
Submitted:
21 June 2024
Posted:
24 June 2024
You are already at the latest version
Abstract
The injury causes resistance in human gastric cancer (GC). This Adverse Outcome Pathway (AOP) entitled “Increases in cellular reactive oxygen species (ROS) and chronic ROS leading to human treatment-resistant gastric cancer (GC)” consists of molecular initiating event (MIE) as “Increases in cellular ROS” and “Chronic ROS”, followed by series of key events (KEs); “porcupine-induced Wnt secretion and Wnt signaling activation”, “beta-catenin activation”, “epithelial-mesenchymal transition (EMT), and adverse outcome (AO) as “human treatment-resistant GC” in the sequence. ROS has multiple roles in disease such as development and progression of cancer, or apoptotic induction causing anti-tumor effects. In this AOP, we focus on the role of sustained levels of chronic ROS to induce the therapy-resistance in human GC. EMT, which is cellular phenotypic change from epithelial to mesenchymal-like features, demonstrates cancer stem cell (CSC)-like characteristics in human GC. EMT is induced by Wnt/beta-catenin signaling, providing the rationale to have Wnt secretion and beta-catenin activation as KEs in the AOP.
Keywords:
Adverse Outcome Pathway (AOP)
; epithelial-mesenchymal transition
; gastric cancer
; reactive oxygen species (ROS)
; Wnt signaling
1. Introduction
Molecular signaling pathway networks are regulated in epithelial-mesenchymal transition (EMT) and cancer stem cells (CSCs), which exhibit anti-cancer drug-resistant features. To reveal the mechanism of human treatment-resistant gastric cancer (GC) and the relationship between oxidative stress response and GC, analysis on oxidative stress response and molecular networks in diffuse- and intestinal-type GC has been performed. The network pathways of GC were analyzed by Ingenuity Pathway Analysis (IPA). NRF2-mediated oxidative stress response network included molecules related to Regulation of EMT by growth factors pathway and Production of nitric oxide and reactive oxygen species in macrophages such as PI3K and AKT.
This research aims to ensure the safety of therapeutics such as anti-cancer drugs by revealing the molecular mechanisms that contribute to the efficacy and side effects or unexpected and off-targeted adverse effects. Chemicals induce molecular alterations and body responses. Recent progress in cellular and molecular network pathway analysis has revealed the activation mechanism of cellular signal transduction upon cancer and chemical stimulation. In developing anti-cancer drugs such as molecular-targeting therapeutics, identifying target molecules and inhibiting or activating the signaling transduction related to the target molecules is important. Anti-cancer therapeutics targeting Wnt/beta-catenin signaling pathway regulating cell self-renewal, Hedgehog signaling pathway, Notch signaling pathway, and EGFR receptor signaling pathway have been developed and approved, however, off-target effects for molecular network pathways are not fully understood. To elucidate the safety of the molecular-targeted therapeutics and the cellular therapeutics using multipotent stem cells, it is critical to predict the off-target network pathways unexpected. Molecular network pathway analysis utilizing the existing abundant data in databases is needed [1,2]. This study aims to predict the side-effects or adverse effects of the therapeutics by analyzing the molecular network pathway dynamism utilizing the data on databases.
ROS consist of free oxygen radicals such as superoxide, hydroxyl radical, nitric oxide, organic radicals, peroxyl radicals, alkoxyl radicals, thiyl radicals, sulfonyl radicals, thiyl peroxyl radicals, or disulfides, and non-radical ROS such as hydrogen peroxide, singlet oxygen, ozone/trioxygen, organic hydroperoxides, hypochlorite, peroxynitrite, nitrosoperoxycarbonate anion, nitrocarbonate anion, dinitrogen dioxide, nitronium, and highly reactive lipid- or carbohydrate-derived carbonyl compounds [3]. ROS have double-edged effects, which may affect tumorigenesis. ROS play crucial roles in defense of human from infection, whereas excess prolonged ROS cause several diseases including cancer, cardiovascular disease, neurological disease, sensory impairment and psychiatric disease [4]. Nicotinamide adenine diphosphate (NADPH) oxidase catalyzes the production of superoxide by the one-electron reduction of oxygen and produces ROS [5] (Figure 1).
