Submitted:
07 May 2024
Posted:
08 May 2024
You are already at the latest version
Abstract
The symbiotic interaction between leguminous and Bradyrhizobium sp. SUTN9-2 mainly relies on the nodulation process through Nod factors (NFs), while the type IV secretion system (T4SS) acts as an alternative pathway in this symbiosis. Two copies of T4SS (T4SS1 and T4SS2) are located on the chromosome of SUTN9-2. ΔT4SS1 reduces both nodule number and nitrogenase activity in all SUTN9-2 nodulating legumes. The functions of three selected genes (copG1, traG1, and virD21) within the region of the T4SS1 were examined. We generated deleted mutants and tested in Vigna radiata cv. SUT4. ΔtraG1 and ΔvirD21 exhibited lower invasion efficiency at the early stages of root infection but could be lately restored. In contrast, ΔcopG1 completely hindered nodule organogenesis and nitrogenase activity in all tested legumes. ΔcopG1 showed low expression of nodulation gene and ttsI, but exhibited high expression levels of T4SS genes, traG1 and trbE1. The secreted proteins from ΔT4SS1 were down-regulated compared to the wild type. Although ΔcopG1 secreted several proteins after flavonoid induction, T3SS (nopP and nopX) and C4-dicarboxylate transporter (dct) were not detected. These results confirm the crucial role of the copG1 gene as a novel key regulator in the symbiotic relationship between SUTN9-2 and legumes.
Keywords:
Symbiosis
; Vigna radiata
; T4SS
; copG
1. Introduction
A complex exchange of signals between legumes and rhizobia bacteria is the key factor in the successful symbiosis between two partners. The establishment of nitrogen-fixing nodules is ensured by this intricate communication system, which is crucial for legumes to thrive in nitrogen-limited environments [1,2]. Two crucial mechanisms are necessary for the nodulation process of many rhizobia: (i) the classical mechanism of perception of the Nod factor (NF) and (ii) the type III secretion system (T3SS) which deliver effector proteins into plant cells [3,4]. NFs are well-studied while, we are just beginning to understand the roles of effectors/secretion systems. The secreted effector proteins from T3SS (T3Es) can have either a positive or negative impact on the symbiosis efficiency, depending on the plant species [5]. For example, Bradyrhizobium vignae ORS3257 contains multiple effector proteins crucial for modulating symbiotic properties in different Vigna species. NopT and NopAB play essential roles in nodulation in V. unguiculata and V. mungo. Whereas, NopP2 displayed incompatibility with V. radiata [6]. Beside the T3SS, others secretion systems have been identified in several rhizobia genera, including type IV and VI secretion systems (T4SS and T6SS) and these systems have also reported as the key determinant to the symbiotic interactions during infection processes. For instance, the T6SS of Rhizobium etli Mim1 and Bradyrhizobium sp. LmicA16 (A16) exhibit a positive effect on nodulation with their host [8,9]. In the case of T4SS, this secretion machinery is found in various bacteria, including Rhizobium. It functions as a molecular channel, allowing bacteria to transport diverse molecules across their cell envelope [10]. Various rhizobia, including Bradyrhizobium, Rhizobium, Sinorhizobium, and Mesorhizobium, possess T4SS which belongs to the tra/trb operon and could be found either in chromosome or plasmid depending on the bacterial strain [11,12,13,14]. Interestingly, most of bradyrhizobia harbor the tra/trb operon on the chromosome. Few studies about the role of T4SS of Bradyrhizobium in the symbiotic process have been reported. Previously, we found two clusters of T4SS (T4SS1 and T4SS2) located on the chromosome of Bradyrhizobium sp. SUTN9-2 with different gene arrangements (Figure S1). Additionally, a specialized gene arrangement consisting of copG, traG, and virD2 genes was observed in the T4SS gene cluster of Bradyrhizobium species. The T4SS evolutionary analysis of the Rhizobiales order, encompassing Bradyrhizobium, Rhizobium, Sinorhizobium, and Mesorhizobium through the traG gene phylogenetic tree, demonstrates a co-evolutionary trend between Bradyrhizobium and Mesorhizobium. Upon phylogenetic examination of the copG, traG, and virD2 combination genes in Bradyrhizobium, two copies of these clusters were divided into two clades within the bradyrhizobia group [15]. Moreover, copy 1 of copG, traG, and virD2 genes exhibits a close evolutionary association with B. yuanmingense BRP09, which are main rhizobia associated with cowpea and mung bean in the subtropical region of China [16], as well as with B. diazoefficiens USDA110, soybean inoculant [17]. Interestingly, T4SS1 mutant (deleted copG1, traG1, and virD21 fragment) retard nodulation in V. radiata cv. SUT4 and Crotalaria juncea and both the number of nodules and nitrogenase activity were decreased compared with the wild type. The results indicated that T4SS1 has a positive effect on the symbiotic interaction with tested plants [15]. To understand the role of T4SS1 in the symbiotic process, individual deletion mutants were generated in copG1, traG1 and virD21 genes (encoding a putative transcriptional factor gene, T4SS structural gene, and relaxase gene, respectively), and examined for their roles in the symbiotic interaction with V. radiata cv. SUT4 [18,19,20]. In addition to V. radiata cv. SUT4, the loss of nodulation ability was observed in different plant species of Genistoids, Dalbergioids, and Millettioids upon infection with ΔcopG1 (Table S1).
2. Materials and Methods
2.1. Bacterial Strains, Plasmids, and Growth Conditions
The bacterial strains and plasmids used in this study are listed in Table 1. Bradyrhizobium sp. SUTN9-2 was grown in arabinose-gluconate (AG) medium [21]. The derivative mutants ΔcopG1, ΔtraG1, and ΔvirD21 were supplemented with streptomycin (sm) at 200 μg/mL. Escherichia coli strains were grown at 37°C in Luria–Bertani (LB) medium. Antibiotic was added in to the medium as required at the following concentrations: 50 μg/mL kanamycin (km), 30 μg/mL nalidixic acid (nal), and 200 μg/mL streptomycin (sm).
2.2. Plasmid Construction and Gene Deletion
The deletion mutants of copG1, copG2, traG1, and virD21 genes in Bradyrhizobium sp. SUTN9-2 (GeneBank accession number LAXE00000001) were obtained as follows. The upstream and downstream regions of copG1 (up: 575 bp, dw: 841 bp), copG2 (up: 874 bp, dw:1043 bp), traG1 (up: 1060 bp, dw:921 bp), and virD21 (up: 944 bp, dw: 738 bp) genes were obtained by PCR using primers listed in Table 2. The target deletion genes in SUTN9-2 were obtained by double crossover. PCR fragments corresponding to the upstream and downstream flanking regions of the gene of interest were merged by overlap extension and introduced into a pNTPS129 plasmid harboring the sacB gene [23]. Then, an Ω cassette fragment (spectinomycin/streptomycin resistance genes) from pHP45 (omega) [24] was introduced between upstream and downstream flanking regions which were already cloned into pNTPS129. The restriction site for antibiotic insertion was HindIII for copG1 and BamHI for copG2, traG1, and virD21. The resulting plasmids were transferred into SUTN9-2 by triparental mating using pRK2013 as a helper plasmid [22] as described previously [15]. A single recombinant clone was obtained by antibiotic selection and PCR verification. Double recombinant clones were selected by culture on AG medium supplemented with 10% sucrose and 200 μg/mL sm. Candidate clones were verified for the loss of the sacB gene from pNTPS129, and the replacement of the Ω cassette was verified by PCR. All mutant strains were further investigated for nodulation efficiency in V. radiata cv. SUT4.
2.3. Nodulation Test and Acetylene Reduction Assay (ARA)
V. radiata cv. SUT4 seeds were surface-sterilized and germinated as previously described [26] and placed on 0.85% water agar at 28°C overnight. One-day-old germinated seedlings were transferred into Leonard’s jars containing sterilized vermiculite and liquid buffered nodulation media (BNM) [27]. At seven days after gemination, seedlings were inoculated with a bacterial suspension of Bradyrhizobium sp. SUTN9-2 or derivative mutants (1 ml per seedling; adjusted to OD600 = 0.8). Five plants per treatment were selected for nodule counting. The symbiotic phenotypes and nitrogen activity were measured at 7, 14, and 21 dpi.
Acetylene reduction assays (ARAs) were used to evaluate the nitrogenase activity. The root samples were transferred into test tubes, which were closed with a plastic stopper. Then, the samples were incubated with 10% (v/v) pure acetylene instead of air, which was withdrawn for 1 h at room temperature. A 1 ml sample was examined using gas chromatography (GC) with a PE-alumina-packed column to measure the conversion of acetylene (C2H2) to ethylene (C2H4). Detection was done at an injection temperature of 150°C and oven temperatures of 200°C and 50°C for flame ionization detection (FID) [28]. The experiment was conducted with five biological replicates per treatment. The nitrogenase activity is presented in nmol ethylene/h/plant dry weight [29].
The symbiotic profiles of ΔcopG1 were compared to SUTN9-2 using a growth pouch test and several leguminous plants, including Genistoids (Crotalaria juncea), Dalbergioids (Aeschynomene americana cv. Thai, Arachis hypogaea cv. Thainan 9 and A. hypogaea cv. Khonkaen 5), and Millettioids (Indigofera tinctoria, Macroptilium atropurpureum, V. radiata cv. SUT1, V. radiata cv. CN72, V. radiata cv. KUML4, V. radiata cv. CN36, V. radiata cv. KPS1, V. mungo cv. U thong 2 and, V. subterranean). Seeds were surface-sterilized and germinated as previously described [26,30]. Pouches were prepared [29] and supplemented with BNM medium. Seedlings were grown at two plants per pouch (Five pouch replicates per treatment) and inoculated with 1 ml per plant of a suspension containing OD600 = 0.8. Plants were grown under the conditions mentioned above.
2.4. Bacterial Induction, RNA Isolation, and qRT-PCR Analysis of Gene Expression
For bacterial induction, the mid-log phase of bacteria cultures including Bradyrhizobium sp. SUTN9-2 and copG1 mutant strains (OD600 = 0.4) were induced by 20 μM genistein at 28°C for 24 hours. Then, bacterial pellets were collected by centrifugation (4,000×g, at 4°C) for total RNA isolation. Total RNA was isolated from bacterial pellets using an RNeasy® Protect Cell Mini Kit (Qiagen, Chatsworth, CA, U.S.A.) according to the manufacturer’s instructions. Total RNA was treated at 37°C for 30 min with RNase-free DNase I (NEB, USA).
cDNA was synthesized using iScript™ Reverse Transcription Supermix for RT-qPCR (Bio-Rad Laboratories, Inc.) according to the manufacturers’ protocols. A cDNA concentration of 50 ng/µL was subjected to real-time PCR using specific primers (Table 2) for nodulation genes (nodA, nodB, nodC, nodD1, and nodD2), transcriptional regulator of T3SS (ttsI), T4SS structural genes (traG1 and trbE1), and other genes. qRT-PCR reactions were performed with Luna® Universal qPCR Master Mix (NEB, USA) by following the manufacturer’s protocol, and thermal cycling was conducted in a CFX Opus 96 Real-Time PCR System (Bio-Rad Laboratories, Inc.). The reactions were done in triplicate for each of the three biological replicates. Relative gene expression was analyzed by the comparative Ct method 2(-ΔΔCT), and 16s rRNA (accession number: JN578804) was used as internal control [11]. Three biological replicates were analyzed.
