
Submitted:
06 December 2024
Posted:
09 December 2024
You are already at the latest version
Abstract
This study investigates Brucella ceti infection in marine mammals stranded along the Lisbon and Tagus Valley coast between 2022 and mid-2024, marking the first report of Brucella presence in Portuguese waters. Out of 59 examined marine mammals, B. ceti was isolated in three common dolphins (5.1%), a prevalence rate consistent with previous studies from other coastlines. Pathological findings varied, with one dolphin exhibiting neurobrucellosis-like symptoms and others showing diverse clinical presentations, complicating diagnosis. PCR-based detection indicated a higher infection rate (23.7%), suggesting underestimation in prevalence due to sample preservation status. Multilocus Sequence Typing (MLST) and Multiple-Locus Variable-Number Tandem-Repeat Analysis (MLVA) revealed distinct genetic profiles and close relationships to B. ceti strains from the Atlantic, supporting the hypothesis of specific host-adapted lineages in dolphins. These findings underscore the need for further research and surveillance to understand B. ceti transmission, host range, and impacts on Atlantic cetaceans, as well as to develop effective diagnostic and management strategies to mitigate infection risks in marine environments.
Keywords:
Atlantic Sea
; Brucella ceti
; Cetaceans
; Comparative Genomics
; Infection surveillance
; Marine environment health
; Portugal
1. Introduction
Brucella infections of terrestrial mammals have long been recognized and extensively researched; however, it was only during the last years of the twentieth century that the first reports on Brucella species from animals living in the marine environment were made, leading later to the inclusion of two novel species in the genus: Brucella ceti and Brucella pinnipedialis [1]. The first reports on Brucella sp. isolates from marine mammals were from wild harbour seals (Phoca vitulina), a harbour porpoise (Phocoena phocoena), and a common dolphin (Delphinus delphi) in Scotland [2] and a captive bottlenose dolphin (Tursiops truncatus) in the USA [3]. B. ceti infections have been frequently described in different dolphin species from the Atlantic and the Pacific Oceans [4], as well as from the Mediterranean Sea [5], and it has been recognized as a significant health concern for cetacean populations [6]. These reports have significantly broadened the range of host species known to be affected. In addition, they have extended the area over which the infection is known to occur, to the point where, if serological evidence is included, it seems likely that Brucella infection among sea mammals has a global occurrence [2].
B. ceti and B. pinnipedialis have been isolated from lungworms and a variety of organs in marine mammals [1,5,6,7,8,9]. In dolphins, B. ceti infections can be a challenging diagnosis, as the symptoms are nonspecific and depending on the affected organ or system. This infection is usually associated to meningoencephalomyelitis [1,3,4,7,8,9], reproductive tract inflammation (orchitis, endometritis, placentitis, endometritis), mastitis, abortion [1,2], discospondylitis, subcutaneous abscesses, and a wide range of other pathological conditions like pneumonia, myocarditis, pericarditis, osteoarthritis, hepatic, splenic, and lymph node necrosis, alongside macrophage infiltration in the liver and spleen, [1,2,3,4,5,6,7,8,9]. Neurobrucellosis represents a common cause of stranding for cetaceans, being associated with disorientation, uncoordinated lateral swimming, buoyancy disturbances, and death [4,9].
Grattarola and colleagues [10], among different bacteria with a potential zoonotic role identified in cetaceans stranded along the Italian Coastline, Brucella spp. were one of the most represented bacterial species, with a prevalence of 4.92%. In fact, there are several reports on B. ceti isolation on the Catalonian coast, in Italy, and on the Spanish Mediterranean coast from striped and bottlenose dolphins [9,10,11,12]. Additionally, a recent review from Jamil and Colleagues [13] highlights the presence of B. ceti and B. pinnipedialis infections in different marine mammal species across European regions, including North Atlantic Ocean, the Russian Bering Island, the Croatian Adriatic Sea, the Netherlands, Germany, and Norway. These epidemiological studies are crucial in elucidating the transmission dynamics, prevalence, and risk factors associated with Brucella spp. infection in marine mammal populations, not only offering valuable insights into the prevalence and distribution of the pathogen but also informing targeted intervention strategies to help mitigate its spread. Although there are no studies on the prevalence of Brucella in wild marine mammals in Portugal, the results above described in surrounding areas, lead us to create the hypothesis that the populations of marine mammals that arrive and pass through Portugal also have the bacteria.
Identifying the etiology of B. ceti infections and conducting comprehensive risk assessments are fundamental steps in developing effective biosafety protocols. By characterizing the virulence factors, antibiotic resistance profiles, and transmission routes of B. ceti, researchers can assess the potential risks posed to marine mammal populations, aquaculture facilities, and Public Health. B. ceti infections in marine mammals can pose a concern for marine conservation efforts [6], as increased pathogen prevalence might interfere with population abundance, by inducing high mortality rates, lowering reproductive success or by synergistically increasing the virulence of other diseases.
In this study, Brucella infection in stranded marine mammals in the region of Lisbon and Tagus Valley (Portugal) was investigated to determine prevalence rates, species identification, phylogenetic relationships and perform comparative genomic analysis. We highlight that this is the first time B. ceti was isolated from dolphins from the Portuguese coastline.
2. Materials and Methods
2.1. Stranding Data Collection, Necropsy and Tissue Sampling
Post-mortem examinations were performed on 59 marine mammals (Balaenoptera acutorostrata n=1; Delphinus delphis n=45; Phoca Vitulina n=1; Phocoena phocoena n=2; Stenella coeruleoalba n=6; Tursiops truncates n=3; Unidentified n=1) stranded in different beaches along the Lisbon and Tagus Valley coastline, specifically between Lourinhã and Setúbal municipalities, between January 2022 and June 2024. Geographical distribution of the stranding events and species composition of the animals under study is shown in Figure 1. Necropsy and macroscopic evaluation, as well as sample collection, were performed by the Lisbon and Tagus Valley Marine Animals' Stranding Network. Timelapse between receiving the stranding alert and performing necropsy and sample collection was under 12 in all of the investigated animals. A detailed post-mortem examination was performed according to standard protocols [14], and sample collection depended on the carcasses’ preservation status. Individual data, including sex, decomposition code and nutritional condition, along with stranding data concerning the geographical distribution, and date of occurrence, were registered. During necropsies, samples from tissues (brain, spleen, liver, lung, mammary gland, testis/uterus, and several limph nodes), vaginal and preputial swabs, and/or fluids (blood, milk) were collected, and kept frozen at <-16ºC until tested. All samples (Supplementary file Table S1) were submitted to the Animal Health National Reference Laboratory for microbiological and molecular investigations focused on Brucella infection diagnosis (INIAV, Oeiras, Portugal).
