Submitted:
21 June 2024
Posted:
25 June 2024
You are already at the latest version
Abstract
Azolla is the only plant with a co-evolving nitrogen-fixing (diazotrophic) cyanobacterial symbiont (cyanobiont), Nostoc azollae, resulting from Whole Genome Duplication (WGD) 80 million years ago in Azolla’s ancestor. Additional genes from the WGD resulted in genetic, biochemical and morphological changes in the plant that enabled transmission of the cyanobiont to successive generations via its megaspores. The resulting permanent symbiosis and co-evolution led to loss, downregulation or conversion of non-essential genes to pseudogenes in the cyanobiont, changing it from a free-living organism to an obligate symbiont. Upregulation of other genes in the cyanobiont increased its atmospheric dinitrogen fixation and provision of nitrogen-based products to the plant. As a result, Azolla can double its biomass in less than two days free-floating on fresh water and sequester large amounts of atmospheric CO2, giving it the potential to mitigate anthropogenic climate change through Carbon Capture and Storage. Azolla’s biomass can also provide local, low-cost food, biofertiliser, feed and biofuel that are urgently needed as our population increases by a billion every twelve years. This paper integrates data from biology, genetics, geology and paleontology to identify the location, timing and mechanism for the acquisition of a co-evolving diazotrophic cyanobiont by Azolla’s ancestor in the Late Cretaceous (Campanian) of North America.
Keywords:
anabaena
; azolla
; climate change
; cyanobacteria
; evolution
; genetics
; nostoc
; sequestration
; symbiosis
1. Introduction
1.1. Nitrogen
Nitrogen is essential for all life on Earth. It is one of the five elements that form nucleotides – the chemical building blocks of deoxyribonucleic acid (DNA) that contains genetic information and biological instructions for the development and functioning of all organisms. It is a component of proteins and photosynthetic pigments, including chlorophyll, that enable algae and plants to harvest energy from the sun, but they are unable obtain it from the atmosphere because of the strength of the dinitrogen (N2) triple bond. The only organisms that can sever the bond and synthesise dinitrogen are a group of prokaryotes, called diazotrophs, that evolved the process more than 2.7 billion years ago using an enzyme called nitrogenase and a process called Biological Nitrogen Fixation (BNF) [1,2].
A few fungi and plants, including some legumes, have temporary symbioses with diazotrophs, but the relationship is severed when the fungus or plant dies and has to be re-established by successive generations. Azolla is the only plant with a co-evolving nitrogen-fixing cyanobacterial symbiont (cyanobiont), Nostoc azollae, that is passed directly to successive generations of the plant via its megaspores. Colonies of the cyanobacteria live in Azolla’s floating leaves, providing nitrogen-based nutrients to the plant that enable it to double its biomass in less than two days free-floating on fresh water [3] (Figure 1). As a result, Azolla has been used for hundreds of years in India and the Far East as a renewable, low-cost livestock feed and nitrogen biofertiliser for paddy rice [4], reducing mosquito breeding populations by 95 % [5,6,7] and emissions of the potent greenhouse gas methane from paddies by up to 50 % [8,9,10].
Azolla can provide local biofertiliser, livestock feed, biofuel and food [12] globally when grown indoors [11], helping alleviate shortages of the ‘three Fs’ that threaten food supplies: fertiliser, feed and fuel. It absorbs and removes phosphates and nitrates from water contaminated by chemical fertilisers, industrial pollutants, and animal and human waste that trigger toxic cyanobacterial (aka blue-green algal) blooms in rivers and lakes. The symbionts’ combined CO2 sequestration increases Azolla’s carbon capture, so that it can sequester large amounts of atmospheric CO2, with the plants being compressed and permanently stored to reduce anthropogenic climate change through Carbon Capture and Storage (CCS) (Figure 2). See 'The Azolla Story' for a review of its uses [11].
Azolla can, therefore, mitigate many of the threats arising from a Perfect Storm as our population increases by more than a million every three days. Its remarkable properties are increasingly recognised, and it was designated as a unique superorganism by Francisco Carrapiço in 2010 [13], but the origin of Azolla’s co-evolving symbiosis with N. azollae has, until recently, been obscure. This paper integrates data from biology, genetics, geology and paleontology to identify the location, timing and mechanism of their unique relationship.
1.2. Azolla’s Suprageneric Classification
The name ‘azolla’ was introduced by the French naturalist Jean-Baptiste Lamarck in his 1783 publication Encyclopédie Méthodique for plants collected from South America by the naturalist Philibert Commerson and his assistant Jeanne Baret (aka Baré, Barret) during Louis-Antoine de Bougainville’s 1767-1768 circumnavigation of the world [14]. Azolla’s suprageneric classification varies, with some authors assigning Azolla to the monogeneric Family Azollaceae [15] and others assigning the genera Azolla and Salvinia to the free-floating Family Salviniaceae, although Azolla can also root in damp soil or mud [16,17,18]. The Family Salviniaceae is assigned to the Order Salviniales, also known as water ferns, that includes the Family Marsileaceae comprising three genera (Marsilea, Pilularia, Regnelidium) that root in mud and are not free-floating.
1.3. The Cyanobacterial Symbiont
Azolla’s cyanobiont has been assigned to both Anabaena azollae and Nostoc azollae because it resembles free-living species of both genera in having chains of cells (filaments) comprising photosynthetic vegetative cells and thicker-walled heterocysts containing the nitrogen-fixing enzyme nitrogenase that is destroyed by oxygen (Figure 3). Akinetes (resting cells) may also be present as their germination ensures survival during stressed conditions [19]. We use the name Nostoc azollae for consistency with other papers in PLANT’S Special Issue "Plant–Cyanobacteria Symbiosis: From Morphology to Practical Uses".
N. azollae‘s filaments are immobilized in a gelatinous mucilage sheath around the periphery of a chamber inside Azolla’s dorsal floating leaves, with pores permitting the exchange of air, but not microorganisms including prokaryotes, with the chamber. Slender rods, called trichomes, transport chemicals and nutrients between the cells of Azolla and N. azollae, with the latter releasing ovoidal capsules, called membrane vesicles (MVs), that contain genetic material [21], so that Azolla and N. azollae are in biochemical and genetic communication.
The vegetative cells of N. azollae use photosynthesis to draw down atmospheric CO2 and produce organic compounds that they utilise and pass to the heterocysts for their metabolism. The heterocysts fix atmospheric dinitrogen and synthesise it into nitrogen-based compounds that they utilise and pass to the vegetative cells. This involves complex biochemical processes in both types of cells and the presence of cyanophycin at their interface [20] (Figure 3). Cyanophycin, abbreviated from cyanophycin granule peptide (CGP), is a nitrogen polymer that acts as a reservoir for fixed nitrogen in the heterocysts, enables the transport of fixed nitrogen to adjacent vegetative cells, and inhibits the transport of soluble nitrogen products from vegetative cells to heterocysts [22].
Differentiation of heterocysts and vegetative cells probably evolved in Anabaena’s and Nostoc’s common ancestor during the Great Oxidation Event (GOE) between 2.5 and 2.1 billion years ago when the Earth’s atmosphere was enriched in oxygen, resulting in the mass extinction of many anaerobic prokaryotes [23,24,25]. The heterocysts enclosed an oxygen-free environment containing nitrogenase, with cyanophycin providing a selective barrier and conduit between the heterocysts and oxygenated vegetative cells. As a result of these ancient adaptations, today’s species of Anabaena and Nostoc, including Azolla’s cyanobiont N. azollae, are the most biochemically and morphologically complex multicellular cyanobacteria with differentiated akinetes, heterocysts and vegetative cells.
1.4. Azolla’s Transmission of N. azollae via Its Megaspores
Azolla is unique in its intergenerational transmission of a diazotroph via its spores. This involves biochemical and morphological processes unknown in any other plant and resulted from Azolla‘s adaptation and utilisation of morphological changes that occur in free-living Anabaena and Nostoc. These enable N. azollae’s movement inside Azolla and its fusion with Azolla‘s female megaspores during their dormancy and germination into a new plant.
Some free-living species of Anabaena and Nostoc change their morphology, mobility and nitrogen-fixation in response to changes in the environment and the emission of chemicals emitted by several plants with which they have a temporary symbiosis [26,27,28]:
- Sessile filaments have vegetative cells that sequester carbon dioxide and heterocysts that fix atmospheric nitrogen. Akinetes may also be present.
- Motile hormogonia only have vegetative cells, so they cannot sequester nitrogen. Hormogonia are produced by free-living species of Anabaena and Nostoc including those that have temporary symbioses with plants like Gunnera manicata. The plants emit hormogonium-inducing factor (HIF) to induce Nostoc in the soil to change into hormogonia that infect specialised stem glands in the plant, after which they change into sessile filaments including heterocysts that provide nitrogen-based compounds to the plant.
- Akinetes, or resting spores, are produced by free-living Anabaena and Nostoc, enabling them to survive adverse conditions.
The ways in which Azolla adapted these processes inside the plant are unique and involved genetic, biochemical and morphological adaptions, with a combination of biology, genetics, geology and paleontology identifying the location, timing and mechanism in Azolla’s ancestor.
2. Evidence for Azolla’s Origin
2.1. Genetic Evidence for Azolla’s Origin
The phylogeny and genetic events shown in Figure 4 are crucial for understanding Azolla’s unique relationship with N. azollae because they document the key genetic event that resulted in their fusion into a unique superorganism – a process called Whole Genome Duplication (WGD). WGDs are important evolutionary events because they ‘provide a source of genetic material for mutation, drift, and selection to act upon, making new evolutionary opportunities possible’ [29, Abstract].
Li et al. (2018) identified two WGD events in the Salviniales based on genetic analysis: an earlier one (WGD1 in Figure 4) 225 million years ago in the Late Triassic and a later one (WGD2) 80 million years ago in the Late Cretaceous [17]. WGD1 may have facilitated the Late Triassic colonisation of aquatic habitats by the Salviniales, after which they evolved into two Families, the Marsileaceae and Salviniaceae, near the end of the Jurassic about 155 million years ago. The Salviniaceae then diverged into two lineages, each having one extant genus: Azolla which has a co-evolving nitrogen-fixing cyanobacterium and Salvinia which does not possess one. Their difference is indicated by the relative size of Azolla and Salvinia’s genomes, with Azolla’s being approximately three times larger than that of Salvinia with more than twice the number of chromosomes (Figure 4). They differ in size because Azolla’s ancestor underwent WGD about 80 million years ago after the two genera diverged from a common ancestor. The 80-million-year-old WGD event, therefore, only affected Azolla. It did not affect any other plant and that is one of the reasons why Azolla is unique.
N. azollae’s coevolution with Azolla resulted in extensive changes to the cyanobiont’s genome compared to free-living species of Anabaena and Nostoc [30,31,32,33]. These alterations involved the downregulation, loss or conversion to pseudogenes of some genes, changing N. azollae’s ancestors from independent free-living organisms into ones that could not survive outside Azolla. The upregulation of other genes enhanced N. azollae’s sequestration of atmospheric nitrogen and provision of nitrogen-based compounds to Azolla, increasing the plant’s speed of growth free-floating on fresh water.
