google Scholar] [CrossRef] [PubMed]
- Lazzeri, L.; Baruzzi, G.; Malaguti, L.; Antoniacci, L. Replacing methyl bromide in annual strawberry production with glucosinolate-containing green manure crops. Pest Management Science 2003, 59, 983. [Google Scholar] [CrossRef] [PubMed]
- Samtani, J.B.; Ajwa, H.A.; Weber, J.B.; Browne, G.T.; Klose, S.; Hunzie, J.; Fennimore, S.A. Evaluation of non-fumigant alternatives to methyl bromide for weed control and crop yield in California strawberries (Fragaria ananassa L.). Crop Protection 2011, 30, 45. [Google Scholar] [CrossRef]
- Koron, D.; Sonjak, S.; Regvar, M. Effects of non-chemical soil fumigant treatments on root colonisation with arbuscular mycorrhizal fungi and strawberry fruit production. Crop Protection 2014, 55, 35. [Google Scholar] [CrossRef]
- Ugolini, L.; Martini, C.; Lazzeri, L.; D’Avino, L.; Mari, M. Control of postharvest grey mould (Botrytis cinerea Per.: Fr.) on strawberries by glucosinolate-derived allyl-isothiocyanate treatments. Postharvest Biology and Technology 2014, 90, 34. [Google Scholar] [CrossRef]
- Giovannini, D.; Brandi, F.; Lanteri, A.P.; Lazzeri, L.; Maltoni, M.L.; Matteo, R.; Minuto, A.; Sbrighi, P.; Stagno, F.; Baruzzi, G. Non-Chemical Soil Fumigation for Sustainable Strawberry Production in Southern Italy. Agronomy 2021, 11, 1678. [Google Scholar] [CrossRef]
- Alaphilippe, A.; Casagrande, M.; Varlet, M.; Laurens, F. Friendly Fruit Outcomes: Environment-friendly innovations in strawberry production; hal-03467291; Inrae, 2021. [Google Scholar]
- Datta, A. Genetic engineering for improving quality and productivity of crops. Agriculture & Food Security 2013, 2, 15. [Google Scholar] [CrossRef]
- Qaim, M.; Kouser, S. Genetically modified crops and food security. PloS one 2013, 8, e64879. [Google Scholar] [CrossRef] [PubMed]
- Parisi, C.; Tillie, P.; Rodríguez-Cerezo, E. The global pipeline of GM crops out to 2020. Nature Biotechnoly 2016, 34, 31. [Google Scholar] [CrossRef]
- ISAAA. Global Status of Commercialized Biotech/GM Crops in 2019: Biotech Crops Drive Socio-Economic Development and Sustainable Environment in the New Frontier. 2019.
- Database, I.s.G.A. GM Approval Database. 2024.
- Asao, H.; Nishizawa, Y.; Arai, S.; Sato, T.; Hirai, M.; Yoshida, K.; Shinmyo, A.; Hibi, T. Enhanced Resistance against a Fungal Pathogen Sphaerotheca humuli in Transgenic Strawberry Expressing a Rice Chitinase Gene. Plant Biotechnology 1997, 14, 145–149. [Google Scholar] [CrossRef]
- Asao, H.; Arai, S.; Nishizawa, Y. Environmental risk evaluation of transgenic strawberry expressing a rice chitinase gene. Journal of Bioscience and Bioengineering 2003, 95, 206. [Google Scholar] [CrossRef]
- Chalavi, V.; Tabaeizadeh, Z.; Thibodeau, P. Enhanced Resistance to Verticillium dahliae in Transgenic Strawberry Plants Expressing a Lycopersicon chilense Chitinase Gene. Journal of the American Society for Horticultural Science jashs 2003, 128, 747. [Google Scholar] [CrossRef]
- Schestibratov, K.A.; Dolgov, S.V. Transgenic strawberry plants expressing a thaumatin II gene demonstrate enhanced resistance to Botrytis cinerea. Scientia Horticulturae 2005, 106, 177. [Google Scholar] [CrossRef]
- Vellicce, G.R.; Ricci, J.C.D.; Hernández, L.; Castagnaro, A.P. Enhanced Resistance to Botrytis cinerea Mediated by the Transgenic Expression of the Chitinase Gene ch5B in Strawberry. Transgenic Research 2006, 15, 57. [Google Scholar] [CrossRef]
- Qin, Y.; Teixeira da Silva, J.A.; Zhang, L.; Zhang, S. Transgenic strawberry: State of the art for improved traits. Biotechnology Advances 2008, 26, 219. [Google Scholar] [CrossRef]
- Mercado, J.A.; Barceló, M.; Pliego, C.; Rey, M.; Caballero, J.L.; Muñoz-Blanco, J.; Ruano-Rosa, D.; López-Herrera, C.; de los Santos, B.; Romero-Muñoz, F.; et al. Expression of the β-1,3-glucanase gene bgn13.1 from Trichoderma harzianum in strawberry increases tolerance to crown rot diseases but interferes with plant growth. Transgenic Research 2015, 24, 979. [Google Scholar] [CrossRef]
- Palomo-Ríos, E.; Quesada, M.A.; Matas, A.J.; Pliego-Alfaro, F.; Mercado, J.A. The History and Current Status of Genetic Transformation in Berry Crops. In The Genomes of Rosaceous Berries and Their Wild Relatives; Hytönen, T., Graham, J., Harrison, R., Eds.; Springer International Publishing: Cham, 2018; pp. 139–160. [Google Scholar]
- Rommens, C.M. Intragenic Crop Improvement: Combining the Benefits of Traditional Breeding and Genetic Engineering. Journal of Agricultural and Food Chemistry 2007, 55, 4281. [Google Scholar] [CrossRef]
- Sticklen, M. Transgenic, Cisgenic, Intragenic and Subgenic Crops. Advances Crop Science Technology 2015, 3. [Google Scholar] [CrossRef]
- Thapa, B.; Joshi, T.; Jangid, K.; Basnet, P. Cisgenesis: Genetic engineering introduced spark to traditional breeding methods. Research Environment Life Science 2015, 8, 757. [Google Scholar]
- Limera, C.; Sabbadini, S.; Sweet, J.B.; Mezzetti, B. New Biotechnological Tools for the Genetic Improvement of Major Woody Fruit Species. Frontiers in Plant Science 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Súnico, V.; Higuera, J.J.; Molina-Hidalgo, F.J.; Blanco-Portales, R.; Moyano, E.; Rodríguez-Franco, A.; Muñoz-Blanco, J.; Caballero, J.L. The Intragenesis and Synthetic Biology Approach towards Accelerating Genetic Gains on Strawberry: Development of New Tools to Improve Fruit Quality and Resistance to Pathogens. Plants 2022, 11, 57. [Google Scholar] [CrossRef] [PubMed]
- Cordova, L.G.; Amiri, A.; Peres, N.A. Effectiveness of fungicide treatments following the Strawberry Advisory System for control of Botrytis fruit rot in Florida. Crop Protection 2017, 100, 163. [Google Scholar] [CrossRef]
- Amil-Ruiz, F.; Blanco-Portales, R.; Munoz-Blanco, J.; Caballero, J.L. The Strawberry Plant Defense Mechanism: A Molecular Review. Plant and Cell Physiology 2011, 52, 1873. [Google Scholar] [CrossRef]
- Barbey, C.R.; Lee, S.; Verma, S.; Bird, K.A.; Yocca, A.E.; Edger, P.P.; Knapp, S.J.; Whitaker, V.M.; Folta, K.M. Disease Resistance Genetics and Genomics in Octoploid Strawberry. G3 Genes, Genomes, Genetics 2019, 9, 3315. [Google Scholar] [CrossRef] [PubMed]
- Encinas-Villarejo, S.; Maldonado, A.M.; Amil-Ruiz, F.; de los Santos, B.; Romero, F.; Pliego-Alfaro, F.; Munoz-Blanco, J.; Caballero, J.L. Evidence for a positive regulatory role of strawberry (Fragaria x ananassa) FaWRKY1 and Arabidopsis AtWRKY75 proteins in resistance. Journal of Experimental Botany 2009, 60, 3043. [Google Scholar] [CrossRef] [PubMed]
- Amil-Ruiz, F.; Garrido-Gala, J.; Gadea, J.; Blanco-Portales, R.; Munoz-Merida, A.; Trelles, O.; de los Santos, B.; Arroyo, F.T.; Aguado-Puig, A.; Romero, F.; et al. Partial Activation of SA- and JA-Defensive Pathways in Strawberry upon Colletotrichum acutatum Interaction. Frontiers in Plant Science 2016, 7. [Google Scholar] [CrossRef]
- Amil-Ruiz, F.; Encinas-Villarejo, S.; Munoz-Blanco, J.; Caballero, J.L.; De Los Santos, B.; Romero, F.; Munoz-Mérida, A.; Trelles, O.; Mercado, J.A.; Pliego-Alfaro, F. Distinctive transcriptome response of two strawberry (Fragaria x ananassa) cultivars to Colletotrichum acutatum infection; Acta Horticulturae 2012, 929, 47, WOS:000314786100005.