2.2. Brucella Isolation and Identification
The primary isolation of Brucella spp. was performed on all tissues, swabs, and fluids available from the 59 animals stranded on different beaches along the Lisbon Coastline between 2022 and June 2024. Samples were homogenized under sterile conditions in the minimum possible amount of sterile buffered saline (PBS pH 6.8) in a Stomacher unit (Seward Medical, Worthing, UK), and 0.2 mL of each tissue homogenate inoculated on two plates of both Farrell’s [15] and CITA [16] selective media. The plates were observed for microbial growth following 5–10 days of incubation at 37°C in both ambient air and a 5% CO₂ environment. A culture was considered positive when at least one Brucella colony forming unit (CFU) was isolated. Suspected colonies were further identified and characterized by standard bacteriological procedures, based on CO2 requirement, H2S production, oxidase test, urea hydrolysis, agglutination with monospecific sera anti-A, anti-M, and anti-R, and fuchsin and thionine dye sensitivity [17].
2.3. Molecular Methods
Bacterial genomic DNA extraction from available tissues was performed in a nucleic acid extraction workstation, Kingfisher Flex (Thermo Fisher Scientific, Waltham, MA, USA), using the IndiMag Kit (Indical Bioscience, Leipzig, Germany), following the manufacturer’s instructions. After extraction, the DNA was stored at 4° C. All samples were tested using RT-PCR targeting bcsp31 and per genes, as described previously [18], for identification of Brucella spp. Briefly, real-time TaqMan PCR was set up in a final volume of 25 µl, 1x TaqMan Universal PCR Master Mix (Applied Biosystems, France), each primer and TaqMan probe at concentrations of 0.3 µM and 0.25 µM, respectively, and 3 ng of DNA template. The reaction mixture was initially incubated for 10 min at 95° C. Amplification was performed for 45 cycles of denaturation at 95° C for 15 s, annealing and extension at 60 ° C for 1 min. The PCR reaction was performed on a Bio-Rad CFX Maestro 2.3 (Bio-Rad, France). A result was considered positive when an amplification curve with a Ct value less than 38 was obtained for both targets.
Genomic DNA from each Brucella spp. isolated from this work, and control strains, were extracted with PureLink Genomic DNA Mini Kit (Invitrogen, USA), stored at -20° C until used. Brucellae isolates were identified using the multiplex PCR Bruce-ladder as described elsewhere [19]. DNA samples were also tested by Multiple loci variable number of tandem repeats analysis (MLVA-16) as previously described [19]. The 16 loci have been classified in three panels, named panel 1, composed of 8 minisatellite (bruce06, bruce08, bruce11, bruce12, bruce42, bruce43, bruce45 and bruce55), panel 2A (bruce18, bruce19 and bruce21) and panel 2B (bruce04, bruce07, bruce09, bruce16 and bruce30) composed of three and five microsatellite markers, respectively. Briefly, PCR reactions were performed in a total volume of 15 μl containing 3 ng of DNA, 1 × PCR Reaction Buffer, 1 U of Taq DNA polymerase (Promega, USA), 200 μM of each dNTPs and 0.3 μM of each flanking primers. An initial denaturation step at 96° C for 5 min was followed by 30 cycles of denaturation at 96° C for 30 s, primer annealing at 60° C for 30 s and elongation at 70° C for 1 min. The final extension step was performed at 70° C for 5 min. Amplification products were loaded on a 3% standard agarose gel to analyze panel 2A and 2B loci (tandem repeats with a unit length shorter than 8 bp), and on a 2% standard agarose gel for panel 1 loci (tandem repeats with a unit length larger than 10 bp), with suitable molecular size markers. The total number of repeats at each locus was determined by the correlation with the amplicon size according to the 2013 Brucella allele assignment table (Le Flèche et al., 2006 version 3.6 available at http://mlva.u-psud.fr). Genomic DNA from B. melitensis biovar 1 strain 16 M (ATCC 23456) and B. ceti Atlantic dolphin type (B14/94) were used as controls for alleles assignment.
2.4. Whole Genome Sequencing and Bioinformatic Analysis
Whole Genome Sequencing (WGS) was used to evaluate the genetic structure of B. ceti isolates to increase knowledge into brucellae genome evolution. The extracted nucleic acids were assessed for quality and quantity using spectrophotometry and dsDNA specific fluorescence-based assays. WGS was performed as previously described [21,22]. Briefly, to recover the genomic structure of these isolates as accurately as possible, the extracted long strand high-grade DNA was directly sequenced using native long-read Nanopore sequencing (GridION X5 sequencing platform, Oxford Nanopore Technologies) at a minimum coverage depth of 200X, using Nanopore V14 kit chemistry with R10.4.1 pore. The sequencing data was analyzed with a customized pipeline developed by BioISI Genomics for genome assembly. The analysis included base-calling, pre-filtering based on read size and quality, and subsequent assembly, which involved the identification of bacterial species using Kraken, plasmid identification with PlasmidFinder [23], clonal origin analysis through phylogenetic methods with ParSNP [24], pathogenicity prediction using PathogenFinder [25], identification of acquired virulence genes via VirulenceFinder and VFDB [26], identification of antibiotic resistance genes with CARD [27], MegaRES [28], and ResFinder [29], as well as Multi-Locus Sequence Typing (MLST) [30,31] and Multiple Locus Variable-number Tandem Repeat Analysis (MLVA) [32].
For bioinformatic analysis, only reads with a Q score > 7 (minimum value from MinKNOW) were retained. Duplex base calling was performed using Dorado (v0.3.1+bb8c5ee) with the dna_r10.4.1_e8.2_400bps_sup@v4.2.0 model. Assembly was conducted with Canu (v2.0) [33] using parameters: genomeSize = 3.4M and -nanopore-raw. A quality report for the assemblies was generated using Quast (v5.0.2) with parameters set for a prokaryotic genome assembly. The Canu assemblies served as input for various bioinformatic tools available at https://genomicepidemiology.org/services/: Abricate v1.0.1 with VFDB [25,33], CARD, MegaRES [27,28], and PlasmidFinder (v2.1, “Gram Negative” database option) [25]. The assemblies were also analyzed using MLST tool (v2.0, with the configuration specific to Brucella spp.) and MLVA bank for Microbes genotyping (v1.4.0, configured for Brucella v4_6_5).