2. Outline of AOP298
2.1. Structure of AOP298
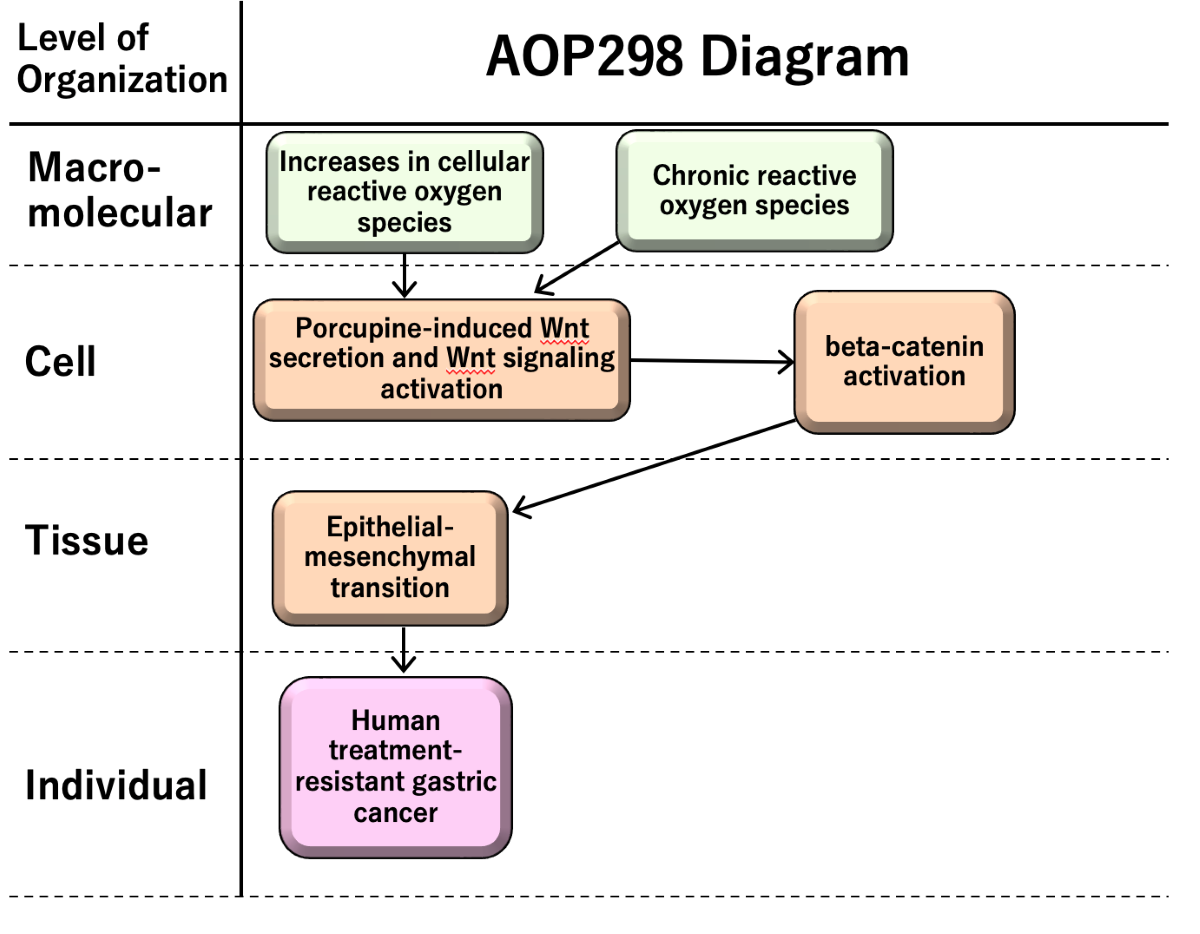
The AOP 298 entitled “Increases in cellular reactive oxygen species (ROS) and chronic ROS leading to human treatment-resistant gastric cancer (GC)” consists of two molecular initiating events; (MIE1; KE1940, Increases in cellular ROS, (MIE2; KE1753, chronic ROS), key events (KEs) as porcupine-induced Wnt secretion and Wnt signaling activation (KE1; KE1754), beta-catenin activation (KE2; KE1755), and epithelial-mesenchymal transition (EMT) (KE3; KE1457), and adverse outcome (AO; KE1651) as human treatment-resistant GC (Figure 2).
While ROS have both benefits and risks in human health, chronic ROS, where excess of ROS prolongs for a long time induces sustained tissue damage, macrophage activation. Porcupine-induced Wnt secretion in macrophages induces proliferation and beta-catenin activation leading to epithelial-mesenchymal transition (EMT). EMT induces cancer migration and drug resistance, causing human treatment-resistant gastric cancer.
AOP298-related information is summarized in Table1.
2.2. Summary of scientific evidence assessment
2.2.1. MIE1; KE1940: Increases in cellular reactive oxygen species (ROS)
Increases in cellular ROS are observed when cells are exposed to various stressors as allergens, ionizing radiation and chemicals [6]. The ROS includes free radicals (e.g., Superoxide anion, hydroxyl radical, nitric oxide, nitrogen dioxide, organic radicals, peroxyl radicals, alkoxyl radicals, thiyl radicals, sulfonyl radicals, thiyl peroxyl radicals, and disulfide) and non-radical ROS (hydrogen peroxide, singlet oxygen, ozone/trioxygen, organic hydroperoxides, hypochloride, peroxynitrite, nitrosoperoxycarbonate anion, nitrocarbonate anion, dinitrogen dioxide, nitronium, and highly reactive lipid- or carbohydrate-derived carbonyl compounds). Increases in ROS are involved in various diseases.
2.2.2. MIE2; KE1753: Chronic reactive oxygen species (ROS)
ROS play important role in physiological status and diseases. The balance between the production of ROS and detoxification maintains the physiological condition, which is in prolonged excess ROS production in cancer progress [3]. The essentiality of chronic ROS is high in cancer.
2.2.3. KE1; KE1754: Porcupine-induced Wnt secretion and Wnt signaling activation
Sustained tissue damage induces inflammation. Wnt/beta-catenin signaling is essential for intestinal homeostasis, where macrophage-derived Wnt in intestinal repair is crucial for rescuing intestinal stem cells from radiation lethality [4].