- Garrido-Gala, J.; Higuera, J.J.; Muñoz-Blanco, J.; Amil-Ruiz, F.; Caballero, J.L. The VQ motif-containing proteins in the diploid and octoploid strawberry. Scientific Reports 2019, 9, 4942. [Google Scholar] [CrossRef]
- Higuera, J.J.; Garrido-Gala, J.; Lekhbou, A.; Arjona-Girona, I.; Amil-Ruiz, F.; Mercado, J.A.; Pliego-Alfaro, F.; Muñoz-Blanco, J.; López-Herrera, C.J.; Caballero, J.L. The Strawberry FaWRKY1 Transcription Factor Negatively Regulates Resistance to Colletotrichum acutatum in Fruit Upon Infection. Frontiers in Plant Science 2019, 10, 17. [Google Scholar] [CrossRef]
- Bai, Y.; Li, Z.; Zhu, J.; Chen, S.; Dong, C.; Gao, Q.; Duan, K. Unraveling NPR-like Family Genes in Fragaria spp. Facilitated to Identify Putative NPR1 and NPR3/4 Orthologues Participating in Strawberry-Colletotrichum fructicola Interaction. Plants 2022, 11, 1589. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Gala, J.; Higuera, J.J.; Rodríguez-Franco, A.; Muñoz-Blanco, J.; Amil-Ruiz, F.; Caballero, J.L. A Comprehensive Study of the WRKY Transcription Factor Family in Strawberry. Plants-Basel 2022, 11, 26. [Google Scholar] [CrossRef] [PubMed]
- Caro, M.D.P.; Pott, D.M.; Tomas-Grau, R.H.; Perato, M.; Albornoz, P.L.; Díaz-Ricci, J.C.; Osorio, S.; Salazar, S.M.; Moschen, S. Defence responses triggered during the plant-pathogen interaction between strawberry (Fragaria x ananassa) and Colletotrichum acutatum. Plant Stress 2023, 10, 100219. [Google Scholar] [CrossRef]
- Lee, M.B.; Han, H.; Lee, S. The role of WRKY transcription factors, FaWRKY29 and FaWRKY64, for regulating Botrytis fruit rot resistance in strawberry (Fragaria × ananassa Duch.). BMC Plant Biology 2023, 23, 420. [Google Scholar] [CrossRef]
- Zhou, P.; Zavaliev, R.; Xiang, Y.; Dong, X. Seeing is believing: Understanding functions of NPR1 and its paralogs in plant immunity through cellular and structural analyses. Current Opinion in Plant Biology 2023, 73, 102352. [Google Scholar] [CrossRef]
- Zavaliev, R.; Dong, X. NPR1, a key immune regulator for plant survival under biotic and abiotic stresses. Molecular Cell 2024, 84, 131. [Google Scholar] [CrossRef]
- Zhang, Y.; Cheng, Y.T.; Qu, N.; Zhao, Q.; Bi, D.; Li, X. Negative regulation of defense responses in Arabidopsis by two NPR1 paralogs. The Plant Journal 2006, 48, 647. [Google Scholar] [CrossRef]
- Després, C.; DeLong, C.; Glaze, S.; Liu, E.; Fobert, P.R. The Arabidopsis NPR1/NIM1 Protein Enhances the DNA Binding Activity of a Subgroup of the TGA Family of bZIP Transcription Factors. The Plant Cell 2000, 12, 279. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fan, W.; Kinkema, M.; Li, X.; Dong, X. Interaction of NPR1 with basic leucine zipper protein transcription factors that bind sequences required for salicylic acid induction of the PR-1 gene. Proceedings of the National Academy of Sciences 1999, 96, 6523. [Google Scholar] [CrossRef]
- Zhou, J.-M.; Trifa, Y.; Silva, H.; Pontier, D.; Lam, E.; Shah, J.; Klessig, D.F. NPR1 Differentially Interacts with Members of the TGA/OBF Family of Transcription Factors That Bind an Element of the PR-1 Gene Required for Induction by Salicylic Acid. Molecular Plant-Microbe Interactions® 2000, 13, 191. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Sun, T.; Ao, K.; Peng, Y.; Zhang, Y.; Li, X.; Zhang, Y. Opposite Roles of Salicylic Acid Receptors NPR1 and NPR3/NPR4 in Transcriptional Regulation of Plant Immunity. Cell 2018, 173, 1454–e1415. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.Q.; Yan, S.; Saleh, A.; Wang, W.; Ruble, J.; Oka, N.; Mohan, R.; Spoel, S.H.; Tada, Y.; Zheng, N.; et al. NPR3 and NPR4 are receptors for the immune signal salicylic acid in plants. Nature 2012, 486, 228. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Sonbol, F.-M.; Huot, B.; Gu, Y.; Withers, J.; Mwimba, M.; Yao, J.; He, S.Y.; Dong, X. Salicylic acid receptors activate jasmonic acid signalling through a non-canonical pathway to promote effector-triggered immunity. Nature Communications 2016, 7, 13099. [Google Scholar] [CrossRef]
- Makandar, R.; Essig, J.S.; Schapaugh, M.A.; Trick, H.N.; Shah, J. Genetically Engineered Resistance to Fusarium Head Blight in Wheat by Expression of Arabidopsis NPR1. Molecular Plant-Microbe Interactions® 2006, 19, 123. [Google Scholar] [CrossRef] [PubMed]
- Malnoy, M.; Jin, Q.; Borejsza-Wysocka, E.E.; He, S.Y.; Aldwinckle, H.S. Overexpression of the Apple MpNPR1 Gene Confers Increased Disease Resistance in Malus × domestica. Molecular Plant-Microbe Interactions® 2007, 20, 1568. [Google Scholar] [CrossRef] [PubMed]
- Meur, G.; Budatha, M.; Srinivasan, T.; Rajesh Kumar, K.R.; Dutta Gupta, A.; Kirti, P.B. Constitutive expression of Arabidopsis NPR1 confers enhanced resistance to the early instars of Spodoptera litura in transgenic tobacco. Physiologia Plantarum 2008, 133, 765. [Google Scholar] [CrossRef] [PubMed]
- Wally, O.; Jayaraj, J.; Punja, Z.K. Broad-spectrum disease resistance to necrotrophic and biotrophic pathogens in transgenic carrots (Daucus carota L.) expressing an Arabidopsis NPR1 gene. Planta 2009, 231, 131. [Google Scholar] [CrossRef]
- Lin, W.-C.; Lu, C.-F.; Wu, J.-W.; Cheng, M.-L.; Lin, Y.-M.; Yang, N.-S.; Black, L.; Green, S.K.; Wang, J.-F.; Cheng, C.-P. Transgenic tomato plants expressing the Arabidopsis NPR1 gene display enhanced resistance to a spectrum of fungal and bacterial diseases. Transgenic Research 2004, 13, 567. [Google Scholar] [CrossRef]
- Zhang, X.; Francis, M.I.; Dawson, W.O.; Graham, J.H.; Orbović, V.; Triplett, E.W.; Mou, Z. Over-expression of the Arabidopsis NPR1 gene in citrus increases resistance to Citrus canker. European Journal of Plant Pathology 2010, 128, 91. [Google Scholar] [CrossRef]
- Silva, K.J.P.; Mahna, N.; Mou, Z.; Folta, K.M. NPR1 as a transgenic crop protection strategy in horticultural species. Horticulture Research 2018, 5, 15. [Google Scholar] [CrossRef] [PubMed]
- Gurr, S.J.; Rushton, P.J. Engineering plants with increased disease resistance: what are we going to express? Trends Biotechnology 2005, 23, 275. [Google Scholar] [CrossRef] [PubMed]
- Chern, M.; Fitzgerald, H.A.; Canlas, P.E.; Navarre, D.A.; Ronald, P.C. Overexpression of a Rice NPR1 Homolog Leads to Constitutive Activation of Defense Response and Hypersensitivity to Light. Molecular Plant-Microbe Interactions® 2005, 18, 511. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Yuan, M.; Ai, C.; Liu, L.; Zhuang, E.; Karapetyan, S.; Wang, S.; Dong, X. uORF-mediated translation allows engineered plant disease resistance without fitness costs. Nature 2017, 545, 491. [Google Scholar] [CrossRef] [PubMed]
