
Submitted:
05 May 2024
Posted:
07 May 2024
You are already at the latest version
Abstract
Zinc (Zn) is the second most abundant metal in the human body, and is essential for the function of 10% of all proteins. As metals cannot be synthesized or degraded, they must be assimilated from diet by specialized transport proteins, which unfortunately also provide an entry route for the toxic metal pollutant cadmium (Cd). The intestinal absorption of Zn depends on the composition of food that is consumed, firstly the amount of Zn itself, and then the quantity of other food constituents such as phytate, protein and calcium (Ca). In cells, Zn is involved in regulation of intermediary metabolism, gene expression, cell growth, differentiation, apoptosis, and antioxidant defense mechanisms. Cellular influx, efflux, subcellular compartmentalization, and trafficking of Zn are coordinated by transporter proteins, the Solute-Linked Carrier 30A and 39A (SLC30A and SLC39A), known as the ZnT and the Zrt-/Irt-like protein (ZIP). Because of its chemical similarity with Zn and Ca, Cd disrupts the physiological functions of both. The concurrent induction of a Zn efflux transporter ZnT1 (SLC30A1) and metallothionein by Cd disrupts the homeostasis and reduces the bioavailability of Zn. The present review highlights the increased mortality and the severity of various diseases, including those induced by COVID-19 infections, among Cd-exposed persons and the roles of Zn and other transport proteins in the manifestation of Cd cytotoxicity. Special emphasis is given to the potential protective effects of Zn against bone, lung and heart diseases associated with Cd exposure. The difficult challenge of determining a permissible intake level of Cd is discussed in relation to the recommended dietary Zn intake levels.
Keywords:
antagonism
; bioavailability
; cadmium
; metal transporters
; recommended dietary allowance
; RDA
; zinc
1. Introduction
Zinc (Zn) is a metal nutrient, which is required for the function of 10% of proteins in the human body, and represents the second most abundant metal in the body, after iron (Fe) [1,2,3,4]. A total of 2-3 g of Zn is present in adults: 50-60% in muscles, 30-36.7% in bones, and 4.2%, 3.4% and < 1% in skin, liver, and the circulation, respectively [2,3,4]. The total plasma Zn concentration ranges between 9.9 and 24.5 µM (65 -160 µg/dL); most of which is bound to albumin (80%) and the remainder to primarily α-2-macroglobulin, a protease inhibitor. The free plasma Zn concentrations are much lower and range between 1 and 3 nM [5,6,7].
Unlike Fe, the body does not have a storage mechanism for Zn, and thus sufficient intake is required to maintain normal cell metabolism and function, and prevent deficiency [4,5]. The recommended dietary allowance (RDA) for Zn is 8 and 11 mg/day for adult women and men, respectively [8]. However, it is noteworthy that the total plasma Zn concentrations do not reliably reflect the Zn body status, as it represents only a miniscule fraction of total body Zn, there are significant diurnal variations, and there are large fluctuations of Zn levels with many disease states that include inflammation and infection [9-11]. Furthermore, Zn contributes to the specialized function of many tissues and organs, including bone turnover [12], hormones thyroxine and insulin [13,14], central nervous system development and function [15], immunity [16] and spermatogenesis [17].
Due to its wide range of biological roles, no specific symptom can be attributable to Zn deficiency, and it cannot be readily measured [2,3]. Thus, the health impact of marginal Zn intake and a subclinical Zn deficiency are not easily recognized. Inadequate dietary Zn intake is believed to be highly prevalent, especially in populations consuming diets low in Zn bioavailability, such as those that contain high amounts of phytate, or are low in protein and calcium (Ca) [5,19-20].
Cadmium (Cd) is a metal contaminant with continuing public health concern worldwide because of its high toxicity. It has no nutritional value or physiological role, and its presence disrupts the homeostasis and function of essential metal nutrients, including Zn, copper (Cu), Fe, Ca, and manganese (Mn) [21-23]. Dietary exposure to Cd is inevitable for most people as it is found in nearly all food types [24-26], polluted air, and tobacco smoke. [27-31]. Children are particularly sensitive to the toxicity of airborne Cd, possibly due to the enhanced pulmonary uptake of Cd in this age group, especially in those with a small airway disease such as asthma [32-34]. Higher risks of having asthma were associated with lower plasma/serum Zn levels in a meta-analysis of 2205 children [35].
As there are no physiologic mechanisms for Cd elimination, continued exposure will lead to it accumulation throughout the body. Thus, the primary public health measure should focus on minimizing exposure. A systematic review and meta-analysis showed that the body burden of Cd was inversely associated with body Fe stores and nutritional status of Zn [36]. The Cd-Zn connection is also supported by the finding that elevated blood and urinary Cd levels are associated with particular genetic variants of the Zn transport proteins, ZIP8 and ZIP14 [37]. However, the current permissible exposure guideline, referred to as a tolerable intake level of Cd, is 0.83 µg/kg body weight per day, (58 µg/day for a 70-kg person), which is outdated and not protective of human health [38]. The benchmark dose limit (BMDL) value, equivalent to the no-observed-adverse effect level (NOAEL) of Cd exposure is extremely low [39]. Of further concern, effective chelation therapy for removal of Cd from cells does not exist.
The primary aim of this review is to provide epidemiological and experimental evidence implicating Zn and its transport proteins in determining the severity of adverse health outcomes due to Cd exposure. Firstly, it summarizes the RDA for Zn and existing exposure guidelines for Cd and its toxicity threshold level. In addition, it discusses current understanding of the specific metal transporters, which are now known to impact the intestinal absorption, cellular acquisition, and manifestations of the cytotoxicity of Cd. Secondly, it summarizes the overall health threat linked with environmental non-workplace Cd exposure as reflected by mortality data recorded in prospective cohort studies. Thirdly, Cd-induced diseases of the bone, lung, and heart are discussed, where the involvement of Zn and its transporters are highlighted.
2. Biomolecule Zinc versus a Contaminant Cadmium
In this section, the RDA for Zn and the metal transporters responsible for its absorption are discussed together with various factors such as diet components that affect Zn absorption and its bioavailability. In comparison, a “tolerable” intake level of Cd and its absorption by enterocytes and uptake by target cells are highlighted. By definition, a tolerable intake level for a chemical with no known biological function is an estimate of the amount that can be ingested over a lifetime without an appreciable health risk [38].
2.1. Recommended Dietary Allowance for Zinc versus Tolerable Intake Level for Cadmium
The RDA values for Zn in various age groups, and current dietary exposure guideline as tolerable intake levels of Cd are provided in Table 1.
2.1.1. Zinc and Its RDA values
According to the U.S. National Academy of Sciences Institute of Medicine (IOM), respective RDA values of Zn for an average adult 60-kg female, and 70-kg male are 8 and 11 mg/day [8]. Additional 3 to 5 mg of Zn is required to meet increased physiological demands during pregnancy and lactating periods. The RDA for Zn may need to be increased further to help reduce the transfer of Cd to breast milk and its inhibitory effect on the secretion of Ca to breast milk, which has been observed in Cd-exposed Bangladeshi women [45,46].
2.1.2. Tolerable Intake Levels of Cadmium: A Threshold-Based Approach
According to the Food and Agriculture Organization and World Health Organization (FAO/WHO) Joint Expert Committee on Food Additives and Contaminants (JECFA), a tolerable intake level of Cd was set originally at 400–500 µg/person/week, which was later revised to 7 µg/kg body weight/week. Later, the weekly intake level was amended to a tolerable monthly intake of Cd at 25 μg per kg body weight per month, equivalent to 0.83 μg per kg body weight per day (58 µg/day for a 70 kg person), and the Cd excretion of 5.24 μg/g creatinine was used as a nephrotoxicity threshold level [38]. These figures were based solely on the excretion rate of β2-microglobumin ≥ 300 μg/g creatinine as a toxic endpoint.
Like JECFA, the tolerable ingestion rates for Cd listed in Table 1 assumed an existence of a toxicity threshold level, while using urinary Cd excretion rate, normalized to creatinine excretion to reflect long-term exposure or body burden of Cd [40-43]. As reviewed in Satarug, 2024, however, this method of normalization of urinary Cd excretion created a high degree of statistical uncertainty, which obscured and nullified the quantification of Cd effects [47]. Consequently, none of the existing guidelines could reliably indicate a dietary intake level that carries a negligible health risk, inferred from the definition for a tolerable intake level of any contaminant.