2.2.4. KE2; KE1755: beta-catenin activation
The oncoprotein beta-catenin stabilizes and translocates to nucleus, followed by induction of ZEB1 transcription factor that induces epithelial-mesenchymal transition (EMT) [7]. One of the important signaling pathways inducing EMT is the canonical Wnt/beta-catenin pathway, where beta-catenin acts as coactivator of T-cell and lymphoid enhancer (TCF-LEF) factors [8]. Beta-catenin/TCF4 binds to the ZEB1 promoter and induces the transcription leading to EMT, a main hallmark of malignant cells [7].
2.2.5. KE3; KE1457: Epithelial-mesenchymal transition (EMT)
It is known that EMT plays an important role in therapeutic resistance and drug responses in human gastric cancer [2,9]. EMT is a critical regulator of the CSC phenotype and drug resistance [9]. EMT program is involved in metastasis of gastric cancer [10,11]. Triggering receptor expressed on myeloid cells 2 (TREM2), a key gene in gastric cancer progression, promotes EMT [12].
2.2.6. AO; KE1651: Treatment-resistant gastric cancer
Gastric cancer can be classified into diffuse- and intestinal-type gastric cancer with mRNA ratio of CDH2 to CDH1 [13]. The diffuse-type gastric cancer, which has a poor prognosis and is treatment-resistant, has up-regulated genes that are involved in EMT compared to intestinal-type gastric cancer [14,15]. Gastric cancer-derived mesenchymal stromal cells-primed macrophages promote metastasis and EMT in gastric cancer [16].
3. Discussion
This AOP298 “Increases in cellular ROS and chronic ROS leading to human treatment-resistant gastric cancer” consists of several components: “Increases in cellular ROS” and “Chronic ROS” as MIEs, “porcupine-induced Wnt secretion and Wnt signaling activation”, “beta-catenin activation” and “Epithelial-mesenchymal transition (EMT)” as intermediate KEs, and “Human treatment-resistant gastric cancer” as an AO. The description of the AOP is based on a mechanism of drug resistance, metastasis, and gastric cancer progression, of which application would be the risk assessment of anti-cancer drugs and the development of the anti-cancer treatment as well.
The chronic low-level increased ROS play crucial role in the development of radioresistant GC via tumor microenvironment alteration and EMT [40]. Radiation causes promotion of metastasis of cancer via ROS and EMT [41]. The extent of ROS level seems to be critical in balancing the cancer cell development and cell death [42].
4. Conclusions
The AOP298 “Increases in cellular ROS and chronic ROS leading to human treatment-resistant gastric cancer” illustrates a pathway beginning at chronic increases of ROS inducing Wnt signaling activation and leading to EMT and treatment-resistant GC in human. The description includes a mechanism of drug resistance, metastasis, and GC progression, of which application would be the risk assessment of anti-cancer drugs and the development of the anti-cancer treatment as well.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Document S1: PDF snapshot of AOP298 (298-2024-06-02T23-53-20+00-00).
Author Contributions
Conceptualization, S.T.; investigation, S.T.; resources, S.T.; writing—original draft preparation, S.T.; writing—review and editing, S.T., S.Q., R.O., H.C., K.A., H.Y., H.S., E.P.; visualization, S.T.; project administration, S.T.; funding acquisition, S.T. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by Japan Agency for Medical Research and Development (AMED), Grant Number JP21mk0101216 (S.T.), JP22mk0101216 (S.T.), JP23mk0101216 (S.T.), and Strategic International Collaborative Research Program, Grant Number JP20jm0210059 (S.T. and S.Q.), and JSPS KAKENHI Grant Number 21K12133 (S.T. and R.O.).
Institutional Review Board Statement
Not applicable.
Acknowledgments
The authors would like to acknowledge Dr. Dan Villeneuve, Dr. Jason O’Brien, Dr. Rex FitzGerald, Dr. Magdalini Sachana, Ms. Nathalie Delrue, Dr. Cinzia La Rocca, Dr. Carlie LaLone, Dr. Judy Choi, Kristie Sullivan, Dr. Julija Filipovska, Dr. Tanja Burgdorf, Prof. Dries Knapen, Dr. Vinita Chauhan, and all OECD AOP Coach Team, and the members of National Institute of Health Sciences, Japan.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
The information on AOP298 can be downloaded at: https://aopwiki.org/aops/298. This AOP report article is for summarizing the content of the AOP298 and scientific review as a part of the OECD AOP project: https://www.oecd.org/chemicalsafety/testing/adverse-outcome-pathways-molecular-screening-and-toxicogenomics.htm.