Approximately one-third of N. azollae’s genome comprises non-functional pseudogenes. These include pseudogenes originating from genes that had previously expressed proteins involved in filament growth and division, reflecting the need to restrict their number in the plant’s confined leaf cavities. Other pseudogenes reflect a loss of function relating to the cyanobiont’s assimilation of nitrogen, so that increased amounts of nitrogen-based compounds are transferred from the cyanobiont’s cells to those of Azolla, facilitating the plant’s growth and its provision of almost three times more nitrogen biofertiliser than other plants including legumes [34].
A third set of pseudogenes originated from genes that previously expressed proteins involved in the synthesis of carotenoid and chlorophyll pigments. Larsson et al. (2011) analysed the genomes of 57 free-living cyanobacteria and N. azollae for shared and unique orthologs and found that two protein groups occur in the genomes of all the cyanobacteria except N. azollae where they are represented by non-functional pseudogenes [32]. These correspond to (1) geranylgeranyl pyrophosphate synthase that is involved in the synthesis of carotenoids and chlorophyll, and (2) uroporphyrinogen-III synthase HemD that generates precursors of tetrapyrroles which provide protection against photooxidative damage. N. azollae is therefore reliant on Azolla’s cellular pigments for protection against photooxidative damage so that it cannot survive outside the plant. The previously independent cyanobacterium became an obligate symbiont and an indispensable partner of the Azolla superorganism. It also resulted in complementary photosynthesis of Azolla and its cyanobiont, with Azolla utilising shorter wavelength pigments including chlorophyll a, chlorophyll b and β-carotene, and N. azollae utilising longer wavelength pigments including phycoerythrin and phycocyanin. As a result, Azolla is a shade plant that needs little illumination for its growth and proliferation, facilitating its growth next to rice plants in paddies, and its cultivation for CCS and renewable food, biofertiliser, livestock feed and biofuel.
In contrast to N. azollae’s pseudogenes, genes that were retained or up-regulated in N. azollae enhance its symbiosis with Azolla and, especially, the sequestration and synthesis of atmospheric nitrogen, reflecting N. azollae’s primary symbiotic role in providing nitrogen-based compounds to Azolla. The entire set of genes related to nitrogen fixation (the nif gene cluster) is, therefore, intact in N. azollae because those genes express proteins involved in the conversion of atmospheric nitrogen into nitrogen-based compounds.
N. azollae’s provision of nitrogen-based compounds is also increased by the number of heterocysts that assimilate atmospheric nitrogen. All twenty-two genes related to heterocyst formation are intact in N. azollae, but the pats gene that expresses proteins involved in the suppression of heterocyst development, is absent. N. azollae, therefore, produces more heterocysts than free-living cyanobacteria, resulting in chains of cells that contain up to 30 % heterocysts compared to less than 10 % in free-living Anabaena and Nostoc. This enables N. azollae to fix between four and eighteen times more atmospheric nitrogen than other species of Anabaena and Nostoc [33,35]. As a result, Azolla provides more nitrogen biofertiliser than any other plant, increasing its potential to augment or replace nitrogen-based chemical fertilisers.
2.2. Paleontological Evidence for Azolla’s Origin
A database compiled by the Geological Survey of Canada listed more than 120,000 published species of fossil pollen and spores, including 89 species of the genus Azolla, and several species of the similar genera Azollopsis, Paleoazolla and Parazolla [36]. Subsequent publications were assessed in the present study, including those documenting the Eocene Arctic Azolla Event that triggered the initial shift from a greenhouse to icehouse climate 49 million years ago [37,38].
The oldest confident record of Azolla is that of Hall (1969) from Campanian rocks of the western interior of North America [39] (Figure 5). Reported occurrences of Azolla and Azollopsis from older strata are questionable because of mis-identification of specimens or erroneous assignments of their age from the Late Cretaceous (Turonian-Cenomanian) of Kazakhstan [40,41] and India [42], and the Jurassic to Early Cretaceous of India [43], Kazakhstan [44], Tajikistan [40], Turkmenistan [45], Ukraine [46] and eastern Russia [44,47,48,49,50] (Appendix A).
Hall (1969) analysed samples from the Claggett and Judith River Formations near the Upper Missouri River Breaks National Monument (UMRBNM) in north-central Montana [39]. Both formations are Campanian, with the Claggett being older, so that together they potentially preserve a fossil record of Azolla’s early development (Figure 6). Hall considered fossils from the Judith River Formation to be the oldest species of Azolla, which he named Azolla simplex and assigned fossils from the Claggett Formation to a new genus and species, Parazolla heterotricha.
Hall did not observe the vegetative remains of either species, but the fossilised spores and sporangia of both have distinctive structures, called floats, and massulae with glochidia that enable the spores to reach and remain on the water’s surface after germination. Extant species of Azolla and A. simplex have distinctive floats and anchor-shaped glochidia, whereas those of P. heterotricha are hair-like which, according to Hall, are more ‘primitive’ [39]. He therefore proposed that P. heterotricha was the ancestor of A. simplex based on their morphology and relative age, and that A. simplex was the oldest species of Azolla.
As discussed earlier, Azolla is unique in having a permanent diazotrophic cyanobiont that is transmitted to subsequent generation of the plant by its megaspores. It is presently uncertain whether P. heterotricha also had a temporary or permanent diazotrophic cyanobiont. We therefore use the informal term ‘azolla’ in the following text for the earliest plant that had a permanent diazotrophic cyanobiont, and the term ‘proto-azolla’ for its immediate ancestor that had a temporary diazotrophic cyanobiont.
2.3. Age of the Oldest Azolla Fossils
The Campanian Stage extends from 83.7 to 72.2 million years (Ma) based on Gradstein et al.’s (2020) Geological Time Scale [52], with the succeeding Maastrichtian Stage extending up to 66.04 Ma, co-eval with the end-Cretaceous Yucatan bolide impact. Rogers (2016) determined the age in millions of years of three horizons in the Judith River Formation using radiometric dating of 40Ar/39Ar argon isotopes in volcanic ash layers called bentonites [53]. The ashes were ejected from volcanoes in the Elkhorn Mountains of southwestern Montana and deposited across freshwater lakes and rivers including Montana’s UMRBNM and the stratotype of the Judith River Formation.
Additional isotope data from Ramezani et al. (2022) provided ages for relevant sections in Alberta, Mexico, Montana and Utah [54], including the earliest recorded occurrences of A. simplex, P. heterotrycha and the genus Azollopsis shown in Figure 6.
Fossils of P. heterotrycha occur near the base of the Claggett Formation ‘not far above the contact with the Eagle Sandstone’ [now the Eagle Formation] at the western end of the UMRBNM [39, page 1177]. There is no radiometric age assignment for the horizon, but regional correlations indicate that the base of the Claggett Formation is Campanian and no older than 83.7 Ma. Rocks containing P. heterotrycha have an age close to 80 Ma based on their thickness and a sedimentation rate of approximately 3.8 cm/1000 years [53]. This is consistent with 80 Ma indicated by genetic analysis for the WGD event that resulted in the earliest Azolla [17].
2.4. The Geological Evidence
The Cretaceous (143.1-66.04 Ma) had a greenhouse climate with polar temperatures that rarely fell below freezing. At the beginning of the Campanian (83.65 Ma) the North American continent was separated by shallow seas into three large islands, with the western isle forming a long narrow landmass extending from today’s Alaska to Mexico [55]. Its western shore bordered the Pacific Ocean, and its eastern coast was lapped by a shallow Western Interior Seaway (WIS) that connected the Arctic Ocean and Gulf of Mexico (Figure 7). Halfway along its length, an eastern arm of the WIS separated two islands before joining a nascent Labrador Sea between North America and Greenland, with most of southern Europe being submerged under a shallow sea punctuated by subtropical islands.
Uplift during the Campanian-Maastrichtian (83.65-66.04 Ma) converted the North American seaways into freshwater rivers, lakes and nutrient-rich land that had been submerged. Previously separated terrestrial fauna, including dinosaurs like Tyrannosaurus, met for the first time and vied for dominance as vegetation rapidly spread across the newly emergent land, including angiosperms, horsetails, cycads and conifers [56]. Beneath the trees, the undergrowth was filled with a dense cover of ferns, some growing on the edges of ponds and lakes where they rooted in the shallow substrate. By 80 Ma, during deposition of the Claggett Formation, these included ‘proto-azolla’ that floated on the surface of lakes in today’s Montana.
3. Evolution of the Earliest Azolla
3.1. Temporary Symbiosis in Azolla’s Ancestor
Integration of data from the past and present provides a model of events that resulted in the evolution of proto-azolla into azolla. Many of today’s freshwater cyanobacterial blooms are triggered by the runoff of phosphate and nitrogen fertilisers [57] and it is probable that the floating leaves of proto-azolla were surrounded by masses of cyanobacteria that thrived on nutrients, and especially phosphates, transported by streams and rivers from newly emergent shales, muds and fertile soils.
Today’s temporary symbioses with diazotrophs occur in several plants and fungi including the hornwort Anthoceros punctatus, the liverwort Blasia, the angiosperm Gunnera, some legumes and cycads [58,59,60]. The most common are legumes that temporarily house nitrogen-fixing soil bacteria called rhizobia in their root nodules, indicating that N. azollae’s presence in the floating leaves of extant Azolla resulted from proto-azolla’s leaves being surrounded by floating cyanobacteria that may have occurred as gelatinous masses like those formed by today’s Nostoc [61].
Gunnera’s symbiosis is similar to that of Azolla as it involves Nostoc. It only occurs when the soil is deficient in nitrogen and begins when the plant emits chemicals called hormogonium-inducing factor (HIF) that induce the cyanobacteria to change from sessile filaments into motile hormogonia [62]. The movement of many prokaryotic hormogonia, including Nostoc punctiforme involves a twitching motility through extension-retraction cycles of type IV pili (T4P) augmented by surface waves generated by the contraction of a fibril layer, with Nostoc hormogonia achieving speeds of ten micrometre per second [63,64,65,66]. This enables the hormogonia to propel themselves to Gunnera’s stems where they enter specialised glands that are only developed when the plants are deprived of combined nitrogen [62].
The symbiosis, therefore, is promoted by the plant’s nitrogen deficiency, confirming the cyanobacteria’s primary symbiotic role to provide nitrogen to the plant, as seen in today’s species of Azolla and, by inference, proto-azolla. During periods of nitrogen deficiency, proto-azolla emitted HIF to trigger a change in the cyanobacteria from sessile filaments into motile hormogonia that propelled themselves towards the floating leaves where the symbiosis occurred in cavities that developed due to low combined nitrogen content in the water. Their entry into the cavities is indicated by Gunnera’s relationship with Nostoc in which the hormogonia follow each other in orderly lines towards the stem glands [62].
Temporary openings in proto-azolla’s leaves enabled entry of the cyanobacteria into cavities where they changed into sessile filaments with nitrogen-fixing heterocysts. This may have been facilitated by the plant’s emission of hormogonium repressing factors (HRF) that are produced by several fungi and plants [28,67,68]. These include Azolla pinnata where HRFs are more common in the plant’s mature leaves than those at the stem apex, providing a reservoir of hormogonia for conversion to filaments in the developing leaves [69].