3. Results
3.1. Brucella Infection Diagnosis and Brucella Isolates Characterization
The isolation of Brucella spp. suspect colonies was obtained in three common dolphins (Delphinus delphis) out of the 59 animals investigated (5.1%). Suspected colonies were isolated from brain in DDE001 and DDE003 dolphins, and from spleen, liver, lung and lymph nodes in all the three animals. All isolates were typed by bacteriological and molecular methods and assigned as B. ceti. The phenotypic features regarding CO2 requirement, H2S production, oxidase and urea tests, agglutination with monospecific sera, and fuchsin and thionine sensitivity, were consistent with previously described B. ceti patterns: none of the three isolates required CO2 for growth or produced H2S, but all presented smooth phenotype, were oxidase and urease positive, agglutinate with anti-A monospecific serum, and grew in the presence of thionine and fuchsin dyes. Additionally, Brucella spp. DNA was detected in tissues from 14 animals (14/59; 23.7%), including the three dolphins with B. ceti isolation, and one Stenella coeruleoalba, and one Foca vitulina. The majority of the positive results were attained from the brain, spleen, liver, lung, lymph nodes, uterus and/or testis. In one suspected animal, DDE040, it was not possible to obtain a pure culture of the isolate, therefore phenotypic tests were inconclusive. However, the PCR carried out on the DNA extracted from the brain sample showed a positive result. B. ceti, isolates were also characterized by the multiplex Bruce-ladder multiplex, and all presented the molecular pattern comprising two fragments of 774 bp and 550 bp. Lastly, MLVA-16 analysis was performed and data compared to results obtained by other authors with B. ceti strains from different origins, which were deposited in the Brucella MLVA database (available at http://mlva.u-psud.frhttp://mlva.u-psud.fr). In this work, MLVA-11 (panels 1 and 2A markers) discriminated 2 genotypes (GT), dividing B. ceti DDE001 into GT8, and DDE002 and DDE003 isolates into GT-17. The results obtained during this study are summarized in Table 1, and in the Supplementary file Table S1.
3.2. Stranding Data and Necropsy Findings from B. ceti Positive Animals
Although post-mortem investigations were carried out on the 59 stranded animals, we will only focus on the observations of the three bacteriologically positive animals. Dolphins DDE001and DDE002 were juveniles, while DDE003 was an adult. DDE001 was a female dolphin that stranded in March 2022 in Sintra, Portugal (38.8545207, -9.4542235). The animal presented moderate nutritional condition and a physical exam revealed skin lacerations and traumatic lesions consistent with bycatch. Macroscopic evaluation of internal organs disclosed moderate pulmonary edema and the presence of gas embolism in the renal, mesenteric and thoracic vasculature. Additionally, mild splenic hyperplasia, with the presence of dark colored papules (0.5 cm diameter) at the organ’s surface and decreased consistency, was also observed. DDE002 was a male dolphin that stranded in March 2023 in Setúbal, Portugal (38.5094191, -8.9217497). The presence of live ectoparasites suggested a recent death. The animal was in a fair nutritional condition and presented a linear single deep laceration (1.4 cm depth, until the muscular layer) in one side of the body, suggesting a traumatic etiology. Pathological findings included a generalized lung hyperemia and a single ulcerative lesion in the oral cavity (2 cm diameter). DDE003 was a female dolphin that live stranded in July 2023 in Sesimbra, Portugal (38.4887533, -9.1840278). Reports indicate that the animal was disoriented and unable to swim and keep floatability alone. Despite efforts from people present on the beach to refloat the animal, it died shortly. The animal was severely emaciated. Pathological findings included moderate splenic hyperplasia, generalized lung hyperemia with associated nematode parasitic infection, the presence of petechiae in the intestinal mucosa, and congested areas in the meninges and the internal side of the skull.
3.3. Comparative genomic analysis and phylogenetic relationship of B. ceti isolates
The characterization of the isolates from dolphins DDE001, DDE002, and DDE003 by whole-genome sequencing, at a minimum coverage depth of 200X, confirmed them as members of B. ceti species. The key findings are summarized in Table 2.
The ParSNP analysis used the 10 assembled contigs of B. ceti strains available on NCBI as of February 2024 (https://blast.ncbi.nlm.nih.gov/) (see Supplementary file Table S2). This analysis revealed that isolates DDE002 and DDE003 are more closely related to each other than to DDE001. Additionally, strains M13/05/1 and M644/93/1 were found to have the closest relationship with all three isolates. Alignments were performed between each isolate and the closely related B. ceti strains M13/05/1 and M644/93/1, as well as among the Portuguese isolates, to evaluate single nucleotide variations (SNVs), insertions, and deletions. This analysis allowed for the construction of a phylogenetic tree. Detailed information on the variations in SNVs, insertions, and deletions in comparison to the closely related B. ceti strains and among the Portuguese isolates is summarized in Table 2 and Table 3, respectively, with the resulting phylogenetic tree illustrated in Figure 2. VirulenceFinder identified acquired virulence genes in Brucella isolates, as detailed in Supplementary file Table S3. For 100% identity and coverage, virulence factors were consistent across all isolates. However, in the core gene manCcore, isolates DDE002 and DDE003 showed a 99% identity, indicating a single mismatch when compared to B. ceti strain M644/93/1, while DDE001 had a 100% match. The identified virulence genes across the isolates were bspE, vceA, btpB, manAoAg, wbkC, virB7, lpxK, lpxA, and manCcore. The ABRicate pipeline, using CARD and MegaRES databases, identified six acquired antibiotic resistance genes (ARGs): mprF, which is commonly linked to resistance to cationic antimicrobial peptides due to its role in modifying cell membrane charge, and genes bepC, D, E, F and bepG, generally associated with resistance mechanisms that may include antibiotic modification or efflux (Supplementary file Table S4).
MLST and MLVA profiles (Table 2, and Supplementary file Table S5), revealed that DDE001 aligned with Sequence Type 49 (ST-49) profile (cg-ST392), while isolates DDE002 and DDE003 matched ST-26 profile (cg-ST340). The difference between ST-49 and ST-26 lies at the cobQ locus, where a single nucleotide polymorphism (SNP) occurs at position 213 bp within the 423-bp gene. This SNP is marked by a 'C' (allele 6) in ST-49 and a 'T' (allele 10) in ST-26. Additionally, MLVA profiling showed that isolate DDE001 exhibited close similarity to B. ceti strains M57/07/1 and M260/03/1, which were isolated from the spleen and brain, respectively, of Atlantic white-sided dolphins (Lagenorhynchus acutus) in Scotland, with both strains falling under the A1 cluster [32]. Isolate DDE002 was closely related to B. ceti strain M654/99/1 from a striped dolphin’s brain, also in Scotland, and classified within the A2 cluster [32]. Meanwhile, DDE003 showed close resemblance to B. ceti strains M654/99/1, M83/07/1, M231/07/3, M267/05/4, M83/07/3, and M267/05/1, which were obtained from the brain, kidney, and colorectal lymph nodes of Striped (Stenella coeruleoalba), Atlantic White-Sided, and bottlenose dolphins, all belonging to the A2 cluster [32].
