
Submitted:
11 June 2024
Posted:
11 June 2024
You are already at the latest version
Abstract
Understanding pollen and ovule fertility as factors influencing fruit and seed set is important in cassava breeding. Red light (RL) and plant growth regulators (PGRs) have been used to induce or enhance flowering and fruit set in cassava without any reference to effects on pollen viability or ovule fertilizability. This study investigated effects of field-applied RL and PGR on pollen viability and ovule fertilizability. Panels of cassava genotypes, with early or moderate flowering responses were used. RL was administered from dusk to dawn. Two PGRs, a cytokinin (6-benzyl adenine, BA) and an anti‑ethylene (silver thiosulphate, STS) were applied. Pollen viability was assessed basing on pollen grain diameter, in vitro stainability, in vivo germinability, ovule fertilizability and ploidy level. Treating flowers with RL led to increased pollen diameter from 143.6 to 148.5µm, stainability from 82.9 to 93.0%, ovule fertilizability from 52.0 to 56.9% in treated female flowers and 47.7 to 64.3% on using pollen from treated male flowers. Fruit set increased from 51.5 to 71.8% in RL-treated female flowers and from 53.7 to 71.1% when pollen from RL-treated male flowers was used. Seed set followed a trend similar to that of fruit set. Ploidy level of pollen from RL-treated flowers increased slightly and positively correlated with pollen diameter (R2 = 0.09*), ovule fertilization (R2 = 0.20*), fruit (R2 = 0.59*) and seed set (R2 = 0.60*); but negatively correlated with pollen stainability. Treating flowers with PGR did not affect pollen diameter and stainability, but increased ovule fertilizability from 42.9 to 64.9% and fruit set from 23.2 to 51.9% in treated female flowers and 31.3 to 54.7% when pollen from treated male flowers was used. Application of combined BA+STS on male flowers caused the highest ovule fertilizability, fruit and seed set efficiency, though this varied among study genotypes. These results show that RL and PGR treatment of flowers increases pollen viability and ovule fertilizability. This is important for planning pollination strategies in cassava breeding programmes.
Keywords:
pollen viability
; in vitro stainability
; in vivo germinability
; ovule fertilizability
; pollen diameter
; pollen ploidy
1. Introduction
Cassava (Manihot esculenta Crantz) is a staple crop with tuberous roots, which are rich in starch, making it important as food for humans and animals as well as raw material for different industries [1,2,3]. It is a crop resilient to unpredictable climatic fluctuations as well as poor soils [4,5]. Thus, it is considered a potential for bridging the food demand gap and a drought-tolerant crop in the forecasted warmer 2030 world. This necessitates that, breeding efforts need to be prioritized towards improvement of important traits such as yield- and nutrient-related traits as well as tolerance to both abiotic and biotic constraints. However, in order to achieve these breeding targets, deep understanding of pollen and ovule fertility as factors influencing fruit and seed set, is needed.
One of the impediments to genetic improvement of cassava is low or no seed set due to high abortion rates of flowers or fruits [6]. High abortions and low fruit or seed set rates, with averagely less than one seed per pollination, in controlled crosses have been reported in cassava crossing programmes [7]. Abortion of a large proportion of flowers and fruits is generally an enormous challenge among a diverse of plant species, with some species producing barely one fruit for every 1000 or more flowers produced [8].
Among key factors underpinning fruit abortions in many plant species include parental genetic differences, unsuccessful pollinations due to poor quality and/or unviable pollen, low ovule fertility rates as well as inadequate or unsynchronized or overproduced amounts or inhibitory effects of growth hormones [8,9]. In conventional plant breeding, successful pollination is a prerequisite for fertilization and seed set [10,11], and this highly depends on good quality and viable pollen as well as stigma receptivity [12]. Thus, insight knowledge on pollen viability enhancement or identification of viable pollen, its germination ability and capacity to fertilize ovules, is essential for increasing fruit and seed set in crop breeding programs [13]. However, knowledge on the factors influencing pollen viability and success or failure of fruit and seed set in cassava is still scanty. It is worth noting that, manipulation of male fertility has proved a useful trait for breeding and increased crop yield [14,15].
Viable pollen is important for species’ survival in the next plant generation and targeted breeding as well as crop improvement [13]. Pollen viability, its functionality or germination and capacity to fertilize ovules depend on various factors, including enzyme-activity, cytoplasmic contents, nutrition status, plant growth regulators, varietal differences, size or ploidy level and environmental factors, including photoperiodism [16,17,18]. The role of plant growth regulators (PGR) and photoperiodism in plant physiology have been extensively studied. However, there are only a few studies on their effects on pollen viability. For example, PGRs like brassinosteroid, have been used to increase pollen viability in pomegranate flowers [16] and Tulipa greigii [19].
On the other hand, application of red and blue lights (ratio of 1:1) were used to increase pollen viability in tomatoes [20] as blue and yellow lights promoted pollen germination in Peltophorum [21]. In cassava, field applications of RL photoperiod extension and PGRs (BA and STS), have been reported to enhance fruit and seed set [22] but there was no any effort made to explore their possible effects on pollen viability and ovule fertilizability. And yet, pollen viability may be an important factor and selection criterion for cassava breeders.
Additionally, pollen grain viability was found to strongly and positively correlate with pollen grain size in strawberry, Fragaria × ananassa Duch [23] and Mimulus guttatus [24]. Relatedly, in Lantana camara, pollen with lower ploidy level (diploids) was found to exhibit higher viability compared to that with higher ploidy levels (tetraploids, pentaploids and hexaploids) [25]. Relatedly, high ploidy level amphiploid pollen grains in oats showed reduced viability compared with their parental species with lower ploidy levels [26].
Pollen viability can be estimated using a variety of methods including, in vivo and in vitro germination as well as the staining techniques [17]. Another, more accurate method involves pollen deposition on stigma followed by evaluation of pollen tube growth and seed set [10,17]. In vitro techniques are mostly used because they are efficient, fast and easy to assess. The chemical staining method is more advantageous as indicator of pollen viability because it is faster and easier compared with pollen germination [10]. A more popular staining technique for assessing pollen viability involves use of Alexander’s stain (AS), which excellently differentiates between aborted and non-aborted pollen [27]. Because different methods give different levels of accuracy in pollen viability evaluations, it is recommended that several test methods be used simultaneously. In this study, a combination of in vivo pollen germination and ovule fertilizability, stainability using AS, pollen grain diameter measurement and ploidy level estimation were used to assess viability of cassava pollen.
The question whether pollen viability and its capacity to fertilize ovules is influenced by treating flowers with red light (RL) and plant growth regulators (PGRs) was the driving force behind this study. Thus, we aimed to assess pollen viability under these treatments using the in vivo pollen germination and seed set, pollen size measurement, and AS staining techniques as well as estimation of ploidy level using flow cytometry.
2. Results
2.1. Effects of RL and PGR Treatments on Pollen Characteristics and Its Capacity to Cause Fruit- and Seed Set
Pollen characteristics including pollen grain diameter and viability, measured as pollen stainability and capacity to fertilize ovules to cause fruit and seed set, were assessed on pollen samples picked from flowers of cassava plants subjected to RL and PGR treatments. Though there was no notable effect on the pollen diameter and its stainability, the treatment effect was significant in the capacity of pollen to fertilize ovules and cause fruit and seed set (Table 1). Treating flowers by exposing them to RL or PGRs increased the capacity of the pollen to fertilize ovules (Figure 1) resulting in increased fruit- and seed set efficiencies compared to the non-treated ones. Generally, the RL treatment was more effective, in influencing pollen diameter, stainability and capacity to cause fruit and seed set, except ovule fertilization.
2.2. Effect of RL and PGR Treatments on Pollen Stainability and Diameter
There was no statistical difference in the effects of PGR treatment on the pollen grain diameter and stainability compared to control (Table 2). However, diameter (148.5µm) and stainability (93.0%) were significantly higher in flowers treated with RL compared to the non-treated ones (control), indicating that RL exposure may be involved in increasing the viability of pollen in cassava. Actually, pollen stainability ranged from 0% in non-treated flowers to 100% in flowers treated with RL.
Pollen grains stained were more in flowers treated with RL (Figure 2A1) compared to PGR- and non-treated flowers (Figure 2B1 and C1). Pollen grains that stained reddish purple were considered viable while those stained pale green were considered non-viable. Additionally, some grains stained dark purple, implying higher viability, while others stained brightly red, implying a relatively lower viability. Pollen stainability was positively correlated with pollen diameter, in both flowers subjected to treatment or not, indicating that pollen viability is dependent on pollen size. The correlation was stronger (R = 0.61) in pollen from non-treated flowers (Figure 2C2) followed by that from PGR-treated flowers (R = 0.52) (Figure 2A2) and was least in the RL-treated flowers (Figure 2B2). Overall, for all treatments, the correlation was R = 0.52.
2.3. Effect of Exposing Flowers to RL on Ovule Fertilizability, Fruit- and Seed-Set Efficiency
Exposing flowers to red light treatment significantly increased the capability of pollen to fertilize ovules leading to enhanced fruit and seed set compared to control (Table 3). Efficiency of ovule fertilization as well as fruit and seed set did not vary significantly when either female or male flowers were treated with red light. However, the efficiencies varied significantly among the genotypes used in this study (Table 4).
In genotype UG15F180P005, treating the male flowers resulted in more ovule fertilization (53 vs 25.4%), fruit- (59.5 vs 53.0%) and seed set (39.4 vs 30.1%) efficiencies compared to when female flowers were treated. Meanwhile, in UG15F302P016, ovule fertilization (62.2 vs 56.9%), fruit- (64.8 vs 60.1%) and seed set 40.5 vs 38.4%) were slightly more efficient when female flowers were treated. Overall, subjecting both male and female flowers to red light treatment significantly increased pollen viability and thus increased ovule fertilization, fruit and seed set efficiency.
2.4. Effect of Treating Flowers with PGR on Ovule Fertilizability, Fruit- and Seed-Set Efficiency
Treating flowers with PGRs significantly increased the efficiencies of ovule fertilization, fruit and seed set compared to control, except ovules fertilized when pollination was done using treated male flowers (Table 5). With exception of ovules fertilized, the efficiencies of fruit and seed set more than doubled under PGR treatment. Whereas treating female flowers with BA resulted in the highest mean number of ovules fertilized, the same treatment on male flowers caused the lowest number of ovules fertilized compared to control (Figure 3). Meanwhile, a combination of BA+STS caused the highest fruit and seed set efficiency.
