
Submitted:
15 June 2024
Posted:
17 June 2024
You are already at the latest version
Abstract
The ethylene-regulated hypocotyl elongation of Arabidopsis thaliana's involves many transcription factors. The specific role of MYC transcription factors in ethylene signal transduction is not completely understood. The results here revealed that two MYCs, MYC2 and MYC3, act as negative regulators in ethylene-suppressed hypocotyl elongation. Etiolated seedlings of the lost-of-function mutant of MYC2 or MYC3 were longer than wild type seedlings. Single- or double-null mutants of MYC2 and MYC3 displayed an enhanced response to ACC(1-aminocyclopropane-1-carboxylate), the ethylene precursor, compared to wild type seedlings. MYC2 and MYC3 directly bind to the promoter zone of ERF1, suppressing its expression. Additionally, EIN3, a key component in ethylene signaling, interacts with MYC2 or MYC3 and suppresses their binding to ERF1’s promoter. MYC2 and MYC3 play crucial roles in the ethylene-regulated expression of functional genes. The results revealed these transcription factors’ novel role and functional mechanism in ethylene signal transduction.
Keywords:
Arabidopsis thaliana
; MYC2
; MYC3
; ethylene
; hypocotyl
; ERF1
1. Introduction
During seed germination, the elongation of the hypocotyl pushes the primary root out of the seed coat and allows it to absorb water and minerals from the soil. Hypocotyl elongation also helps in uncovering the cotyledons, which is essential for photomorphogenesis and the development of autotrophic seedlings [1,2]. The hypocotyl is sensitive to various internal and external signals [3], making it an ideal model for studying the regulation of plant growth and development [4,5].
When elongating hypocotyls encounter obstacles, ethylene levels increase to suppress the elongation rate and boost the jacking force, which is necessary for pushing seedlings out of the soil [6]. In ethylene-regulated growth and development, five ethylene receptors in the endoplasmic reticulum (ER) membrane, including ETR1, ERS1, ETR2, ERS2, and EIN4, are involved in responding to ethylene and triggering downstream signaling events [7,8,9]. When ethylene is absent, these receptors bind to CTR1 to form a complex. As a serine/threonine protein kinase, CTR1 phosphorylates EIN2. Since phosphorylated EIN2 is unable to trigger EIN3/EIL1, which act as transcription factors, the signaling cascade is blocked. When ethylene content increases, it binds and disassembles the receptor complex, releasing the inhibitory effect of CTR1 on EIN2. Activated EIN2 migrates from the ER membrane to the nucleus and stimulates EIN3/EIL1, which ultimately alters functional gene expression related to growth and development [9,10,11].
The growth of Arabidopsis hypocotyl is influenced by ethylene in relation to light exposure. Ethylene suppresses elongation of etiolated hypocotyls in the dark but promotes elongation in the light [3,12]. In darkness, hypocotyls of the ethylene-overproducing mutant eto1-1 and the null mutant of CTR1 (ctr1-1) are shorter than the wild type, whereas hypocotyls of the ethylene-insensitive mutant ein2-5 and ein3eil1 are longer than the wild type [13,14]. In the light, hypocotyls of the EIN3-overexpression line and ctr1-1 are longer than the wild type, whereas hypocotyls of ein2-5 and ein3eil1 are shorter than the wild type [12]. The expression of Ethylene Response Factor 1 (ERF1) is regulated by ethylene, and overexpression of ERF1 significantly inhibits elongation of etiolated hypocotyls, thereby positively regulating ethylene-inhibited elongation [15,16].
MYC transcription factors belong to the plant bHLH transcription factor family. The first plant MYC gene was discovered in maize, and it is involved in regulating anthocyanin biosynthesis [17]. In Arabidopsis, AtMYC2 and its three homologues, AtMYC3, AtMYC4, and AtMYC5, have been extensively studied [18,19]. MYCs are involved in four physiological processes: (1) Plant growth and development. MYC2, MYC3, MYC4, and MYC5 inhibit cell proliferation of integument, thus negatively regulating seed enlargement [20]. MYC3 interacts with the DELLA protein, antagonizes the regulation of CO (CONSTANS) on FT (FLOWERING LOCUS T) gene expression, and participates in the flowering process which is regulated by photoperiod and gibberellin(GA) [21]. MYC2, MYC3, and MYC4 negatively regulate Jasmonic Acid(JA)-inhibited root and hypocotyl growth, by regulating the expression of HY5 [22,23]. MYC2 promotes the expression of a senescence-related gene, Dof2.1, thus enhancing JA-induced leaf senescence [24]. (2) Plants’ response to biotic stresses. In Solanum lycopersicum, magnesium oxide triggers the expression of resistance genes by stimulating JA signaling and activating MYC2, thus inducing the immune response to Fusarium wilt [25]. In rice, overexpression of OsMYC2 induces up-regulation of pathogen resistance genes and enhances resistance to bacterial blight [26]. In Arabidopsis, AtPIFs and AtMYC2 form a homodimer that binds the promoter of the TPS (terpene synthase) gene. An increase in terpene biosynthesis enhances the defensive ability of whitefly (Bemisia tabaci) [27]. (3) Plants’ response to abiotic stresses. MYC1 inhibits the binding of FIT (FER-like Iron Deficiency-induced transcription factor) with bHLH38/39 heterodimer, suppressing the expression of its target genes (FRO2 and IRT1), thus inhibiting iron absorption and tolerance to iron deficiency [28]. MYC2 enhances Arabidopsis' tolerance to salinity by regulating proline biosynthesis [29]. The MYC2/MYC3/MYC4-dependent transcription factor network regulates water spray-responsive gene expression, as well as JA accumulation, responding to water spray stress caused by simulated rain [30]. (4) Regulating the synthesis of secondary metabolites. Some MYCs regulate gene expression of components in the biosynthesis of tryptophan, alkaloids, or flavonoids, promoting the accumulation of these secondary metabolites [26,31,32]. The accumulation of flavonoids inhibits the growth of insects and enhances the recovery ability of plant cells, increasing resistance to insect attacks [33].
The growth and development of plants are regulated by phytohormones, which also induce stress response by stimulating or suppressing the expression of functional genes using multiple transcription factors. In ethylene signaling, several agents can bind and regulate the activity of some bHLH transcription factor members. Phytochrome-interacting factors (PIFs), a group of well-characterized bHLH transcription factors, are involved in the cross-talk of ethylene- and photo-signaling [34]. Overexpression of PIF5 increases ethylene levels in etiolated seedlings by stimulating the expression of ACC synthase [35,36]. Ethylene promotes hypocotyl elongation of green seedlings by increasing PIF3 expression [12]. PIF1 and EIN3/EIL1 inhibit photobleaching by inhibiting the expression of protochlorophyllide biosynthetic genes and activating the expression of POR genes [37]. EIN3 and EIL1 participate in skotomorphogenesis by regulating the activity of PIF3 [16]. PIFs and EIN3/EIL1 are involved in apical hooks development, which is being regulated by phytohormones (e.g., ethylene, gibberellin, JA, salicylic acid, cytokinin) and light, by regulating the expression of HLS140 [38,39,40]. MYCs are involved in antagonism of JA- and ethylene-signaling in Arabidopsis. MYC2/3/4 repress the transcriptional activity of EIN3/EIL1, thus suppressing ethylene-promoted resistance to necrotic fungi and apical hook formation [41], vice versa, EIN3/EIL1 inhibits the transcriptional activity of MYC2/3/4 to inhibit the expression of wound response genes (TAT3, VSP1, and VSP2) and herbivore-induced genes (CYP79B3, BCAT4, and BAT5) induced by JA [42].
Several MYCs have been studied for their physiological functions and functional mechanisms, but the specific role of MYC2 and MYC3 in ethylene signaling, is not fully understood. This study confirms the involvement of MYC2 and MYC3 in regulating hypocotyl growth in response to ethylene and explores their relationship with various ethylene signaling agents. The findings offer new insights into the role of MYC2 and MYC3 in the ethylene signal transduction pathway and shed light on the mechanism behind ethylene-regulated seedling growth.
2. Results
2.1. MYC2 and MYC3 Negatively Regulate the Ethylene-Inhibited Elongation of Etiolated Hypocotyls
To analyze the role of MYC3 and MYC3 in ethylene-inhibited etiolated hypocotyl elongation, the response of seedlings, including Col-0 and two MYC2 single null mutants (myc2-1 and myc2-2) and a MYC3 null mutant (myc3), to ethylene precursor ACC was detected. The hypocotyl length of myc2-1, myc2-2, and myc3 seedlings were 13.86±0.74, 14.00±1.07, and 13.84±1.09 mm, respectively, significantly longer than hypocotyl of Col-0 seedlings (11.83±0.74 mm) (p<0.05) (Figure 1A), indicating that MYC2 and MYC3 may play negative roles in elongation of etiolated hypocotyls. The etiolated hypocotyl of the double null mutant (myc2myc3) seedlings was longer than myc2 and myc3 seedlings (p<0.01), indicating that MYC2 and MYC3 may regulate hypocotyl elongation synergistically (Figure 1A,B).
On a medium containing ACC, the growth rate of etiolated hypocotyls significantly decreased. The effect of 25 μΜ ACC was stronger than that of 10 μΜ ACC, and the effect of 50 μΜ ACC was similar to that of 25 μΜ ACC. The length of etiolated hypocotyls of myc2-1, myc2-2, myc3, and myc2myc3 seedlings was similar to that of Col-0 seedlings after treatment with 10, 25, or 50 μΜ ACC (p>0.05) (Figure 1A–C). To clarify the role of MYC2 or MYC3 in ethylene-inhibited etiolated hypocotyl elongation, the ratio of etiolated hypocotyl length under ACC to control was calculated. Results showed that the ratio of any null mutant was higher than that of Col-0, indicating that mutants respond to ACC more sensitively than the wild type (Figure 1 and Figure S1).
To elucidate the role of MYCs in etiolated seedlings’ responses to ethylene, growth parameters of seedlings that were cultured on ACC-containing medium were detected. 0.1 μΜ ACC significantly inhibited elongation of etiolated hypocotyls of myc2-2, myc3, and myc2myc3, while did not affect etiolated seedlings of myc2-1 and Col-0. On 0.25 μΜ ACC containing medium, all myc mutants’ seedlings were notably suppressed, while Col-0 seedlings were only slightly suppressed. Furthermore, on a medium containing 0.5 μΜ ACC, the elongation of hypocotyls in myc2myc3 displayed increased sensitivity compared to Col-0 and the three single null mutants. Col-0 and all mutants’ seedlings responded to 0.75 or 1 μΜ ACC in a similar manner and to a similar extent. The results indicated that the effectiveness of MYC2 and MYC3 in etiolated hypocotyls’ response to ethylene depends on ethylene dosage (Figure S1).
To further verify the role of MYC2 and MYC3 in ethylene-regulated seedling growth, the response of etiolated seedlings of MYC2 or MYC3 complemention lines to ACC was detected. Results showed that seedlings of two MYC2 complemention lines (MYC2#COM1, MYC2#COM2) and two MYC3 complemention lines (MYC3#COM1, MYC3#COM2) responded to ACC in similar sensitivity with Col-0 seedlings (Figure S2).
It was reported that while ethylene suppresses the elongation of etiolated hypocotyls, it promotes the elongation of hypocotyls in green seedlings [3,12]. To further investigate the role of MYC2 and MYC3 in ethylene-regulated seedling growth, seedlings were cultured under light on ACC-containing medium. The results showed that the seedling growth rate was similar in Col-0 and MYC2 or MYC3 null mutants. In other words, the hypocotyls of Col-0, myc2-1, myc2-2, myc3, and myc2myc3 seedlings were all significantly promoted by ACC (Figure S3), suggesting that MYC2 and MYC3 may not be involved in the ethylene-promoted elongation of hypocotyls in green seedlings.
To further investigate the role of MYC2 and MYC3 in ethylene-suppressed etiolated hypocotyl elongation, scanning electron microscopy (SEM) was used to detect etiolated hypocotyl cells. The epidermal cell count showed that the number of hypocotyl cells in both Col-0 and all mutant seedlings is similar, with each row containing 20-22 epidermal cells. Etiolated seedlings grown on ACC-free medium showed that the hypocotyl cells' length of myc2-1, myc2-2, and myc3 were significantly longer than Col-0, but shorter than myc2myc3. On ACC-containing medium, the hypocotyl cells' length decreased significantly, and the cell length of myc2-1, myc2-2, and myc3 was not significantly different from Col-0 (Figure 2). This suggests that mutants were more sensitive to ACC than the wild type, indicating that MYC2 and MYC3 may play a negative role in ethylene-regulated cell elongation in etiolated hypocotyls.