- Shu, L.-J.; Liao, J.-Y.; Lin, N.-C.; Chung, C.-L. Identification of a strawberry NPR-like gene involved in negative regulation of the salicylic acid-mediated defense pathway. PLOS ONE 2018, 13, e0205790. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Withers, J.; Li, H.; Zwack, P.J.; Rusnac, D.-V.; Shi, H.; Liu, L.; Yan, S.; Hinds, T.R.; Guttman, M.; et al. Structural basis of salicylic acid perception by Arabidopsis NPR proteins. Nature 2020, 586, 311. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Gao, Y.; Yan, Q.; Chen, W. Salicylic acid promotes autophagy via NPR3 and NPR4 in Arabidopsis senescence and innate immune response. Acta Physiologiae Plantarum 2016, 38, 241. [Google Scholar] [CrossRef]
- Eulgem, T.; Somssich, I.E. Networks of WRKY transcription factors in defense signaling. Special Issue on Biotic Interactions Current Opinion in Plant Biology 2007, 10, 366. [Google Scholar] [CrossRef] [PubMed]
- Birkenbihl, R.P.; Liu, S.; Somssich, I.E. Transcriptional events defining plant immune responses. 38 Biotic interactions 2017 2017, 38, 1. [Google Scholar] [CrossRef]
- Zheng, Z.; Qamar, S.A.; Chen, Z.; Mengiste, T. Arabidopsis WRKY33 transcription factor is required for resistance to necrotrophic fungal pathogens. The Plant Journal 2006, 48, 592. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, F.; Jiang, L.; Chen, C.; Wu, L.; Liu, Z. Different Pathogen Defense Strategies in Arabidopsis: More than Pathogen Recognition. Cells 2018, 7, 252. [Google Scholar] [CrossRef] [PubMed]
- Tao, Z.; Liu, H.; Qiu, D.; Zhou, Y.; Li, X.; Xu, C.; Wang, S. A Pair of Allelic WRKY Genes Play Opposite Roles in Rice-Bacteria Interactions. Plant Physiology 2009, 151, 936. [Google Scholar] [CrossRef]
- Zheng, Z.; Mosher, S.L.; Fan, B.; Klessig, D.F.; Chen, Z. Functional analysis of Arabidopsis WRKY25 transcription factor in plant defense against Pseudomonas syringae. BMC Plant Biology 2007, 7, 2. [Google Scholar] [CrossRef] [PubMed]
- Glazebrook, J. Contrasting Mechanisms of Defense Against Biotrophic and Necrotrophic Pathogens. Annual Review of Phytopathology 2005, 43, 205. [Google Scholar] [CrossRef]
- Zeier, J.; Pink, B.; Mueller, M.J.; Berger, S. Light conditions influence specific defence responses in incompatible plant–pathogen interactions: uncoupling systemic resistance from salicylic acid and PR-1 accumulation. Planta 2004, 219, 673. [Google Scholar] [CrossRef] [PubMed]
- Aerts, N.; Pereira Mendes, M.; Van Wees, S.C.M. Multiple levels of crosstalk in hormone networks regulating plant defense. The Plant Journal 2021, 105, 489. [Google Scholar] [CrossRef]
- Maier, F.; Zwicker, S.; Hückelhoven, A.; Meissner, M.; Funk, J.; Pfitzner, A.J.P.; Pfitzner, U.M. NONEXPRESSOR OF PATHOGENESIS-RELATED PROTEINS1 (NPR1) and some NPR1-related proteins are sensitive to salicylic acid. Molecular Plant Pathology 2011, 12, 73. [Google Scholar] [CrossRef]
- Aharoni, A.; O’Connell, A.P. Gene expression analysis of strawberry achene and receptacle maturation using DNA microarrays. Journal of Experimental Botany 2002, 53, 2073. [Google Scholar] [CrossRef]
- Karimi, M.; Inzé, D.; Depicker, A. GATEWAY vectors for Agrobacterium-mediated plant transformation. Trends Plant Science 2002, 7, 193. [Google Scholar] [CrossRef]
- Koncz, C.; Martini, N.; Mayerhofer, R.; Koncz-Kalman, Z.; Körber, H.; Redei, G.P.; Schell, J. High-frequency T-DNA-mediated gene tagging in plants. Proceedings of the National Academy of Sciences 1989, 86, 8467. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: a simplified method for Agrobacterium -mediated transformation of Arabidopsis thaliana. The Plant Journal 1998, 16, 735. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, A.M.; Doerner, P.; Dixon, R.A.; Lamb, C.J.; Cameron, R.K. A putative lipid transfer protein involved in systemic resistance signalling in Arabidopsis. Nature 2002, 419, 399. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, M.a.E.; Pennell, R.I.; Meijer, P.-J.; Ishikawa, A.; Dixon, R.A.; Lamb, C. Reactive Oxygen Intermediates Mediate a Systemic Signal Network in the Establishment of Plant Immunity. Cell 1998, 92, 773. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT. Methods. 2001, 25, 402. [Google Scholar] [CrossRef]
- Amil-Ruiz, F.; Garrido-Gala, J.; Blanco-Portales, R.; Folta, K.M.; Munoz-Blanco, J.; Caballero, J.L. Identification and Validation of Reference Genes for Transcript Normalization in Strawberry (Fragaria x ananassa) Defense Responses. Plos One 2013, 8. [Google Scholar] [CrossRef]
Figure 1.
Gene expression pattern of FaNPR3 genes in strawberry fruit after agroinfiltration with pFRN::FaNPR3all.RNAi construct. qRT-PCR analysis was accomplished in agroinfiltrated strawberry fruit before (0 and 2 days) and after (3, 5 and 7 days) C. acutatum inoculation (Ca). The time scale is shown as for agroinfiltration fruit samples (dai). For all the genes, arrow and horizontal bar indicate the time of Ca inoculation. Black and grey columns, pFRN (control) and pFRN::FaNPR3all.RNAi (silencing) agroinfiltrations, respectively. Data from all time points are referred to data in time cero, represented as 1 (white column). Bars, mean ± standard error. Note the different scales in the relative expression level axis. Statistical significance was determined by one-way ANOVA. Letters indicate significant differences (p < 0.05) as per HSD Tukey's post-hoc test.
Figure 1.
Gene expression pattern of FaNPR3 genes in strawberry fruit after agroinfiltration with pFRN::FaNPR3all.RNAi construct. qRT-PCR analysis was accomplished in agroinfiltrated strawberry fruit before (0 and 2 days) and after (3, 5 and 7 days) C. acutatum inoculation (Ca). The time scale is shown as for agroinfiltration fruit samples (dai). For all the genes, arrow and horizontal bar indicate the time of Ca inoculation. Black and grey columns, pFRN (control) and pFRN::FaNPR3all.RNAi (silencing) agroinfiltrations, respectively. Data from all time points are referred to data in time cero, represented as 1 (white column). Bars, mean ± standard error. Note the different scales in the relative expression level axis. Statistical significance was determined by one-way ANOVA. Letters indicate significant differences (p < 0.05) as per HSD Tukey's post-hoc test.