A temporary symbiosis was, therefore, established in proto-azolla with entry into its leaf cavity being sealed. The cyanobacteria’s heterocysts provided the plant with nitrogen-based nutrients that promoted its growth, and the plant supplied the cyanobacteria with metabolites. Photosynthesis by cyanobacteria’s vegetative cells was facilitated by light penetrating the plant’s floating leaves, with narrow pores enabling air exchange with the atmosphere. The temporary symbiosis ended when the plant died and had to be re-established by successive generations of proto-azolla.
3.2. Whole Genome Duplication (WGD)
Whole Genome Duplication (WGD) occurred in a single plant approximately 80 million years ago, with the additional genes enabling its descendants to transmit the cyanobacteria via their megaspores. This was achieved in three ways:
- Chemical compounds were secreted inside the plant to induce changes in cyanobacterial mode involving motile hormogonia, sessile filaments and resting akinetes, facilitating their movement, retention and viability during the plant’s sexual reproduction. Similar chemical and environmental triggers induce the same changes in today’s free-living Anabaena and Nostoc.
- A series of passages inside the plant enabled the cyanobacteria to travel from the plant’s leaf cavities to a chamber, called the indusium, next to its developing megaspore. After germination of the megaspore, a channel opened in the embryonic leaf (cotyledon), enabling the cyanobacteria to travel from the indusium to dorsal leaf cavities as they developed in the new plant.
- The cyanobionts’ movement through the passages was directed by chemicals secreted inside the plant, augmented by a negative nitrogen gradient, similar to the attraction of today’s Nostoc in the soil to Gunnera’s stem nodules.
Azolla’s sexual reproduction illustrates the complex strategies and processes that Azolla’s and N. azollae’s ancestors evolved, enabling them to remain together and co-evolve for 80 million years (Figure 8). Azolla produces two types of spores during sexual reproduction in structures called sporocarps: (1) female megaspores that occur in conical megasporocarps about half a millimetre in length and (2) male microspores that occur in larger, spherical microsporocarps. Each megasporocarp produces 32 female megaspores, but only one survives inside the megasporocarp which has a cavity, called the indusium, inside its narrow end (Figure 8).
Prior to Azolla’s sexual reproduction, sessile filaments of N. azollae are immobilised in mucilage that fills the periphery of Azolla’s floating-leaf cavities, similar to gelatinous masses surrounding free-living Nostoc [61]. When Azolla’s spores begin to develop, Azolla secretes HIF into its leaf cavities that induces N. azollae to change into motile hormogonia, in the same way that some species of Gunnera secrete HIF outside the plant to induce free-living Nostoc to convert to hormogonia and enter the plant’s stem glands. Other chemicals are emitted that dissolve or disaggregate the mucilage, freeing the hormogonia to move inside the plant.
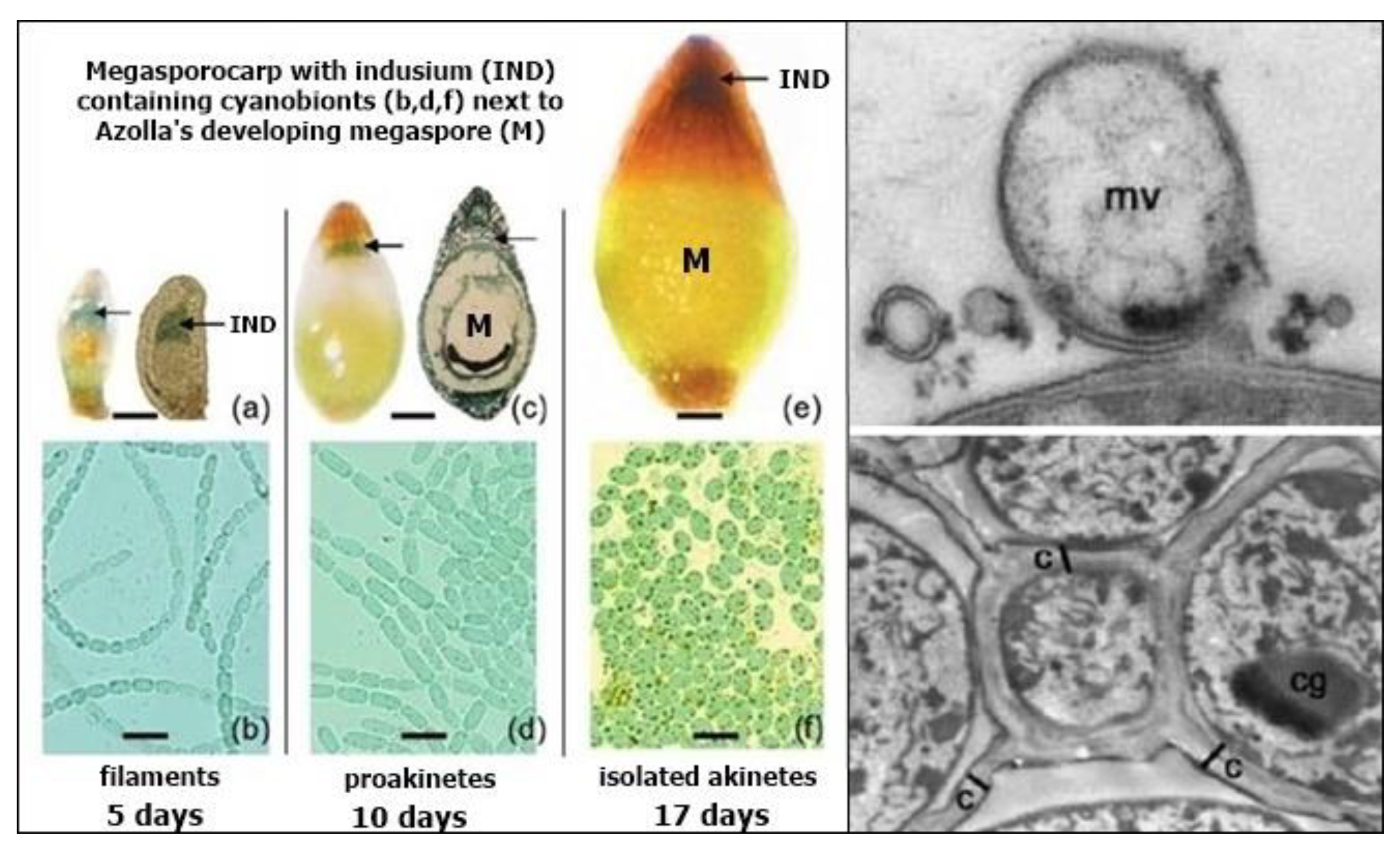
N. azollae’s hormogonia enter a passage that opens at one end of the leaf’s cavity, providing a conduit for them to travel to Azolla’s megasporocarp on the lower, submerged lobe of the leaf where they enter a cavity called the indusium that encloses them next to the developing megaspore (Figure 9). The hormogonia change back into filaments that excrete membrane vesicles containing genetic material, including DNA, that they exchange with Azolla [70]. The filaments of vegetative cells then change into chains of proakinetes before separating into unicellular akinetes that are immobilised by a structured biofilm filling the indusium, with the network of channels and akinetes resembling cells that are fused to the megaspore (Figure 9). Akinetes are unknown inside any plant except for Azolla, and their resistance to adverse conditions enables their survival next to Azolla’s megaspore as it develops and germinates into a new plant [70].
3.3. Germination and Propagation
Azolla’s male microsporocarps are larger than its female megasporocarps and have an indusium chamber that does not contain N. azollae, so the cyanobacteria are always transmitted via Azolla’s female reproductive organs. Each microsporocarp produces eight motile antherozoids (male gametes) that have flagella, enabling them to approach and fertilise the oosphere (female gamete) produced by the megaspore. This is facilitated by the microsporocarp’s excretion of loose masses of organic material (massulae) that enclose the microspores and antherozoids. The massulae trap air to aid their floatation and extrude anchor-shaped protuberances, called glochidia, that hook onto the megaspore and facilitate its fertilisation.
Megasporocarp floats that emerge from beneath its indusium cap enable the fertilised oosphere to float to the water’s surface where it develops into a seedling. This begins with the formation of an embryonic leaf (cotyledon), after which the young plant develops a root, detaches from the megasporocarp and floats on the water’s surface.
The final stage in N. azollae’s transmission to the next generation of the plant is its entry and enclosure in Azolla’s developing leaf cavities, described by Becking (1987) [21]. This is enabled by conversion of N. azollae into motile hormogonia that move into developing cavities of the dorsal leaves where they change into sessile filaments with photosynthetic vegetative cells and nitrogen-fixing heterocysts. The cycle has now been completed. N. azollae has been transmitted to a new generation of Azolla via its female megaspores, enabling their co-evolution for approximately 80 million years into a unique superorganism.
4. Identifying Azolla’s Ancestor
Hall (1969) recorded Parazolla heterotricha just above the base of the Campanian Claggett Formation (approximately 81 Ma) and suggested that it was ancestral to Azolla [39]. This is consistent with estimates based on genetics for the earliest Azolla at 80 Ma and radiometric dating of the oldest observed Azolla fossils at 78.6 Ma (Figure 6) but needs to be confirmed because confident recognition of the earliest azolla is based on its permanent retention of a nitrogen-fixing cyanobiont. Other candidate precursors of Azolla are fossils assigned to Paleoazolla and Parazolla, but all are geologically younger than A. simplex [16, Table 3]. The oldest is Azollopsis coccoides (77.5 Ma) which is 1.1 million years younger than A. simplex (Figure 6), with a Cenomanian (100.5-93.9 Ma) record of Azollopsis polyancyra from South America being re-assigned to the Maastrichtian (72.17-66.04 Ma) [71] (Appendix A).
Detailed sampling of the Claggett and Judith River Formations could provide a record of events that led to the earliest Azolla, including the timing and way in which its ancestor developed a permanent symbiosis with N. azollae‘s ancestor. This depends on the cyanobacteria being transmitted with its megaspores to subsequent generations of the plant, so that the cyanobacteria should occur with megaspores of the earliest Azolla, but not with megaspores of its ancestor. The earliest occurrence of cyanobacteria with Azolla’s megaspores, therefore, pinpoints the precise time when the permanent relationship began. Microscopic observation of cyanobacterial filaments or akinetes in the plant’s megasporocarps would confirm their intergenerational symbiosis.
Geochemical analyses could also identify cyanobacterial biomarkers associated with megaspores of the earliest Azolla. The presence of 2-methylhopane lipids characterises modern cyanobacteria and has been used to infer their oldest occurrence in sediments deposited close to the GOE at about 2,500 Ma [72]. The earliest association of 2-methylhopane biomarkers with Azolla’s megaspores, therefore, indicates their transmission to subsequent plant generations and their co-evolution into a unique nitrogen-fixing superorganism. Upper Cretaceous sediments exposed along the banks of the Judith and Missouri rivers in Montana’s UMRBNM may contain this evidence.
5. Discussion
Azolla’s intergenerational transmission of N. azollae via its megaspore is unknown in any other plant and resulted from a WGD event that occurred 80 million years ago in the western interior of North America. Their subsequent co-evolution converted the cyanobiont into a permanent obligate diazotroph unknown in any other plant, enabling Azolla to undertake sustainable CCS and provide local renewable food, biofertiliser, livestock feed and biofuel globally – essentials that are urgently needed by today’s population of more than eight billion people.