4. Discussion
This study investigates Brucella infection in marine mammals stranded along the Lisbon and Tagus Valley coast from 2022 to mid-2024. In out of the 59 marine mammals examined, B. ceti was isolated in three common dolphins (5.1%). This finding highlight, for the first time, the presence of Brucella in marine mammals in Portugal coastline, expanding the dispersion area of this bacteria. The observed prevalence (5.1%) is like the 4.92% prevalence obtained by Grattarola and colleagues [10] in cetaceans stranded along the Italian Coastline.
As referred previously, B. ceti has been associated with a range of pathological changes in cetaceans. In this study, dolphins in which B. ceti isolation was achieved exhibited distinct pathological findings, with some being coincident with the ones expected for Brucella infections. For instance, dolphin DDE003, with isolation of B. ceti and Brucella DNA detection on the central nervous system, presented meningitis, associated to disorientation and inability to swim and maintain equilibrium and flotation, suggesting a situation of neurobrucellosis, like the findings in a dolphin of the Mediterranean Catalonian coast [4]. It also displayed severe emaciation, parasitic infection, and multiple signs of systemic distress, including splenic hyperplasia. DDE001 and DDE002 presented pathological changes typically not consistent with B. ceti infection, underlying the diverse clinical presentations of Brucella infections, already referred by other authors, and complicating the diagnosis in marine mammals. Furthermore, it is interesting to highlight that the three cases display different likely death causes, reinforcing the idea that susceptibility to this pathogen can differ between individuals and be linked to different ecological drivers.
Although B. ceti isolation was achieved in 5,1% (3/59) of dolphins the results obtained by PCR for Brucella spp. detection directly from the same tissues were higher, with molecular confirmation in 14 animals (23.7%), including those three Brucella-positive, suggesting that the incidence of this agent might be higher in these species. In fact, the three isolates here identified were obtained in samples from animals with code 1 (recently died) or 2 (fresh carcass) of preservation status [14], highlighting that Brucella isolation success may decrease with increased time between death and sampling.
The MLST analysis revealed distinct genetic profiles, indicating diversity within B. ceti strains. ST-26 was attributed to the strains from animals DDE002 and DDE003, also previously identified in isolates from dolphins of the Mediterranean Sea and North America. This sequence type is exclusively observed in dolphin isolates, forming the cluster A [4,5,8]. The isolate from DDE001 was assigned to ST-49, which was also observed in 17 B. ceti isolates, two in Spain and the remaining in Scotland, between 2006 and 2019 (PubMLST, available online: https://pubmlst.org/organisms?title=Brucella+spp. Accessed on 15/11/2024). Additionally, MLVA showed a close relation between the isolates from this work and isolates from the Atlantic Sea, although DDE001 falling under the A1 cluster, and DDE002 and DDE003 falling under A2 cluster [32]. Up to now, all B. ceti Mediterranean strains stem in a separate branch from the main MLVA A1 and A2 clusters of B. ceti isolates from dolphins inhabiting the Atlantic Ocean [4,5]. This close MLVA similarity to other dolphin-derived strains supports the hypothesis of specific host-adapted Brucella lineages in marine environments [35]. Also, the comparative genomic analysis showed a close relationship with other B. ceti strains previously isolated from dolphins, namely those obtained from dolphins in the Scottish sea [32], indicating possible common sources or transmission pathways within marine environments. In fact, the phylogenetic analysis based on the SNVs revealed that isolates DDE002 and DDE003 are more closely related to each other than to DDE001, belonging respectively to the cluster A2 and A1, suggesting potential differences in infection sources or transmission routes. Virulence genes were consistent across the isolates, reinforcing the pathogenic potential of B. ceti, with minor variations in the manCcore gene suggesting slight genetic diversity. Eight virulence associated genes were identified in the B. ceti isolates: bspE gene, associated with pathogenesis, and likely aids in host interaction, enhancing B. ceti's ability to cause disease; vceA gene, part of the virB operon, is essential for the Type IV secretion system, crucial for the intracellular survival as it allows Brucella to manipulate host cells; btpB gene is involved in host interactions and the modulation of the host immune response, probably contributing to the ability of B. ceti to evade the host immune system; manAoAg gene, involved in the biosynthesis of mannose-containing O-antigen; wbkC gene, associated with O-polysaccharide biosynthesis, which contributes to the structure of the bacterial cell surface; virB7 gene, also part of the Type IV secretion system, which is crucial for intracellular survival and virulence as this system allows Brucella to translocate effector molecules into host cell; lpxK and lpxA genes, involved in lipid A biosynthesis; and manCcore gene (required for synthetizing LPS core oligosaccharide), which is vital to Brucella's outer membrane and pathogenicity. Furthermore, the analysis performed using the CARD and MegaRES databases highlighted the presence of six genes potentially involved in AMR in the three B. ceti genomes: the multiple peptide resistance factors mprF, and the outer membrane efflux proteins bep C, D, E, F, and G. These results are in accordance with other authors that in a recent study, investigated and compared a panel of B. ceti and B. pinnipedialis genome sequences [36]. The identification of six antibiotic resistance genes (ARGs) in these isolates raises concerns regarding treatment challenges, highlighting the importance of monitoring ARGs in B. ceti to better understand and manage the risks associated with resistance in marine mammal pathogens.
5. Conclusions
In conclusion, the demonstration of the existence of B. ceti in dolphins for the first time in Portugal, highlights the need of further studies with greater geographic coverage to assess the infection's true prevalence along this coastline. These findings shed light on the occurrence, and characteristics of B. ceti infection in dolphins, and emphasize the need for continuous surveillance and advanced diagnostic methods to better understand and manage Brucella infections in marine mammals. Continued research on B. ceti infection is crucial to further understand its transmission dynamics, host range, and its impact on cetacean populations in the Atlantic Sea. Additionally, there is a need for the development of effective diagnostic tools and sustainable management strategies to mitigate the spread of this infection and preserve the health of affected cetaceans.
Supplementary Materials
Table S1: Cetacean strandings along the Lisbon and Tagus Valley Coastline (January 2022 – June 2024): Animal details, Sample collection, and Bacteriological and Molecular results. Table S2: Assembled Brucella ceti contigs from NCBI utilized in ParSNP analysis. Table S3: Identification of acquired virulence genes. Table S4: Identification of acquired antibiotic resistance genes. Table S5: Brucella ceti Multi-Locus Sequence Typing (MLST) and Multiple-Locus Variable Number Tandem-Repeat Analysis (MLVA) alleles profiles and clustering.
