Submitted:
29 May 2024
Posted:
30 May 2024
Read the latest preprint version here
Abstract
Mango twig-tip dieback (MTTD) is a new type of mango decline, but its impact on the trees’ mineral nutrition is unknown. This study was conducted to determine the effect of MTTD infection on the nutrient status, balance, and resorption efficiency (RE) of mango. Leaf nutrient concentrations and deviation from the optimum percentage (DOP) indices of ‘Kensington Pride’ (KP) mango trees with low (LD) and high (HD) levels of MTTD infections were analyzed to compare the foliar nutrition status and nutrient balance between the LD and HD trees. Moreover, the nutrient resorption efficiency of MTTD-infected dried leaves (RED) was compared with the resorption efficiency of healthy (RES) leaves of KP mangoes. The concentrations of Ca, Mg, Cu, Fe, Mn, and Zn were lower in the HD trees than in the LD trees. But the K content was higher in the HD trees, and its DOP index was sufficient, while the K concentration was low and deficient level in LD trees. Moreover, DOP indices for Ca, Mn, and Zn were less deficient in LD trees than in HD trees, and overall nutrient imbalances were exacerbated in HD trees. RED was significantly lower than RES for N, P, S, Cu, Fe, and Zn but significantly higher than RES for K. The study underscores the significant influence of MTTD on the mineral nutrition of KP mangoes, revealing distinct nutrient variations between trees with low and high MTTD infection levels. These findings have important implications for mango crop management, emphasizing the need for targeted nutrient interventions to address imbalances induced by MTTD and enhance the overall health and resistance of mango trees against MTTD infection.
Keywords:
deviation from optimum percentage
; foliar nutrition
; mango sudden decline
; nutrient imbalance
; remobilization
1. Introduction
Mangoes are the main horticultural crop of the Northern Territory (NT), the largest producer of mangoes in Australia. NT growers produce 52% of the nation's mangoes and give Australian customers their first crop harvest of the season. The mango industry provides approximately $128 million to the NT economy and employs about 3500 people [1]. However, plant diseases pose a significant threat to maintaining or increasing mango production in the NT. Plant diseases can significantly reduce farm profitability owing to yield losses, higher production costs, and poorer crop quality [2,3,4]. Some of the common mango diseases in the NT include anthracnose [5], stem end rot [2,6], resin canal discoloration [3,7], and sudden decline or common dieback [8,9,10].
Common dieback causes serious production issues for mango growers [8,9,10,11,12,13,14]. It is characterized by a progressive and downward drying out or dying back of the twig or branch, sometimes with the death of the whole branch. Twig/branch/stem cankers are also a typical symptom. In very severe cases, this dieback can lead to the death of the mango tree, particularly when trees are under prolonged periods of drought with inadequate irrigation. Tree death is not common in managed orchards. Common die-back symptoms are not frequently found on young trees, its intensity appears to in-crease with the plant age [14]. Some visible symptoms of common dieback include gummosis, vascular discoloration, death of twigs from the top, streaking, and bark splitting. The leaves of the affected tree dry up and remain attached to the dying tree, and the infected trunk oozes badly with a smelling exudate [10,11,12]. Both biotic and abiotic factors are likely to be responsible for the symptoms of mango dieback. For biotic factors, reports identified several genera and species in the family Botryosphaeriaceae as organisms associated with dieback in mangoes [8,10]. Moreover, abiotic factors that may play a role in dieback include relative humidity, high temperatures, sun scald, water stress, drought, and cultural management practices [11,13].
Mango growers in the Darwin region of NT reported a type of dieback that showed different symptoms compared with that of common dieback. The symptom was a dark longitudinal lesion that was only on one side of the last twig flush above the last node. The apical bud dies as the lesion spreads upward and downward. The leaves of the last flush dried out and temporarily stayed affixed to the twig. Occasionally, only the apical bud had dried leaves on one side, and a black lesion was visible on the second and third flushes, with or without gum exudation. The middle of the twig, either between two nodes or between the apical bud and terminal node, appears to be where the majority of lesions are located. Moreover, the internal lesion extended a few centimeters beyond the visible symptoms. Transversely cutting the twig reveals that the lesion is not in the center but on the outside. Thus, it is called twig-tip dieback (MTTD), which distinguishes it from common dieback (Figure 1). The etiology of MTTD, if biotic or abiotic, is presently unknown. The intensity of MTTD differed among orchards, with certain locations experiencing severe impacts. MTTD severity varied across orchards, impacting some areas more severely than others. MTTD occurrence was not solely dependent on plant stress, as it had been found in orchards with healthy trees and even in previously abandoned settings. Remarkably, MTTD affected young mango trees, particularly the KP cultivar, showing its susceptibility regardless of tree age [15].
Mineral nutrition is important not only for the optimum growth and yield of mangoes but also plays a significant role in plant and disease interactions [16,17]. The nutrient composition and imbalances affect plant growth, flowering traits, fruit development [18], and disease susceptibility through metabolic changes that influence the vascular system of plants [10,19,20]. Adequate levels of essential nutrients are also needed for key physiological processes such as photosynthesis and stomatal regulation, which determine how trees cope with stress [21]. Furthermore, because of the direct role of mineral elements in plant defense, comprehensive and balanced nutrition has traditionally been the first line of defense for plants by modifying root exudates, the dynamics of the microflora population, and the production of phytoalexins, all of which have a direct impact on plant health [16]. Thus, ensuring comprehensive and balanced nutrition stands as the cornerstone of safeguarding plant health, highlighting its pivotal role as the primary defense mechanism for plants against adversity and disease challenges.
However, there are no reports on the effect of MTTD on mineral nutrition in mangoes. In common dieback, studies have shown that nutrient stress predisposes and contributes to the common dieback problem in sugar maples [22], cherries [23], eucalypts [24], and other trees [25]. Saeed et al. [11] found that common dieback disturbs the level of important phytochemicals and minerals in the stem bark of mango. In citrus, the effects of Huanglongbing (HLB) pathogen infection on P resorption efficiency (PRE) varied depending on the species of citrus [26]. Therefore, it is important to elucidate the effects of MTTD on the nutrient dynamics and resorption efficiency (RE) of mangoes. Because leaves are the most dynamic and responsive plant part for metabolic activity and nutrient availability [27], leaf nutrient analysis provides the actual uptake of nutrients and indicates the existing nutrient levels when compared to the standard optimum leaf nutrient content [28]. Leaf analysis can identify imbalances and aid in formulating fertilizer recommendations to address nutritional deficiencies [16]. Moreover, nutrient analysis of senescing leaf samples provides information on the nutrient resorption efficiency [29].
The investigation into the cause of MTTD is ongoing. In this study, we present the analysis of nutrient levels in a commercial mango orchard featuring trees exhibiting varying degrees of MTTD. This study aimed to (1) assess leaf nutrient levels in trees affected by differing MTTD intensities, (2) ascertain if MTTD intensity contributes to elevated nutrient imbalances, and (3) understand how MTTD intensity influences the nutrient resorption efficiency of mango trees.
2. Materials and Methods
2.1. Study Site
This study was conducted in a commercial mango orchard in the Litchfield municipality of NT, Australia (14° 27′ 52.6′′ S, 132° 18′ 48.5′′ E) with a population density of 100 trees per hectare. The orchard was one of the 15 mango orchards surveyed for the occurrence of MTTD [15]. The soil was classified as red Kandosol with sandy loam soil texture. The climate is characterized by a pronounced rainy season from July to October, with annual rainfall ranging from 800 to 1100 mm. The average monthly temperature was 25 °C from November to January and 33 °C from April to May.