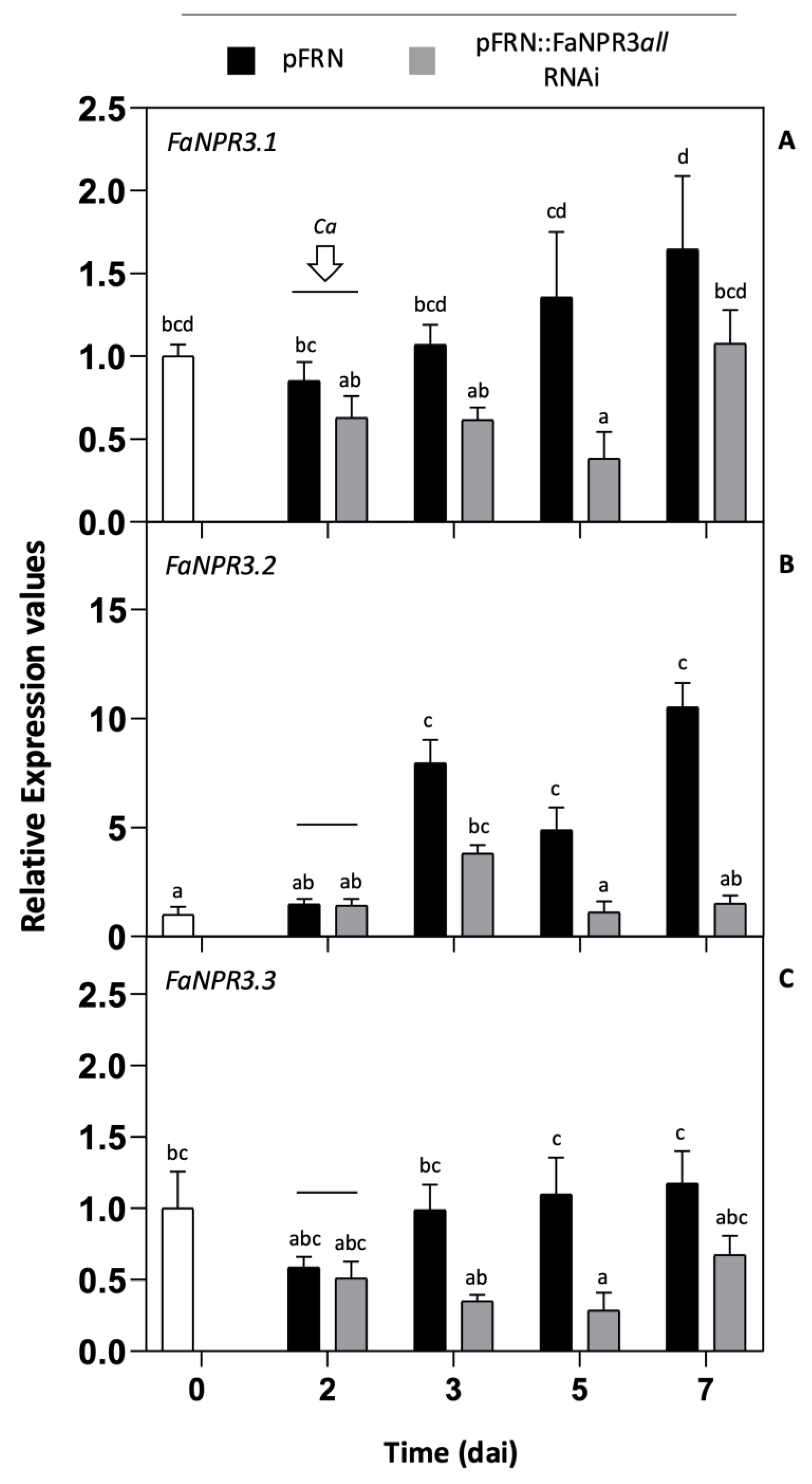
Figure 2.
Gene expression pattern of FaNPR3 genes in strawberry fruit after agroinfiltration with pB7GWIWG2::FaNPR32.RNAi. qRT-PCR analysis was accomplished in agroinfiltrated strawberry fruit before (0 and 2 days) and after (3, 5 and 7 days) C. acutatum inoculation (Ca). The time scale is shown as for agroinfiltration fruit samples (dai). Arrow indicates the time of Ca inoculation. Black and grey columns, pB7GWIWG2 and pB7GWIWG2::FaNPR32.RNAi agroinfiltrations, respectively. Data from all time points are referred to data in time cero, represented as 1 (white column). Bars, mean ± standard error. Note the different scales in the relative expression level axis. Statistical significance was determined by one-way ANOVA. Letters indicate significant differences (p < 0.05) as per HSD Tukey's post-hoc test.
Figure 2.
Gene expression pattern of FaNPR3 genes in strawberry fruit after agroinfiltration with pB7GWIWG2::FaNPR32.RNAi. qRT-PCR analysis was accomplished in agroinfiltrated strawberry fruit before (0 and 2 days) and after (3, 5 and 7 days) C. acutatum inoculation (Ca). The time scale is shown as for agroinfiltration fruit samples (dai). Arrow indicates the time of Ca inoculation. Black and grey columns, pB7GWIWG2 and pB7GWIWG2::FaNPR32.RNAi agroinfiltrations, respectively. Data from all time points are referred to data in time cero, represented as 1 (white column). Bars, mean ± standard error. Note the different scales in the relative expression level axis. Statistical significance was determined by one-way ANOVA. Letters indicate significant differences (p < 0.05) as per HSD Tukey's post-hoc test.
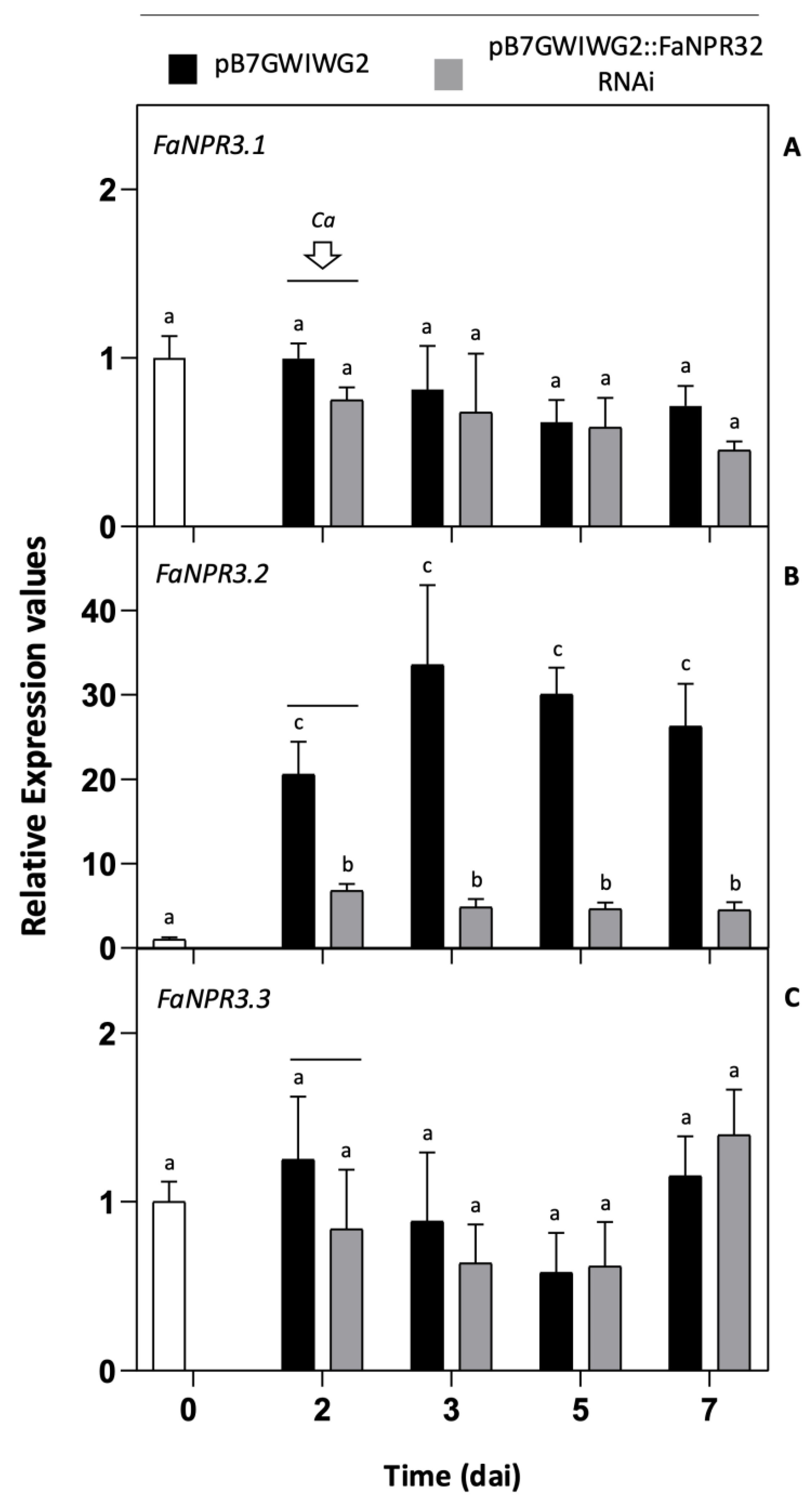
Figure 3.
Silencing effect of pFRN::FaNPR3all.RNAi and ectopic overexpression of FaNPR3.1 in strawberry fruit after C. acutatum infection. (A) and (B) upper panels, external surface disease symptoms on the two agroinfiltrated opposite halves of the same fruit, after silencing and overexpression, respectively. (A) and (B) lower panels, internal tissue damage of the same fruit shown in the corresponding upper panels. pFRN::FaNPR3all.RNAi and pFRN, silencing construct and its corresponding empty vector, as control. pK7::FaNPR31.OE and pK7, overexpression construct and its corresponding empty vector, as control. A relevant fruit is shown for each condition, as an example. (C) Statistically analysis of internal tissue damage ratio of the two opposite halves of the same fruit, according to the 1 to 5 severity scale; striped and plain blue bars, pFRN/pFRN and pFRN/pFRN::FaNPR31-RNAi agroinfiltrated values, respectively; striped and plain red bars, pK7/pK7 and pK7/pK7::FaNPR31-OE agroinfiltrated values, respectively. Data correspond to mean ± SD. Within each bar, means with different letters are significantly different by LSD test at p < 0.05. A ratio value of 1, indicate no differences between opposite halves of the same fruit.
