Submitted:
21 July 2024
Posted:
23 July 2024
Read the latest preprint version here
Abstract
The study aimed to evaluate the nutritional value of watermelon plant silage (WPS) and to assess the effect of replacing alfalfa hay (AH) with WPS in a diet formulated for dairy goats. The study used four goats as donors of rumen microbiota to analyze in vitro fermentation parameters. The chemical composition analysis revealed that WPS and AH had similar protein content (21.1 vs. 18.9 g CP/100 g DM), but WPS had lower contents of organic matter (77.4 vs. 89.6 g/100 g DM), NDF (36.3 vs. 49.4 g/100 g DM), ADF (26.4 vs. 34.6 g/100 g DM), LAD (6.40 vs. 8.01 g/100 g DM), and amino acids (716 vs. 874 g AA/kg N) compared to AH. Conversely, WPS had higher fat content (3.09 vs. 1.29 g CF/100 g DM) and different macro- and micromineral content values than AH. In a first in vitro experiment, WPS and AH were incubated independently to compare their fermentation behaviour, indicating that the metabolizable energy (ME) tended to be higher for WPS compared to the AH diet (6.67 vs. 5.72 MJ/kg DM; p = 0.058). WPS fermentation produced lower concentrations of total VFA than AH (66.6 vs. 115 mM; p = 0.011), lower proportions of propionate and valerate (p ≤ 0.022), and higher proportions of isobutyrate (p = 0.001), without altering gas production (GP) kinetics as degradation rate and its potential release. In a second in vitro trial, a formulated goat diet (consisting of commercial concentrate and AH in a 1:1 ratio) was used as a control to assay the impact of AH replacing by WPS at 25% and 50% respectively. Trends were observed in several parameters, such as a linear increase in GP rate (p = 0.088), organic matter digestibility (p = 0.067), and CH4 concentration in the gas produced (p = 0.094) as AH was replaced by WPS. The study concluded that WPS could serve as a viable fodder to replace AH in conventional dairy goat diets while simultaneously reducing agricultural waste and serving as a regenerative model for implementing the circular economy strategy in the affected sectors.
Keywords:
watermelon
; silage
; fermentation
; rumen
; goats
; byproducts
1. Introduction
In recent years, the idea of the circular economy has gained importance to meet the requirements of sustainable livestock farming. In March 2020, the European Commission adopted a new Circular Economy Action Plan to increase the growth of sustainability that includes a “waste and recycling” policy aimed to protect the environment and human health [1]. The main targets of this policy are to stimulate innovation in recycling and improve waste management. These policies become increasingly more important because the world’s population is expected to increase by 33 % over the current population by 2050 [2], which will increase the demand for agricultural products [3], while the natural resources and provision of services from agriculture will not grow [4]. Furthermore, in light of the increasing global population feeding animals with ecological leftovers, instead of human-edible products is currently proposed as a means to increase the food supply [5]. In parallel, livestock is suffering from the negative effects of the warmer climate, which generates the scarcity of forage availability and, in consequence, the increasing of the prices, causing the need for searching alternative strategies to feed animals must be sought for the sector’s economic sustainability [6]. Thus, including by-products in livestock feeding could help address the challenges of this emerging scenario by reducing the cost of feed and improving sustainability on-farm, both from the economic and environmental point of view [7].
Specifically, the biomass generated in the case of the watermelon plant (Citrullus lanatus) was 100 million tons/year in 2019 worldwide [8]. Watermelon fruit and even its seeds have been assayed [9] but, to our knowledge the study of [10], is the only one published on the suitability of using the watermelon plant as forage, and this has been carried out in vitro using sheep as an animal model for meat production systems. However, several studies concluded that there were substantial interspecific differences between goats and sheep in their ability to digest and utilize the nutrients of a number of by-products (e.g. [11,12]). Another aspect to consider is that, the high moisture content may be the main challenge for the practical use and conservation of watermelon plants, because it usually contains certain amounts of discarded fruits left on the plant after harvesting. Nonetheless, ensiling is a technique usually chosen to address this problem and, in addition, it can also enhance the substrate’s nutritional value and increase its palatability [13]. We hypothesized that watermelon plant silage, because of their intrinsic properties, that resemble those of the medium-good quality forage [10], could be beneficial for dairy goats feeding. This work aimed to study the nutritional value of watermelon plant silage (WPS), as well as to assess the impact of replacing alfalfa hay with watermelon plant silage on different rumen in vitro fermentation parameters using goats as donors of inoculum of the rumen microbiota.
2. Materials and Methods
2.1. Animals
Animal procedures and care were conducted in accordance with the Spanish regulations (RD 53/2013) that transpose the European Directive (2010/63/EU) on the protection of animals used in experiments or alternative scientific purposes. Experimental protocols were approved previously by the CSIC Ethical Committee for Experimental Animal Protection and authorized by the regional Andalusian government (procedure 06/07/2023/61) as the competent body in the matter. Four Murciano-Granadina goats (46 ± 4 kg body weight) fitted with rumen cannula were used in this study as inoculum donors. An in vitro non-renewed culture system of ruminal microorganisms [14] inoculated with rumen fluid from goats was carried out for 24 h.
2.2. Diets and Experimental Design
The entire watermelon plant (including branches and leaves and excluding roots) was used, considered as waste in the field after the harvesting of its corresponding fruits, although, in our case, containing small quantities of them that remained as a worthless surplus. Watermelon plant silage (WPS) was prepared by pressing and wrapping with four to six layers of “bale wrap plastic” (25 μm stretch film). This was performed using a bale wrapper machine with a front-loader (Vicon RF 135 Balepack 3D Opticut 23, Brazil). Formic acid (0.45 % of fresh matter) was previously added to enhance the pH drop. Bales were opened after 68 days of ensiling and pH was measured (3.61 ± 0.53). When opened, the colour, aroma and lack of mould were checked as an indicator of the quality of the ensiling process and some samples were squeezed to obtain separate juice fractions for the determination of the concentration and molar proportions of volatile fatty acids (VFA). Representative samples of watermelon plant and WPS were freeze-dried and ground before chemical analysis and in vitro experiments. Two in vitro trials were carried out: in the first trial samples of WPS and alfalfa hay (AH) were incubated independently to compare their fermentative behaviour; in the second trial, the incubation consisted of using a control diet formulated for dairy goats, composed of AH and commercial concentrate (50:50 concentrate to forage ratio), and two experimental diets similar to the control diet but in which an AH fraction (25 and 50 %, respectively) was substituted by WPS. Approximately 500 mL of rumen content was collected from the rumen of four fistulated fasting (12 h) goats. The incubation solution including rumen fluid, from each animal and strained through double cheesecloth layers, and buffer solution (1:3) [15] was used as a biological replicate (n = 4). In vitro incubations were carried out in 120 mL Wheaton bottles containing 400 mg DM of diet and 40 mL of incubation solution. One bottle per buffered inoculum was incubated as blank (without diet). Bottles were incubated at 39 ˚C for 24 h and gas pressure was measured in each bottle at 2, 4, 6, 8, 12, and 24 h using a wide range Pressure Meter (Sper Scientific LTD, Scottdale, AZ, USA). The gas production (GP, mL) in each of these intervals was obtained by coupling a 40 mL capacity syringe to the pressure meter through a three-way stopcock. After 24 h of incubation, bottles were opened, the pH of each bottle was measured and samples (0.8 mL) of each bottle were obtained for VFA quantification as indicated forward.