2.2. Twig-Tip Dieback Evaluation
One hundred trees with varying levels of MTTD infection were selected from the middle part of the orchard. Trees were all affected by MTTD and the severity of MTTD infection was scored from one to four, with Level 1 having the lowest infection and Level 4 having the highest infection, based on the number of twigs infected in the north-south and east-west orientations of the tree. Level 1 trees had less than 10% TTD, Level 2 trees had 25% TTD, Level 3 trees had 50% TTD, and Level 4 trees had more than 50% TTD. From the 100 trees evaluated, 10 trees with low TTD infection (LD trees) and 10 trees with high TTD infection (HD trees) were selected and tagged for sample collection (Figure 2).
2.3. Sample Collection
Leaf samples were collected from each of the ten LD and HD trees in January 2022 to compare the nutritional status of trees with low- and high-dieback infections. Twenty mature, healthy green leaf samples were randomly selected from healthy twigs located at the top, bottom, east, and west sides of each tree. In addition, MTTD-affected leaves were collected to assess the effects of dieback on nutrient absorption. The MTTD-affected leaves were those that dried because of MTTD but were still attached to the twig and surrounded by green leaves on the opposite side of the twig. Composite soil samples were collected from five LD and five HD trees. Each composite sample comprises soil samples taken from four points around the tree, approximately 1.5 meters away from the trunk, using a soil auger with a sampling depth of 20cm. All samples were placed in a tagged bag, immediately put in an esky with ice, and transported to the laboratory at Berrimah Farm Science Precinct in Darwin (12° 26′ 38.79′′ S, 130° 55′ 46.61′′ E) for processing.
Figure 2.
KP mango trees with low MTTD infection (LD trees) symptoms (a) and high MTTD infection (b) symptoms. LD trees had less than 10% MTTD, while HD trees had 50% MTTD, and Level 4 trees had more than 50% MTTD. The left side shows the whole tree picture, while the right side shows a close up of the MTTD infection and its symptoms.
Figure 2.
KP mango trees with low MTTD infection (LD trees) symptoms (a) and high MTTD infection (b) symptoms. LD trees had less than 10% MTTD, while HD trees had 50% MTTD, and Level 4 trees had more than 50% MTTD. The left side shows the whole tree picture, while the right side shows a close up of the MTTD infection and its symptoms.

2.4. Sample Processing and Analysis
Soil samples were air-dried in the laboratory, and leaf samples were washed with tap water and rinsed with reverse osmosis water before oven-drying at 60 °C until a constant dry weight was attained. Samples were sent to the CSBP Soil and Plant Analysis Laboratory (Perth, Australia) and analyzed according to the company’s laboratory procedure (https://csbp-fertilisers.com.au/services/lab). The analyzed soil parameters were pH, electrical conductivity (EC), available nitrogen (N), available phosphorus (P), available potassium (K), organic carbon (OC), exchangeable calcium (Ca), exchangeable magnesium (Mg), available sulfur (S), extractable boron (B), extractable copper (Cu), extractable iron (Fe), extractable manganese (Mn), and extractable zinc (Zn). For the leaf samples, the nutrients analyzed were the macronutrients N, P, K, Ca, Mg, and S, as well as the micronutrients B, Cu, Fe, Mn, and Zn.
2.5. Sufficient Levels of Nutrient Status of the Leaves
The nutrient status of the mango trees was evaluated based on the following sufficiency levels: N = 1-1.5%, P = 0.08-0.18%, K = 0.3-1.2%, Ca = 2.0-3.5%, Mg = 0.15-0.40%, S = 0.5-0.6%, B = 50-80 mg/kg, Cu = 10-200 mg/kg, Fe = 70-200 mg/kg, Mn = 60-500 mg/kg and Zn = 2-150 mg/kg [30].
2.6. Deviation from the Optimum Percentage (DOP) Index
The DOP index was calculated to diagnose imbalances in leaf minerals, according to Heras and Montañés [31]. The DOP index was computed using the formula:
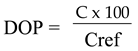
2.7. Nutrient Resorption Efficiency
The impact of MTTD severity on RE (RED) was computed using the formula:
where NG and ND are the nutrient concentrations of green leaves and dried leaves in NTT-affected trees, respectively; and MCF is the mass loss correction factor used to compensate for the loss of leaf mass during leaf drying, which is the ratio of the dry mass of ND to the dry mass of NG (Figure 3a). Moreover, RED was compared with the mean RE of senesced (RES) leaves from five orchards (12° 33′ 39.38′′ S, 131° 18′ 17.05′′ E; 12° 47′ 28.07′′ S, 131° 09′ 30.67′′ E; 12° 44′ 52.80′′ S, 131° 10′ 38.97′′ E; 14° 27′ 59.91′′ S, 131° 18′ 45.39′′ E; 14° 32′ 24.90′′ S, 132° 28′ 6.20′′ E), where there were no symptoms of MTTD. Senesced leaves were yellow leaves that were still attached to the twig, but were easily knocked off with a slight touch (Figure 3b). RES was computed using the formula:
where NG and NS are the nutrient concentrations in green and senesced leaves, respectively, and MCF is the ratio of the dry mass of NS to the dry mass of NG [34].
2.8. Statistical Analysis
The data on soil and leaf nutrient concentrations, DOP, RES, and RED were statistically analyzed by comparing the means between LD and HD trees using a two-sample independent t-test at the 5% level of significance. Normality and homogeneity of variance were checked using Bartlett’s test and Shapiro-Wilk’s test, respectively. Statistical analysis and visualization were done using GraphPad Prism version 10.0.0 software (www.graphpad.com).
Figure 3.
Photograph of dried leaves from dieback infection (a) and senescing leaves of KP mango.

3. Results
3.1. Soils and Lef Nutrient Analysis
Soil analysis showed that the soil properties of LD and HD trees were comparable (Table 1), but leaf analysis displayed significant differences in leaf nutrient contents between LD and HD trees (Figure 4). The results also showed that MTTD infection caused significant variations in certain nutrients; however, some nutrients maintained relatively consistent levels, regardless of MTTD severity. For example, the concentrations of Ca and Mg were higher (p < 0.001) in the LD trees (Ca = 1.37 ± 0.05%, Mg = 0.50 ± 0.05%) than in the HD trees (Ca = 1.07%, Mg = 0.37%).
A similar trend was also observed for micronutrients Cu, Fe, Mn, and Zn, where their concentrations (mg/kg) were higher (p < 0.05) in the LD (Cu = 31 ± 8, Fe = 74.2 ± 3, Mn = 130 ± 7, and Zn = 17 ± 1) than in the HD (Cu = 10 ± 0.3, Fe = 58 ±4, Mn = 83 ± 7, and Zn = 12 ± 0.7) trees. Conversely, the K content was lower (p < 0.01) in LD (0.63 ± 0.02%) trees than in HD (0.75 ± 0.02%) trees, whereas the levels of the macronutrients N, P, and S, as well as the micronutrient B content, were unaffected by MTTD infection because they were present in comparable (p > 0.05) amounts between LD and HD trees.