Figure 3.
Silencing effect of pFRN::FaNPR3all.RNAi and ectopic overexpression of FaNPR3.1 in strawberry fruit after C. acutatum infection. (A) and (B) upper panels, external surface disease symptoms on the two agroinfiltrated opposite halves of the same fruit, after silencing and overexpression, respectively. (A) and (B) lower panels, internal tissue damage of the same fruit shown in the corresponding upper panels. pFRN::FaNPR3all.RNAi and pFRN, silencing construct and its corresponding empty vector, as control. pK7::FaNPR31.OE and pK7, overexpression construct and its corresponding empty vector, as control. A relevant fruit is shown for each condition, as an example. (C) Statistically analysis of internal tissue damage ratio of the two opposite halves of the same fruit, according to the 1 to 5 severity scale; striped and plain blue bars, pFRN/pFRN and pFRN/pFRN::FaNPR31-RNAi agroinfiltrated values, respectively; striped and plain red bars, pK7/pK7 and pK7/pK7::FaNPR31-OE agroinfiltrated values, respectively. Data correspond to mean ± SD. Within each bar, means with different letters are significantly different by LSD test at p < 0.05. A ratio value of 1, indicate no differences between opposite halves of the same fruit.
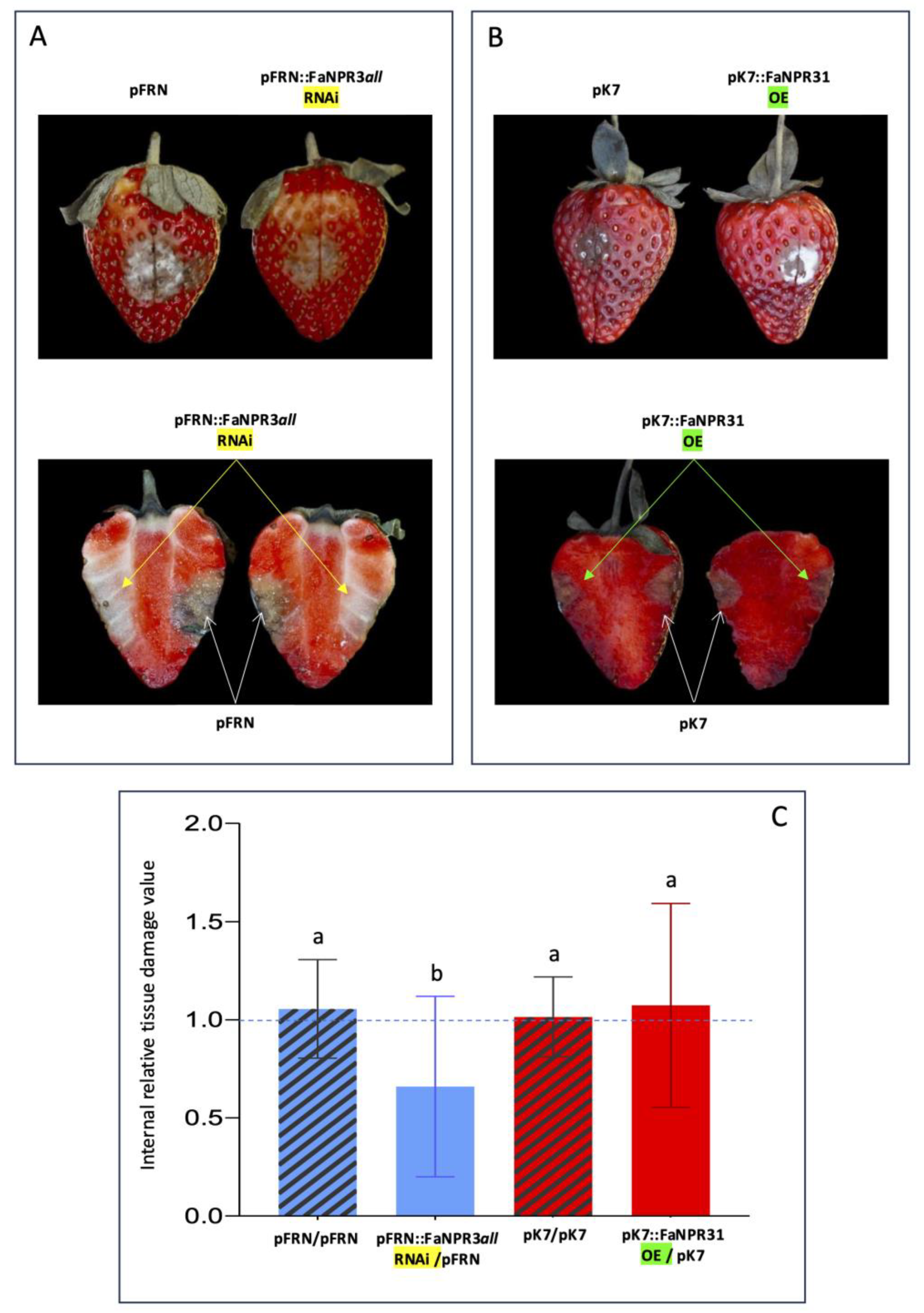
Figure 4.
Silencing effect of pB7GWIWG2::FaNPR32.RNAi and ectopic overexpression of FaNPR3.2 in strawberry fruit after C. acutatum infection. (A) and (B) upper panels, external surface disease symptoms on the two agroinfiltrated opposite halves of the same fruit, after silencing and overexpression, respectively. (A) and (B) lower panels, internal tissue damage, of the same fruit shown in the corresponding upper panels. pB7GWIWG2::FaNPR32.RNAi and pB7GWIWG2, silencing construct and its corresponding empty vector, as control. pB7WG2::FaNPR32.OE and pB7WG2, overexpression construct and its corresponding empty vector, as control. A relevant fruit is shown for each condition, as an example. (C) Statistically analysis of internal tissue damage ratio of the two opposite halves of the same fruit, according to the 1 to 5 severity scale; striped and plain blue bars, pB7GWIWG2/pB7GWIWG2 and pB7GWIWG2/pB7GWIWG2::FaNPR32-RNAi agroinfiltrated values, respectively; striped and plain red bars, pB7WG2/pB7WG2 and pB7WG2/pB7WG2::FaNPR32-OE agroinfiltrated values, respectively. Data correspond to mean ± SD. Within each bar, means with different letters are significantly different by LSD test at p < 0.05. A ratio value of 1, indicate no differences between opposite halves of the same fruit.
Figure 4.
Silencing effect of pB7GWIWG2::FaNPR32.RNAi and ectopic overexpression of FaNPR3.2 in strawberry fruit after C. acutatum infection. (A) and (B) upper panels, external surface disease symptoms on the two agroinfiltrated opposite halves of the same fruit, after silencing and overexpression, respectively. (A) and (B) lower panels, internal tissue damage, of the same fruit shown in the corresponding upper panels. pB7GWIWG2::FaNPR32.RNAi and pB7GWIWG2, silencing construct and its corresponding empty vector, as control. pB7WG2::FaNPR32.OE and pB7WG2, overexpression construct and its corresponding empty vector, as control. A relevant fruit is shown for each condition, as an example. (C) Statistically analysis of internal tissue damage ratio of the two opposite halves of the same fruit, according to the 1 to 5 severity scale; striped and plain blue bars, pB7GWIWG2/pB7GWIWG2 and pB7GWIWG2/pB7GWIWG2::FaNPR32-RNAi agroinfiltrated values, respectively; striped and plain red bars, pB7WG2/pB7WG2 and pB7WG2/pB7WG2::FaNPR32-OE agroinfiltrated values, respectively. Data correspond to mean ± SD. Within each bar, means with different letters are significantly different by LSD test at p < 0.05. A ratio value of 1, indicate no differences between opposite halves of the same fruit.
