
Submitted:
15 May 2024
Posted:
16 May 2024
You are already at the latest version
Abstract
Dinoflagellates belonging to Hematodinium are important parasites of marine crustaceans, primarily dec-apods. In this study, we document the first report of H. perezi Chatton & Poisson, 1930 on the African Atlantic coast. This is also the first parasite record in the invasive American blue crab Callinectes sapidus Rathbun, 1896 in Morocco. Specimens of C. sapidus were sampled in winter 2023 from two Ramsar sites on the Atlantic coast of Morocco, namely Merja Zerga and Oualidia Lagoons, and were screened to detect the presence of parasites in their hemolymph. Based on staining fresh hemolymph smears, we did not de-tect Hematodinium in any of the 36 investigated individuals (20 and 16 from Merja Zerga and Oualidia lagoons, respectively), probably due to methodological artifacts. The PCR-based method was revealed to be more accurate in diagnosing the Hematodinium parasite. It showed that 13 blue crabs were infected by the parasite at Merja Zerga Lagoon (prevalence 65%) and four individuals at Oualidia Lagoon (25%). Ge-netic analysis, based on the ITS1 rDNA gene from Hematodinium, confirmed the sequences as being those of Hematodinium perezi.
Keywords:
bioinvasion
; parasites
; ITS1 rDNA
; coastal lagoon
; North Africa
1. Introduction
The American blue crab Callinectes sapidus Rathbun, 1896, belonging to Portunidae, is native to the estuaries and coastal waters of the western Atlantic Ocean [1,2]. Its natural geographical distribution ranges from Nova Scotia in Canada down to northern Argentina, including Bermuda and the Antilles [1,3,4]. This species was accidentally or intentionally introduced into Asia, Europe and Africa, and this widened the current biogeographic distribution of the species worldwide. The first record from European waters was in 1900 along the Atlantic coast of France [5]. In the Mediterranean Sea, blue crab first occurred in the Aegean Sea between 1935 and 1945, but its first confirmed and documented presence was reported from Italy in 1949 and was commonly ascribed to [6]. To date, the species has been recorded almost ubiquitously in the Mediterranean and the Black Sea [1,7,8,9] where it is classified as one of the 100 “worst invasive” species [10]. While ballast water is considered the most likely introduction pathway for this species, some of these introductions have resulted from deliberate releases [1,6]. In Morocco, C. sapidus was first reported on its Mediterranean coast, in the Marchica coastal lagoon in 2017 [11] while its first record on the Atlantic coast of Morocco was in the Merja Zerga Lagoon in 2019 [12]. The species is now established in many localities along the Mediterranean and Atlantic coasts of Morocco [12,13,14].
Invasive alien species are considered to be a severe threat to marine biodiversity and functioning of invaded coastal and marine ecosystems [15]. In addition, they pose a risk as possible carriers of invasive parasites and/or enhancers for native parasites, which can lead to loss of native biodiversity, disease and mortality of indigenous species, and risks to human health and the economy [16,17,18]. The establishment of introduced hosts and their parasites may also impact native parasite life cycles. Native hosts could contract parasites carried with an introduced species; i.e. ‘host switching’, more specifically, spill-over [17,19,20,21,22]. For example, the invasive nematode Spirocamallanus istiblenni Noble, 1966, was introduced to the Hawaiian archipelago along with Lutjanus kasmira Forsskål, 1775 from French Polynesia, and this introduction led to the spread of the nematode into native hosts [23]. The monogenean Nitzschia sturionis (Abildgaard, 1794) Krøyer, 1852 was co-introduced with Acipenser stellatus Pallas, 1771 from the Caspian Sea to the Aral Sea and caused severe mass mortalities of the native bastard sturgeon Acipenser nudiventris Lovetsky, 1828. The parasite infected the gills of the fish, causing the fish to die on the beaches [24]. The eel nematode Anguillicoloides crassus (Kuwahara, Niimi & Itagaki, 1974) Moravec & Taraschewski, 1988 has infected seven different eel species on four different continents via the global eel trade, causing severe damage to the swim bladder of the eels, which resulted in significant mortality in most eel species [25,26,27,28]. The rhizocephalan barnacle Loxothylacus panopaei Gissler, 1884, which infects native flatback mud crabs Eurypanopeus depressus Smith, 1869, was probably co-introduced with infected mud crabs (species unknown) in batches of oysters transferred from the Gulf of Mexico to Chesapeake Bay (North America) [29].
In native habitats, C. sapidus is known to host a wide range of pathogens, including viral, bacterial and microalgal agents [30], which have been implicated in causing diseases, mortalities or reducing fecundity [31]. Along invaded Mediterranean and north-eastern Atlantic coasts, although the blue crab constitutes an important model organism for studying invasions related to the spread of microorganisms, the majority of scientific research is focused on the population dynamics, structure, and fisheries of the species [32]. The occurrence of parasites in the species has been very little explored, even though it is an appreciated shellfish product in many countries such as Greece and Turkey, Italy, and Egypt [33].
Among pathogens of C. sapidus, the parasitic dinoflagellates belonging to Hematodinium (Dinophyceae: Syndiniales: Syndiniaceae) are an important group of disease-causing agents infecting over 40 species of crustaceans worldwide [34,35,36]. This parasite is known to cause bitter crab disease in a number of crab species such as Chionoecetes opilio Fabricius, 1788 and Chionoecetes bairdi Rathbun, 1924 [37], resulting in a crab meat flavor resembling bitterness with an aspirin-like taste due to biochemical alterations [34]. Also due to the massive proliferation of parasitic cells in the hemolymph of affected tissues, Hematodinium-infected hosts generally experience malfunction or dysfunction of major tissues or organs, and consequent mortality in the late stages of infection [38,39,40]. Despite not causing biochemical alteration in blue crabs, it does cause mass mortalities [41].
In this study, hemolymph of C. sapidus specimens from Merja Zerga and Oualidia coastal lagoons, located on the Atlantic coast of Morocco, was screened to detect the presence of parasites. Here, we document the first detection of dinoflagellates belonging to Hematodinium in C. sapidus on the African Atlantic coast.
2. Materials and Methods
2.1. Study Sites
Merja Zerga and Oualidia Lagoons are two semi-enclosed coastal systems (SECS) situated along the Atlantic coast of Morocco (Figure 1). Both sites are recognized as Site of Biological and Ecological Interest (SIBE) [42] and Ramsar sites as Wetlands of International Importance.
The Merja Zerga Lagoon (34°47′N–6°13′W) is an elliptically shaped lagoon of 9 km long and 5 km wide, with a depth from 0.50 to 1.50 m for a total surface of 35 km2 [43]. The lagoon is connected with the Atlantic Ocean through a narrow, sinuous and relatively deep gully (up to 6 m), which branches out in the direction of the lagoon into shallow subtidal channels, ensuring the circulation of water during the flood and ebb. The freshwater supply is provided by two rivers: Canal of Nador in the south and Oued Drader in the northeast of the lagoon [44]. Tides are semi-diurnal, with an average amplitude of 0.15 to 1.50 meters [45]. Salinity in this lagoon fluctuates between 8 PSU and 36 PSU, with mean water temperature varying between 14.6°C and 24.15°C [46].
The Oualidia Lagoon (32°74′N–9°03′W) is over 7 km long and 1 km wide, with a mean depth of 2 m for a total surface of 3 km2 [43]. This coastal basin is made up by the morphology of a depression, in a north-south direction, limited by a continental cliff and by a coastal consolidated dune ridge [47]. Tides are semi-diurnal, with amplitudes ranging from 0.8 to 3.6 meters [48]. The average water temperature ranges from 16.1°C to 21.1°C, and the lagoon's salinity varies between 20 PSU and 35 PSU at low tide, while at high tide it can reach 30 PSU to 36 PSU throughout the year [49].