2.5. Protein Preparation, Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE) Analysis, and Protein Identification
Wild type (WT) and ΔcopG1 Bradyrhizobium sp. SUTN9-2 were grown in AG medium with shaking at 200 rpm and 30°C until they reached an OD600 of 1. One percent (v/v) of each starter was inoculated into 100 ml of AG medium with and without 20 µM genistein induction. Then, cultures were incubated at 30°C until they reached an OD600 of approximately 1.0. The bacterial supernatants were harvested by centrifugation at 4,000xg and 4°C for 1 h, followed by 8,000xg for 30 min. One milliliter of 1 M dithiothreitol (DTT) and 7.5 ml of phenol solution (equilibrated with 10 mM Tris HCl at pH 8.0 with ESTA) were added into 25 ml of fresh supernatant. The solution was vigorously mixed with a vortex before centrifugation at 8,000xg and 4°C for 30 min. The water phase was discarded, and the phenol phase was added into another 25 ml of supernatant, followed by vigorous mixing and centrifugation at 8,000xg and 4°C for 30 min. Next, 20 ml of methanol containing 300 µl of 8 M ammonium acetate and 400 µl of 1 M dithiothreitol were added to remove the phenol phase. The secreted protein was precipitated for overnight at -20°C. The solution was then centrifuged at 8,000xg and 4°C for 1 h, and the supernatant was discarded. After precipitation, the protein was washed with chilled 70% (v/v) ethanol and air-dried in a laminar flow before being dissolved in phosphate-buffered saline (PBS). Protein concentrations were determined using a plate reader and the manufacturer’s protocol (PanReac) according to the method of Bradford [31]. A standard calibration curve was constructed using 0 to 2 µg of bovine serum albumin (BSA). Denaturing SDS-PAGE was carried out according to the method of Laemmli [32], in which 10 µg each lane of protein were analyzed on a 12% SDS-PAGE gel. The protein samples were mixed with loading buffer containing β-mercaptoethanol and heated for 10 min before loading. Protein bands were stained with colloidal Coomassie brilliant blue R-250 for visualization of the expression of secreted protein. The protein bands which were observed on WT lane but not observed on ∆T4SS1 lane were cut protein identification by mass spectrometry. Breiftly, the protein bands were performed in-gel digestion by 12.5 ng/µL trypsin (mass spectrometry grade; Promega, USA). The extacted peptides were collected and dried in the Nitrogen Evaporator (Organomation, USA). Then, the peptide were reconstituted in 15 µl of 0.1% formic acid (FA) for LC/MS analysis. The LC–MS/MS system consists of a liquid chromatography part (Dionex Ultimate 3000, RSLCnano System) in combination with an captivespray ionization / mass spectrometer (Model Q-ToF Compact, Bruker, Germany) at the Proteomics Services, Faculty of Medical Technology, Mahidol University (Salaya Campus), Nakonpathum, Thailand. Mass spectral data from 300 to 1500 m/z was collected in the positive ionization mode. The most abundant peptide ions were analysed using MS/MS, to determine the peptide sequence. The peptide sequence was searched on the Uniprot database using the Mascot daemon version 2.6.0 (Matrix Science, London, UK) searching engine. The search parameters in Mascot daemon MS/MS Ions search included carbamidomethyl at cysteine residues as a fixed modification and oxidation on methionine as variable modifications. The peptide tolerance was set at ±1.6 Da, and the MS/MS fragment tolerance was set at ±0.8 Da. Protein hits were selected with a p-value of ≤ 0.05. The obtained results were examined against protein-NCBI database to identify and annotate proteins.
2.6. Microscopy
The nodule phenotypes and cross sections of representative nodules generated by the wild type (WT) or mutants were examined under stereomicroscope (LEIGA EZ4). For in-situ live/dead cell staining, the nodules were harvested and embedded with 5% agarose [33]. Three plants per treatment were selected for nodule sections with thickness of 40-50 µm were prepared with a VT1000S vibratome (Leica Nanterre, France) and incubated with live/dead staining solution (5 µM SYTO9 and 30 µM propidium iodide (PI) in PBS pH 7.0 buffer) for 30 min, followed by staining with 1X calcofluor white stain for 20 min. Sections were washed to remove the staining solution and mounted in 10% glycerol in 1X PBS buffer. After staining, nodules were observed by confocal microscopy using a Nikon Inverted Eclipse Ti-E Confocal Laser Scanning Microscope. Calcofluor was detected with emission at 460-500 nm, while SYTO9 and PI were detected at 510-570 nm and 600-650 nm, respectively [15]. Three nodules were randomly selected for nodule imaging and bacteroid observation.
2.7. Bioinformatics
Bradyrhizobium sp. SUTN9-2 genome sequences were obtained from the NCBI database (https://www.ncbi.nlm.nih.gov) and Genoscope (https://mage.genoscope.cns.fr) [35]. Multiple sequence alignments were determined by CLUSTALW (2.1) (https://www.genome.jp/tools-bin/clustalw). Domain architecture analysis was performed with the Simple Modular Architecture Research Tool (SMART) (https://smart.embl.de) [36] and InterPro (https://www.ebi.ac.uk/interpro) [37]. The annotation features and whole genome sequences were analyzed by SnapGene software (www.snapgene.com).
2.8. Statistical Analysis
All data were obtained from experiments performed in triplicate. For statistical analyses, one-way analysis of variance (ANOVA) followed by Tukey’s honestly significant difference (HSD) test (Tukey’s tests at P ≤ 0.05) and Student’s t-tests (P ≤ 0.05) were performed using SPSS software (SPSS version 22.0 windows: SPSS Inc., Chicago, IL) and GraphPad Prism statistical software (Version 10.0.3).
3. Results
3.1. Symbiotic Properties of ΔcopG1, ΔtraG1, and ΔvirD21 in Vigna Radiata cv. SUT4 Symbiosis
Differences between the wild type and mutants of Bradyrhizobium sp. SUTN9-2 were observed in terms of nodulation and nitrogenase activity in V. radiata cv. SUT4 (Figure 1). ΔtraG1 and ΔvirD21 induced a higher number of nodules on the plant tested than the wild type at 7 days post inoculation (dpi) (Figure 1Q), although the nodules produced displayed a white color (Figure 1I,M) instead of pink, indicating problems in nodule development. Moreover, there were higher numbers of dead cells in nodules inoculated with ΔtraG1 and ΔvirD21 in the symbiosome area (Figure 1J,N). At 21 dpi, there was no difference in the number of nodules obtained with ΔtraG1, ΔvirD21, and the wild type (Figure 1R).
In addition, there were different results of the nitrogenase activities in each mutant at 21 dpi, ΔtraG1 was identical to that of the wild type, while low nitrogenase activity was obtained with ΔvirD21 (Figure 1T). Interestingly, ΔcopG1 showed a significant effect on nodulation in that nodule formation was abolished (Figure 1E,G,Q-R). Although nodule organogenesis was not observed in the plant inoculated with ΔcopG1, both live and dead cells were detected in the cortex and vascular tissue instead (Figure 1H). According to the results, ΔcopG1 was able to infect plant cells, but it was no longer capable of surviving in host cells.
3.2. The copG Genes Are Involved in Nodulation Efficiency of Bradyrhizobium sp. SUTN9-2
The copG gene typically encodes the CopG protein, which is a transcriptional factor that consists of ribbon helix-turn-helix (RHH) motifs [18,34,38]. Bradyrhizobium sp. SUTN9-2 has two copies of copG genes located downstream of the traG and virD2 genes. These gene clusters are located in distinct locations on the SUTN9-2 chromosome. Although two copies of copG genes were present on the chromosome, they did not share the same gene sequences. The copG gene copies 1 and 2 (copG1 and copG2) revealed a low degree of similarity with 52.35% DNA sequence identity (Figure S2) and 51.77 % amino-acid sequence identity (Figure S3), which are different in both the N- and C-terminals. Domain architecture analysis identified CopG1 as an unidentified domain, which shows similarities with B. yuanmingense BRP09, CCBAU05623, and, more distantly, in P10 130. While, CopG2 was identified as a Pfam:RHH domain that is also found in B. diazoefficiens USDA110, B. diazoefficiens SEMIA5080, and B. japonicum J5 (Figure S4).
To gain a more complete understanding of the function of copG1 and copG2, we constructed ΔcopG2, inoculated it into V. radiata cv. SUT4, and compared it with ΔcopG1 and wild type strains (Figure 2). Contrary to ΔcopG1 (Figure 2B), ΔcopG2 was able to produce pink nodules (Figure 2C) that were smaller than those generated by the wild type (Figure 2A). At 14 and 21 dpi, ΔcopG2 produced the highest number of nodules compared to other strains (Figure 2G,H). Despite the high number of nodules generated by ΔcopG2, the nitrogenase activity was significantly lower than that of the wild type (Figure 2I,J). Confocal microscopic examination showed dead cells in nodules generated by ΔcopG2 (Figure 2F) in the symbiosome, in contrast with the wild type (Figure 2D). These findings indicated both copG1 and copG2 genes are essential for the symbiotic relationship between SUTN9-2 and legume plants. copG1 is necessary for nodulation, while copG2 is crucial for nitrogenase efficiency. Lack of copG2 leads to decreased nitrogenase activity, despite the presence of high nodule numbers.
3.3. The copG1 Gene Plays a Crucial Role in the Expression of Nodulation (nod) Genes and Transcriptional Regulator TtsI (ttsI)
To examine if copG1 affects the structuring of the NF backbone, transcript levels of nodABC and transcriptional activator nodD (nodD1 and nodD2) were examined with and without 20 μM genistein induction (Figure 3). The nodA, nodC, nodD1, and nodD2 genes were almost not expressed in ΔcopG1 (Figure 3A,C,D,E), while the expression of nodB was not affected by a mutation in copG1 (Figure 3B). The results indicated that copG1 modulate, either directly or indirectly, the expression of nod genes, especially the nodD gene (Figure 3D,E), which is a transcriptional activator of NFs [39]. Beside NF biosynthesis, NodD1 also activated the transcriptional regulator TtsI (ttsI), gene encoding for T3SS secretion and synthesis [40]. Similar to the nod genes, the expression level of ttsI was not determined in ΔcopG1 in all conditions (Figure 3F). The loss of nodule formation in ΔcopG1 may be caused by the suppression of NF synthesis and also T3SS due to the absence of nodD expression.
3.4. Bradyrhizobium sp. SUTN9-2 copG1 is Involved in the Repression of T4SS Structural Genes traG1 and trbE1
To understand the relationship of T4SS and copG1 gene more clearly, the gene expression fold changes were examined. T4SS with trbE1 and traG1 genes showed high expression levels under non-symbiotic conditions when copG1 was deleted (Figure 4A,B). These results indicated that copG1 of SUTN9-2 may act as a suppressor of trbE1 and traG1 under non-symbiotic conditions. However, under mimicked symbiotic conditions with 20 μM genistein induction, the expression of the trbE1 gene significantly decreased in ΔcopG1 compared to the wild type. The expression of the traG1 gene did not differ in both the wild type and the ΔcopG1 (Figure 4C,D). However, it is noteworthy that under mimicked symbiosis conditions, CopG1 can induce the expression of trbE1 and traG1, resulting in significantly higher levels than those observed under non-induction conditions. These results suggest that copG1 might act as a synergistic regulator of the T4SS gene in SUTN9-2 under flavonoid induction (Figure 4C,D).