2.2. MYC2 and MYC3 Inhibit Expression of ERF1 by Binding Its Promoter
To verify the possible mechanism of MYC2 and MYC3-regulated hypocotyl growth, expression of ERF1 in myc2 or myc3 was detected. In 4-days-old etiolated seedlings, expression of ERF1 in myc2, myc3 was markedly higher than in Col-0 (p<0.01), the expression level in myc2-2 was the highest among the three mutants, which was 7-8 times of Col-0 (Figure 3A). The results indicate that ERF1 expression is negatively regulated by MYC2 or MYC3.
MYC2 and MYC3 can bind G-box, a cis-acting element in the promoter of target genes, to regulate gene expression [23]. In the promoter of ERF1, there are 4 potential MYC2-binding sites, located at -1233, -1279, -1830, and -1921 upstream of the transcription start point; and 2 potential MYC3-binding sites, located at -1789 and -1921 upstream of transcription start point, respectively (Figure 3B). MYC2 or MYC3 may binds these elements and regulates ERF1 expression.
To verify the binding of MYC2 or MYC3 with the ERF1 promoter, GST-MYC2 and GST-MYC3 proteins were purified. Probe primers were designed based on the potential binding sequence in the ERF1 promoter, and EMSA (electrophoretic mobility shift assay) was performed. The results showed that GST-MYC2 or GST-MYC3 bound to P1 (Figure 3C,D), while they did not bind to P2, P3, P4, or P5 of the ERF1 promoter (Figure S4). The binding was abolished by adding an unlabeled P1 probe, but it was not affected by mutated probes.
To confirm the binding of MYC2 or MYC3 with the ERF1 promoter, MYC2 or MYC3 was cloned to pSuper::pCAMBIA1300GFP binary vector and transformed into Col-0, DNA was extracted and purified from transformant seedlings for chromatin immunoprecipitation assay, which was performed by using GFP (green fluorescent protein) antibody. Results showed that ERF1 promoter sequences in genomic DNA fragments were co-immunoprecipitated by MYC2 or MYC3 (Figure 3E,F), proving the in vivo binding of MYC2 or MYC3 with ERF1 promoter.
To evaluate the effect of MYC2 or MYC3 on the expression of ERF1, the ERF1 promoter-drived GUS(β-glucuronidase) reporter was transiently expressed. pSuper::MYC2 and pSuper::MYC3 were used as effectors. GUS vectors driven by ERF1 promoter, including G-box or mutated G-box, were used as reporters (Figure 4A). Genes were transiently expressed in Nicotiana benthamiana leaves. Results showed that MYC2 or MYC3 inhibited ERF1 expression through the G-box region in the ERF1 promoter, since the inhibitory effect was not detected when G-box was mutated (Figure 4B,C).
2.3. EIN3 Suppresses the Binding of MYC2 or MYC3 with ERF1’s Promoter
Interaction between some MYCs (e.g. MYC2, MYC3, MYC4) and EIN3 has been reported [41,42]. Yeast two hybridization result primarily showed that MYC2 and MYC3 did not interact with EIN2 (Figure S5). To investigate whether EIN3 can affect the binding and inhibitory effect of MYC2 or MYC3 on ERF1 expression, fusion genes including EIN3-MYC, MYC2-GFP, MYC3-GFP, ERF1pro::GUS, and ERF1mpro::GUS were separately combined and co-expressed in N. benthamiana leaves. Results showed that the inhibitory effect of MYC2 or MYC3 on ERF1 expression was further enhanced by EIN3 (Figure 4B,C).
It was reported that EIN3 can bind ERF1’s promoter [15]. To further explore the impact of EIN3 on the association between MYC2, MYC3, and ERF1’s promoter, the EIN3 binding site, MYC2 and MYC3 binding site (P1) in ERF1’s promoter were mutated for following EMSA. Results showed that when the EIN3 binding site was mutated, the expression of ERF1 decreased, when the EIN3 binding site and P1 were both mutated, the expression of ERF1 was not altered (Figure 4B,C), indicating that EIN3 inhibits the transcriptional activity of MYC2 or MYC3. EMSA results showed that, EIN3 inhibited the binding of MYC2 or MYC3 with ERF1’s promoter, along with the increasing of EIN3 dosage, the binding of MYC2 with P1 was gradually weakened (Figure 4D,E).
2.4. MYC2 and MYC3 Genetically Act Upstream of ERF1
To verify the role of ERF1 in ethylene-suppressed etiolated hypocotyl elongation, seedings’ response of 4 ERF1 null mutants (erf1-1 and erf1-2 were generated using CRISPR/Cas9, as showed in Figure 5A; erf1-3 and erf1-4 were generated using RNAi) and two ERF1 overexpression lines (ERF1-OE1 and ERF1-OE2) to ACC was detected. On ACC-free medium, hypocotyl length of the ERF1 null mutants was similar to wild type, while hypocotyls of the ERF1 OE lines were significantly shorter than Col-0. On ACC-containing medium, the hypocotyl length of the null mutants’ seedlings was similar to Col-0, while hypocotyls of the ERF1 OE lines were significantly shorter than Col-0 (Figure 5B,C). The ratio of hypocotyl length for seedlings grown on ACC/ACC-free medium showed that the ratio of ERF1 OE lines were significantly higher than Col-0, indicating overexpression of ERF1 increased seedlings’ sensitivity to ethylene (Figure 5D).
To further investigate the genetic link of MYC2, MYC3, and ERF1, erf1-3 and ERF1-OE1 were hybridized with null mutants of MYC2 or MYC3, materials with different genetic backgrounds were identified from their offsprings. The response of etiolated seedlings to ACC showed that the sensitivity of three double null mutants (myc2-1erf1-3, myc2-2erf1-3 and myc3erf1-3) was similar to erf1-3. However, seedlings of myc2-1ERF1-OE1, myc2-2ERF1-OE1, and myc3ERF1-OE1 showed sensitivity to ACC similar to ERF1-OE1 (Figure 6). These results further demonstrate that MYC2 and MYC3 act upstream of ERF1.
2.5. MYC2 and MYC3 Are Involved in Ethylene-Regulated Gene Expression
To better understand the role of MYC2 and MYC3 in ethylene-regulated hypocotyl growth, high-throughput RNA-sequencing was used to analyze changes in gene expression pattern after ACC treatment in both Col-0 and myc2myc3. Following 25 μM ACC treatment, 1651 differentially expressed genes (DEGs) (fold change > 2) were identified in Col-0 seedlings, with 853 genes up-regulated and 798 genes down-regulated. In myc2myc3 seedlings, 1559 DEGs were identified, with 636 genes up-regulated and 923 genes down-regulated (Figure 7A). Of these genes, 724 were differentially expressed both in Col-0 and myc2myc3 seedlings (Figure 7B). Gene ontology (GO) analysis revealed that in Col-0 seedlings, DEGs were enriched in response to stress, plant hormone signaling, cell wall organization, and cellular metabolism; while in myc2myc3 seedlings, DEGs were enriched in response to ethylene, water or salt stress, plant hormone signaling, and secondary metabolism (Figures 7C and S6).
To further verify the role of MYC2 and MYC3 in ethylene-regulated gene expression, DEGs with a fold change >5 in both Col-0 and myc2myc3 were analyzed in detail. DEGs that were up-regulated in Col-0 while not up-regulated in myc2myc3 are involved in cell wall organization, oxidation & reduction, stress response, and gene expression regulation, etc. (Table 1) This suggests that MYC2 and MYC3 may have a positive role in the ethylene-regulated expression of these genes. DEGs that were down-regulated in Col-0 while not down-regulated in myc2myc3 are involved in cellular metabolism, growth & development, signal transduction, oxidation & reduction, and gene expression regulation, etc. (Table 2) This suggests that MYC2 and MYC3 may play a negative role in the ethylene-regulated expression of these genes. Further analysis revealed that DEGs that were up-regulated in myc2myc3 while not up-regulated in Col-0 are involved in cell wall organization, cellular metabolism, signal transduction, and gene expression regulation, etc. (Table S1) DEGs that were down-regulated in myc2myc3 while not down-regulated in Col-0 are involved in cellular metabolism, hormone signaling, oxidation & reduction, stress response, and detoxification, etc. (Table S2)
3. Discussion
MYCs are important transcription factors involved in JA, ABA, and GA signaling, as well as photoreceptor-mediated light signaling [43,44,45]. In 2020, Yi et al. reported that light induces JA synthesis, and the binding of JA to its receptor COI1 activates MYC2, MYC3, and MYC4. These transcription factors bind to the promoter of HY5, leading to suppressed hypocotyl elongation [23]. The role of MYCs in skotomorphogenesis has been rarely reported. We investigated the growth of etiolated seedlings of MYCs null mutants (including myc2, myc3, myc4, myc5) and found that the etiolated hypocotyls of myc2 and myc3 were significantly longer than those of wild type, indicating their novel role in regulating etiolated hypocotyl elongation.
As a typical ethylene-induced response, the "triple response" had been used to verify the involvement of ethylene in physiological processes. As a precursor of ethylene synthesis, ACC has been used in studies about the physiological function and mechanism of ethylene [11,46,47]. In this work, etiolated hypocotyls of myc2 or myc3 seedlings showed increased sensitivity to ACC, illustrating that MYC2 and MYC3 play negative roles in ethylene-inhibited etiolated hypocotyl elongation. In un-stimulated seedlings, endogenous growth substrates, including ethylene, finely modulate seedling growth, ensuring the steady growth of seedlings in darkness [3,4,48]. MYC2 and MYC3 may be downstream targets of ethylene signaling, acting as growth suppressors, and playing synergistic roles.
The signal transduction process of ethylene-induced triple response has been illustrated. Ethylene binds its receptors, releases their inhibitory effect on CTR1 and EIN2, and then regulates cell elongation through transcription factors EIN3/EIL1 and ERFs, which ultimately inhibits hypocotyl elongation [6]. Results here showed that ethylene inhibited cell elongation rather than proliferation since cell number was not altered after ACC treatment, consistent with reported data [49]. Following the illustration of the framework of ethylene signal transduction, more detailed mechanisms were revealed, e.g. EIN3 induced the transcription of PIF3, PIF3 regulated expression of a microtubule-related protein, MDP60, led microtubule re-organization, and enhanced hypocotyl growth [50]. Cross-talk between ethylene-CTR1 and Glc-TOR signaling pathway regulated the etiolated hypocotyl elongation by affecting multiple phosphorylation sites in EIN2 [51]. Moreover, ethylene regulates hypocotyl elongation by crosstalk with light signaling components, e.g. PhyB, PIF3, COP1, and HY5. Ethylene stimulates PIF3 to promote hypocotyl elongation under light, whereas photoactivated phyB may attenuate the promoting effect of ethylene. In darkness, ethylene inhibited hypocotyl elongation by stimulating ERF1 and WDL5. When seedlings grow up to the soil surface, increased light inhibit ethylene synthesis and eliminate the inhibitory effect of EIN3 on hypocotyl elongation [6]. Dozens of transcription factors are involved in ethylene signal transduction, e.g. PIFs, MYCs, HSFs, and CBFs, some of which are bHLH family members (PIF1/3/4/5, MYC2/3/4, RLS2/4, RHD6, etc.) [52]. Nevertheless, so far, the position and role of MYC2 or MYC3 in the ethylene signaling pathway have not been elucidated. Results here are of good help for understanding the role of the two transcription factors in ethylene signaling, as well as their role in the regulation of growth and development.