Author Contributions
Conceptualization, S.C., M.L.G., R.D., and A.C.F.; Formal analysis S.C, M.L.G., M.N., P.P., M.P., R.D., and A.C.F.; Investigation, S.C., M.L.G., M.N., P.P., M.P., C.F., A.B.C., S.P., R.D. and. A.C.F.; Methodology, S.C., M.L.G, M.N., P.P., M.P., R.D., and A.C.F.; Writing—original draft preparation, S.C., and A.C.F.; Writing—review and editing, S.C., M.L.G, M.N., R.D., and A.C.F. All authors have read and agreed to the published version of the manuscript.
Funding
Not applicable.
Institutional Review Board Statement
The study protocol was approved by the National Competent Authority (ICNF, ), under permits: LICENÇA Nº 170 / 2022 / CAPT, LICENÇA Nº 171 / 2022 / CAPT, LICENÇA Nº 883 / 2022 / CAPT, LICENÇA Nº 464 / 2023 / CAPT, LICENÇA Nº 465 / 2023 / CAPT, LICENÇA Nº 463 / 2023 / CAPT, LICENÇA Nº 462 / 2023 / CAPT, LICENÇA Nº 011 / 2024 / CAPT, LICENÇA Nº 014 / 2024 / CAPT, LICENÇA Nº 012 / 2024 / CAPT, LICENÇA Nº 009 / 2024 / CAPT, LICENÇA Nº 013 / 2024 / CAPT, LICENÇA Nº 010 / 2024 / CAPT.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article. The WGS data is submitted at NCBI GenBank with the following accession number: SAMN45131997, SAMN45131998, and SAMN45131999 for DDE001, DDE002 and DDE003 genome sequences, respectively.
Acknowledgments
Work supported by UIDB/04046/2020 and UIDP/04046/2020 Centre grants from FCT, Portugal (to BioISI).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Foster G., Osterman B. S., Godfroid J., Jacques I., Cloeckaert A. Brucella ceti sp. Nov. and Brucella pinnipedialis sp. nov. for Brucella strains with cetaceans and seals as their preferred hosts. Int. J. Syst. Evol. Microbiol. 2007, 57, 2688–2693. [CrossRef]
- Foster G., MacMillan A.P., Godfroid J., Howie F., Ross H.M., Cloeckaert A., Reid R. J., Brew S., Patterson I. A. A review of Brucella spp. infection of sea mammals with particular emphasis on isolates from Scotland. Vet. Microbiol. 2002, 90, 563–580. [CrossRef]
- Ewalt D.R., Payeur J. B., Martin B. M., Cummins D.R., Miller G. M. Characteristics of a Brucella species from a bottlenose dolphin (Tursiops truncates). J. Vet. Diagn. Invest. 1994, 6, 448–452. [CrossRef]
- Isidoro-Ayza, M. , Ruiz-Villalobos, N., Pérez, L., Guzmán-Verri, C., Muñoz, P.M., Alegre, F., Barberán, M., Chacón-Díaz, C., Chaves-Olarte, E., González-Barrientos, R., Moreno, E., Blasco, J.M., Domingo, M. Brucella ceti infection in dolphins from the Western Mediterranean sea. BMC Vet. Res, 2014; 10, 206. [Google Scholar] [CrossRef]
- Garofolo, G., Petrella, A., Lucifora, G., Di Francesco, G., Di Guardo, G., Pautasso, A., Iulini, B., Varello, K., Giorda, F., Goria, M., et al. Occurrence of Brucella ceti in Striped Dolphins from Italian Seas. PloS ONE 15, 2020, e0240178.
- Guzmán-Verri, C. , González-Barrientos, R., Hernández-Mora, G., Morales, J.Á., Baquero-Calvo, E., Chaves-Olarte, E., Moreno, E. Brucella ceti and brucellosis in cetaceans. Front Cell Infect Microbiol, 2012. [Google Scholar] [CrossRef] [PubMed]
- Nymo, I.H., Tryland, M., Godfroid, J. A Review of Brucella Infection in Marine Mammals, with Special Emphasis on Brucella pinnipedialis in the Hooded Seal (Cystophora cristata). Vet. Res. 2011, 42, 93.
- Whatmore, A.M., Dawson, C., Muchowski, J., Perrett, L.L., Stubberfield, E., Koylass, M., Foster, G., Davison, N.J., Quance, C., Sidor, I.F., et al. Characterisation of North American Brucella Isolates from Marine Mammals. PloS ONE, 2017, 12, e0184758.
- Grattarola, C. , Petrella, A., Lucifora, G., Di Francesco, G., Di Nocera, F., Pintore, A., Cocumelli, C., Terracciano, G., Battisti, A., Di Renzo, L., et al. Brucella ceti Infection in Striped Dolphins from Italian Seas: Associated Lesions and Epidemiological Data. Pathogens. [CrossRef]
- Grattarola, C., Pietroluongo, G., Belluscio, D., Berio, E., Canonico, C, Centelleghe, C., Cocumelli, C., Crotti, S., Denurra, D., Di Donato, A., et al. Pathogen Prevalence in Cetaceans Stranded along the Italian Coastline between 2015 and 2020. Pathogens. 2024, 13, 762. [CrossRef]
- Alba, P., Terracciano, G., Franco, A., Lorenzetti, S., Cocumelli, C., Fichi, G., Eleni, C., Zygmunt, M.S., Cloeckaert, A., Battisti, A. The presence of Brucella ceti ST26 in a striped dolphin (Stenella coeruleoalba) with meningoencephalitis from the Mediterranean Sea. Vet Microbiol. 2013 May 31;164(1-2):158-63. [CrossRef] [PubMed]
- Isidoro-Ayza, M. , Ruiz-Villalobos, N., Pérez, L., Guzmán-Verri, C., Muñoz, P.M., Alegre, F., Barberán, M., Chacón-Díaz, C., Chaves-Olarte, E., González-Barrientos, R., Moreno, E., Blasco, J.M., Domingo, M. Brucella ceti, 2014; 10. [Google Scholar] [CrossRef]
- Jamil, T. , Akar, K. , Erdenlig, S., Murugaiyan, J., Sandalakis, V., Boukouvala, E., Psaroulaki, A., Melzer, F., Neubauer, H., Wareth, G. Spatio-Temporal Distribution of Brucellosis in European Terrestrial and Marine Wildlife Species and Its Regional Implications. Microorganisms 2022, 10, 1970. [Google Scholar] [CrossRef]
- IJsseldijk, L.L., Brownlow, A.C. & Mazzariol, S. Best practice on cetacean post mortem investigation and tissue sampling – Joint ACCOBAMS and ASCOBANS. document.https://www.ascobans.org/sites/default/files/document/ascobans_ac25_inf3.2_rev1_best-practice-cetacean-post-mortem-investigation.pdf [Accessed August 3, 2023].