2.3. Chemical Analyses and Calculations
The dry matter (DM) (method 934.01), organic matter (OM) (method 942.05) and crude fat (CF) (method 920.39) in both WPS and AH were evaluated in triplicate according the AOAC procedures [16]. The nitrogen (N, AOAC method 990.03) was determined by Dumas procedure (Leco TruSpec CN®, St. Joseph, Michigan, USA) to obtain the crude protein (CP = total N g/100 g DM × 6.25). Neutro detergent fibre (NDF) and acid detergent fibre (ADF) were determined following the sequential procedure of Van Soest [17] using the Ankom 220 Fiber Analyzer (ANKOM Technology). The cellulose was solubilized with 72 % sulfuric acid for acid detergent lignin (ADL) analysis. These fibre fractions were expressed exclusive of residual ash.
Heavy metal and mineral contents were determined by atomic spectroscopy inductively coupled plasma-optical emission spectrometry (ICP-OES) analysis. The analyses were carried out in duplicate. The content of some essential and non-essential minerals was measured. The method was based on the addition of a mixture of Milli-Q water and 65 % HNO3 to the sample (1:3), after that, the digestion was carried out in an ultra-wave digester at 220 ˚C for 15 min and cooling to 60 ˚C under high pressure. The product was then transferred to a volumetric flask and diluted with Milli-Q water. Determinations were conducted using an ICP-OES 720-ES system with a sea spray nebulizer and axial torch. Two spectral lines per element were selected, with linearity (correlation coefficient ≥0.995) ensured by utilizing standard solutions. Each analytical result was obtained by calculating the average of the two spectral line readings.
The amino acid (AA) composition of forages was determined by high-performance liquid chromatography using a Waters Lambda-Max LC Spectrophotometer detector (Waters Corporation, USA) using the Waters® Pico-Tag method, which involves precolumn derivatisation with phenylisothiocyanate. Protein hydrolysis was then carried out with 6 N HCl using evacuated tubes at 110 ˚C for 24 h [18].
Rumen fluid samples were analysed for total volatile fatty acids (VFA) as well as their individual molar proportions. Immediately after opening the bottles at 24 h of incubation, a 0.8 mL aliquot of rumen content was mixed with an equal amount of an acid solution consisting of metaphosphoric acid (20 % wt/vol in 0.5 N HCl) and crotonic acid (0.8 g/L, internal standard), then the samples were centrifuged at 2700 g for 20 min. The VFA concentration was determined as described by a gas chromatography (GC) system coupled with a Flame Ionization Detector (Autosystem Perkin-Elmer Cor., Norwalk, Connecticut, USA).
To measure the methane (CH4) production, 4 mL of headspace gas were taken at 24 h at atmospheric pressure and stored in an evacuated tube (Terumo Europe N.V., Leuven, Belgium) at 4 ˚C until the determination by GC using a HP Hewlett 5890 Packard Series II gas chromatograph (Waldbronn, Germany) equipped with a flame ionization detector and with the methodology described by Kheddouma et al. [19]. The gas dynamics generated throughout the incubation was adjusted using the exponential model y = A·(1-e-c·t) described by France et al. [20], where “y” represents the gas production (GP, ml/g); “A” represents the asymptote (ml); “c” represents the rate of gas production (h-1) and “t” represents the time of incubation (h). Feeds chemical composition (CP and OM) and fermentation GP data were used for the estimation of the metabolizable energy (ME) and organic matter digestibility (OMD) following the models described by Menke and Steingass [15]:
ME (MJ/kg DM) = 2.20 + 0.136GP + 0.0057CP + 0.00029 CP2
OMD (g/kg OM) = 148.8 + 8.89GP + 0.448CP + 0.651Ash
2.4. Statistical Analysis
The analysis of data was performed using the SPSS software (IBM Corp. IBM SPSS Statistics for Windows, Version 29.0.0.0 Armonk, New York USA). The comparison between AH and WPS fermentation parameters was analysed by a one-way analysis of variance (ANOVA). The effect of substituting AH with WPS was also analysed with ANOVA, but following an orthogonal contrast. This analysis allowed us to study the effect of the type of forage and the substitution of AH by WPS on the parameters of degradation kinetics related to GP, ME, OMD, pH, VFA and CH4 production. Fisher’s Least Significant Difference LSD test was used to compare mean values and data were considered significant differences at p < 0.05, and p < 0.10 values were considered as trends.
3. Results
3.1. Chemical and Amino Acid Composition
The analysis of VFA in the silage juice showed values of 2.86 ± 0.002 and 0.171 ± 0.005 g/100 g DM of WPS, respectively for acetate and propionate (Mean ± SD), while the values for the rest of the VFA were negligible (< 0.03 g/100 g DM). The chemical and AA composition of WPS and AH are shown in Table 1 and Table 2. Predictably, chemical and AA composition was highly variable between them. The DM content was low, both in WPS and in the starting material used for the ensiling process (15.7 and 14.7 g/100 g FM, respectively), compared to the value obtained for the properly hayed alfalfa (92.8 g/100 g FM) used in this comparative study. The organic matter (OM) varied from 77.4 (WPS) to 89.6 g/100 g DM (AH). Crude protein (CP) was also variable, with values from 21.1 (WPS) to 18.9 g /100 g DM (AH). In the same way, crude fat (CF) varied widely, from 3.09 (WPS) to 1.29 g/100 g DM (AH). Neutral detergent fiber (NDF) content varied, from 36.3 (WPS) to 49.4 g/100 g DM (AH), acid detergent fiber (ADF) from 26.4 (WPS) to 34.9 g/100 g DM (AH) and acid detergent lignin (ADL) from 6.40 (WPS) to 8.01 g/100 g DM (AH). Consequently, hemicellulose varied from 9.91 (WPS) to 14.6 g/100 g DM (AH) and cellulose from 20 (WPS) to 26.9 g/100 g DM (AH). Total carbohydrates changed from 53.2 (WPS) to 69.4 g/100 g DM (AH) and crude energy (CE) from 15.5 (WPS) to 17.6 MJ/kg DM (AH). A large amount of variation between diets was observed in the macrominerals content, with WPS having a higher Ca content (44.7 g/kg) compared to AH (4.40 g/kg). Furthermore, the Mg and P content was higher for WPS (6.07 and 2.91 g/kg, respectively) compared to AH (2.30 and 2.00 g/kg, respectively).
Regarding the AA composition, the content in relation to the DM was lower for WPS compared to AH (151 and 165 g AA/kg DM, respectively) and up to 18 % lower with respect to the total N content (716 and 874 g AA/kg N, respectively). The WPS had a higher content of Glutamic acid, Phenylalanine, Serine, Threonine and Leucine, while the content in Glycine + Histidine, Aspartic acid, Proline, Lysine, Tyrosine, Arginine, Isoleucine and Valine were higher for AH. However, the sum of essential (EAA) and non-essential (NEAA) amino acids resulted balanced and similar for both forages (48.8 and 50.3 g EAA/100 g AA and 51.2 and 49.7 g NEAA/100 g AA, respectively for WPS and AH).