3.2. DOP Index
The DOP index of mineral elements was determined to evaluate the deviation of the nutrient concentration from the optimum level [27,35–37]. Regardless of the MTTD level, most of the analyzed nutrients (N, P, Ca, Mg, S, B, Cu, Fe, and Zn) had a negative DOP index, indicating that their concentrations were below the optimum concentration (Figure 5a), and only Mg had a higher concentration than the optimum level owing to its positive DOP index. Comparison of DOP indices among LD and HD trees showed that the DOP index of K was below the optimum at low MTTD infection level (-15.5) and at the optimum level at high MTTD infection level (0.5), while the DOP index of Mn was optimum at low MTTD (-0.3) and below the optimum at high MTTD level (-35.9).
The results also showed that the DOP indices of N, P, S, and B were comparable (p > 0.05) in both LD and HD trees, but the former had a significantly lower negative DOP index for Ca (-57.4), Fe (-45.0), Mn (-0.3), and Zn (-67.8) than the latter (Ca = -61.0, Fe = -56.6, Mn = -35.9, and Zn = -83.6). Conversely, the DOP index for Mg was more positive in the LD (80.4) trees than the HD trees (36.0). Figure 5b illustrates that both LD and HD trees had imbalanced nutrients, but ƩDOP was more pronounced (p < 0.01) in the HD trees (477.7) than the LD (427.1) trees, indicating that MTTD increases nutrient imbalances in mangoes.
3.3. Nutrient Resorption Efficiency
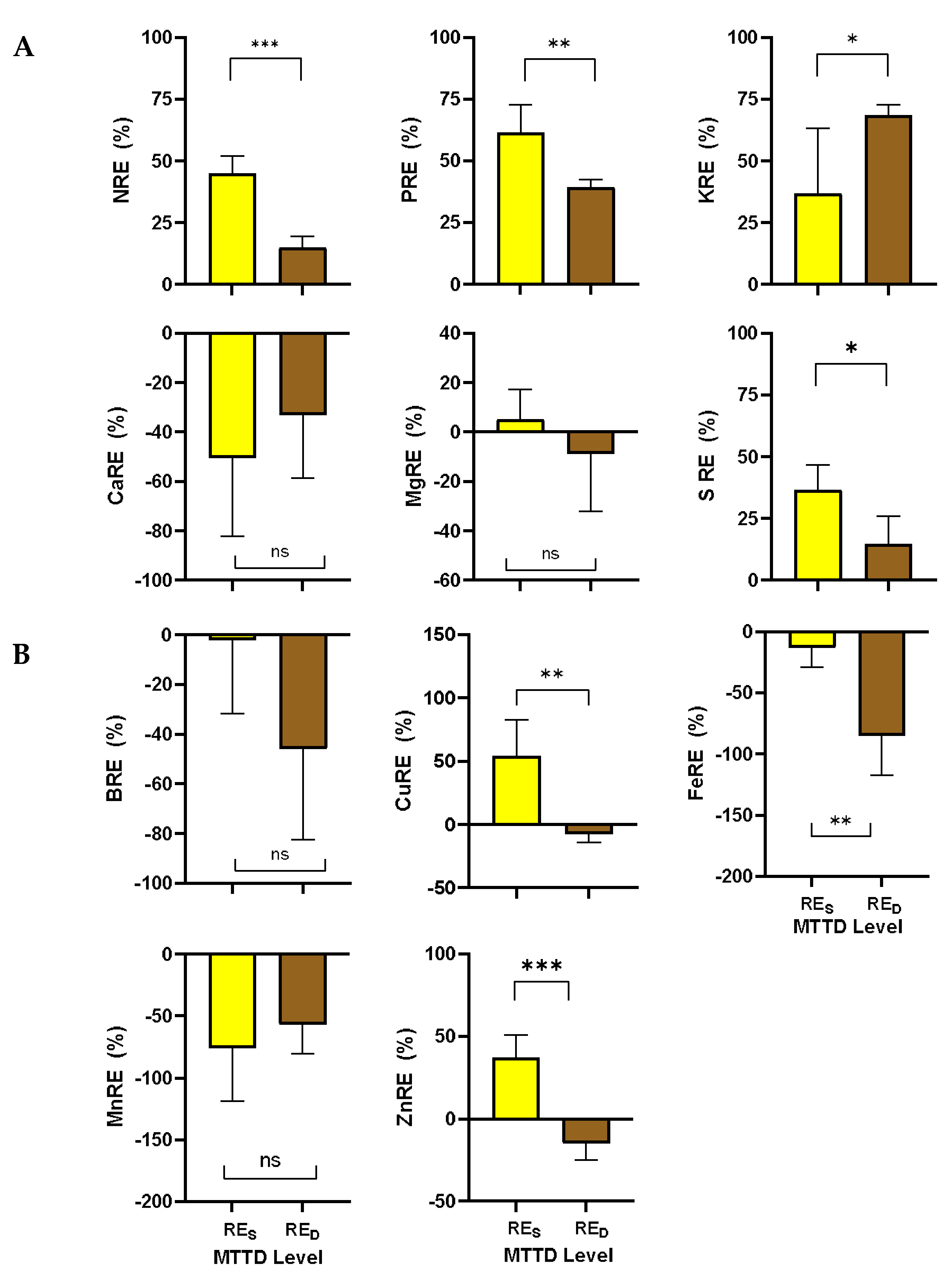
Leaf RES plays an important role in determining plant nutrient-use strategies. It is a mechanism used by plants to recover nutrients from senescing leaves before abscission, allowing the plants to conserve nutrients by remobilizing them and making them available for reuse [38,39,40]. This is an important resource conservation strategy for mangoes in the NT, where the soil has a low fertility level [41]. Thus, determining the RED provides information on the influence of MTTD on nutrient movement and resource economy in mangoes.
In this study, we found that the RED among macro- and micronutrients significantly (p < 0.05) varied between LD and HD trees except for Ca, Mg, B, and Mn (Figure 6). RED was significantly lower for N (14.9%), P (39.4%), and S (14.6%) than RES (N = 45.1%, P = 61%, and S = 36.5%) but significantly higher for K (RED = 68.63%, RES = 36.8%). Among micronutrients, RED was significantly lower for Cu (-7.5%), Fe (-85%), and Zn (-14.7%) than RES (Cu = 53.9%, Fe = -13.1%, and Zn = 37.4%).
Figure 4.
Macronutrients (A) and micronutrients (B) concentration in the healthy leaves of trees with low infection (LD) and high infection (HD) of twig-tip dieback on the KP mango cultivar. Vertical bars represent the standard error of 10 replicates. ns = no significant difference, * = significant at the 0.05 level, ** = significant at the 0.01 level, and *** = significant at the 0.001 level of significance.
Figure 4.
Macronutrients (A) and micronutrients (B) concentration in the healthy leaves of trees with low infection (LD) and high infection (HD) of twig-tip dieback on the KP mango cultivar. Vertical bars represent the standard error of 10 replicates. ns = no significant difference, * = significant at the 0.05 level, ** = significant at the 0.01 level, and *** = significant at the 0.001 level of significance.