The utility of urinary β2M excretion levels ≥ 300 µg/g creatinine, termed tubular proteinuria, as a toxic endpoint, is another serious conceptual flaw. This endpoint is associated with severe kidney pathologies, resulted from tubular cell injury and death and malfunction and nephron destruction by Cd accumulation (Figure 1).
In phase 1, a submicroscopic tubular cell perturbation induced by Cd is evident from the appearance of KIM1 in the urine [22]. In phase 2, GFR begins to decline as tubular cell damage and death are intensified with continuing Cd influx [48]. In phase 3, albuminuria and tubular proteinuria ensue because of defective reabsorption and a substantial loss of functioning nephrons.
Phase 1 appears to be reversible through cellular repair mechanisms if there is no further influx of Cd. Cessation of exposure may delay progression to phase 3, which is irreversible, and can advance to end-stage kidney disease, when dialysis is necessary for survival.
2.1.3. Loss of GFR as a Sensitive Toxic Endpoint
In theory, an exposure level that is more likely to produce discernable health risk needs to be based on the most sensitive endpoint [49]. Kidney tubular epithelial cell damage and death, depicted in Figure 1, have been most frequently reported as signs of the nephrotoxicity of Cd in non-occupationally exposed conditions. However, a declining eGFR, is a common sequela of ischemic acute tubular necrosis, acute and chronic tubulointerstitial fibrosis, all of which create impediments to filtration such as the destruction of post-glomerular peritubular capillaries, amputation of glomeruli from tubules, and obstruction of nephrons with cellular debris [50,51].
When GFR loss was used as a toxic endpoint, the NOAEL equivalent value or a toxicity threshold level of urinary Cd excretion ranged between 0.01 and 0.02 µg/g creatinine [39,52], which is at least 250 times less than 5.24 µg/g creatinine used by the JECFA to derive a tolerable intake level for Cd [38]. There is no safe level of Cd exposure.
In the China Health and Nutrition Survey (n = 8429), in which 641 (7.6%) had chronic kidney disease (CKD), the likelihood of having CKD increased 1.73-, 2.93- and 4.05-fold when dietary Cd exposure rose from 16.7 to 23.2, 29.6 and 36.9 μg/day, respectively [53]. A dietary Cd exposure level of 23.2 µg/day is 40% of the JECFA tolerable exposure guideline, and yet it was associated with 1.73-fold increase in risk of eGFR to fall below 60 mL/min/1.73m2. This is not a not negligible health risk.
2.1.4. Summary of a “Tolerable” Intake Level of Cadmium
Current dietary Cd exposure guidelines range between 0.21 to 0.83 µg/kg body weight per day (Table 1). These guidelines assumed Cd excretion rates of 0.5−5.24 µg/g creatinine as the threshold level for toxicity to kidneys or bones. However, the NOAEL equivalent value of urinary Cd excretion of 0.01 and 0.02 µg/g creatinine was obtained when a declining eGFR was employed as a toxic endpoint. It is now apparent that all existing tolerable dietary Cd intake levels are not low enough to protect human health due to exposure to Cd. Also, it should be noted that most or all excreted Cd originates from injured or dying kidney tubular epithelial cells. Thus, the excretion of Cd itself quantifies the severity of the kidney injury due to Cd accumulation at the present time, not the risk of injury in the future. We therefore argue whether a “tolerable” intake of a cumulative toxicant like Cd is a scientifically valid concept. At very least, exposure to environmental Cd has been found to enhance the mechanisms underlying the cellular senescence, involving surtuin-1 (SIRT1) [54-56].
2.2. Absorption of Metal Nutrients versus Contaminant Cadmium: An Overview
Living organisms cannot synthesize nor destroy any metals, and transport proteins have consequently been evolved to obtain all their required metals from an external environment. Generally, for humans, the intestinal absorption of metals from diet involves two main processes. Firstly, transport of metals into enterocytes, mediated by influx transporters, localized to the brush border membrane. Secondly, transport of metals to the basolateral membrane, where metals are extruded from enterocytes to portal blood, mediated by efflux transporters.
Table 2 enlists the influx and efflux transporters that the body uses to acquire metal nutrients, Zn, Fe, Mn, Cu and Ca.
2.2.1. Role of ZIP4, ZnT5, and ZnT1 in Dietary Zn Absorption
ZIP4, ZnT5 and ZnT1 are the transport proteins responsible for the intestinal absorption of Zn. Due to unique Zn-histidine pairing, not present in the bacterial ortholog ZnT1, Zn is absorbed through highly selective transporters ZIP4 and ZnT1 [57,58,79]. In adults, an approximate of 2.5-3.5 mg of Zn is absorbed per day to meet physiological requirements although several mg of Zn are secreted in pancreatic juices [5,9,10,18]. An amount of absorbable dietary Zn depends on phytate (myo-inositol hexakisphosphate), Ca and protein intake. Dietary phytate chelates Zn, and Ca fortification of phytate-rich soy milk has been shown to increase Zn available for absorption. Protein intake may increase Zn absorption through the Zn-amino acid co-transport mechanisms [5]. The intestinal absorption may be enhanced also by Zn ionophores [80].
Zn supplementation has many challenges, given the co-existence of Fe and Zn deficiencies and their interactions if supplemented together [4,5,20,70,80]. A high-dose Zn supplement necessitates an additional 2 mg of Cu to prevent anemia due to Cu deficiency induced by a high-dose Zn (≥ 80 mg/day) [5,81]. Decreased ZnT5 and ZIP4 protein levels were observed in ileal biopsies collected from volunteers, who consumed 25 mg of Zn as zinc sulfate with food daily for two weeks [60]. Increased systemic blood pressure and reduced kidney function, measured by inulin clearance, were observed in rats fed with diet containing 40 times higher Zn than in normal diet for 4 weeks [82].
2.2.2. High Absorption Rate of Cadmium: Role of Multiple Transporters
As listed in Table 2, dietary Cd gains an access to the systemic circulation and reaches cells throughout the body through several metal transporters, including those for Fe, Zn, Cu and Ca. Furthermore, Cd complexed with MT and phytochelatin can be absorbed through transcytosis and receptor-mediated endocytosis [83-85]. Studies from Japan reported the absorption rates of Cd among women to be as high as 24–45% [86,87].
It is noteworthy that Cd absorption rates of 3–7% were used in the estimation of a tolerable intake level of Cd by JECFA (Table 1). Such assumption of low intestinal Cd absorption rates has led to miscalculation of a tolerable exposure level of dietary Cd.
Cd can reach most cells in the body in the same way as metal nutrients, Zn, Fe, Ca, and Cu. Indeed, the cellular acquisition of Cd appeared to be through many more transporters than those mediate Cd entry into the enterocytes [88,89]. Of note, however, no excretory mechanism has been found for Cd. ZnT1 mediates the efflux of Zn, but not Cd [61,62]. Similarly, FPN1 mediates the efflux of Fe, Zn and Co, but not of Cu, Cd or Mn [69,70]. Consequently, most Cd acquired are retained within the cell after its entrance. In the absence of any efflux transporters for Cd removal exploitation, avoidance/minimization of exposure is an essential measure to prevent many health threats, elaborated in Section 3 and Section 4.
3. Antioxidative Function of Zinc and the Cytotoxicity of Cadmium
Zn is a type 2 nutrient that plays role in fundamental biological mechanisms, namely protein and DNA syntheses, gene transcription and cell proliferation, division, and differentiation [3,70,90]. The maintenance of normal intermediary metabolism and cell function by Zn is accomplished through its role in cellular redox signaling and antioxidant defense mechanisms [91-93].
In this section, we highlight fundamental role of Zn in the retention of cell redox state, which is pivotal to normal cell metabolism and cell function. Also, we provide insights into the molecular basis for the cytotoxicity of Cd, focusing on its disruption of cellular Zn homeostasis and the antioxidative function of Zn.
3.1. Zinc Homeostasis and Antioxidative Function of Zinc
In cells, half of Zn is present in the cytoplasm, where it is bound to many proteins, and sequestered into subcellular organelles such as mitochondria, the endoplasmic reticulum (ER), vesicles, and the Golgi [94-96]. The cytosolic labile Zn concentration is in a picomolar to low nanomolar range, which is very low compared to the total intracellular Zn concentrations of 10 -100 μM [94].