Efficiency of ovule fertilization, fruit and seed set varied among the study genotypes. In genotype UG15F192P012, the efficiencies were consistently higher in PGR-treated female flowers than the treated male flowers (Table 6). While in genotype UG15F302P016, the efficiencies were higher when pollen from PGR-treated flowers was used, except for ovules fertilized.
2.5. Comparison of Effects of RL and PGR Treatments on Pollen Viability, Fruit and Seed Set Efficiency
Exposing flowers to either RL or PGR treatments was more efficient in increasing ovule fertilization, fruit and seed set compared to non-treated flowers (Figure 4). Treating both female and male flowers with RL was most efficient, except in ovules fertilized, where treating the female flowers with PGR was more efficient (Figure 4A). According to these results, efficiency of ovule fertilization, fruit and seed set is higher when flowers are exposed to RL.
2.6. Pollen Ploidy and Its Relationship with Diameter and Viability in RL-Treated Flowers
Pollen from flowers exposed to red light treatment exhibited a slight increase in ploidy level compared to control (Figure 5A). Spearman’s correlation coefficients matrix was calculated for ploidy, pollen characteristics as well as fruit and seed set efficiencies. Pollen ploidy level was positively correlated with pollen diameter (R2 = 0.09*), ovule fertilization (R2 = 0.20*), fruit (R2 = 0.59*) and seed set (R2 = 0.60*) efficiency; but negatively correlated with pollen stainability (R2 = -0.37*) (Figure 5B).
Variation in amounts of nuclear DNA and ploidy levels in pollen samples used in the flow cytometric analyses in this study is shown in Figure 6. The analyses showed that the DNA content in pollen grains ranged from 2C (diploid) amount to polyploid amounts of 4C (doubled), 6C and 8C (more-than doubled), indicating increased ploidy levels due to endoreduplication of DNA. In determining the ploidy levels of the pollen samples, ploidy histograms were plotted and compared with ploidy level of young cassava leaves (Figure 7). As shown in Figure 7A, the young leaf samples displayed one major 2C peak, which clearly indicated a diploid state, and one minor 4C peak. The minor 4C peak being most likely due to nuclei in the G2 stage of the cell cycle. Comparatively, the histograms of the pollen samples predominantly displayed one main ploidy peak within the 2C DNA or diploid region just as the control (Figure 7B-D) corresponding to the G1 (DNA doubling) phase of the cell cycle. However, the peaks corresponded to a DNA content greater than 200, a channel mean set to correspond to 2n DNA content of a diploid state. This shows that the pollen exhibited a slightly higher ploidy level. Taken together, the results indicate that the pollen grains picked at anthesis in this study contained diploid nuclei but had portions of higher ploidy nuclei.
3. Discussion
3.1. Effect of RL on Pollen Diameter, Stainability and Ovule Fertilizability
The results of this study indicate that RL treatment exerted an effect on pollen viability, by increasing pollen grain diameter, stainability and capacity to fertilize ovules to cause fruit and seed set. The involvement of RL in the flowering process of crop plants has been extensively studied. It is involved in the regulation of various stages of plant development, including pollen development [28,29]. However, there are limited studies on the effect of RL on pollen viability or stainability in plants including cassava.
In a previous study, RL increased pollen viability measured as in vitro pollen germination and pollen tube growth in Pinus roxburghii [30], while a mixture of red and blue light increased pollen viability in tomato [20]. These results closely agree with the findings of the current study in which pollen viability measured as pollen stainability and ovule fertilizability were higher in flowers exposed to RL treatment. The physiological effect of RL in plants is linked to phytochromes, the light-sensitive pigments, existing in two interconvertible forms: Pr (an inactive, RL-absorbing) and Pfr (an active, far-red light-absorbing) [31,32]. These interconversions have correlative photomorphogenic effects that result in flower formation [28]. The Pfr, interacts with other proteins to trigger light-dependent photomorphogenic flowering responses [33,34]. It initiates signal transduction pathways within plant cells, which then influence gene expression and various physiological processes associated with floral initiation and development including pollen development and viability [29]. Thus, the results in this study suggest that pollen stainability, in vivo germination and ovule fertilization may also be phytochrome-controlled, and this is perhaps, being reported for the first time. The observed variation in ovule fertilizability among genotypes could be attributed to physiological and/or genotypic differences in genes that are involved in photoperiod and hormone systems that are involved in regulating flowering.
Pollen diameter was greater in RL-treated flowers compared to control (. Though not directly comparable, positive correlation between pollen grain diameter and pollen viability was reported in sunflower [35] and Mimulus guttatus [24]. The larger pollen size could be attributed to evolutionary selection pressure imposed by pollinating honeybees which prefer large-sized pollen grains [35,36], unreduced gametes [24] as well as a high protein content, mainly enzymes that function during pollen germination and subsequent fertilization [36]. Since exposure to RL influences Pr and Pfr interconversions culminating in protein synthesis, it can be hypothesized that the increase in pollen diameter observed in this study was due to increased protein synthesis in pollen of flowers exposed to RL treatment.
3.2. Effect of PGR on Pollen Diameter, Stainability and Ovule Fertilizability
In this study, the effect of two PGRs, BA and STS, on pollen diameter, stainability and ovule fertilizability were investigated. The results indicate that there was no significant difference in pollen grain diameter and stainability between pollen of PGR-treated and the non-treated flowers. However, PGR treatment increased capability of the pollen to fertilize ovules leading to increased fruit and seed set. The involvement of plant growth regulators (PGR) in modulating various morphogenetic processes, including flowering has been reported [37,38]. However, the effect of PGR, especially BA and STS, on pollen viability or stainability and capability of pollen to cause ovule fertilization in plants has not been widely explored, and besides literature about such is scanty. Whereas BA was reported to induce flowering or feminization of flowers, STS increased abundance and prevented abortion of flowers, thus enhancing fruit and seed set in cassava [22,39,40]. However, the effects of these PGRs on pollen viability and ovule fertilizability in cassava were not analysed.
PGRs such as brassinosteroids were used to increase pollen viability and in vitro germination in male flowers of pomegranate [16], while gibberellins were reported to be essential for pollen germination and pollen tube elongation in rice [41]. PGRs, especially cytokinins, promote protein synthesis and nutrient mobilization, cell division and differentiation leading to ovule and pollen development and viability, including pollen germination [42,43,44]. Application of BA, a cytokinin, slightly improved flowering and pollen viability in shallots (Allium cepa var. ascalonicum) [45]. On the contrary, in this study, a decrease in pollen viability and increased ovule fertilizability were observed when female and male flowers were treated with BA only compared to control. However, fruit and seed set increased compared to control.
Application of STS increased pollen quality in male flowers [46], and contrary to the findings of this study, produced large-diameter pollen grains in both male and masculinized female genotypes of Cannabis sativa L. [47]. Relatedly, STS induced production of male flowers with viable pollen grains, able to germinate on stigmas to effect seed set in genetically female Cannabis sativa plants [48,49]. All this can be explained by the fact that STS is known to prevent ethylene-induced senescence and to inhibit ethylene production by outcompeting copper ion cofactors in the ethylene receptors, allowing CONSTITUTIVE TRIPLE RESPONSE1 (CTR1) to be active and signaling a slowed ethylene response [50]. This in turn initiates male flower induction and maintains overall plant health and viability of flowers, including their pollen [51]. Similarly, the results of this study show increased pollen viability, exhibited through increased ovule fertilizability when both male and female flowers were treated with STS and the consequent increase in fruit and seed set. These results confirm effects of STS on pollen viability, thus corroborating the results of previous studies [46,49].
In the current study, much more promotive effects on the pollen’s capability to fertilize ovules and fruit and seed set were observed when BA and STS were applied in combination, on the male and female flowers, than when these growth regulators were applied singly. This suggests that BA and STS had additive and/or synergistic or interactive effects which increased pollen viability and ovule fertilizability. However, the variation in ovule fertilizability among genotypes could be attributed to physiological and/or genotypic differences in genes that are involved in photoperiod and hormone systems that regulate flowering.
3.3. Relationship between Ploidy, Diameter and Viability of Pollen in RL-Treated Flowers
Red light regulates developmental processes throughout plant life, including flower development [29,52]. Effect of RL on pollen ploidy has not been previously reported. However, in a previous study, extended photoperiod improved pollen quality in hexaploids better than in diploid and tetraploids and promoted formation of apomictic seeds in Ranunculus auricomus complex (Ranunculaceae) [53]. In this study, pollen of flowers exposed to red light treatment exhibited a slightly increased ploidy level. Moreover, DNA content in pollen grains of treated flowers ranged from 2C (diploid) amount to polyploid amounts of 4C (tetraploid), 6C (hexaploidy) and 8C (octaploid), indicating an increased amount of cellular DNA and the ploidy peaks stabilized slightly beyond the control channel mean of 200. This suggests that red light may have played a role in stimulating increased DNA amounts. The increase in the DNA content indicates cell somatic polyploidization, which could be attributed to either endomitosis or endoreduplication. Endoreduplication, a more common mode of cell polyploidization in plants, involves one or several rounds of nuclear DNA synthesis in the absence of mitosis [54,55]. It occurs during biological processes, including cell metabolism, differentiation and cell expansion. Endoduplication is presumably important for increasing availability of DNA templates for gene expression [56].
Pollen size is often positively correlated with ploidy [57], and thus, is often used as a biological parameter to estimate the ploidy and viability of pollen [58]. It was previously reported that there is a tendency for polyploid pollen grains to be larger than diploid pollen grains [59,60]. In Camellia oleifera, pollen size was found to progressively increase with ploidy level, from diploids to octaploids while pollen viability as well as germination rate were relatively high in hexaploids than in pollen with lower ploidy levels [58]. Similarly, in this study, pollen ploidy correlated positively with pollen diameter (Figure 5B). This is consistent with the results of a study in which it was found that pollen grain size of octoploids was larger, thus suggesting that pollen size is influenced by ploidy levels [61]. Very little is known about the causal relationship between ploidy level and pollen size. In theory, polyploidy causes increase in amount of DNA and a consequent increase in size of a cell [62]. Thus, the increased pollen diameter in relation to ploidy observed in this study could be due to increased DNA amount or other genetic factors that may also play a significant role in determining pollen size.