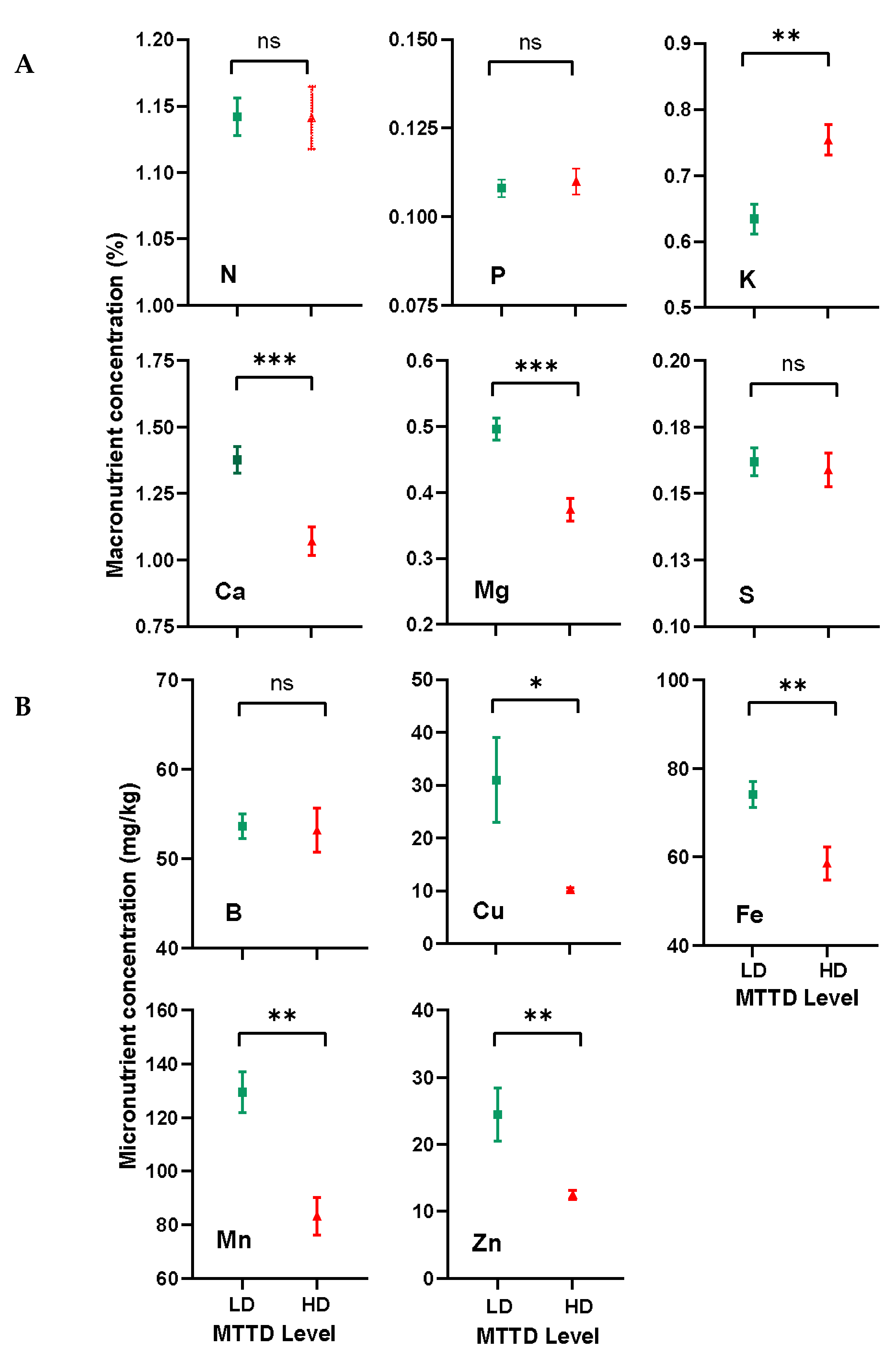
Figure 5.
DOP indices of healthy leaf nutrients (A) and summation of DOP (ƩDOP) of all nutrients (B) of trees with low infection (LD) and high infection (HD) of twig-tip dieback on the KP mango cultivar. Vertical bars represent the standard error of 10 replicates. ns = no significant difference, * = significant at the 0.05 level, ** = significant at the 0.01 level, and *** = significant at the 0.001 level of significance.
Figure 5.
DOP indices of healthy leaf nutrients (A) and summation of DOP (ƩDOP) of all nutrients (B) of trees with low infection (LD) and high infection (HD) of twig-tip dieback on the KP mango cultivar. Vertical bars represent the standard error of 10 replicates. ns = no significant difference, * = significant at the 0.05 level, ** = significant at the 0.01 level, and *** = significant at the 0.001 level of significance.
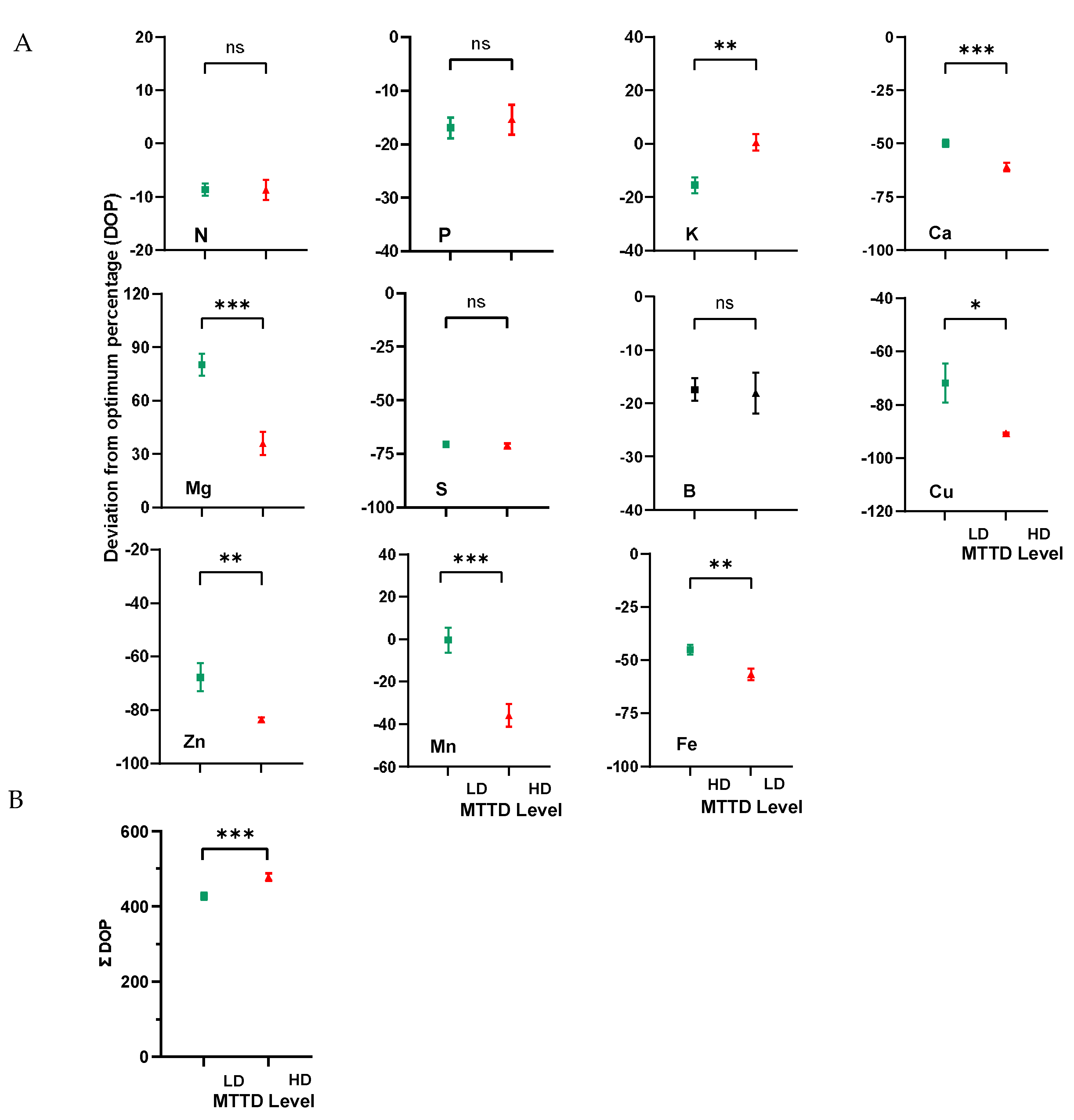
Figure 6.