Cellular acquisition of Zn, its levels in cells and trafficking between subcellular organelles are regulated tightly by specialized transport proteins, the SLC30A (ZnT) and SLC39A (ZIP) [94-96]. These transport proteins are fundamental to Zn homeostasis. Zn itself contributes also to its homeostasis through induction of ZnT1 [97] and MT to which 5-15% of cytosolic Zn is bound [98]. ZnT1 is a unique efflux transporter that functions as Zn/Ca exchanger, which protects against a rise of cellular Zn [99,100].
In normal intermediary metabolism, including mitochondrial ATP synthesis, reactive oxygen species (ROS) is produced and antioxidative mechanisms have been evolved to protect against oxidative damage, due to excessive ROS [91-93,101].
An overview of the molecular entities responsible for Zn homeostasis and its antioxidative function are presented in Figure 2.
As Figure 2 indicates, the entry of Zn, its distribution to subcellular localization and extrusion from cells are mediated by influx transporters, working in concert with ZnT efflux transporters. Modulation of superoxide dismutase (SOD) and NADPH oxidase (Nox) activities are well-known Zn roles [102,103], while the antioxidative function of Zn through bilirubin has been demonstrated recently, detailed below.
Because δ-aminolevulinic acid dehydratase (ALD), an enzyme in heme biosynthesis is Zn-dependent, anemia in Zn deficiency conditions was due to insufficient amount of heme for hemoglobin synthesis [104]. A methodological breakthrough in measuring bilirubin in cells was made, following a discovery of a protein capable of binding unconjugated bilirubin [105]. Subsequently, it was demonstrated that heme was synthesized de novo and used continuously to produce bilirubin as a cytoprotective biomolecule [106], given it is a potent antioxidant and a lipid peroxidation chain breaker [107]. Production of bilirubin may be diminished due to in reduced Zn availability, in the case of Cd, or displacement of Zn in ALD by lead (Pb) [108].
3.2. Cadmium-Induced Disruption of Cellular Zinc Homeostasis and Redox State
More than a decade ago, Moulis (2010) published the hypothesis that disruption of the homeostasis of physiologically required metals is a central toxic mechanism of Cd [109]. Because the ionic radius and electronegativity of Cd are close to Zn and Ca, the homeostasis of Zn and Ca are the primary Cd toxicity targets [110,111].
An Australian autopsy study has provided evidence for the impacts of Cd exposure on Zn and Cu levels in livers and kidneys [112]. Similar effects of Cd on hepatic and renal Zn and Cu levels have been observed in rats 6 months after exposure [113].
By the same way as Zn, Cd induces expression of MT and ZnT1 simultaneously through its interaction with the metal-response element binding transcription factor-1(MTF-1) [98]. Satarug et al. (2021) used UROtsa cells, a cell culture model of human urothelium [114], to explore the effects of an acute exposure to Cd on the ZnT and ZIP expression profiles (Table 3).
At basal state, ZnT7 was most abundantly expressed among the ZnT family members, followed by ZnT5 and ZnT1. For ZIP family, ZIP7 was most abundantly expressed, followed by ZIP14, ZIP6, ZIP1, ZIP10, and ZIP3A.
The expression levels of ZnT3, ZnT4, ZIP1, ZIP2, ZIP3A, ZIP5, ZIP7 and ZIP10, all fell 24 h after Cd exposure, but the expression of ZnT1 rose markedly.
In the same study, expression profiles of ZnT and ZIP in UROtsa cells treated with buthionine sulfoximine, an inhibitor of glutathione (GSH) synthesis were compared with those cells treated with 5-aza-2'-deoxycytidine, a DNA methylation inhibitor. Results indicated that Cd-induced ZnT1 expression was influenced mostly by GSH levels. Similarly, the effects of Cd on ZnT5, ZnT7, and ZIP8 expression levels appeared to vary with cellular redox status, but not with DNA methylation status. However, the effects of Cd on ZIP14 gene expression seemed to depend on both GSH levels and the DNA methylation state.
Experimental data described above suggested that that redox state and epigenic mechanisms were key regulators of ZIP and ZnT expression and that the relationships of influx (ZIP) and efflux (ZnT) were highly sophisticated, noted also in a recent study [ 115].
4. Global Health Threat of Environmental Cadmium
In this section, we summarize results from cross-sectional and longitudinal cohort studies, where overall health impact of Cd has been observed at the exposure levels below existing health-based guidelines presently exist. Experimental studies that revealed the interactions of Cd, Zn, and various Zn transport proteins, which may mitigate Cd-induced damage to bones are highlighted. Arguably, an increased dietary intake of bioavailable Zn ions will have a significant health benefit.
4.1. Cadmium and the World’s Leading Causes of Death
In the WHO global health report, cardiovascular disease (CVD), particularly ischemic heart disease was the world’s top cause of death, which accounted for 16% of total global deaths in 2019 (https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death). The 2nd, 3rd and 9th leading causes of death in 2019 were stroke, chronic obstructive pulmonary disease (COPD), and diabetes, respectively. Death from kidney disease rose from the 13th in 2000 to the 10th in 2019.
Verzelloni et al. (2024) conducted a dose-response meta-analysis of data from 26 studies, published during 2005 to 2023, to evaluate the contribution of Cd exposure to the risk of CVD [116]. They found that the risks of heart failure, coronary heart disease, and stroke were all increased with Cd exposure in a dose-dependent manner. Specifically, blood Cd level of 1 μg/L and urinary Cd excretion rate of 0.5 μg/g creatinine appeared to be sufficient to increase the risk of having CVD 2.58-fold and 2.79-fold, respectively.
Epidemiological studies implicating Cd exposure as a significant contributor to total global mortality and morbidity can be found in Table 4.
In the U.S. hypertension cohort [117], blood Cd levels ≥ 0.80 μg/L were associated 1.85-,1.76- and 3.41-fold increases in the mortality from any causes, CVD and Alzheimer’s disease, respectively. Cd appeared to increase CVD mortality markedly (OR 2.12) among non-smokers who had hypertension [117]. In other two U.S. diabetes and CKD cohorts, urinary Cd levels > 0.60 μg/L were associated with a 49% increase in all-cause mortality among the diabetics [118], while urinary Cd levels ≥ 0.60 μg/g creatinine were associated with a 75% increase in deaths from any cause among those with CKD [119].
In a cohort of Swedish women [120], Cd exposure was associated with 38% and 20% increases in risk of death from any cause and having bone fracture, respectively. These results were obtained, when top tertile of urinary Cd (median 0.54 µg/g creatinine) was compared with the bottom tertile (median urinary Cd of 0.20 µg/g creatinine).
In a Taiwanese cohort of the general population [121], there was a 35% increase in all-cause mortality per 1 μg/L increment of urinary Cd. Also, in this study, death from any cause rose 35% for every 1 μg/dL increment of urinary Cu. The association between mortality and urinary Cu in Taiwanese cohort study may at least reflect kidney Cd toxicity, given that Cu is a redox active metal and an ROS generator [101]. Furthermore, increased urinary excretion of Cu among Cd-exposed subjects has been noted in studies from Japan [124,125], Thailand [126] and Korea [127]. Urinary Cu was associated with a reduced eGFR in Mexican children [128].
In a Taiwanese COVID-19 cohort [122], urinary Cd levels > 2.05 μg/g creatinine were associated with 5.35-fold increase in the likelihood of suffering from severe symptoms from infections with COVID-19 [122].
In a Chinese COPD cohort [123], Cd exposure was found to contribute significantly to disease morbidity and mortality. The likelihood of having acute exacerbation and death rose, respectively 2.26-fold and 2.12-fold per 1 µg/L rise of blood Cd with adjustment for potential confounders and co-morbidity factors.
4.2. Public Health Significance of Environmental Cadmium
As data in Table 4 indicate, chronic exposure to Cd has been found to contribute substantially to mortality and the morbidity of highly prevalent health conditions and diseases, like hypertension, diabetes, and CKD. Cd exposure levels producing adverse effects on mortality and morbidity, inferred from blood and urinary Cd levels, were low, and these exposure levels were found in a significant proportion of the populations, documented below.