Appendix A. Questionable pre-Campanian records of Azolla and Azollopsis
The oldest confident occurrence of Azolla is in Campanian rocks of the western interior of North America [39]. Questionable pre-Campanian occurrences of Azolla and Azollopsis are listed below from youngest to oldest age.
Azolla sp.: Turonian of Kazakhstan, central Asia [41]. Review paper without original data for occurrence of Azolla sp.
Azolla sp.: Turonian of Kazakhstan, central Asia [40]. Poor control for the Turonian age assignment.
Azolla polyancyra, now Azollopsis polyancyra Hall (Sweet & Hills, 1974) [73]: Cenomanian of Argentina [71]. The Cerro Fortaleza Formation that contains A. polyancyrea has been re-assigned to the Maastrichtian [74].
Azolla hamata Srivastava (1968) [75]: Cenomanian of Andhra Pradesh, India [42]. Data from a subsurface well without cores. Azolla probably caved from younger strata.
Azolla circinata Hall (1968) [76] and Azolla pondicherriana Ramanujam (1996) [43]: Albian of Tamil Nadu, India. Incorrect Albian age assignment. Probable Maastrichtian age.
Azolla sp.: Albian of Tajikistan, central Asia [77]. Review paper without original data for occurrence of Azolla sp.
Azolla sp.: Albian of Turkmenistan, central Asia [45]. Review paper without original data for occurrence of Azolla sp.
Azolla sp.: Valanginian of the Zabaykalsky Krai region, eastern Russia [47]. Review paper without original data for occurrence of Azolla sp.
Azolla sp.: Valanginian of the Buryatia Republic, south-central Siberia, Russia [50]. Review paper without original data for occurrence of Azolla sp.
Azolla sp.: Middle Jurassic of Ukraine [46]. Review paper without original data for occurrence of Azolla sp.
Azolla sp.: Middle Jurassic of Kazakhstan, central Asia [44]. Incorrect identification of specimens assigned to Azolla.
Azolla sp.: Middle Jurassic of Orenburg Oblast, eastern Europe, Russia [48]. Incorrect identification of specimens assigned to Azolla.
Azolla sp. and Azollopsis sp.: Early Jurassic to Early Cretaceous. No region specified, probably eastern Russia [49]. Incorrect identification of specimens assigned to Azolla.
References
- Pi, H.-W.; Lin, J.-J.; Chen, C.-A.; Wang, P.-H.; Chiang, Y.-R.; Huang, C.-C.; Young, C.-C.; Li, W.-H. Origin and Evolution of Nitrogen Fixation in Prokaryotes. Mol. Biol. Evol. 2022, 39, msac181. [Google Scholar] [CrossRef] [PubMed]
- Stüeken, E.E.; Buick, R.; Guy, B.M.; Koehler, M.C. Isotopic Evidence for Biological Nitrogen Fixation by Molybdenum-Nitrogenase from 3.2 Gyr. Nature 2015, 520, 666–669. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, I.; Berja, N.S. The Growth of Four Species of Azolla as Affected by Temperature. Aquat. Bot. 1983, 15, 175–185. [Google Scholar] [CrossRef]
- Bujak, A.; Bujak, J. Azolla’s Use as a Biofertilizer and Livestock Feed. In Ferns; Marimuthu, J., Fernández, H., Kumar, A., Thangaiah, S., Eds.; Springer Nature Singapore: Singapore, 2022; pp. 671–695. ISBN 9789811661693. [Google Scholar]
- Ansari, M.A.; Sharma, V.P. Role of Azolla in Controlling Mosquito Breeding in Ghaziabad District Villages (U.P.). Indian J. Malariol. 1991, 28, 51–54. [Google Scholar] [PubMed]
- Mwingira, V.; Mayala, B.; Senkoro, K.; Rumisha, S.; Shayo, H., Elizabeth; Mlozi, P.; Mboera, L. Mosquito Larval Productivity in Rice-Fields Infested with Azolla in Mvomero District, Tanzania. Tanzan. J. Health Res. 2009, 11, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, R.; Reuben, R. Evaluation of the Water Fern Azolla Microphylla for Mosquito Population Management in the Rice-Land Agro-Ecosystem of South India. Med. Vet. Entomol. 1991, 5, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Bharati, K. Influence of Incorporation or Dual Cropping of Azolla on Methane Emission from a Flooded Alluvial Soil Planted to Rice in Eastern India. Agric. Ecosyst. Amp Environ. 2000. [Google Scholar] [CrossRef]
- Mujiyo, Sunarminto, B.; Hanudin, E.; Widada, J.; Syamsiyah, J. Methane Emission on Organic Rice Experiment Using Azolla. Int. J. Appl. Environ. Sci. 2016, 11, 295–308.
- Xu, H.; Zhu, B.; Liu, J.; Li, D.; Yang, Y.; Zhang, K.; Jiang, Y.; Hu, Y.; Zeng, Z. Azolla Planting Reduces Methane Emission and Nitrogen Fertilizer Application in Double Rice Cropping System in Southern China. Agron. Sustain. Dev. 2017, 37, 29. [Google Scholar] [CrossRef]
- Bujak, J.; Bujak, A. The Azolla Story: A Message from the Future.; The Azolla Foundation, 2020; ISBN 1-5272-8335-6.
- Winstead, D.; Di Gioia, F.; Jauregui, M.; Jacobson, M. Nutritional Properties of Raw and Cooked Azolla Caroliniana Willd., an Aquatic Wild Edible Plant. Food Sci. Nutr. 2024, 12, 2050–2060. [Google Scholar] [CrossRef]
- Carrapiço, F. Azolla as a Superorganism. Its Implication in Symbiotic Studies. In Symbioses and Stress: Joint Ventures in Biology; Seckbach, J., Grube, M., Eds.; Springer Netherlands: Dordrecht, 2010; pp. 225–241. ISBN 978-90-481-9449-0. [Google Scholar]
- Carrapico, F. Azolla and Bougainville’s Voyage Around the World. In Current Advances in Fern Research; 2018; pp. 251–267 ISBN 978-3-319-75102-3.
- Saunders, R.M.K.; Fowler, K. The Supraspecific Taxonomy and Evolution of the Fern Genus Azolla (Azollaceae). 1993. [CrossRef]
- De Benedetti, F.; Zamaloa, M.D.C.; Gandolfo, M.A.; Cúneo, N.R. Reinterpretation of Paleoazolla: A Heterosporous Water Fern from the Late Cretaceous of Patagonia, Argentina. Am. J. Bot. 2020, 107, 1054–1071. [Google Scholar] [CrossRef] [PubMed]
- Li, F.-W.; Brouwer, P.; Carretero-Paulet, L.; Cheng, S.; de Vries, J.; Delaux, P.-M.; Eily, A.; Koppers, N.; Kuo, L.-Y.; Li, Z.; et al. Fern Genomes Elucidate Land Plant Evolution and Cyanobacterial Symbioses. Nat. Plants 2018, 4, 460–472. [Google Scholar] [CrossRef] [PubMed]
- Metzgar, J.S.; Schneider, H.; Pryer, K.M. Phylogeny and Divergence Time Estimates for the Fern Genus Azolla (Salviniaceae). Int. J. Plant Sci. 2007, 168, 1045–1053. [Google Scholar] [CrossRef]
- Singh, P.; Khan, A.; Srivastava, A. Chapter 16 - Heterocyst and Akinete Differentiation in Cyanobacteria: A View toward Cyanobacterial Symbiosis. In Advances in Cyanobacterial Biology; Singh, P.K., Kumar, A., Singh, V.K., Shrivastava, A.K., Eds.; Academic Press, 2020; pp. 235–248 ISBN 978-0-12-819311-2.
- Malatinszky, D.; Steuer, R.; Jones, P.R. A Comprehensively Curated Genome-Scale Two-Cell Model for the Heterocystous Cyanobacterium Anabaena Sp. PCC 7120. Plant Physiol. 2017, 173, 509–523. [Google Scholar] [CrossRef] [PubMed]
- Becking, J.H. Endophyte Transmission and Activity in the Anabaena-Azolla Association. Plant Soil 1987, 100, 183–212. [Google Scholar] [CrossRef]
- Watzer, B.; Forchhammer, K.; Watzer, B.; Forchhammer, K. Cyanophycin: A Nitrogen-Rich Reserve Polymer. In Cyanobacteria; IntechOpen, 2018 ISBN 978-1-78923-705-4.