2.2. Sampling and Microscopic Analysis
Blue crabs were collected from Merja Zerga (February 2023) and Oualidia (March 2023) Lagoons using a seine net. Crabs were transferred to the laboratory in refrigerated containers; we immersed them in ice water for about 15 to 20 minutes to induce anesthesia and reduce mobility and sensitivity to pain. Captured crabs were identified and numbered, measured for their carapace length (CL), carapace width (CW) and fresh weight (W). Sex and maturity were determined for each crab.
The hemolymph was extracted from each specimen from the right swimming leg (based on dorsal view) at the uncalcified joint near the carapace which was cleaned with a cotton swab and 70% ethanol. To ensure complete sterilization of the joint, the leg was sterilized twice. A disposable 1ml syringe coupled to a 26-G needle was inserted into the leg. The hemolymph of each specimen was analysed by preparation of wet smears, one drop of hemolymph was mixed (1:1) with 0.3% neutral red solution on a glass slide and directly observed under optical microscope Leica® DM 2500. The hemolymph was also collected and placed in EDTA tubes containing 95% ethanol and frozen at -20 °C for DNA extraction to detect the presence of Hematodinium by polymerase chain reaction (PCR).
2.3. DNA Extraction, Amplification and Sequencing
Prior to DNA extraction, 200 µl of ethanol-preserved hemolymph was centrifuged at 1500 g for 1 min and the excess ethanol removed [50]. Samples were dried for at least 30 min to allow residual ethanol to evaporate [50]. DNA was extracted using Invitrogen TM Kit Blood and Tissue kits per the manufacturer’s recommendations, with overnight lysis of hemolymph samples, and two 5-minute elution incubations and two 50 µl elutions.
For amplification, HITS1F and HITS1R primers that target the ITS1 rDNA were used according to [51], with the expected length of 299 bp [52].
The PCR reaction was performed in a final volume of 25µl containing: 5 µl Taq (Gquence) reaction buffer, 1.5 mM MgCl2, 0.1 mM dNTPs, 0.5 µM HITS1F (5′CATTCACCGTGAACCTTAGCC3′), 0.5 µM HITS1R (5′CTAGTCATACGTTTGAAGAAAGCC3′), 1 unit of Platinum™ Taq DNA Polymerase and 5 µl of extracted DNA [50]. The mix was brought to the final volume with Nanopure water. Amplification reactions were performed in a thermal gradient PCR MultiGene OptiMax Thermal Cycler according to the following program: 95.0 °C for 10 min; 40 cycles of 94.0 °C for 30 s, 56.0 °C for 30 s, 72.0 °C for 1 min; and a final extension at 72.0 °C for 10 min. Amplified products were separated by 1% agarose gel electrophoresis stained with ethidium bromide. Successful PCR products were sent to the National Centre for Scientific and Technical Research (CNRST) in Rabat for purification and sequencing. The sequencing was carried out using a Genomix sequencer (MGX) and the same forward and reverse primers as for the PCR.
The obtained sequences were cleaned manually with MEGA version XI [53] and aligned using the computer program ClustalW [54], running in MEGA version XI. Sequences obtained were identified by the Basic Local Alignment Search Tool (BLAST) [55]. Genetic distances (uncorrected – ‘p’) between Moroccan sequences and all published sequences generated from GenBank were calculated using MEGA version XI. Sequences generated from this study were deposited to GenBank under accession numbers xxxxxxxx-xx (will be made available after revision).
3. Results
3.1. Biometric Characteristics of Analysed Specimens of Callinectes sapidus
Overall, 36 C. sapidus specimens were collected in winter 2023. The 20 specimens from Merja Zerga Lagoon were five adult females, five adult males, five female juveniles and five male juveniles. Among the 16 specimens from Oualidia Lagoon, there were four adult females, three adult males, five female juveniles and four male juveniles. Their biometric characteristics are summarized in Table 1.
3.2. The Hemolymph Smear Assay with Neutral Red
Based on staining fresh hemolymph smears, we did not detect Hematodinium in any of the 36 investigated individuals (20 and 16 from Merja Zerga and Oualidia lagoons, respectively) sampled in winter 2023.
3.3. PCR-Based Method and Sequence Analysis
Overall, 17 samples out of the 36 individuals investigated were successfully amplified, revealing that 13 blue crabs were infected by the parasite at Merja Zerga Lagoon (prevalence 65%) and four individuals at Oualidia Lagoon (25%).
15 ITS1 rDNA sequences were generated (13 sequences from Merja Zerga Lagoon and two from Oualidia Lagoon), and the alignment was 295 bp in length. The comparison of ITS1 rDNA sequences with those already present in the GenBank database using BLAST search confirmed the identification of our parasites’ species sequences as being those of Hematodinium perezi, sharing 98.99% of similarity with the sequence having the accession number EF065716, 98.65% with EF065708 and EF065711, all of them from the host Liocarcinus depurator Linnaeus, 1758 (Polybiidae), harbour crab, collected from Rye Bay on the South Coast of England [56], and 98.32% with the sequence with accession number PP056127 from the host Callinectes sapidus Rathbun, 1896 (Portunidae), American blue crab, collected from Greece [33].
The uncorrected p-distances between Moroccan sequences varied between 0% and 0.6%, and the uncorrected p-distances between Moroccan sequences and published sequences generated from GenBank [33,56,57] (Table 2), varied between 0.6% from H. perezi in Liocarcinus depurator collected off the South Coast of England and 4% from H. perezi in Callinectes sapidus from the United States of America (Table 3).
The measurements of specimens of Callinectes sapidus parasitized by Hematodinium perezi (as identified using PCR) from Merja Zerga Lagoon and Oualidia Lagoon of Morocco are reported in Table 4. All the four groups (male adult, female adult, male juveniles and female juveniles) were parasitized.
4. Discussion
The first report of Hematodinium perezi (Dinophyceae: Syndiniales) on the African Atlantic coast, which also represents the first parasite recorded in the invasive blue crab Callinectes sapidus in Morocco, has been documented in the present study.
The blue crab is known to be infected by two parasitic dinoflagellates belonging to the genus Hematodinium [58], namely Hematodinium sp. and H. perezi. Hematodinium sp. was reported for the first time in blue crab C. sapidus sampled from coastal areas of North Carolina, Georgia, and Florida (USA) [59], and many habitats in the USA [50,51,60,61,62,63,64,65,66], and in Adana, Turkey [67]. Hematodinium perezi was first described from Carcinus maenas Linnaeus, 1758 and Liocarcinus depurator from the French coastline [68]. Since then, the number of host species and distribution records have increased gradually in the past decades [69]. Gallien [70] reported its spread in French host Portunus latipes Pennant, 1777. In the Mid-Atlantic, it showed rare infections in the cancer crabs Cancer irroratus Say, 1817 and Cancer borealis Stimpson, 1859, and in the portunid crab Ovalipes oceallatus Herbst, 1799 from the New York bight area of the northeastern United States [71]. Hematodinium perezi was documented in the American blue crab Callinectes sapidus for the first time in coastal Maryland and Virginia, USA [72] and afterwards in many habitats in the USA [62,73,74,75,76,77]. In the Mediterranean and north-eastern Atlantic, regions where C. sapidus invaded, H. perezi was reported as endoparasite of C. sapidus in the eastern Mediterranean, Greece [33] and now also on the African Atlantic coast (this study) (Figure 2).