3.2. The Effect of Different Forages on Rumen Fermentation Parameters
The results of the fermentation parameters obtained from the first in vitro trial with AH and WPS incubated independently are shown in Table 3. In the present study, we found no significant differences (p ≥ 0.679) between the diets in the GP, the asymptotic value of total gas production (A, mL) and in the rate of degradability (c, h-1). However, we found differences in pH increasing with the HA diet compared to WPS (7.04 and 7.00, respectively; p = 0.017). Furthermore, the metabolisable energy (ME) tended to be higher for WPS compared to the AH diet (6.67 and 5.72 MJ/kg DM, respectively; p = 0.058). The organic matter digestibility (OMD) was also numerically higher for the WPS than for the AH diet, but the difference was not significant (471 and 410 g/kg, respectively; p = 0.126).
The CH4 production was not affected by the inclusion of WPS compared to the AH (47.1 and 43.4 µl/ml GP, respectively; p = 0.287).
According to the total VFA measurement, the AH was more fermentable compared to WPS (115 and 66.6, respectively; p = 0.011). In terms of VFA molar proportions, no differences were detected in the acetate, whereas the propionate decreased (17.3 and 19.0 %, respectively for WPS and AH; p = 0.022) and the isobutyrate increased for WPS compared to AH (1.78 and 1.17 %, respectively for WPS and AH; p = 0.001). The valerate decreased with the WPS incubation compared to the AH diet (1.35 and 1.57 %; p = 0.014). We did not find significant differences in the molar proportions of butyrate and isovalerate (p ≥ 0.183). Consequently, the acetate:propionate ratio tended to be higher in the WPS fermentation compared to the AH (4.08 and 3.51 %, respectively; p = 0.057).
The graph of the accumulated GP for both WPS and AH forages is represented in Figure 1. The WPS had the highest cumulative GP compared to AH, regarding the asymptote of the kinetics curve. Furthermore, the slope was similar for both forages but after 20 hours of fermentation, the WPS diets exceed the GP threshold of the AH.
3.3. Effects of Replacing Alfalfa Hay with Watermelon Plant Silage on the Ruminal Fermentation Parameters of the Experimental Diet.
The results of the second trial, in which the replacement of AH by WPS was evaluated, are depicted in Table 4. No significant differences were found in the parameters of rumen fermentation kinetics nor the total VFA or their molar proportions when 25 or 50 % of AH was replaced by WPS. However, we did find trends in different parameters such as an increase in the degradability rate (c, h-1; p = 0.088), the OMD, (p = 0.067) and the CH4 concentration in the gas produced (p = 0.094). Such trends seemed to follow a linear response to the replacement level of AH by WPS performed.
The effect of replacing different percentages of AH with WPS in GP is graphically represented in Figure 2. Diets with 25 and 50 % substitution of AH with WPS showed a slightly lower GP asymptote, while the slope until 20 h of incubation resulted quite similar among treatments.
4. Discussion
The use of plant silages as an alternative source in animal feed has been considered during the last decades, showing a wide variation in its chemical composition due to several factors such as plant species, climatic conditions during cultivation and treatments to improve ensiling processing. The principle of ensiling is based on preserving green fodder under anaerobic conditions to support the growth of lactic acid-producing bacteria, which generate lactic acid and cause a decrease in the pH of the conserved material [21]. Several studies have shown promising results when using plant silages to feed ruminants [22,23], but is scarce the scientific literature on the suitability and adequacy of the watermelon plant ensiling, especially considering the entire plant and not only surplus fruits. The silage of a similar plant species such as melon (Cucumis melo L.) under different technical conditions, has been recently characterized from a physico-chemical and microbiological approach but without studying the rumen fermentation of this product [24]. Mean values of 3 % of DM in acetate and 0.5 % in propionate have been summarized by Kung et al. [25] as the upper limit in silages that have had adequate fermentation and without proliferation of clostridia or enterobacteria. In our case, these values were 2.86 and 0.171 % of DM, respectively for acetic and propionic. These same authors also argued that high concentrations of these acids are related to a low voluntary feed intake of silage in cows, although it may not be directly attributable to these compounds, rather because they can be considered indicators of an inadequate ensiling process or an incorrect conservation or storage. In any case, VFA concentrations are proportional to the moisture content of the silage, a fact that in the case of WPS (almost 85 %, conferred by the presence of watermelon fruits in the pre-silage mixture) was much higher than the average of the studies reviewed in the cited work (75 % upper limit)
Similar to what Lin et al. [26] found when analyzing the impact of silage on the chemical composition of the AH, the nutrient composition of the WPS hardly changed concerning what the plant had before the ensiling process. They reported a significant reduction in water-soluble carbohydrates, probably for a reason that can explain the decrease in hemicellulose in exchange for the increase in cellulose (in a magnitude close to 20 %) observed in our study. The main explanation could be that hemicellulose is susceptible to being used to a greater extent by the microorganisms involved in the fermentation that occurred during the ensiling process. Some authors reported variations in the saturation of fatty acids due to the ensiling, but there is no available information on the impact of this process on the plant CF content that could explain the increase (by more than 100 %) of this nutrient observed in the WPS. However, Hassan et al. [10], reported a WSF CF content similar to that in our product (2.80 vs. 3.09 g/100 g DM), whereas Ibrahim et al. [27] found a range of CF content in the intact watermelon plant (from 1.19 to 1.82 g/100 g DM) which covered the value of our observation (1.40 g/100 g DM) before ensiling.
Alfalfa stands out among forage crops due to its high levels of CP and energy, which minimize the necessity for additional supplements in feed and makes it ideal for inclusion in the diets of high-yield dairy ruminants. The comparison between the composition of WPS and AH used in our trial reflected that the former showed a higher content of minerals, CP and CF, while AH contained higher NDF, ADF and ADL proportions. Considering this, and that the NDF content (nearly 50%) of the AH used in our study, it can be regarded as a medium-quality forage legume, while WPS could be expected to offer a higher potential for providing ME. The ADF represents the percentage of highly indigestible and slowly digestible components in a feed or forage, including cellulose, lignin, pectin, and ash. A lower ADF value in WPS, approximately 25% less than in AH, suggests that this forage might be more digestible and thus suitable for dairy goats. The lower OM and carbohydrates content in WPS likely resulted from the presence of earth in the plant samples obtained, suggesting that the collection method used in this experiment may have been suboptimal. This same reason would explain the very high calcium (Ca) content since the WPS cultivation soil in this study was made up of quaternary deposits of the Ca luvisol type. Minerals are necessary for metabolic processes in small ruminants and Ca is required for lactation, maintenance and growth in small ruminants. It has been reported to be between 1.2 to 2.6 g/kg DM [28] and, while AH is considered rich in Ca source (4.40 g Ca/kg DM), the concentration of this element in WSP exceeded almost twenty times the upper threshold of the requirements range (44.7 g Ca/kg DM WPS). However, if the Ca is in the form of carbonates of soil contamination, the bioavailability of this element may be lower and also, it could depress the absorption of Fe [29]. The WPS showed higher concentrations of K, Mg and Na, and lower of Fe, Al, Zn, Ti, As, B, Si and Sr than AH. In comparison with AH and contrarily to described by Hassan et al. [10], the low heavy metals content of WPS draws attention, which would constitute it as a safe fodder for animals.