- Hodgskiss, M.S.W.; Crockford, P.W.; Peng, Y.; Wing, B.A.; Horner, T.J. A Productivity Collapse to End Earth’s Great Oxidation. Proc. Natl. Acad. Sci. U. S. A. 2019, 116, 17207–17212. [Google Scholar] [CrossRef]
- Knoll, A.H.; Nowak, M.A. The Timetable of Evolution. Sci. Adv. 2017, 3, e1603076. [Google Scholar] [CrossRef] [PubMed]
- Schirrmeister, B.E.; de Vos, J.M.; Antonelli, A.; Bagheri, H.C. Evolution of Multicellularity Coincided with Increased Diversification of Cyanobacteria and the Great Oxidation Event. Proc. Natl. Acad. Sci. 2013, 110, 1791–1796. [Google Scholar] [CrossRef] [PubMed]
- Campbell, E.L.; Christman, H.; Meeks, J.C. DNA Microarray Comparisons of Plant Factor- and Nitrogen Deprivation-Induced Hormogonia Reveal Decision-Making Transcriptional Regulation Patterns in Nostoc Punctiforme. J. Bacteriol. 2008, 190, 7382–7391. [Google Scholar] [CrossRef] [PubMed]
- Meeks, J.C. Symbiosis between Nitrogen-Fixing Cyanobacteria and Plants: The Establishment of Symbiosis Causes Dramatic Morphological and Physiological Changes in the Cyanobacterium. BioScience 1998, 48, 266–276. [Google Scholar] [CrossRef]
- Risser, D.D. Hormogonium Development and Motility in Filamentous Cyanobacteria. Appl. Environ. Microbiol. 2023, 89, e0039223. [Google Scholar] [CrossRef] [PubMed]
- Crow, K.D.; Wagner, G.P.; SMBE Tri-National Young Investigators Proceedings of the SMBE Tri-National Young Investigators’ Workshop 2005. What Is the Role of Genome Duplication in the Evolution of Complexity and Diversity? Mol. Biol. Evol. 2006, 23, 887–892. [Google Scholar] [CrossRef] [PubMed]
- Ekman, M.; Tollbäck, P.; Klint, J.; Bergman, B. Protein Expression Profiles in an Endosymbiotic Cyanobacterium Revealed by a Proteomic Approach. Mol. Plant-Microbe Interactions® 2006, 19, 1251–1261. [Google Scholar] [CrossRef]
- Ekman, M.; Tollbäck, P.; Bergman, B. Proteomic Analysis of the Cyanobacterium of the Azolla Symbiosis: Identity, Adaptation, and NifH Modification. J. Exp. Bot. 2008, 59, 1023–1034. [Google Scholar] [CrossRef] [PubMed]
- Larsson, J.; Nylander, J.A.; Bergman, B. Genome Fluctuations in Cyanobacteria Reflect Evolutionary, Developmental and Adaptive Traits. BMC Evol. Biol. 2011, 11, 187. [Google Scholar] [CrossRef] [PubMed]
- Ran, L.; Larsson, J.; Vigil-Stenman, T.; Nylander, J.A.A.; Ininbergs, K.; Zheng, W.-W.; Lapidus, A.; Lowry, S.; Haselkorn, R.; Bergman, B. Genome Erosion in a Nitrogen-Fixing Vertically Transmitted Endosymbiotic Multicellular Cyanobacterium. PLoS ONE 2010, 5, e11486. [Google Scholar] [CrossRef]
- Watanabe, I. Azolla-Symbiosis - Its Physiology and Use in Tropical Agriculture. In Microbiology of Tropical Soils and Plant Productivity; Dommergues, Y.R., Diem, H.G., Eds.; Springer Netherlands: Dordrecht, 1982; pp. 169–185. ISBN 978-94-009-7529-3. [Google Scholar]
- Bergman, B.; Johansson, C.; Söderbäck, E. The Nostoc-Gunnera Symbiosis. New Phytol. 1992, 122, 379–400. [Google Scholar] [CrossRef] [PubMed]
- White, J. Palynodata Datafile: 2006 Version. Geol. Surv. Can. Open File 2008, 5793. [Google Scholar]
- Brinkhuis, H.; Schouten, S.; Collinson, M.E.; Sluijs, A.; Damsté, J.S.S.; Dickens, G.R.; Huber, M.; Cronin, T.M.; Onodera, J.; Takahashi, K. Episodic Fresh Surface Waters in the Eocene Arctic Ocean. Nature 2006, 441, 606–609. [Google Scholar] [CrossRef]
- Collinson, M.E.; Barke, J.; van der Burgh, J.; van Konijnenburg-van Cittert, J.H.A. A New Species of the Freshwater Fern Azolla (Azollaceae) from the Eocene Arctic Ocean. Rev. Palaeobot. Palynol. 2009, 155, 1–14. [Google Scholar] [CrossRef]
- Hall, J.W. Studies on Fossil Azolla; Primitive Types of Megaspores and Massulae from the Cretaceous. Am. J. Bot. 1969, 56, 1173–1180. [Google Scholar] [CrossRef]
- Boytsova, E.P. Upper Cretaceous Spore-Pollen Complexes of the Turgay Depressions. (In: Atlas of Upper Cretaceous, Paleocene and Eocene Spore-Pollen Complexes of Various Regions of the USSR). [ Verkhne Melovye Sporovo-Pyl’tsevye Kompleksy Turgayskogo Progiba. (In: Atlas Verkhne Melovykh Paleotsenorvykh i Eotsenovykh Sporov Pyl’tsevykh Kompleksov Nekotorykh Rayonov SSSR.). Tr. VSEGEI Novaya SeriyaLeningrad 1960, 30, 28–49. [Google Scholar]
- Romanovskaya, G.M.; Stelmak, N.K. Short Description of Spores and Pollen from Upper Cretaceous Deposits of the Turgay Basin. (In: Atlas of Upper Cretaceous, Paleocene and Eocene Spore-Pollen Complexes of Various Regions of the USSR). [ Kratkoe Opisanie Spor i Pyl’tsy Iz Verkhnemelovykh Otlozheniy Turgayskog Progiba. (In: Atlas Verkhne Melovykh Paleotsenorvykh i Eotsenovykh Sporov Pyl’tsevykh Kompleksov Nekotorykh Rayonov SSSR.). Tr. VSEGEI Novaya SeriyaLeningrad 1960, 30, 200–269. [Google Scholar]
- Juyal, N.P.; Sharma, J.; Chopra, A.S.; Misra, C.M. Palynostratigraphy and Paleoenvironment Analysis of the Mesozoic - Cenozoic Succession of the Wells Bhavadevarapalli, Bobbarlanka and Peddapalem in Krishna Delta. Contrib. XV Indian Colloq. Micropaleontol. Stratigr. 1996, 607–614. [Google Scholar]
- Ramanujam, C.J.K. Remain of Azollaceae from Late Albian of Cauvery Basin and Their Evolutionary Significance. Geophytology 1996, 25, 105–111. [Google Scholar]
- Faddeeva, I.Z. Palynological Characteristics of Lower Mesozoic Coal Deposits of Kazakhstan. (In: History of Lower Mesozoic Coal Accumulation in Kazakhstan. Part 3) [Palinologicheshaya Kharacteristika Nizhnemezozoic Kikh Uglenoshykh Otlozhenii Kazakhstana (In-Istoriya Nizhnemezozoiskogo Uglenakopleniya v Kazakastane]. Tr. Lab. Geol. UglyaAkademiya Nauk SSR 1963, 20, 143–185. [Google Scholar]
- Panova, L.A. Lower Cretaceous Spore-Pollen Complexes of Central Asia. (In: Atlas of Lower Cretaceous Spore-Pollen Complexes of Some Regions of USSR. I.M.Pokyovsksys and N.K.Stel’mak, Editors) [Nizhnemelovye Sporovo-Pyl’tsevye Kompleksy Sredney Azii. (In: Atlas Nizhnemelovykh Sporovo-Pyl’tsevykh Kompleksov Nekotorykh Reyonov SSSR. I.M.Pokyovsksys and N.K.Stel’mak,Editors)]. Tr. VSEGEI Novaya Seriya 1964, 124, 40–48. [Google Scholar]
- Andreeva, E.M.; Boytsova, E.P.; Koltsova, T.T.; Komarova, N.O.; Kruchinina, N.V.; Lyuber, A.A. Palaeopalynology. (Three Volumes: Volume I: Methods. Volume II: Complexes of Palynomorphs of the Precambrian-Holocene of the USSR. Volume III: Figures and Plates. 1966, 141. [Google Scholar]
- Bondarenko, N.M.; Bocharnikova, A.D.; Kara-Murza, E.N.; Koltsova, T.T.; Kopytova, E.A. A List of Plant Spores and Pollen of Valanginian Deposits in Some Districts of the USSR. (In: Atlas of Lower Cretaceous Spore-Pollen Complexes of Some Regions of USSR. I.M.Pokyovsksys and N.K.Stel’mak, Editors) [Spisok Rastenii Spory i Pyl’tsa Kotorykh Vstrecheny v Valankhinskikh i Valankhii-Goterivski Kh Otlozheniyakh Nekotorykh Raionov SSSR. (In: Atlas Nizhnemelovykh Sporovo-Pyl’tsevykh Kompleksov Nekotorykh Reyonov SSSR. I.M.Pokyovsksys and N.K.Stel’mak,Editors)]. Tr. VSEGEI Novaya Seriya 1964, 124, 322–339. [Google Scholar]
- Faddeeva, I.Z. Part 5. Lower Mesozoic Megaspores of the Or-Ilek Region and Their Stratigraphic Significance. (In: Palynological Validation of the Stratigraphic Division of the Lower Mesozoic Coal-Bearing Deposits of Or’-Ilek Region.) [5.Nizhnemezozoiskie Megaspory Or’-Ilekskogo Raiona i Ikh Stratigraficheskoe Znachenie In(Palinologischeskoe Obosnovanie Stratigraficheskogo Raschlene Nizhnemezozoickikh Uglenosnykh Otlozhenii Or’-Ilekskogo Raiona]. Geol. InstitutOtdelenie Geol. Uglya Goryuchikh SlantsevAkademiya Nauk SSRVNIGI 1965, 1–116.
- Petrosyants, M.A. A Study of Microfossils for Revealing Old Sedimentation Environment [Issledovanie Mikrofossilii Dlya Byyableniya Obstanovok Drevnego Osadkonakopleniya. Izv. Akad. Nauk SSSR Seriya Geol. 1984, 1984, 73–79. [Google Scholar]
- Sedova, M.A. Description of Spores and Pollen in Neocomian Deposits of the Vitimsk Plateau and Transbaikal. (In: Atlas of Lower Cretaceous Spore-Pollen Complexes of Some Regions of USSR. I.M.Pokyovsksys and N.K.Stel’mak, Editors) [Opisanie Spor i Pyl’tsy Iz Nedkomskikh Otlozhenii Vitimskogo Plostogor’Ya i Zabaikal’Ya. (In: Atlas Nizhnemelovykh Sporovo-Pyl’tsevykh Kompleksov Nekotorykh Reyonov SSSR. I.M.Pokyovsksys and N.K.Stel’mak,Editors)]. Tr. VSEGEI Novaya Seriya 1964, 124, 280–295. [Google Scholar]
- Freeman, M. Scouting Magazine. Available online: https://scoutingmagazine.org/2019/08/paddling-in-the-wake-of-lewis-and-clark/ (accessed on 24 May 2024).
- Gradstein, F.M.; Ogg, J.G.; Schmitz, D.; Ogg, M. Geologic Time Scale 2020 - 1st Edition; Elsevier, 2020; ISBN 978-0-12-824360-2.
- Rogers, R.R.; Kidwell, S.M.; Deino, A.L.; Mitchell, J.P.; Nelson, K.; Thole, J.T. Age, Correlation, and Lithostratigraphic Revision of the Upper Cretaceous (Campanian) Judith River Formation in Its Type Area (North-Central Montana), with a Comparison of Low- and High-Accommodation Alluvial Records. J. Geol. 2016, 124, 99–135. [Google Scholar] [CrossRef]
- Ramezani, J.; Beveridge, T.L.; Rogers, R.R.; Eberth, D.A.; Roberts, E.M. Calibrating the Zenith of Dinosaur Diversity in the Campanian of the Western Interior Basin by CA-ID-TIMS U-Pb Geochronology. Sci. Rep. 2022, 12, 16026. [Google Scholar] [CrossRef]
- Blakey, R. Deep Time MapsTM. Available online: https://deeptimemaps.com/ (accessed on 24 May 2024).
- Dalman, S.; Lucas, S. Tyrannosaurid Teeth from the Claggett Formation of the Elk Basin, Late Cretaceous of Western North America. N. M. Mus. Nat. Hist. Sci. Bull. 2016, 71, 83–89. [Google Scholar]
- Paerl, H.W.; Gardner, W.S.; McCarthy, M.J.; Peierls, B.L.; Wilhelm, S.W. Algal Blooms: Noteworthy Nitrogen. Science 2014, 346, 175–175. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.G.; Bergman, B.; Nierzwicki-Bauer, S.A.; Rai, A.N.; Schüßler, A. Cyanobacterial-Plant Symbioses. In The Prokaryotes: Volume 1: Symbiotic associations, Biotechnology, Applied Microbiology; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, 2006; pp. 331–363. ISBN 978-0-387-30741-1. [Google Scholar]
- Adams, D.G.; Duggan, P.S.; Jackson, O. Cyanobacterial Symbioses. In Ecology of Cyanobacteria II: Their Diversity in Space and Time; Whitton, B.A., Ed.; Springer Netherlands: Dordrecht, 2012; pp. 593–647. ISBN 978-94-007-3855-3. [Google Scholar]
- Adams, D.G.; Duggan, P.S. Cyanobacteria–Bryophyte Symbioses. J. Exp. Bot. 2008, 59, 1047–1058. [Google Scholar] [CrossRef] [PubMed]
- Nicoletti, M. Chapter 10 - The Nutraceutical Potential of Cyanobacteria. In The Pharmacological Potential of Cyanobacteria; Lopes, G., Silva, M., Vasconcelos, V., Eds.; Academic Press, 2022; pp. 287–330 ISBN 978-0-12-821491-6.