Studies have shown that ploidy level affects pollen viability and pollen tube growth [59,63], with viability being higher in higher ploidy pollens [60]. In this study, pollen ploidy correlated positively with ovule fertilization, but negatively with pollen stainability. Partly in agreement with the findings of this study, is that pollen stainability was higher in lower ploidy pollen compared to higher ploidy pollen in Hydrangea macrophylla [63] and Lantana camara [25], indicating a positive correlation. Conversely, germination rate was higher in the high ploidy, hexaploid pollen than in the lower ploidy, tetraploid and diploid pollens in Camellia oleifera, indicating a negative correlation [58]. Although, it has been reported that low ploidy levels confer lower pollen viability or stainability [60], in this study, it was not possible to establish a distinct relationship between ploidy level and the pollen viability or stainability of pollen and its capacity to fertilize ovules. Thus, from the findings of this study, it is not possible to clearly relate pollen ploidy level with pollen viability.
4. Materials and Methods
4.1. Plant Materials and Treatments
This experiment was conducted using plant materials established under field conditions at the National Crops Resources Research Institute (NaCRRI), Namulonge. The selection included profusely or early to moderately flowering cassava genotypes. Three sets of plant materials (each consisting 10 genotypes), were subjected to red light (RL-), plant growth regulators (PGR-) and non-treatment (Non). The RL treatment was administered as described in [22]. In this case the study genotypes were subjected to a RL system setup using 50W light emitting diode (LED) lamps with red LEDs (model 5-10x5w, China) (illumination range 640-660nm) and with reflectors (339 × 350mm, model ISL-RFGB, CCS Inc., China) as sources of RL for extension of the photoperiod during the night. The lamps were placed horizontally at 3m above the ground in the center of a plot to cast red light over the plants up to a radius of 6m. Plants beyond this radius were regarded as non-treated plants, and thus, served as control. The illumination was administered daily from dusk to dawn until the plants reached 2nd to 4th level of flowering.
The PGR-treated plants were established in a randomized split-plot design in which each variety was represented by 20 plants, spaced at 1m between plants and rows, in three blocks or replications. Each main plot (genotype) was split into four plots of four plants each. Two PGRs, silver thiosulphate (STS) and 6-benzyl adenine (BA), in four treatment regimens; 1) BA only; 2) STS only; 3) combination of BA and STS and 4) control (non-treatment) were used. One treatment was administered per split plot, following a method described by [22]. Briefly, 2.5ml of 4 mM STS were applied through the petiole at a 14-day interval while 0.5 mM BA was applied by hand spray at shoot tips until just wet, at a 7-day interval. The treatments were commenced at the earliest notice of forking in any one genotype, and this was routinely continued up to the fourth tier/level of branching, 5 to 8 months after planting, as this varied with genotype.
4.2. Pollen Collection
The pollen was picked in batches from each of the RL-, PGR- and non-treated plants and each batch was assessed for viability using techniques of in-vivo germinability and capacity to set fruit and seed, in-vitro staining as well as measurement of grain diameter and ploidy level. The pollen was sampled from forking/branching levels between one and four. In each case, mature and ripe male flower buds (anthers) were hand-picked in the mornings before midday. To allow for replication, batches of pollen samples were picked at least two times from a particular flowering level.
4.3. Assessment of Pollen Germination, Stainability and Measurement of Grain Diameter
4.3.1. Pollination
To assess the ability of RL-, PGR-, and non- treated pollen to germinate on stigmas and its capacity to effect seed set (ovule fertilizability), self-, out- and hybridization crosses were done through manual pollinations. In this case, mature female flowers under similar treatments were bagged using muslin bags for 1 to 3 days prior to anthesis to avoid contamination by pollen from unknown sources. Pollen samples collected in section 2.2 from same and different genotypes were used to pollinate fully opened flowers (at anthesis) by gently brushing the opened anthers on the stigma. All the pollinated flowers were re-bagged for at least three days to keep away stray pollen.
4.3.2. In Vivo Pollen Germination and Ovule Fertilization
To determine the germinability of the pollen and ovule fertilization, about three hand-pollinated flowers were picked at two-to-three days after pollination (DAP), fixated in a solution containing glacial acetic acid and 96% ethanol (in a ratio of 1:3) and then stained with 0.1% aniline blue in 0.1% K2HPO4 before being examined for pollen tube growth as previously described in [64]. The pistils were removed from the stain, placed in a drop of glycerol on a glass slide, stigmas cut off and the ovaries dissected to extract ovules. Ovary wall tissues were discarded, a drop of basic 0.1% aniline blue solution added on the ovules, covered with a cover slip and then gently squashed. Observations of pollen germination and pollen tube growth were made with a fluorescence microscope (Nikon Alphaphot-2 YS2, Japan) and images taken using a camera head (Nikon DS-L3, Japan).
4.3.3. In Vitro Pollen Stainability
In the in vitro viability test using the staining assay, a portion of the fresh anther buds were fixated in a Carnoy solution (containing Absolute ethanol, Chloroform and Glacial acetic acid in ratio of 6:3:1 respectively) for not less than 2 hours. Stainability was determined by using a modified solution of AS prepared according to a protocol described by Peterson et al., (2010). The final stain solution, that was kept in the dark, contained the following constituents, added in the order they are presented: 10mL 95% alcohol, 1mL Malachite green (1% solution in 95% alcohol), 50mL distilled water, 25mL glycerol, 5mL acid fuchsin (1% solution in water), 0.5mL Orange G (1% solution in water), 4 mL Glacial acetic acid and distilled water (4.5 mL) added to make a total volume of 100mL solution.
The flower buds were carefully removed from fixative solution and placed on absorbent paper to dry off excess solution. Two-to-three buds were then placed on the glass slide, dissected under a dissecting microscope to release the pollen and the leftover plant debris carefully removed. Two-to-four drops of the AS stain were added before the sample completely dried, and the slide was slowly heated over a spirit burner in a fume hood until the stain solution was near boiling (~10 seconds). This was to enable better penetration of the dye into the cellulose and protoplasm of the pollen. The sample was left to stand for 10 to 15 minutes after heating to allow stain to be completely absorbed into the pollen grains. A cover-slip was placed over the sample and even pressure applied on the cover-slip to ensure that pollen converged to one plane. The slides were then examined using a microscope (Nikon, Model C - PS 1092883, Japan) and images taken using a camera head (Nikon DS-L3, Japan). Pollen grains that stained red to purple were regarded as viable while those that stained green were non-viable. Viable and non-viable pollen were manually quantified by counting differentially stained pollen in three views in three replications (slides). Percentage (%) pollen stainability per view was calculated as follows:
4.3.4. Estimation of Pollen Grain Diameter
To estimate pollen size, the temporary slides prepared for the in vitro stainability tests above were first used to measure grain diameter, because fixation did not affect grain size when compared with the fresh pollen (data not shown). The choice to measure diameter was based on the fact that cassava pollen is basically spherical in shape. In this case, 20 pollen grains were randomly sampled per slide and their diameters measured under an ocular microscope (Optika, SZM-1LED, Italy) using a calibrated eyepiece lens at a magnification of 10x. For purposes of replication, sampling and measurement were done three times per slide.
4.4. Assessment of Fruit and Seed Set Efficiency
To determine fruit and seed set following controlled pollination, the pollinated flowers were observed and monitored for fruit set until harvest. At two-to-three months after pollination, the fruits were harvested to determine seed set. Fruit (FE) and seed efficiencies (SE) were calculated in percentage as follows:
4.5. Determination of Pollen Ploidy Level
Pollen samples picked in section 2.2 were used to measure ploidy levels. Since the samples were collected in small bits, they were dried in silica gel and kept until a sizeable amount for analysis was obtained. The dry samples were kept (on silica gel) at room temperature for not more four months. Flow cytometry method was used to determine the amount of DNA and ploidy level of the preserved pollen following a modification of the method described by [65]. In brief, two-to-five male flower buds with dehisced anthers were placed in a 2mL Eppendorf tube, 1mL of cold Otto1 buffer (0.1M citric acid monohydrate and 0.5% v/v of Tween-20) added, allowed to soak for five minutes, and then vortexed slightly for a few seconds to shed off the pollen. The pollen suspension was placed on a clean glass petri dish and the pollen grains gently crushed against the glass for 10–15 s, using a glass rod with a flattened end. Then the homogenate was filtered through a 50µm nylon filter into a cuvette. The samples were incubated for about 20-60 minutes before 1ml of Otto II buffer (0.4M anhydrous Na2HPO4, 4µg/ml of DAPI (4, 6’-diamidino-2-phenylindole), and 1µl/ml β-metcaptoethanol) were added and ran according to standard plant flow cytometry protocols using Sysmex Partec Ploidy Analyser Machine (the CyFlow® ploidy analyser, Software- CyViewTM 1.6, REF-CY-S-3039, Germany). For control purposes, young leaves from one test plant were used. To extract nuclei and DNA, the leaves were chopped with a sharp razor blade in a cold buffer and then taken through the procedure for ploidy determination as described above. The G1 (2n DNA content) peak of the control was set at channel 200. The ploidy level of the pollen sample was determined by comparing the relative position of its G1 peak and that of the control. Data analyses were controlled and performed within the ploidy analyser by the CyView™ software (CyViewTM 1.6).
4.6. Statistical Analysis
All statistical analyses were done using statistical packages in R (v.4.2.2) [66]. A generalized linear mixed-effects model (GLMM) was used to investigate determinants on the number of fertilized ovules, stained pollen grains, fruit and seed set assuming Poisson distribution. Data on pollen diameter were assumed continuous and thus, were analysed using one-way analysis of variance (ANOVA). The graphics were prepared using Grammar of Graphics (ggplot2) package. Spearman coefficient of rank correlation was used to examine relationships among traits. Where applicable, the means were compared using TukeyHSD grouping test with a 5% probability. Percentage pollen stainability, fruit and seed set efficiencies were computed using the expressions shown in sections 4.3.3 and 4.3.4 respectively.
5. Conclusions
Enhancing pollen viability boosts crossing programmes in cassava breeding. Application of RL and PGRs (BA and STS) improved pollen viability, and hence fruit and seed set. Red light treatment increased pollen diameter, stainability, ovule fertilizability and ploidy level. PGR treatment did not affect pollen diameter and stainability, but increased ovule fertilizability. Further studies are necessary to clarify the influence of RL and PGR (BA and STS) on pollen viability.
Supplementary Materials
Supporting raw data information can be accessed at https://cassavabase.org/breeders/trial/6707?format=
Author Contributions
RSK and YB, fund acquisition; JKB, methodology, investigation, data collection and curation and visualization, and writing draft manuscript; RSK, supervision of fieldwork and project administration; SBM, RSK, MSO, WE, EN, TA and CO, supervision of the general progress of the study and manuscript write-up; PTH and TLS, provision of lamps for red light and protocols for RLE and PGRs application; BY, leader of Root Crops Program under which study was conducted. PI, EW and MK played a role in data analysis. All authors contributed to the article and approved the submission for publication.