To verify the role of the two MYCs in ethylene signaling, the relationship of MYC2 or MYC3 with ERF1 and EIN3 was intensively investigated. The interaction between EIN3 and MYC2, MYC3, and MYC4, have been reported; as well as EIN3 binds and regulates transcription of ERF1 [15,53]. However, the relation between MYC2 or MYC3 and ERF1 was not revealed so far. As a crucial component in ethylene signaling, EIN3 can be stimulated by EIN2, and then stimulates multiple downstream signaling agents, including the apical curvature regulator HLS1 [38], the root hair elongation factor RSL4 [54], the senescence-related genes ORE1, SAG29 [55], ESE1 [56], cold stress-related genes CBF1/2/3 [57], and pathogenicity-related genes ERF1 and ORA59 [42], which in turn regulate growth of vegetative organs (e.g. apical hook, root hair, and leaves), as well as response to (a)biotic stresses [58]. The results in this work suggest that MYC2 and MYC3 are closely associated with EIN3, and involved in the ethylene-regulated hypocotyl growth, providing new evidence to clarify the mechanism of EIN3-involved regulation of plant growth.
AP2/ERF are essential transcription factors, which are involved in multiple physiological functions’ regulation [59,60,61]. ERF1 is involved in plant growth, development, and stress responses, which were modulated by various phytohormones, e.g. ethylene, JA, auxin, and ABA(Abscisic Acid) [42,62,63,64,65]. Expression of ERF1 is regulated by EIN3/EIL1, ABI3, or NIGT1.4, at the transcriptional level [65,66,67]. After translation, UBC18, MPK3/MPK6 may regulate ERF1 activity by protein modification, e.g. ubiquitination and phosphorylation. The abundance and activity of ERF1 directly affect cell metabolism, proliferation, and differentiation [68,69]. The regulatory mechanism of ERF1 expression was also intensively explored [64,65,70]. In this work, ERF1 expression was significantly increased in MYC2 and MYC3 null mutants, indicating that MYC2 and MYC3 may be involved in regulating ERF1 expression. Subsequently, EMSA and ChIP assays showed that MYC2 and MYC3 directly bind the promoter of ERF1 and inhibit its expression. The data indicated that MYCs may act at upstream of ERF1, providing novel clues for understanding the regulation mechanism of ERF1 expression. An assay of the relationship among EIN3, MYCs, and ERF1 showed that EIN3 suppressed the effect of MYC2 or MYC3 on ERF1 expression. MYC2 and MYC3 have been revealed to antagonize the effect of EIN3 on the expression of HLS1 [41]. Here, it is verfied that EIN3 inhibits the binding of MYC2 or MYC3 with ERF1 promoter, clarifying the inhibitory mechanism of EIN3 on MYCs-regulated ERF1 expression.
The analysis of DEGs after ACC treatment indicated that MYC2 and MYC3 may be essential components in ethylene-regulated gene expression. The complicated data showed that MYC2 and MYC3 may play positive roles in some genes’ expression, while playing negative roles in other genes’ expression. The data suggests that MYC2 and MYC3 affect the expression of multiple sets of functional genes that are involved in cellular metabolism, cell wall organization, growth and development, and stress responses. These effects may ultimately impact the elongation of hypocotyls. In the double-null mutant, impaired regulation of gene expression by MYC2 and MYC3 resulted in altered gene expression pattern and the enhanced sensitivity to ethylene. The mechanism needs to be intensively investigated in the future.
The data in the work suggest that EIN3 may be located upstream of the two MYCs and inhibit their transcription activity. MYC2 and MYC3 inhibit ERF1 expression. When MYC2 and MYC3 were mutated, ERF1 gene expression may be enhanced and the hypocotyls’ ethylene responsiveness is markedly increased (Figure 8).
4. Materials and Methods
4.1. Plant Materials
All Arabidopsis materials were derived from ecotype Col-0 background. The mutants, including myc2-1 (SALK _017005), myc2-2 (SALK_061267), and myc3 (SALK_01 2763), were obtained from the Arabidopsis Biological Resource Center (Columbus, OH, USA). The double null mutant myc2myc3 was obtained by crossing myc2-1 and myc3. The ERF1 knock-out mutants, erf1-1 and erf1-2, were obtained by CRISPR/Cas9 mutation in Col-0. Two ERF1 knock-out mutants, erf1-3 and erf1-4, were obtained by RNAi.
Seeds were surface sterilized with 70% ethanol, after being washed with sterilized water 3 times, seeds were sown on a solid medium consisting of 1/2 MS salt, 1% (w/v) agar, and 3% (w/v) sucrose, and then cultured under light (22°C, 16/8h light/dark, 130 μmol∙m-2∙s-1). For etiolated hypocotyl growth measurement, culture dishes were stored at 22°C under light for 6 h, and then coated with foil and cultured at 22 °C for 4 days.
After being cultured for some time, seedlings were photographed using a scanner (Perfection V39II, Epson), images were analyzed by using Image J software to measure hypocotyl length. Statistical analysis of the data was performed using IBM SPSS Statistics 22.0 software (IBM, New York, NY, USA).
4.2. Gene Transformation
The pSuper::MYC2 and pSuper::MYC3 plasmids were introduced into Agrobacterium tumefaciens (strain GV3101) using the freeze-thaw method. The fused genes were then transformed into wild type (Col-0) plants through floral dipping. Transgenic plants were screened using 1/2 MS medium containing 100 mg∙mL-1 kanamycin.
4.3. PCR Analysis
Total RNA was isolated from seedlings for quantitative real-time PCR analysis, which were performed using CFX96TM real-time PCR system (Bio-Rad, Hercules, CA, USA). Primers used for subsequent detection of ERF1 expression were 5’-ACACGTGCGGGATATCAAATA-3’ and 5’-ACACGTGCGGGATATCAAATA-3’. ACTIN2 was used as a reference (5’-GGTAACATTGTGCTCAGTGGTGG-3’ and 5’-AACGACCTTAATCTTCATGCTGC-3’). Three biological replicates and three technical replicates were used each time. Mean±SE was calculated and statistically analyzed.
4.4. Electrophoretic Mobility Shift Assay (EMSA)
EMSA was performed according to the reported method [49]. The recombinant GST-MYC2 and GST-MYC3 protein was expressed in E. coli and then extracted and purified. EMSA was conducted using the Light Shift Chemiluminescent EMSA Kit (Thermo Scientific, Rockford, IL, USA) with biotin-labeled and cold probes.
Nucleotide sequences of the double-stranded oligonucleotides were as follows:ERF1 P1: 5’-AAAACTTTGAACACGTGCGGGATATCAA-3’ and 5’-TTGATATCCCGCACGTGTTCAAAGTTTT-3’. ERF1 P2: 5’-GAAAAAATGGCACATGAAGTATCTTT-3’ and 5’-AAAGATACTTCATGTGCCATTTTTTC-3’. ERF1 P3: 5’-ACTCAGGATGCATGTGATGATGTGTA-3’ and 5’-TACACATCATCACATGCATCCTGAGT-3’. ERF1 P4: 5’-TTATCTTCTAAACATGAGATGGCTCG-3’ and 5’-CGAGCCATCTCATGTTTAGAAGATAA-3’. ERF1 P5: 5’-TTAGTTGCGTATCCGTTCGAATAATT-3’ and 5’-AATTATTCGAACGGATACGCAACTAA-3’.
4.5. Chromatin Immunoprecipitation (ChIP)
10-days-old seedlings of pSuper::MYC2-GFP or pSuper::MYC3-GFP transformant were fixed in 1% formaldehyde, and then were homogenized and sonicated. The sheared chromatins were immunoprecipitated, washed, reverse cross-linked, and amplified. The monoclonal anti-GFP antibody was used for immunoprecipitation. Approximately 10% of sonicated but non-immunoprecipitated chromatin was reverse cross-linked and used as an input DNA control. The immunoprecipitated DNA and input DNA were analyzed by using real-time q-PCR. Primer used to detect the MYC2 and MYC3 target ERF1 promoter was 5’-ACACGTGCGGGATATCAAATA-3’ and 5’-CATGCATCCTGAGTCC AGTAG-3’, ACTIN2 as a control (5’-GGTAACATTGTGCTCAGTGGTGG-3’ and 5’-AACGACCTTAATCTTCATGCTGC-3’).
4.6. Gene Transient Expression in N. benthamiana
ERF1pro::GUS or ERF1mpro::GUS with pCAMBIA1391, pSuper::MYC2, pSuper::MYC3 or pSuper::EIN3 with pCAMBIA1300 constructs were transformed into A. tumefaciens (strain GV3101). Cultured A. tumefaciens cells were harvested by centrifugation and suspended and diluted with a solution consisting of 10 mM MES, 10 mM MgCl2, and 200 mM acetosyringone, pH 5.6, to an optical density (600 nm, OD=0.7). Cells were injected into N. benthamiana leaves by using a needle-free syringe. GUS activity of the infiltrated leaves was quantitatively assessed.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure S1. The effect of serial concentration of ACC on elongation of etiolated hypocotyls of wild type and null mutants of MYC2 or MYC3. Figure S2. The effect of serial concentration of ACC on elongation of etiolated hypocotyls of wild type and the complementation lines of MYC2 or MYC3. Figure S3. MYC2 or MYC3 is not involved in ethylene-promoted hypocotyl elongation of seedlings grown under light. Figure S4. MYC2 or MYC3 did not bind P2-P5 of ERF1 promoter. Figure S5. MYC2 or MYC3 did not interact with EIN2. Figure S6. KEGG analysis of ethylene-regulated DEGs in Col-0 and myc2myc3. Table S1. DEGs that were up-regulated in myc2myc3 while not up-regulated in Col-0 after ACC treatment. Table S2. DEGs that were down-regulated in myc2myc3 while not down-regulated in Col-0 after ACC treatment.
Author Contributions
E.K. and Z.S designed the experiment and finished writing the article. Y.L.(Yuke Li), Y.C., F.W., Y.L.(Yingxiao Liu), and R.Z. performed research. C.X., P.Z., and J.Z. provided partial material.
Funding
This work was supported by the National Natural Science Foundation of China (Grant No. 31800233, 31871409), the Science Research Project of Hebei Education Department (BJK2023093), the Advanced Talents Foundation of Hebei Education Department (Grant No. GCC2014037), the Graduate Innovation Research Project of Hebei Normal University (Grant No. XCXZZBS202309), the Provincial Fundamental Research Funds for Universities (JYZ202007), the Outstanding Youth Fund of Hebei University of Traditional Chinese Medicine (YQ2020002).
Data Availability Statement
Data available on request from the authors.