The AA analysis plays a crucial role in examining the make up of proteins, as well as in studying the constituents of foods and animal feeds and their potential to cover the protein requirements of the different animal species. Moreover, EAA is defined as AA whose carbon skeletons are not synthesized de novo by animal, cells or AA that are insufficiently synthesized de novo by animal cells relative to metabolic needs [30], and although many species of rumen bacteria that are capable of de novo synthesizing them, ruminants’ diets must provide sufficient protein and EAA when high rates of growth and lactation are required [31]. Ibrahim et al. [32] studied the AA composition of different parts of the watermelon plant (Citrullus vulgaris) and their results regarding proportions of AA in stem and leaves can be considered equivalent to those found in our study, including the observation related to a proportion higher in NEAA than in EAA. They also concluded that most of the AA values of the watermelon plant are comparable with those of most vegetable proteins, an observation that can also be extended to the comparison of WPS with AH performed in our study. Nevertheless, it should be noted that the ratio of AA-N in WPS was 18 % lower than in AH, probably indicating a worse protein value for WPS, although this fact could partially be compensated by a higher total N content of more than 11.5 % for this fooder.
The in vitro gas technique, based on volume and pressure measurements, has been used for decades to assess the degree of fermentation of diets in ruminants [33] and monogastric animals [34]. Using the in vitro gas technique allows to reduce the cost, time and use of in vivo experiments [35] and is considered a replacement and reduction system to implement two of the three Rs (replacement, reduction and refinement) principles to address the ethical protection of animal experimentation.
When evaluating the quality of silage, it is important to consider the rumen fermentation parameters, which adequacy indicates that the silage can be conveniently degraded by the animal and supplies the required nutrients. In the present study, in terms of rumen fermentation dynamics, the WPS was shown to lead a fermentative process similar to AH regarding the rate (c), the asymptotic limit (A) of GP, indicating the suitability of its inclusion in the diet of goats. Although the range of ME values for medium quality AH reported in the literature, determined through in vivo tests, typically falls between 8 and 9 MJ/kg DM, and differs from the 5.72 MJ/kg DM found in this study, it is important to recognize that the in vitro values obtained are still valid for comparing the energy availability of AH and WPS. This allows for an assessment of the suitability of WPS as a potential substitute for AH. Moreover, the ME calculations following the model proposed by Menke and Steingass [15], which considered the GP and the chemical composition (CP and OM) of the substrates, revealed a value favourable to WPS in comparison to AH. Thus, the underestimation obtained in the calculation of ME was due to the low GP records, which resulted from a lower ruminal inoculum to buffer ratio in our study (1:3) compared to that used in the work of Menke & Steingass (1:2). If we apply the same proportion of our underestimation of AH in reverse, the actual ME of the WPS could fall within the range of 9.3 to 10.5 MJ/kg DM. This would lead us to classify it as a good quality forage. The significant variations in pH did not represent relevant biological importance because of the small deviations observed between the diets, since the culture media were suitably buffered, and because these values were adequate to allow rumen microbial activity.
The end products of dietary carbohydrate fermentation in ruminants are VFA, mainly acetate, propionate and butyrate [36]. All of them are the main source of energy in ruminants, accounting for more than 70 % of total metabolic energy [36]. Acetate and propionate are essential for fat synthesis and gluconeogenesis [37]. In the present study, when comparing WPS to AH in vitro fermentation, we found that WPS fermentation promoted similar acetate and butyrate proportions to the AH but lower propionate rate. The acetate: propionate ratio is used to characterize the type of fermentation that has predominated, when the ratio is lower, it is considered more efficient [34]. In our trial, we found that WPS showed a higher ratio compared to the AH. Considering the total VFA production, the energy potential showed by the rumen fermentation of the WPS was barely 60 % of the AH. Total VFA production data are usually corrected to express them based on degraded OM, but, although the content of this nutrient was lower for the WPS, its OMD was numerically higher, so the VFA production potential was not compensated by this calculation (24.4 vs. 41.7 mmol/g degraded OM, respectively for WPS and AH). If we examine the GP and its CH4 proportion, along with the total VFA production from the fermentation of WPS, the data appear to be controversial and difficult to interpret. In the case of isobutyrate and isovalerate, they increased with WPS compared to AH. These VFAs are iso-acids and are involved in the stimulation of microbial protein synthesis [38]. The lower proportion of valerate promoted by WPS fermentation could indicate per se a decreased potential of microbial protein synthesis of this forage compared to AH.
The trial carried out to study the effect of replacing AH with WPS did not reflect major impact on the substitution levels of 25 and 50 % compared to 0 % on the rumen fermentation parameters, except for trends to linearly increase the rate of degradation (c), the OMD and the methanogenic capacity of the diet. However, no effect was observed on the parameters that were different (especially ME and total VFA and their molar proportions) when the WPS and AH were incubated alone in the trial described above. These differences could probably be explained by synergistic interactions of the microbial utilization of WPS with that of the other components of the diet (AH and concentrate) that did not occur when forages were incubated individually.
In an in vivo trial, Soliman et al. [39] used a mixed ration for lactating cows (40:60 forage to concentrate ratio) to study the replacement of berseem (leguminous plant) hay by dried watermelon vine in different percentages (0, 25, 50, 75 and 100 %) and observed, contrary to our observations, that OMD and total VFA production decreased from a 50 % of hay replacement. From this substitution level, they also observed a detrimental effect on productive parameters (i.e. milk yield and its dry extract). Nevertheless, it is necessary to consider that the quality of the watermelon plant used by these authors had significantly lower nutritional characteristics compared to the WPS in our study, as it had lower protein contents and higher cellulose and, especially, hemicellulose contents.
Recently, Hassan et al. [10], when studied the impact of the replacement (10, 20 and 30 %) of sunflower meal (protein concentrate that represented 6 % of the diet) with WPS using an in vitro approach with rumen inoculum from goats, no effect was observed on GP or total and molar proportion of VFA. This agrees with our results, although the total proportion of inclusion of WPS in the diet was much higher in our case (6 versus 25 % of the diet), which indicates that a considerable replacement of the standard diet with WPS has no harmful effects on rumen fermentation.
5. Conclusions
The present study shows that WPS could be used as a suitable ingredient in the formulation of diets for dairy goats, given the positive effects observed on ruminal fermentation and the ability to provide nutrients and energy. There are few results in the literature on the use of this source in the feeding of small ruminants; Therefore, the evaluation of the use of this by-product in animals, including the evaluation of WPS voluntary intake, is crucial in the feeding of lactating animals, would be favoured by further studies under practical farm conditions.
Given that conventional forages in many cases present problems because the demand of livestock farmers exceeds existing production or because prices are excessive, the use of watermelon plant silage could be an alternative to improve livestock sustainability through a strategy that meets the axioms of the circular bio-economy.