References
- Tanabe, S.; Quader, S.; Ono, R.; Cabral, H.; Aoyagi, K.; Hirose, A.; Yokozaki, H.; Sasaki, H. Molecular Network Profiling in Intestinal- and Diffuse-Type Gastric Cancer. Cancers (Basel) 2020, 12. [Google Scholar] [CrossRef]
- Tanabe, S.; Quader, S.; Cabral, H.; Ono, R. Interplay of EMT and CSC in Cancer and the Potential Therapeutic Strategies. Front Pharmacol 2020, 11, 904. [Google Scholar] [CrossRef]
- Liou, G.Y.; Storz, P. Reactive oxygen species in cancer. Free Radic Res 2010, 44, 479–496. [Google Scholar] [CrossRef]
- Brieger, K.; Schiavone, S.; Miller, F.J., Jr.; Krause, K.H. Reactive oxygen species: from health to disease. Swiss Med Wkly 2012, 142, w13659. [Google Scholar] [CrossRef]
- Babior, B.M. NADPH oxidase: an update. Blood 1999, 93, 1464–1476. [Google Scholar] [CrossRef]
- Tanabe, S.; O’Brien, J.; Tollefsen, K.E.; Kim, Y.; Chauhan, V.; Yauk, C.; Huliganga, E.; Rudel, R.A.; Kay, J.E.; Helm, J.S. , et al. Reactive Oxygen Species in the Adverse Outcome Pathway Framework: Toward Creation of Harmonized Consensus Key Events. Frontiers in Toxicology 2022, 4. [Google Scholar] [CrossRef]
- Sanchez-Tillo, E.; de Barrios, O.; Siles, L.; Cuatrecasas, M.; Castells, A.; Postigo, A. beta-catenin/TCF4 complex induces the epithelial-to-mesenchymal transition (EMT)-activator ZEB1 to regulate tumor invasiveness. Proc Natl Acad Sci U S A 2011, 108, 19204–19209. [Google Scholar] [CrossRef]
- Clevers, H. Wnt/beta-catenin signaling in development and disease. Cell 2006, 127, 469–480. [Google Scholar] [CrossRef]
- Shibue, T.; Weinberg, R.A. EMT, CSCs, and drug resistance: the mechanistic link and clinical implications. Nat Rev Clin Oncol 2017, 14, 611–629. [Google Scholar] [CrossRef]
- Huang, Y.N.; Xu, Y.Y.; Ma, Q.; Li, M.Q.; Guo, J.X.; Wang, X.; Jin, X.; Shang, J.; Jiao, L.X. Dextran Sulfate Effects EMT of Human Gastric Cancer Cells by Reducing HIF-1alpha/ TGF-beta. J Cancer 2021, 12, 3367–3377. [Google Scholar] [CrossRef]
- Sohn, S.H.; Kim, B.; Sul, H.J.; Kim, Y.J.; Kim, H.S.; Kim, H.; Seo, J.B.; Koh, Y.; Zang, D.Y. INC280 inhibits Wnt/beta-catenin and EMT signaling pathways and its induce apoptosis in diffuse gastric cancer positive for c-MET amplification. BMC Res Notes 2019, 12, 125. [Google Scholar] [CrossRef]
- Li, C.; Hou, X.; Yuan, S.; Zhang, Y.; Yuan, W.; Liu, X.; Li, J.; Wang, Y.; Guan, Q.; Zhou, Y. High expression of TREM2 promotes EMT via the PI3K/AKT pathway in gastric cancer: bioinformatics analysis and experimental verification. J Cancer 2021, 12, 3277–3290. [Google Scholar] [CrossRef]
- Tanabe, S.; Aoyagi, K.; Yokozaki, H.; Sasaki, H. Gene expression signatures for identifying diffuse-type gastric cancer associated with epithelial-mesenchymal transition. Int J Oncol 2014, 44, 1955–1970. [Google Scholar] [CrossRef]
- Alberts, S.R.; Cervantes, A.; van de Velde, C.J. Gastric cancer: epidemiology, pathology and treatment. Ann Oncol 2003, 14 Suppl 2, ii31–36. [Google Scholar] [CrossRef]
- Perrot-Applanat, M.; Vacher, S.; Pimpie, C.; Chemlali, W.; Derieux, S.; Pocard, M.; Bieche, I. Differential gene expression in growth factors, epithelial mesenchymal transition and chemotaxis in the diffuse type compared with the intestinal type of gastric cancer. Oncol Lett 2019, 18, 674–686. [Google Scholar] [CrossRef]
- Li, W.; Zhang, X.; Wu, F.; Zhou, Y.; Bao, Z.; Li, H.; Zheng, P.; Zhao, S. Gastric cancer-derived mesenchymal stromal cells trigger M2 macrophage polarization that promotes metastasis and EMT in gastric cancer. Cell Death Dis 2019, 10, 918. [Google Scholar] [CrossRef]
- Vallée, A.; Lecarpentier, Y. Crosstalk Between Peroxisome Proliferator-Activated Receptor Gamma and the Canonical WNT/β-Catenin Pathway in Chronic Inflammation and Oxidative Stress During Carcinogenesis. Front Immunol 2018, 9, 745. [Google Scholar] [CrossRef]
- Ahmad, A.; Sarkar, S.H.; Bitar, B.; Ali, S.; Aboukameel, A.; Sethi, S.; Li, Y.; Bao, B.; Kong, D.; Banerjee, S. , et al. Garcinol regulates EMT and Wnt signaling pathways in vitro and in vivo, leading to anticancer activity against breast cancer cells. Mol Cancer Ther 2012, 11, 2193–2201. [Google Scholar] [CrossRef]
- Clevers, H.; Nusse, R. Wnt/β-catenin signaling and disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef]
- Pearlman, R.L.; Montes de Oca, M.K.; Pal, H.C.; Afaq, F. Potential therapeutic targets of epithelial-mesenchymal transition in melanoma. Cancer Lett 2017, 391, 125–140. [Google Scholar] [CrossRef]