Funding
This research was funded by the “Next Generation Cassava Breeding Project” through a grant by the Bill & Melinda Gates Foundation (Grant INV- 007637) and the UK’s Foreign, Commonwealth & Development Office (FCDO), managed by Cornell University, through a sub-award agreement (grant number OPP1048542) between NaCRRI and Cornell University.
Data Availability Statement
All relevant datasets generated during and/or analysed during the current study can be found in online repositories and are available from Cassavabase, https://www.cassavabase.org/breeders/trial/6707?format= a website maintained by the Next Generation Cassava Breeding Project.
Acknowledgments
We thank the Root Crops Program and the management of NaCRRI for providing office and field and laboratory space for conducting experiments or analyses, and the field staff who assisted with the field work. We thank Ms. Catherine Nasuuna of the International Institute of Tropical Agriculture (IITA Uganda) for technical assistance in ploidy analysis. This research was also part of the Ph.D. thesis of the first author at Makerere University, and he thanks his supervisors and colleagues for the invaluable advice and support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Alves, A.A.C. , Cassava botany and phyiology, In Cassava: Biology, production and utilization, R.J. Hillocks, J.M. Thresh, and A.C. Bellotti, Editors. 2002. p. 67-89.
- Fathima, A.A. , et al., Cassava (Manihot esculenta) dual use for food and bioenergy: A review. Food and Energy Security 2022, 12. [Google Scholar]
- Amelework, A.B. , et al., Adoption and promotion of resilient crops for climate risk mitigation and import substitution: A case analysis of cassava for South African agriculture. Frontiers in Sustainable Food Systems 2021, 5, 617783. [Google Scholar] [CrossRef]
- Pushpalatha, R. and B. Gangadharan, Is cassava (Manihot esculenta Crantz) a climate “smart” crop? A review in the context of bridging future food demand gap. Tropical Plant Biology 2020, 13, 201–211. [Google Scholar] [CrossRef]
- Jarvis, A. , et al., Is cassava the answer to African climate change adaptation? Tropical Plant Biology 2012, 5, 9–29. [Google Scholar] [CrossRef]
- Bandeira, E. , et al., Reproductive barriers in cassava: Factors and implications for genetic improvement. PLoS One 2021, 16, e0260576. [Google Scholar]
- Ndubuisi, N.D. , et al., Crossability and germinability potentials of some elite cassava progenitors. Journal of Plant Breeding and Crop Science 2015, 7, 61–66. [Google Scholar]
- Stephenson, A.G. , Flower and fruit abortion: Proximate causes and ultimate functions. Annual Review of Ecology and Systematics 1981, 12, 253–279. [Google Scholar] [CrossRef]
- Palupi, E.R. , et al., Importance of fruit set, fruit abortion and pollination success in teak. Canadian Journal of Forest Research 2010, 40, 2204–2214. [Google Scholar] [CrossRef]
- Rathod, V. , et al., Pollen viability and in vitro pollen germination studies in Momordica species and their intra and interspecific hybrids. International Journal of Chemical Studies 2018, 6, 32–40. [Google Scholar]
- Lankinen, A., S. A.M. Lindstrom, and T. D’Hertefeldt, Variable pollen viability and effects of pollen load size on components of seed set in cultivars and feral populations of oilseed rape. PLoS One 2018, 13, e0204407. [Google Scholar] [CrossRef]
- He, G. , et al., Pollen viability and stigma receptivity in Lilium during anthesis. Euphytica 2017, 213. [Google Scholar]
- Impe, D. , et al., Assessment of pollen viability for wheat. Front Plant Sci 2019, 10, 1588. [Google Scholar] [CrossRef] [PubMed]
- Younis, A., Y. J. Hwang, and K.B. Lim, Exploitation of induced 2n-gametes for plant breeding. Plant Cell Rep 2014, 33, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Farinati, S. , et al., Current insights and advances into plant male sterility: new precision breeding technology based on genome editing applications. Front Plant Sci 2023, 14, 1223861. [Google Scholar] [CrossRef] [PubMed]
- Gökbayrak, Z. and H. Engin, Effects of foliar-applied brassinosteroid on viability and in vitro germination of pollen collected from bisexual and functional male flowers of pomegranate. International Journal of Fruit Science 2018, 18, 226–230. [Google Scholar] [CrossRef]
- Dafni, A. and D. Firmage, Pollen viability and longevity. Plant Systematics and Evolution 2000, 222, 113–132. [Google Scholar] [CrossRef]
- Ottaviano, E. and D.L. Mulcahy, Genetics of angiosperm pollen. 1989. p. 1-64.
- Akçal, A., Z. Gokbayrak, and H. Engin, Determination of the effects of growth regulators on pollen viability and germination level of tulip. Acta Horticulturae, 2019(1242), 549-552.
- Fan, X., Y. Yang, and Z. Xu, Effects of different ratio of red and blue light on flowering and fruiting of tomato, in The 4th International Conference on Agricultural and Food Science. 2021, IOP Publishing.
- Jha, C.V., N. B. Patel, and V.A. Patel, Effects of the lights of different spectral composition on in vitro pollen germination and tube growth in Peltophorum. Life sciences Leaflets 2011, 12, 396–402. [Google Scholar]
- Baguma, J.K. , et al., Flowering and fruit-set in cassava under extended red-light photoperiod supplemented with plant-growth regulators and pruning. BMC Plant Biol 2023, 23, 335. [Google Scholar] [CrossRef] [PubMed]
- Zebrowska, J. , Factors affecting pollen grain viability in the strawberry (Fragaria x ananassa Duch.). Journal of Horticultural Science 2015, 72, 213–219. [Google Scholar] [CrossRef]
- Kelly, J.K., A. Rasch, and S. Kalisz, A method to estimate pollen viability from pollen size variation. Am J Bot 2002, 89, 1021–1023. [Google Scholar] [CrossRef]
- Czarnecki II, D.M. , et al., Ploidy levels and pollen stainability of Lantana camara cultivars and breeding lines. Hortscience 2014, 49, 1271–1276. [Google Scholar] [CrossRef]
- Tomaszewska, P. and R. Kosina, Variability in the quality of pollen grains in oat amphiploids and their parental species. Brazilian Journal of Botany 2022, 45, 987–1000. [Google Scholar] [CrossRef]
- Peterson, R., J. P. Slovin, and C. Chen, A simplified method for differential staining of aborted and non-aborted pollen grains. International Journal of Plant Biology 2010, 1. [Google Scholar] [CrossRef]
- Harshitha, H. , et al., Photoperiod manipulation in flowers and ornamentals for perpetual flowering. The Pharma Innovation Journal 2021, 10, 127–134. [Google Scholar]
- Demotes-Mainard, S. , et al., Plant responses to red and far-red lights, applications in horticulture. Environmental and Experimental Botany 2016, 121, 4–21. [Google Scholar] [CrossRef]
- Dhawan, A.K. and C.P. Malik, Effect of growth regulators and light on pollen germination and pollen tube growth in Pinus roxburghii Sarg. Ann. Bot. 1981, 47, 239–248. [Google Scholar] [CrossRef]
- Kimball, J.W. , Photoperiodism and phytochrome. Biology 2020.
- Tripathi, S. , et al., Regulation of photomorphogenic development by plant phytochromes. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Wu, F. and Y. Hanzawa, Photoperiodic control of flowering in plants, in Handbook of Plant and Crop Physiology. 2014. p. 33-62.
- Li, J. , et al., Phytochrome signaling mechanisms. Arabidopsis Book 2011, 9, e0148. [Google Scholar] [CrossRef]
- Elena, B. , Aspects of the pollen grains diameter variability and the pollen viability to some sunflower genotypes. Journal of Horticulture, Forestry and Biotechnology 2013, 17, 161–165. [Google Scholar]
- Roulston, T.a.H., J. H. Cane, and S.L. Buchmann, What governs protein content of pollen: pollinator preferences, pollen–pistil interactions, or phylogeny? Ecological Monographs 2000, 70, 617–643. [Google Scholar]
- Dias, J.P.T. , Plant growth regulators in horticulture: practices and perspectives. Biotecnología Vegetal 2019, 19, 3–14. [Google Scholar]
- Aliyu, O.M., O. O. Adeigbe, and J.A. Awopetu, Foliar application of the exogenous plant hormones at pre-blooming stage improves flowering and fruiting in cashew (Anacardium occidentale L.). Journal of Crop Science and Biotechnology 2011, 14, 143–150. [Google Scholar] [CrossRef]
- Oluwasanya, D. , et al., Flower development in cassava is feminized by cytokinin, while proliferation is stimulated by anti-ethylene and pruning: Transcriptome responses. Front. Plant Sci, 6: 12, 6662. [Google Scholar]
- Hyde, P.T. , et al., The anti-ethylene growth regulator silver thiosulfate (STS) increases flower production and longevity in cassava (Manihot esculenta Crantz). Plant Growth Regul 2020, 90, 441–453. [Google Scholar] [CrossRef] [PubMed]
- Chhun, T. , et al., Gibberellin regulates pollen viability and pollen tube growth in rice. Plant Cell 2007, 19, 3876–3888. [Google Scholar] [CrossRef] [PubMed]
- Terceros, G.C. , et al., The importance of cytokinins during reproductive development in Arabidopsis and beyond. Int J Mol Sci 2020, 21. [Google Scholar] [CrossRef]
- Emery, R.J.N. and A. Kisiala, The roles of cytokinins in plants and their response to environmental stimuli. Plants 2020, 9. [Google Scholar] [CrossRef]
- Bairwa, S. and J.S. Mishra, Effect of NAA, BA and kinetin on yield of african marigold (Tagetes erecta Linn.). International Journal of Current Microbiology and Applied Sciences 2017, 6, 1236–1241. [Google Scholar] [CrossRef]
- Rosliani, R., E. R. Palupi, and Y. Hilman, Benzyl amino purine and boron application for improving production and quality of true shallots seed (Allium cepa var. ascalonicum) in highlands. J. Hort. 2012, 22, 242–250. [Google Scholar] [CrossRef]
- Owen, L.C., D. H. Suchoff, and H. Chen, A novel method for stimulating Cannabis sativa L. male flowers from female plants. Plants 2023, 12. [Google Scholar] [CrossRef]
- DiMatteo, J., L. Kurtz, and J.D. Lubell-Brand, Pollen appearance and in vitro germination varies for five strains of female hemp masculinized using silver thiosulfate. HortScience 2020, 55, 547–549. [Google Scholar] [CrossRef]
- Ram, M.H.Y. and R. Sett, Induction of fertile male flowers in genetically female Cannabis sativa plants by silver nitrate and silver thiosulphate anionic complex. Theor. Appl. Genet 1982, 62, 369–375. [Google Scholar]
- Flajsman, M., M. Slapnik, and J. Murovec, Production of feminized seeds of high CBD Cannabis sativa L. by manipulation of sex expression and its application to breeding. Front Plant Sci 2021, 12, 718092. [Google Scholar] [CrossRef] [PubMed]
- Veen, H. , Silver thiosulphate: An experimrntal tool in plant science. Scientia Horticulturae 1983, 20, 211–224. [Google Scholar] [CrossRef]
- Li, Q. , et al., Integrated transcriptome and hormone analyses provide insights into silver thiosulfate-induced “maleness” responses in the floral sex differentiation of pumpkin (Cucurbita moschata D.). Front Genet 2022, 13, 960027. [Google Scholar] [CrossRef] [PubMed]
- Gautam, P. , et al., Red and blue light effects on morphology and flowering of Petunia × hybrida. Scientia Horticulturae, 2015, 184, 171–178. [Google Scholar] [CrossRef]
- Ulum, F.B., C. Costa Castro, and E. Horandl, Ploidy-dependent effects of light stress on the mode of reproduction in the Ranunculus auricomus Complex (Ranunculaceae). Front Plant Sci.