- Chiu, W.-L.; Peters, G.A.; Levieille, G.; Still, P.C.; Cousins, S.; Osborne, B.; Elhai, J. Nitrogen Deprivation Stimulates Symbiotic Gland Development in Gunnera Manicata. Plant Physiol. 2005, 139, 224–230. [Google Scholar] [CrossRef]
- Conradi, F.D.; Mullineaux, C.W.; Wilde, A. The Role of the Cyanobacterial Type IV Pilus Machinery in Finding and Maintaining a Favourable Environment. Life Basel Switz. 2020, 10, 252. [Google Scholar] [CrossRef] [PubMed]
- Faluweki, M.K.; Goehring, L. Structural Mechanics of Filamentous Cyanobacteria. J. R. Soc. Interface 2022, 19, 20220268. [Google Scholar] [CrossRef] [PubMed]
- Khayatan, B.; Meeks, J.C.; Risser, D.D. Evidence That a Modified Type IV Pilus-like System Powers Gliding Motility and Polysaccharide Secretion in Filamentous Cyanobacteria. Mol. Microbiol. 2015, 98, 1021–1036. [Google Scholar] [CrossRef] [PubMed]
- Wilde, A.; Mullineaux, C.W. Motility in Cyanobacteria: Polysaccharide Tracks and Type IV Pilus Motors. Mol. Microbiol. 2015, 98, 998–1001. [Google Scholar] [CrossRef] [PubMed]
- Campbell, E.L.; Wong, F.C.Y.; Meeks, J.C. DNA Binding Properties of the HrmR Protein of Nostoc Punctiforme Responsible for Transcriptional Regulation of Genes Involved in the Differentiation of Hormogonia. Mol. Microbiol. 2003, 47, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Splitt, S. Sucrose Analog Sucralose Is Potent Inhibitor of Hormogonium Differentiation in Nostoc Punctiforme. Pac. Undergrad. Res. Creat. Conf. PURCC 2015. [Google Scholar]
- Cohen, M.F.; Sakihama, Y.; Takagi, Y.C.; Ichiba, T.; Yamasaki, H. Synergistic Effect of Deoxyanthocyanins from Symbiotic Fern Azolla Spp. on hrmA Gene Induction in the Cyanobacterium Nostoc Punctiforme. Mol. Plant-Microbe Interact. MPMI 2002, 15, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Bergman, B.; Chen, B.; Zheng, S.; Xiang, G.; Rasmussen, U. Cellular Responses in the Cyanobacterial Symbiont during Its Vertical Transfer between Plant Generations in the Azolla Microphylla Symbiosis. New Phytol. 2009, 181, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Stough, J.B. Palynomorphs from South America | KU Biodiversity Institute and Natural History Museum. Univ. Kans. Publ. 1968, 32, 1–12. [Google Scholar]
- Hoshino, Y.; Nettersheim, B.J.; Gold, D.A.; Hallmann, C.; Vinnichenko, G.; van Maldegem, L.M.; Bishop, C.; Brocks, J.J.; Gaucher, E.A. Genetics Re-Establish the Utility of 2-Methylhopanes as Cyanobacterial Biomarkers before 750 Million Years Ago. Nat. Ecol. Evol. 2023, 7, 2045–2054. [Google Scholar] [CrossRef]
- Sweet, A.R.; Hills, L.V. A Detailed Study of the Genus Azollopsis. Can. J. Bot. 1974, 52, 1625–1642. [Google Scholar] [CrossRef]
- Paulina-Carabajal, A.; Barrios, F.T.; Méndez, A.H.; Cerda, I.A.; Lee, Y.-N. A Late Cretaceous Dinosaur and Crocodyliform Faunal Association–Based on Isolate Teeth and Osteoderms–at Cerro Fortaleza Formation (Campanian-Maastrichtian) Type Locality, Santa Cruz, Argentina. PLoS ONE 2021, 16, e0256233. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.K. Azolla from the Upper Cretaceous Edmonton Formation, Alberta, Canada. Can. J. Earth Sci. 1968, 5, 915–919. [Google Scholar] [CrossRef]
- Hall, J.W. A New Genus of Salviniaceae and a New Species of Azolla from the Late Cretaceous. Am. Fern J. 1968, 58, 77–88. [Google Scholar] [CrossRef]
- Boytsova, E.P. Lower Cretaceous Spore-Pollen Complexes of the Central Asia Trans-Urals. >(In: Atlas of Lower Cretaceous Spore-Pollen Complexes of Some Regions of USSR. I.M.Pokyovsksys and N.K.Stel’mak, Editors) [Nizhnemelovye Sporovo-Pyl’tsevye Kompleksy Sredney Azii. (In- Atlas Nizhnemelovykh Sporovo-Pyl’tsevykh Kompleksov Nekotorykh Reyonov SSSR. I.M.Pokyovsksys and N.K.Stel’mak,Editors)]. 1964, 124, 71–81.
Figure 1.
Azolla filiculoides showing its floating leaves and root-like tendrils. Photograph from 'The Azolla Story' [11].
Figure 1.
Azolla filiculoides showing its floating leaves and root-like tendrils. Photograph from 'The Azolla Story' [11].

Figure 2.
Azolla’s CO2 and N2 sequestration (1,2) and its products: (3) Captured carbon for Carbon Capture & Storage, (4) food and livestock feed, (5) biofuel, (6) biofertiliser, (7) nitrogen-enriched water to fertilise other plants. Illustration rendered by Victor Leshyk (https://www.victorleshyk.com/).
Figure 2.
Azolla’s CO2 and N2 sequestration (1,2) and its products: (3) Captured carbon for Carbon Capture & Storage, (4) food and livestock feed, (5) biofuel, (6) biofertiliser, (7) nitrogen-enriched water to fertilise other plants. Illustration rendered by Victor Leshyk (https://www.victorleshyk.com/).
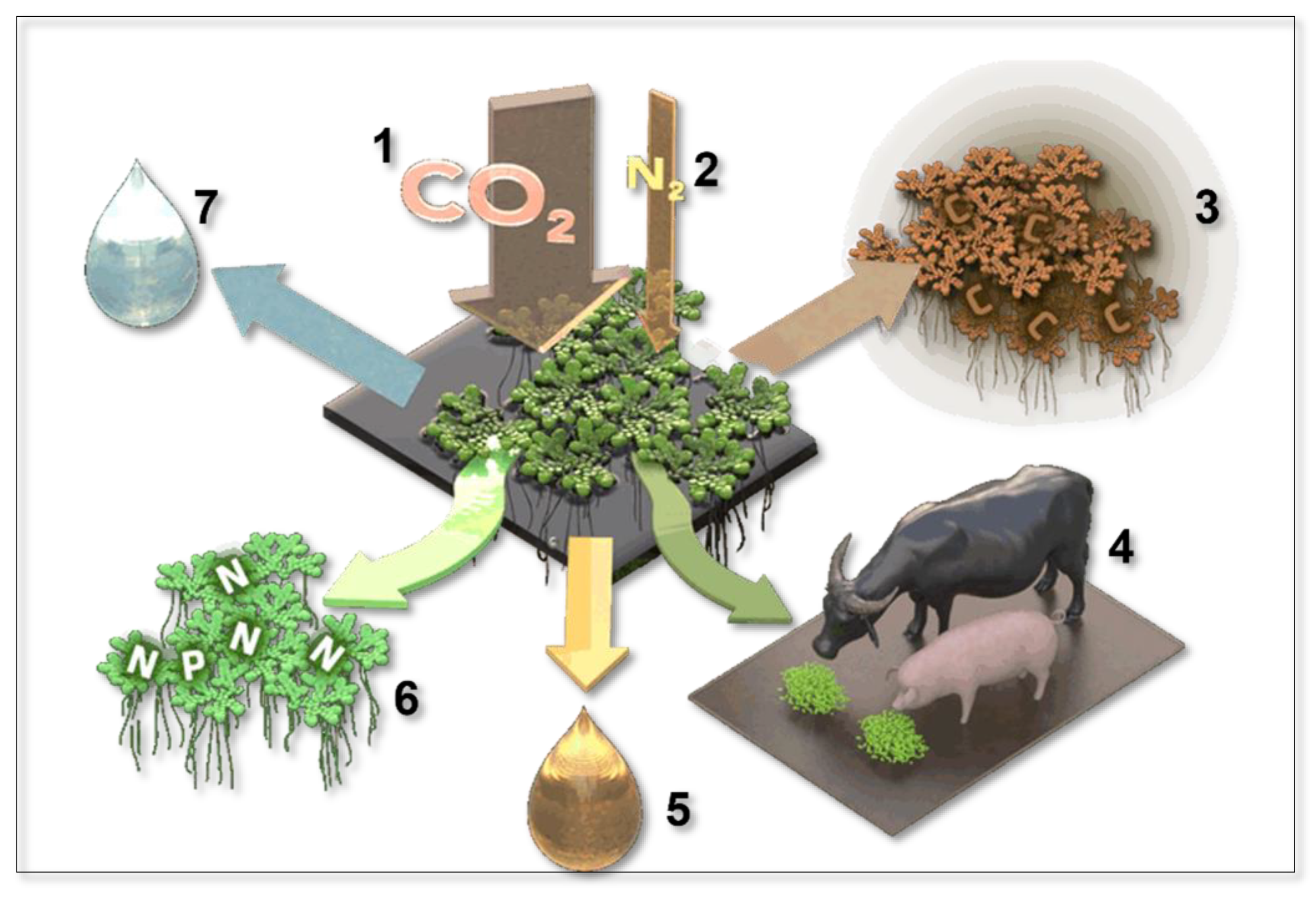
Figure 3.
Nostoc azollae’s vegetative cells and heterocysts showing the complex biochemical pathways used for photosynthesis and nitrogen fixation. Cyanophycin acts as a reservoir for fixed nitrogen in the heterocysts and enables transport of fixed nitrogen to adjacent vegetative cells. Details of N. azollae’s cells from Malatinszky et al. (2017) [20].
Figure 3.
Nostoc azollae’s vegetative cells and heterocysts showing the complex biochemical pathways used for photosynthesis and nitrogen fixation. Cyanophycin acts as a reservoir for fixed nitrogen in the heterocysts and enables transport of fixed nitrogen to adjacent vegetative cells. Details of N. azollae’s cells from Malatinszky et al. (2017) [20].