Author Contributions
Conceptualization, A.I.M.-G.; methodology, Z.P. and J.M.P.H.; validation, Z.P. and J.M.P.H.; formal analysis, A.I.M.G. and Z.P.; investigation, Z.P. and A.M.S.G.; data curation, A.I.M.G. and Z.P.; writing original draft preparation, Z.P. and A.I.M.G..; writing, review and editing, Z.P., A.I.M.G., J.M.P.-H. and A.M.S.G.; visualization, Z.P.; supervision, A.I.M.G.; project administration, A.I.M.G.; funding acquisition, A.I.M.G. All authors have read and agreed to the published version of the manuscript.
Funding
“This research was funded by MCIN/AEI/ 10.13039/501100011033 and by the “European Union””.
Institutional Review Board Statement
The CSIC Ethical Committee for Experimental Animal Protection and the re-gional Andalusian government (procedure 06/07/2023/61) approved all experimental protocols.
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors upon request.
Acknowledgments
The authors thank the Technical Scientific Services "Animal Experimentation" and "C and N Analysis" of the Estación Experimental del Zaidín (CSIC). Also, thanks for their help in laboratory procedures and advice to Isabel Jiménez and Elisabeth Jiménez.
Conflicts of Interest
The authors report no declarations of interest.
References
- European Commision. A new Circular Economy Action Plan. For a cleaner and more competitive Europe (COM/2020/98 final) 2020. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=COM:2020:98:FIN (accessed on 22 June 2024).
- FAO. How to Feed the World in 2050. Available online: https://www.fao.org/fileadmin/templates/wsfs/docs/expert_paper/How_to_Feed_the_World_in_2050.pdf (accessed on 21 June 2024).
- FAO. The future of food and agriculture:Trends and challenges. Rome 2017. Available online: https://www.fao.org/3/i6583e/i6583e.pdf (accessed on 22 June 2024).
- United Nations. Department of Economic and social affairs 2019. Available online: https://population.un.org/wpp/ (accessed on 22 June 2024).
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, P.C.; Balzer, C.; Bennett, E.M.; Carpenter, S.R.; Hill, J.; Monfreda, C.; Polasky, S.; Rockström, J.; Sheehan, J.; Siebert, S.; Zaks, D.P.M.; et al. Solutions for a cultivated planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef]
- Godde, C.M.; Mason-D’Croz, D.; Mayberry, D.E.; Thornton, P.K.; Herrero, M. Impacts of climate change on the livestock food supply chain; a review of the evidence. Glob Food Sec 2021, 28, 100488. [Google Scholar] [CrossRef]
- Arco-Pérez, A.; Ramos-Morales, E.; Yáñez-Ruiz, D.R.; Abecia, L.; Martín-García, A.I. Nutritive evaluation and milk quality of including of tomato or olive by-products silages with sunflower oil in the diet of dairy goats. Anim Feed Sci Technol 2017, 232, 57–70. [Google Scholar] [CrossRef]
- Méndez, D.A.; Martínez-Abad, A.; Martínez-Sanz, M.; López-Rubio, A.; Fabra, M.J. Tailoring structural, rheological and gelling properties of watermelon rind pectin by enzymatic treatments. Food Hydrocoll 2023, 135, 108119. [Google Scholar] [CrossRef]
- Chharang, D.; Tribhuwan, S.; Vijay, P.; Hemant, J.; Surendra, S.S. Effect of Replacement of Conventional Feeds by Prosopis juliflora Pods and Citrullus lanatus Seed Cake on Nutrient Utilization in Marwari Goats. J Anim Res 2020, 10, 551–555. [Google Scholar] [CrossRef]
- Hassan, M.; Belanche, A.; Jiménez, E.; Rivelli, I.; Martín-García, A.I.; Margolles, A.; Yáñez-Ruiz, D.R. Evaluation of the nutritional value and presence of minerals and pesticides residues in agro-industrial by-products to replace conventional ingredients of small ruminant diets. Small Rumin Res 2023, 229, 107117. [Google Scholar] [CrossRef]
- Martı́n-Garcı́a, A.I.; Moumen, A.; Yáñez-Ruiz, D.R.; Molina-Alcaide, E. Chemical composition and nutrients availability for goats and sheep of two-stage olive cake and olive leaves. Anim Feed Sci Technol 2003, 107, 61–74. [Google Scholar] [CrossRef]
- Molina-Alcaide, E.; Ruiz, D.R.Y.; Moumen, A.; Garcı́a, A.I.M. Ruminal degradability and in vitro intestinal digestibility of sunflower meal and in vitro digestibility of olive by-products supplemented with urea or sunflower meal. Anim Feed Sci Technol 2003, 110, 3–15. [Google Scholar] [CrossRef]
- Meneses, M.; Megías, M.D.; Madrid, J.; Martínez-Teruel, A.; Hernández, F.; Oliva, J. Evaluation of the phytosanitary, fermentative and nutritive characteristics of the silage made from crude artichoke (Cynara scolymus L.) by-product feeding for ruminants. Small Rumin. Res 2007, 70, 292–296. [Google Scholar] [CrossRef]
- Theodorou, M.K.; Williams, B.A.; Dhanoa, M.S.; McAllan, A.B.; France, J. A simple gas production method using a pressure transducer to determine the fermentation kinetics of ruminant feeds. Anim Feed Sci Technol 1994, 48, 185–197. [Google Scholar] [CrossRef]
- Menke, K.H.; Steingass, H. Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim. Res. Rural Develop 1988, 28, 7–55. [Google Scholar]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 18th ed.; AOAC International: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Fígares, I.; Prieto, C.; Nieto, R.; Aguilera, J.F. Free amino acid concentrations in plasma, muscle and liver as indirect measures of protein adequacy in growing chickens. Anim. Sci 1997, 64, 529–539. [Google Scholar] [CrossRef]