Hematodinium is considered to be a generalist parasite [34,56,69], reflecting its host range as well as its ability to switch between host species, allowing it to maintain a presence in the environment when the preferred host may become scarce. Positive infections of Hematodinium were reported in 13 crustacean species belonging to two orders, Decapoda and Amphipoda [78]. Several factors affect the epidemiology of Hematodinium, including environmental conditions such as salinity and temperature. It is known that this parasite tends to infect hosts in high salinity waters [79]. For example, in Europe (Wadden Sea), no detection of Hematodinium in 1252 individuals of eight crustacean species from six sites was reported due to lower salinity [78]. Epidemics of these parasites have damaged commercial stocks of Nephrops norvegicus (Linnaeus, 1758) langoustines, Chionoecetes opilio (Fabricius, 1788) snow crabs, Chionoecetes bairdi Rathbun, 1924 Tanner crabs, C. sapidus American blue crabs and Necora puber (Linnaeus, 1767) velvet swimming crabs [34]. Moreover, it is thought to affect fisheries and host populations in a way that is similar to how viral diseases of crustaceans affect fisheries and host populations [56], as they cause significant mortality in the host [80].
In the present research, diagnosis of infection by representatives of Hematodinium in C. sapidus was performed using the fresh hemolymph smear essay with neutral red and molecular analysis (PCR-based method and sequencing). The hemolymph smear assay with neutral red has been used as an initial assessment tool due to its cost-effective and time-efficient diagnostic method for detection of members of Hematodinium [78] as well as to its specificity and sensitivity [81]. The active absorption of neutral red by Hematodinium lysosomes results in distinct staining, providing visual contrast to host hemocytes [34]. In our case, based on staining fresh hemolymph smears, we did not detect Hematodinium in any of the 36 investigated individuals (20 and 16 from Merja Zerga and Oualidia lagoons, respectively), probably due to methodological artifact. Indeed, as smears are rated positive when abnormal cells, i.e., cells that could not be identified as crab hemocytes that were consistent with Hematodinium in some features, are encountered, it requires expertise in parasite identification [82]. Some pathogens of crustaceans such as parasitic dinoflagellates and rhizocephalans may be more difficult to identify for the non-specialist [83]. Moreover, certain stages of Hematodinium parasites can be very difficult to detect in fresh hemolymphs smears, because the trophic stages resemble hemocytes. The vermiform plasmodium (cf. filamentous trophont) is the most easily identified form, but the vegetative, amoeboid stage is that most frequently observed and to the uninitiated, is easily confused with a hemocyte and they may be present at relatively low densities, making microscopic diagnosis difficult [34,63].
The use of molecular analysis has become increasingly widespread in pathogen identification and disease detection and monitoring and is particularly useful in the diagnosis of cryptic organisms such as dinoflagellates and parasitic stages [51,84,85,86]. The PCR-based method offers 1000 times greater sensitivity than histological methods. The sensitivity of PCR diagnosis has been estimated at one parasite cell in 300,000 crab hemocytes [51]. Use of the PCR test eliminates the need for visual identification of cells with ambiguous characteristics. Overall, a combination of morphological and molecular characterization is often used to ensure accurate detection and monitoring of Hematodinium infections in crustacean populations.
The PCR assay, adopted in our study to detect the Hematodinium infecting the blue crab Callinectes sapidus, based on the amplification of the parasite’s first internal transcribed spacer region (ITS1), has been developed by [63]. It was revealed to be more accurate in identifying crabs parasitized by Hematodinium than histological methods. The difference in parasite prevalence between Merja Zerga (65%) and Oualidia Lagoon (25%) could be explained by environmental factors [60,78,80] or biological factors [66,87]. According to [80], several studies have shown that temperature and salinity conditions favor the invasion of Hematodinium. The prevalence of Hematodinium decreased at lower temperatures [80], and lower salinity can also limit the distribution of the parasite [78]. For example, the highest prevalence (69%) of the parasite in blue crab collected from the USA was found at salinities of 26 PSU to 30 PSU and water temperature >25 °C and no infected crabs were found in salinities below 11 PSU [60]. Hematodinium may also be more prevalent due to a large variety of environmental reservoirs and high densities of hosts [87], and the absence of host immune response may also be the reason of the high prevalence of Hematodinium in crustaceans [87]. Furthermore, according to [66], Hematodinium infection in blue crabs may vary due to additional factors that differ from year to year and river to river.
Calculated pairwise uncorrected p-distances from sequences of Hematodinium perezi parasitizing Callinectes sapidus in Morocco and all published sequences of Hematodinium perezi drawn from GenBank show that Moroccan sequences are closely similar to those from Licocarcinus depurator collected from the South Coast of England with accession numbers (EF065716, EF065711, EF065708) and to that with accession number PP056127 from the host Callinectes sapidus Rathbun, 1896 (Portunidae), American blue crab, collected from Greece, and differ more from the sequence from Callinectes sapidus collected from the USA with accession number KX758132. The uncorrected p-distances between Moroccan sequences and sequences having accession numbers (KX244637, KX244644, KX244641) from Portunus trituberculatus from China range from 1.7% to 2.3% (Table 3). According to [56], the mean interspecific genetic distances between H. perezi from L. depurator and the Hematodinium sp. infecting Portunus trituberculatus Miers, 1876 and Scylla serrata Forskål, 1775 is 0.025, and 0.046 between H. perezi from L. depurator and the Hematodinium sp. from C. sapidus. In our study, the mean intraspecific genetic distances between H. perezi from C. sapidus collected from Morocco and all published sequences of H. perezi from GenBank is 0.011. In comparison with the above-mentioned interspecific distances, this confirms that our sequences belong to parasites that are conspecific with H. perezi.
Ultimately, in-depth studies are desirable for understanding the interactions between the invasive American blue crab and its parasites in the coastal areas of Morocco and for assessing their effects on native biodiversity, associated marine diseases and risks to human health. In addition, more elaborate genetic analysis based on a large-scale sampling of native and introduced populations of the American blue crab and its associated Hematodinium parasites will provide relevant insights into the invasion history of the American blue crab in Morocco.
Author Contributions
Conceptualization, A.L.; I.R.; N.K.; M.P.M.V and H.B.; methodology, A.L.; I.R.; N.K.; M.P.M.V and H.B.; software, A.L and I.R.; formal analysis, A.L.; I.R.; H.B.; N.K and M.P.M.V.; investigation, A.L.; I.R.; M.S and A.H.; resources, A.L.; M.S.; A.H and H.B.; data curation, A.L.; I.R.; N.K.; M.P.M.V and H.B.; writing—original draft preparation, A.L.; writing—review and editing, A.L.; I.R.; H.B.; N.K. and M.P.M.V.; supervision, I.R.; H.B.; N.K and M.P.M.V.; funding acquisition, A.L. M.S and H.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Special Research Fund of Hasselt University (BOF23BL07 to A.L.; BOF21PD01 to N.K.; BOF20TT06 to M.P.M.V.; BOF21INCENT09), by research grant 1513419N of the Research Foundation – Flanders (FWO-Vlaanderen) and by a grant of the National Centre for Scientific and Technical Research (CNRST), Morocco.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The genetic data are available on GenBank under the accession numbers xxxxxxxx-xx (will be made available after revision).
Acknowledgments
We thank Dr. Jeffrey Shields, Professor at Virginia Institute of Marine Science, for his helpful suggestions and advices on preparing neutral red dye, and for sharing with us a video on crab bleeding and smears preparation as well as valuable documentation on the topic. Authors would to thank the National Centre of Scientific and Technical Research (CNRST) of Morocco for putting at their disposal the technical facilities of the UATRS Division.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nehring, S. Invasion history and success of the American blue crab Callinectes sapidus in European and adjacent waters. In the wrong place-alien marine crustaceans: distribution, biology and impacts. Dordrecht: Springer Netherlands. 2011, pp. 607-426. [CrossRef]
- Nehring, S. Callinectes sapidus. In: NOBANIS- Invasive Alien Species Fact Sheet. Online Database of the European Network on Invasive Alien Species, NOBAMIS. 2012. http://www.nobanis.org.