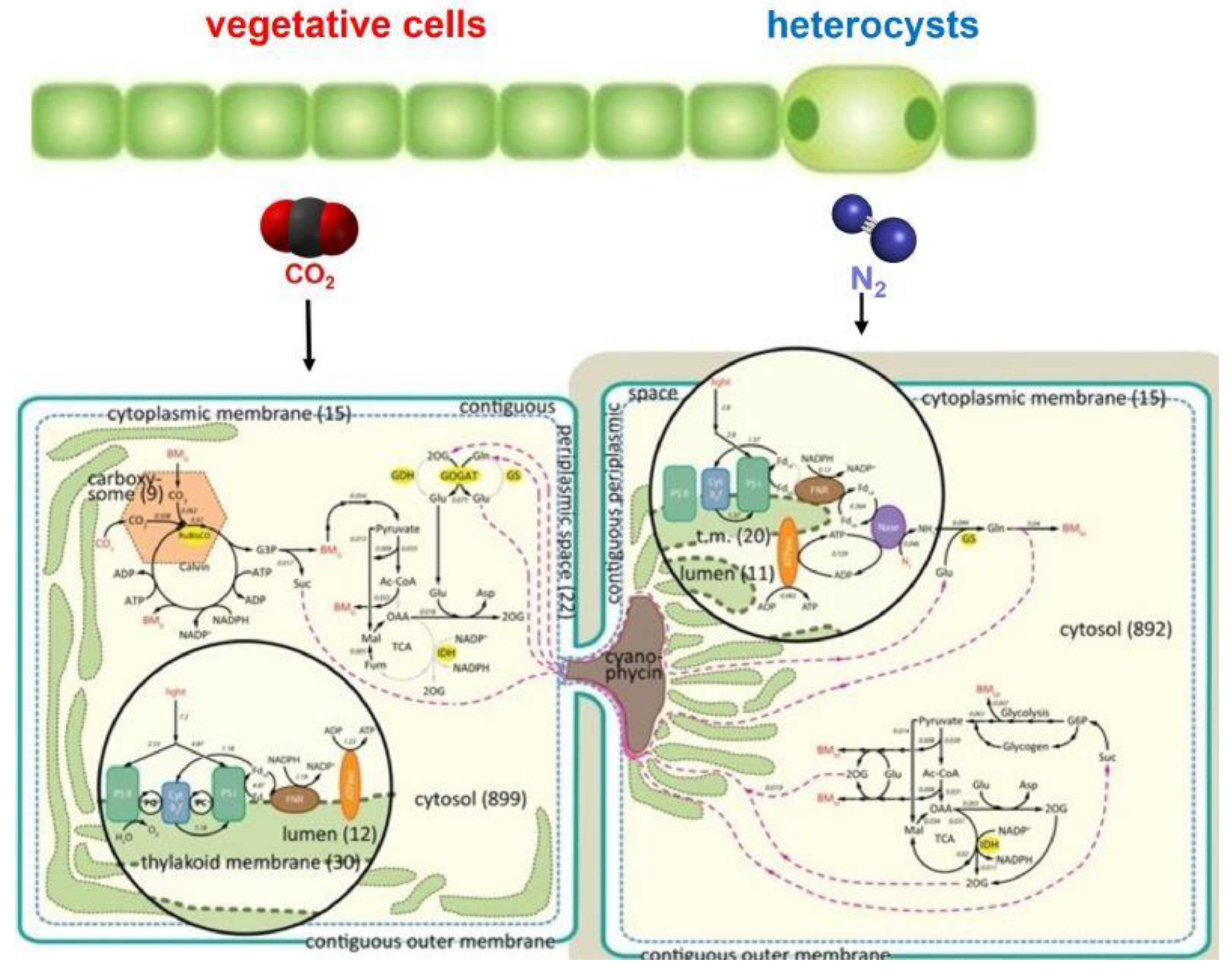
Figure 4.
Genome size of the Order Salviniales (water ferns), divergence times and two Whole Genome Duplication (WGD) events of the Salviniales based on phylogenetic mapping. Geological scale on the left is in millions of years. Figure redrawn and expanded from Li et al. (2018) [17].
Figure 4.
Genome size of the Order Salviniales (water ferns), divergence times and two Whole Genome Duplication (WGD) events of the Salviniales based on phylogenetic mapping. Geological scale on the left is in millions of years. Figure redrawn and expanded from Li et al. (2018) [17].
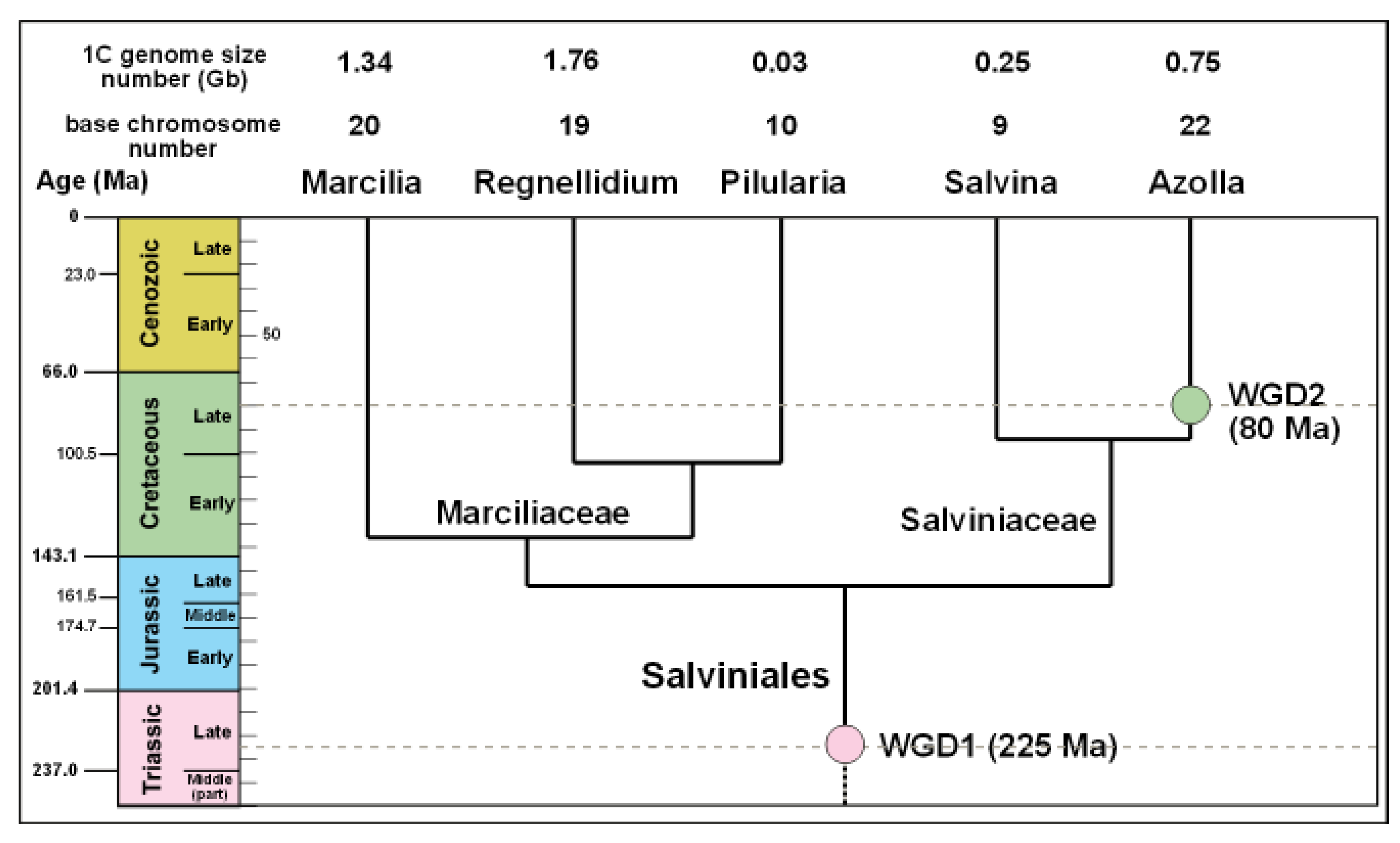
Figure 5.
Location of rocks sampled by Hall (1969) [39] containing (1) Parazolla heterotricha and (2) Azolla simplex. UMRBNM shown in green. JR = Judith River. Map modified from Scouting Magazine [51].
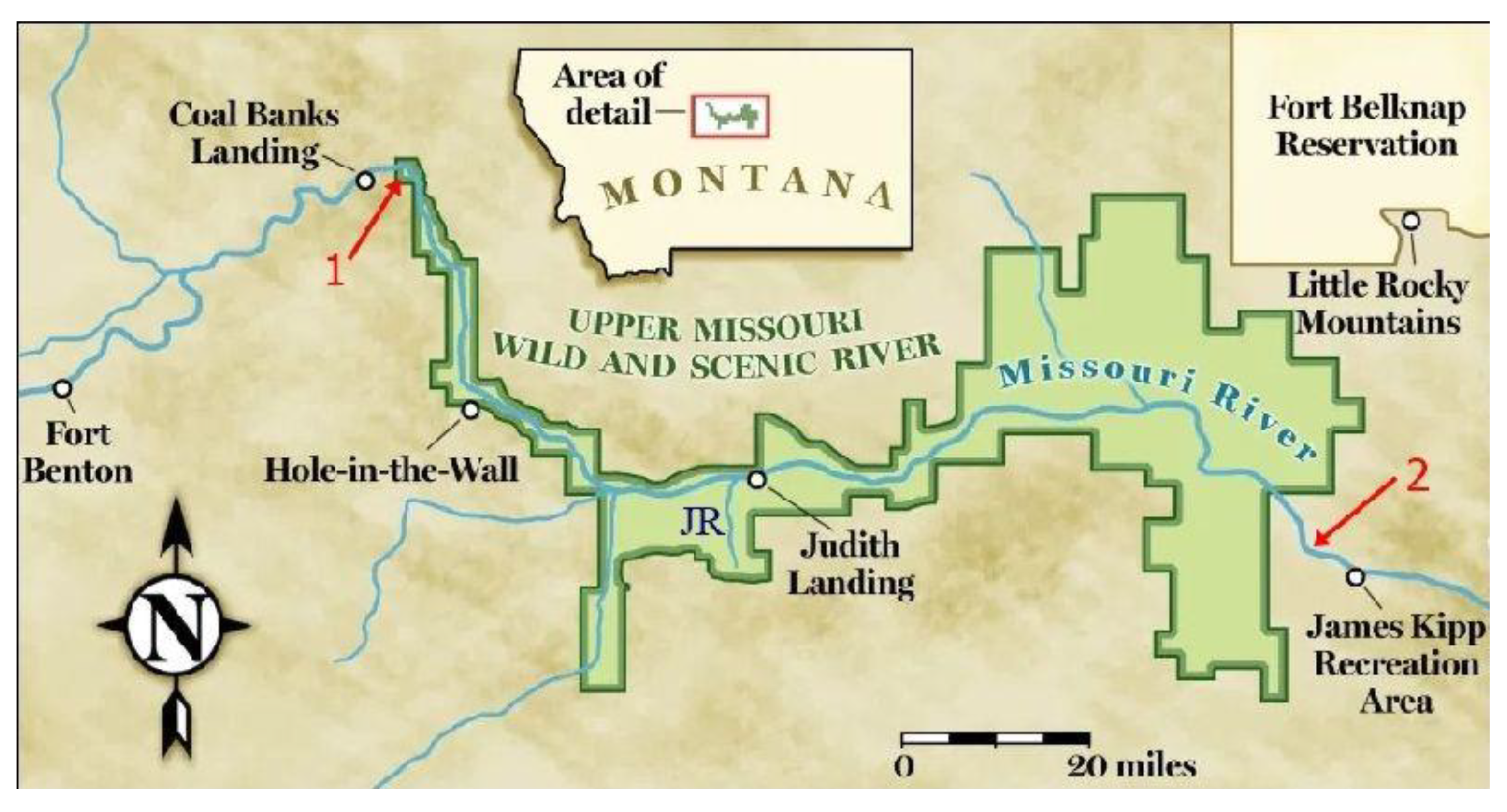
Figure 6.