A study from Thailand reported the percentage of Cd excretion ≥ 1 µg/g creatinine among non-smoking women who had low body iron stores to be 22.5% [129]. In the U.S. general population with an overall mean urinary Cd excretion of 0.5 µg/g creatinine, 2.5%, 7.1%, and 16% of non-smoking women (aged ≥ 20 years) had urinary Cd levels > 1, > 0.7, and > 0.5 μg/g creatinine, respectively [130]. Thus, the proportion of U.S. women at risk of adverse health effects of Cd was not negligible. The estimated mean Cd intake among U.S. women (n = 1002, mean age 63.4) was 10.4 μg/day and the mean urinary excretion rate was 0.62 μg/g creatinine [131]. These dietary exposure and urinary Cd excretion were 21% and 12% of a tolerable intake level of 49.8 µg/day for an average 60-kg female (0.83 µg/kg body weight/day) and a urinary Cd threshold level of 5.24 µg/g creatinine, respectively.
In summary, current population exposure to environmental Cd presents a global public health significance and many challenges because the metal Cd is detectable in virtually all food types, especially in those frequently consumed in large quantities such as rice, potatoes, wheat, leafy salad vegetables, and other cereal crops. Current environmental Cd exposure has now reached toxic levels in a significant proportion of many populations. Alarmingly, CKD is predicted to become the 5th leading cause of years life lost by 2040 should an upward trend in its prevalence continue [132].
4.3. Evidence for Protetive Effects of Zinc
Lower Zn intake levels were found to be associated with higher urinary Cd levels in both men and women [133], and a 2.48-fold increase in risk of COPD was associated with urinary Cd levels > 0.79 μg/g creatinine [134]. There was a 1.89-fold increase in risk of having COPD, when Zn intake levels < 8.35 mg/day. This dietary Zn level was lower than the RDA values for women and men of 8 and 11 mg/day, respectively (Table 1).
An increased risk of having low eGFR was associated with serum Zn levels <74 μg/dL and blood Cd levels > 0.53 μg/L [135]. Among participants, enrolled in the U.S. NHANES 2003-2018 (n = 37,195), a biphasic dose-response relationship was noted between Zn intake levels and CKD risk, indicated by albuminuria or low eGFR [136]. The overall mean Zn intake level was 11.85 mg/day, while a significant decrease in risk of CKD was observed at Zn intake levels not higher 16.46 mg/day.
As expected, data above indicate the significance of optimum dietary intake of Zn, while suggesting the precaution regarding potential adverse outcomes of high intake of Zn. It is noteworthy, however, dietary Cd exposure levels among participants in the referred studies were considerably low, and yet health benefit was observed.
4.4. Cadmium, Zinc and Bone Resorption
4.4.1. Human Studies
Chronic ingestion of a high-dose Cd (>100 µg/day) is a known cause of itai-itai disease, marked by severe damage to kidneys and bones with multiple fractures due to osteoporosis and osteomalacia [137]. Notably, however, evidence for adverse effects of Cd on kidney and bone have been observed in residents of a Cd-contaminated area of Mae Sot, Tak Province of Thailand even though their Cd exposure levels were moderate [138-140]. Furthermore, as data in Table 4 indicate, the effects of Cd on the onset of osteoporosis and fracture have been observed also in a prospective cohort of a population exposed to a very low level of Cd like Sweden [120]. An increased excretion of the biomarkers indicative of defective mitochondrial function has been found in studies of Mae Sot residents and Swedish women [141,142] although exposure levels in Sweden were very low, compared to the residents of Cd contamination of Thailand. A bone effect in a Swedish study of Cd was observed in the absence of any effects on kidneys [120]. This may reflect a very high sensitivity of bone to mitochondrial effects of Cd, leading to enhanced ROS production and oxidative damage. Bone tissues from patients with osteoporosis had higher Cd levels, compared to controls without osteoporosis [143].
4.4.2. Experimental Studies
For osteoblast entry, Cd and Zn both utilize ZIP8 and ZIP14, which are expressed on the osteoblast surface [144]. Unlike Zn, however, no excretory mechanism exists for Cd, and Cd thus accumulates within osteoblasts [145]. Although this phenomenon has not been specifically investigated for bone cells, induction of ZnT1 expression by Cd may result in removal of Zn from bone cells, while Cd is retained (Section 3.1).
Ou et al. examined the effects of Cd on pre-osteoblasts (MC3T3-E1 subclone14 cell lines) at different concentrations for 60 hours. They demonstrated that the presence of Cd was associated with lower alkaline phosphatase (ALP) secretion, attenuated cell viability, and decreased mRNA expression of Runx2 and type I collagen [146]. Moreover, exposure to Cd led to DNA damage and induced apoptosis via the caspase-dependent pathway [146 7]. Zheng et al. cultivated osteoblasts obtained from fetal rat cranial bones with Cd at 1, 2, and 5 μM for two periods of time: 6 and 12 hours. They showed that Cd decreased cell viability in a dose-dependent manner. The authors found that the rate of osteoblast apoptosis was 10% after exposure to 1 μM CdCl2, while this rate rose to 30% with 5 μM CdCl2 [147]. Exposure to Cd led to the release of mitochondrial cytochrome c, resulting in Cd-mediated apoptosis linked with the mitochondrial p53 signaling pathway. On the other side, Zn reversed most of Cd effects and increased the viability of osteoblasts, the synthesis of ALP, collagen and bone mass [148].
Tian et al. exposed Saos-2 cell line to Cd as CdCl2 at 0.5, 1, 10, 20, 40, and 80 μM for 24-48 hours. Their results indicated that Cd higher than 10 μM decreased cell viability. They also observed a notable change in the morphology of the osteoblast nuclei, characterized by bending, fragmentation, and eventual nuclear collapse at 20 μM [149].
Chen et al. conducted a study in which they exposed osteoblasts obtained from rat calvarias to various concentrations of Cd (0, 0.125, 0.5, and 2.0 mol/L) for 24 hours, while osteoclasts obtained from long bone of rats were exposed to different Cd concentrations (0, 0.03 mol/L) for 72 hours. They found that Cd decreased the viability of osteoblasts and the activity of ALP in a dose-dependent manner. Moreover, the presence of Cd altered mineralization [150]. In another study, Cd at 0.03 mol/L potentiated RANKL expression which consequently increased the number of osteoclasts. In comparison, Zn down regulated RANKL/RANK expression and the osteoclastogenesis was diminished as a consequential result [151].
The mechanisms by which Cd induced osteoblast death were examined by Liu and colleagues. They utilized osteoblasts obtained from Sprague–Dawley rat fetuses treated with 0, 1, 2, and 5 μM Cd. They observed changes in osteoblast nucleus morphology, upregulation of Bax, but downregulation of Bcl-2 (which has an antiapoptotic activity) [152]. In another in vitro study utilizing MC-3T3-E1 cells, exposure to CdCl2 at 0-20 μM decreased cell viability, and promoted osteoblast apoptosis, which was resulted from a decreased Bcl-2 level and accumulation of Bax mRNA and protein, and interfered with osteoblast formation by decreasing the expression of RANKL [153].
A diagrammatically representation of Cd-induced bone resorption is provided in Figure 3.
As depicted in Figure 3, Cd causes osteoblast premature cell death and it stimulates the genesis of osteoclasts, thereby promoting bone resorption. Cd increases ZnT1 expression, leading extrusion of Zn and Zn deficiency conditions, while retaining Cd [143]. In comparison, Zn reduces osteoclast resorption activities and increases number of osteoblasts [154,155].
In summary, it is evident from series of experimental studies that Cd adversely affected osteoblast function. Cd and Zn appeared to impact the genesis of osteoclasts via the RANKL-RANK axis. Because Cd and Zn interact with the same transport proteins, an increased dietary Zn intake could potentially mitigate some detrimental effects of Cd on bone primarily through decreasing the number of osteoclasts.
5. Conclusions
Most dietary Zn is absorbed through highly specific transport proteins, namely ZIP4, ZnT5 and ZnT1, but Zn-amino acid co-transport mechanisms may also be involved. In comparison, Cd is absorbed through many transport proteins such as ZIP14, DMT1, ATP7A, TRPV5, TRPV6 which are necessary for the assimilation of essential metals Fe, Mn, Cu and Ca.
As a mechanism to prevent toxicity from Zn overload, Zn is extruded from cells by ZnT1, an efflux transporter for Zn only. No equivalent exit route for Cd exists and most acquired Cd is thus retained within cells
Zn is involved in the fundamental biological mechanisms that maintain normal cell metabolism and function. It is vital for cellular redox signaling and antioxidant defense mechanisms. Its antioxidative function is through the activity of SOD, Nox, and MT, and bilirubin, all of which prevent oxidative damage from excessive ROS. Through its interaction with MTF-1, like Zn, Cd induces simultaneously, MT and ZnT1, which can result in a reduction in Zn bioavailability, disruption cellular Zn homeostasis and induced Zn deficiency conditions.