3.5. Effect of T4SS and copG1 on Secreted Protein Pattern after 48 h With Genistein Induction
The secreted protein patterns of Bradyrhizobium sp. SUTN9-2, ∆T4SS1, and ∆copG1 with 20 µM genistein after 48 h induction were analyzed by SDS-PAGE. The results revealed distinct protein band patterns in the different conditions (Figure 5). The amino acid sequences of each selected band were examined using mass spectrometry (MS) with the MASCOT program. In the wild type with genistein induction (band 1) contain a protein matched with the T3SS translocon protein, NopX (27%), while this band was absent at the same position of ∆T4SS1. Similarly, a protein band was observed in the wild type with genistein induction (band 6), a match was found with the T3SS host specificity protein, NopP (18%), this protein was not observed in ∆T4SS1 (band 7) at the same position. While protein bands from the wild type (band 3) and ∆T4SS1 (band 4) were found a protein matched with Dct; C4-dicarboxylate ABC transporter (31%), but it was not observed in ∆copG1 (band 5). Nevertheless, ∆copG1 (band 5) was associated with amino acid ABC transporter substrate-binding protein glutamate/aspartate transporter subunit (38%).
qRT-PCR was performed to identify the gene expression of T3SS (nopP and nopX) and C4-dicarboxylate transporter (dct), which was not in secreted protein from ∆copG1. The results showed that the expression of T3Es (nopP and nopX) and C4-dicarboxylate transporter (dct) under genistein induction was down-regulated in ∆copG1 (Figure 6). This indicated that these genes required copG1 to mediate the regulation under genistein induction.
4. Discussion
At an early nodulation stage of V. radiata cv. SUT4, ΔtraG1 and ΔvirD21 generated a high number of nodules with smaller size compared to the wild type (Figure 1I,M,Q-R). The symbiosome space of ΔtraG1 and ΔvirD21 infecting nodules revealed some dead cells that were not found in the wild type under confocal microscopy (Figure 1J,N). According to the findings, T4SS is beneficial in the early stages of symbiotic interaction between SUTN9-2 and legumes. Bradyrhizobia have a TraG/Trb operon on the chromosome in the symbiosis island that is similar to that of mesorhizobia based on the traG gene’s phylogenetic and gene organization [15]. Beside the structural protein, various bacteria containing T4SS also identified ATPase/Coupling protein, VirD4/TraG and relaxase, VirD2 [20]. The traG is commonly found in conjugative plasmids that are responsible for horizontal gene transfer between bacteria. The traG gene required for encode T4SS component served as ATPase to generate energy during secretion [19]. In addition, TraG also act as a substrate receptor of T4SS called coupling protein, a substrate receptor that mediate the substrate such as effector proteins, DNA or DNA-protein complex through T4SS channel [7,10,41,42]. In the Pfam prediction, TraG protein was matched with Pfam family T4SS-DNA_transfer (PF02534), TrwB_AAD_bound (PF10412), and TraG-D_C (PF12696) (Figure S8A). The C-terminal of this protein is able to interact with the relaxosome, which is essential for DNA transfer and conjugation in bacteria [19,43,44]. In mesorhizobia, traG plays an important role in an early stage of infection, and its expression was observed during induction with root exudate and early nodules generated by M. mediterraneum Ca36T. Corresponding to mesorhizobia, traG1 of SUTN9-2 may play a crucial role in the beginning of symbiotic interaction with legumes [7]. In Agrobacterium, VirD2 protein is a part of the relaxase family that plays a crucial in conjugating and mobilizing plasmids that are required for translocation and integration of T-strands into recipient plant cells [45,46]. The conjugative transfer of ICEMlSymR7A in M. loti R7A requires VirD2 relaxase to initiate the rolling-circle replication [47]. VirD21 of SUTN9-2 possesses a domain of unknown function (DUF), DUF3363, which is an uncharacterized protein (Figure S8B). Although ΔvirD21 had no effect on the number of nodules of V. radiata cv. SUT4, but the activity of nitrogen fixation was reduced (Figure 1S,T).
Nodules generated by ΔvirD21 showed many uninfected cells (Figure 1N,P). This finding showed that the communication between SUTN9-2 and the legume at the beginning of nodule organogenesis plays an important role in enhancing infection efficiency and nitrogenase activity after infection. These results strongly indicated that the traG1 and virD21 genes may be necessary for symbiotic interaction during an early infection stage. Unlike ΔtraG1 and ΔvirD21, ΔcopG1 has an impact on the symbiotic interaction between SUTN9-2 and legumes because this mutant was unable to generate nodules with the tested plant (Figure 1E,G). Since, copG1 was located downstream of traG1 and virD21 within the same cluster, it is assumed that copG1 may share a common promoter with traG1 and virD21. This observation was supported by the previous study, where T4SS complementation successfully restored nodule formation [15]. Several bacteria, such as Pseudomonas aeruginosa [48], Streptococcus agalactiae [38], Vibrio cholerae [49], Bradyrhizobium sp., and Mesorhizobium sp. contain the copG gene in their genomes [15]. This gene encodes CopG protein, a small transcriptional repressor containing a helix-turn-helix motif domain, which is similar to that of regulatory repressors such as Mnt, Arc, and MetJ in Salmonella typhimurium bacteriophage P22 and Escherichia coli [18,50,51]. The CopG protein was first discovered in streptococcal plasmid pMV158 as a transcriptional repressor that interacts with RepB to control the copy number of the plasmid [34,38,52]. In addition, copper resistance was also demonstrated to be influenced by CopG in P. aeruginosa and V. cholerae [48,49].
In SUTN9-2, the CopG1 protein was classified as an uncharacterized conserved protein, while CopG2 was annotated as the Pfam;RHH_1 domain (PF01402), which may serve as a transcriptional regulator within the CopG family (Figure S4) [34]. The removal of both copG genes in SUTN9-2 resulted in distinct nodulation efficiency. Even without a nodule generated by ΔcopG1, it can still infect plants because we could monitor both live and dead cells within plant tissues. Surprisingly, live cells were found mostly in the vascular bundle tissue, which is similar to how endophytic bacteria behave. These findings imply that copG1 may be crucial for SUTN9-2 in protecting the survival of bacterial cells in the host plant. Bacteria can evolve and adapt to their environment through horizontal gene transfer, which is usually facilitated by conjugation. Conjugation is a significant biological process as it is the primary way to spread antibiotic resistance genes [53]. Integrative and conjugative elements (ICEs) are another essential mechanism that contributes to conjugation. ICEs are recognized as elements encoded for excision and transferred by conjugation and integration, regardless of the specific mechanisms involved [54]. The T4SS found in SUTN9-2 is classified as a tra/trb operon and is recognized for its crucial role in facilitating conjugal transfer. Although SUTN9-2 lacks a conjugation plasmid, ICE is still present on the chromosome. The genes encoding the T4SS1 cluster are presented in this ICE, which is an alternative mechanism of genetic exchange in this bacterial strain [15]. In order to study the impaired nodulation phenotype of ΔcopG1 in V. radiata cv. SUT4, we analyzed the expression of nod genes with and without genistein induction. Common nod genes in SUTN9-2 including nodA and nodC genes were not expressed even with a lack of copG1 under flavonoid induction condition, but it did not affect nodB expression. Additionally, copG1 acts as a stimulator for nodD1 and nodD2, which are the primary transcription factors responsible for NF production. In addition to nod genes, the expression level of ttsI, which was not determined in ΔcopG1. TtsI protein is a transcriptional regulator (previously called y4xI) is activated by flavonoids and NodD1 that bind to conserved sequences called tts-boxes [40,55,56]. This protein controlled the genes responsible for T3SS synthesis and effector protein secretion which have an effect for symbiosis interaction between rhizobium-legumes interaction depends on plant host [6,57]. During symbiotic interaction, SUTN9-2 required copG1 to mediate the expression of nod genes and ttsI under flavonoid induction. These results indicate that CopG1 may positively mediate the expression of nod genes via NodD activation before stimulating NF production, nodule organogenesis and T3SS. Furthermore, copG1 plays a role as a repressor in T4SS gene expression, suppressing trbE and traG gene expression under flavonoid stimulation (Figure 4C,D). In contrast, these genes were not affected by flavonoids in the absence of the copG1 gene.
The protein expression profiles of SUTN9-2, ∆T4SS1 and ∆copG1 with genistein treatment were analyzed by SDS-PAGE. The results revealed distinct protein band patterns in the different conditions (Figure 5). ∆copG1 exhibited a deficiency in producing nodules in various plant species and a striking increase in protein expression compared to the wild type. According to the analysis of copG1 domain protein (Figure S4), CopG1 was predicted to be a transcriptional regulator which might play a role in the regulation of gene expression. The ΔcopG1 lane appears to have much more protein intensity overall because the proportion of protein in this lane might be less than other lanes. So, 10 µg might be shown higher band intensity. A comparative proteomic analysis of the whole secretome should be conducted further to identify additional target proteins involved in this interaction. It was found that several proteins were secreted, but the C4-dicarboxylate transport system (dct) protein was not identified in ∆copG1 and this result was corresponded to the down-regulated of dct gene quantified by qRT-PCR (Figure 6). The dct gene play a crucial role for symbiosis in numerous rhizobia [58]. For example, the dct mutant of S. meliloti and R. trifolii can generate ineffective nodules with the host legume [59,60]. Besides the dct gene, there are other genes that are expressed in the same pattern, including nopX and nopP, which are also essential for symbiosis (Figure 6). NopX is a component of T3SS as a translocation pore (translocon) apparatus that is important for host-specific interaction between the rhizobium and host plant. The NGRΔnopX has a significant effect on the nodule number because this mutant forms fewer nodules in all plant species tested, including Flemingia congesta, Tephrosia vogelii, Pachyrhizus tuberosus, and Lablab purpureus [61]. NopP is a T3SS effector protein that is phosphorylated by plant kinases [62]. A lack of NopP in Rhizobium sp. NGR234 reduces the capacity of nodule organogenesis in tropical legumes. This indicates a positive effect of NopP on symbiosis [61]. NopP of B. diazoefficiens USDA122 is necessary and causes Rj2-dependent incompatibility [63]. The T3SS of SUTN9-2 has no impact on the symbiotic relationship with V. radiata [11]. However, based on the protein secretion results of T3Es (NopP and NopX) and nodD gene expression, it is evident that copG1 regulates the function of nodD and T3SS. Previous reports indicated that nodD controls the function of nod cluster by binding to the nod box region. Similarly, nodD can regulate the T3SS function by binding to ttsI [55]. Therefore, the results of this experiment confirm that CopG1 controls the function of nodD, influencing the expression of the nod cluster genes and T3SS. Perhaps, CopG1 is a crucial factor in the early stage of legume and SUTN9-2 communication. It is plausible that the regulatory system governing the expression of nod genes does not solely depend on the interaction between flavonoids and NodD. Another factor, CopG1, also collaborates with flavonoids and NodD in regulating the expression of nod genes and T3SS. Carbon and nitrogen metabolism are the primary mechanisms that are necessary for the exchange of nutrients between plant and bacteria partners. The proteins secreted from ∆copG1 matched with the periplasmic binding proteins of the glutamate/aspartate ABC transporter. Glutamate is a significant contributor to the total metabolite content, which plays an essential role in nitrogen metabolism, amino acid metabolism, transamination, and carbon sources [64,65]. During symbiosis, the main carbon source utilized by rhizobia is C4-dicarboxylic acid [66,67]. In the mimicked symbiotic conditions, ∆copG1 lost the ability to establish symbiotic interaction. Thereafter, the increasing of glutamate/aspartate ABC transporter may promote carbon and nitrogen uptake to support bacterial cell survival, but it is not necessary for symbiotic interaction. This again suggests that copG1 may act as a regulator of nodD and T4SS gene expression under symbiosis conditions.