- Kheddouma, A.; Arhab, R.; Martín-García, AI.; Aouidane, L.; Bouraiou, A. Effects of the methane-inhibitors Nitrophenol, 5-Nitrobenzimidazol and two new synthetic nitrocompounds on in vitro ruminal fermentation. Biocatal Agric Biotechnol 2018, 14, 160–165. [Google Scholar] [CrossRef]
- France, J.; Dijkstra, J.; Dhanoa, M.S.; Lopez, S.; Bannink, A. Estimating the extent of degradation of ruminant feeds from a description of their gas production profiles observed in vitro: derivation of models and other mathematical considerations. Br. J. Nutr 2000, 83, 143–150. [Google Scholar] [CrossRef]
- Ahmed, M.G.; Al-Sagheer, A.A.; El-Waziry, A.M.; El-Zarkouny, S.Z.; Elwakeel, E.A. Ensiling Characteristics, In Vitro Rumen Fermentation Patterns, Feed Degradability, and Methane and Ammonia Production of Berseem (Trifolium alexandrinum L.) Co-Ensiled with Artichoke Bracts (Cynara cardunculus L.). Animals 2023, 131543. [Google Scholar] [CrossRef]
- Cui, Y.; Liu, H.; Gao, Z.; Xu, J.; Liu, B.; Guo, M.; Yang, X.; Niu, J.; Zhu, X.; Ma, S.; Li, D.; Sun, Y.; Shi, Y. Whole-plant corn silage improves rumen fermentation and growth performance of beef cattle by altering rumen microbiota. Appl. Microbiol. Biotechnol 2022, 106, 4187–4198. [Google Scholar] [CrossRef] [PubMed]
- Monllor, P.; Romero, G.; Sendra, E.; Atzori, A.S.; Díaz, J.R. Short-term effect of the inclusion of silage artichoke by-products in diets of dairy goats on milk quality. Animals 2020, 10, 339. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, R.R.d.; Edvan, R.L.; Nascimento, K.d.S.; Alves Barros, D.M.; Barros, L.d.S.; Camboim, L.F.R.; Dias e Silva, T.P.; Miranda, R.d.S.; Araújo, M.J.d.; Lima Neto, A.F.; et al. Characterization of Melon, (Cucumis melo L.) Silage with Different Biomass Mixtures and Dry Matter Contents. Agriculture 2023, 13, 1536. [Google Scholar] [CrossRef]
- Limin, K.R.D.; Shaver, R.D.; Grant, R.J.; Schmidt, R.J. Silage review: Interpretation of chemical, microbial, and organoleptic components of silages. J. Dairy. Sci 2018, 5, 4020–4033. [Google Scholar]
- Lin, J.; Li, G.; Sun, L.; Wang, S.; Meng, X.; Sun, L. Varieties and ensiling: Impact on chemical composition, fermentation quality and bacterial community of alfalfa. Front Microbiol 2023, 13. [Google Scholar] [CrossRef]
- Ibrahim, S.E.; Sulieman, A.M.E.; Hassan, E.N.; Ali, N.A.; Hakeem, B.S.A.E.; Muhsin, A.M.A.A. Proximae chemical composition of watermelon (Citrullus vulgaris). Plant. Cell. Biotechnol. Mol. Bio 2021, 22, 114–121. [Google Scholar]
- Kazemi, M. In vitro ruminal-microbial fermentation pattern: nutritional insights about some agricultural crop mesocarps (peel) in ruminant nutrition. AMB. Express 2023, 13, 144. [Google Scholar] [CrossRef] [PubMed]
- Prater, T.A.; Miller, D.D. Calcium carbonate depresses iron bioavailability in rats more than calcium sulfate or sodium carbonate. J Nutr 1992, 122, 327–332. [Google Scholar]
- Wu, G. Amino Acids: Biochemistry and Nutrition (2nd ed.); CRC Press, 2021. [Google Scholar] [CrossRef]
- Hou, Y.; Wu, G. Nutritionally Essential Amino Acids. Adv Nutr 2018, 9, 849–851. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S. E.; Sulieman, A.M.E.; Ali, N.A.; Bothaina.; Hakeem, S.A.; Amin, H.B.; Abdelmuhsin, A.A.; Veettil, V.N. Amino acid Profile of the Watermelon, Citrullus vulgaris and Detection of its Antimicrobial Activity. Biotech. Res. Comm 2019, 12. [Google Scholar] [CrossRef]
- Belanche, A.; Palma-Hidalgo, J.M.; Nejjam, I.; Serrano, R.; Jiménez, E.; Martín-García, I.; Yáñez-Ruiz, D.R. In vitro assessment of the factors that determine the activity of the rumen microbiota for further applications as inoculum. J Sci Food Agric 2019, 99, 163–172. [Google Scholar] [CrossRef]
- Pardo, Z.; Mateos, I.; Campos, R.; Francisco, A.; Lachica, M.; Ranilla, M.J.; Fernández-Fígares, I. Heat stress increases in vitro hindgut fermentation of distinct substrates in iberian pigs. Animals 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Amanzougarene, Z.; Fondevila, M. Fitting of the in vitro gas production technique to the study of high concentrate diets. Animals 2020, 10, 1935. [Google Scholar] [CrossRef]
- E.N. Bergman. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev 1990, 70, 567–590. [Google Scholar] [CrossRef]
- Baldwin, R.L.; McLeod, K.R.; Klotz, J.L.; Heitmann, R.N. Rumen development, intestinal growth and hepatic metabolism in the pre- and postweaning ruminant. J Dairy Sci 2004, 87, E55–E65. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, C.; Huang, Y.X.; Dong, K.H.; Yang, W.Z.; Zhang, S.L.; Wang, H. Effects of isovalerate on ruminal fermentation, urinary excretion of purine derivatives and digestibility in steers. J. Anim. Physiol. Anim. Nutr 2009, 93, 716–725. [Google Scholar] [CrossRef] [PubMed]
- Soliman, S.; El-Morsy, A.; El-Shinnawy, A.; El-Ashry, G.; Osman, M. Utilization of Biologically Treated Watermelon Vine in Rations for Dairy Cows. J. Anim. Poult. Prod 2020, 11, 117–123. [Google Scholar] [CrossRef]
Figure 1.
Cumulative gas production (mL/g DM) from forage sources in vitro fermentation in buffered rumen fluid obtained from Murciano-Granadina goats.
Figure 1.
Cumulative gas production (mL/g DM) from forage sources in vitro fermentation in buffered rumen fluid obtained from Murciano-Granadina goats.
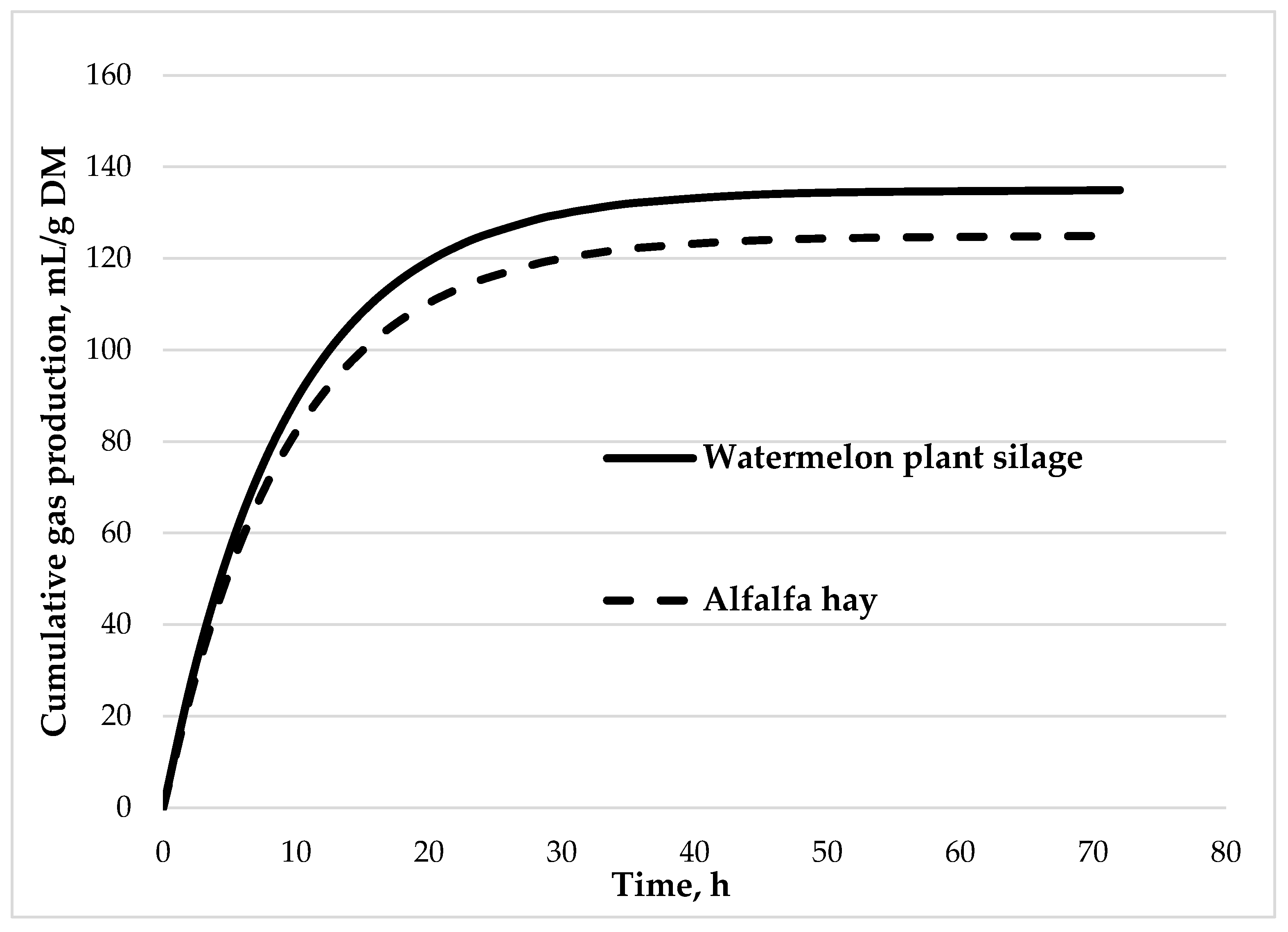
Figure 2.