Acknowledgments
We thank all the colleagues who helped with the development of the different parts of this manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Li, X.; Liu, C.; Zhao, Z.; Ma, D.; Zhang, J.; Yang, Y.; Liu, Y.; Liu, H. COR27 and COR28 are novel regulators of the COP1-HY5 regulatory hub and photomorphogenesis in Arabidopsis. Plant Cell 2020, 32, 3139–3154. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Bao, Y. PIF4: Integrator of light and temperature cues in plant growth. Plant Sci. 2021, 313, 111086. [Google Scholar] [CrossRef] [PubMed]
- Ahammed, G. J.; Gantait, S.; Mitra, M.; Yang, Y. X.; Li, X. Role of ethylene crosstalk in seed germination and early seedling development: A review. Plant Physiol. Biochem. 2020, 151, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Kris Vissenberg, B. A. K. The Arabidopsis thaliana hypocotyl, a model to identify and study control mechanisms of cellular expansion. Plant Cell Rep. 2014, 33, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Tavridou, E.; Pireyre, M.; Ulm, R. Degradation of the transcription factors PIF4 and PIF5 under UV-B promotes UVR8-mediated inhibition of hypocotyl growth in Arabidopsis. Plant J. 2019, 101, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.W.; Huang, R.F. Integration of ethylene and light signaling affects hypocotyl growth in Arabidopsis. Front. Plant Sci. 2017, 8, 57. [Google Scholar] [CrossRef] [PubMed]
- Hua, J.; Meyerowitz, E.M. Ethylene responses are negatively regulated by a receptor gene family in Arabidopsis thaliana. Cell 1998, 94, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Bakshi, A.; Piya, S.; Fernandez, J. C.; Chervin, C.; Hewezi, T.; Binder, B.M. Ethylene receptors signal via a noncanonical pathway to regulate abscisic acid responses. Plant Physiol. 2018, 176, 910–929. [Google Scholar] [CrossRef] [PubMed]
- Shi, J. J.; Zhu, Z. Q. Seedling morphogenesis: when ethylene meets high ambient temperature. aBIOTECH 2021, 3, 40–48. [Google Scholar] [CrossRef]
- Binder, B. M. Ethylene signaling in plants. J. Biol. Chem. 2020, 295, 7710–7725. [Google Scholar] [CrossRef]
- Hao, D.; Jin, L.; Wen, X.; Yu, F.; Xie, Q.; Guo, H. The RING E3 ligase SDIR1 destabilizes EBF1/EBF2 and modulates the ethylene response to ambient temperature fluctuations in Arabidopsis. Proc. Natl. Acad. Sci. USA 2021, 118, e2024592118. [Google Scholar] [CrossRef]
- Zhong, S. W.; Shi, H.; Xue, C.; Wang, L.; Xi, Y. P.; Li, J. G.; Quail, P. H.; Deng, X.; Guo, H. W. A molecular framework of light-controlled phytohormone action in Arabidopsis. Curr. Biol. 2012, 22, 1530–1535. [Google Scholar] [CrossRef] [PubMed]
- Roman, G.; Lubarsky, B.; Kieber, J. J.; Rothenberg, M.; Ecker, J.R. Genetic analysis of ethylene signal transduction in Arabidqsis thaliana: five novel mutant loci integrated into a stress response pathway. Genetics 1995, 139, 1393–1409. [Google Scholar] [CrossRef]
- Ortigosa, A.; Fonseca, S.; Franco-Zorrilla, J. M.; Fernández-Calvo, P.; Zander, M.; Lewsey, M. G.; García-Casado, G.; Fernández-Barbero, G.; Ecker, J. R.; Solano, R. The JA-pathway MYC transcription factors regulate photomorphogenic responses by targeting HY5 gene expression. Plant J. 2020, 102, 138–152. [Google Scholar] [CrossRef] [PubMed]
- Solano, R.; Stepanova, A.; Chao, Q. M.; Ecker, J. R. Nuclear events in ethylene signaling: a transcriptional cascade mediated by ETHYLENE-INSENSITIVE3 and ETHYLENE-RESPONSE-FACTOR1. Genes Dev. 1998, 12, 3703–3714. [Google Scholar] [CrossRef]
- Zhong, S.; Shi, H.; Xue, C.; Wei, N.; Guo, H.; Deng, X.W. Ethylene-orchestrated circuitry coordinates a seedling’s response to soil cover and etiolated growth. Proc. Natl. Acad. Sci.USA 2014, 111, 3913–3920. [Google Scholar] [CrossRef]
- Ludwig, S.R.; Habera, L.F.; Dellaporta, S.L.; Wessler, S.R. Lc, a member of the maize R gene family responsible for tissue-specific anthocyanin production, encodes a protein similar to transcriptional activators and contains the myc-homology region. Genetics 1989, 86, 7092–7096. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Calvo, P.; Chini, A.; Fernández-Barbero, G.; Chico, J.M.; Gimenez-Ibanez, S.; Geerinck, J.; Eeckhout, D.; Schweizer, F.; Godoy, M.; Franco-Zorrilla, J.M.; et al. The Arabidopsis bHLH transcription factors MYC3 and MYC4 are targets of JAZ repressors and act additively with MYC2 in the activation of jasmonate responses. Plant Cell 2011, 23, 701–715. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Du, J.; Han, X.; Li, H.; Kui, M.; Zhang, J.; Huang, Z.; Fu, Q.; Jiang, Y.; Hu, Y. PHOSPHATE STARVATION RESPONSE1 (PHR1) interacts with JASMONATE ZIM-DOMAIN (JAZ) and MYC2 to modulate phosphate deficiency-induced jasmonate signaling in Arabidopsis. Plant Cell 2023, 35, 2132–2156. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Yang, H.; Gao, H.; Yan, J.; Xie, D. Control of seed size by jasmonate. Sci. China Life Sci. 2021, 64, 1215–1226. [Google Scholar] [CrossRef]
- Bao, S.J.; Hua, C.M.; Huang, G.Q.; Cheng, P.; Gong, X.M.; Shen, L.S.; Yu, H. Molecular basis of natural variation in photoperiodic flowering responses. Dev. Cell 2019, 50, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Sun, J.Q.; Zhai, Q.Z.; Zhou, W.K.; Qi, L.L.; Xu, L.; Wang, B.; Chen, R.; Jiang, H. L.; Qi, J.; et al. The basic Helix-Loop-Helix transcription factor MYC2 directly represses PLETHORA expression during jasmonate-mediated modulation of the root stem cell niche in Arabidopsis. Plant Cell 2011, 23, 3335–3352. [Google Scholar] [CrossRef] [PubMed]
- Yi, R.; Yan, J.B.; Xie, D.X. Light promotes jasmonate biosynthesis to regulate photomorphogenesis in Arabidopsis. Sci. China Life Sci. 2020, 63, 943–952. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, M.; Sakuraba, Y.; Yanagisawa, S.C. A jasmonate-activated MYC2-Dof2.1-MYC2 transcriptional loop promotes leaf senescence in Arabidopsis. Plant Cell 2020, 32, 242–262. [Google Scholar] [CrossRef] [PubMed]
- Fujikawa, I.; Takehara, Y.; Ota, M.; Imada, K.; Sasaki, K.; Kajihara, H.; Sakai, S.; Jogaiah, S.; Ito, S.I. Magnesium oxide induces immunity against Fusarium wilt by triggering the jasmonic acid signaling pathway in tomato. J. Biotech. 2021, 325, 100–108 https: //doiorg/101016/jjbiotec202011012. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Wang, Y.; Qiu, L.; Han, X.; Zhu, Y.; Liu, L.; Man, M.; Li, F.; Ren, M.; Xing, Y. MYC2: A master switch for plant physiological processes and specialized metabolite synthesis. Int. J. Mol. Sci. 2023, 24, 3511. [Google Scholar] [CrossRef]
- Zhao, P.Z.; Zhang, X.; Gong, Y.Q.; Wang, D.; Xu, D.Q.; Wang, N.; Sun, Y.W.; Gao, L.B.; Liu, S.S.; Deng, X.W.; et al. Red-light is an environmental effector for mutualism between begomovirus and its vector whitefly. PLOS Pathog. 2021, 17, e1008770 https:// doiorg/101371/journalppat1008770. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Geng, Q.; Wu, X.; Hu, M.; Ye, M.; Yu, X.; Chen, Y.; Xu, J.; Jiang, L.; Cao, S. The transcription factor MYC1 interacts with FIT to negatively regulate iron homeostasis in Arabidopsis thaliana. Plant J. 2023, 114, 193–208. [Google Scholar] [CrossRef]
- Verma, D.; Jalmi, S.K.; Bhagat, P.K.; Verma, N.; Sinha, A.K. A bHLH transcription factor, MYC2, imparts salt intolerance by regulating proline biosynthesis in Arabidopsis. FEBS J. 2020, 287, 2560–2576. [Google Scholar] [CrossRef]
- Moerkercke, A.V.; Duncan, O.; Zander, M.; Šimura, J.; Broda, M.; Bossche, R.V.; Lewsey, M.G.; Lama, S.; Singh, K.B.; Ljung, K.; et al. A MYC2/MYC3/MYC4-dependent transcription factor network regulates water spray-responsive gene expression and jasmonate levels. Proc. Natl. Acad. Sci. USA 2019, 116, 23345–23356. [Google Scholar] [CrossRef]
- Kazan, K.; Manners, J.M. MYC2: The master in action. Mol. Plant 2013, 6, 686–703. [Google Scholar] [CrossRef]
- Chen, X.Y.; Wang, D.D.; Fang, X.; Chen, X.Y.; Mao, Y.B. Plant specialized metabolism regulated by jasmonate signaling. Plant Cell Physiol. 2019, 60, 2638–2647. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.D.; Li, P.; Chen, Q.Y.; Chen, X.Y.; Yan, Z.W.; Wang, M.Y.; Mao, Y.B. Differential contributions of MYCs to insect defense reveals flavonoids alleviating growth inhibition caused by wounding in Arabidopsis. Front. Plant Sci. 2021, 12, 700555. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Kim, K.; Kim, M.E.; Kim, H.G.; Heo, G.S.; Parka, O.K.; Park, Y.I.; Choi, G.; Oh, E. Phytochrome and ethylene signaling integration in occurs via the transcriptional regulation of genes co-targeted by PIFs and EIN3. Front. Plant Sci. 2016, 7, 1055. [Google Scholar] [CrossRef] [PubMed]
- Khanna, R.; Shen, Y.; Marion, C.M.; Tsuchisaka, A.; Theologis, A.; Schäfer, E.; Quail, P.H. The basic helix-loop-helix transcription factor PIF5 acts on ethylene biosynthesis and phytochrome signaling by distinct mechanisms. Plant Cell 2007, 19, 3915–3929. [Google Scholar] [CrossRef] [PubMed]