5. Conclusions
The T4SS1 containing copG1, traG1, and virD21 has beneficial effects on symbiotic interactions with diverse legumes. The deletion of each gene in a unique cluster showed a distinctive effect on symbiosis. The early stage of infection and nodulation is influenced by traG1 and virD21, while nodulation with legume hosts was not efficient in the absence of copG1. While, copG1 is necessary for nodulation, the ΔcopG2 results in reduced nitrogenase activity, despite maintaining high nodule numbers. This underscores the essential role of copG2 in nitrogenase efficiency. Overall, both copG1 and copG2 genes play crucial roles in the symbiotic relationship between SUTN9-2 and legume plants, contributing to effective nitrogen fixation during symbiosis. The expression of nodulation (nod) genes including common nodA, nodC, nodD1, and nodD2 and ttsI were not detected with and without genistein induction in ΔcopG1, while the T4SS genes in ΔcopG1 were highly expressed in non-induction conditions. Interestingly, the absence of copG1 with the induction of flavonoid led to a low expression level of T4SS genes. The results suggest that copG1 may play an important role in nodulation, which is a crucial for the symbiotic interaction between Bradyrhizobium sp. SUTN9-2 and legumes. Additionally, copG1 served as a suppressor of T4SS genes under non-induction conditions and was required to stimulate the expression of T4SS genes through flavonoid induction. copG1 also acted as a suppressor of secreted protein under flavonoid induction conditions. In addition, a lack of copG1 led to suppressed expression of nopX, nopP, and dct, which are important for infection and nodulation during symbiosis. Thus, copG1 is most likely responsible for regulation via functions in T3SS, nodD regulation, and the carbon and nitrogen exchange systems, which are significant for SUTN9-2 during symbiosis.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure S1: title; Table S1: title; Video S1: title.
Acknowledgments
This work was supported by (i) The Royal Golden Jubilee Ph.D. Programme (RGJ) scholarship under the Thailand Research Fund (TRF), (ii) Suranaree University of Technology (SUT), National Science, Research, and Innovation Fund (NSRF), (iii) the NSRF via the Program Management Unit for Human Resources & Institutional Development, Research, and Innovation (grant number B16F640113), (iv) the Office of the Permanent Secretary of the Ministry of Higher Education, Science, Research, and Innovation, and (v) National Research Council of Thailand (NRTC) and Suranaree University of Technology (grant number N42A650322).
References
- Dénarié, J.; Debellé, F.; Promé, J.-C. Rhizobium lipo-chitooligosaccharide nodulation factors: signaling molecules mediating recognition and morphogenesis Annu. Rev. Biochem. 1996, 65, 503–535.
- Hanin, M.; Jabbouri, S.; Quesada-Vincens, D.; Freiberg, C.; Perret, X.; Promé, J. -C.; Broughton, W.J.; Fellay, R. Sulphation of Rhizobium sp. NGR234 Nod-Factors is dependent on noeE, a new host-specificity gene. Mol. Microbiol. 1997, 24, 1119–1129.
- Fauvart, M.; Michiels, J. Rhizobial secreted proteins as determinants of host specificity in the rhizobium-legume symbiosis. FEMS Microbiol. Lett. 2008, 285, 1-9.
- Nelson, M.S.; Sadowsky, M.J. Secretion systems and signal exchange between nitrogen-fixing rhizobia and legumes. Front. Plant Sci. 2015, 6, 491. [CrossRef]
- Jiménez-Guerrero, I.; Medina, C.; Vinardell, J.M.; Ollero, F.J.; López-Baena, F.J. The rhizobial type 3 secretion system: The Dr. Jekyll and Mr. Hyde in the rhizobium–legume symbiosis. Int. J. Mol. Sci. 2022, 23, 11089. [CrossRef]
- Songwattana, P.; Chaintreuil, C.; Wongdee, J.; Teulet, A.; Mbaye, M.; Piromyou, P.; Gully, D.; Fardoux, J.; Zoumman, A.M.A.; Camuel, A.; et al. Identification of type III effectors modulating the symbiotic properties of Bradyrhizobium vignae strain ORS3257 with various Vigna species. Sci. Rep. 2021, 11, 4874.
- Paço, A.; Da-Silva, J.R.; Eliziário, F.; Brígido, C.; Oliveira, S.; Alexandre, A. traG gene is conserved across Mesorhizobium spp. able to nodulate the same host plant and expressed in response to root exudates. BioMed Res. Int. 2019, 2019, 3715271. [CrossRef]
- Salinero-Lanzarote, A.; Pacheco-Moreno, A.; Domingo-Serrano, L.; Durán, D.; Ormeño-Orrillo, E.; Martínez-Romero, E.; Albareda, M.; Palacios, J.M.; Rey, L. Type VI secretion system of Rhizobium etli Mim1 has a positive effect in symbiosis. FEMS Microbiol. Ecol. 2019, 95, fiz054. [CrossRef]
- Tighilt, L.; Boulila, F.; De Sousa, B.F.S.; Giraud, E.; Ruiz-Argüeso, T.; Palacios, J.M.; Imperial, J.; Rey, L. The Bradyrhizobium sp. LmicA16 type VI secretion system is required for efficient nodulation of Lupinus spp. Microb. Ecol. 2022, 84, 844–855. [CrossRef]
- Fronzes, R.; Christie, P.J.; Waksman, G. The structural biology of type IV secretion systems. Nat. Rev. Microbiol. 2009, 7, 703–714. [CrossRef]
- Piromyou, P.; Songwattana, P.; Teamtisong, K.; Tittabutr, P.; Boonkerd, N.; Tantasawat, P.A.; Giraud, E.; Göttfert, M.; Teaumroong, N. Mutualistic co-evolution of T3SSs during the establishment of symbiotic relationships between Vigna radiata and bradyrhizobia. MicrobiologyOpen 2019, 8, e00781.
- Kaneko, T.; Maita, H.; Hirakawa, H.; Uchiike, N.; Minamisawa, K.; Watanabe, A.; Sato, S. Complete genome sequence of the soybean symbiont Bradyrhizobium japonicum strain USDA6T. Genes 2011, 2, 763–787.
- Okazaki, S.; Noisangiam, R.; Okubo, T.; Kaneko, T.; Oshima, K.; Hattori, M.; Teamtisong, K.; Songwattana, P.; Tittabutr, P.; Boonkerd, N.; et al. Genome analysis of a novel Bradyrhizobium sp. DOA9 carrying a symbiotic plasmid. PLOS ONE 2015, 10, e0117392. [CrossRef]
- Cytryn, E.J.; Jitacksorn, S.; Giraud, E.; Sadowsky, M.J. Insights learned from pBTAi1, a 229-Kb accessory plasmid from Bradyrhizobium sp. strain BTAi1 and prevalence of accessory plasmids in other Bradyrhizobium sp. strains. ISME J. 2008, 2, 158–170.
- Wangthaisong, P.; Piromyou, P.; Songwattana, P.; Wongdee, J.; Teamtaisong, K.; Tittabutr, P.; Boonkerd, N.; Teaumroong, N. The type IV secretion system (T4SS) mediates symbiosis between Bradyrhizobium sp. SUTN9-2 and legumes. Appl. Environ. Microbiol. 2023, 89, e00040-23.
- Zhang, Y.F.; Wang, E.T.; Tian, C.F.; Wang, F.Q.; Han, L.L.; Chen, W.F.; Chen, W.X. Bradyrhizobium elkanii , Bradyrhizobium yuanmingense and Bradyrhizobium japonicum are the main rhizobia associated with Vigna unguiculata and Vigna radiata in the subtropical region of China. FEMS Microbiol. Lett. 2008, 285, 146–154.
- Albareda, M.; Rodríguez-Navarro, D.N.; Temprano, F.J. Soybean inoculation: dose, N fertilizer supplementation and rhizobia persistence in sSoil. Field Crops Res. 2009, 113, 352–356.
- Costa, M.; Solà, M.; Del Solar, G.; Eritja, R.; Hernández-Arriaga, A.M.; Espinosa, M.; Gomis-Rüth, F.X.; Coll, M. Plasmid transcriptional repressor CopG oligomerises to render helical superstructures unbound and in complexes with oligonucleotides. J. Mol. Biol. 2001, 310, 403–417. [CrossRef]
- Schröder, G.; Krause, S.; Zechner, E.L.; Traxler, B.; Yeo, H.-J.; Lurz, R.; Waksman, G.; Lanka, E. TraG-like proteins of DNA transfer systems and of the Helicobacter pylori type IV secretion system: inner membrane gate for exported substrates? J. Bacteriol. 2002, 184, 2767–2779.
- Tegtmeyer, N.; Linz, B.; Yamaoka, Y.; Backert, S. Unique TLR9 activation by Helicobacter pylori depends on the Cag T4SS, but not on VirD2 relaxases or VirD4 coupling proteins. Curr. Microbiol. 2022, 79, 121. [CrossRef]
- Sadowsky, M.J.; Tully, R.E.; Cregan, P.B.; Keyser, H.H. Genetic Diversity in Bradyrhizobium japonicum serogroup 123 and its relation to genotype-specific nodulation of soybean. Appl. Environ. Microbiol. 1987, 53, 2624–2630. [CrossRef]
- Ditta, G.; Stanfield, S.; Corbin, D.; Helinski, D.R. Broad host range DNA cloning system for Gram-negative bacteria: construction of a gene bank of Rhizobium meliloti. Proc. Natl. Acad. Sci. 1980, 77, 7347–7351. [CrossRef]
- Tsai, J.-W.; Alley, M.R.K. Proteolysis of the McpA chemoreceptor does not require the Caulobacter major chemotaxis operon. J. Bacteriol. 2000, 182, 504–507. [CrossRef]
- Blondelet-Rouault, M.-H.; Weiser, J.; Lebrihi, A.; Branny, P.; Pernodet, J.-L. Antibiotic resistance gene cassettes derived from the π interposon for use in E. coli and Streptomyces. Gene 1997, 190, 315–317.
- Muyzer, G.; De Waal, E.C.; Uitterlinden, A.G. Profiling of complexmicrobial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700.