- Williams, A.B. The swimming crabs of the genus Callinectes (Decapoda: Portunidae). Fish. Bull. 1974, 72, 685–798. [Google Scholar]
- Castriota, L.; Andaloro, F.; Costantini, R.; De Ascentiis, A. First record of the Atlantic crab Callinectes sapidus Rathbun, 1896 (Crustacea: Brachyura: Portunidae) in Abruzzi waters, central Adriatic Sea. Acta Adriat. 2012, 53, 467–471. [Google Scholar]
- Bouvier, E.L. Sur un Callinectes sapidus M. Rathbun trouvé à Rocheford. Bull. Mus. Natl. Hist. Nat. 1901, 7, 16–17. [Google Scholar]
- Giordani Soika, A. II Neptunus pelagicus (L. ) nell’Alto Adriatico. Natura 1951, 42, 18–20. [Google Scholar]
- Pashkov, A.N.; Reshetnikov, S.I.; Bondarev, K.B. The capture of the blue crab (Callinectes sapidus, Decapoda, Crustacea) in the Russian sector of the Black Sea. Russ. J. Biol. Invasions 2012, 3, 22–28. [Google Scholar] [CrossRef]
- Castejón, D.; Guerao, G. A new record of the American blue crab, Callinectes sapidus Rathbun, 1896 (Decapoda: Brachyura: Portunidae), from the Mediterranean coast of the Iberian Peninsula. BioInvasions Rec. 2013, 2, 141–143. [Google Scholar] [CrossRef]
- Öztürk, R.Ç.; Terzi, Y.; Feyzioğlu, A.M.; Şahin, A.; Aydın, M. Genetic Characterization of the Invasive Blue Crab, Callinectes sapidus (Rathbun 1896), in the Black Sea. Reg. Stud. Mar. Sci. 2020, 39, 101412. [Google Scholar] [CrossRef]
- Streftaris, N.; Zenetos, A. Alien marine species in the Mediterranean - the 100 ‘Worst Invasives’ and their impact. Mediterr. Mar. Sci. 2006, 7, 87–118. [Google Scholar] [CrossRef]
- Chartosia, N.; Anastasiadis, D.; Bazairi, H.; Crocetta, F.; Deidun, A.; Despalatović, M.; Di Martino, V.; Dimitriou, N.; Dragičević, B.; Dulčić, J.; Durucan, F.; Hasbek, D.; Ketsilis-Rinis, V.; Kleitou, P.; Lipej, L.; Macali, A.; Marchini, A.; Ousselam, M.; Piraino, S.; Stancanelli, B.; Theodosiou, M.; Tiralongo, F.; Todorova, V.; Trkov, D.; Yapici, S. New Mediterranean Biodiversity Records. Mediterr. Mar. Sci. 2018, 19, 398–415. [Google Scholar] [CrossRef]
- Oussellam, M.; Benhoussa, A.; Pariselle, A.; Rahmouni, I.; Salmi, M.; Agnèse, J.F.; Selfati, M.; El Ouamari, N.; Bazairi, H. First and southern-most records of the American blue crab Callinectes sapidus Rathbun, 1896 (Decapoda, Portunidae) on the African Atlantic coast. Bioinvasions Rec. 2023, 12, 403–416. [Google Scholar] [CrossRef]
- Chaouti, A.; Belattmania, Z.; Nadri, A.; Serrão, E.; Encarnação, J.; Teodósio, A.; Reani, A.; Sabour, B. The Invasive Atlantic Blue Crab Callinectes sapidus Rathbun 1896 Expands its Distributional Range Southward to Atlantic African Shores: First Records Along the Atlantic Coast of Morocco. BioInvasions Rec. 2022, 11, 227–237. [Google Scholar] [CrossRef]
- Chairi, H. , González-Ortegón, E. Additional records of the blue crab Callinectes sapidus Rathbun, 1896 in the Moroccan Sea, Africa. BioInvasions Rec. 2022, 11, 776–784. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Wallentinus, I.; Zenetos, A.; Leppäkoski, E.; Çinar, M.E.; Oztürk, B.; Grabowski, M.; Golani, D.; Cardoso, A.C. Impacts of marine invasive alien species on ecosystem services and biodiversity: a pan-European review. Aquat. Invasions 2014, 9, 391–423. [Google Scholar]
- Crowl, T.A.; Crist, T.O.; Parmenter, R.R.; Belovsky, G.; Lugo, A.E. The spread of invasive species and infectious disease as drivers of ecosystem change. Front. Ecol. Environ. 2008, 2008. 6, 238–246. [Google Scholar] [CrossRef]
- Goedknegt, M.A.; Feis, M.E.; Wegner, K.M.; Luttikhuize, P.C.; Buschbaum, C.; Camphuysen, K.C.J.; Vander Meer, J.; Thieltges, DW. Parasites and marine invasions: Ecological and evolutionary perspectives. J. Sea Res. 2016, 113, 11–27. [Google Scholar] [CrossRef]
- Bojko, J.; Burgess, A.L.; Baker, A.G.; Orr, C.H. Invasive Non-Native Crustacean Symbionts: Diversity and Impact. J. Invertebr. Pathol. 2021, 186, 107482. [Google Scholar] [CrossRef]
- Hall, S.R.; Becker, C.R.; Simonis, J.L.; Duffy, M.A.; Tessier, A.J.; Cáceres, C.E. Friendly competition: evidence for a dilution effect among competitors in a planktonic host–parasite system. Ecology 2009, 90, 791–801. [Google Scholar] [CrossRef]
- Kelly, D.W.; Paterson, R.A.; Townsend, C.R.; Poulin, R.; Tompkins, D.M. Parasite spillback: A neglected concept in invasion ecology? Ecology 2009, 90, 2047–2056. [Google Scholar] [CrossRef]
- Ortega, N.; Roznik, E.A.; Surbaugh, K.L.; Cano, N.; Price, W.; Campbell, T.; Rohr, J.R. Parasite spillover to native hosts from more tolerant, supershedding invasive hosts: Implications for management. J. Appl. Ecol. 2021, 59, 39–51. [Google Scholar] [CrossRef]
- Goedknegt, M.A.; Havermans, J.; Waser, A.M.; Luttikhuizen, P.C.; Vellila, E.; Camphuysen, K.C.J.; Vander Meer, J.; Thieltges, D.W. Cross-species comparison of parasite richness, prevalence, and intensity in a native compared to two invasive brachyuran crabs. Aquat. Invasions 2017, 12, 201–212. [Google Scholar] [CrossRef]
- Gaither, M.R.; Aeby, G.; Vignon, M.; Meguro, Y.I.; Rigby, M.; Runyon, C.; Bowen, B.W. An invasive fish and the time-lagged spread of its parasite across the Hawaiian archipelago. PloS ONE 2013, 8, e56940. [Google Scholar] [CrossRef]
- Dogiel, V.A.; Lutta, A.S. On the mortality of Acipenser nudiventris of the Aral Sea in 1936. J. Rybn. Choz. 1937, 12, 25–27. [Google Scholar]
- Barse, A.M.; McGuire, S.A.; Vinores, M.A.; Eierman, L.E.; Weeder, J.A. The swimbladder nematode Anguillicola crassus in American eels (Anguilla rostrata) from middle and upper regions of Chesapeake Bay. J. Parasitol. 2001, 87, 1366–1370. [Google Scholar] [CrossRef]
- Køie, M. Swimbladder nematodes (Anguillicola spp.) and gill monogeneans (Pseudoactylogyrus spp.) parasitic on the European eel (Anguilla anguilla). ICES J. Mar. Sci. 1991, 47, 391–398. [Google Scholar] [CrossRef]
- Kvach,Y. ; Skóra, K.E. Metazoan parasites of the invasive round goby Apollonia melanostoma (Neogobius melanostomus) (Pallas) (Gobiidae: Osteichthyes) in the Gulf of Gdansk, Baltic Sea, Poland: a comparison with the Black Sea. Parasitol. Res. 2007, 100, 767–774. [Google Scholar] [CrossRef]
- Sasal, P.; Taraschewski, H.; Valade, P.; Grondin, H.; Wielgoss, S.; Moravec, F. Parasite communities in eels of the Island of Reunion (Indian Ocean): a lesson in parasite introduction. Parasitol. Res. 2008, 102, 1343–1350. [Google Scholar] [CrossRef]
- Van Engel, W. A.; Dillon, W. A.; Zwerner, D.; Eldridge, D. Loxothylacus panopaei (Cirripedia, Sacculinidae) an introduced parasite on a xanthid crab in Chesapeake Bay, USA. Crustacean 1966, 10, 110–112. [Google Scholar]
- Flowers, E.M.; Simmonds, K.; Messick, G.A.; Sullivan, L.; Schott, E.J. PCR-based prevalence of a fatal reovirus of the blue crab, Callinectes sapidus (Rathbun) along the northern Atlantic coast of the USA. J. Fish Dis. 2016, 39, 705–714. [Google Scholar] [CrossRef]
- Messick, G.A.; Sindermann, C.J. Synopsis of principal diseases of the blue crab, Callinectes sapidus. NOAA Tech. Memo. 1992, Woods Hole.