Comparison of the macronutrients (A) and micronutrients (B) resorption efficiency of healthy leaves (RES) and dieback-infected (RED) leaves of the KP mango cultivar. Vertical bars represent the standard error of 10 replicates. ns = no significant difference, * = significant at the 0.05 level, ** = significant at the 0.01 level. and *** = significant at the 0.001 level of significance.
Figure 6.
Comparison of the macronutrients (A) and micronutrients (B) resorption efficiency of healthy leaves (RES) and dieback-infected (RED) leaves of the KP mango cultivar. Vertical bars represent the standard error of 10 replicates. ns = no significant difference, * = significant at the 0.05 level, ** = significant at the 0.01 level. and *** = significant at the 0.001 level of significance.
4. Discussion
Plant disease management is a major concern for mango growers, and knowledge about the interplay between mineral nutrition and plant diseases is indispensable for effective crop management and promoting sustainability in agriculture moving forward. Both plants and pathogens require mineral nutrients, which play a significant role in their interactions [16,42]. This pilot study focuses on understanding the influence of MTTD on the mineral nutrition of mango. We found that there was no relationship between soil nutrients and MTTD severity, but leaf nutrient contents varied significantly between LD and HD trees (Table 1, Figure 4). The result indicates a direct and significant impact on the nutrient content of tree leaves, independent of soil nutrient availability, as all trees may have similar opportunities for accessing nutrients from the soil and applied fertilizer. In oak tree decline, Azim Nejad et al. [43] also reported that the physical and chemical properties of soil are not determinant factors. Moreover, there were no significant differences in the soil properties of samples collected from healthy trees and those infected with HLB, thus, it is unlikely that the difference in foliar mineral concentrations between infected and PCR-negative trees was due to the differences in soil [44].
The findings also demonstrated that MTTD severity significantly alters several nutrients levels, while others remain at comparatively constant amounts, regardless of the severity of MTTD (Figure 4). This result indicates that the impact of MTTD is nutrient-specific. It also corroborates a study showing the variable effects of diseases on the nutrient content of citrus [36], coconut palm [45], and cacao [46]. For example, in Valencia orange, the severity of the citrus decline index is negatively correlated with Ca, Mg, S, and B and positively correlated with high leaf N, P, K, Cu, and Mn indices in the leaves [36]. Moreover, healthy coconut palms had higher macronutrient (N, P, K, Ca, Mg, and S) and micronutrient (B and Cu) uptake than those with root wilt disease trees, while Mn and Zn were comparable between healthy and diseased trees [45]. In cacao, leaves infected with witches’ broom disease caused by Monoliophtora perniciosa had lower Ca, Mg, S, Fe, and Mn contents in healthy leaves than diseased leaves. Moreover, P and Cu contents were higher in diseased leaves than in healthy leaves, but N, K, and Zn were comparable between healthy leaves and leaves of vegetative broom, indicating differential changes in the concentration of nutrients in the presence of M. perniciosa infection [46].
The increase in K in the leaves of HD trees (Figure 4 and Figure 5) is in line with the report of Putiska et al. [44], showing that a significant negative relationship (r = -0.71) existed between the symptom severity of HLB and concentrations of K in Siem mandarin (Citrus reticulata) in Indonesia. Spann and Schumann [42] also observed an elevated concentration of potassium (K) in citrus (C. sinensis) infected with HLB in three out of four orchards. They attributed this increase to the diminished growth of the trees and an apparent concentration of K induced by HLB. Furthermore, the findings in this study, where HD trees exhibited high K and low Ca leaf contents, suggest a negative interaction between these nutrients. This interaction could be explained by the competition for the same absorption sites in the plant [47]. Maintaining a balanced level of Ca and K has been shown to reduce the severity of disease and defoliation caused by the fungus Calonectria pteridis in eucalyptus plants. The elevation in K levels may have contributed to a decrease in the stomatal closure response and an increase in stomatal opening movement, potentially facilitating infection by the pathogen. Under insufficient Ca concentration, substances such as sugars and amino acids may leak into the apoplast, providing the pathogen with resources for its growth and development [47,48,49].
Our study showed that HD trees experienced more pronounced nutrient imbalances (Figure 5), which may have predisposed the plants to disease susceptibility. Studies have shown that orchards with higher nutrient imbalance indices recorded higher disease severity [15,36,47,50]. The specific nutrient imbalances observed in HD trees may have implications for their health and growth, highlighting the importance of integrated nutrient and disease management strategies in mango production to address these imbalances and promote tree health. The emphasis on balanced nutrition as the first line of defence against diseases in crops stresses the significance of providing plants with the necessary nutrients to increase their natural defences. It is a preventive measure that supports plant health, resilience, and sustainable crop management strategies [16,51]. According to Tadayon and Sadeghi [36], balanced nutrition management could reduce the severity of citrus decline in Valencia orange orchards by increasing the leaf concentrations of Ca, B, S and Mg as higher concentrations of these nutrients were positively correlated with a lower citrus decline severity index. In our earlier report, we found that the imbalance of nutrients such as Ca/Cu, B/Mn, and Fe/Zn ratios may have contributed to the development of RCD mango, a quality defect that appears during fruit ripening, reduces the market price and consumer appeal of mango fruits [7].
This study showed that MTTD significantly reduced the nutrient RE of macronutrients N, P, and S and micronutrients Cu, Fe, and Zn in dieback affected leaves (Figure 6). However, MTTD infection increased the resorption efficiency of K but did not affect Ca, Mg, and Mn, indicating that MTTD influences most of the mobile elements and less of the immobile elements. According to reports, Ca and Mn have poor mobility in the phloem sap, while N, P, K, S, Cu, Fe, and Zn are mobile elements [52,53,54]. Though not as low as that of Ca, Mg transport is likewise low in both xylem and phloem [52,55]. A high concentration of nutrients in leaves coupled with low nutrient resorption typically indicates a strategy where plants invest resources extensively for rapid growth and development. Conversely, a low concentration of nutrients in leaves along with high nutrient resorption signifies a strategy where plants conserve nutrients, prioritizing survival and reproduction over immediate growth [40]. Moreover, the increase in KRE in this study could be explained by the increase in K concentration in the leaves as affected by MTTD infection (Figure 4). The reduction in nutrient resorption efficiency would also mean that plants would have to increase their nutrient intake from the soil, which would be more costly in low-nutrient environments [38]. This is detrimental to the nutritional economics of trees because it can shorten leaf lifetime, limit photosynthetic capacity, and lessen internal nutrient cycling [56]. Furthermore, the current season's slower growth will have a cumulative effect on the upcoming years as nutrient reserves in perennial tissues are depleted [57]. Boercher [58] reported that infection of Arabidopsis thaliana leaves with Pseudomonas syringae significantly reduced nutrient remobilization, which resulted in a significant fitness cost. In citrus, the effects of HLB pathogen infection on PRE varied depending on the species of citrus, such that HLB increased PRE in Citrus limon. In contrast, HLB had no significant effect on C. maxima species and substantially decreased in HLB-infected Citrus reticulata, which may account for the marked decrease in average fruit yield [25].