Current environmental Cd concentrations, mean that many populations are now exposure to toxic levels of this metal and there is compelling evidence that this is impacting on the global mortality and morbidity. Risks of heart failure, coronary heart disease, and stroke are all increased with Cd exposure in a dose-dependent manner. A blood Cd level of 1 μg/L and urinary Cd excretion rate of 0.5 μg/g creatinine are associated with an increased risk of cardiovascular disease of 2.58-fold and 2.79-fold, respectively. Death from any causes among those with CKD or diabetes also rose with Cd exposure in dose-dependent manner.
The main source of Cd exposure in the non-smoking and non-occupationally exposed people is the food that is eaten. However, as the current dietary exposure guidelines are not low enough to reduce the risk of adverse health effects, public health measures should be instigated to help minimize Cd contamination of food chains. It would also be prudent to establish new health protective exposure guidelines. Ensuring adequate nutritional levels of essential metals, especially Zn, in populations at risk of Cd exposure is warranted.
Author Contributions
Conceptualization, S.S. and D.A.V.; writing—original draft preparation, Ana C.; Alek. C.; S.Y. and S.S.; writing—review and editing, S.S. and D.A.V. All authors have read and agreed to the published version of the manuscript.
Funding
It was supported with resources from the Centre for Kidney Disease Research, Translational Research Institute, and the Faculty of Medicine, University of Belgrade, Serbia.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Jean-Marc Moulis for his insightful comments on the Zn efflux transporter ZnT1 which is highly Zn selectivity over other metals and his conceptual framework for investigating metal homeostasis disruption as a primary target of Cd toxicity [109].
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Andreini, C.; Banci, L.; Bertini, I.; Rosato, A. Counting the zinc-proteins encoded in the human genome. J. Proteome Res. 2006, 5, 196–201. [Google Scholar] [CrossRef]
- King, J.C.; Shames, D.M.; Woodhouse, L.R. Zinc homeostasis in humans. J. Nutr. 2000, 130, 1360S–1366S. [Google Scholar] [CrossRef]
- King, J.C. Zinc: an essential but elusive nutrient. Am. J. Clin. Nutr. 2011, 94, 679S–684S. [Google Scholar] [CrossRef] [PubMed]
- Stiles, L.I.; Ferrao, K.; Mehta, K.J. Role of zinc in health and disease. Clin. Exp. Med. 2024, 24, 38. [Google Scholar] [CrossRef]
- Hall, A.G.; King, J.C. The molecular basis for zinc bioavailability. Int. J. Mol. Sci. 2023, 24, 6561. [Google Scholar] [CrossRef] [PubMed]
- Andrew, D.; Gail, R.; Morag, B.; Kishor, R. Recommended reference intervals for copper and zinc in serum using the US National Health and Nutrition Examination surveys (NHANES) data. Clin. Chim. Acta 2023, 546, 117397. [Google Scholar] [CrossRef]
- Barman, N.; Salwa, M.; Ghosh, D.; Rahman, M.W.; Uddin, M.N.; Haque, M.A. Reference value for serum zinc level of adult population in Bangladesh. EJIFCC 2020, 31, 117–124. [Google Scholar] [PubMed]
- IOM. Institute of Medicine, The National Academy of Sciences, Dietary reference intakes for vitamin A, vitamin K, arsenic, boron, chromium, copper, iodine, iron, manganese, molybdenum, nickel, silicon, vanadium, and zinc. Institute of Medicine. Food and Nutrition Board, NRC. Washington, DC: National Academies Press, USA, 2001; pp. 442–501. [CrossRef]
- King, J.C. Does zinc absorption reflect zinc status? Int. J. Vitam. Nutr. Res. 2010, 80, 300–306. [Google Scholar] [CrossRef]
- King, J.C. Yet again, serum zinc concentrations are unrelated to zinc intakes. J. Nutr. 2018, 148, 1399–1401. [Google Scholar] [CrossRef]
- McDonald, C.M.; Suchdev, P.S.; Krebs, N.F.; Hess, S.Y.; Wessells, K.R.; Ismaily, S.; Rahman, S.; Wieringa, F.T.; Williams, A.M.; Brown, K.H.; King, J.C. Adjusting plasma or serum zinc concentrations for inflammation: Biomarkers reflecting inflammation and nutritional determinants of anemia (BRINDA) project. Am. J. Clin. Nutr. 2020, 111, 927–937. [Google Scholar] [CrossRef]
- Molenda, M.; Kolmas, J. The role of zinc in bone tissue health and regeneration-A review. Biol. Trace Elem. Res. 2023, 201, 5640–5651. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, K. The interaction of zinc with the multi-functional plasma thyroid hormone distributor protein, transthyretin: evolutionary and cross-species comparative aspects. Biometals 2021, 34, 423–437. [Google Scholar] [CrossRef] [PubMed]
- Dabravolski, S.A.; Sadykhov, N.K.; Kartuesov, A.G.; Borisov, E.E.; Sukhorukov, V.N.; Orekhov, A.N. Interplay between Zn2+ homeostasis and mitochondrial functions in cardiovascular diseases and heart ageing. Int. J. Mol. Sci. 2022, 23, 6890. [Google Scholar] [CrossRef] [PubMed]
- Wessels, I.; Maywald, M.; Rink, L. Zinc as a gatekeeper of immune function. Nutrients 2017, 9, 1286. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Quan, Z.; Ni, J.; Li, H.; Qing, H. The many faces of the zinc finger protein 335 in brain development and immune system. Biomed. Pharmacother. 2023, 165, 115257. [Google Scholar] [CrossRef] [PubMed]
- Vickram, S.; Rohini, K.; Srinivasan, S.; Nancy Veenakumari, D.; Archana, K.; Anbarasu, K.; Jeyanthi, P.; Thanigaivel, S.; Gulothungan, G.; Rajendiran, N.; et al. Role of zinc (Zn) in human reproduction: A journey from initial spermatogenesis to childbirth. Int. J. Mol. Sci. 2021, 22, 2188. [Google Scholar] [CrossRef] [PubMed]
- King, J.C.; Brown, K.H.; Gibson, R.S.; Krebs, N.F.; Lowe, N.M.; Siekmann, J.H.; Raiten, D.J. Biomarkers of nutrition for development (BOND)-zinc review. J. Nutr. 2015, 146, 858S–885S. [Google Scholar] [CrossRef]
- Trame, S.; Wessels, I.; Haase, H.; Rink. L. A short 18 items food frequency questionnaire biochemically validated to estimate zinc status in humans. J. Trace Elem. Med. Biol. 2018, 49, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Lowe, N.M.; Hall, A.G.; Broadley, M.R.; Foley, J.; Boy, E.; Bhutta, Z.A. Preventing and controlling zinc deficiency across the life course: A call to action. Adv. Nutr. 2024, 15, 100181. [Google Scholar] [CrossRef]
- Satarug, S.; Phelps, K.R. Chapter 14: Cadmium Exposure and Toxicity. In Metal Toxicology Handbook; Bagchi, D., Bagchi, M., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 219–274. [Google Scholar]
- Satarug, S.; Vesey, D.A.; Gobe, G.C.; Phelps, K.R. Estimation of health risks associated with dietary cadmium exposure. Arch. Toxicol. 2023, 97, 329–358. [Google Scholar] [CrossRef]
- Satarug, S.; Baker, J.R.; Reilly, P.E.; Moore, M.R.; Williams, D.J. Changes in zinc and copper homeostasis in human livers and kidneys associated with exposure to environmental cadmium. Hum. Exp. Toxicol. 2001, 20, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Boon, P.E.; Pustjens, A.M.; Te Biesebeek, J.D.; Brust, G.M.H.; Castenmiller, J.J.M. Dietary intake and risk assessment of elements for 1- and 2-year-old children in the Netherlands. Food Chem. Toxicol. 2022, 161, 112810. [Google Scholar] [CrossRef] [PubMed]