- Kampouris, T.E.; Kouroupakis, E.; Batjakas, I.E. Morphometric relationships of the global invader Callinectes sapidus Rathbun,1896 (Decapoda, brachyura, portunidae) from Papapouli lagoon, NW Aegean Sea, Greece. with notes on its ecological preferences. Fishes 2020, 5, 5. [Google Scholar] [CrossRef]
- Lattos, A.; Papadopoulos, D.K.; Giantsis, I.A.; Stamelos, A.; Karagiannis, D. Histopathology and Phylogeny of the Dinoflagellate Hematodinium perezi and the Epibiotic Peritrich Ciliate Epistylis sp. Infecting the Blue Crab Callinectes sapidus in the Eastern Mediterranean. Microorganisms 2024, 12, 456. [Google Scholar] [CrossRef]
- Stentiford, G.D.; Shields, J.D. A review of the parasitic dinofagellates Hematodinium species and Hematodinium-like infections in marine crustaceans. Dis. Aquat. Org. 2005, 66, 47–70. [Google Scholar] [CrossRef]
- Shields, J.D. The impact of pathogens on exploited populations of decapod crustaceans. J. Invertebr. Pathol. 2012, 110, 211–224. [Google Scholar] [CrossRef]
- Small, H.J. Advances in our understanding of the global diversity and distribution of Hematodinium spp. significant pathogens of commercially exploited crustaceans. J. Invertebr. Pathol. 2012, 110, 234–246. [Google Scholar] [CrossRef]
- Shields, J.D.; Overstreet, R. Diseases, parasites, and others Symbionts. In The blue crab Callinectes sapidus; Kennedy, V., Cronin, L., Eds.; Maryland Sea Grant, College Park, Maryland. 2007,1, pp. 299−417.
- Field. R.; Appleton. P.; A Hematodinium-like dinoflagellate infection of the Norway lobster Nephrops norvegicus: observations on pathology and progression of infection. Dis. Aquat. Org. 1995, 22, 115–128.
- Taylor, A.; Field, R.; Parslow-Williams, P. The effects of Hematodinium sp.-infection on aspects of the respiratory physiology of the Norway lobster, Nephrops norvegicus (L.). J. Exp. Mar. Biol. Ecol. 1996, 207, 217–228. [Google Scholar] [CrossRef]
- Wheeler, K.; Shields, J.D.; Taylor, D.M. Pathology of Hematodinium infections in snow crabs (Chionoecetes opilio) from Newfoundland. Canada. J. Invertebr. Pathol. 2007, 95, 93–100. [Google Scholar] [CrossRef]
- Albalat, A.; Collard, A.; Cadam, B.; Coates, C.J.; Fox, C.J. Physiological condition, short-term survival, and predator avoidance behavior of discraded Norway lobsters (Nephrops norvegicus). J. Shellfish Res. 2012, 35, 1053–1065. [Google Scholar] [CrossRef]
- PDAPM. Plan directeur des Aires protégées du Maroc. Les sites d’intérêt biologique et écologique de domaine littoral. BCEOM/SECA, BAD, EPHE, ISR, IB. 1996, 3, 166.
- Bououarour, O.; El Kamcha, R.; Boutoumit, S.; Pouzet, P.; Maanan, M.; Bazairi, H. Effects of the Zostera noltei meadows on benthic macrofauna in North Atlantic coastal ecosystems of Morocco: spatial and seasonal patterns. Biologia 2021, 76, 2263–2275. [Google Scholar] [CrossRef]
- Boutoumit, S.; Bououarour, O.; El Kamcha, R.; Pouzet, P.; Zourarah, B.; Benhoussa, A.; Maanan, M.; Bazairi, H. Spatial patterns of macrozoobenthos assemblages in a sentinel coastal Lagoon: Biodiversity and environmental drivers. J. Mar. Sci. Eng. 2021, 9, 461. [Google Scholar] [CrossRef]
- Gam, M. Dynamique des systèmes parasites-hôte, entre trématodes digènes et coque Cerastoderma edule: Comparaison de la lagune de Merja Zerga avec le bassin d'Arcachon. Doctoral dissertation. Université Bordeaux 1, 2008.
- Labbardi, H.; Ettahiri, O.; Lazar, S.; Massik, Z.; El Antri, S. Etude de la variation spatio-temporelle des paramètres physico-chimiques caractérisant la qualité des eaux d'une lagune côtière et ses zonations écologiques : cas de Moulay Bousselham, Maroc. C R Geoscience 2005, 337, 504–514. [Google Scholar]
- Maanan, M.; Landesman, C.; Maanan, M.; Zourarah, B.; Fattal, P.; Sahabi, M. Evaluation of the anthropogenic influx of metal and metalloid contaminants into the Moulay Bousselham lagoon, Morocco, using chemometric methods coupled to geographical information systems. Environ. Sci. Pollut. Res. 2013, 20, 4729–4741. [Google Scholar] [CrossRef]
- Hilmi, K.; Orbi, A.; Lakhdar Idrissi, J. Hydrodynamisme de la lagune de Oualidia (Maroc) durant l’été et l’automne 2005. Bull. Inst. Sci. Rabat Sect. Sci. Terre 2009, 31, 29–34. [Google Scholar]
- Makaoui, A.; Idrissi, M.; Agouzouk, A.; Larissi, J.; Baibai, T.; El Ouehabi, Z.; Laamal, M.A.; Bessa, I.; Ettahiri, O.; Hilmi, K. Etat océanographique de la lagune de Oualidia, Maroc (2011-2012). Eur. Sci. J. 2018, 14, 93. [Google Scholar] [CrossRef]
- Lohan, K.M.P.; Reece, K.S.; Miller, T.L.; Wheeler, K.N.; Small, H.J.; Shields, J.D. The role of alternate hosts in the ecology and life history of Hematodinium sp., a parasitic dinoflagellate of the blue crab (Callinectes sapidus). J. Parasitol. 2012, 98, 73–84. [Google Scholar] [CrossRef]
- Gruebl, T.; Frischer, M.E.; Sheppard, M.; Neumann, M.A.; Maurer, A.N.; Lee, R.F. Development of an 18S rRNA gene-targeted PCR-based diagnostic for the blue crab parasite Hematodinium sp. Dis. Aquat. Org. 2002, 49, 61–70. [Google Scholar] [CrossRef]
- Huang, Q.; Li, M.; Wang, F.; Li, C. The parasitic dinoflagellate Hematodinium perezi infecting mudflat crabs, Helice tientsinensis, in polyculture system in China. J. Invertebr. Pathol. 2019, 166, 107229. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish,W. ; Miller,W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar]
- Small, H.J.; Shields, J.D.; Reece, K.S.; Bateman, K.; Stentiford, G.D. Morphological and Molecular Characterization of Hematodinium perezi (Dinophyceae: Syndiniales), a Dinoflagellate Parasite of the Harbour Crab, Liocarcinus depurator. J. Eukaryot. Microbiol. 2012, 59, 54–66. [Google Scholar] [CrossRef]
- Xiao, J.; Miao, X.; Li, C.; Xu, W.; Zhang, X.; Wang, Z. Genetic variations of the parasitic dinoflagellate Hematodinium infecting cultured marine crustaceans in China. Protist 2016, 167, 597–609. [Google Scholar] [CrossRef]
- Shields, J.D. Infection and mortality studies with Hematodinium perezi in blue crabs. In: Proceedings of the Blue Crab Mortality Symposium; Guillory, V., Perry, H., VanderKooy, S., Eds.; Gulf States Mar. Fish. Comm, Ocean Springs. 2001; pp. 50–60.