Effect of substituting different percentages of alfalfa hay (0, 25, 50 %) with watermelon plant silage on the cumulative gas production (mL/g DM) from the in vitro fermentation of experimental diets.
Figure 2.
Effect of substituting different percentages of alfalfa hay (0, 25, 50 %) with watermelon plant silage on the cumulative gas production (mL/g DM) from the in vitro fermentation of experimental diets.
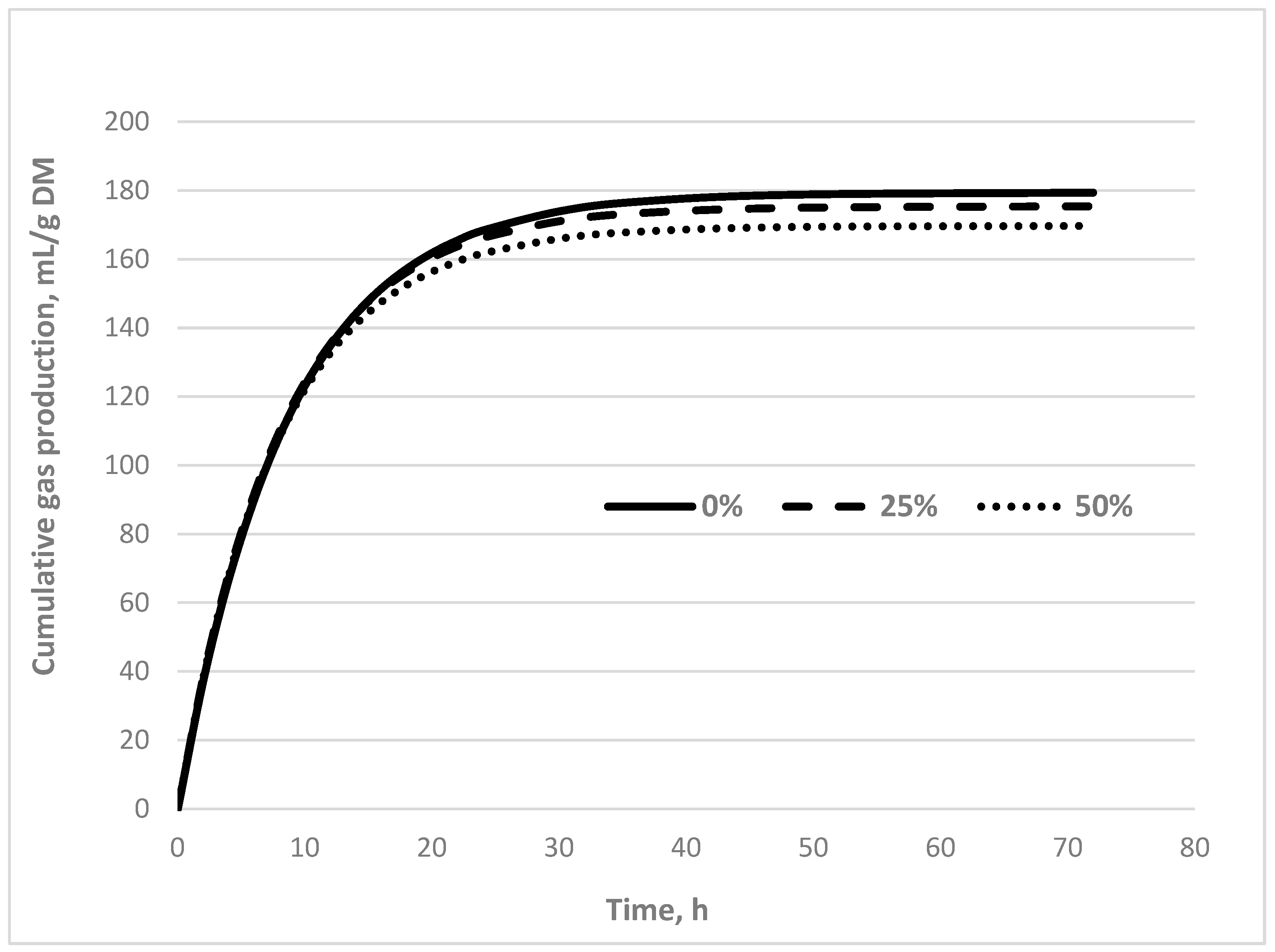

Table 1.