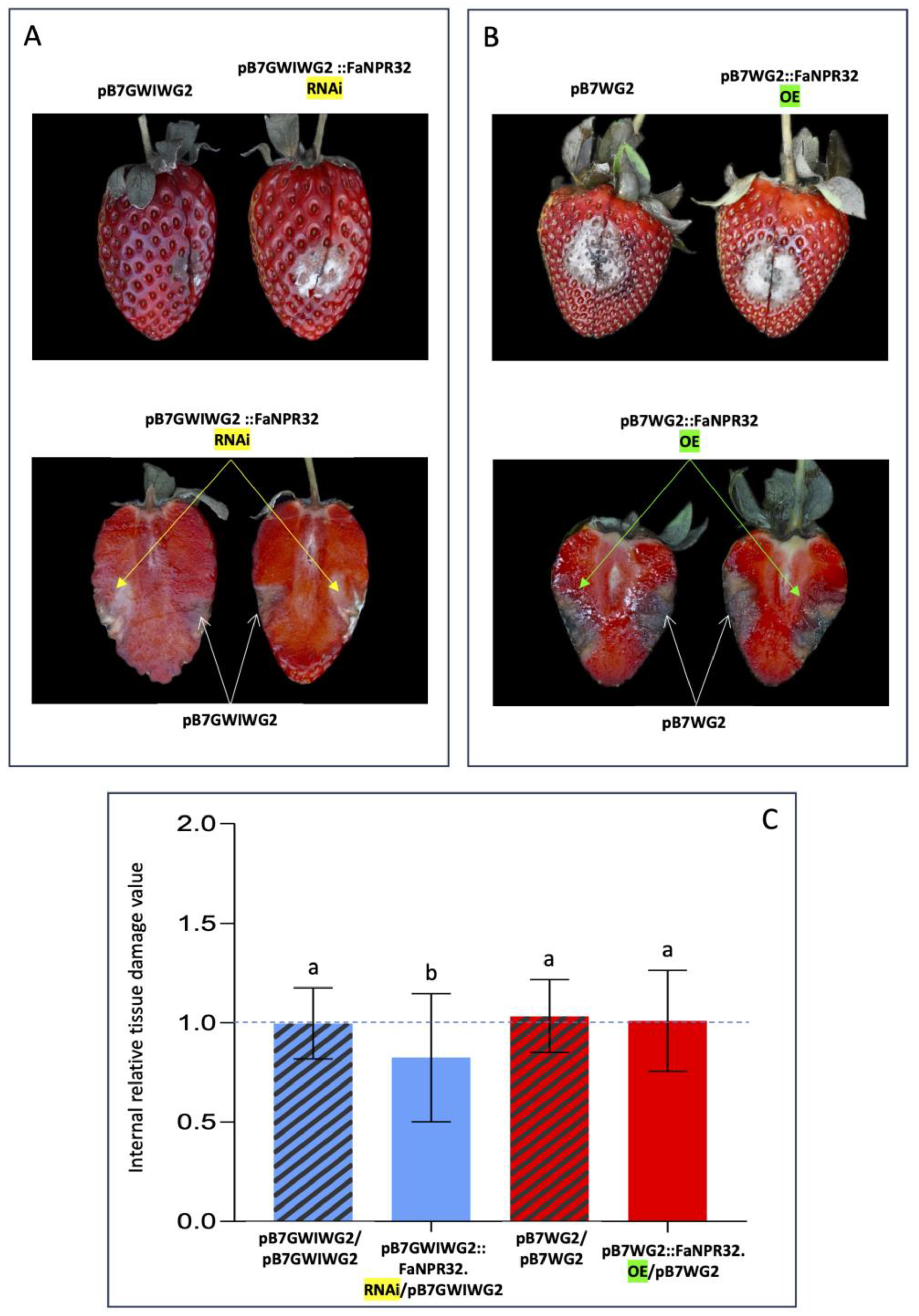
Figure 5.
Silencing effect of FaNPR3 members on the expression of FaWRKY19 and FaWRKY24 genes in strawberry fruit after C. acutatum infection. (A) Silencing effect of pFRN::FaNPR3all.RNAi. (B) Silencing effect pB7GWIWG2::FaNPR32.RNAi. Black columns, the expression of FaWRKY19 and FaWRKY24 genes in half fruit agroinfiltrated with pFRN (A) or pB7GWIWG2 (B) control vectors. Grey columns, the expression of FaWRKY19 and FaWRKY24 genes in half fruit agroinfiltrated with pFRN::FaNPR3all.RNAi (A) or pB7GWIWG2::FaNPR32.RNAi (B). qRT-PCR analysis was accomplished in agroinfiltrated strawberry fruit before (0 and 2 days) and after (3, 5 and 7 days) C. acutatum inoculation (Ca). The time scale is shown as for agroinfiltration fruit samples (dai). Arrow indicates the time of Ca inoculation. Data from all time points are referred to data in time cero, represented as 1 (white column). Bars, mean ± standard error. Note the different scales in the relative expression level axis. Statistical significance was determined by one-way ANOVA. Letters indicate significant differences (p < 0.05) as per HSD Tukey's post-hoc test.
Figure 5.
Silencing effect of FaNPR3 members on the expression of FaWRKY19 and FaWRKY24 genes in strawberry fruit after C. acutatum infection. (A) Silencing effect of pFRN::FaNPR3all.RNAi. (B) Silencing effect pB7GWIWG2::FaNPR32.RNAi. Black columns, the expression of FaWRKY19 and FaWRKY24 genes in half fruit agroinfiltrated with pFRN (A) or pB7GWIWG2 (B) control vectors. Grey columns, the expression of FaWRKY19 and FaWRKY24 genes in half fruit agroinfiltrated with pFRN::FaNPR3all.RNAi (A) or pB7GWIWG2::FaNPR32.RNAi (B). qRT-PCR analysis was accomplished in agroinfiltrated strawberry fruit before (0 and 2 days) and after (3, 5 and 7 days) C. acutatum inoculation (Ca). The time scale is shown as for agroinfiltration fruit samples (dai). Arrow indicates the time of Ca inoculation. Data from all time points are referred to data in time cero, represented as 1 (white column). Bars, mean ± standard error. Note the different scales in the relative expression level axis. Statistical significance was determined by one-way ANOVA. Letters indicate significant differences (p < 0.05) as per HSD Tukey's post-hoc test.
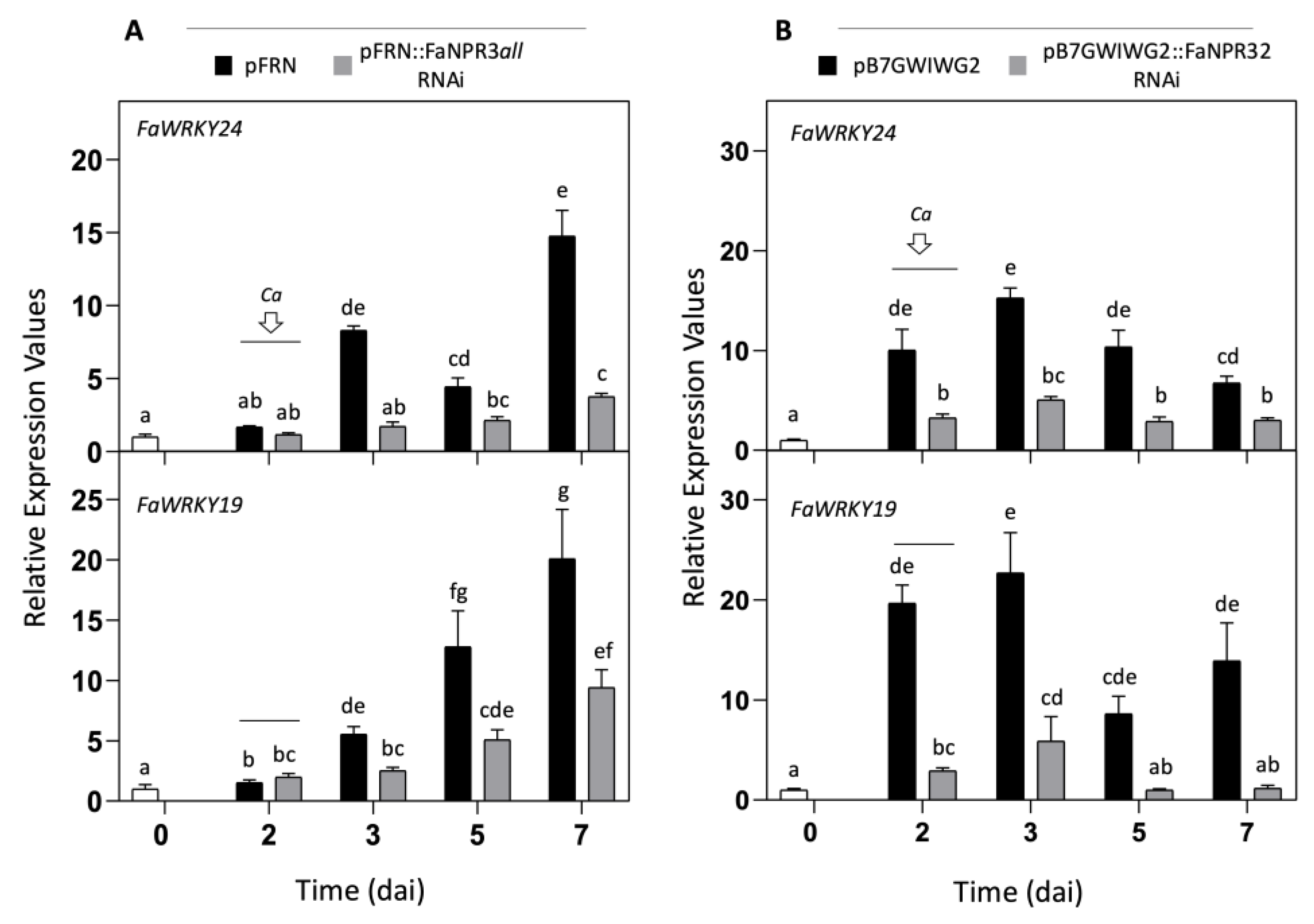
Figure 6.