- Yang, W.; Wu, P.F.; Ma, J.X.; Liao, M.J.; Wang, X.H.; Xu, L.S.; Xu, M.H.; Yi, L. Sortilin promotes glioblastoma invasion and mesenchymal transition through GSK-3β/β-catenin/twist pathway. Cell Death Dis 2019, 10, 208. [Google Scholar] [CrossRef]
- Diaz, V.M.; Vinas-Castells, R.; Garcia de Herreros, A. Regulation of the protein stability of EMT transcription factors. Cell Adh Migr 2014, 8, 418–428. [Google Scholar] [CrossRef]
- Tanabe, S. Origin of cells and network information. World J Stem Cells 2015, 7, 535–540. [Google Scholar] [CrossRef]
- Tanabe, S. Signaling involved in stem cell reprogramming and differentiation. World J Stem Cells 2015, 7, 992–998. [Google Scholar] [CrossRef]
- Tanabe, S.; Aoyagi, K.; Yokozaki, H.; Sasaki, H. Regulated genes in mesenchymal stem cells and gastric cancer. World J Stem Cells 2015, 7, 208–222. [Google Scholar] [CrossRef]
- Tanabe, S. Perspectives of gene combinations in phenotype presentation. World J Stem Cells 2013, 5, 61–67. [Google Scholar] [CrossRef]
- Smith, B.N.; Bhowmick, N.A. Role of EMT in Metastasis and Therapy Resistance. J Clin Med 2016, 5. [Google Scholar] [CrossRef]
- Gao, Q.; Zhou, G.; Lin, S.J.; Paus, R.; Yue, Z. How chemotherapy and radiotherapy damage the tissue: Comparative biology lessons from feather and hair models. Exp Dermatol 2019, 28, 413–418. [Google Scholar] [CrossRef]
- Perez, S.; Talens-Visconti, R.; Rius-Perez, S.; Finamor, I.; Sastre, J. Redox signaling in the gastrointestinal tract. Free Radic Biol Med 2017, 104, 75–103. [Google Scholar] [CrossRef]
- Jiang, X.; Charlat, O.; Zamponi, R.; Yang, Y.; Cong, F. Dishevelled promotes Wnt receptor degradation through recruitment of ZNRF3/RNF43 E3 ubiquitin ligases. Mol Cell 2015, 58, 522–533. [Google Scholar] [CrossRef]
- Janda, C.Y.; Waghray, D.; Levin, A.M.; Thomas, C.; Garcia, K.C. Structural basis of Wnt recognition by Frizzled. Science 2012, 337, 59–64. [Google Scholar] [CrossRef]
- Nile, A.H.; Mukund, S.; Stanger, K.; Wang, W.; Hannoush, R.N. Unsaturated fatty acyl recognition by Frizzled receptors mediates dimerization upon Wnt ligand binding. Proc Natl Acad Sci U S A 2017, 114, 4147–4152. [Google Scholar] [CrossRef]
- Wawruszak, A.; Kalafut, J.; Okon, E.; Czapinski, J.; Halasa, M.; Przybyszewska, A.; Miziak, P.; Okla, K.; Rivero-Muller, A.; Stepulak, A. Histone Deacetylase Inhibitors and Phenotypical Transformation of Cancer Cells. Cancers (Basel) 2019, 11. [Google Scholar] [CrossRef]
- Peinado, H.; Olmeda, D.; Cano, A. Snail, Zeb and bHLH factors in tumour progression: an alliance against the epithelial phenotype? Nat Rev Cancer 2007, 7, 415–428. [Google Scholar] [CrossRef]
- Batlle, E.; Clevers, H. Cancer stem cells revisited. Nat Med 2017, 23, 1124–1134. [Google Scholar] [CrossRef]
- Saxena, M.; Stephens, M.A.; Pathak, H.; Rangarajan, A. Transcription factors that mediate epithelial-mesenchymal transition lead to multidrug resistance by upregulating ABC transporters. Cell Death Dis 2011, 2, e179. [Google Scholar] [CrossRef]
- Pirozzi, G.; Tirino, V.; Camerlingo, R.; Franco, R.; La Rocca, A.; Liguori, E.; Martucci, N.; Paino, F.; Normanno, N.; Rocco, G. Epithelial to mesenchymal transition by TGFbeta-1 induction increases stemness characteristics in primary non small cell lung cancer cell line. PLoS One 2011, 6, e21548. [Google Scholar] [CrossRef]
- Kudo-Saito, C.; Shirako, H.; Takeuchi, T.; Kawakami, Y. Cancer metastasis is accelerated through immunosuppression during Snail-induced EMT of cancer cells. Cancer Cell 2009, 15, 195–206. [Google Scholar] [CrossRef]
- Chen, L.; Gibbons, D.L.; Goswami, S.; Cortez, M.A.; Ahn, Y.H.; Byers, L.A.; Zhang, X.; Yi, X.; Dwyer, D.; Lin, W. , et al. Metastasis is regulated via microRNA-200/ZEB1 axis control of tumour cell PD-L1 expression and intratumoral immunosuppression. Nat Commun 2014, 5, 5241. [Google Scholar] [CrossRef]
- Gu, H.; Huang, T.; Shen, Y.; Liu, Y.; Zhou, F.; Jin, Y.; Sattar, H.; Wei, Y. Reactive Oxygen Species-Mediated Tumor Microenvironment Transformation: The Mechanism of Radioresistant Gastric Cancer. Oxid Med Cell Longev 2018, 2018, 5801209. [Google Scholar] [CrossRef]
- Lee, S.Y.; Jeong, E.K.; Ju, M.K.; Jeon, H.M.; Kim, M.Y.; Kim, C.H.; Park, H.G.; Han, S.I.; Kang, H.S. Induction of metastasis, cancer stem cell phenotype, and oncogenic metabolism in cancer cells by ionizing radiation. Mol Cancer 2017, 16, 10. [Google Scholar] [CrossRef]
- Jiang, H.; Zuo, J.; Li, B.; Chen, R.; Luo, K.; Xiang, X.; Lu, S.; Huang, C.; Liu, L.; Tang, J. , et al. Drug-induced oxidative stress in cancer treatments: Angel or devil? Redox Biol 2023, 63, 102754. [Google Scholar] [CrossRef]
Figure 1.