- Fechner, C.; Hackethal, C.; Höpfner, T.; Dietrich, J.; Bloch, D.; Lindtner, O.; Sarvan, I. Results of the BfR MEAL Study: In Germany, mercury is mostly contained in fish and seafood while cadmium, lead, and nickel are present in a broad spectrum of foods. Food Chem. X 2022, 14, 100326. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Kataoka, Y.; Hayashi, K.; Matsuda, R.; Uneyama, C. Dietary exposure of the Japanese general population to elements: Total diet study 2013–2018. Food Saf. 2022, 10, 83–101. [Google Scholar] [CrossRef] [PubMed]
- Almerud, P.; Zamaratskaia, G.; Lindroos, A.K.; Bjermo, H.; Andersson, E.M.; Lundh, T.; Ankarberg, E.H. , Lignell, S. Cadmium, total mercury, and lead in blood and associations with diet, sociodemographic factors, and smoking in Swedish adolescents. Environ. Res. 2021, 197, 110991. [Google Scholar] [CrossRef] [PubMed]
- Pappas, R.S.; Fresquez, M.R.; Watson, C.H. Cigarette smoke cadmium breakthrough from traditional filters: Implications for exposure. J. Anal. Toxicol. 2015, 39, 45–51. [Google Scholar] [CrossRef]
- Kim, J.; Song, H.; Lee, J.; Kim, Y.J.; Chung, H.S.; Yu, J.M.; Jang, G.; Park, R.; Chung, W.; Oh, C.M.; Moon, S. Smoking and passive smoking increases mortality through mediation effect of cadmium exposure in the United States. Sci. Rep. 2023, 13, 3878. [Google Scholar] [CrossRef]
- Świetlik, R.; Trojanowska, M. Chemical fractionation in environmental studies of potentially toxic particulate-bound elements in urban air: A critical review. Toxics 2022, 10, 124. [Google Scholar] [CrossRef] [PubMed]
- Hill, D.T.; Jandev, V.; Petroni, M.; Atallah-Yunes, N.; Bendinskas, K.; Brann, L.S.; Heffernan, K.; Larsen, D.A.; MacKenzie, J.A.; Palmer, C.D.; et al. Airborne levels of cadmium are correlated with urinary cadmium concentrations among young children living in the New York state city of Syracuse, USA. Environ. Res. 2023, 223, 115450. [Google Scholar] [CrossRef]
- Willers, S.; Gerhardsson, L.; Lundh, T. Environmental tobacco smoke (ETS) exposure in children with asthma-relation between lead and cadmium, and cotinine concentrations in urine. Respir. Med. 2005, 99, 1521–1527. [Google Scholar] [CrossRef]
- Hsieh, C.Y.; Jung, C.R.; Lin, C.Y.; Hwang, B.F. Combined exposure to heavy metals in PM2.5 and pediatric asthma. J. Allergy Clin. Immunol. 2021, 147, 2171–2180. [Google Scholar] [CrossRef] [PubMed]
- Madrigal, J.M.; Persky, V.; Jackson, B.P.; Bain, A.; Siemer, M.; Pappalardo, A.A.; Argos, M. Assessment of metal concentrations and associations with pulmonary function among children with asthma in Chicago, Illinois. Int. J. Environ. Res. Public Health 2021, 18, 7279. [Google Scholar] [CrossRef] [PubMed]
- Xue M, Wang Q, Pang B, Zhang X, Zhang Y, Deng X, Zhang Z, Niu W. Association Between Circulating Zinc and Risk for Childhood Asthma and Wheezing: A Meta-analysis on 21 Articles and 2205 Children. Biol Trace Elem Res. 2024 Feb;202(2):442-453.
- Peng, X.; Li, C.; Zhao, D.; Huang, L. Associations of micronutrients exposure with cadmium body burden among population: A systematic review. Ecotoxicol. Environ. Saf. 2023, 256, 114878. [Google Scholar] [CrossRef] [PubMed]
- Rentschler, G.; Kippler, M.; Axmon, A.; Raqib, R.; Skerfving, S.; Vahter, M.; Broberg, K. Cadmium concentrations in human blood and urine are associated with polymorphisms in zinc transporter genes. Metallomics 2014, 6, 885–891. [Google Scholar] [CrossRef] [PubMed]
- JECFA. JECFA. In Proceedings of the Joint FAO/WHO Expert Committee on Food Additives and Contaminants, Seventy-Third Meeting, Geneva, Switzerland, 8–17 June 2010; In Summary and Conclusions; JECFA/73/SC; Food and Agriculture Organization of the United Nations/World Health Organization: Geneva, Switzerland, In Proceedings of the Joint FAO/WHO Expert Committee on Food Additives and Contaminants, Seventy-Third Meeting, Geneva, Switzerland, 8–17 June 2010; In Summary and Conclusions; JECFA/73/SC; Food and Agriculture Organization of the United Nations/World Health Organization: Geneva, Switzerland, 2011. Available online: https://apps.who.int/iris/handle/10665/44521 (accessed on 17 April 2024).
- Satarug, S.; Đorđević, A.B.; Yimthiang, S.; Vesey, D.A.; Gobe, G.C. The NOAEL equivalent of environmental cadmium exposure associated with GFR reduction and chronic kidney disease. Toxics 2022, 10, 614. [Google Scholar] [CrossRef] [PubMed]
- EFSA. European Food Safety Agency, Statement on tolerable weekly intake for cadmium. EFSA J. 2011, 9, 1975. [Google Scholar]
- Qing, Y.; Yang, J.; Zhu, Y.; Li, Y.; Zheng,W. ; Wu, M.; He, G. Dose-response evaluation of urinary cadmium and kidney injury biomarkers in Chinese residents and dietary limit standards. Environ. Health 2021, 20, 75. [Google Scholar] [CrossRef] [PubMed]
- Leconte, S.; Rousselle, C.; Bodin, L.; Clinard, F.; Carne, G. Refinement of health-based guidance values for cadmium in the French population based on modelling. Toxicol. Lett. 2021, 340, 43–51. [Google Scholar] [CrossRef]
- Schaefer, H.R.; Flannery, B.M.; Crosby, L.M.; Pouillot, R.; Farakos, S.M.S.; Van Doren, J.M.; Dennis, S.; Fitzpatrick, S.; Middleton, K. Reassessment of the cadmium toxicological reference value for use in human health assessments of foods. Regul. Toxicol. Pharmacol. 2023, 144, 105487. [Google Scholar] [CrossRef] [PubMed]
- Nishijo, M.; Nogawa, K.; Suwazono, Y.; Kido, T.; Sakurai, M.; Nakagawa, H. Lifetime cadmium exposure and mortality for renal diseases in residents of the cadmium-polluted Kakehashi River Basin in Japan. Toxics 2020, 8, 81. [Google Scholar] [CrossRef]
- Kippler, M.; Lönnerdal, B.; Goessler, W.; Ekström, E.C.; Arifeen, S.E.; Vahter, M. Cadmium interacts with the transport of essential micronutrients in the mammary gland - A study in rural Bangladeshi women. Toxicol. 2009, 257, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Castro, F.; Harari, F.; Llanos, M.; Vahter, M.; Ronco, A.M. Maternal-child transfer of essential and toxic elements through breast milk in a mine-waste polluted area. Am. J. Perinatol. 2014, 31, 993–1002. [Google Scholar] [CrossRef] [PubMed]
- Satarug, S. Is chronic kidney disease due to cadmium exposure inevitable and can it be reversed? Biomedicines 2024, 12, 718. [Google Scholar] [CrossRef] [PubMed]
- Satarug, S.; Vesey, D.A.; Ruangyuttikarn, W.; Nishijo, M.; Gobe, G.C.; Phelps, K.R. The source and pathophysiologic significance of excreted cadmium. Toxics 2019, 7, 55. [Google Scholar] [CrossRef] [PubMed]
- Moffett, D.B.; Mumtaz, M.M.; Sullivan, D.W., Jr.; Whittaker, M.H. Chapter 13, General Considerations of Dose-Effect and Dose-Response Relationships. In Handbook on the Toxicology of Metals, 5th ed.; Volume I: General, Considerations, Nordberg, G., Costa, M., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 299–317. [Google Scholar]
- Satarug, S.; Gobe, G.C.; Vesey, D.A.; Phelps, K.R. Cadmium and lead exposure, nephrotoxicity, and mortality. Toxics 2020, 8, 86. [Google Scholar] [CrossRef] [PubMed]
- Lang, S.M.; Schiffl, H. Smoking status, cadmium, and chronic kidney disease. Renal Replacement Therapy 2024, 10, 17. [Google Scholar] [CrossRef]