- Gallego-Bartolomé, J.; Arana, M.V.; Vandenbussche, F.; Zádníková, P.; Minguet, E.G.; Guardiola, V.; Van Der Straeten, D.; Benkova, E.; Alabadí, D.; Blázquez, M.A. Hierarchy of hormone action controlling apical hook development in Arabidopsis. Plant J. 2011, 67, 622–634. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.W.; Zhao, M.T.; Shi, T.Y.; Shi, H.; An, F.Y.; Zhao, Q.; Guo, H.W. EIN3/EIL1 cooperate with PIF1 to prevent photo-oxidation and to promote greening of seedlings. Proc. Natl. Acad. Sci. USA 2009, 106, 21431–21436. [Google Scholar] [CrossRef]
- Zhang, X.; Ji, Y.S.; Xue, C.; Ma, H.H.; Xi, Y.L.; Huang, P.X.; Wang, H.; An, F.Y.; Li, B.S.; Wang, Y.C.; et al. Integrated regulation of apical hook development by transcriptional coupling of EIN3/EIL1 and PIFs in Arabidopsis. Plant Cell 2018, 30, 1971–1988. [Google Scholar] [CrossRef]
- Wang, Y.C.; Guo, H.W. On hormonal regulation of the dynamic apical hook development. New Phytol. 2019, 222, 1230–1234. [Google Scholar] [CrossRef] [PubMed]
- Aizezi, Y.; Shu, H.Z.; Zhang, L.L.; Zhao, H.M.; Peng, Y.; Lan, H.X.; Xie, Y.P.; Li, J.; Wang, Y.C.; Guo, H.W.; Jiang, K. Cytokinin regulates apical hook development via the coordinated actions of EIN3/EIL1 and PIF transcription factors in Arabidopsis. J. Exp. Bot. 2022, 73, 213–227. [Google Scholar] [CrossRef]
- Zhang, X.; Zhu, Z.Q.; An, F.Y.; Hao, D.D.; Li, P.P.; Song, J.H.; Yi, C.Q.; Guo, H.W. Jasmonate-activated MYC2 represses ETHYLENE INSENSITIVE3 activity to antagonize ethylene-promoted apical hook formation in Arabidopsis. Plant Cell 2014, 26, 1105–1117. [Google Scholar] [CrossRef] [PubMed]
- Song, S.S.; Huang, H.; Gao, H.; Wang, J.J.; Wu, D.W.; Liu, X.L.; Yang, S.H.; Zhai, Q.Z.; Li, C.Y.; Qi, T.C.; et al. Interaction between MYC2 and ETHYLENE INSENSITIVE3 modulates antagonism between jasmonate and ethylene signaling in Arabidopsis. Plant Cell 2014, 26, 263–279. [Google Scholar] [CrossRef] [PubMed]
- Abe, H.; Urao, T.; Ito, T.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in abscisic acid signaling. Plant Cell 2003, 15, 63–78. [Google Scholar] [CrossRef] [PubMed]
- Breeze, E. , Master MYCs: MYC2, the jasmonate signaling “master switch”. Plant Cell 2019, 31, 9–10. [Google Scholar] [CrossRef]
- Chakraborty, M.; Gangappa, S.N.; Maurya, J.P.; Sethi, V.; Srivastava, A.K.; Singh, A.; Dutta, S.; Ojha, M.; Gupta, N.; Sengupta, M.; et al. Functional interrelation of MYC2 and HY5 plays an important role in Arabidopsis seedling development. Plant J. 2019, 99, 1080–1097. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.Y.; Li, Z.Q.; Zhao, Y.D.; Shen, W.J.; Chen, M.S.; Gao, H.Q.; Ge, X.M.; Wang, H.Q.; Li, X.; He, J.M. Ethylene-induced stomatal closure is mediated via MKK1/3<italic>-</italic>MPK3/6 cascade to EIN2 and EIN3. J. Integr. Plant Biol. 2021, 63, 1324–1340. [Google Scholar] [CrossRef] [PubMed]
- Gamalero, E.; Lingua, G.; Glick, B.R. Ethylene, ACC, and the plant growth-promoting enzyme ACC deaminase. Biology (Basel) 2023, 12, 1043. [Google Scholar] [CrossRef]
- Liu, K.; Li, Y.; Chen, X.; Li, L.; Liu, K.; Zhao, H.; Wang, Y.; Han, S. ERF72 interacts with ARF6 and BZR1 to regulate hypocotyl elongation in Arabidopsis. J. Exp. Bot. 2018, 69, 3933–3947. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Ma, Q.; Mao, T. Ethylene regulates the Arabidopsis microtubule-associated protein WAVE-DAMPENED2-LIKE5 in etiolated hypocotyl elongation. Plant Physiol. 2015, 169, 325–337. [Google Scholar] [CrossRef]
- Ma, Q.; Wang, X.; Sun, J.; Mao, T. Coordinated regulation of hypocotyl cell elongation by light and ethylene through a microtubule destabilizing protein. Plant Physiol. 2018, 176, 678–690. [Google Scholar] [CrossRef]
- Fu, L.W.; Liu, Y.L.; Qin, G.C.; Wu, P.; Zi, H.L.; Xu, Z.T.; Zhao, X.D.; Wang, Y.; Li, Y.X.; Yang, S.H.; et al. The TOR-EIN2 axis mediates nuclear signalling to modulate plant growth. Nature 2021, 591, 288–292. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.Y.; Zhao, X.B.; Bürger, M.; Chory, J.; Wang, X.C. The role of ethylene in plant temperature stress response. Trends Plant Sci. 2023, 28, 808–824. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.; Munné-Bosch, S. Ethylene response factors: a key regulatory hub in hormone and stress signaling. Plant Physiol. 2015, 169, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Xu, P.; Li, B.S.; Li, P.P.; Wen, X.; An, F.Y.; Gong, Y.; Xin, Y.; Zhu, Z.Q.; Wang, Y.C.; et al. Ethylene promotes root hair growth through coordinated EIN3/EIL1 and RHD6/RSL1 activity in Arabidopsis. Proc. Natl. Acad. Sci. USA 2017, 114, 13834–13839. [Google Scholar] [CrossRef]
- Wang, C.Q.; Dai, S.Y.; Zhang, Z.L.; Lao, W.Q.; Wang, R.Y.; Meng, X.Q.; Zhou, X. Ethylene and salicylic acid synergistically accelerate leaf senescence in Arabidopsis. J. Integr. Plant Biol. 2021, 63, 828–833. [Google Scholar] [CrossRef] [PubMed]
- Quan, R.D.; Wang, J.; Yang, D.X.; Zhang, H.W.; Zhang, Z.J.; Huang, R.F. EIN3 and SOS2 synergistically modulate plant salt tolerance. Sci. Rep. 2017, 7, 44637. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.T.; Tian, S.W.; Hou, L.Y.; Huang, X.Z.; Zhang, X.Y.; Guo, H.W.; Yang, S.H. Ethylene signaling negatively regulatesfreezing tolerance by repressing expression of CBF and type-A ARR Genes in Arabidopsis. Plant Cell 2012, 24, 2578–2595. [Google Scholar] [CrossRef] [PubMed]
- Dolgikh, V.A.; Pukhovaya, E.M.; Zemlyanskaya, E.V. Shaping ethylene response: The role of EIN3/EIL1 transcription factors. Front. Plant Sci. 2019, 10, 1030. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.; Guo, Z.H.; Hao, P.P.; Wang, G.M.; Jin, Z.M.; Zhang, S.L. Multiple regulatory roles of AP2/ERF transcription factor in angiosperm. Bot. Stud. 2017, 58, 6. [Google Scholar] [CrossRef]
- Phukan, U.J.; Jeena, G.S.; Tripathi, V.; Shukla, R.K. Regulation of apetala2/ethylene response factors in plants. Front. Plant Sci. 2017, 8, 150. [Google Scholar] [CrossRef]
- Shoji, T.; Yuan, L. ERF gene clusters: working together to regulate metabolism. Trends Plant Sci. 2021, 26, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.C.; Liao, P.M.; Kuo, W.W.; Lin, T.P. , The Arabidopsis ETHYLENE RESPONSE FACTOR1 regulates abiotic stress-responsive gene expression by binding to different cis-acting elements in response to different stress signals. Plant Physiol. 2013, 162, 1566–1582. [Google Scholar] [CrossRef] [PubMed]
- Corbineau, F.; Xia, Q.; Bailly, C.; El-Maarouf-Bouteau, H. Ethylene, a key factor in the regulation of seed dormancy. Front. Plant Sci. 2014, 5, 539. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.J.; Mao, J.L.; Miao, Z.Q.; Wang, Z.; Yu, L.H.; Cai, X.T.; Xiang, C.B. <italic>Arabidopsis</italic> ERF1 mediates cross-talk between ethylene and auxin biosynthesis during primary root elongation by regulating ASA1 expression. PLOS Genet. 2016, 12, e1005760. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhao, P.X.; Chen, S.Y.; Sun, L.Q.; Mao, J.L.; Tan, S.T.; Xiang, C.B. The ABI3-ERF1 module mediates ABA-auxin crosstalk to regulate lateral root emergence. Cell Rep. 2023, 42, 112809. [Google Scholar] [CrossRef] [PubMed]
- Takaoka, Y.; Iwahashi, M.; Chini, A.; Saito, H.; Ishimaru, Y.; Egoshi, S.; Kato, N.; Tanaka, M.; Bashir, K.; Seki, M.; et al. A rationally designed JAZ subtype-selective agonist of jasmonate perception. Nat. Commun. 2018, 9, 3654. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.F.; Zeng, L.; Lv, X.D.; Guo, J.H.; Li, X.Y.; Zhang, X.H.; Wang, D.; Wang, J.Y.; Bi, J.L.; Julkowska, M.M.; Li, B. NIGT1.4 maintains primary root elongation in response to salt stress through induction of ERF1 in Arabidopsis. Plant J. 2023, 116, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.C.; Kuo, W.C.; Wang, Y.M.; Chen, H.Y.; Lin, T.P. UBC18 mediates ERF1 degradation under light-dark cycles. New Phytol. 2016, 213, 1156–1167. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.G.; Mu, Q.; Wang, X.Y.; Zhang, J.; Yu, H.Z.; Huang, T.Z.; He, Y.X.; Dai, S.J.; Meng, X.Z. Multilayered synergistic regulation of phytoalexin biosynthesis by ethylene, jasmonate, and MAPK signaling pathways in Arabidopsis. Plant Cell 2022, 34, 3066–3087. [Google Scholar] [CrossRef]
- Chen, Y.L.; Zhang, L.P.; Zhang, H.Y.; Chen, L.G.; Yu, D.Q. ERF1 delays flowering through direct inhibition of FLOWERING LOCUS T expression in Arabidopsis. J. Integr. Plant Biol. 2021, 63, 1712–1723. [Google Scholar] [CrossRef]
Figure 1.
MYC2 and MYC3 negatively regulates ethylene-inhibited elongation of etiolated hypocotyls. (A). Photograph of etiolated seedlings which were grown on 1/2 MS medium containing serial concentration of ACC for 4 days. Scale bar =1 mm. (B-C). Mean±SE value of hypocotyl length (B) and the ratio ((ACC treatment/control)*100%) of hypocotyl length (C). In each experiment, at least 40 seedlings were measured. Data from 3 replicates were calculated and statistically analyzed using one-way ANOVA.
Figure 1.
MYC2 and MYC3 negatively regulates ethylene-inhibited elongation of etiolated hypocotyls. (A). Photograph of etiolated seedlings which were grown on 1/2 MS medium containing serial concentration of ACC for 4 days. Scale bar =1 mm. (B-C). Mean±SE value of hypocotyl length (B) and the ratio ((ACC treatment/control)*100%) of hypocotyl length (C). In each experiment, at least 40 seedlings were measured. Data from 3 replicates were calculated and statistically analyzed using one-way ANOVA.
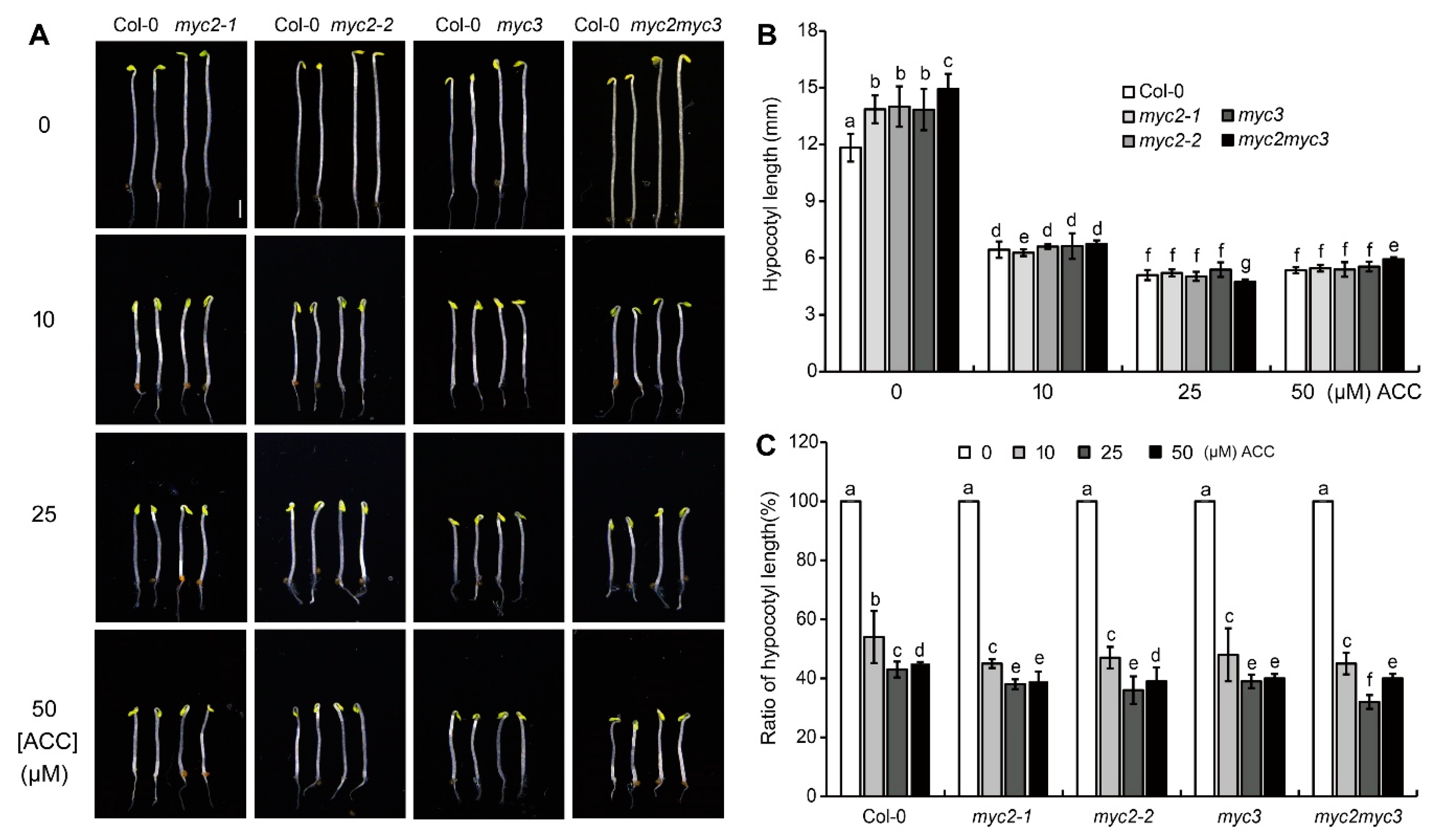
Figure 2.