Overall, information on the nutrient-disease interaction is important owing to the various roles of nutrients in disease management, including forming robust cell walls, generating natural defense compounds like phytoalexins and antioxidants, fostering flavonoid production, and activating enzymes involved in phenol and lignin synthesis [16,51,59]. N stands as a vital element for mango trees, crucial in improving vigor, promoting vegetative growth, and supporting physiological development. P plays a pivotal role in cell division, developmental processes, and the creation of sugar phosphate molecules, while K aids in pathogen and insect resistance, regulating water uptake, and influencing the quality of fruits. Ca contributes to membrane stability and reinforces cell walls. It also holds particular significance during the initial growth stages following mango harvesting, preserving fruit firmness. Mg is indispensable for chlorophyll production, while S supports enzymatic functions and protein synthesis [15,52,60]. Micronutrients such as B, Cu, Fe, Mn, and Zn are crucial for overall plant health. Fe and Mn aid in chlorophyll synthesis, and Zn plays a key role in protein formation and regulating water movement in mango plants. Moreover, B facilitates cell division, ensuring effective pollination and fruit development, while Cu is vital for photosynthesis by lignifying cell walls to enable efficient carbohydrate and water transport within the plant [15,52,60,61].
5. Conclusions
This study focused on MTTD and its impact on the mineral nutrition of KP mango trees, shedding light on significant differences in nutrient concentrations between trees with low and high MTTD infection levels. HD trees had lower concentrations of essential elements such as Ca, Mg, Cu, Fe, Mn, and Zn compared to LD trees, while showing higher K levels, albeit deficient in LD trees. Nutrient imbalance was more pronounced in HD trees, and comparisons between RED and RES showed substantial differences. The results indicate the need for targeted nutrient interventions to rectify MTTD-induced imbalances and bolster mango tree health and resilience against MTTD infections.
Author Contributions
Conceptualization, C.A. and A.N.; methodology, C.A. and A.N.; software, C.A.; formal analysis, C.A. and A.N.; investigation, C.A. and A.N.; data curation, C.A.; writing—original draft preparation, C.A. and A.N.; writing—review and editing, C.A. and A.N.; visualization, C.A. All authors have read and agreed to the published version of the manuscript.
Acknowledgments
We acknowledge the management of an NT grower for providing access to the mango trees in Lambell’s Lagoon orchard and the NT DITT for the internal fund to conduct the study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- NT (Northern Territory) Farmers. NT Mangoes. Available online: https://ntfarmers.org.au/commodities/mangoes/ (accessed on 2 November 2023).
- Jabeen, A. 2016. Field and Postharvest Biology of Dendritic Spot and Stem End Rot of Mango. PhD Thesis, School of Agriculture and Food Sciences, The University of Queensland, Gatton, Queensland, Australia, 2016. [CrossRef]
- Umar, M.; McConchie, C.; Tran-Nguyen, L.; Asis, C.A.; Eyles, A.; Stanley, R; Gracie, A. Perception of Australian consumers on resin canal discouloration (RCD) in mango. Acta Hortic 2020, 1299, 13-18. [CrossRef]
- Grice, K.R.E.; Bally, I.S.E.; Wright, C.L.; Maddox, C.; Ali, A.; Dillon, N.L. Mango germplasm screening for the identification of sources of tolerance to anthracnose. Australasian Plant Pathol 2023 52, 27-41. [CrossRef]
- Giblin, F.R.; Tan, Y.P.; Mitchell, R.; Coates, L.M.; Irwin, J.A.G.; Shivas, R.G. Colletotrichum species associated with pre-and post-harvest diseases of avocado and mango in eastern Australia. Australasian Plant Pathol 2018, 47, 269–276. [CrossRef]
- Johnson, G.I.; Cooke, A.W.; Mead, A.J.; Wells, I.A. Stem end rot of mango in Australia: Causes and control. Acta Hortic 1991, 291, 288-295. [CrossRef]
- Asis, C.A.; Meschiari, L.; McConchie, C. Ionome balance analysis of mango fruit from orchard with and without resin canal discolouration. Acta Hortic 2019, 1244, 221-228. [CrossRef]
- Sakalidis, M.L.; Ray, J.D.; Lanoiselet, V.; Hardy, G.E. StJ.; Burges, T.I. Pathogenic Botryosphaeriaceae associated with Mangifera indica in the Kimberley Region of Western Australia. Eur J Plant Pathol 2011, 130, 379–391. [CrossRef]
- McTaggart, A.R.; Grice, K.R.; Shivas, R.G. First report of Vialaea minutella in Australia, its association with mango branch dieback and systematic placement of Vialaea in the Xylariales. Australasian Plant Dis Notes 2013, 8, 63–66. [CrossRef]
- Umar, U.D.; Ahmed, N.; Zafar, M.Z.; Rehman, A.; Naqvi, S.A.H.; Zulfiqar, M.A.; Malik, M.T.; Ali, B.; Saleem, M.H.; Marc, R.A. Micronutrients foliar and drench application mitigate mango sudden decline disorder and impact fruit yield. Agron 2022, 12, 2449. [CrossRef]
- Saeed, A.; Shad, M.A.; Nawaz, H.; Shafqat, M.N.; Muneer, Z.; Shaheen, A.; Shah, S.T.A. Quick decline disease disturbs the levels of important phytochemicals and minerals in the stem bark of mango (Mangifera indica). Int J Agron 2016, 8219356. [CrossRef]
- Saeed, E.E.; Sham, A.; AbuZarqa, A.; Al Shurafa, K.A.; Al Naqbi, T.S.; Iratni, R.; El-Tarabily, K.; AbuQamar, S.F. Detection and management of mango dieback disease in the United Arab Emirates. Int J Mol Sci 2017, 18, 2086. PMID: 29053600; PMCID: PMC5666768. [CrossRef]
- Kamil, F.H.; Saeed, E.E.; El-Tarabily, K.A.; Abu Qamar, S.F. Biological control of mango dieback disease caused by Lasiodiplodia theobromae using stretomycete and non-streptomycete Actinobacteria in the United Arab Emirates. Front Microb 2018, 9, 829. [CrossRef]
- NT Rural Review. 2021. Mango common dieback. Available online https://industry.nt.gov.au/publications/primary-industry-publications/newsletters/regional-newsletters/rural-review/nt-rural-review-november-2021/mango-common-dieback (accessed on 5 November 2023).
- NT Rural Review. 2021. Mango twig tip dieback. Available online https://industry.nt.gov.au/publications/primary-industry-publications/newsletters/regional-newsletters/rural-review/nt-rural-review-november-2021/mango-twig-tip-dieback (accessed on 5 November 2023).
- Ram, R.A.; Rahim, M.A.; Alam, M.S. Diagnosis and management of nutrient constraints in mango. In Fruit Crops: Diagnosis and Management of Nutrient Constraints, Srivastava, A.K., Hu, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 12, pp. 629–650. [CrossRef]
- Tripathi, R.; Terwai, R.; Sing, K.P.; Keswani, C.; Minkina, T.; Srivastava, A.K.; De Corato, U.; Sansinenea, E. Plant mineral nutrition and disease resistance: A significant linkage for sustainable crop protection. Front Plant Sci 2022, 13, 883970. [CrossRef]
- Kumar, R.; Kumar, V. Physiological disorders in perennial woody tropical and subtropical fruit crops: A review. Indian J Agric Sci 2016, 86, 703–17.