- Farrell, I.D. The development of a new selective medium for the isolation of Brucella abortus from contaminated sources. Res Vet Sci, 1974, 16:280–286.
- De Miguel, M.J., Marín, C.M., Muñoz, P.M., Dieste, L., Grilló, M.J., Blasco, J.M. Development of a selective culture medium for primary isolation of the main Brucella species. J Clin Microbiol, 2011, 49:1458–1463.
- World Organization for Animal Health. Brucellosis (infection with <i>B. abortus, B. World Organization for Animal Health. Brucellosis (infection with B. abortus, B. melitensis and B. suis). OIE Terrestrial Manual 2022.
- Bounaadja, L., Albert, D., Chénais, B., Hénault, S., Zygmunt, M.S., Poliak, S., Garin-Bastuji , B. Real-time PCR for identification of Brucella spp.: a comparative study of IS711, bcsp31 and per target genes. Vet Microbiol. 2009, 28;137(1-2):156-64. [CrossRef] [PubMed]
- Ferreira, A.C. , Corrêa de Sá, M.I., Dias, R. and Tenreiro, R. MLVA-16 typing of Brucella suis biovar 2 strains circulating in Europe. Vet Microbiol. [CrossRef]
- López-Goñi, I. , García-Yoldi, D., Marín, C.M., de Miguel, M.J., Barquero-Calvo, E., Guzmán-Verri, C., Albert, D., Garin-Bastuji, B. New Bruce-ladder multiplex PCR assay for the biovar typing of Brucella suis and the discrimination of Brucella suis and Brucella canis. Vet. Microbiol. 2011, 154, 152–155.
- Molina-Mora, J. A. , Campos-Sánchez, R., Rodríguez, C., Shi, L., & García, F. High quality 3C de novo assembly and annotation of a multidrug resistant ST-111 Pseudomonas aeruginosa genome: Benchmark of hybrid and non-hybrid assemblers. Scientific reports, 2020; 10, 1392. [Google Scholar] [CrossRef]
- Ruan, Z. , Wu, J., Chen, H., Draz, M. S., Xu, J., & He, F. Hybrid Genome Assembly and Annotation of a Pandrug-Resistant Klebsiella pneumoniae Strain Using Nanopore and Illumina Sequencing. Infection and drug resistance, 2020; 13, 199–206. [Google Scholar] [CrossRef]
- Carattoli, A., Hasman, H. PlasmidFinder and In Silico pMLST: Identification and Typing of Plasmid Replicons in Whole-Genome Sequencing (WGS). In: de la Cruz, F. (eds) Horizontal Gene Transfer. Methods in Molecular Biology, 2020, vol 2075. Humana, New York, NY. [CrossRef]
- Kille, B. , Nute, M.G., Huang, V., Kim, E., Phillippy, A.M., Treangen, T.J. Parsnp 2.0: Scalable Core-Genome Alignment for Massive Microbial Datasets. bioRxiv, 2024. [Google Scholar] [CrossRef]
- Cosentino S, Voldby Larsen M, Møller Aarestrup F, Lund O. PathogenFinder--distinguishing friend from foe using bacterial whole genome sequence data. PLoS One. 2013, 8(10):e77302. doi: 10.1371/journal.pone.0077302. Erratum in: PLoS One. 2013;8(12). doi:10.1371/annotation/b84e1af7-c127-45c3-be22-76abd977600f. PMID: 24204795; PMCID: PMC3810466.
- Chen, L. VFDB: a reference database for bacterial virulence factors. Nucleic Acids Research, 2004. [Google Scholar] [CrossRef]
- Alcock, B.P. , Huynh, W., Chalil, R., Smith, K.W., Raphenya, A.R., Wlodarski, M.A., Edalatmand, A., Petkau, A., Syed, S.A., Tsang, K.K., et al. CARD 2023: expanded curation, support for machine learning, and resistome prediction at the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. [CrossRef] [PubMed]
- Lakin, S.M. , Dean, C., Noyes, N.R., Dettenwanger, A., Ross, A.S., Doster, E., Rovira, P., Abdo, Z., Jones, K.L., Ruiz, J., Belk, K.E., Morley, P.S., Boucher, C. MEGARes: an antimicrobial resistance database for high throughput sequencing. Nucleic Acids Res, 2017. [Google Scholar] [CrossRef]
- Larsen, M. V. , Cosentino, S., Lukjancenko, O., Saputra, D., Rasmussen, S., Hasman, H., Sicheritz -Pontén, T., Aarestrup, F. M., Ussery, D. W., & Lund, O. Benchmarking of Methods for Genomic Taxonomy. J Cli Microbiol, 2014, 52(5), 1529–1539. [CrossRef]
- Larsen, M.V. , Cosentino, S., Rasmussen, S., Friis, C., Hasman, H., Marvig, R.L., Jelsbak, L., Sicheritz-Pontén, T., Ussery, D.W., Aarestrup, F.M., Lund, O. Multilocus Sequence Typing of Total-Genome-Sequenced Bacteria. J Cli Microbiol, 2012; 50, 1355–1361. [Google Scholar] [CrossRef]
- Florensa, A.F. , Kaas, R.S., Clausen, P.T.L.C., Aytan-Aktug, D., Aarestrup, F.M. ResFinder - an open online resource for identification of antimicrobial resistance genes in next-generation sequencing data and prediction of phenotypes from genotypes. Microb Genom, 2022. [Google Scholar] [CrossRef] [PubMed]
- Maquart, M. , Le Flèche, P., Foster, G. et al. MLVA-16 typing of 295 marine mammal Brucella isolates from different animal and geographic origins identifies 7 major groups within Brucella ceti and Brucella pinnipedialis. BMC Microbiol. [CrossRef]
- Koren, S. , Walenz, B.P., Berlin, K., Miller, J.R., Bergman, N.H., Phillippy, A.M. Canu: scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res, 2017; 27, 722–736. [Google Scholar] [CrossRef] [PubMed]
- Seemann T, A. (n.d.). Abricate. https://github.com/tseemann/abricate.