- Newman, M.W.; Johnson, C.A. A disease of blue crabs (Callinectes sapidus) caused by a parasitic dinofagellate Hematodinium Sp. J. Parasitol. 1975, 61, 554–557. [Google Scholar] [CrossRef]
- Messick, G.A.; Shields, J.D. Epizootiology of the parasitic dinofagellate Hematodinium sp. in the American blue crab Callinectes sapidus. Dis. Aquat. Org. 2000, 43, 139–152. [Google Scholar] [CrossRef]
- Sheppard, M.; Walker, A.; Frischer, M.E.; Lee, R.F. Histopathology and prevalence of the parasitic dinofagellate, Hematodinium sp, in crabs (Callinectes sapidus, Callinectes similis, Neopanopesayi, Libinia emarginata, Menippe mercenaria) from a Georgia estuary. J. Shellfsh Res. 2003, 22, 873–880. [Google Scholar]
- Frischer, M.; Lee, R.; Sheppard, M.; Mauer, A.; Rambow, F.; Neumann, M.; Broft, J.; Wizenmann, T.; Danforth, J. Evidence for a freeliving life stage of the blue crab parasitic dinofagelate Hematodinium sp. Harmful Algae 2006, 5, 548–557. [Google Scholar] [CrossRef]
- Small, H.J.; Shields, J.D.; Hudson, K.L.; Reece, K.S. Molecular detection of Hematodinium sp infecting the blue crab Callinectes sapidus. J. Shellfish Res. 2007, 26, 131–139. [Google Scholar] [CrossRef]
- Troedsson, C.; Lee, R.F.; Walters, T.; Stokes, V.; Brinkley. K.; Naegele, V.; Frischer, M.E. Detection and discovery of crustacean parasites in blue crabs (Callinectes sapidus) by using 18S rRNA gene-targeted denaturing high-performance liquid chromatography. Appl. Environ. Microbiol. 2008, 74, 4346–4353. [Google Scholar] [CrossRef]
- Nagle, L.; Place, A.; Schott, E.; Jagus, R.; Messick, G.; Pitula, J. Real-time PCR-based assay for quantitative detection of Hematodinium sp. in the blue crab Callinectes sapidus. Dis. Aquat. Org. 2009, 84, 79–87. [Google Scholar] [CrossRef]
- Parmenter, K.J.; Vigueira, P.A.; Morlok, C.K.; Micklewright, J.A.; Smith, K.M.; Paul, K.S.; Childress, M.J. Seasonal prevalence of Hematodinium sp. infections of blue crabs in three South Carolina (USA) rivers. Estuar. Coast 2013, 36, 174–191. [Google Scholar] [CrossRef]
- Aldik, R.; Cengizler, İ. The investigation of bacteria, parasite and fungi in blue crabs (Callinectes sapidus, Rathbun 1896) caught from Akyatan Lagoon in east Mediterranean Sea. J. VetBio Sci. Tech. 2017, 2, 11–17. [Google Scholar]
- Chatton, E.; Poisson, R. Sur l’existence, dans le sang des crabs, de péridiniens parasites : Hematodinium perezi n. g., n.sp. (Syndinidae). C.R. Séances. Soc. Biol. Paris. 1931, 105, 553–557. [Google Scholar]
- Li, M.; Huang, Q.; Lv, X.; Song, S.; Li, C. The parasitic dinolagellate Hematodinium infects multiple crustaceans in the polyculture systems of Shandong Province, China. J. Invertebr. Pathol. 2021, 178, 107523. [Google Scholar] [CrossRef]
- Gallien, L.; Sur la presence dans le sang de Platyonychus latipes Penn. d’un Peridinien parasite Hematodinium perezi Chatton et Poisson. Bull. Biol. Fr. Belg. 1938, 72, 1–7. [Google Scholar]
- MacLean, S.A.; Ruddell, C.L. Three new crustacean hosts for the parasitic dinofagellate Hematodinium perezi (Dinofagellata: Syndinidae). J. Parasitol. 1978, 64, 158–160. [Google Scholar] [CrossRef]
- Messick, G.A. Hematodinium perezi infections in adult and juvenile blue crabs Callinectes sapidus from coastal bays of Maryland and Virginia, USA. Dis. Aquat. Org. 1994, 19, 77–82. [Google Scholar] [CrossRef]
- Shields, J.D.; Squyars, C.M. Mortality and hematology of blue crabs, Callinectes sapidus, experimentally infected with the parasitic dinofagellate Hematodinium perezi. Fish. Bull. 2000, 98, 139. [Google Scholar]
- Hanif, A.W.; Dyson, W.D.; Bowers, H.A.; Pitula, J.S.; Messick, G.A.; Jagus, R.; Schott, E.J. Variation in spatial and temporal incidence of the crustacean pathogen Hematodinium perezi in environmental samples from Atlantic Coastal Bays. Aquat. Biosyst. 2013, 9, 1–13. [Google Scholar] [CrossRef]
- Butler, M.J.; Tiggelaar, J.M.; Shields, J.D. Effects of the parasitic dinofagellate Hematodinium perezi on blue crab (Callinectes sapidus) behavior and predation. J. Exp. Mar. Biol. Ecol. 2014, 461, 381–388. [Google Scholar] [CrossRef]
- Lycett, K.A.; Chung, J.S.; Pitula, J.S. The relationship of blue crab (Callinectes sapidus) size class and molt stage to disease acquisition and intensity of Hematodinium perezi infections. PLoS ONE 2018, 13, e0192237. [Google Scholar] [CrossRef]
- Small, H.J.; Huchin-Mian, J.; Reece, K.; Lohan, K.P.; Butler, M.J.; Shields, J.D. Parasitic dinofagellate Hematodinium perezi prevalence in larval and juvenile blue crabs Callinectes sapidus from coastal bays of Virginia. Dis. Aquat. Org. 2019, 134, 215–222. [Google Scholar] [CrossRef]
- Huang, Q.; Waser, A.M.; Li, C.; Thieltges, D.W. Lack of Hematodinium microscopic detection in crustaceans at the northern and southern ends of the Wadden Sea and an update of its distribution in Europe. Mar. Biol. 2024, 171, 1–8. [Google Scholar] [CrossRef]
- Alimin, A.W.F.; Yusoff, N.A.H.; Kadriah, I.A.K.; Anshary, H.; Abdullah, F.; Jabir, N.; Hassan, M. Parasitic dinoflagellate Hematodinium in marine decapod crustaceans: a review on current knowledge and future perspectives. Parasitol. Res. 2024, 123, 49. [Google Scholar] [CrossRef]
- Barbosa, H.; Soares, A. M.; Pereira, E.; Freitas, R. Are the consequences of lithium in marine clams enhanced by climate change? Environ. Pollut. 2023, 326, 121416. [Google Scholar] [CrossRef]
- Shields, J.D., Sullivan, S.E.; Small, H.J. Overwintering of the parasitic dinoflagellate Hematodinium perezi in dredged blue crabs (Callinectes sapidus) from Wachapreague Creek, Virginia. J. Invertebr. Pathol. 2015, 130, 124–132. [CrossRef]
- Morado, J.F.; Jensen, P.; Hauzer, L.; Lowe, V.; Califf, K.; Roberson, N.; shavey, C.; Woodby, D. Species identity and life history of Hematodinium, the causitive agent of bitter crab syndrome in north east pacific snow, Chionoecetes opilio, and tanner, C. bairdi, crabs. 2005. PROJECT 0306 FINAL REPORT. Anchorage, AK, USA: North Pacific Research Board.