- Dordas, C. Role of nutrients in controlling plant diseases in sustainable agriculture. A review. Agron Sustain Dev 2008, 28, 33–46. [CrossRef]
- Gupta, N.; Debnath, S.; Sharma, S.; Sharma, P.; Purohit, J. Role of nutrients in controlling plant diseases in sustainable agriculture. In Agriculturally Important Microbes for Sustainable Agriculture, Meena, V.S., Mishra, P., Bisht, J., Pattanayak, A. Eds., Springer, Singapore. 2017; Volume 2, pp. 217-2162. [CrossRef]
- Gonzalez de Andrés, E.; Gazol, A.; Querejeta, J.I.; Igual, J.M.; Colangelo, M.; Sánchez-Salguero, R.; Linares, J.C.; Camarero, J.J. The role of nutritional impairment in carbon-water balance of silver fir drought-induced dieback. Glob Chang Biol 2022, 28, 4439-4458. [CrossRef]
- Bal, T.L.; Storer, A.J.; Jurgensen, M.F.; Doskey, P.V.; Amacher, M.C. Nutrient stress predisposes and contributes to sugar maple dieback across its northern ranges: A review. Forestry 2015, 18, 64-83. [CrossRef]
- Akillioglu, A. Micronutrient levels of cherry orchards in the Aegean region of Turkey. Acta Hortic 1997, 448, 199-202. [CrossRef]
- Nageli, P.; Grant, J.; Nichols, J.D.; Sheil, D.; Horton, B. Bell miner associated dieback: Nutrient cycling and herbivore crown damage in Eucalytus propinqua. Austral For 2016, 79, 74-82. https://10.1080/00049158.2015.1123565.
- Gerrish, G.; Dombois, D.M.; Bridges, K.W. Nutrient limitation and Metrosideros forest dieback in Hawai’i. Ecol 1988, 69, 723-727. [CrossRef]
- Cao, J.; Cheng, C.; Yang, J.; Wang Q. Pathogen infection drives patterns of nutrient resorption in citrus plants. Sci Rep 2015, 5, 14675. [CrossRef]
- Huang, C.; Xu, C.; Ma, Y.; Song, T.; Xu, Z.; Li, S.; Liang, J.; Zhang, L. Nutritional diagnosis of the mineral elements in Tainong mango leaves during flowering in karst areas. Land 2022, 11, 1311. [CrossRef]
- Sarkhosh, A.; Shahkoomahally, S.; Asis, C.; McConchie, C. Influence of rootstocks on scion leaf mineral content in mango tree (Mangifera indica L.). Hortic Environ Biotechnol 2021, 62, 725–735. [CrossRef]
- Ganeshamurthy, A.N.; Reddy, Y.T.N. Fitness of mango for colonization in low fertility soils and dry lands: Examination of leaf life-span, leaf nutrient resorption, and nutrient use efficiency in elite mango varieties. Agric Res 2015, 4, 254–260. [CrossRef]
- Reuter, D.J.; Robinson, J.B. Plant Analysis, an Interpretation Manual, 2nd ed.; CSIRO Publishing, Collingwood, Australia, 1997; pp. 366-367.
- Heras, L.; Montañés, L. Desviación del óptimo porcentual (DOP): Nuevo índice para la interpretación del análisis vegetal. An Estac Exp Aula Dei 1991, 20, 93–108.
- Kumar, P.; Sharma, S.K.; Kumar, A. Foliar nutritive fluids affect generative potential of apples: Multilocation DOP indexing and PCA studies under dry temperate agro-climatic conditions of north-west Himalaya. Sci Hortic 2017, 218, 265–274. [CrossRef]
- Zarrouk, O.; Gogorcena, Y.; Gómez-Aparisi, J.; Betrán, J.; Moreno, M. Influence of almond× peach hybrids rootstocks on flower and leaf mineral concentration, yield and vigour of two peach cultivars. Sci Hortic 2005, 106, 502–514. [CrossRef]
- Zhang, Y., Yang, J., Wei, X., Ni, X., & Wu, F. Monthly dynamical patterns of nitrogen and phosphorus resorption efficiencies and C: N: P stoichiometric ratios in Castanopsis carlesii (Hemsl.) Hayata and Cunninghamia lanceolata (Lamb.) Hook. Plantations. Forests 2022, 13, 1458.
- Montañés, L.; Heras, L.; Abadía, J.; Sanz, M. Plant analysis interpretation based on a new index: Deviation from optimum percentage (DOP). J Plant Nutr 1993, 16, 1289–308. [CrossRef]
- Tadayon, M.S.; Sadeghi, S. The relation between compositional nutrient diagnosis indices and susceptibility of Valencia orange to citrus decline. J Plant Nutr 2023, 46, 261-274. [CrossRef]
- Milošević, T., Moreno, M.Á., Milošević, N., Milinković, M. Regulation of yield, fruit size, and leaf mineral nutrients of the ‘Šumadinka’ sour cherry cultivar with help of rootstocks. J Plant Growth Regul 2023, 42, 5587–5599. [CrossRef]
- Wright, I.J.; Westoby, M. Nutrient concentration, resorption and lifespan: Leaf traits of Australian sclerophyll species. Funct Ecol 2003, 17, 10–19. [CrossRef]
- Prieto, I.; Querejeta, J.I. Simulated climate change decreases nutrient resorption from senescing leaves. Glob Change Biol 2020, 26, 1795-1807.
- Zhang, M.; Zhang, L.; Yao, X.; Li, J.; Deng, Q. Co-evaluation of plant leaf nutrient concentrations and resorption in response to fertilization under different nutrient-limited conditions. Diversity 2022, 14, 385. [CrossRef]
- Smith, S.; Hill, J. Supporting sustainable development–risks and impacts of plant industries on soil condition. Tech Bull 2011, 340, 1-27. https://industry.nt.gov.au/__data/assets/pdf_file/0005/233258/tb340.pdf.
- Spann, T.M.; Schumann, A.W. The role of plant nutrients in disease development with emphasis on citrus and huanglongbing. Proc Fla State Hort Soc 2009, 22, 169-171. https://swfrec.ifas.ufl.edu/hlb/database/pdf/00001871.pdf.
- Azim Nejad, Z.; Badehian, Z.; Rezaei Nejad, A.; Bazot, S. Do soil properties and ecophysiological responses of aak (Quercus brantii Lindl.) correlate with the rate of dieback? Trees 2021, 35, 1639–1650. [CrossRef]
- Pustika, A.B.; Subandiyah, S.; Holford, P.; Beattie, G.A.C.; Iwanami, T.; Masaoka, Y. Interactions between plant nutrition and symptom expression in mandarin trees infected with the disease huanglongbing. Australas Plant Dis Notes 2008, 3, 112-115. [CrossRef]
- Mathew, J.; Haris, A.A.; Bhat, R.; Kumar, V.K.; Muralidharan, K.; John, K.S.; Surendran, U. A comparative assessment of nutrient partitioning in healthy and root (wilt) disease affected coconut palms grown in an Entisol of humid tropical Kerala. Trees 2021, 35, 621-635. [CrossRef]
- Sousa Filho, H.R.; Jesus, R.M.D.; Bezerra, M.A.; da Silva, V.H.; da Silva Jr, A.L.; Alves, J.P.; Santana, G; Souza Jr, J.O.D. Mineral nutrients and plant-fungal interaction in cocoa trees (Theobroma cacao L.). J Braz Chem Soc 2021, 32, 337-346. [CrossRef]
- Soares, T.P.; Pozza, E.A.; Pozza, A.A.; Mafia, R.G.; Ferreira, M.A. Calcium and potassium imbalance favours leaf blight and defoliation caused by Calonectria pteridis in Eucalyptus plants. Forests 2018, 9, 782. [CrossRef]
- Pervez, H.; Ashraf, M.; Makhdum, M.I.; Mahmood, T. Potassium nutrition of cotton (Gossypium hirsutum L.) in relation to cotton leaf curl virus disease in aridisols. Pak J Bot 2007, 39, 529-539. https://www.pakbs.org/pjbot/PDFs/39(2)/PJB39(2)529.pdf.