- Suárez-Esquivel, M. , Baker, K.S., Ruiz-Villalobos, N., Hernández-Mora, G., Barquero-Calvo, E., González-Barrientos, R., Castillo-Zeledón, A., Jiménez-Rojas, C., Chacón-Díaz, C., Cloeckaert, A., et al. Brucella Genetic Variability in Wildlife Marine Mammals Populations Relates to Host Preference and Ocean Distribution. Genome Biol. Evol, 2017; 9, 1901–1912. [Google Scholar]
- Girault, G. , Freddi, L., Jay, M., Perrot, L., Dremeau, A., Drapeau, A., Delannoy, S., Fach, P., Ferreira Vicente, A., Mick, V., Ponsart, C., Djokic, V. Combination of in silico and molecular techniques for discrimination and virulence characterization of marine Brucella ceti and Brucella pinnipedialis. Front Microbiol, 2024; 15, 1437408. [Google Scholar] [CrossRef]
Figure 1.
Geographical distribution of the stranding events and species composition of the animals under study.
Figure 1.
Geographical distribution of the stranding events and species composition of the animals under study.
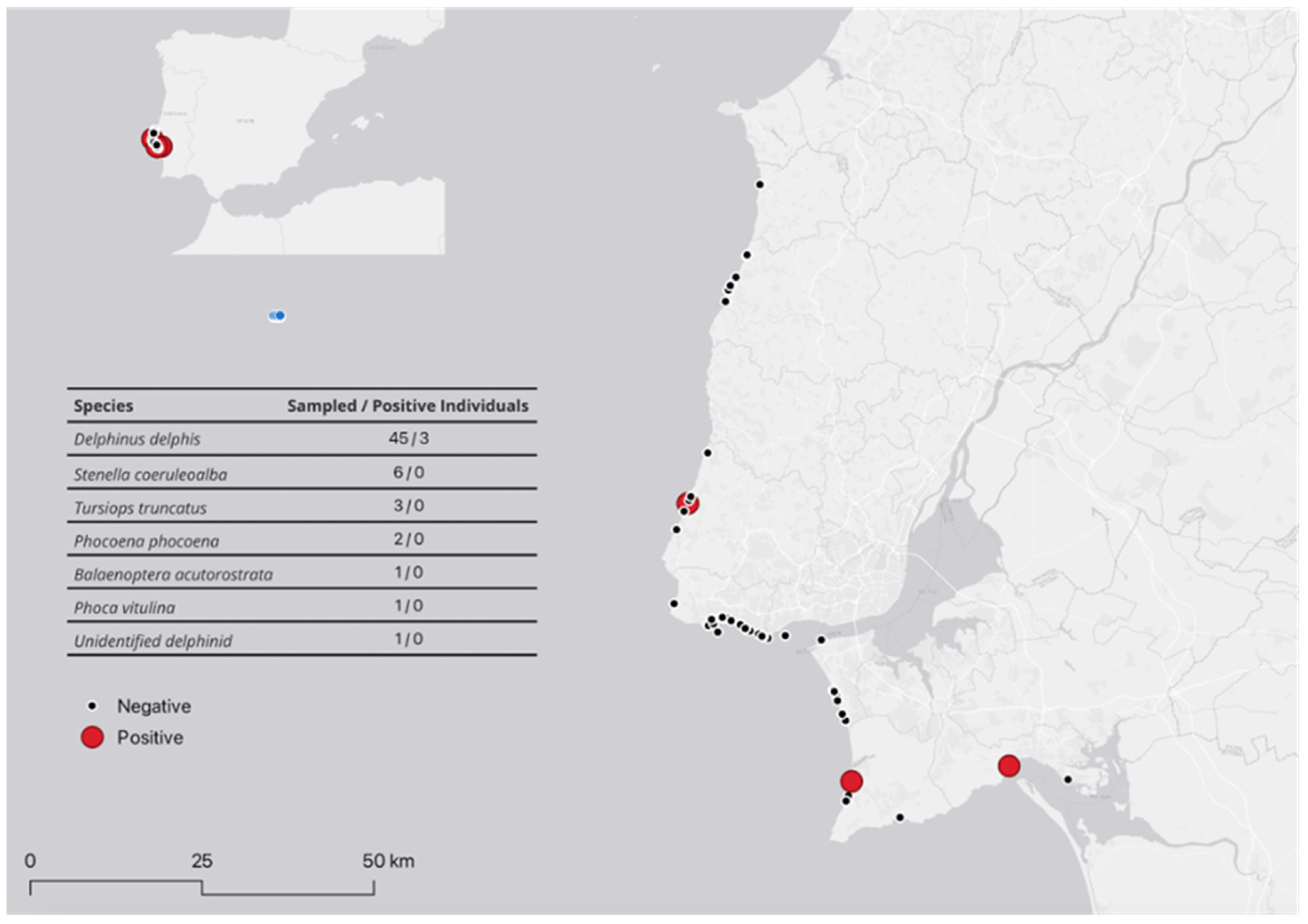
Figure 2.
Phylogenetic tree generated by ParSNP. The phylogenetic tree was based on 689 SNVs and is rooted with Brucella ceti TE28753-12. The branch length is proportional to the number of SNVs (the scale bar represents the difference in SNVs). The strains from this study are marked with a yellow square.
Figure 2.
Phylogenetic tree generated by ParSNP. The phylogenetic tree was based on 689 SNVs and is rooted with Brucella ceti TE28753-12. The branch length is proportional to the number of SNVs (the scale bar represents the difference in SNVs). The strains from this study are marked with a yellow square.