Characterization of Arabidopsis lines. (A) FaNPR3.2 expression in Arabidopsis WT, npr3npr4 mutant and FaNPR3.2 overexpressing lines npr3npr4::FaNPR32 and WT::FaNPR32 (black, grey, white and striped bars, respectively). Transcript accumulation was monitored by qRT-PCR in non-infected plants as described in the Materials and methods. Expression levels were normalized with respect to the internal control ACTIN2 and displayed relative to the threshold value of the wild type (no expression of FaNPR3.2) that was given a value of 1 for convenience. Note that no amplification of FaNPR3.2 strawberry gene is detected in neither WT nor double mutant npr3npr4 using the specific primers (Supplementary Table S1). Bars represent the mean levels of transcript quantified from three independent biological experiments (±SD). (B-D) Disease resistance phenotype of Arabidopsis lines upon Pst inoculation (B) Symptoms development on leaves 3, 5, and 7 days post inoculation (dpi) (106 CFU mL-1); (C) Trypan blue staining for the detection of cell death 1 dpi (105 CFU mL-1); (D) In planta bacterial growth monitored 3, 5 and 7 day post inoculation (106 CFU mL-1). CFU, colony-forming units. Statistically significant differences are labelled (one-way ANOVA, Tukey’s multiple comparisons test, P < 0.05). Pst were pressure infiltrated into fully expanded mature leaves from 4-5 weeks old Arabidopsis plants. As control, leaves were infiltrated with 10mM MgCl2. The whole experiment was performed three times with similar results.
Figure 6.
Characterization of Arabidopsis lines. (A) FaNPR3.2 expression in Arabidopsis WT, npr3npr4 mutant and FaNPR3.2 overexpressing lines npr3npr4::FaNPR32 and WT::FaNPR32 (black, grey, white and striped bars, respectively). Transcript accumulation was monitored by qRT-PCR in non-infected plants as described in the Materials and methods. Expression levels were normalized with respect to the internal control ACTIN2 and displayed relative to the threshold value of the wild type (no expression of FaNPR3.2) that was given a value of 1 for convenience. Note that no amplification of FaNPR3.2 strawberry gene is detected in neither WT nor double mutant npr3npr4 using the specific primers (Supplementary Table S1). Bars represent the mean levels of transcript quantified from three independent biological experiments (±SD). (B-D) Disease resistance phenotype of Arabidopsis lines upon Pst inoculation (B) Symptoms development on leaves 3, 5, and 7 days post inoculation (dpi) (106 CFU mL-1); (C) Trypan blue staining for the detection of cell death 1 dpi (105 CFU mL-1); (D) In planta bacterial growth monitored 3, 5 and 7 day post inoculation (106 CFU mL-1). CFU, colony-forming units. Statistically significant differences are labelled (one-way ANOVA, Tukey’s multiple comparisons test, P < 0.05). Pst were pressure infiltrated into fully expanded mature leaves from 4-5 weeks old Arabidopsis plants. As control, leaves were infiltrated with 10mM MgCl2. The whole experiment was performed three times with similar results.
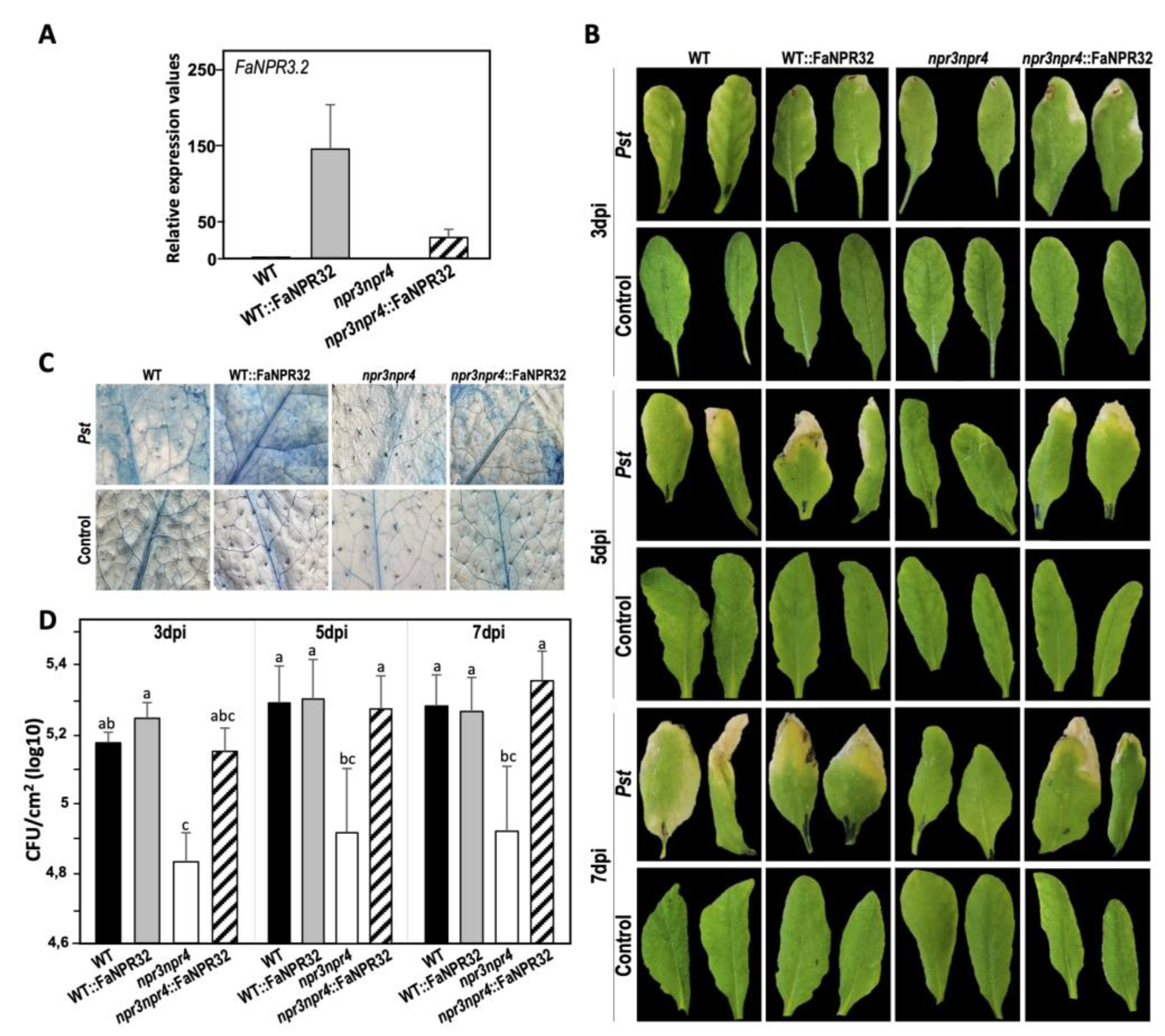
Figure 7.
Expression of the defense-related genes PR1, PR2, and PR5 in Arabidopsis lines. Relative expression level was monitored by qRT-PCR in control (A) and infected plants 1 and 2 days post inoculation (dpi) with Pst (106 CFU mL-1) (B), in WT, WT::FaNPR32, double mutant npr3npr4, and npr3npr4::FaNPR32 (black, grey, white and striped bars, respectively). Expression levels were normalized with respect to the internal control ACTIN2 and displayed relative to the expression in mock-treated wild-type samples (A) or to the expression in mock-treated samples of each line (B) that were given a value of 1. Bars, mean ± standard error. Note the different scales in the relative expression level axis. Statistical significance was determined by one-way ANOVA. Letters indicate significant differences (p < 0.05) as per HSD Tukey's post-hoc test.
Figure 7.
Expression of the defense-related genes PR1, PR2, and PR5 in Arabidopsis lines. Relative expression level was monitored by qRT-PCR in control (A) and infected plants 1 and 2 days post inoculation (dpi) with Pst (106 CFU mL-1) (B), in WT, WT::FaNPR32, double mutant npr3npr4, and npr3npr4::FaNPR32 (black, grey, white and striped bars, respectively). Expression levels were normalized with respect to the internal control ACTIN2 and displayed relative to the expression in mock-treated wild-type samples (A) or to the expression in mock-treated samples of each line (B) that were given a value of 1. Bars, mean ± standard error. Note the different scales in the relative expression level axis. Statistical significance was determined by one-way ANOVA. Letters indicate significant differences (p < 0.05) as per HSD Tukey's post-hoc test.