Nutrient composition of the materials used in the study.
| Composition | Watermelon plant | Watermelon plant silage | Alfalfa hay | Concentrate |
|---|---|---|---|---|
| DM 1, g/100 g FM 2 | 14.7 | 15.7 | 92.8 | 91.3 |
| Nutrients, g/100 g DM | ||||
| OM 3 | 79.8 | 77.4 | 89.6 | 91.6 |
| CP 4 | 22.6 | 21.1 | 18.9 | 19.7 |
| CF 5 | 1.40 | 3.09 | 1.29 | 3.29 |
| NDF 6 | 36.7 | 36.3 | 49.4 | 30.3 |
| ADF 7 | 24.2 | 26.4 | 34.9 | 15.3 |
| ADL 8 | 7.86 | 6.40 | 8.01 | 4.75 |
| Hemicellulose | 12.6 | 9.91 | 14.6 | 15.0 |
| Cellulose | 16.3 | 20.0 | 26.9 | 10.6 |
| Total Carbohydrates | 55,8 | 53,2 | 69,4 | 68,6 |
| Non Fibrous Carbohydrates | 19.1 | 16.9 | 20.0 | 38.3 |
| CE 11, MJ/kg DM | 16.0 | 15.5 | 17.6 | 17.1 |
| Macrominerals, g/kg DM | ||||
| Ca | 44.7 | 4.40 | ||
| K | 22.5 | 11.1 | ||
| Mg | 6.07 | 2.30 | ||
| P | 2.91 | 2.00 | ||
| S | 2.28 | 1.30 | ||
| Microminerals, mg/kg DM | ||||
| Na | 198 | 40.0 | ||
| Fe | 119 | 180 | ||
| Al | 103 | 227 | ||
| Mn | 39.6 | 43.1 | ||
| Zn | 14.6 | 46.8 | ||
| Cu | 5.62 | 5.33 | ||
| Ti | 3.42 | 10.6 | ||
| As | n.d. 12 | 0.340 | ||
| B | n.d. 12 | 11.9 | ||
| Si | n.d. 12 | 129 | ||
| Sr | n.d. 12 | 46.0 |
1 DM: Dry matter; 2 FM: Fresh matter; 3 OM: Organic matter; 4 CP: Crude protein; 5 CF: Crude fat; 6 NDF: Neutral detergent fiber; 7 ADF: Acid detergent fiber; 8 ADL: Acid detergent lignin; 9 Total Carbohydrates = (100-(CP+CF+Ash); 10 Non Fibrous Carbohydrates = Total Carbohydrates - NDF; 11 CE: Crude energy; 12 n.d.: not detected.
Table 2.
Amino acid (AA) composition of forage protein compared.
| g AA/100 g AA | Watermelon plant silage | Alfalfa Hay |
|---|---|---|
| Aspartic acid | 8.56 | 12.5 |
| Serine | 5.86 | 4.69 |
| Glutamic acid | 14.9 | 9.66 |
| Glycine+Histidine* | 4.96 | 8.15 |
| Arginine | 5.67 | 6.15 |
| Threonine* | 5.38 | 4.63 |
| Alanine | 8.10 | 6.20 |
| Proline | 8.10 | 10.5 |
| Tyrosine* | 3.22 | 3.92 |
| Valine* | 6.61 | 6.67 |
| Lysine* | 5.88 | 7.40 |
| Isoleucine* | 4.59 | 4.84 |
| Leucine* | 9.16 | 8.66 |
| Phenylalanine* | 9.03 | 5.98 |
| EAA 1 | 48.8 | 50.3 |
| NEAA 2 | 51.2 | 49.7 |
| g AA/kg DM | 151 | 165 |
| g AA/kg N | 716 | 874 |
1 EAA: essential amino acids (Hou and Wu et al, 2018; Cys, Met and Trp were not determined), 2NEAA: non-essential amino acids.
Table 3.
Fermentation parameters, after 24 h of in vitro culture of rumen microorganisms, of the compared forages.
Table 3.
Fermentation parameters, after 24 h of in vitro culture of rumen microorganisms, of the compared forages.
| Watermelon plant silage | Alfalfa hay | SEM 1 | p-Value | |
|---|---|---|---|---|
| A 2, ml | 135 | 125 | 6.71 | 0.679 |
| c 3, h-1 | 0.108 | 0.107 | 0.006 | 0.901 |
| pH | 7.00 | 7.04 | 0.015 | 0.017 |
| GP 4 24h, ml/g DM | 120 | 111 | 5.07 | 0.506 |
| ME 5, MJ/kg DM | 6.67 | 5.72 | 0.167 | 0.058 |
| OMD 6, g/kg | 471 | 410 | 13.8 | 0.126 |
| CH4 µl/ml GP | 47.1 | 43.4 | 1.27 | 0.287 |
| Total VFA 7, mM | 66.6 | 115 | 6.70 | 0.011 |
| Acetate, % | 70.1 | 70.2 | 0.809 | 0.963 |
| Propionate, % | 17.3 | 19.0 | 0.283 | 0.022 |
| Butyrate, % | 7.40 | 6.17 | 0.381 | 0.170 |
| Isobutyrate, % | 1.78 | 1.17 | 0.042 | 0.001 |
| Valerate, % | 1.35 | 1.57 | 0.047 | 0.014 |
| Isovalerate, % | 2.07 | 1.77 | 0.102 | 0.210 |
| Acetate/Propionate | 4.08 | 3.51 | 0.103 | 0.057 |
1 SEM: Standard Error of Mean, 2 A: Asymptotic value of total gas production, 3 c: the rate of degradability, 4 GP: gas production, 5 ME: Metabolizable energy, 6 OMD: Organic matter digestibility, 7 VFA: Volatile fatty acids.
Table 4.
Effect of substituting different percentages of alfalfa hay (0, 25 and 50 %) with watermelon plant silage on fermentation parameters after 24 h in vitro rumen microorganisms culture.
Table 4.
Effect of substituting different percentages of alfalfa hay (0, 25 and 50 %) with watermelon plant silage on fermentation parameters after 24 h in vitro rumen microorganisms culture.
| Inclusion rate, % | 0 | 25 | 50 | SEM 1 | p-value |
|---|---|---|---|---|---|
| A 2, ml | 179 | 175 | 170 | 9.01 | 0.549 |
| c 3, h-1 | 0.116 | 0.123 | 0.127 | 0.01 | 0.088 |
| pH | 6.71 | 6.72 | 6.72 | 0.16 | 0.469 |
| GP 4 24h, ml/g DM | 162 | 169 | 170 | 4.65 | 0.558 |
| ME 5, MJ/kg DM | 6.81 | 6.80 | 6.80 | 0.28 | 0.436 |
| OMD 6, g/kg | 645 | 692 | 769 | 59.9 | 0.067 |
| CH4 µl/ml GP | 48.3 | 54.4 | 55.3 | 1.00 | 0.094 |
| CH4, µl/mol VFA | 723 | 710 | 799 | 39.8 | 0.595 |
| Total VFA 7, mM | 66.8 | 81.1 | 70.5 | 5.14 | 0.529 |
| Acetate, % | 66.3 | 66.6 | 66.5 | 0.33 | 0.594 |
| Propionate, % | 20.5 | 20.7 | 20.9 | 0.24 | 0.183 |
| Butyrate, % | 8.16 | 7.89 | 7.73 | 0.32 | 0.895 |
| Isobutyrate, % | 1.42 | 1.39 | 1.38 | 0.03 | 0.345 |
| Valerate, % | 1.53 | 1.52 | 1.51 | 0.02 | 0.183 |
| Isovalerate, % | 1.86 | 1.82 | 1.81 | 0.05 | 0.531 |
| Acetate/Propionate | 3.22 | 3.22 | 3.18 | 0.03 | 0.138 |
1 SEM: Standard Error of Mean, 2 A: Asymptotic value of total gas production, 3 c: the rate of degradability, 4 GP: gas production, 5 ME: Metabolizable energy, 6 OMD: Organic matter digestibility, 7 VFA: Volatile fatty acids.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