- Satarug, S.; Vesey, D.A.; Khamphaya, T.; Pouyfung, P.; Gobe, G.C.; Yimthiang, S. Estimation of the cadmium nephrotoxicity threshold from loss of glomerular filtration rate and albuminuria. Toxics 2023, 11, 755. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Taylor, A.W.; Riley, M.; Byles, J.; Liu, J.; Noakes, M. Association between dietary patterns, cadmium intake and chronic kidney disease among adults. Clin. Nutr. 2018, 37, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Chou, X.; Li, X.; Min, Z.; Ding, F.; Ma, K.; Shen, Y.; Sun, D.; Wu, Q. Sirtuin-1 attenuates cadmium-induced renal cell senescence through p53 deacetylation. Ecotoxicol. Environ. Saf. 2022, 245, 114098. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, M.; Xie, R. Associations between cadmium exposure and whole-body aging: mediation analysis in the NHANES. BMC Public Health 2023, 23, 1675. [Google Scholar] [CrossRef]
- Chen, L.; Zhao, Y.; Liu, F.; Chen, H.; Tan, T.; Yao, P.; Tang, Y. Biological aging mediates the associations between urinary metals and osteoarthritis among U.S. adults. BMC Med. 2022, 20, 207. [Google Scholar] [CrossRef] [PubMed]
- Kuliyev, E.; Zhang, C.; Sui, D.; Hu, J. Zinc transporter mutations linked to acrodermatitis enteropathica disrupt function and cause mistrafficking. J. Biol. Chem. 2021, 296, 100269. [Google Scholar] [CrossRef] [PubMed]
- Wiuf, A.; Steffen, J.H.; Becares, E.R.; Gronberg, C.; Mahato, D.R.; Rasmussen, S.G.F.; Andersson, M.; Croll, T.; Gotfryd, K.; Gourdon, P. The two-domain elevator-type mechanism of zinc-transporting ZIP proteins. Sci. Adv. 2022, 8, eabn4331. [Google Scholar] [CrossRef] [PubMed]
- Valentine, R.A.; Jackson, K.A.; Christie, G.R.; Mathers, J.C.; Taylor, P.M.; Ford, D. ZnT5 variant B is a bidirectional zinc transporter and mediates zinc uptake in human intestinal Caco-2 cells. J. Biol. Chem. 2007, 282, 14389–14393. [Google Scholar] [CrossRef] [PubMed]
- Cragg, R.A.; Phillips, S.R.; Piper, J.M.; Varma, J.S.; Campbell, F.C.; Mathers, J.C.; Ford, D. Homeostatic regulation of zinc transporters in the human small intestine by dietary zinc supplementation. Gut 2005, 54, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Nishito, Y.; Kambe, T. Absorption mechanisms of iron, copper, and zinc: An overview. J. Nutr. Sci. Vitaminol. 2018, 64, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Hoch, E.; Lin, W.; Chai, J.; Hershfinkel, M.; Fu, D.; Sekler, I. Histidine pairing at the metal transport site of mammalian ZnT transporters controls Zn2+ over Cd2+ selectivity. Proc. Natl. Acad. Sci. USA 2012, 109, 7202–7207. [Google Scholar] [CrossRef] [PubMed]
- Aydemir, T.B. , Cousins, R.J. The multiple faces of the metal transporter ZIP14 (SLC39A14). J. Nutr. 2018, 148, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Guthrie, G.J.; Aydemir, T.B.; Troche, C.; Martin, A.B.; Chang, S.M.; Cousins, R.J. Influence of ZIP14 (slc39A14) on intestinal zinc processing and barrier function. Am. J. Physiol.-Gastrointest. Liver Physiol. 2015, 308, G171–G178. [Google Scholar] [CrossRef]
- Park, J.D. , Cherrington, N.J.; Klaassen, C.D. Intestinal absorption of cadmium is associated with divalent metal transporter 1 in rats. Toxicol. Sci. 2002, 68, 288–294. [Google Scholar] [CrossRef]
- Garrick, M.D.; Dolan, K.G.; Horbinski, C.; Ghio, A.J.; Higgins, D.; Porubcin, M.; Moore, E.G.; Hainsworth, L.N.; Umbreit, J.N.; Conrad, M.E.; et al. DMT1: A mammalian transporter for multiple metals. Biometals 2003, 16, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Fujishiro, H.; Hamao, S.; Tanaka, R.; Kambe, T.; Himeno, S. Concentration-dependent roles of DMT1 and ZIP14 in cadmium absorption in Caco-2 cells. J. Toxicol. Sci. 2017, 42, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, Y. Iron from the gut: the role of divalent metal transporter 1. J. Clin. Biochem. Nutr. 2024, 74, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.J.; Shawki, A.; Ganz, T.; Nemeth, E.; Mackenzie, B. Functional properties of human ferroportin, a cellular iron exporter reactive also with cobalt and zinc. Am. J. Physiol. Cell Physiol. 2014, 306, C450–C459. [Google Scholar] [CrossRef] [PubMed]
- Kondaiah, P.; Yaduvanshi, P.S.; Sharp, P.A.; Pullakhandam, R. Iron and zinc homeostasis and interactions: Does enteric zinc excretion cross-talk with intestinal iron absorption? Nutrients 2019, 11, 1885. [Google Scholar] [CrossRef] [PubMed]
- Nose, Y.; Kim, B.E.; Thiele, D.J. Ctr1 drives intestinal copper absorption and is essential for growth, iron metabolism, and neonatal cardiac function. Cell Metab. 2006, 4, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Ohta, H.; Ohba, K. Involvement of metal transporters in the intestinal uptake of cadmium. J. Toxicol. Sci. 2020, 45, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Diaz de Barboza, G.; Guizzardi, S.; Tolosa de Talamoni, N. Molecular aspects of intestinal calcium absorption. World J. Gastroenterol. 2015, 21, 7142–7154. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, G.; Danko, T.; Bergeron, M.J.; Balazs, B.; Suzuki, Y.; Zsembery, A.; Hediger, M.A. Heavy metal cations permeate the TRPV6 epithelial cation channel. Cell Calcium 2011, 49, 43–55. [Google Scholar] [CrossRef]
- Kovacs, G.; Montalbetti, N.; Franz, M.C.; Graeter, S.; Simonin, A.; Hediger, M.A. Human TRPV5 and TRPV6: Key players in cadmium and zinc toxicity. Cell Calcium 2013, 54, 276–286. [Google Scholar] [CrossRef]
- Wasserman, R.H.; Chandler, J.S.; Meyer, S.A.; Smith, C.A.; Brindak, M.E.; Fullmer, C.S.; Penniston, J.T.; Kumar, R. Intestinal calcium transport and calcium extrusion processes at the basolateral membrane. J. Nutr. 1992, 122(3 Suppl), 662–671. [Google Scholar] [CrossRef]
- Wasserman, R.H. Vitamin D and the dual processes of intestinal calcium absorption. J. Nutr. 2004, 134, 3137–3139. [Google Scholar] [CrossRef] [PubMed]
- Bronner, F. Recent developments in intestinal calcium absorption. Nutr. Rev. 2009, 67, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Hoch, E.; Levy, M.; Hershfinkel, M.; Sekler, I. Elucidating the H+ Coupled Zn2+ Transport Mechanism of ZIP4; Implications in acrodermatitis enteropathica. Int. J. Mol. Sci. 2020, 21, 734. [Google Scholar] [CrossRef] [PubMed]
- Pérez de la Lastra, J.M.; Andrés-Juan, C.; Plou, F.J.; Pérez-Lebeña, E. Theoretical three-dimensional zinc complexes with glutathione, amino acids and flavonoids. Stresses 2021, 1, 123–141. [Google Scholar] [CrossRef]
- Knez, M.; Graham, R.D.; Welch, R.M.; Stangoulis, J.C. New perspectives on the regulation of iron absorption via cellular zinc concentrations in humans. Crit. Rev. Food Sci. Nutr. 2017, 57, 2128–2143. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, H.; Miyazaki, T.; Nodera, M.; Miyajima, Y.; Suzuki, T.; Kido, T.; Suka, M. Zinc-excess intake causes the deterioration of renal function accompanied by an elevation in systemic blood pressure primarily through superoxide radical-induced oxidative stress. Int. J. Toxicol. 2014, 33, 288–296. [Google Scholar] [CrossRef]