Absolute ages determined for strata in Alberta, Montana, Utah and New Mexico showing the earliest recorded occurrences of Azolla simplex (As), Parazolla heterotricha (Ph) and Azollopsis coccoides (Ac). Small yellow circles show radiometric dates based on 40Ar/39Ar isotopes. The 80 Ma age indicated by genome analysis for the oldest Azolla is circled in red. Figure modified from Ramezani et al. (2022) [54] to include details of taxon occurrences and absolute ages.
Figure 6.
Absolute ages determined for strata in Alberta, Montana, Utah and New Mexico showing the earliest recorded occurrences of Azolla simplex (As), Parazolla heterotricha (Ph) and Azollopsis coccoides (Ac). Small yellow circles show radiometric dates based on 40Ar/39Ar isotopes. The 80 Ma age indicated by genome analysis for the oldest Azolla is circled in red. Figure modified from Ramezani et al. (2022) [54] to include details of taxon occurrences and absolute ages.
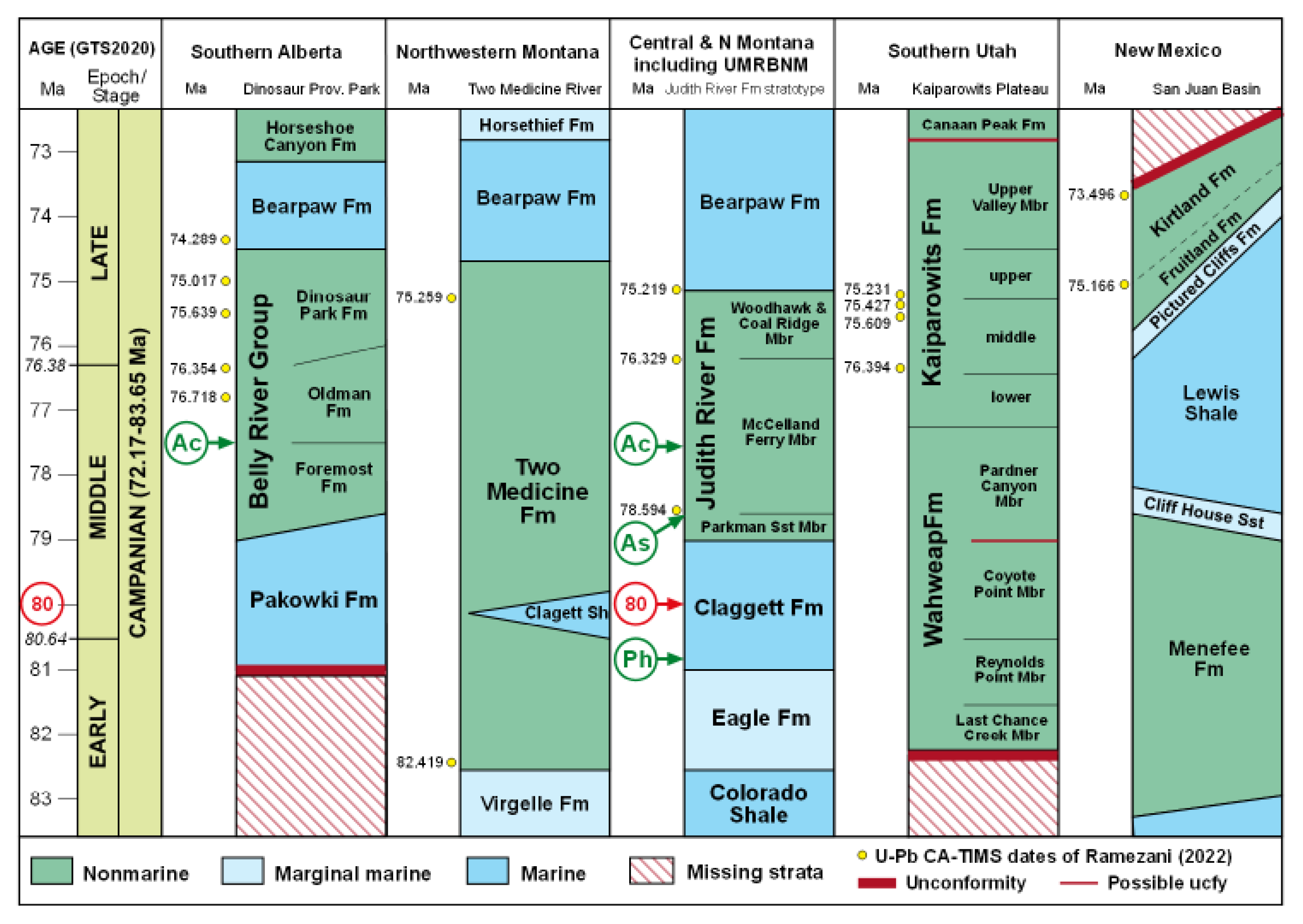
Figure 7.
North America paleogeography at the beginning of the Campanian (83.7 Ma, left) and the end of the Maastrichtian (66.04 Ma, right). Star shows the location of the Yucatan bolide impact. Reconstructions from Deep Time Maps™ [55] ©2020 Colorado Plateau Geosystems, Inc.
Figure 7.
North America paleogeography at the beginning of the Campanian (83.7 Ma, left) and the end of the Maastrichtian (66.04 Ma, right). Star shows the location of the Yucatan bolide impact. Reconstructions from Deep Time Maps™ [55] ©2020 Colorado Plateau Geosystems, Inc.
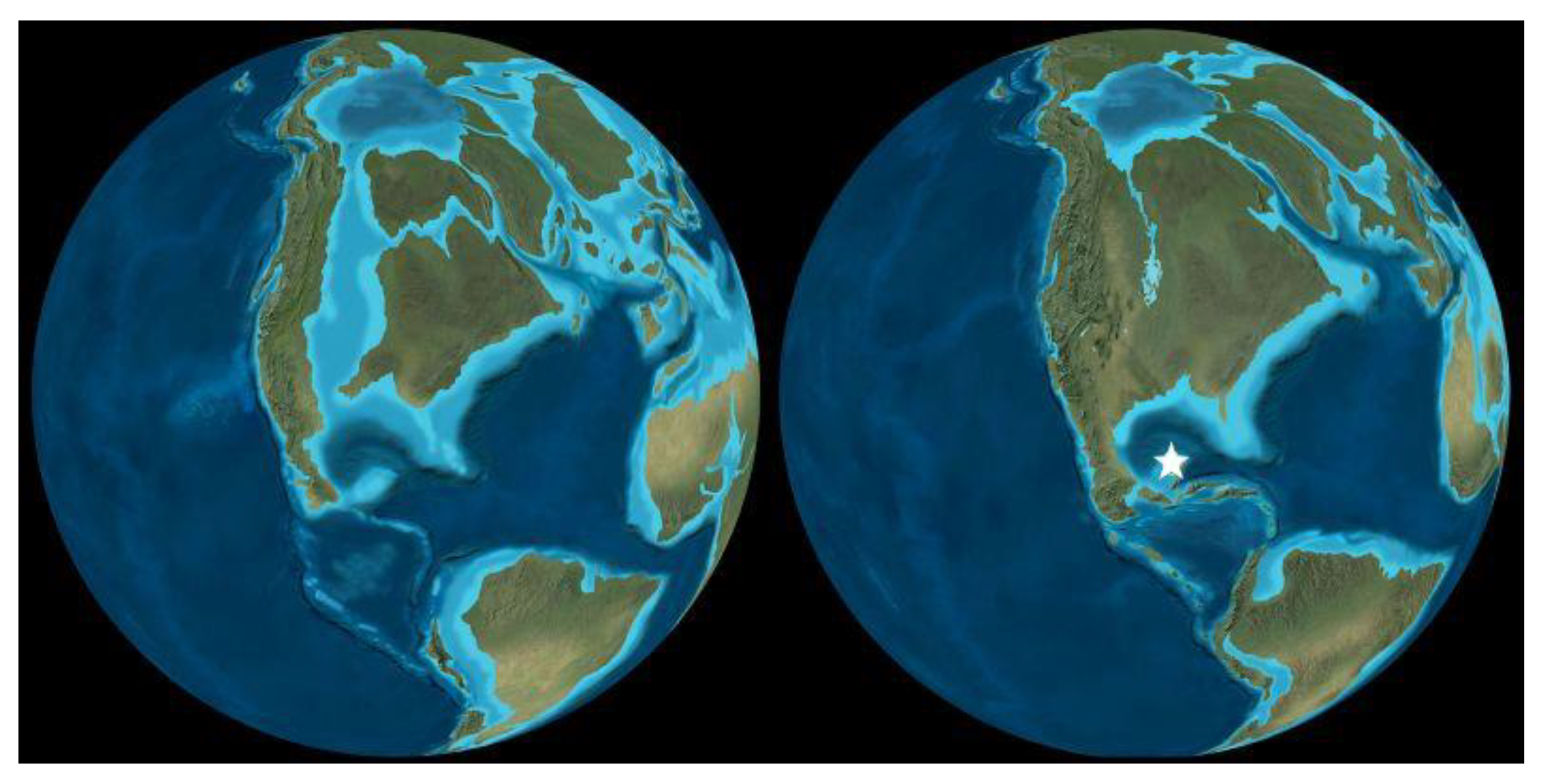
Figure 8.
Azolla’s intergenerational transmission of Nostoc azollae via its megaspores. See text for details.
Figure 8.
Azolla’s intergenerational transmission of Nostoc azollae via its megaspores. See text for details.
Figure 9.
Left: Azolla’s megasporocarp (a,c,e) colonised by N. azollae in the indusium (IND) next to Azolla’s developing megaspore (M), showing N. azollae’s change from filaments (b) to proakinetes (d) to isolated akinetes (f). Upper right: Membrane vesicles (mv) released from N. azollae contain genetic material. Lower right: Isolated akinetes are immobilised inside a structured biofilm that fills the indusium so that they resemble cells separated by channels (c). Cyanophycin granules (cg) in the immobilised akinetes provide reserves of nitrogen, carbon and energy facilitating their dormancy, survival and germination at the same time as Azolla’s megaspore. Illustrations modified from Zheng et al. (2009) [70].
Figure 9.
Left: Azolla’s megasporocarp (a,c,e) colonised by N. azollae in the indusium (IND) next to Azolla’s developing megaspore (M), showing N. azollae’s change from filaments (b) to proakinetes (d) to isolated akinetes (f). Upper right: Membrane vesicles (mv) released from N. azollae contain genetic material. Lower right: Isolated akinetes are immobilised inside a structured biofilm that fills the indusium so that they resemble cells separated by channels (c). Cyanophycin granules (cg) in the immobilised akinetes provide reserves of nitrogen, carbon and energy facilitating their dormancy, survival and germination at the same time as Azolla’s megaspore. Illustrations modified from Zheng et al. (2009) [70].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