- Shields, J.D. Collection techniques for the analyses of pathogens in crustaceans. J. Crus. Biol. 2017, 37, 753–763. [Google Scholar] [CrossRef]
- Cunningham, C.O. Molecular diagnosis of fish and shellfish diseases: present status and potential use in disease control. Aquaculture 2002, 206, 19–55. [Google Scholar] [CrossRef]
- Lee, R.F.; Frischer, M.E. The decline of the blue crab. Am. Sci. 2004, 92, 548–553. [Google Scholar]
- Small, H.J.; Neil, D.M.; Taylor, A.C.; Atkinson, R.J.A.; Coombs, G.H. Molecular detection of Hematodinium spp. in Norway lobster Nephrops norvegicus and other crustaceans. Dis. Aquat. Org. 2006, 69, 185–195. [Google Scholar] [CrossRef]
- Davies, C.E.; Thomas, J.E.; Malkin, S.H.; Batista, F.M.; Rowley, A.F.; Coates, C.J. Hematodinium sp. infection does not drive collateral disease contraction in a crustacean host. Elife 2022, 11, e70356. [Google Scholar] [CrossRef]
Figure 1.
Map showing the localization of Merja Zerga (A) and Oualidia (B) Lagoons on the Atlantic coast of Morocco.
Figure 1.
Map showing the localization of Merja Zerga (A) and Oualidia (B) Lagoons on the Atlantic coast of Morocco.
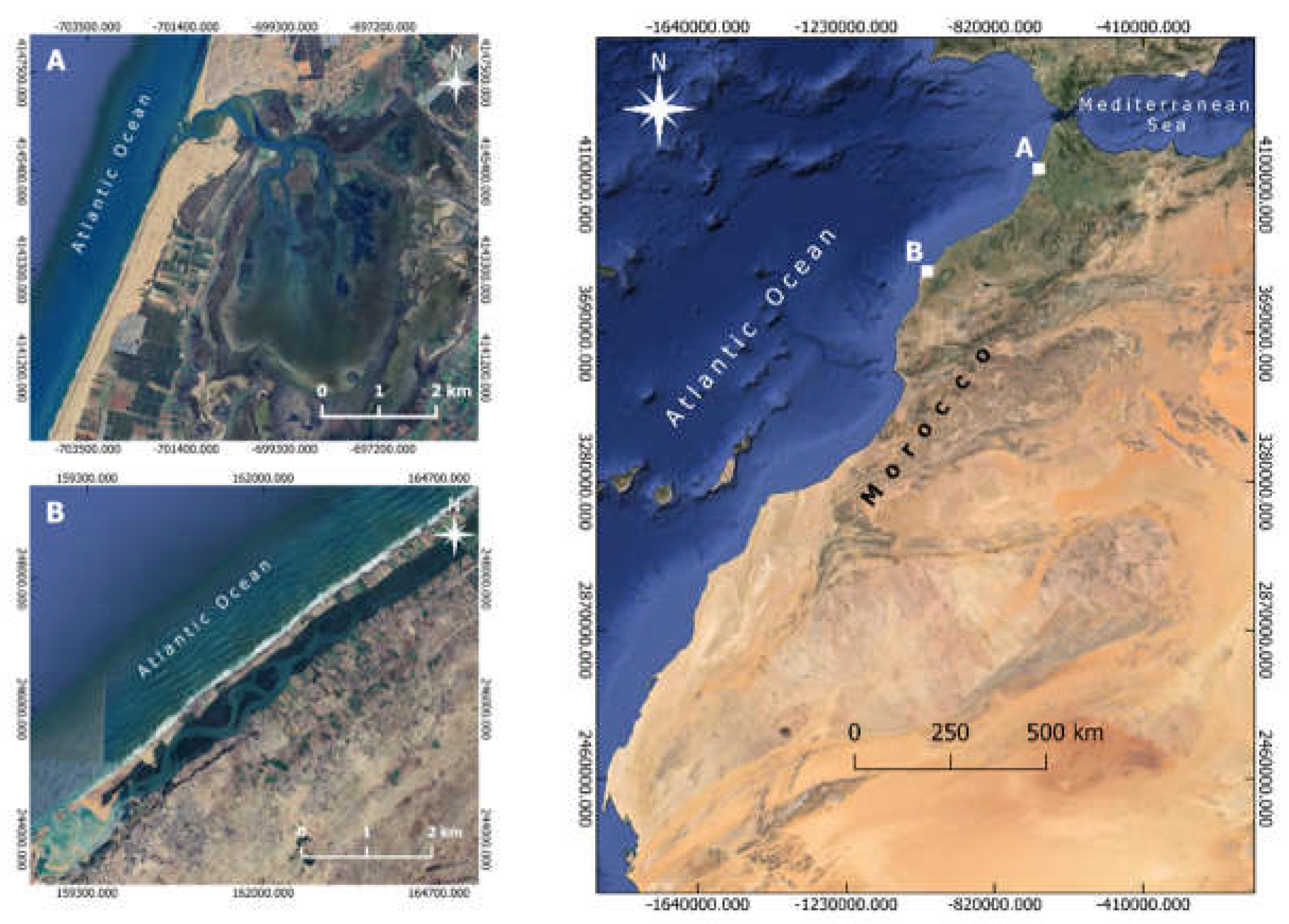
Figure 2.
Map showing the distribution of Hematodinium species infecting Callinectes sapidus. Note: Hematodinium perezi (red dot); Hematodinium sp. (green dot); and Hematodinium-like disease (blue dot) [33,78].