MYC2 and MYC3 negatively regulated ethylene-inhibited elongation of etiolated hypocotyl epidermal cell. (A). SEM photograph of epidermal cells of etiolated seedlings which were grown on MS medium containing 25 μM ACC for 4 days. (B-C). Mean±SE of hypocotyl epidermal cell length (B) and the ratio of hypocotyl epidermal cell length after/before ACC treatment (C). In each experiment, at least 10 seedlings were investigated. Data from 3 replicates were calculated and statistically analyzed using one-way ANOVA.
Figure 2.
MYC2 and MYC3 negatively regulated ethylene-inhibited elongation of etiolated hypocotyl epidermal cell. (A). SEM photograph of epidermal cells of etiolated seedlings which were grown on MS medium containing 25 μM ACC for 4 days. (B-C). Mean±SE of hypocotyl epidermal cell length (B) and the ratio of hypocotyl epidermal cell length after/before ACC treatment (C). In each experiment, at least 10 seedlings were investigated. Data from 3 replicates were calculated and statistically analyzed using one-way ANOVA.
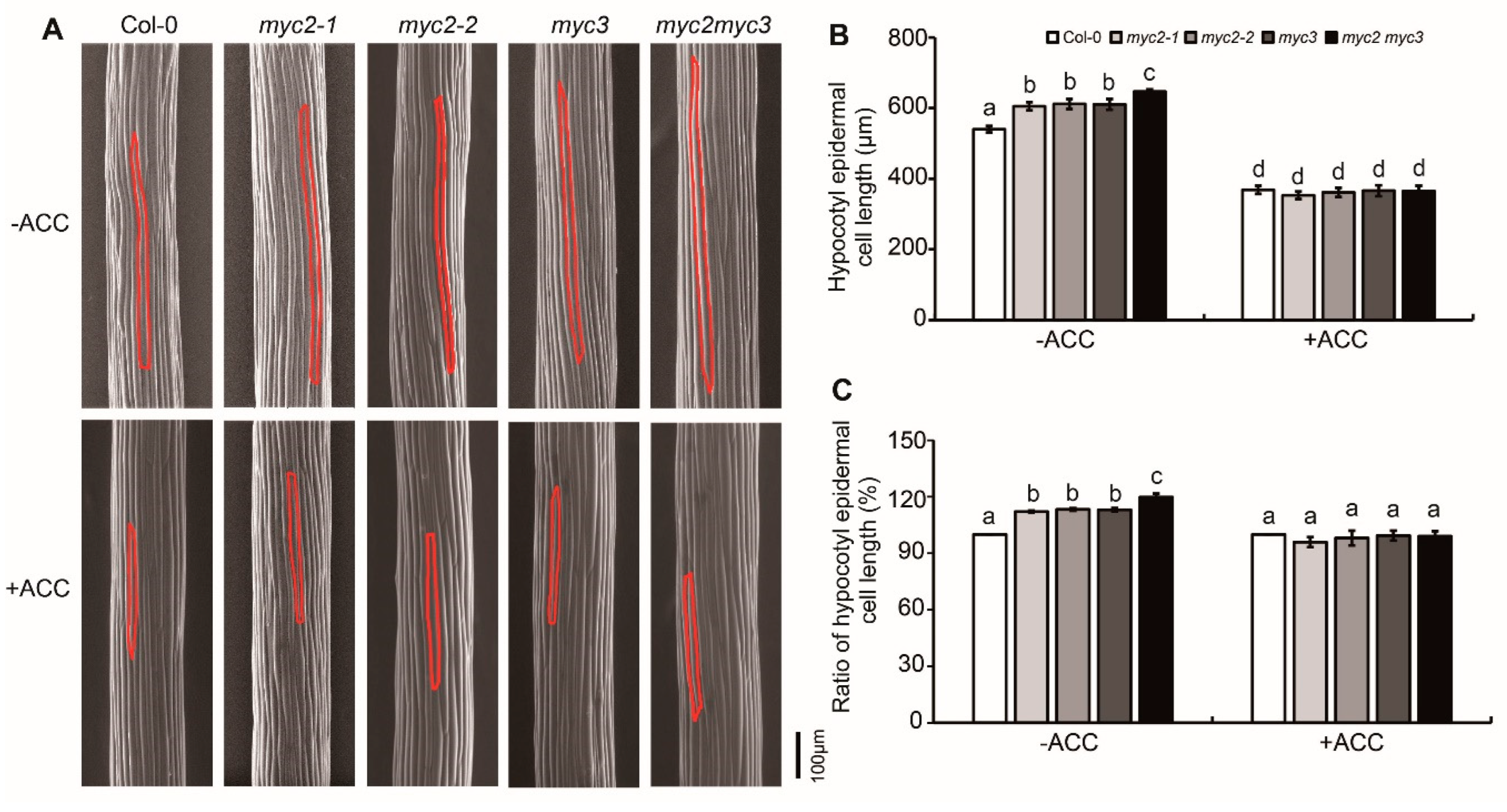
Figure 3.
ERF1 is a target of MYC2 and MYC3. (A). ERF1 expression level in etiolated wild type and mutants’ seedlings. (B). Five predicted MYCs-binding sites in the promoter region of ERF1. (C-D). EMSA results of the binding of MYC2 (C) or MYC3(D) with ERF1 promoter. (E-F). ChIP-qPCR assay of MYC2 and MYC3 binding to ERF1 in vivo. In figure A, E and F, mean±SE of data from three replicates were calculated and statistically analyzed using one-way ANOVA.
Figure 3.
ERF1 is a target of MYC2 and MYC3. (A). ERF1 expression level in etiolated wild type and mutants’ seedlings. (B). Five predicted MYCs-binding sites in the promoter region of ERF1. (C-D). EMSA results of the binding of MYC2 (C) or MYC3(D) with ERF1 promoter. (E-F). ChIP-qPCR assay of MYC2 and MYC3 binding to ERF1 in vivo. In figure A, E and F, mean±SE of data from three replicates were calculated and statistically analyzed using one-way ANOVA.
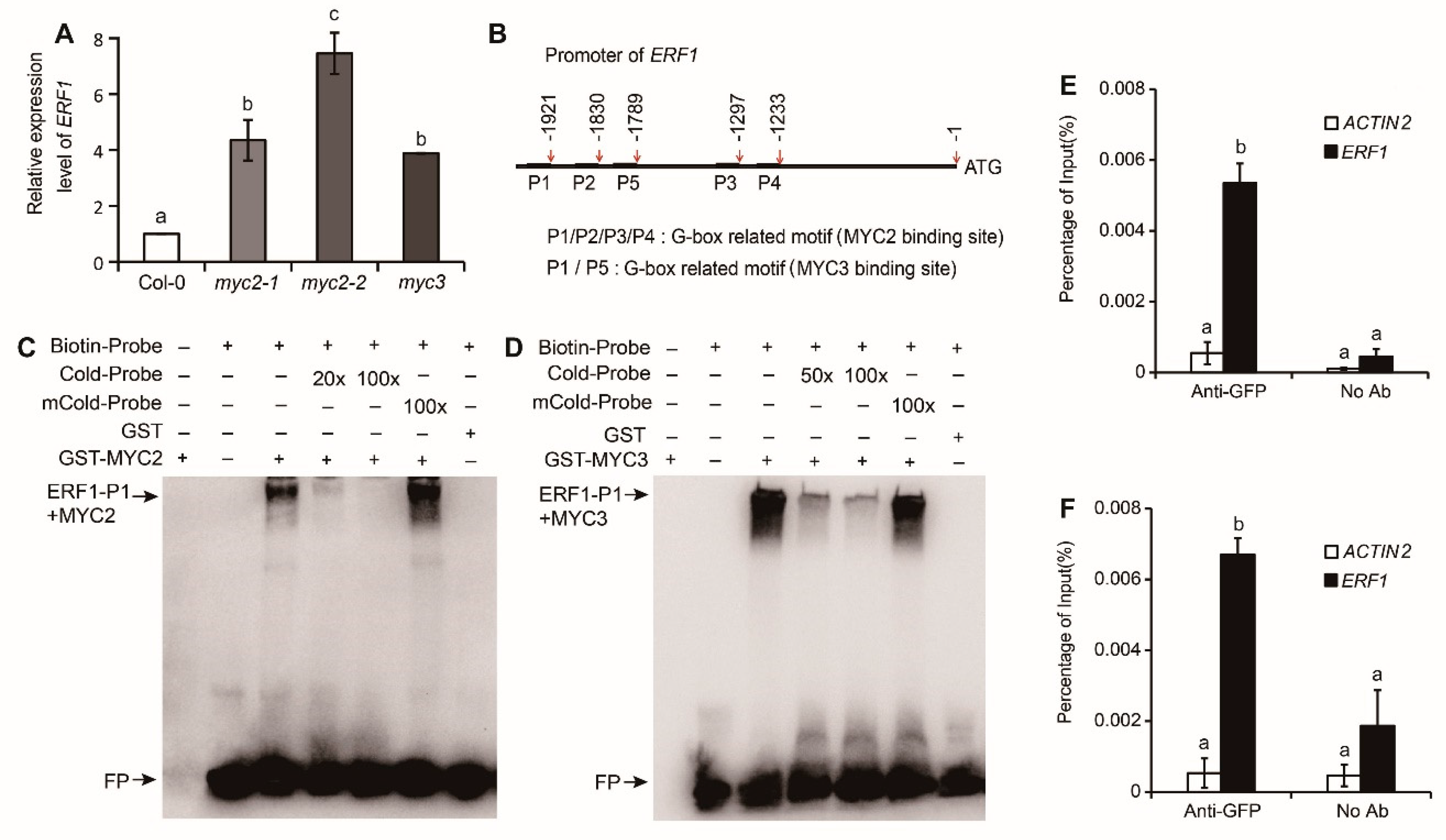
Figure 4.
EIN3 suppressed the transcriptional activity of MYC2 or MYC3. (A). Schematic image of the reporter and effector used in transient transactivation assays. (B-C). Transient expression of MYC2, MYC3, EIN3 and ERF1pro::GUS or ERF1mpro::GUS (ERF1m1pro: EIN3 binding site was mutated; ERF1m2pro: P1 was mutated; ERF1m3pro: EIN3 binding site and P1 were both mutated) in N. benthamiana leaves. ERF1pro::GUS or ERF1mpro::GUS expression level was set to 1. Mean±SE of data from three replicates were calculated and statistically analyzed using one-way ANOVA. (D-E). EIN3 blocked the binding of MYC2 or MYC3 with ERF1 promoter. The binding of MYC2 (D) or MYC3 (E) with P1 was detected using EMSA.
Figure 4.
EIN3 suppressed the transcriptional activity of MYC2 or MYC3. (A). Schematic image of the reporter and effector used in transient transactivation assays. (B-C). Transient expression of MYC2, MYC3, EIN3 and ERF1pro::GUS or ERF1mpro::GUS (ERF1m1pro: EIN3 binding site was mutated; ERF1m2pro: P1 was mutated; ERF1m3pro: EIN3 binding site and P1 were both mutated) in N. benthamiana leaves. ERF1pro::GUS or ERF1mpro::GUS expression level was set to 1. Mean±SE of data from three replicates were calculated and statistically analyzed using one-way ANOVA. (D-E). EIN3 blocked the binding of MYC2 or MYC3 with ERF1 promoter. The binding of MYC2 (D) or MYC3 (E) with P1 was detected using EMSA.

Figure 5.
ERF1 is involved in ethylene regulated elongation of etiolated hypocotyl. (A). Schematic diagram of ERF1 gene structure. The black boxes note the exons. Red frame note the point of a T base insertion. Sequence of gRNA target site of ERF1 was noted below. (B). Overexpression of ERF1 suppressed elongation of etiolated hypocotyls. Seedlings were grown on 1/2 MS medium in darkness for 4 days. (C-D). The effect of ACC treatment on elongation of etiolated hypocotyl of ERF1 null mutants or OE lines. Hypocotyl length (C) and ratio of hypocotyl length (D) were showed, respectively. In each experiment, at least 40 seedlings were measured. Data from 3 replicates were calculated and statistically analyzed using one-way ANOVA.
Figure 5.