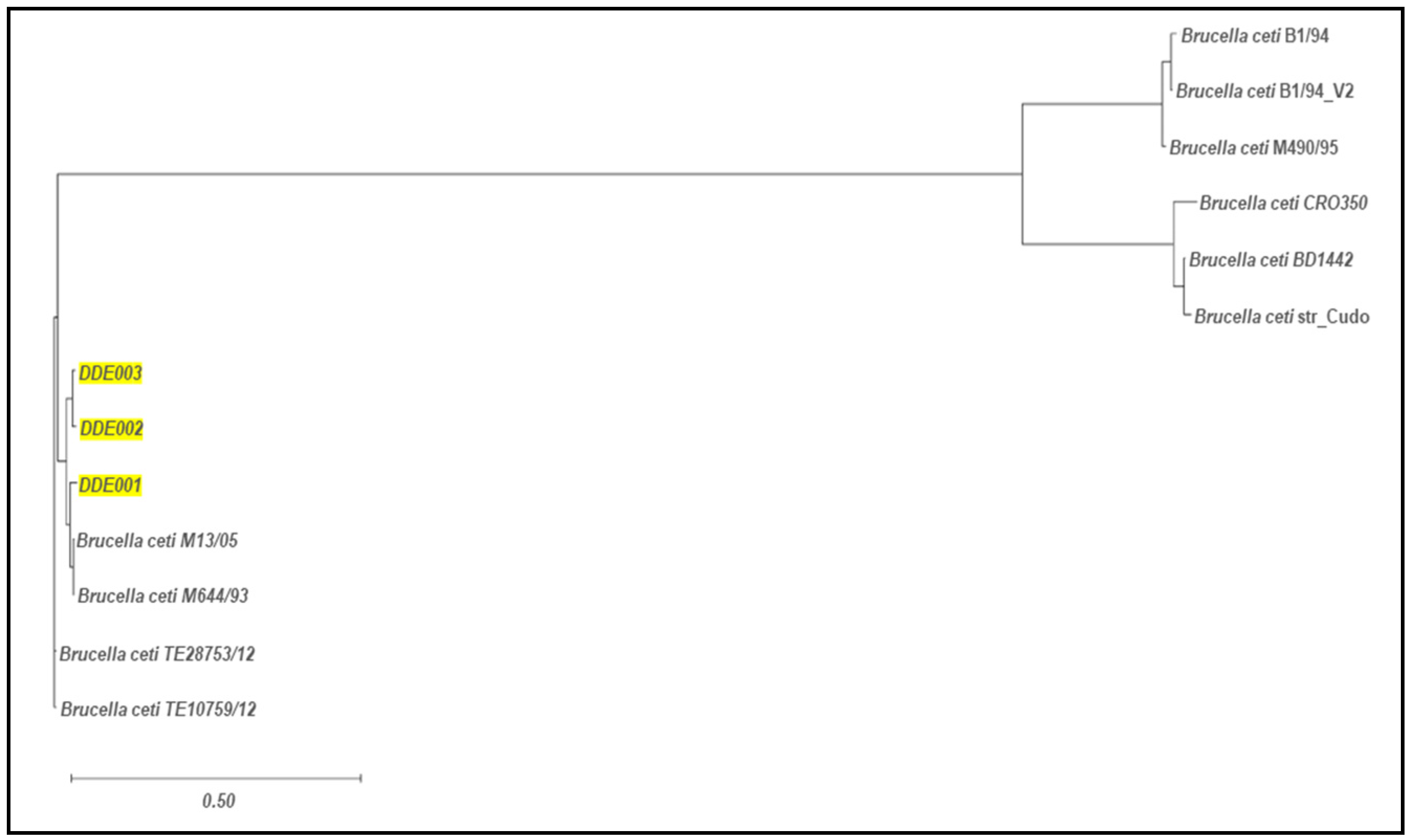

Table 1.
Confirmed and suspected positive animals, and respective qPCR and bacteriological results.
| Animal Identification1 | Sex/Age2 | Status3 | Region | Year of stranding | qPCR positive samples 4 | Bacteriological suspected samples | Brucella characterization |
|---|---|---|---|---|---|---|---|
| DDE001 | F/J | 2 | Sintra | 2022 | B,L,Lg,LN,S | B,L,Lg,S | B. ceti |
| DDE002 | M/J | 2 | Setúbal | 2023 | B,L,Lg,LN,S | L,Lg,LN,S | B. ceti |
| DDE003 | F/A | 1 | Sesimbra | 2023 | B,C | B,L,Lg,LN,S | B. ceti |
| DDE005 | F/A | 4 | Setúbal | 2022 | B,L,Lg,S | N | N |
| DDE008 | M/A | 2 | Sesimbra | 2022 | B,L,Lg,S | N | N |
| DDE013 | M/J | 2 | Cascais | 2022 | LN | N | N |
| SCO016 | M/A | 2 | Almada | 2023 | B | N | N |
| DDE027 | M/J | 2 | Cascais | 2023 | B,L,Lg,LN,S,T | N | N |
| DDE033 | F/A | 1 | Cascais | 2023 | LN | N | N |
| PVI035 | M/J | 4 | Cascais | 2023 | T | N | N |
| DDE036 | M/A | 4 | Cascais | 2023 | LN | N | N |
| DDE037 | F/A | 4 | Cascais | 2023 | LN | N | N |
| DDE040 | M/J | 2 | Cascais | 2023 | B | B | Inconclusive |
| DDE043 | M/J | 2 | Cascais | 2023 | PS | N | N |
1 DDE: Delphinus delphis; PVI: Phoca vitulina; SCO: Stenella coeruleoalba. 2 A: adult; F: female; J: juvenil; M: male. 3 Carcasses’preservation status: 1 (just died), 2 (fresh carcass), 3 (moderate decomposition), 4 (advanced decomposition). 4 B: brain; C: cervix; L: liver; Lg: lung; LN: limph nodes; N: negative; PS: prepucial swab; S: spleen; T: testis.
Table 2.
Summary of the main genomic results for dolphins B. ceti isolates.
| B. ceti isolates | DDE001 | DDE002 | DDE003 | |
|---|---|---|---|---|
| B. ceti strains M13/05/1 alignment | 3077 SNVs, 279 insertions, 257 deletions |
2956 SNVs, 248 insertions, 241 deletions |
2478 SNVs, 212 insertions, 215 deletions |
|
| B. ceti strain M644/93/1 alignment | 5855 SNVs, 505 insertions, 562 deletions |
4996 SNVs, 439 insertions 457 deletions |
4997 SNVs, 439 insertions, 464 deletions |
|
| Virulence genes | bspE, vceA, btpB, manAoAg, wbkC, virB7, lpxA, lpxK, manCcore | |||
| Antibiotic resistance genes | mprF, bepC and D, E, F, G | |||
| MLST 1 | ST-49 | ST-26 | ||
| cgMLST 2 | cgST-392 | cgST-340 | ||
| MLVA 3 | Cluster A1 | Cluster A2 | ||
1 MLST: Multlocus sequence typing (ST: Sequence Type from the typing database htps://pubmlst.org/ for the species Brucella spp.);. 2 cgMLST: core genome MLST (cgST from the typing database htps://pubmlst.org/ for the species Brucella spp.);. 3 MLVA: Multiple-Locus Variable-Number Tandem Repeat Analysis; cluster attribution based on: Maquart et al. 2009 [32].
Table 3.
Single Nucleotide Variants (SNVs), insertions and deletions within the Portuguese isolates.
Table 3.
Single Nucleotide Variants (SNVs), insertions and deletions within the Portuguese isolates.
| B. ceti isolates | DDE001 | DDE002 | DDE003 |
|---|---|---|---|
| DDE001 | --------------------------------------- | 213 SNVs, 12 insertions, 18 deletions | 216 SNVs, 16 insertions, 20 deletions |
| DDE002 | 213 SNVs, 12 insertions, 18 deletions | ------------------------------------ | 107 SNVs, 13 insertions, 11 deletions |
| DDE003 | 216 SNVs, 16 insertions, 20 deletions | 107 SNVs, 13 insertions, 11 deletions | -------------------------- |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