- Teamtisong, K.; Songwattana, P.; Noisangiam, R.; Piromyou, P.; Boonkerd, N.; Tittabutr, P.; Minamisawa, K.; Nantagij, A.; Okazaki, S.; Abe, M.; et al. Divergent nod-containing Bradyrhizobium sp. DOA9 with a megaplasmid and its host range. Microbes Environ. 2014, 29, 370–376. [CrossRef]
- Ehrhardt, D.; Atkinson, E.; Long depolarization of alfalfa root hair membrane potential by Rhizobium meliloti Nod factors. Science 1992, 256, 998–1000. [CrossRef]
- Renier, S.; Hébraud, M.; Desvaux, M. Molecular biology of surface colonization by Listeria monocytogenes : an additional facet of an opportunistic Gram-positive foodborne pathogen. Environ. Microbiol. 2011, 13, 835–850. [CrossRef]
- Somasegaran, P.; Hoben, H.J. Handbook for rhizobia; Springer New York: New York, NY, 1994; ISBN 978-1-4613-8377-2.
- Phimphong, T.; Sibounnavong, P.; Phommalath, S.; Wongdee, J.; Songwattana, P.; Piromyou, P.; Greetatorn, T.; Boonkerd, N.; Tittabutr, P.; Teaumroong, N. Selection and evaluation of Bradyrhizobium inoculum for peanut, Arachis hypogea production in the Lao People’s Democratic Republic. J. Appl. Nat. Sci. 2023, 15, 137–154.
- Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254.
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [CrossRef]
- Haag, A.F.; Baloban, M.; Sani, M.; Kerscher, B.; Pierre, O.; Farkas, A.; Longhi, R.; Boncompagni, E.; Hérouart, D.; Dall’Angelo, S.; et al. Protection of Sinorhizobium against host cysteine-rich antimicrobial peptides is critical for symbiosis. PLoS Biol. 2011, 9, e1001169. [CrossRef]
- Acebo, P.; García De Lacoba, M.; Rivas, G.; Andreu, J.M.; Espinosa, M.; Solar, G.D. Structural features of the plasmid pMV158-encoded transcriptional repressor CopG, a protein sharing similarities with both helix-turn-helix and β-sheet DNA binding proteins. Proteins Struct. Funct. Genet. 1998, 32, 248–261.
- Vallenet, D.; Calteau, A.; Dubois, M.; Amours, P.; Bazin, A.; Beuvin, M.; Burlot, L.; Bussell, X.; Fouteau, S.; Gautreau, G.; et al. MicroScope: an integrated platform for the annotation and exploration of microbial gene functions through genomic, pangenomic and metabolic comparative analysis. Nucleic Acids Res. 2020, gkz926. [CrossRef]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: recent updates, new developments and status in 2020. Nucleic Acids Res. 2021, 49, D458–D460. [CrossRef]
- Paysan-Lafosse, T.; Blum, M.; Chuguransky, S.; Grego, T.; Pinto, B.L.; Salazar, G.A.; Bileschi, M.L.; Bork, P.; Bridge, A.; Colwell, L.; et al. InterPro in 2022. Nucleic Acids Res. 2023, 51, D418–D427.
- Gomis-Ruth, F.X. The structure of plasmid-encoded transcriptional repressor CopG unliganded and bund to its operator. EMBO J. 1998, 17, 7404–7415.
- Heidstra, R.; Bisseling, T. Nod factor-induced host responses and mechanisms of Nod factor perception. New Phytol. 1996, 133, 25–43. [CrossRef]
- Marie, C.; Deakin, W.J.; Ojanen-Reuhs, T.; Diallo, E.; Reuhs, B.; Broughton, W.J.; Perret, X. TtsI, a key regulator of Rhizobium species NGR234 is required for type III-dependent protein secretion and Synthesis of Rhamnose-Rich Polysaccharides. Mol. Plant-Microbe Interactions. 2004, 17, 958–966. [CrossRef]
- Cascales, E.; Atmakuri, K.; Sarkar, M.K.; Christie, P.J. DNA Substrate-Induced Activation of the Agrobacterium VirB/VirD4 Type IV Secretion System. J. Bacteriol. 2013, 195, 2691–2704. [CrossRef]
- Christie, P.J.; Whitaker, N.; González-Rivera, C. Mechanism and structure of the bacterial type IV secretion systems. Biochim. Biophys. Acta BBA - Mol. Cell Res. 2014, 1843, 1578–1591. [CrossRef]
- Gunton, J.E.; Gilmour, M.W.; Baptista, K.P.; Lawley, T.D.; Taylor, D.E. Interaction between the co-inherited TraG coupling protein and the TraJ membrane-associated protein of the H-plasmid conjugative DNA transfer system resembles chromosomal DNA translocases. Microbiology 2007, 153, 428–441. [CrossRef]
- Lu, J.; Wong, J.J.W.; Edwards, R.A.; Manchak, J.; Frost, L.S.; Glover, J.N.M. Structural basis of specific TraD-TraM recognition during F plasmid-mediated bacterial conjugation. Mol. Microbiol. 2008, 70, 89–99.
- Byrd, D.R.; Matson, S.W. Nicking by transesterification: the reaction catalysed by a relaxase. Mol. Microbiol. 1997, 25, 1011–1022. [CrossRef]
- Van Kregten, M.; Lindhout, B.I.; Hooykaas, P.J.J.; Van Der Zaal, B.J. Agrobacterium -mediated T-DNA transfer and integration by minimal VirD2 consisting of the relaxase domain and a type IV secretion system translocation signal. Mol. Plant-Microbe Interactions. 2009, 22, 1356–1365.
- Ramsay, J.P.; Sullivan, J.T.; Stuart, G.S.; Lamont, I.L.; Ronson, C.W. Excision and transfer of the Mesorhizobium loti R7A symbiosis island requires an integrase IntS, a novel recombination directionality factor RdfS, and a putative relaxase RlxS. Mol. Microbiol. 2006, 62, 723–734.
- Hausrath, A.C.; Ramirez, N.A.; Ly, A.T.; McEvoy, M.M. The bacterial copper resistance protein CopG contains a cysteine-bridged tetranuclear copper cluster. J. Biol. Chem. 2020, 295, 11364–11376. [CrossRef]
- Marrero, K.; Sánchez, A.; González, L.J.; Ledón, T.; Rodríguez-Ulloa, A.; Castellanos-Serra, L.; Pérez, C.; Fando, R. Periplasmic proteins encoded by VCA0261–0260 and VC2216 genes together with copA and cueR products are required for copper tolerance but not for virulence in Vibrio cholerae. Microbiology 2012, 158, 2005–2016. [CrossRef]
- Breg, J.N.; Van Opheusden, J.H.J.; Burgering, M.J.M.; Boelens, R.; Kaptein, R. Structure of Arc repressor in solution: evidence for a family of β-sheet DNA-binding proteins. Nature 1990, 346, 586–589.
- Somers, W.S.; Phillips, S.E.V. Crystal structure of the Met repressor–operator complex at 2.8 Å resolution reveals DNA recognition by β-strands. Nature 1992, 359, 387–393.
- del Solar, G.; Hernández-Arriaga, A.M.; Gomis-Rüth, F.X.; Coll, M.; Espinosa, M. A Genetically economical family of plasmid-encoded transcriptional repressors involved in control of plasmid copy number. J. Bacteriol. 2002, 184, 4943–4951. [CrossRef]
- Guglielmini, J.; Quintais, L.; Garcillán-Barcia, M.P.; de la Cruz, F.; Rocha, E.P.C. The repertoire of ICE in prokaryotes underscores the unity, diversity, and ubiquity of conjugation. PLoS Genet. 2011, 7, e1002222. [CrossRef]
- Bellanger, X.; Payot, S.; Leblond-Bourget, N.; Guédon, G. Conjugative and mobilizable genomic islands in bacteria: evolution and diversity. FEMS Microbiol. Rev. 2014, 38, 720–760. [CrossRef]
- Krause, A.; Doerfel, A.; Göttfert, M. Mutational and transcriptional analysis of the type III secretion system of Bradyrhizobium japonicum. Mol. Plant-Microbe Interactions. 2002, 15, 1228–1235. [CrossRef]
- Kobayashi, H.; Graven, Y.N.; Broughton, W.J.; Perret, X. Flavonoids induce temporal shifts in gene-expression of nod -box controlled loci in Rhizobium sp. NGR234. Mol. Microbiol. 2004, 51, 335–347.
- Teulet, A.; Camuel, A.; Perret, X.; Giraud, E. Theversatile roles of type III secretion systems in Rhizobium-legume symbioses. Annu. Rev. Microbiol. 2022, 76, 45–65.
- Yurgel, S.N.; Kahn, M.L. Dicarboxylate transport by rhizobia. FEMS Microbiol. Rev. 2004, 28, 489–501. [CrossRef]
- Ronson, C.W.; Lyttleton, P.; Robertson, J.G. C4 -dicarboxylate transport mutants of Rhizobium trifolii form ineffective nodules on Trifolium Repens. Proc. Natl. Acad. Sci. 1981, 78, 4284–4288.
- Yurgel, S.N.; Kahn, M.L. Sinorhizobium meliloti dctA mutants with partial ability To transport dicarboxylic acids. J. Bacteriol. 2005, 187, 1161–1172. [CrossRef]
- Marie, C.; Deakin, W.J.; Viprey, V.; Kopciñska, J.; Golinowski, W.; Krishnan, H.B.; Perret, X.; Broughton, W.J. Characterization of Nops, nodulation outer proteins, secreted via the type III secretion system of NGR234. Mol. Plant-Microbe Interactions. 2003, 16, 743–751. [CrossRef]
- Bartsev, A.V.; Boukli, N.M.; Deakin, W.J.; Staehelin, C.; Broughton, W.J. Purification and phosphorylation of the effector protein NopL from Rhizobium sp. NGR234. FEBS Lett. 2003, 554, 271–274. [CrossRef]
- Sugawara, M.; Takahashi, S.; Umehara, Y.; Iwano, H.; Tsurumaru, H.; Odake, H.; Suzuki, Y.; Kondo, H.; Konno, Y.; Yamakawa, T.; et al. ariation in bradyrhizobial NopP effector determines symbiotic incompatibility with Rj2-soybeans via effector-triggered immunity. Nat. Commun. 2018, 9, 3139.
- Streeter, J.G. Effect of nitrate on the organic acid and amino acid composition of legume nodules. Plant Physiol. 1987, 85, 774–779. [CrossRef]
- Forde, B.G.; Lea, P.J. Glutamate in plants: metabolism, regulation, and signalling. J. Exp. Bot. 2007, 58, 2339–2358. [CrossRef]
- Finan, T.M.; Wood, J.M.; Jordan, D.C. Symbiotic properties of C4-dicarboxylic acid transport mutants of Rhizobium leguminosarum. J. Bacteriol. 1983, 154, 1403–1413. [CrossRef]
- Jording, D.; Sharma, P.K.; Schmidt, R.; Engelke, T.; Uhde, C.; Pühler, A. Regulatory aspects of the C4-dicarboxylate transport in Rhizobium meliloti: transcriptional activation and dependence on effectave symbiosis. J. Plant Physiol. 1993, 141, 18–27. [CrossRef]
Figure 1.