ERF1 is involved in ethylene regulated elongation of etiolated hypocotyl. (A). Schematic diagram of ERF1 gene structure. The black boxes note the exons. Red frame note the point of a T base insertion. Sequence of gRNA target site of ERF1 was noted below. (B). Overexpression of ERF1 suppressed elongation of etiolated hypocotyls. Seedlings were grown on 1/2 MS medium in darkness for 4 days. (C-D). The effect of ACC treatment on elongation of etiolated hypocotyl of ERF1 null mutants or OE lines. Hypocotyl length (C) and ratio of hypocotyl length (D) were showed, respectively. In each experiment, at least 40 seedlings were measured. Data from 3 replicates were calculated and statistically analyzed using one-way ANOVA.
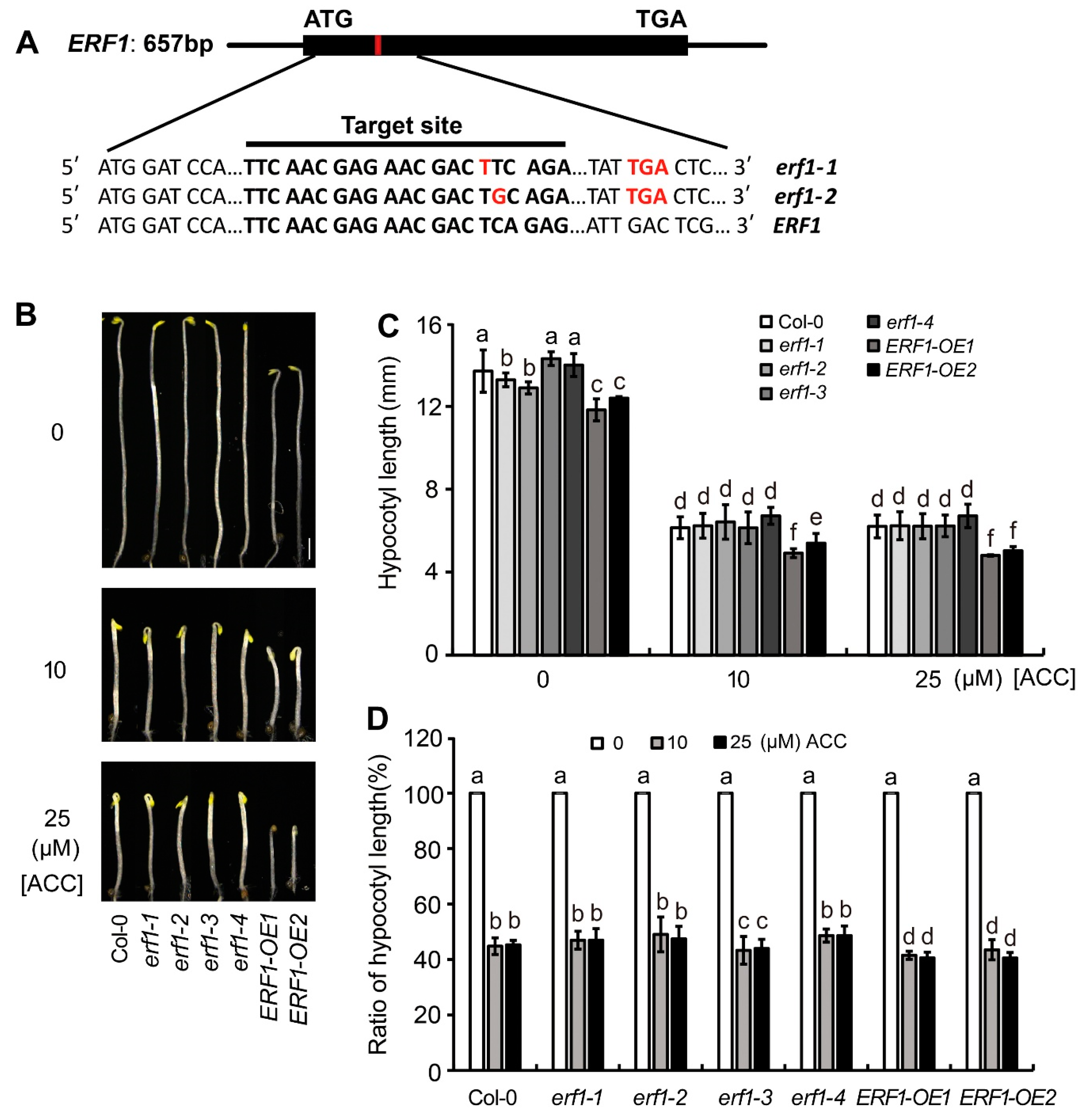
Figure 6.
MYC2 and MYC3 function at upstream of ERF1. Null mutants of MYC2 or MYC3 were hybridized with erf1-3 or ERF1-OE1. Effect of ACC on seedlings of hybrid progenies was detected. (A). Photograph of etiolated seedlings which were grown on ACC-containing medium for 4 days. In figure B and C, hypocotyl length (B) and ratio of hypocotyl length (C) after/before ACC treatment were showed, respectively. In each experiment, at least 40 seedlings were measured. Data from 3 replicates were calculated and statistically analyzed using one-way ANOVA.
Figure 6.
MYC2 and MYC3 function at upstream of ERF1. Null mutants of MYC2 or MYC3 were hybridized with erf1-3 or ERF1-OE1. Effect of ACC on seedlings of hybrid progenies was detected. (A). Photograph of etiolated seedlings which were grown on ACC-containing medium for 4 days. In figure B and C, hypocotyl length (B) and ratio of hypocotyl length (C) after/before ACC treatment were showed, respectively. In each experiment, at least 40 seedlings were measured. Data from 3 replicates were calculated and statistically analyzed using one-way ANOVA.
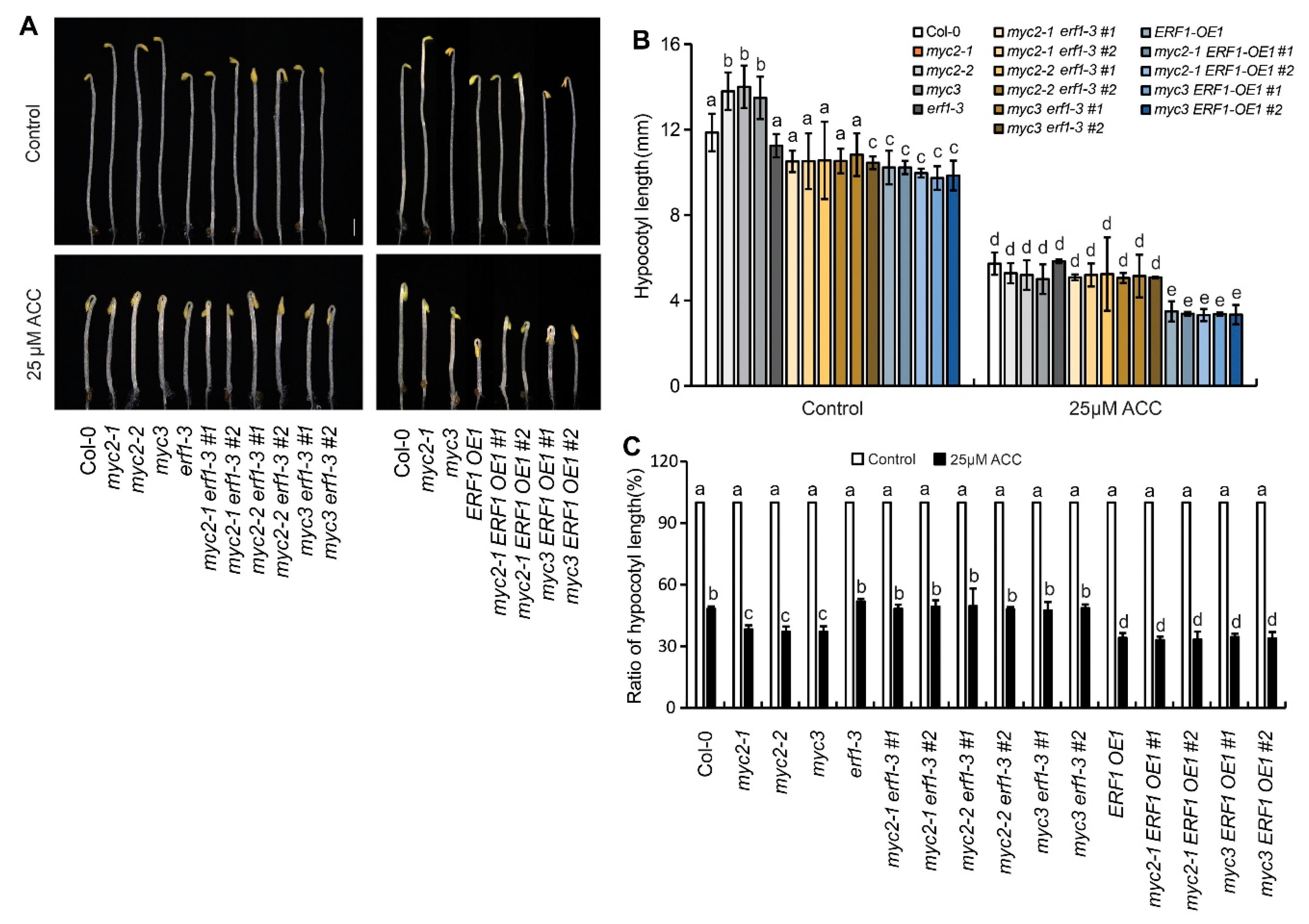
Figure 7.
MYC2 and MYC3 are involved in ethylene-regulated expression of functional genes. Etiolated seedlings were grown on a medium containing 25 μM ACC for 4 days. High-throughput RNA-seq was used to analyze the expression pattern of functional genes. (A). Number of ethylene-regulated DEGs in Col-0 and myc2myc3. (B). Venn analysis of DEGs. (C). GO (gene ontology) analysis of DEGs.
Figure 7.
MYC2 and MYC3 are involved in ethylene-regulated expression of functional genes. Etiolated seedlings were grown on a medium containing 25 μM ACC for 4 days. High-throughput RNA-seq was used to analyze the expression pattern of functional genes. (A). Number of ethylene-regulated DEGs in Col-0 and myc2myc3. (B). Venn analysis of DEGs. (C). GO (gene ontology) analysis of DEGs.
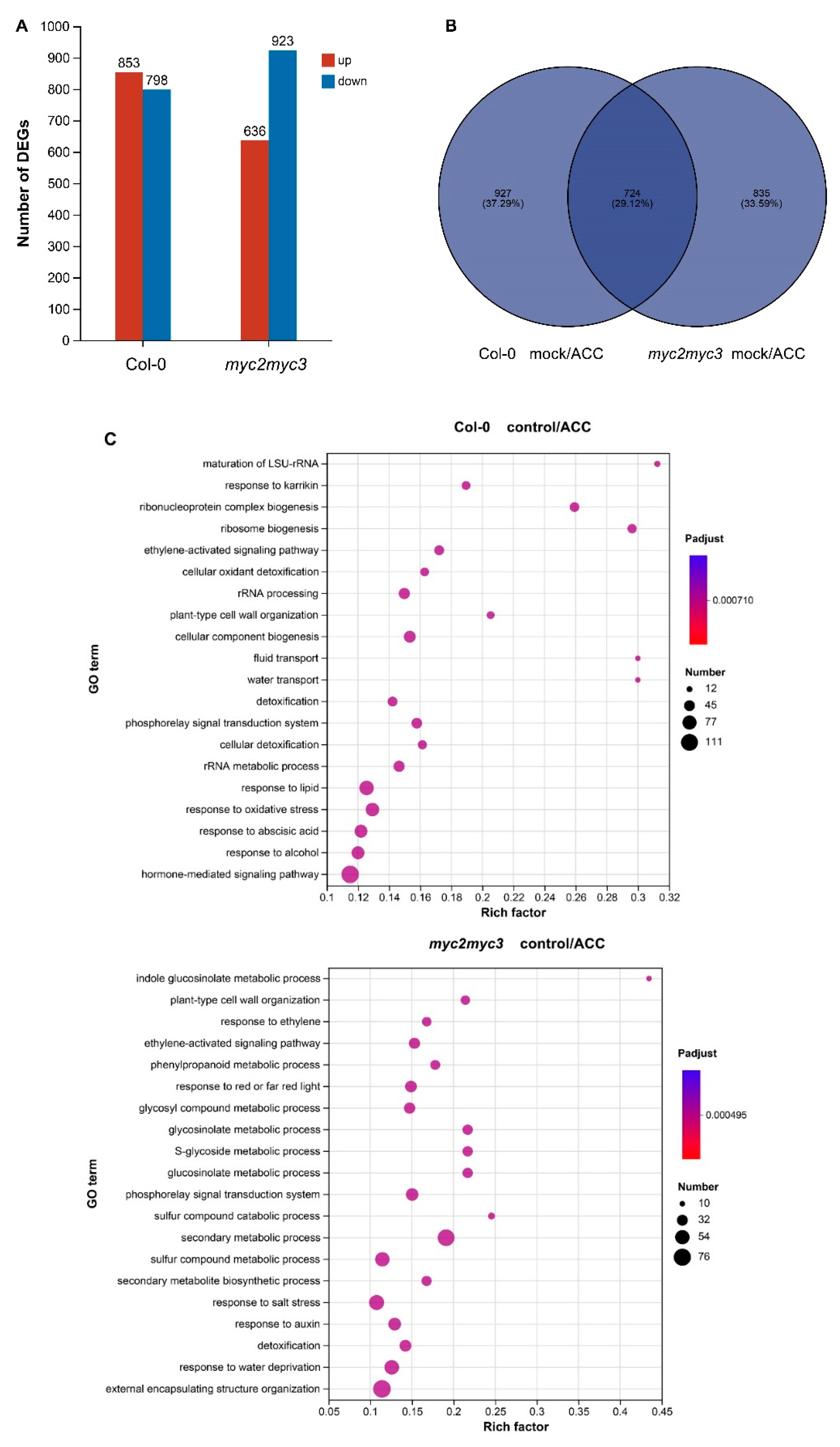
Figure 8.