- Joubès, J. and C. Chevalier, Endoreduplication in higher plants, In The Plant Cell Cycle. 2000. p. 191-201.
- Traas, J. , et al., Endoreduplication and development: rule without dividing? Curr Opin Plant Biol 1998, 1, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Larkins, B.A. , et al., Investigating the hows and whys of DNA endoreduplication. J Exp Bot 2001, 52, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Qing-Hua, S.H.I., L. I.U. Ping, and L.I.U. Meng-Jun, Advances in ploidy breeding of fruit trees. Acta Horticulturae Sinica 2012, 39, 1639. [Google Scholar]
- Zhao, R. , et al., Comparative study on pollen viability of Camellia oleifera at four ploidy levels. Agronomy 2022, 12. [Google Scholar] [CrossRef]
- Jan, F., L. Schüler, and H. Behling, Trends of pollen grain size variation in C3 and C4 Poaceae species using pollen morphology for future assessment of grassland ecosystem dynamics. Grana 2014, 54, 129–145. [Google Scholar] [CrossRef]
- Rotreklová, O. and A. Krahulcová, Estimating paternal efficiency in an agamic polyploid complex: pollen stainability and variation in pollen size related to reproduction mode, ploidy level and hybridogenous origin in Pilosella (Asteraceae). Folia Geobotanica 2016, 51, 175–186. [Google Scholar] [CrossRef]
- Souza-Pérez, M. , et al., Pollen grain performance in Psidium cattleyanum (Myrtaceae): a pseudogamous polyploid species. Flora 2021, 281. [Google Scholar] [CrossRef]
- Sanders, H. , Polyploidy and pollen grain size: Is there a correlation? Graduate Review 2021, 1, 15. [Google Scholar]
- Alexander, L. , Ploidy level influences pollen tube growth and seed viability in interploidy crosses of Hydrangea macrophylla. Front Plant Sci, 2020, 11, 100. [Google Scholar] [CrossRef] [PubMed]
- Baguma, J.K. , et al., Fruit set and plant regeneration in cassava following interspecific pollination with castor bean. African Crop Science Journal 2019, 27. [Google Scholar] [CrossRef]
- Kron, P. and B.C. Husband, Using flow cytometry to estimate pollen DNA content: Improved methodology and applications. Ann Bot 2012, 110, 1067–1078. [Google Scholar] [CrossRef]
- R-Core-Team, R: A language and environment for statistical computing. R Foundation for Statistical Computing. 2021: Vienna, Austria.
Figure 1.
PGR- or RL-treated flowers, pollen germination and ovule fertilization: A) Ovule of untreated female flower x pollen of RL-treated male flower; B) Ovule of RL-treated female flower x pollen of untreated male flower; C) Ovule of untreated female flower x pollen of PGR-treated male flower; D) PGR-treated female flower x pollen from untreated male flower; E) pollen penetration into embryo sac (ES) and fertilization; F) Ovule of untreated female flower x pollen of untreated male flower. NB – Nucellar beak; NU – Nucellus; red arrows point at pollen tubes; and yellow circle surrounds an embryo sac region within the nucellus.
Figure 1.
PGR- or RL-treated flowers, pollen germination and ovule fertilization: A) Ovule of untreated female flower x pollen of RL-treated male flower; B) Ovule of RL-treated female flower x pollen of untreated male flower; C) Ovule of untreated female flower x pollen of PGR-treated male flower; D) PGR-treated female flower x pollen from untreated male flower; E) pollen penetration into embryo sac (ES) and fertilization; F) Ovule of untreated female flower x pollen of untreated male flower. NB – Nucellar beak; NU – Nucellus; red arrows point at pollen tubes; and yellow circle surrounds an embryo sac region within the nucellus.
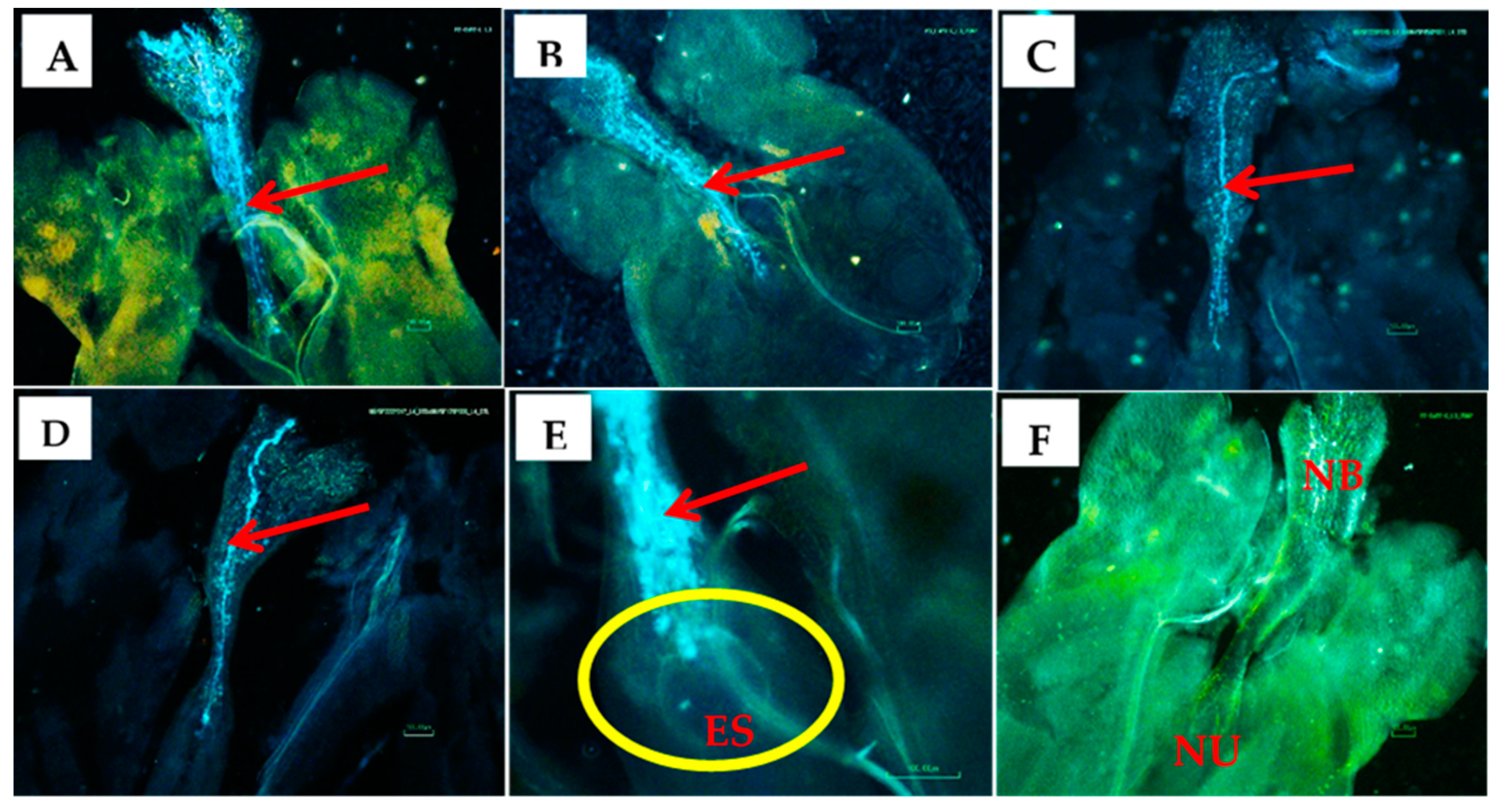
Figure 2.
Pollen stainability and its correlation with diameter in flowers subjected to different treatments: A) plant growth regulators (PGR) treatment; B) red light (RL) treatment; C) no treatment. A1, B1 and C1 show pollen stained with Alexander stain and observed at 10x magnification using an inverted microscope and images taken with a camera head, Nikon DS-L3. Pollen grains stained reddish purple are viable while those stained pale green are unviable.
Figure 2.
Pollen stainability and its correlation with diameter in flowers subjected to different treatments: A) plant growth regulators (PGR) treatment; B) red light (RL) treatment; C) no treatment. A1, B1 and C1 show pollen stained with Alexander stain and observed at 10x magnification using an inverted microscope and images taken with a camera head, Nikon DS-L3. Pollen grains stained reddish purple are viable while those stained pale green are unviable.