Symbiotic phenotype of Bradyrhizobium sp. SUTN9-2 mutants during symbiosis with Vigna radiata cv. SUT4. Nodule phenotype at 7 and 21 dpi generated by wild type (A, C), ΔcopG1 (E, G), ΔtraG1 (I, K) and ΔvirD21 (M, O). Cytological analysis of the nodules (at 7 dpi and 21 dpi) induced by SUTN9-2 with wild type (B, D), ΔcopG1 (F, H), ΔtraG1 (J, L) and ΔvirD21 (N, P) observed by confocal microscopy after staining with propidium iodide, PI (red; infected plant nuclei and dead bacteria), SYTO9 (green: live bacteria), and calcofluor (blue: plant cell wall). Number of nodules at 7 dpi (Q) and 21 dpi (R). Nitrogen fixation activity determined by the acetylene reduction assay (ARA) of plants infected with the indicated bacterial mutants at 7 dpi (S) and 21 dpi (T). Values represent mean ± SD (n = 5). Scale bars; white bars indicate 1 mm, yellow bars indicate 100 µm (20X) and 50 µm (40X). P values based on Tukey’s test (* P <0.05,** P<0.01,*** P < 0.001).
Figure 1.
Symbiotic phenotype of Bradyrhizobium sp. SUTN9-2 mutants during symbiosis with Vigna radiata cv. SUT4. Nodule phenotype at 7 and 21 dpi generated by wild type (A, C), ΔcopG1 (E, G), ΔtraG1 (I, K) and ΔvirD21 (M, O). Cytological analysis of the nodules (at 7 dpi and 21 dpi) induced by SUTN9-2 with wild type (B, D), ΔcopG1 (F, H), ΔtraG1 (J, L) and ΔvirD21 (N, P) observed by confocal microscopy after staining with propidium iodide, PI (red; infected plant nuclei and dead bacteria), SYTO9 (green: live bacteria), and calcofluor (blue: plant cell wall). Number of nodules at 7 dpi (Q) and 21 dpi (R). Nitrogen fixation activity determined by the acetylene reduction assay (ARA) of plants infected with the indicated bacterial mutants at 7 dpi (S) and 21 dpi (T). Values represent mean ± SD (n = 5). Scale bars; white bars indicate 1 mm, yellow bars indicate 100 µm (20X) and 50 µm (40X). P values based on Tukey’s test (* P <0.05,** P<0.01,*** P < 0.001).
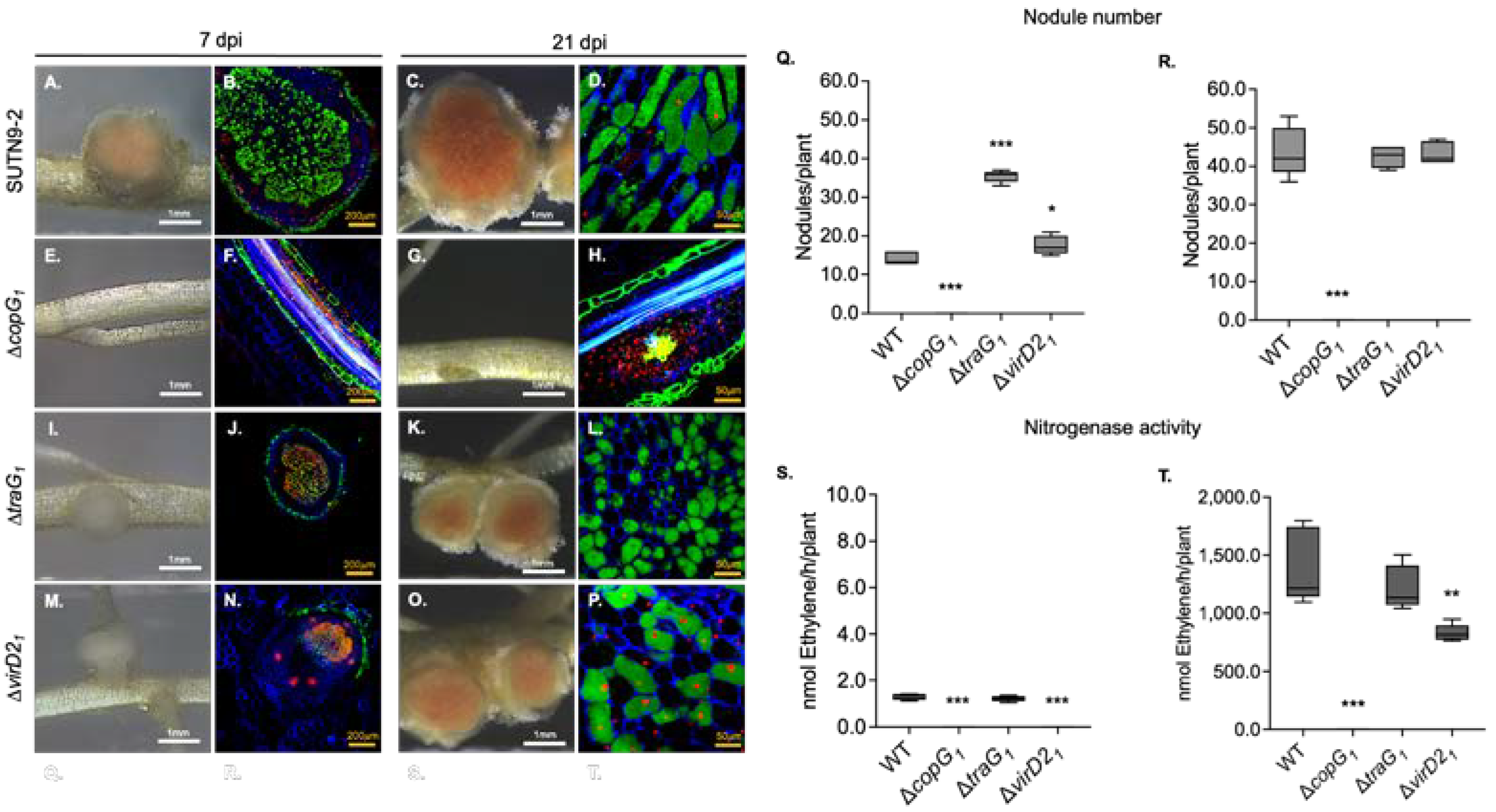
Figure 2.
Derivative copG mutants of Bradyrhizobium sp. SUTN9-2 presenting different symbiotic interaction with Vigna radiata cv. SUT4. Nodule phenotypes induced by wild type (A), ΔcopG1 (B), and ΔcopG2 (C). Cytological analysis of live/dead cells of section nodule infected with wild type (D), ΔcopG1 (E), and ΔcopG2 (F) at 14 dpi were observed with confocal microscopy, and bacteroids were stained with PI, SYTO9, and calcofluor-white. Number of nodules at 14 dpi (G) and 21 dpi (H). Nitrogen fixation activity determined by ARA of plants infected with the indicated bacterial mutants at 14 (I) and 21 dpi (J). Values represent mean ± SD (n = 5). Scale bars; white bars indicate 1 mm and yellow bars indicate 50 µm (40X). P values based on Tukey’s test (** P <0.01, *** P <0.001).
Figure 2.
Derivative copG mutants of Bradyrhizobium sp. SUTN9-2 presenting different symbiotic interaction with Vigna radiata cv. SUT4. Nodule phenotypes induced by wild type (A), ΔcopG1 (B), and ΔcopG2 (C). Cytological analysis of live/dead cells of section nodule infected with wild type (D), ΔcopG1 (E), and ΔcopG2 (F) at 14 dpi were observed with confocal microscopy, and bacteroids were stained with PI, SYTO9, and calcofluor-white. Number of nodules at 14 dpi (G) and 21 dpi (H). Nitrogen fixation activity determined by ARA of plants infected with the indicated bacterial mutants at 14 (I) and 21 dpi (J). Values represent mean ± SD (n = 5). Scale bars; white bars indicate 1 mm and yellow bars indicate 50 µm (40X). P values based on Tukey’s test (** P <0.01, *** P <0.001).
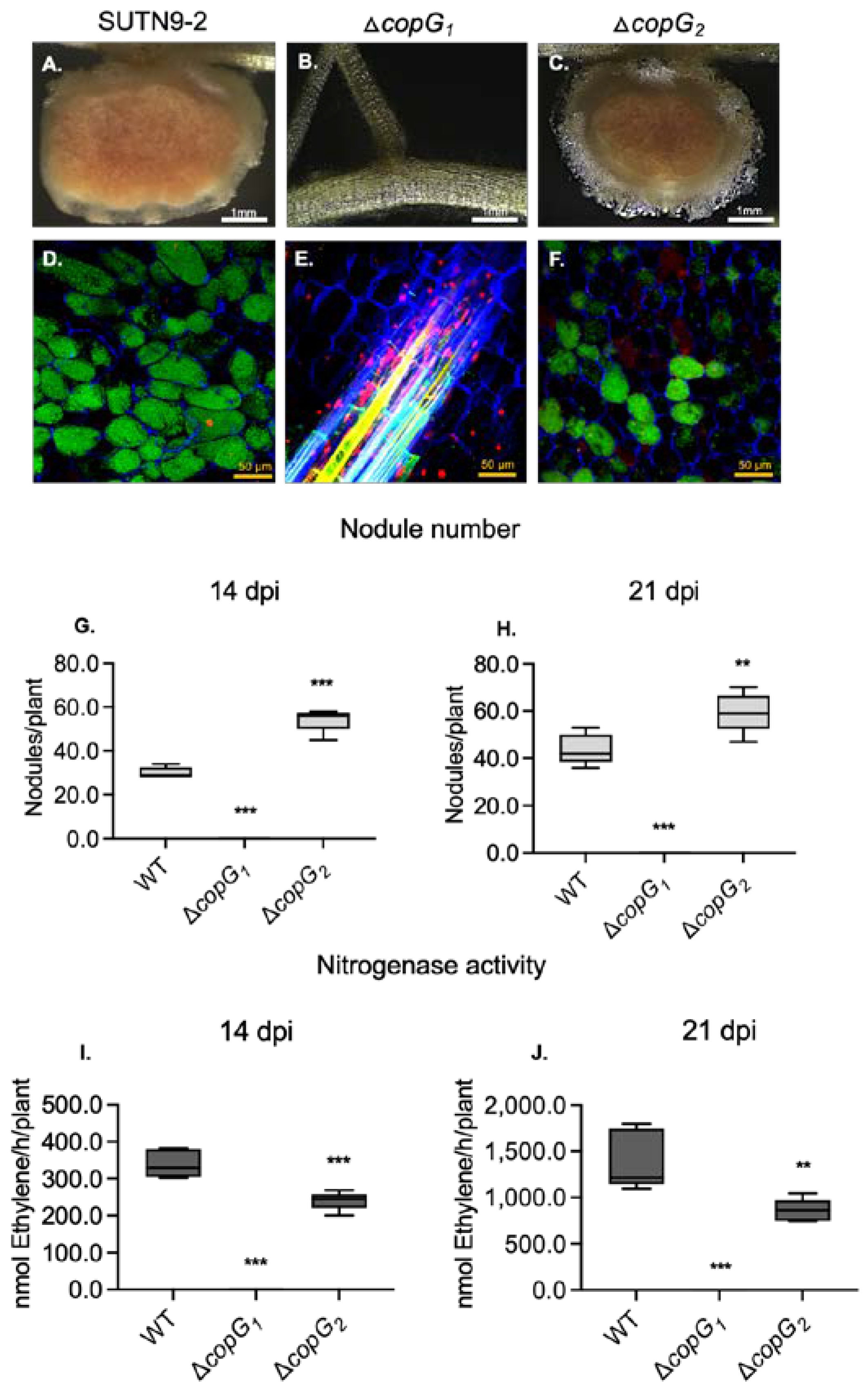
Figure 3.
qRT-PCR analysis of nod genes from wild type Bradyrhizobium sp. SUTN9-2 (WT) and ΔcopG1 grown in absence and presence of 20 μM genistein (G). Expression of structural genes nodA (A), nodB (B), and nodC (C) and regulatory genes nodD1 (D), nodD2 (E) and transcriptional regulator of T3SS, ttsI (F). Data were normalized in relation to the endogenous control (16S rRNA). Values represent mean ± SD (n = 3). P values based on Tukey’s test (*** P < 0.001).