Canonical pathway of production of nitric oxide and reactive oxygen species in macrophages, and its molecular relationship to epithelial-mesenchymal transition (analyzed in Ingenuity Pathway Analysis (IPA)). Analysis of 12- stomach cancer 14264, as of June 2023.
Figure 1.
Canonical pathway of production of nitric oxide and reactive oxygen species in macrophages, and its molecular relationship to epithelial-mesenchymal transition (analyzed in Ingenuity Pathway Analysis (IPA)). Analysis of 12- stomach cancer 14264, as of June 2023.
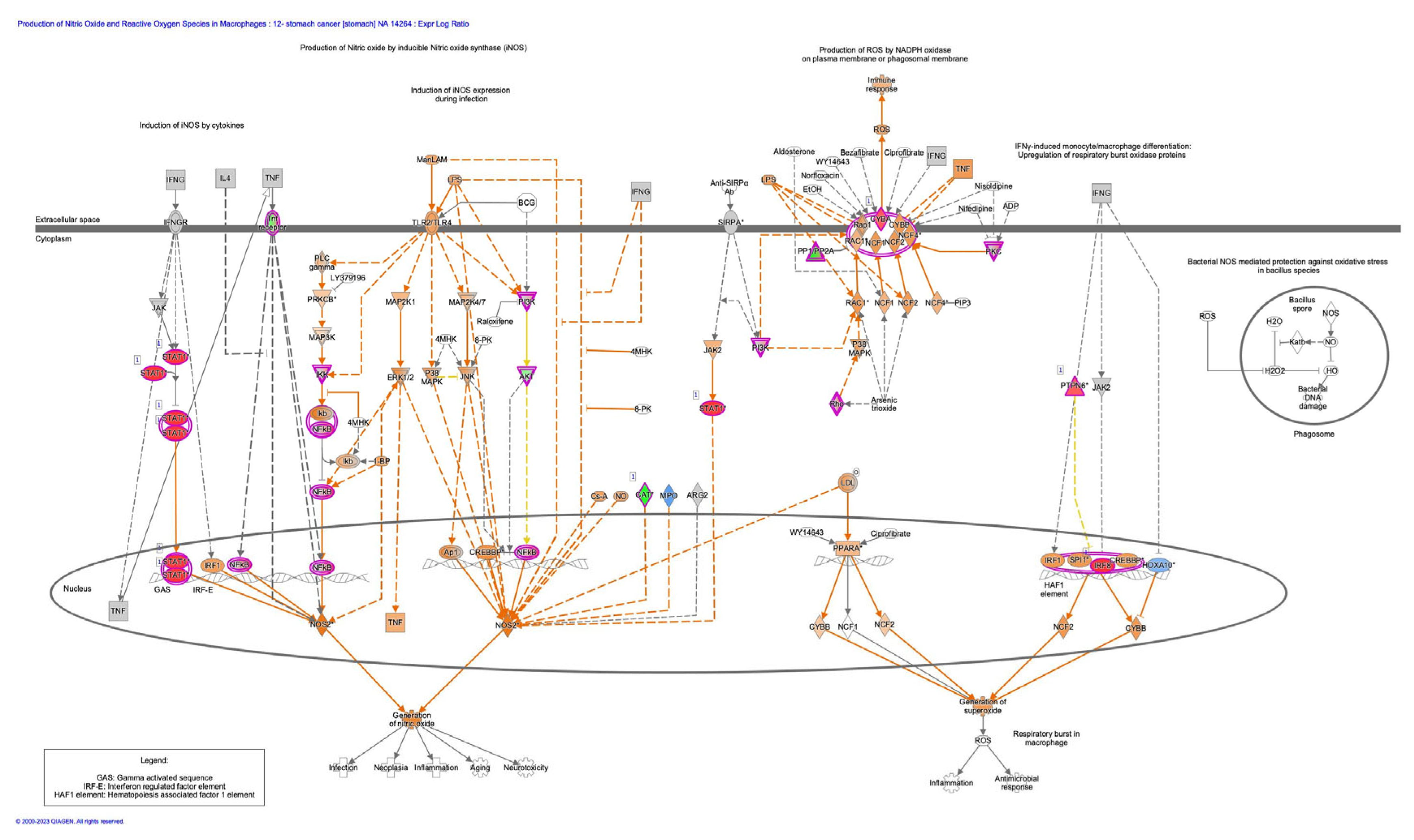
Figure 2.
AOP 298 “Increases in cellular ROS and chronic ROS leading to human treatment-resistant gastric cancer”.
Figure 2.