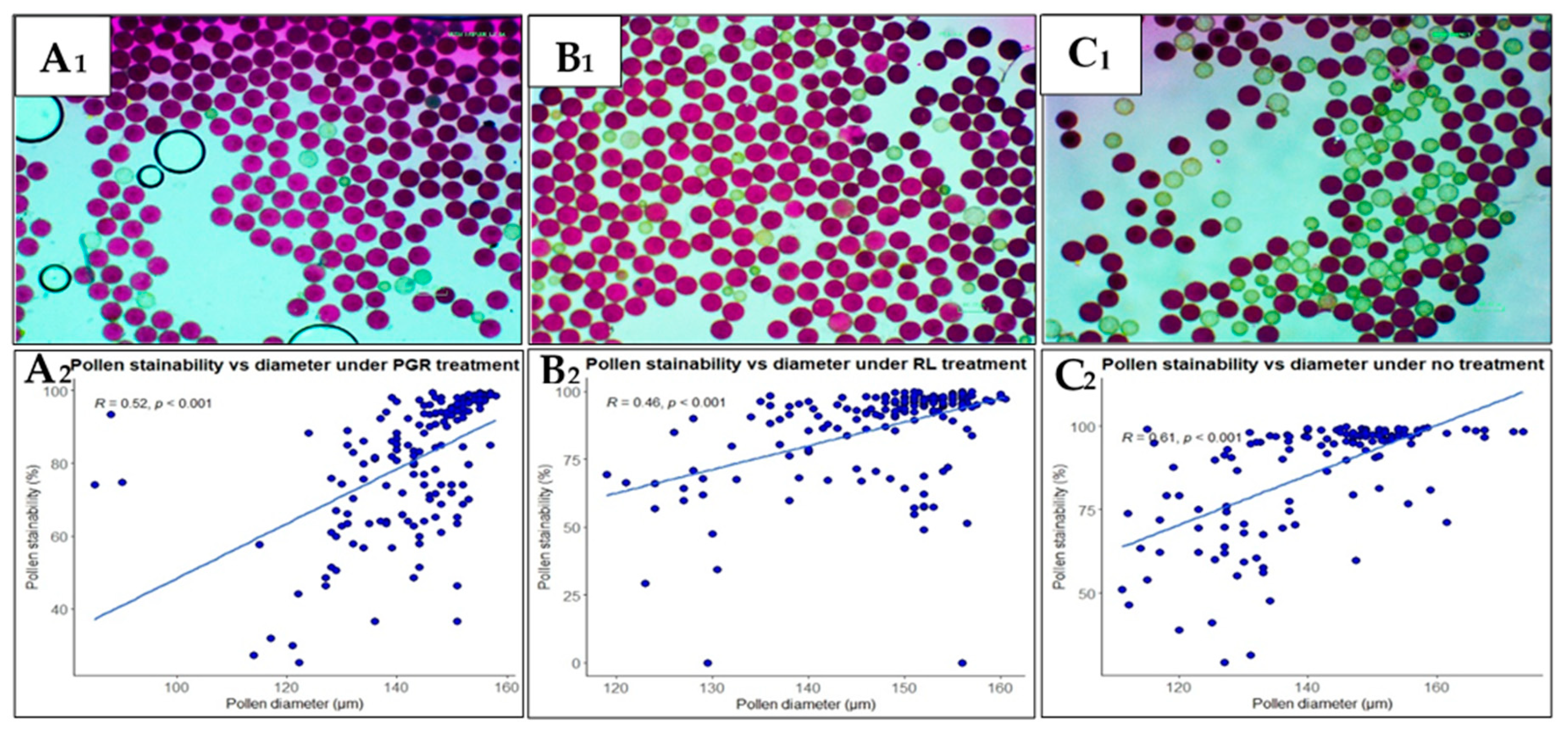
Figure 3.
Mean efficiency of ovule fertilization, fruit and seed set in flowers subjected to PGR treatments: A) Ovules fertilized in treated female flowers; B) Ovules fertilized using pollen from treated flowers; C & D) Fruit set when female and male flowers were respectively treated; E & F) Seed set when female and male flowers were respectively treated. The black lines inside each boxplot represent the median value and the dots inside each boxplot represent the mean value.
Figure 3.
Mean efficiency of ovule fertilization, fruit and seed set in flowers subjected to PGR treatments: A) Ovules fertilized in treated female flowers; B) Ovules fertilized using pollen from treated flowers; C & D) Fruit set when female and male flowers were respectively treated; E & F) Seed set when female and male flowers were respectively treated. The black lines inside each boxplot represent the median value and the dots inside each boxplot represent the mean value.
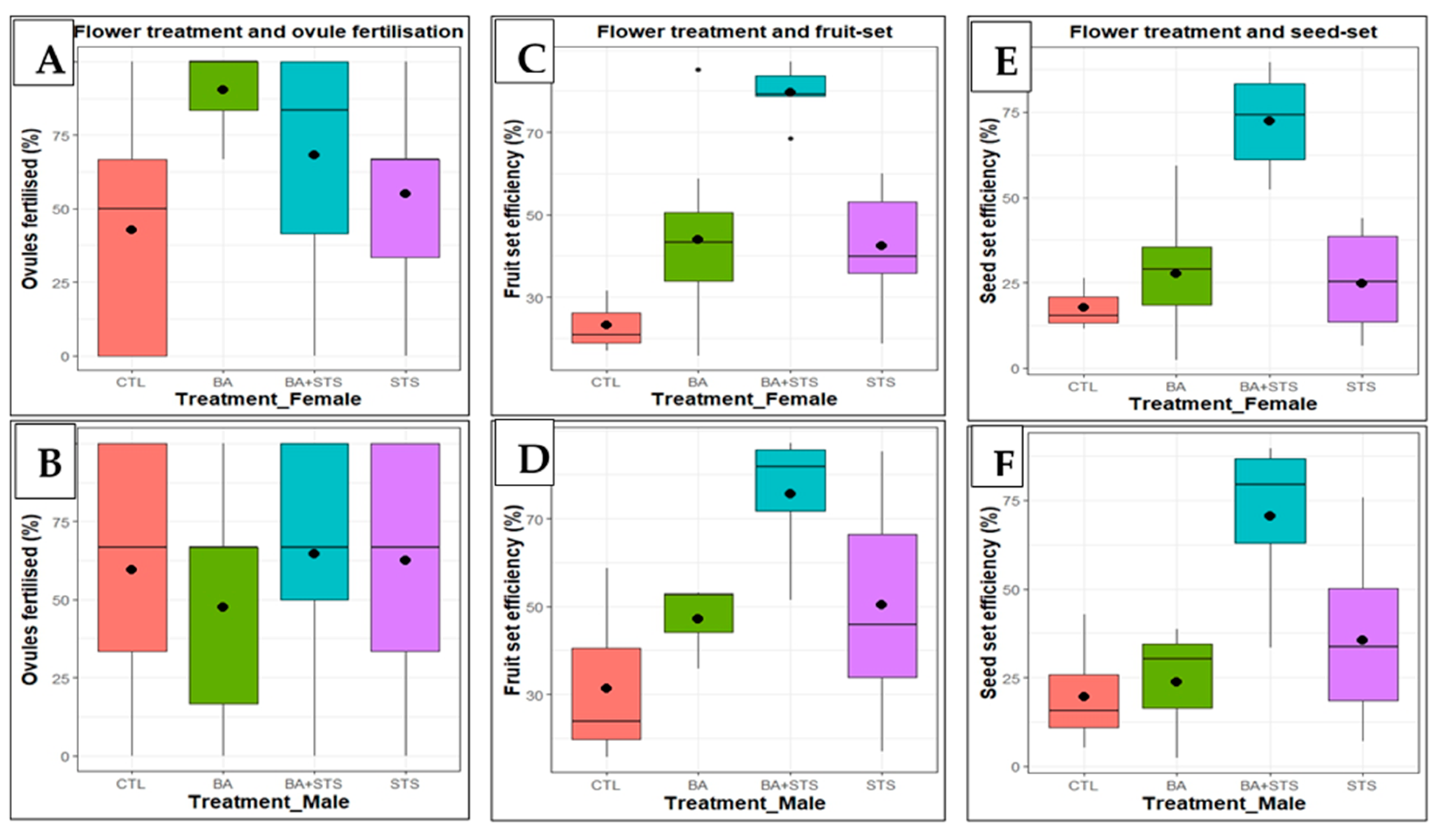
Figure 4.
Comparison of effects of subjecting flowers to PGR and RL treatments on efficiency of ovule fertilization, fruit and seed set: A) Ovules fertilized in treated female flowers; B) Ovules fertilized using pollen from treated male flowers; C & D) Fruit set when female and male flowers were respectively treated; E & F) Seed set when female and male flowers were respectively treated.
Figure 4.
Comparison of effects of subjecting flowers to PGR and RL treatments on efficiency of ovule fertilization, fruit and seed set: A) Ovules fertilized in treated female flowers; B) Ovules fertilized using pollen from treated male flowers; C & D) Fruit set when female and male flowers were respectively treated; E & F) Seed set when female and male flowers were respectively treated.
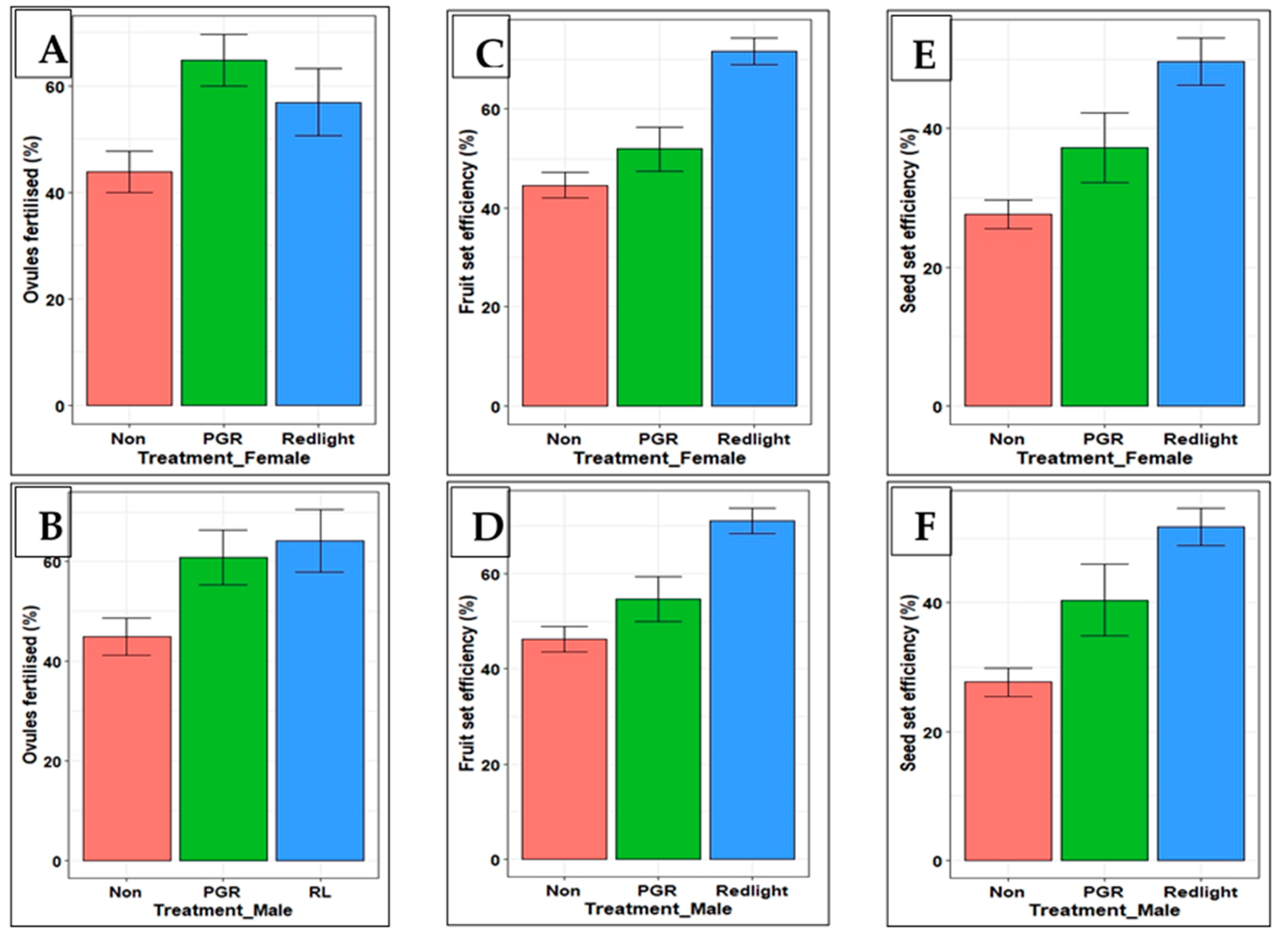
Figure 5.