- Graça, R.N.; Alfenas, A.C.; Maffia, L. A.; Titon, M.; Alfenas, R.F.; Lau, D.; Rocabado, J.M.A. Factors influencing infection of eucalypts by Cylindrocladium pteridis. Plant Pathol 2009, 58, 971-981. [CrossRef]
- Maity, A.; Sharma, J.; Sarkar, A.; More, A.K.; Pal, R.K. Nutrient imbalance indices are closely related with susceptibility of pomegranate to bacterial blight disease. Sci Hortic 2016, 211, 79-86. [CrossRef]
- Gopi, R.; Madhavi, G.B.; Kapoor, C.; Raj, C.; Singh, S.; Ramprakash, T. Role of mineral nutrients in the management of plant diseases. In Plant Disease Management Strategies, Nehra, S., Trivedi, P.C., Ed., Agrobios Research: Rajasthan, India, 2021; Chapter 4, pp. 87-117.
- Weinmann, M.; Bradáčová, K.; Nikolic, M.. Relationship between mineral nutrition, plant diseases, and pests. In Marschner's Mineral Nutrition of Higher Plants, 4th ed.; Marschner, P., Ed.; Elsevier/Academic Press: Amsterdam, Netherlands, 2023; Chapter 10, pp. 445-476. [CrossRef]
- Maillard, A.; Diquélou, S.; Billard, V.; Laîné, P.; Garnica, M.; Prudent, M.; Garcia-Mina, J.M.; Yvin ,J.C.; Ourry, A. Leaf mineral nutrient remobilization during leaf senescence and modulation by nutrient deficiency. Front Plant Sci 2015, 6, 317. [CrossRef]
- Etienne, P.; Diquelou, S.; Prudent, M.; Salon, C.; Maillard, A.; Ourry, A. Macro and micronutrient storage in plants and their remobilization when facing scarcity: The case of drought. Agriculture, 2018, 8, 14. [CrossRef]
- Achat, D.L.; Pousse, N.; Nicolas, M.; Augusto, L. Nutrient remobilization in tree foliage as affected by soil nutrients and leaf life span. Ecol Monogr 2018, 88, 408-428. [CrossRef]
- McIntire, C.D.; Huggett, B.A.; Dunn, E.; Munck, I.A.; Vadeboncoeur, M.A.; Asbjornsen, H. Pathogen-induced defoliation impacts on transpiration, leaf gas exchange, and non-structural carbohydrate allocation in eastern white pine (Pinus strobus). Trees 2021, 35, 357-373. [CrossRef]
- Gortari, F.; Guiamet, J.J.; Cortizo, S.C.; Graciano, C. Poplar leaf rust reduces dry mass accumulation and internal nitrogen recycling more markedly under low soil nitrogen availability, and decreases growth in the following spring, Tree Physiol 2019, 39, 19–30. [CrossRef]
- Boercker, M.A. The Effects of Pathogen Infection on Nitrogen Remobilization in Arabidopsis thaliana. Master's Thesis, University of Tennessee, Knoxville, Tennessee, USA, December 2006. https://trace.tennessee.edu/utk_gradthes/1506.
- Rahman, A.; Albadrani, G.M.; Waraich, E.A.; Awan, T.H.; Yavaş, İ.; Hussain, S. Plant secondary metabolites and abiotic stress tolerance: Overview and implications. In Plant Abiotic Stress Responses and Tolerance Mechanisms, Hussain, S., Hussain Awan, T., Ahmad Waraich, E., Iqbal Awan, M., Eds.; IntechOpen: Rijeka, Croatia, 2023; Chapter 3, pp. 133-206. [CrossRef]
- McMahon, P. Effect of nutrition and soil function on pathogens of tropical tree crops. In Plant Pathology, Cumagon, C.J.R., Ed., IntechOpen: Rijeka, Croatia, 2012; Chapter 10, pp. 241–272. https://www.intechopen.com/chapters/34847.
- Ahmed, N.; Zhang, B.; Chachar, Z.; Li, J.; Xiao, G.; Wang, Q.; Faisal Hayat, F.; Deng, L.; Narejo, M.; Bozdar, B.; Tu, P. Micronutrients and their effects on horticultural crop quality, productivity and sustainability. Sci Hortic 2024, 323, 112512. [CrossRef]
Figure 1.
Photographs of MTTD infection symptoms in KP mango show a dark longitudinal lesion on one side of the last twig flush (a, c, d) with dried levels and green leaves on the unaffected side (a) that later dried out at the advanced stage (b).
Figure 1.
Photographs of MTTD infection symptoms in KP mango show a dark longitudinal lesion on one side of the last twig flush (a, c, d) with dried levels and green leaves on the unaffected side (a) that later dried out at the advanced stage (b).


Table 1.
Mean ± standard deviation (n = 5) of the soil properties of the commercial mango orchard with varying levels of twig-tip dieback infection, Lambells Lagoon, Litchfield municipality, Northern Territory. The ns indicates that there is no significant difference in the soil properties collected from LD and HD trees at the 5% level of significance.
Table 1.
Mean ± standard deviation (n = 5) of the soil properties of the commercial mango orchard with varying levels of twig-tip dieback infection, Lambells Lagoon, Litchfield municipality, Northern Territory. The ns indicates that there is no significant difference in the soil properties collected from LD and HD trees at the 5% level of significance.
| Soil Properties | Dieback Level | p value | |
|---|---|---|---|
| LD | HD | ||
| Soil pH (H2O) | 7.36±0.13 | 7.26±0.05 | 0.49 ns |
| Organic C (%) | 1.06±0.11 | 0.93±0.04 | 0.33 ns |
| EC (dS/m) | 0.04±0.008 | 0.03±0.003 | 0.46 ns |
| N (mg/kg) | 1.23±0.36 | 2.42±0.39 | 0.06 ns |
| P (mg/kg) | 10.80±1.93 | 12.6±2.44 | 0.57 ns |
| S (mg/kg) | 4.64±1.71 | 5.22±1.42 | 0.80 ns |
| K (cmol(+)/kg) | 0.16±0.04 | 0.11±0.01 | 0.23 ns |
| Ca (cmol(+)/kg) | 4.01±0.55 | 3.66±0.31 | 0.59 ns |
| Mg (cmol(+)/kg) | 2.70±0.52 | 2.45±0.27 | 0.68 ns |
| B (mg/kg) | 0.22±0.02 | 0.21±0.01 | 0.94 ns |
| Cu (mg/kg) | 2.19±0.20 | 2.44±0.31 | 0.54 ns |
| Fe (mg/kg) | 14.84±0.75 | 16.26±0.74 | 0.21 ns |
| Mn (mg/kg) | 6.74±2.38 | 4.97±0.66 | 0.49 ns |
| Zn (mg/kg) | 8.26±4.37 | 0.93±0.04 | 0.33 ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