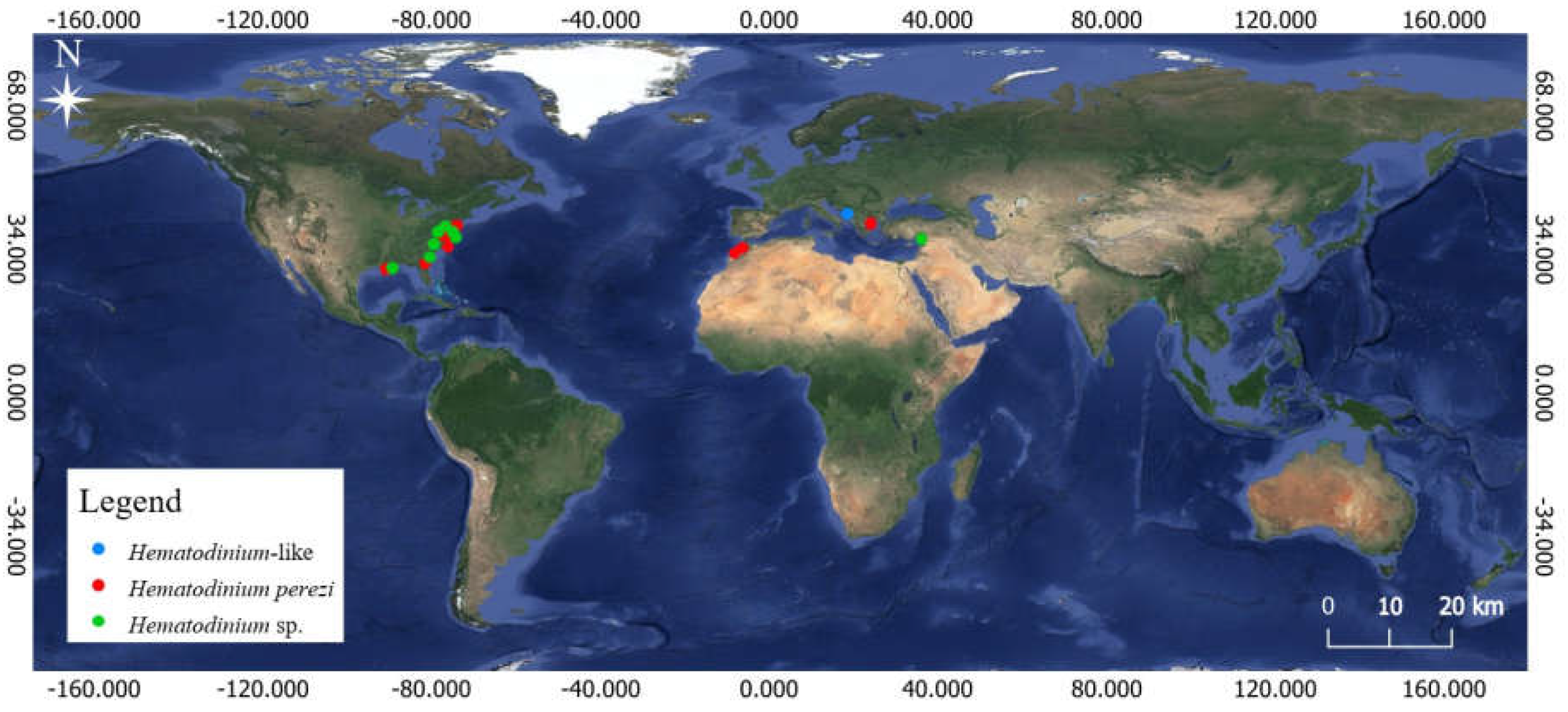

Table 1.
Biometric measurements of specimens of Callinectes sapidus from Merja Zerga and Oualidia Lagoons on the Moroccan Atlantic. CL: carapace length; CW: carapace width; W: body weight; SD: standard deviation.
Table 1.
Biometric measurements of specimens of Callinectes sapidus from Merja Zerga and Oualidia Lagoons on the Moroccan Atlantic. CL: carapace length; CW: carapace width; W: body weight; SD: standard deviation.
| Lagoon | Sampling period | Number of individuals | W (g) | CW (cm) | CL (cm) | |||
|---|---|---|---|---|---|---|---|---|
| Min-Max | Mean ±SD | Min-Max | Mean ±SD | Min-Max | Mean ±SD | |||
| Merja Zerga | February 2023 | 20 | 4.6 - 237.5 | 88.9 ± 78.7 | 3.1 - 14.8 | 9.2 ± 4.1 | 2.2 - 6.6 | 4.6 ± 1.8 |
| Oualidia | March 2023 | 16 | 18.8 -207 | 57.0 ± 52.4 | 5.8 - 14.9 | 8.6 ± 2.6 | 3.1 - 7.4 | 4.5 ± 1.4 |
Table 2.
List of Hematodinium perezi sequences used in this study, their GenBank accession numbers, host species, localities and references.
Table 2.
List of Hematodinium perezi sequences used in this study, their GenBank accession numbers, host species, localities and references.
| GenBank accession number | Host | Locality | References |
|---|---|---|---|
| PP056127 | Callinectes sapidus (Portunidae) | Greece | [33] |
| EF065716 | Liocarcinus depurator (Polybiidae) | South Coast of England | [56] |
| EF065708 | Liocarcinus depurator | South Coast of England | [56] |
| EF065711 | Liocarcinus depurator | South Coast of England | [56] |
| KX244637 | Portunus trituberculatus (Portunidae) | China | [57] |
| KX244644 | Portunus trituberculatus | China | [57] |
| KX244641 | Portunus trituberculatus | China | [57] |
| KX244634 | Callinectes sapidus | USA | [57] |
Table 3.
Range of uncorrected pairwise genetic distances (p-distances) between ITS1 rDNA sequences of Hematodinium perezi infecting Callinectes sapidus collected from Morocco and all published sequences from GenBank.
Table 3.
Range of uncorrected pairwise genetic distances (p-distances) between ITS1 rDNA sequences of Hematodinium perezi infecting Callinectes sapidus collected from Morocco and all published sequences from GenBank.
| Merja Zerga | Oualidia | South Coast of England | China | USA | Greece | |
|---|---|---|---|---|---|---|
| Merja Zerga Lagoon | - | - | - | - | - | - |
| Oualidia Lagoon | 0%-0.6% | - | - | - | - | - |
| South Coast of England EF065716, EF065708, EF065711 |
0.6%-1.3% | 0.6%-1.3% | - | - | - | - |
| China KX244637, KX244644, KX244641 |
1.7%-2.3% | 1.7%-2.3% | 1%-1.7% | - | - | - |
| USA KX244634 |
3.7%-4% | 3.7%-4% | 3.6%-4% | 3.7%-4% | - | - |
| Greece PP056127 |
0.6%-1.6% | 0.6%-1.6% | 0.3%-0.6% | 1.3%-1.7% | 3.4% | - |
Table 4.
Biometric measurements of specimens of Callinectes sapidus parasitized by Hematodinium perezi from Merja Zerga and Oualidia Lagoons on the Moroccan Atlantic. CL: carapace length; CW: carapace width; W: body weight; SD: standard deviation.
Table 4.
Biometric measurements of specimens of Callinectes sapidus parasitized by Hematodinium perezi from Merja Zerga and Oualidia Lagoons on the Moroccan Atlantic. CL: carapace length; CW: carapace width; W: body weight; SD: standard deviation.
| Lagoon | Groups | Total individuals | W (g) | CW (cm) | CL (cm) | |||
|---|---|---|---|---|---|---|---|---|
| Min-Max | Mean ±SD | Min-Max | Mean ±SD | Min-Max | Mean ±SD | |||
| Merja Zerga | male adult | 2 | 159.2 – 175.7 | 167.4 ± 11.6 | 11.7 – 12.1 | 11.9 ± 0.2 | 6.2 - 6.5 | 6.3 ± 0.2 |
| female adult | 5 | 109.9 - 153.1 | 131 ± 18.2 | 12.5 - 13.3 | 12.9 ± 0.3 | 5.8 - 6.3 | 6.1 ± 0.2 | |
| male juveniles | 1 | 4.6 | 3.1 | 2.2 | ||||
| female juveniles | 5 | 10.4 - 22.2 | 17.0 ± 5.0 | 4.3 - 6.4 | 5.4 ± 0.9 | 2.6 - 3.2 | 3.0 ± 0.3 | |
| Oualidia | male adult | 1 | 70.1 | 9.7 | 5.0 | |||
| female adult | 0 | - | - | - | - | - | - | |
| male juveniles | 2 | 21.6 - 30.5 | 25.8 ± 6.6 | 6.2 - 7.1 | 6.6 ± 0.6 | 3.3 - 3.7 | 3.5 ± 0.3 | |
| female juveniles | 1 | 18.7 | 6.1 | 3.1 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