Figure 3.
qRT-PCR analysis of nod genes from wild type Bradyrhizobium sp. SUTN9-2 (WT) and ΔcopG1 grown in absence and presence of 20 μM genistein (G). Expression of structural genes nodA (A), nodB (B), and nodC (C) and regulatory genes nodD1 (D), nodD2 (E) and transcriptional regulator of T3SS, ttsI (F). Data were normalized in relation to the endogenous control (16S rRNA). Values represent mean ± SD (n = 3). P values based on Tukey’s test (*** P < 0.001).
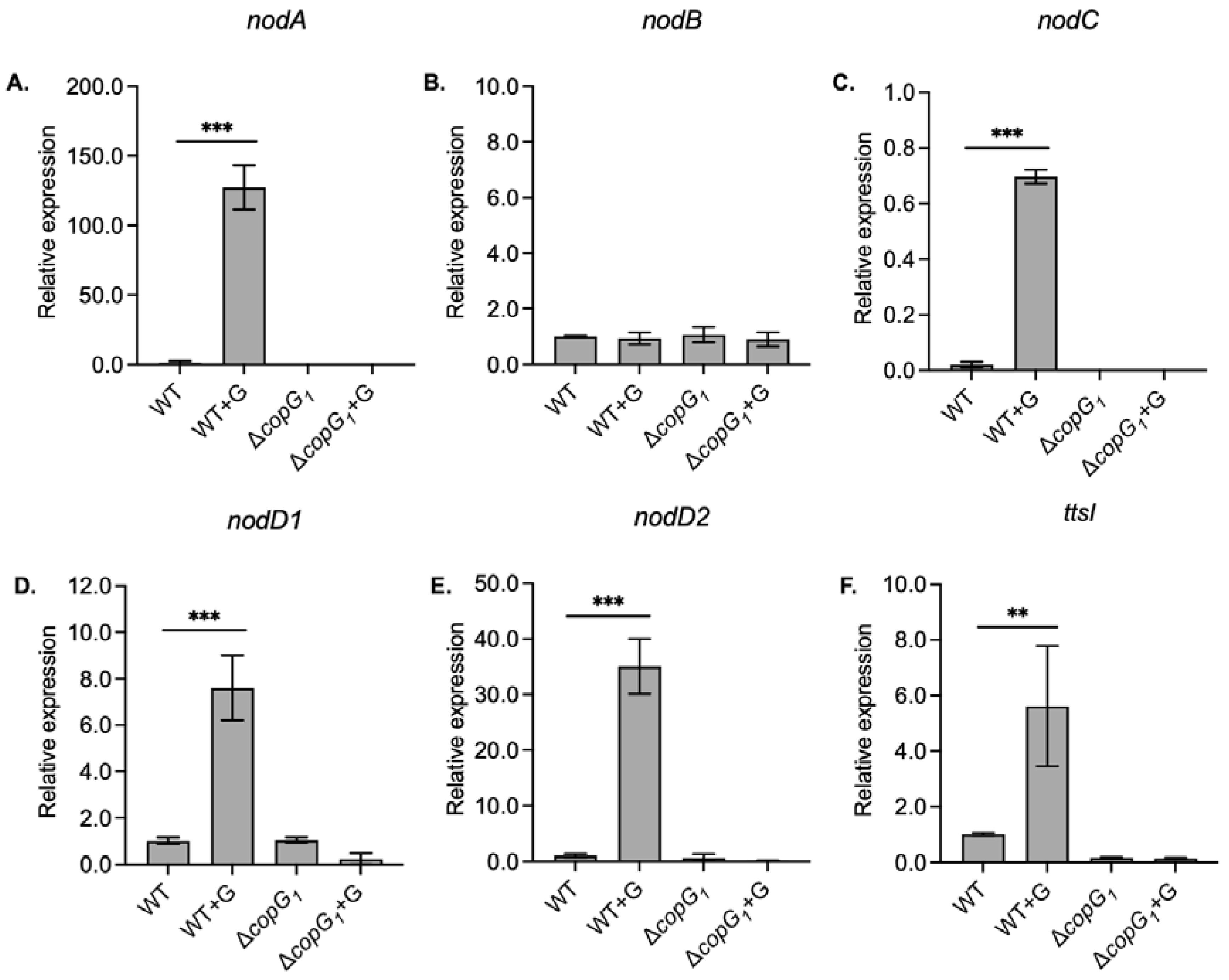
Figure 4.
Relative expression of representatives T4SS structural genes, including trbE1 (A, C), and traG1 genes (B, D) in Bradyrhizobium sp. SUTN9-2 (WT) and ΔcopG1 with and without 20 μM genistein (G) induction. The 16S rRNA gene was used as an internal control. Values represent mean ± SD (n = 3). P values based on Student’s t-test (ns P > 0.05, * P < 0.05, ** P < 0.01, *** P < 0.001). The green and red arrows represent a statistical increase and decrease, respectively, in gene expressions when comparing experiments with the presence and absence of genistein.
Figure 4.
Relative expression of representatives T4SS structural genes, including trbE1 (A, C), and traG1 genes (B, D) in Bradyrhizobium sp. SUTN9-2 (WT) and ΔcopG1 with and without 20 μM genistein (G) induction. The 16S rRNA gene was used as an internal control. Values represent mean ± SD (n = 3). P values based on Student’s t-test (ns P > 0.05, * P < 0.05, ** P < 0.01, *** P < 0.001). The green and red arrows represent a statistical increase and decrease, respectively, in gene expressions when comparing experiments with the presence and absence of genistein.
Figure 5.
SDA-PAGE analysis of proteins secretion into the external medium of Bradyrhizobium sp. SUTN9-2 (WT), ΔT4SS1, and ΔcopG1 with 20 μM genistein (+Gen) and without 20 μM genistein (-Gen) induction. Numbers on the left indicate molecular size markers in kilodaltons. The arrowhead indicates bands identified by mass spectrometry (MS) analysis of WT (1, 3, 6), ΔT4SS1 (2, 4, 7), and ΔcopG1 (5).
Figure 5.
SDA-PAGE analysis of proteins secretion into the external medium of Bradyrhizobium sp. SUTN9-2 (WT), ΔT4SS1, and ΔcopG1 with 20 μM genistein (+Gen) and without 20 μM genistein (-Gen) induction. Numbers on the left indicate molecular size markers in kilodaltons. The arrowhead indicates bands identified by mass spectrometry (MS) analysis of WT (1, 3, 6), ΔT4SS1 (2, 4, 7), and ΔcopG1 (5).
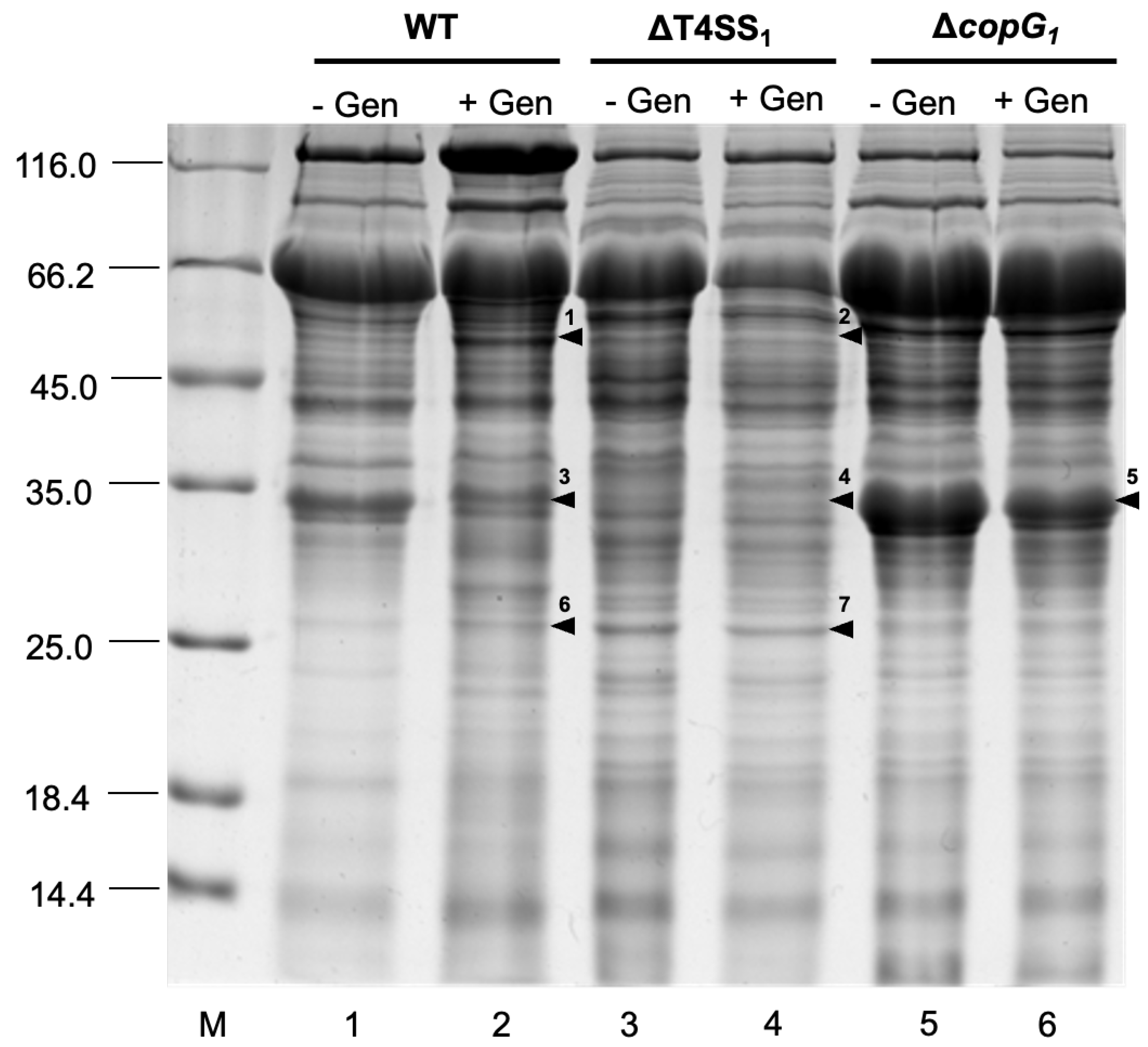
Figure 6.
Relative expression of Nodulation outer protein X (nopX) (A), C4-dicarboxylate transporter (dct) (B), and nopP genes (C) in Bradyhizobium sp. SUTN9-2 (WT) and ΔcopG1 under 20 μM genistein induction (+G). Data were normalized in relation to endogenous control (16S rRNA). Values represent mean ± SD (n = 3). P values based on Student’s t-test (* P <0.05, ** P < 0.01,*** P < 0.001).