Cassava pollen ploidy and correlations: A) Effect of RL treatment on ploidy level; the dots inside each boxplot represent the mean value. B) Correlation between ploidy and pollen characteristics.
Figure 5.
Cassava pollen ploidy and correlations: A) Effect of RL treatment on ploidy level; the dots inside each boxplot represent the mean value. B) Correlation between ploidy and pollen characteristics.
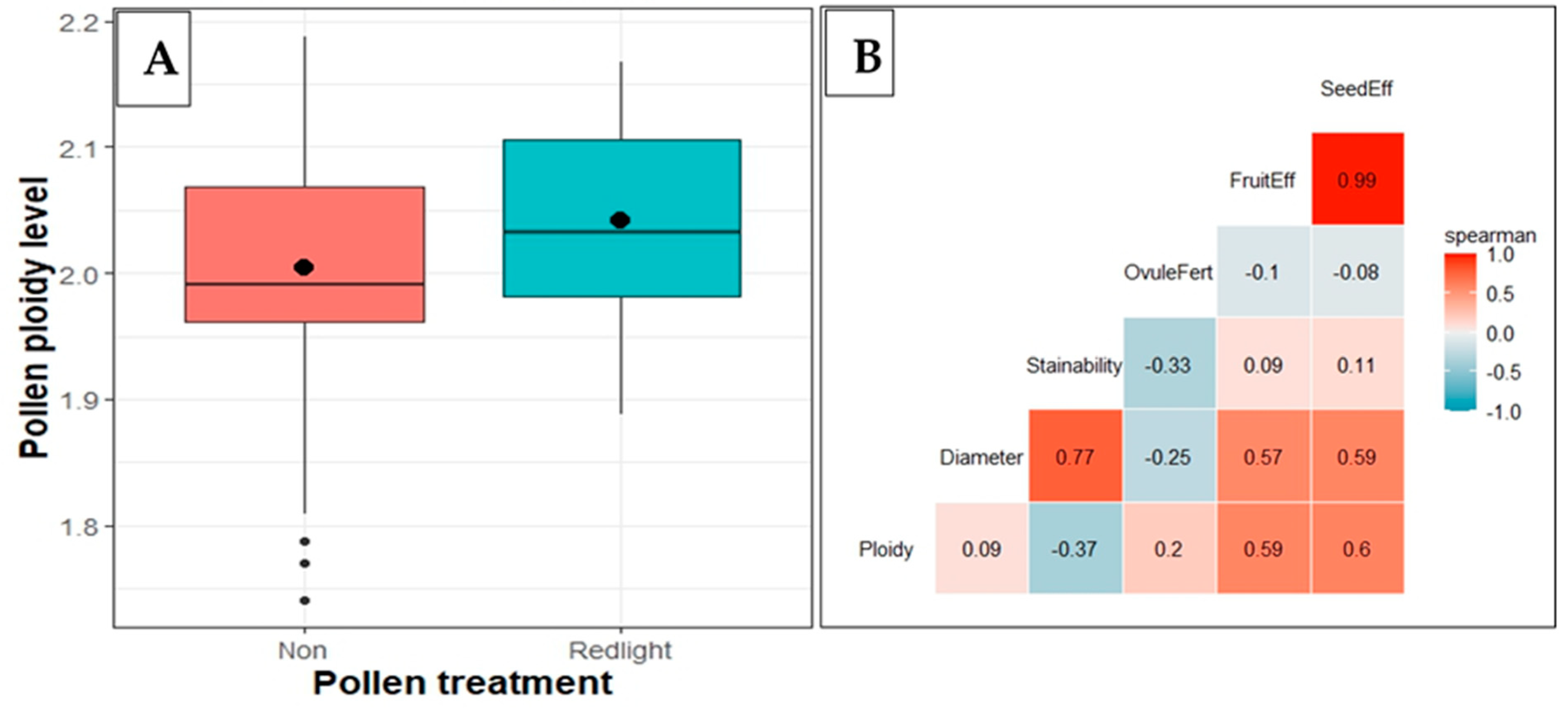
Figure 6.
Flow cytometry profile for cassava pollen samples analysed for DNA content and ploidy levels. 2C represents the diploid amount of DNA at the G1 stage of the cell cycle while the 4C is the doubled amount at the G2 stage following DNA synthesis. 6C and 8C show DNA amounts due to endoreduplication.
Figure 6.
Flow cytometry profile for cassava pollen samples analysed for DNA content and ploidy levels. 2C represents the diploid amount of DNA at the G1 stage of the cell cycle while the 4C is the doubled amount at the G2 stage following DNA synthesis. 6C and 8C show DNA amounts due to endoreduplication.
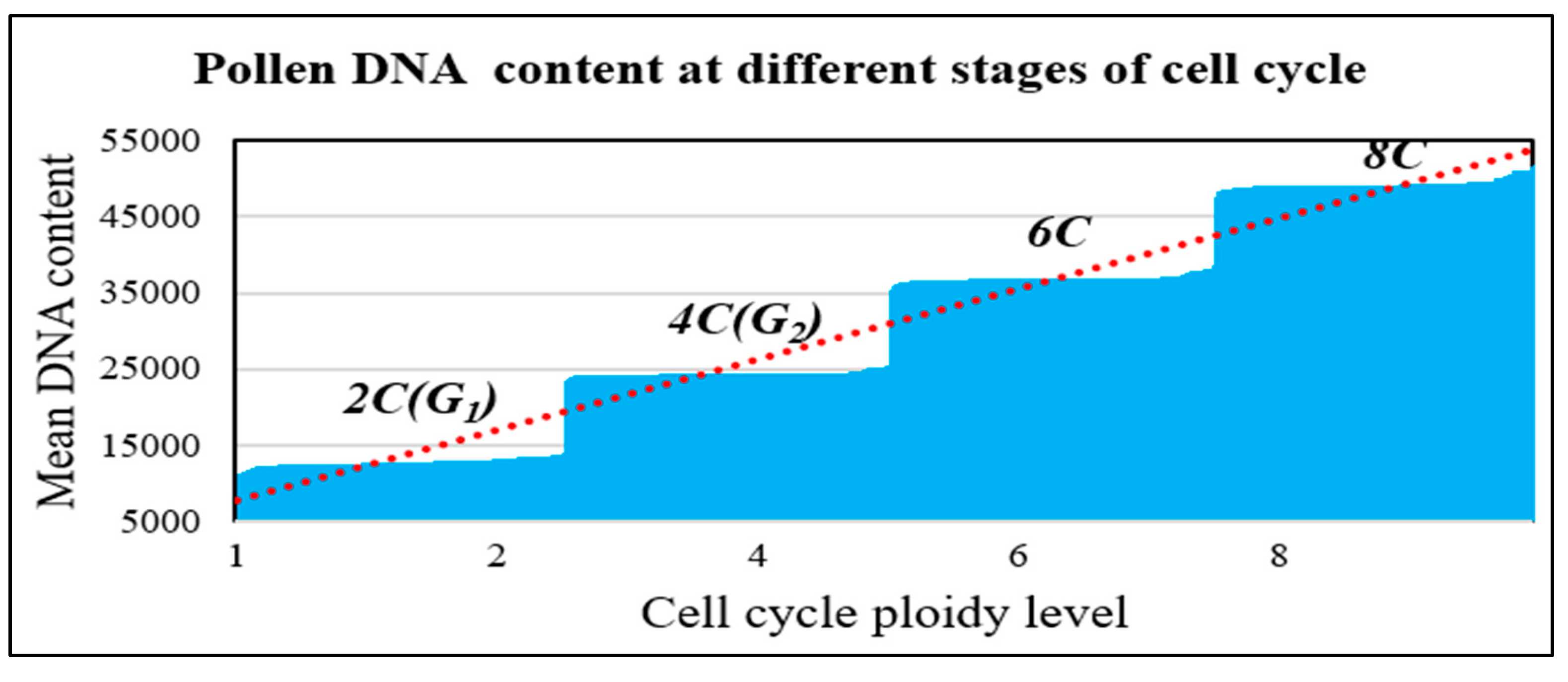
Figure 7.
Representative flow cytometric histograms of pollen and leaf nuclear preparations for cassava: A) Cassava young leaf (control); B & C) Pollen samples picked at anthesis from RL-treated flowers; D) Pollen sample picked at anthesis from non-treated plants.
Figure 7.
Representative flow cytometric histograms of pollen and leaf nuclear preparations for cassava: A) Cassava young leaf (control); B & C) Pollen samples picked at anthesis from RL-treated flowers; D) Pollen sample picked at anthesis from non-treated plants.