AOP 298 “Increases in cellular ROS and chronic ROS leading to human treatment-resistant gastric cancer”.
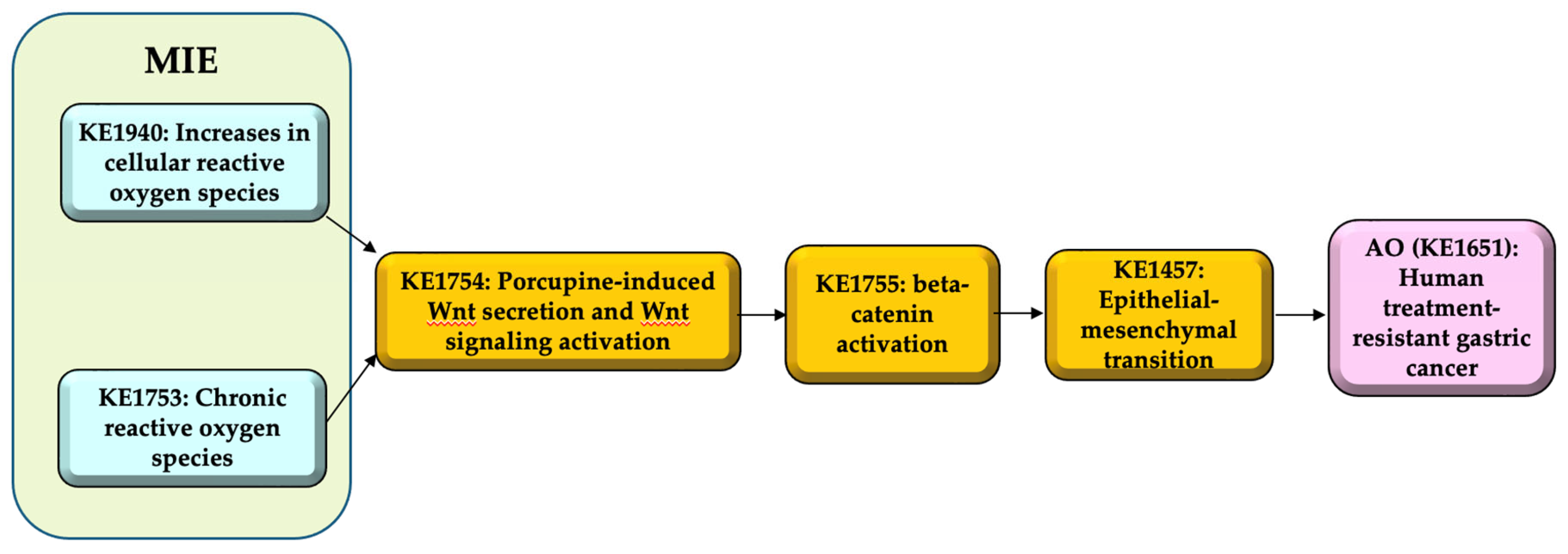

Table 1.
AOP298-related information.
| Item | Title |
|---|---|
| AOP | Increases in cellular reactive oxygen species (ROS) and chronic ROS leading to human treatment-resistant gastric cancer (GC) |
| MIE1 | KE1940: Increases in cellular reactive oxygen species (ROS) |
| MIE2 | KE1753: Chronic reactive oxygen species (ROS) |
| KE1 | KE1754: Porcupine-induced Wnt secretion and Wnt signaling activation |
| KE2 | KE1755: beta-catenin activation |
| KE3 | KE1457: Epithelial-mesenchymal transition |
| AO | KE1651: Treatment-resistant gastric cancer |
Table 2.
Support for Biological Plausibility of KERs in AOP298.
| Item | Evidence |
|---|---|
| MIE1 => KE1: Increases in cellular ROS leads to Porcupine-induced Wnt secretion and Wnt signaling activation |
Biological Plausibility of the MIE1 => KE1 is moderate. Rationale: Increases in cellular ROS caused by/causes DNA damage, which will alter several signaling pathways including Wnt signaling. ROS stimulate inflammatory factor production and Wnt/beta-catenin signaling [17]. |
| MIE2 => KE1: Chronic ROS leads to Porcupine-induced Wnt secretion and Wnt signaling activation |
Biological Plausibility of the MIE2 => KE1 is moderate. Rationale: Sustained ROS increase caused by/causes DNA damage altering several signaling pathways including Wnt signaling. Macrophages accumulate into the injured tissue to recover the tissue damage, which may be followed by porcupine-induced Wnt secretion. ROS stimulate inflammatory factor production and Wnt/ beta-catenin signaling [17]. |
| KE1 => KE2: Porcupine-induced Wnt secretion and Wnt signaling activation leads to beta-catenin activation |
Biological Plausibility of the KE1 => KE2 is moderate. Rationale: Secreted Wnt ligand stimulates Wnt/beta-catenin signaling, in which beta-catenin is activated. Wnt ligand binds to Frizzled receptor, which leads to GSK3beta inactivation. GSK3beta inactivation leads to beta-catenin dephosphorylation, which avoids the ubiquitination of the beta-catenin and stabilize the beta-catenin [8] |
| KE2 => KE3: beta-catenin activation leads to Epithelial-mesenchymal transition (EMT) |
Biological Plausibility of the KE2 => KE3 is moderate. Rationale: Beta-catenin activation, which includes stabilizing the dephosphorylated beta-catenin and translocation of beta-catenin into the nucleus, induces the formation of beta-catenin-TCF complex and transcription of transcription factors, such as Snail, Zeb and Twist [11,18,19,20,21]. EMT-related transcription factors including Snail, ZEB, and Twist, are up-regulated in cancer cells [22]. The transcription factors such as Snail, ZEB, and Twist bind to E-cadherin (CDH1) promoter and inhibit the CDH1 transcription via the consensus E-boxes (5’-CACCTG-3’ or 5’-CAGGTG-3’), which leads to EMT [22]. |
| KE3 => AO: Epithelial-mesenchymal transition (EMT) leads to human treatment-resistant gastric cancer |
Biological Plausibility of the KE3 => AO is moderate. Rationale: Some population of the cells exhibiting EMT demonstrates the feature of cancer stem cells (CSCs), which are related to cancer malignancy [9,23,24,25]. EMT phenomenon is related to cancer metastasis and cancer therapy resistance [26,27]. The increase in expression of enzymes that degrade the extracellular matrix components and the decrease in adhesion to the basement membrane in EMT induce the cell to escape from the basement membrane and metastasis [27]. Morphological changes observed during EMT are associated with therapy resistance [27]. |
Table 3.