Figure 6.
Relative expression of Nodulation outer protein X (nopX) (A), C4-dicarboxylate transporter (dct) (B), and nopP genes (C) in Bradyhizobium sp. SUTN9-2 (WT) and ΔcopG1 under 20 μM genistein induction (+G). Data were normalized in relation to endogenous control (16S rRNA). Values represent mean ± SD (n = 3). P values based on Student’s t-test (* P <0.05, ** P < 0.01,*** P < 0.001).
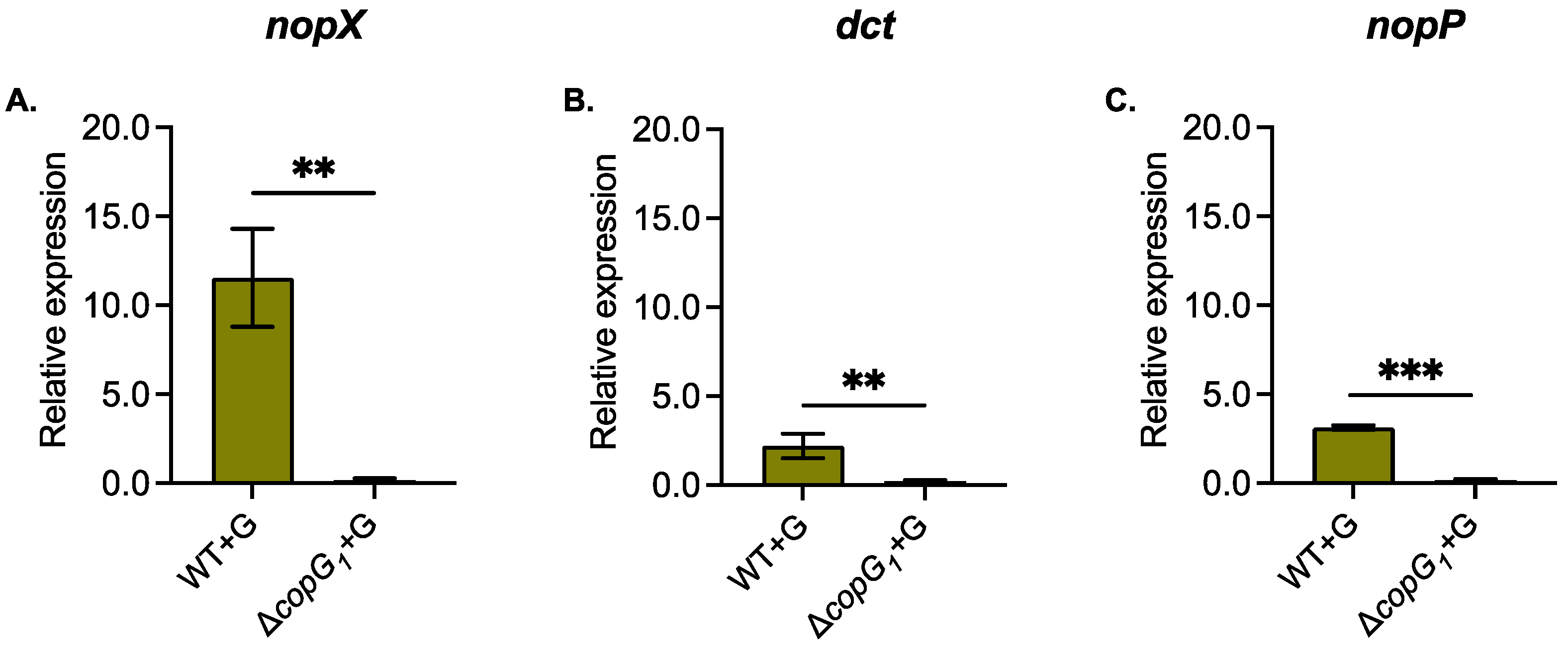

Table 1.
Bacterial strains and plasmids used in this study.
| Strain or Plasmid | Relevant Characteristics | Reference or |
|---|---|---|
| Source | ||
| Strain | ||
| Bradyrhizobium sp. | ||
| SUTN9-2 | A. americana nodule isolate (paddy crop) | |
| ∆copG1 | SUTN9-2 derivative containing an Ω cassette insertion at HindIII site, copG copy 1::sm/sp; Smr, Spr | This study |
| ∆copG2 | SUTN9-2 derivative containing an Ω cassette insertion at BamHI site, copG copy 2::sm/sp; Smr, Spr | This study |
| ∆traG1 | SUTN9-2 derivative containing an Ω cassette insertion at BamHI site, traG copy 1::sm/sp; Smr, Spr | This study |
| ∆virD21 | SUTN9-2 derivative containing an Ω cassette insertion at BamHI site, virD2 copy 1::sm/sp; Smr, Spr | This study |
| Escherichia coli | ||
| DH5α | supE44 ΔlacU169 hsdR17 recA1 endA1 gyrA96 thi-1 relA1 | Toyobo Inc. |
| Plasmid | ||
| pRK2013 | ColE1 replicon carrying RK2 transfer genes; Kmr; Helper plasmid | [22] |
| pNTPS129 | Cloning vector harboring sacB gene under the control of the constitutive npt2 promoter; Kmr | [23] |
| pNTPS129-∆copG1 | pNTPS129-npt2-sacB containing the flanking region of copG copy 1 | This study |
| pNTPS129-∆copG2 | pNTPS129-npt2-sacB containing the flanking region of copG copy 2 | This study |
| pNTPS129-∆traG1 | pNTPS129-npt2-sacB containing the flanking region of traG copy 1 | This study |
| pNTPS129-∆virD21 | pNTPS129-npt2-sacB containing the flanking region of virD2 copy 1 | This study |
Table 2.
Primers used in this study.
| Name | Sequences (5’-3’) | Descriptions |
|---|---|---|
| Primers for gene deletion | ||
| Up.copG1. XbaI.F | CCT TGA GAT CTA GAT GTA GTC TGC CCC GAA GTA GC | This primer sets used to obtain the deletion of copG1 gene of Bradyrhizobium sp. SUTN9-2 by double crossing over. |
| Up. copG1. overl. HindIII. R | GAG GCG GAC ATG AAA GCT TAA TGA AGG CGG ACG GCC ACT AG | |
| Dw. copG1. overl. HindIII. F | GTC CGC CTT CAT TAA GCT TTC ATG TCC GCC TCA CAG TCC GA | |
| Dw. copG1. EcoRI.R | AGA TCG GGA ATT CGT TGA CCG AGG ATC TTC AGG CCA | |
| Up. copG2. XbaI.F | GCC GTT TCT AGA ATT GCG ACA ACG GAC CAG GGC AA | This primer sets used to obtain the deletion of copG2 gene of Bradyrhizobium sp. SUTN9-2 by double crossing over. |
| Up. copG2. overl. HindIII. R | GCG CGA CCG AAT GAA GCT TAA GCT GGT CAC GCT ATC GGC T | |
| Dw. copG2. overl. HindIII. F | GCG TGA CCA GCT TAA GCT TCA TTC GGT CGC GCA TAT TGC C | |
| Dw. copG2. EcoRI. R | CTG TCC GAA TTC ATG TCG TTC CTC GGG TTG TAC C | |
| Up. traG1. XbaI. F | TTC GGG TCT AGA TGT AGT CTG CCC CGA AGT AGC | This primer sets used to obtain the deletion of traG1 gene of Bradyrhizobium sp. SUTN9-2 by double crossing over. |
| Up. traG1. overl. BamHI | TCC CTC CAA TCA CGG ATC CAT CCT GGT GAC GAT CTC GGA C | |
| Dw. traG1. overl. BamHI | TCG TCA CCA GGA TGG ATC CGT GAT TGG AGG GAT CGT TCA CAG | |
| Dw. traG1.EcoRI.R | CCG GCT GAA TTC CTT GGA AAG CCT TGG TCT CG | |
| Up. virD21. XbaI. F | ACC GGC TTC TAG AAG ATG CGC AGT CCG CAT CAT C | This primer sets used to obtain the deletion of virD21 gene of Bradyrhizobium sp. SUTN9-2 by double crossing over. |
| Up. virD21. overl. BamHI | GAG GAG AAG GAA TGG ATC CTG AAC GAT CCC TCC AAT CAC CG | |
| Dw. virD21. overl. BamHI | GAG GGA TCG TTC AGG ATC CAT TCC TTC TCC TCA GCC ATG GC | |
| Dw. virD21. EcoRI. R | CCA TCG GAA TTC TTG TCG ATG CGG AGG AGG CAT C | |
|
Primers for qRT-PCR analysis |
||
| SUTN9-2. nodA. F | GTT CAA TGC GCA GCC CTT TGA G | Specific primers for nodA gene expression in SUTN9-2 on chromosome |
| SUTN9-2. nodA. R | ATT CCG AGT CCT TCG AGA TCC G | |
| SUTN9-2. nodC. F | ATT GGC TCG CGT GCA ACG AAG A | Specific primers for nodC gene expression in SUTN9-2 on chromosome |
| SUTN9-2. nodC. R | AAT CAC TCG GCT TCC CAC GGA A | |
| SUTN9-2. nodD1. F | ATT CGT CTC CTC AGA CCG TGC T | Specific primers for nodD1 gene expression in SUTN9-2 on chromosome |
| SUTN9-2. nodD1. R | TTC ATG TCG AGT GCG CAC CCT A | |
| SUTN9-2. nodD2. F | TGC TTA ACT GCA ACG TGA CCC | Specific primers for nodD2 gene expression in SUTN9-2 on chromosome |
| SUTN9-2. nodD2. R | ATG AGC ACG AGG AGC TTC TC | |
| SUTN9-2. trbE1. F | GAT TGC AGG AGA ACC GTG AGG C | Specific primers for trbE1 gene expression in SUTN9-2 on chromosome |
| SUTN9-2. trbE1. R | AAC AGC GCC GAG GAT TCA GTC T | |
| SUTN9-2. traG1. F | TTC TCG ATC TGG TTC AGC GAC TG | Specific primers for traG1 gene expression in SUTN9-2 on chromosome |
| SUTN9-2. traG1. R | TTG ACC GAG GAT CTT CAG GCC A | |
| SUTN9-2. ttsI. F | ATG AGT TCG TCG GTG GAC AC | Specific primers for transcriptional regulator TtsI (ttsI) gene expression in SUTN9-2 on chromosome |
| SUTN9-2. ttsI. R | CCA CAT GGT CCT GCT CGA AT | |
| 16s. F | ATT ACC GCG GCT GCT GG | Universal primers for 16S rRNA used as internal control for bacterial gene expression [25] |
| 16s. R | ACT CCT ACG CGA GGC AGC AG | |
| dct. F | CGA CTA TCA GGG CGT GAA AT | Specific primers for C4-dicarboxylate transport (dct) gene expression in SUTN9-2 on chromosome |
| dct. R | TCC AGC AAT CAG ACC TGT G | |
| nopX. F | GGGTGGTCGAGGAAGTATTG |
Specific primers for Type III secretion system (T3SS) gene expression in SUTN9-2 on chromosome |
| nopX. R | GGTTATGACCCAGACCGATG | |
| nopP. F | GGTCACACCGACGAAGATAC | |
| nopP. R | CCGAAGATCCACTTGGGATG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