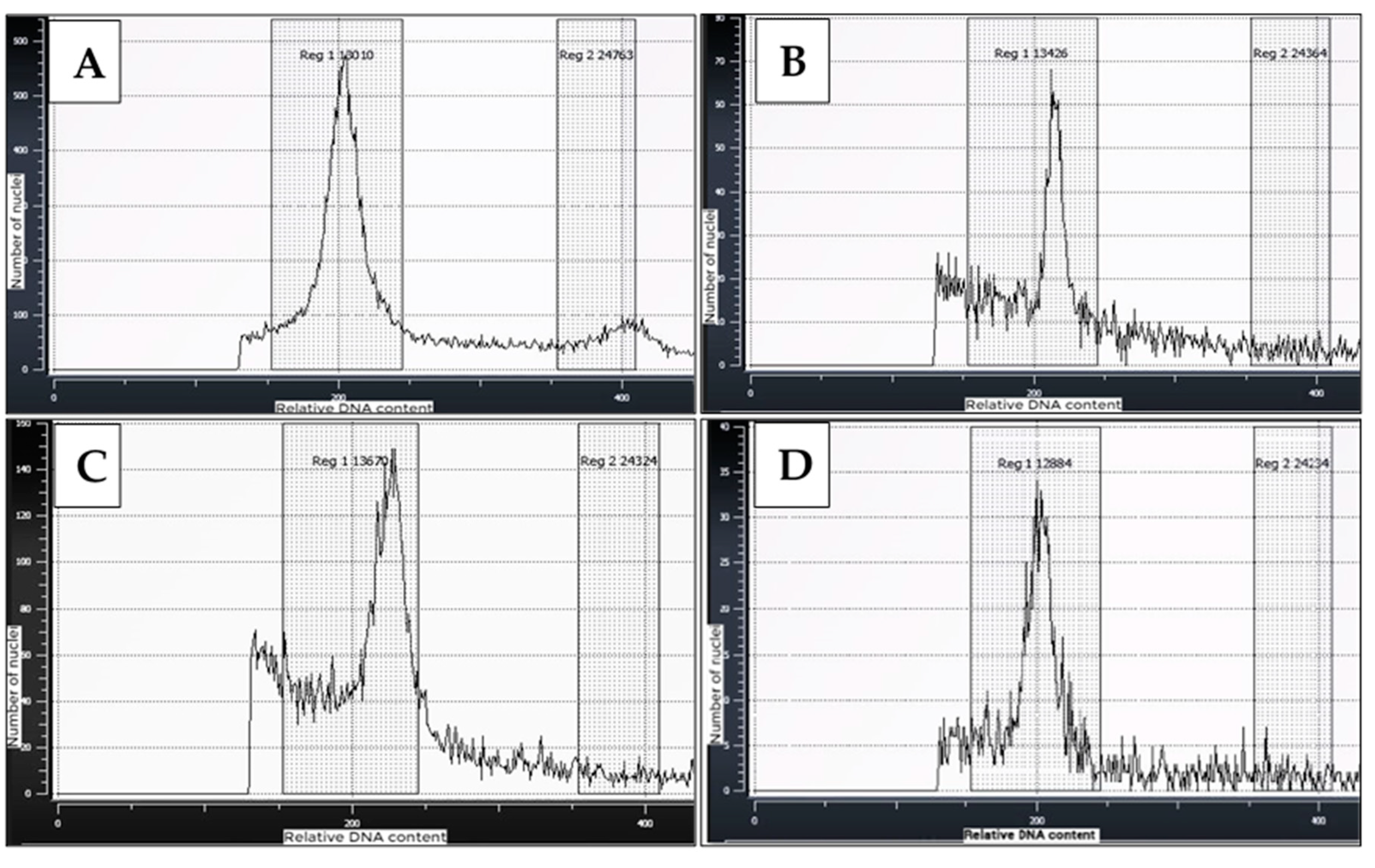

Table 1.
Variation of pollen characteristics and its ability to cause fruit and seed set when treated with RL and PGR.
Table 1.
Variation of pollen characteristics and its ability to cause fruit and seed set when treated with RL and PGR.
| Variable | Pollen diameter (µm) | Pollen stainability (%) | Ovules fertilized (%) |
Fruit set efficiency (%) | Seed set efficiency (%) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatment | RL | PGR | Non | RL | PGR | Non | RL | PGR | Non | RL | PGR | Non | RL | PGR | Non |
| Mean | 147.6 | 143.0 | 141.5 | 86.6 | 80.7 | 86.5 | 54.2 | 60.4 | 34.0 | 60.7 | 48.8 | 26.5 | 39.8 | 35.2 | 16.5 |
| SEM | 0.7 | 1.0 | 1.2 | 1.4 | 1.4 | 1.5 | 4.2 | 4.5 | 6.0 | 2.2 | 4.3 | 4.2 | 2.3 | 4.7 | 3.0 |
| CI | 1.4 | 1.9 | 2.4 | 2.7 | 2.8 | 2.9 | 8.3 | 8.9 | 12.2 | 4.4 | 8.8 | 9.2 | 4.6 | 9.5 | 6.7 |
| Variance | 83.5 | 150.7 | 191.9 | 305.5 | 318.7 | 284.3 | 1856.2 | 1381.9 | 1714.9 | 368.3 | 511.0 | 209.9 | 393.2 | 606.0 | 111.0 |
| Std | 9.1 | 12.3 | 13.9 | 17.5 | 17.9 | 16.9 | 43.1 | 37.2 | 41.4 | 19.2 | 22.6 | 14.5 | 19.8 | 24.6 | 10.5 |
| CV | 0.1 | 0.1 | 0.1 | 0.2 | 0.2 | 0.2 | 0.8 | 0.6 | 1.2 | 0.3 | 0.5 | 0.5 | 0.5 | 0.7 | 0.6 |
RL = Red light; PGR = Plant growth regulator; SEM = Standard error of mean; CI = Confidence interval; CV = Coefficient of variation.
Table 2.
Pollen diameter and stainability under PGR and RL treatments.
| Treatment | Pollen diameter (µm) | Pollen stainability (%) |
|---|---|---|
| PGR | 142.4 b | 82.1 b |
| Red light | 148.5 a | 93.0 a |
| Control (Non) | 143.6 b | 82.9 b |
Different lower-case letters within columns indicate significant differences among treatments by Tukey’s HSD test at P ≤ 0.05.
Table 3.
Efficiency of ovule fertilization, fruit and seed set in flowers subjected to RL treatment.
Table 3.
Efficiency of ovule fertilization, fruit and seed set in flowers subjected to RL treatment.
| Treatment | Ovules fertilised (%) | Fruit set efficiency (%) | Seed set efficiency (%) | |||
|---|---|---|---|---|---|---|
| Female | Male | Female | Male | Female | Male | |
| Red light | 56.9 a | 64.3 a | 71.8 a | 71.1 a | 49.7 a | 51.8 a |
| Control | 52.0 b | 47.7 b | 51.5 b | 53.7 b | 31.7 b | 31.9 b |
| Significance (α = 0.05) | *** | *** | *** | *** | *** | *** |
Different lower-case letters within columns indicate significant differences among treatments by Tukey’s HSD test at P ≤ 0.05. ANOVA tests of significant main effects are indicated: P ≤ 0.001 (***).
Table 4.
Efficiency of ovule fertilization, fruit and seed set in flowers of cassava genotypes subjected to RL treatment.
Table 4.
Efficiency of ovule fertilization, fruit and seed set in flowers of cassava genotypes subjected to RL treatment.
| Genotype | Ovules fertilised (%) | Fruit set efficiency (%) |
Seed set efficiency (%) | |||
|---|---|---|---|---|---|---|
| Female | Male | Female | Male | Female | Male | |
| UG15F178P001 | 65.1 a | 66.7 a | 54.7 b | 58.1 c | 37.0 c | 41.2 a |
| UG15F180P005 | 25.4 c | 53.0 b | 53.8 b | 59.5 bc | 30.1 d | 39.4 ab |
| UG15F192P012 | 57.1 b | 43.3 c | 63.9 a | 65.9 a | 49.6 a | 40.7 a |
| UG15F302P016 | 62.2 a | 56.9 b | 64.8 a | 60.1 b | 40.5 b | 38.4 b |
| Significance (α = 0.05) | ns | *** | *** | *** | *** | *** |
Different lower-case letters within columns indicate significant differences among genotypes by Tukey’s HSD test at P ≤ 0.05. ANOVA tests of significant main effects are indicated: P ≤ 0.001 (***), n.s.: not significant.
Table 5.
Efficiency of ovule fertilization, fruit and seed set in flowers subjected to PGR treatment.
Table 5.
Efficiency of ovule fertilization, fruit and seed set in flowers subjected to PGR treatment.
| Treatment | Ovules fertilized (%) | Fruit set efficiency (%) | Seed set efficiency (%) | |||
|---|---|---|---|---|---|---|
| Female | Male | Female | Male | Female | Male | |
| PGR | 64.9 a | 60.9 a | 51.9 a | 54.7 a | 37.3 a | 40.4 a |
| Control | 42.9 b | 59.4 a | 23.2 b | 31.3 b | 17.7 b | 19.6 b |
| Significance (α = 0.05) | *** | ns | *** | *** | *** | *** |
Different lower-case letters within columns indicate significant differences among treatments by Tukey’s HSD test at P ≤ 0.05. ANOVA tests of significant main effects are indicated: P ≤ 0.001 (***), n.s.: not significant.
Table 6.
Efficiency of ovule fertilization, fruit and seed set in flowers of cassava genotypes treated with PGR.
Table 6.
Efficiency of ovule fertilization, fruit and seed set in flowers of cassava genotypes treated with PGR.
| Genotype | Ovules fertilized (%) | Fruit set efficiency (%) | Seed set efficiency (%) | |||
|---|---|---|---|---|---|---|
| Female | Male | Female | Male | Female | Male | |
| NASE14 | - | 90.0 a | 49.5 b | 55.7 b | 30.7 bc | 42.0 ab |
| UG15F056P001 | - | 79.2 b | 79.5a | 35.9 e | 57.3 a | 30.2 cd |
| UG15F192P012 | 81.5 a | 33.4 e | 72.2 a | 41.8 cde | 68.4 a | 25.1 a |
| UG15F222P017 | 48.7 c | - | 32.8 d | 58.6 ab | 25.0 c | 26.4 cd |
| UG15F228P016 | 57.5 b | - | 40.4 c | - | 23.8 c | - |
| UG15F302P016 | 83.3 a | 41.7 e | 49.5 b | 63.1 a | 36.0 b | 43.7 ab |
| Significance (α = 0.05) | * | ns | * | *** | *** | *** |
Different lower-case letters within columns indicate significant differences among genotypes by Tukey’s HSD test at P ≤ 0.05. ANOVA tests of significant main effects are indicated: P ≤ 0.05 (*), P ≤ 0.001 (***), n.s.: not significant.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