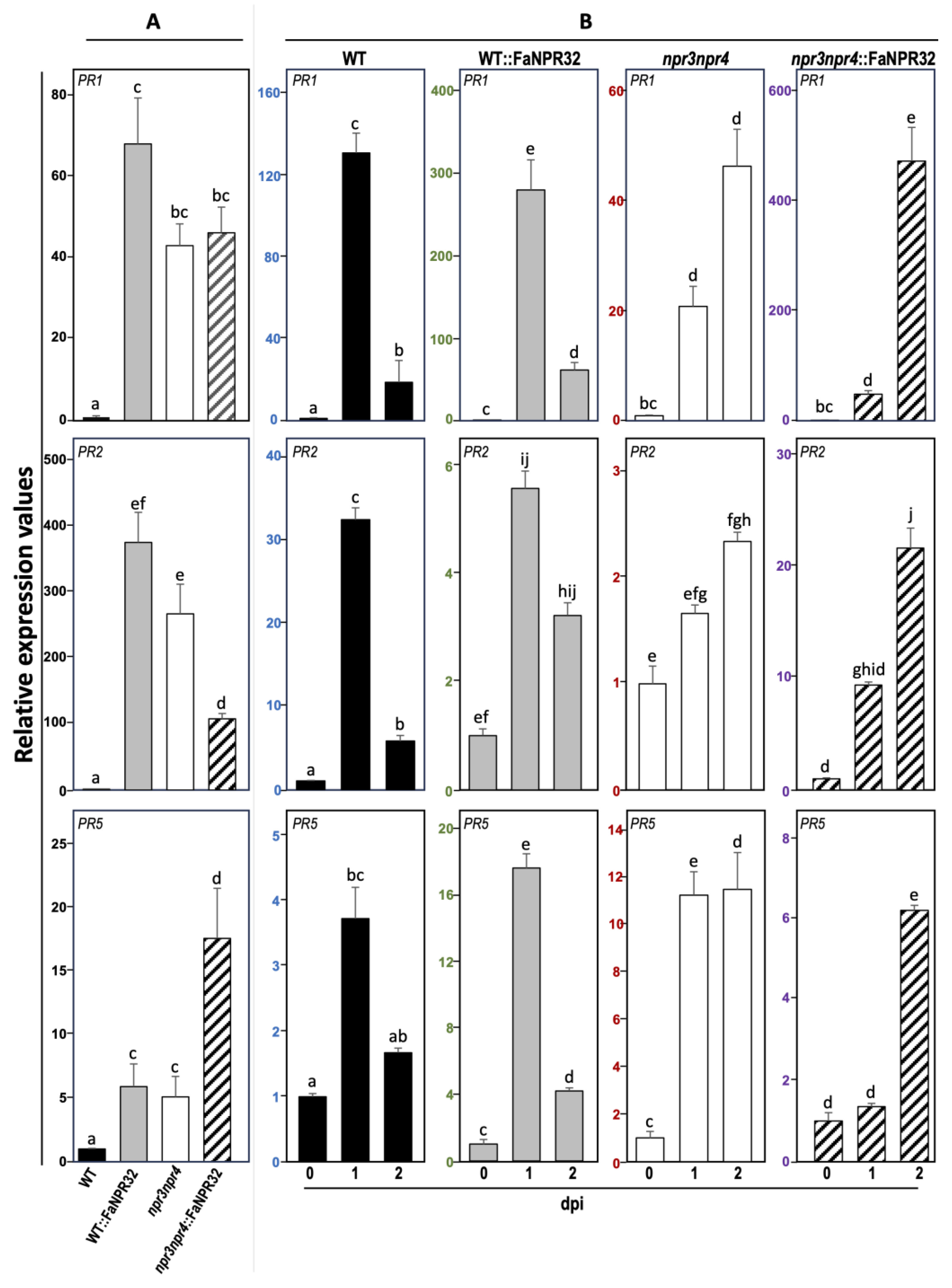
Figure 8.
Expression of the endogenous AtNPR1, AtNPR3 and AtNPR4 genes in Arabidopsis. Relative expression level was monitored as described in Figure 7. Note that the absence of transcript for AtNPR3 and AtNPR4 in the overexpressing lines in the double mutant npr3npr4 background proves that Arabidopsis primers do not amplify strawberry orthologous genes.
Figure 8.
Expression of the endogenous AtNPR1, AtNPR3 and AtNPR4 genes in Arabidopsis. Relative expression level was monitored as described in Figure 7. Note that the absence of transcript for AtNPR3 and AtNPR4 in the overexpressing lines in the double mutant npr3npr4 background proves that Arabidopsis primers do not amplify strawberry orthologous genes.
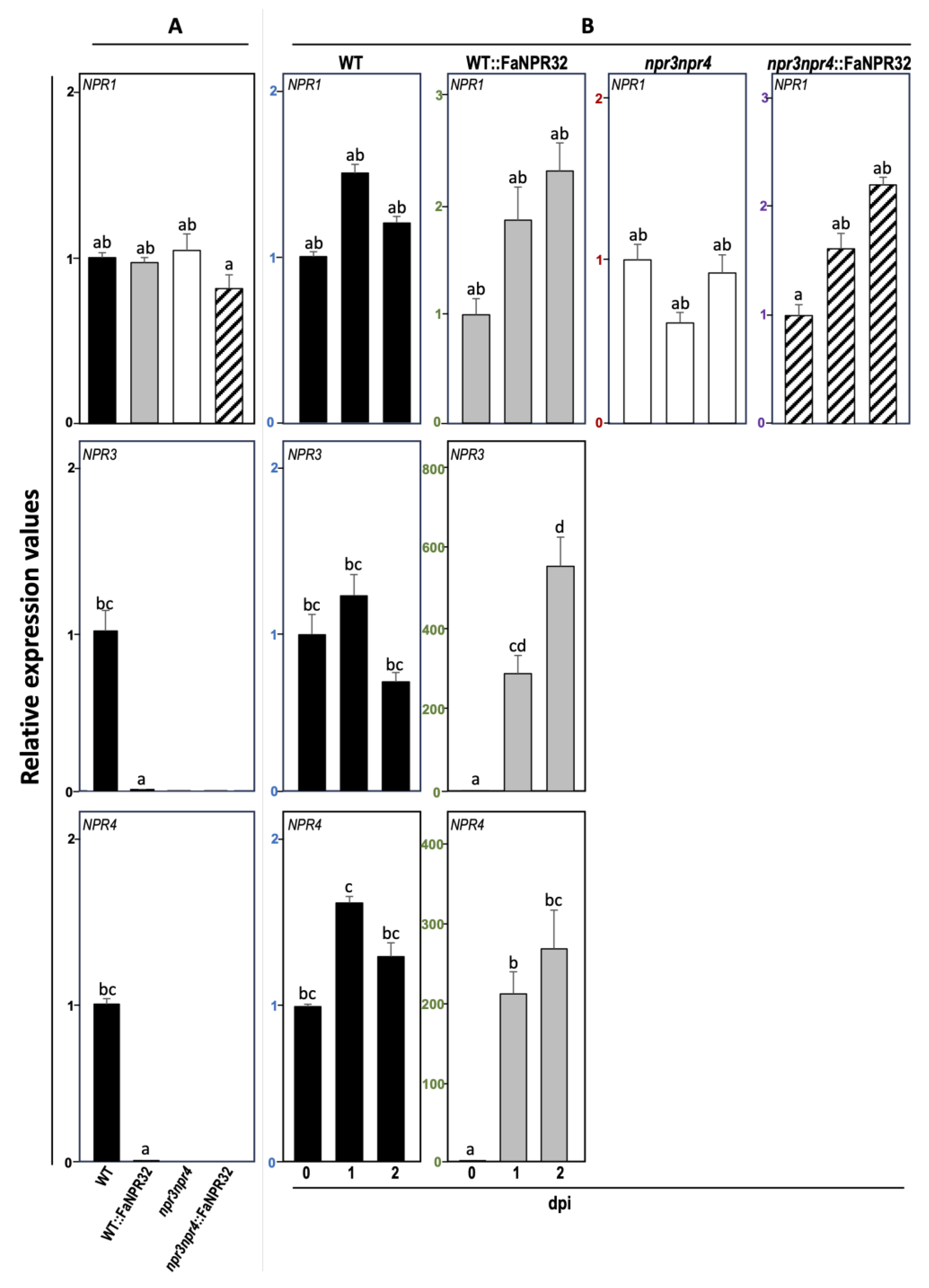
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