Model of the function of MYC2 or MYC3 in ethylene-inhibited elongation of etiolated hypocotyls. Arrows and bars indicate positive and negative regulation, respectively.
Figure 8.
Model of the function of MYC2 or MYC3 in ethylene-inhibited elongation of etiolated hypocotyls. Arrows and bars indicate positive and negative regulation, respectively.
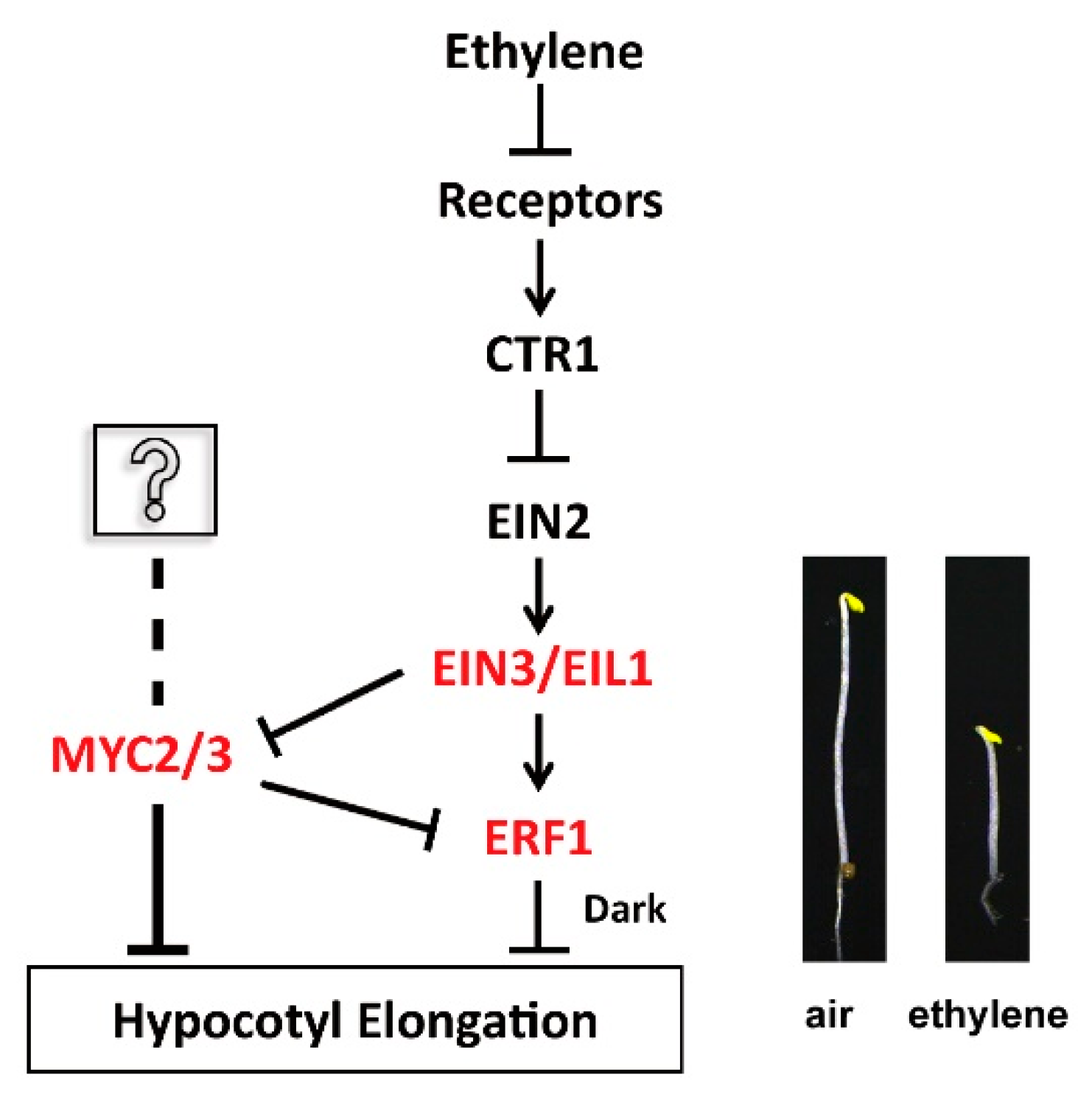

Table 1.
DEGs which were up-regulated in Col-0 while not up-regulated in myc2myc3 after ACC treatment.
Table 1.
DEGs which were up-regulated in Col-0 while not up-regulated in myc2myc3 after ACC treatment.
| Functional category | Gene_id | Gene name | Gene description |
|---|---|---|---|
| cell wall organization | AT1G54970 | PRP1 | involved in cell wall structure formation, inducible by ethylene or auxin |
| AT1G26250 | proline-rich extensin-like protein | ||
| AT1G02460 | pectin lyase-like superfamily protein | ||
| AT1G62980 | EXPA18 | Expansin-A18, involved in cell wall loosening | |
| growth & development | AT5G49870 | JAL48 | involved in plant growth and development |
| AT1G18630 | RBG6 | involved in regulation of growth & development | |
| hormone metabolism or signaling | AT1G04180 | YUC9 | involved in auxin synthesis |
| AT4G00880 | SAUR-like auxin-responsive protein | ||
| AT5G25350 | EBF2 | EIN3-binding F-box protein 2, involved in ethylene signaling | |
| oxidation reduction | AT3G49960 | PER35 | Peroxidase 35 |
| AT1G69880 | TRX8 | Thioredoxin H8, involved in cell redox homeostasis | |
| AT5G25180 | CYP71B14 | Cytochrome P450 family protein | |
| AT4G20240 | CYP71A27 | Cytochrome P450 family protein | |
| AT4G26010 | a peroxidase | ||
| signal transduction | AT4G04220 | AtRLP46 | Receptor-like protein 46 |
| AT3G23120 | AtRLP38 | Receptor-like protein 38 | |
| AT1G03010 | a NPH3 family protein, involved in response to light stimulus | ||
| stress response | AT2G05230 | DNA-J heat shock protein | |
| AT1G53130 | GRI | involved in extracellular ROS-induced cell death | |
| AT4G09435 | involved in disease resistance | ||
| AT4G38410 | dehydrin family protein, involved in response to water stress | ||
| AT5G14200 | IMDH3 | 3-isopropylmalate dehydrogenase, involved in glucosinolate synthesis | |
| transcription factor | AT5G57520 | ZFP2 | Zinc finger protein 2 |
| AT1G03790 | SOM | Zinc finger CCCH domain-containing protein 2 | |
| AT3G62090 | PIF6 | Myc-related bHLH transcription factor | |
| AT5G17430 | BBM | AP2-domain containing protein | |
| AT3G60490 | ERF035 | Ethylene-responsive transcription factor 035 |
Table 2.
DEGs which were down-regulated in Col-0 while not down-regulated in myc2myc3 after ACC treatment.
Table 2.
DEGs which were down-regulated in Col-0 while not down-regulated in myc2myc3 after ACC treatment.
| Functional category | Gene_id | Gene name | Gene description |
|---|---|---|---|
| cell wall organization | AT3G50220 | IRX15 | involved in xylan synthesis and deposition |
| AT3G55090 | ABCG16 | required for synthesis of cell wall | |
| cellular metabolism | AT1G05675 | UGT74E1 | UDP-Glycosyltransferase superfamily protein |
| AT4G25835 | AAA-ATPase, P-loop containing NTP hydrolases | ||
| AT4G25150 | Acid phosphatase-like protein | ||
| AT1G06520 | GPAT1 | Glycerol-3-phosphate acyltransferase 1 | |
| AT1G66040 | ORTH4 | E3 ubiquitin-protein ligase | |
| AT1G06030 | fructokinase-2 | ||
| AT5G05270 | CHI3 | chalcone--flavonone isomerase 3, involved in response to karrikin | |
| AT1G65060 | 4CL3 | 4-coumarate-CoA ligase 3, involved in phenylpropanoid synthesis | |
| AT1G72520 | LOX4 | Lipoxygenase 4, involved in growth and defense response | |
| AT2G28210 | ATACA2 | alpha carbonic anhydrase 2 | |
| growth & development | AT2G29950 | EFL1 | ELF4-LIKE 1, involved in circadian and flowering |
| AT3G18217 | MIR157C | MIR157C, involved in regulating vegetative phase | |
| AT5G51720 | NEET | involved in plant development | |
| AT1G43790 | TED6 | involved in tracheary element differentiation | |
| AT2G46830 | CCA1 | involved in regulating circadian rhythms | |
| AT1G65620 | AS2 | involved in formation of a symmetric flat leaf lamina | |
| AT1G52690 | LEA7 | Late embryogenesis abundant protein 7 | |
| hormone metabolism or signaling | AT2G14960 | GH3.1 | IAA-amido synthetase |
| AT2G16660 | involved in response to karrikin | ||
| AT5G66260 | SAUR-like auxin-responsive protein | ||
| oxidation reduction | AT3G49110 | PER33 | Peroxidase 33 |
| AT1G32780 | Alcohol dehydrogenase-like 3 | ||
| AT3G44970 | Cytochrome P450 family protein | ||
| AT2G30540 | GRXS9 | Monothiol glutaredoxin-S9 | |
| AT5G25130 | CYP71B12 | Cytochrome P450 family protein | |
| signal transduction | AT3G15050 | IQD10 | a calmodulin binding protein |
| AT5G15430 | a calmodulin-binding protein-like protein | ||
| AT3G01830 | CML40 | Calmodulin-like protein 40 | |
| AT1G21550 | CML44 | Calmodulin-like protein 44 | |
| AT2G24850 | TAT3 | a tyrosine aminotransferase that is responsive to jasmonic acid | |
| AT5G43300 | GDPD3 | PLC-like phosphodiesterase | |
| AT3G45430 | LECRK15 | L-type lectin-domain containing receptor kinase I.5 | |
| AT3G45390 | LECRK12 | L-type lectin-domain containing receptor kinase I.2 | |
| AT5G55830 | LECRKS7 | L-type lectin-domain containing receptor kinase S.7 | |
| stress response | AT1G12280 | SUMM2 | a disease resistance protein, involved in defense response |
| AT3G59930 | Defensin-like protein 206 | ||
| AT2G03020 | Heat shock protein HSP20 | ||
| transcription factor | AT5G61620 | myb-like transcription factor family protein | |
| AT1G73870 | COL7 | zinc finger protein CONSTANS-LIKE 7 | |
| AT1G62700 | NAC026 | NAC (No apical meristem) domain transcriptional regulator | |
| AT5G64750 | ABR1 | ethylene-responsive transcription factor | |
| AT1G22810 | ERF019 | ethylene-responsive transcription factor |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