Support for Essentiality of KEs in AOP298.
| Item | Evidence |
| KE1: Porcupine-induced Wnt secretion and Wnt signaling activation | Essentiality of the KE1 is moderate. Rationale for Essentiality of KEs in the AOP: The Wnt signaling activation is essential for the subsequent beta-catenin activation and cancer resistance. |
| KE2: beta-catenin activation | Essentiality of the KE2 is moderate. Rationale for Essentiality of KEs in the AOP: beta-catenin activation is essential for the Wnt-induced cancer resistance. |
| KE3: Epithelial-mesenchymal transition (EMT) | Essentiality of the KE3 is moderate. Rationale for Essentiality of KEs in the AOP: EMT is essential for the Wnt-induced cancer promotion and acquisition of resistance to anti-cancer drug. |
Table 4.
Empirical support for KERs in AOP298.
| Item | Evidence |
| MIE1 => KE1: Increases in cellular ROS leads to Porcupine-induced Wnt secretion and Wnt signaling activation |
Empirical Support of the MIE1 => KE1 is moderate. Rationale: Production of ROS and DNA double-strand break causes the tissue damages [28]. ROS-related signaling induces Wnt/beta-catenin pathway activation [29]. |
| MIE2 => KE1: Chronic ROS leads to Porcupine-induced Wnt secretion and Wnt signaling activation |
Empirical Support of the MIE2 => KE1 is moderate. Rationale: Production of ROS and DNA double-strand break causes the tissue damages [28]. ROS-related signaling induces Wnt/beta-catenin pathway activation [29]. |
| KE1 => KE2: Porcupine-induced Wnt secretion and Wnt signaling activation leads to beta-catenin activation |
Empirical Support of the KE1 => KE2 is moderate. Rationale: Dishevelled (DVL), a positive regulator of Wnt signaling, form the complex with FZD and lead to trigger the Wnt signaling together with Wnt coreceptor low-density lipoprotein (LDL) receptor-related protein 6 (LRP6) [19,30]. Wnt binds to FZD and activate the Wnt signaling [19,31,32]. Wnt binding towards FZD induce the formation of the protein complex with LRP5/6 and DVL, leading to the down-stream signaling activation including beta-catenin [8]. |
| KE2 => KE3: beta-catenin activation leads to Epithelial-mesenchymal transition (EMT) |
Empirical Support of the KE2 => KE3 is moderate. Rationale: The inhibition of c-MET, which is overexpressed in diffuse-type gastric cancer, induced increase in phosphorylated beta-catenin, decrease in beta-catenin and Snail [11]. The garcinol, which has an anti-cancer effect, increases phosphorylated beta-catenin, decreases beta-catenin and ZEB1/ZEB2, and inhibits EMT [18]. The inhibition of sortilin by AF38469 (a sortilin inhibitor) or small interference RNA (siRNA) results in a decrease in beta-catenin and Twist expression in human glioblastoma cells [21]. Histone deacetylase inhibitors affect EMT-related transcription factors including, ZEB, Twist, and Snail [33]. Snail and Zeb induces EMT and suppress E-cadherin (CDH1) [22,34,35]. |
| KE3 => AO: Epithelial-mesenchymal transition (EMT) leads to human treatment-resistant gastric cancer |
Empirical Support of the KE3 => AO is moderate. Rationale: EMT activation induces the expression of multiple members of the ATP-binding cassette (ABC) transporter family, which results in the resistance to doxorubicin [9,36]. TGFbeta-1 induced EMT results in the acquisition of cancer stem cell (CSC) like properties [9,37]. Snail-induced EMT induces cancer metastasis and resistance to dendritic cell-mediated immunotherapy [38]. Zinc finger E-box-binding homeobox (ZEB1)-induced EMT results in the relief of miR-200-mediated repression of programmed cell death 1 ligand (PD-L1) expression, a major inhibitory ligand for the programmed cell death protein (PD-1) immune-checkpoint protein on CD8+ cytotoxic T lymphocyte (CTL), subsequently the CD8+ T cell immunosuppression and metastasis [39]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