- Fujita, Y.; el Belbasi, H.I.; Min, K.S.; Onosaka, S.; Okada, Y.; Matsumoto, Y.; Mutoh, N.; Tanaka, K. Fate of cadmium bound to phytochelatin in rats. Res. Commun. Chem. Pathol. Pharmacol. 1993, 82, 357–365. [Google Scholar] [PubMed]
- Langelueddecke, C.; Roussa, E.; Fenton, R.A.; Thévenod, F. Expression and function of the lipocalin-2 (24p3/NGAL) receptor in rodent and human intestinal epithelia. PLoS ONE 2013, 8, e71586. [Google Scholar] [CrossRef] [PubMed]
- Langelueddecke, C.; Lee, W.K.; Thévenod, F. Differential transcytosis and toxicity of the hNGAL receptor ligands cadmium-metallothionein and cadmium-phytochelatin in colon-like Caco-2 cells: Implications for in vivo cadmium toxicity. Toxicol. Lett. 2014, 226, 228–235. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Nomiyama, T.; Kumagai, N.; Dekio, F.; Uemura, T.; Takebayashi, T.; Nishiwaki, Y.; Matsumoto, Y.; Sano, Y.; Hosoda, K.; et al. Uptake of cadmium in meals from the digestive tract of young non-smoking Japanese female volunteers. J. Occup. Health 2003, 45, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Horiguchi, H.; Oguma, E.; Sasaki, S.; Miyamoto, K.; Ikeda, Y.; Machida, M.; Kayama, F. Comprehensive study of the effects of age, iron deficiency, diabetes mellitus, and cadmium burden on dietary cadmium absorption in cadmium-exposed female Japanese farmers. Toxicol. Appl. Pharmacol. 2004, 196, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Thévenod, F.; Fels, J.; Lee, W.K.; Zarbock, R. Channels, transporters and receptors for cadmium and cadmium complexes in eukaryotic cells: myths and facts. Biometals 2019, 32, 469–489. [Google Scholar] [CrossRef]
- Thévenod, F.; Lee, W.K. Cadmium transport by mammalian ATP-binding cassette transporters. Biometals 2024. [Google Scholar] [CrossRef]
- Beyersmann, D.; Haase, H. Functions of zinc in signaling, proliferation and differentiation of mammalian cells. Biometals 2001, 14, 331–341. [Google Scholar] [CrossRef]
- Krezel, A.; Hao, Q.; Maret, W. The zinc/thiolate redox biochemistry of metallothionein and the control of zinc ion fluctuations in cell signaling. Arch. Biochem. Biophys. 2007, 463, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Hübner, C.; Haase, H. Interactions of zinc- and redox-signaling pathways. Redox Biol. 2021, 41, 101916. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Smith, M.J.; Siow, R.C.M.; Aarsland, D.; Maret, W.; Mann, G.E. Interactions between zinc and NRF2 in vascular redox signalling. Biochem. Soc. Trans. 2024, 52, 269–278. [Google Scholar] [CrossRef]
- Kambe, T.; Tsuji, T.; Hashimoto, A.; Itsumura, N. The physiological, biochemical, and molecular roles of zinc transporters in zinc homeostasis and metabolism. Physiol. Rev. 2015, 95, 749–784. [Google Scholar] [CrossRef]
- Hara, T.; Takeda, T.A.; Takagishi, T.; Fukue, K.; Kambe, T.; Fukada, T. Physiological roles of zinc transporters: molecular and genetic importance in zinc homeostasis. J. Physiol. Sci. 2017, 67, 283–301. [Google Scholar] [CrossRef]
- Hara, T.; Yoshigai, E.; Ohashi, T.; Fukada, T. Zinc transporters as potential therapeutic targets: An updated review. J. Pharmacol. Sci. 2022, 148, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Nishito, Y.; Kambe, T. Zinc transporter 1 (ZNT1) expression on the cell surface is elaborately controlled by cellular zinc levels. J. Biol. Chem. 2019, 294, 15686–15697. [Google Scholar] [CrossRef] [PubMed]
- Laity, J.H.; Andrews, G.K. Understanding the mechanisms of zinc-sensing by metal-response element binding transcription factor-1 (MTF-1). Arch. Biochem. Biophys. 2007, 463, 201–210. [Google Scholar] [CrossRef]
- Mor, M.; Beharier, O.; Cook, D.I.; Campbell, C.R.; Gheber, L.A.; Katz, A.; Moran, A.; Etzion, Y. ZnT1 induces a crosstalk between T-type and L-type calcium channels through interactions with Raf-1 kinase and the calcium channel beta2 subunit. Metallomics 2023, 15, mfad031. [Google Scholar] [CrossRef]
- Sun, C.; He, B.; Gao, Y.; Wang, X.; Liu, X.; Sun, L. Structural insights into the calcium-coupled zinc export of human ZnT1. Sci. Adv. 2024, 10, eadk5128. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Jomova, K.; Rhodes, C.J.; Kuča, K.; Musílek, K. Redox- and non-redox-metal-induced formation of free radicals and their role in human disease. Arch. Toxicol. 2016, 90, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Harraz, M.M.; Marden, J.J.; Zhou, W.; Zhang, Y.; Williams, A.; Sharov, V.S.; Nelson, K. Luo, M.; Paulson, H.; Schoneich, C.; et al. SOD1 mutations disrupt redox-sensitive Rac regulation of NADPH oxidase in a familial ALS model. J. Clin. Invest.
- Li, M.S.; Adesina, S.E.; Ellis, C.L.; Gooch, J.L.; Hoover, R.S.; Williams, C.R. NADPH oxidase-2 mediates zinc deficiency-induced oxidative stress and kidney damage. Am. J. Physiol. Cell Physiol. 2017, 312, C47–C55. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, J.; Ryu, M.S. Cellular zinc deficiency impairs heme biosynthesis in developing erythroid progenitors. Nutrients 2023, 15, 281. [Google Scholar] [CrossRef]
- Kumagai, A.; Ando, R.; Miyatake, H.; Greimel, P.; Kobayashi, T.; Hirabayashi, Y.; Shimogori, T.; Miyawaki, A. A bilirubin-inducible fluorescent protein from eel muscle. Cell 2013, 153, 1602–1611. [Google Scholar] [CrossRef]
- Takeda, T.A.; Mu, A.; Tai, T.T.; Kitajima, S.; Taketani, S. Continuous de novo biosynthesis of haem and its rapid turnover to bilirubin are necessary for cytoprotection against cell damage. Sci. Rep. 2015, 5, 10488. [Google Scholar] [CrossRef]
- Vitek, L.; Hinds, T.D. Jr.; Stec, D.E.; Tiribelli, C. The physiology of bilirubin: health and disease equilibrium. Trends Mol. Med. 2023, 29, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Petering, D.H. Reactions of the Zn proteome with Cd2+ and other xenobiotics: Trafficking and toxicity. Chem. Res. Toxicol. 2016, 30, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Moulis, J.M. Cellular mechanisms of cadmium toxicity related to the homeostasis of essential metals. Biometals 2010, 23, 877–896. [Google Scholar] [CrossRef] [PubMed]
- Nzengue, Y.; Candéias, S.M.; Sauvaigo, S.; Douki, T.; Favier, A.; Rachidi, W.; Guiraud, P. The toxicity redox mechanisms of cadmium alone or together with copper and zinc homeostasis alteration: its redox biomarkers. J. Trace Elem. Med. Biol. 2011, 25, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Moulis, J.M.; Bourguinon, J.; Catty, P. Chapter 23 Cadmium. In RSC Metallobiology Series No. 2, Binding, Transport. and Storage of Metal. Ions in Biological Cells; Wolfgang, M., Anthony, W., Eds.; The Royal Society of Chemistry: London, UK, 2014; pp. 695–746. [Google Scholar]
- Satarug, S.; Baker, J.R.; Reilly, P.E.; Moore, M.R.; Williams, D.J. Changes in zinc and copper homeostasis in human livers and kidneys associated with exposure to environmental cadmium. Hum. Exp. Toxicol. 2001, 20, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Kusak, R.; Nasiadek, M.; Stragierowicz, J.; Hanke, W.; Kilanowicz, A. Changes in endogenous essential metal homeostasis in the liver and kidneys during a six-month follow-up period after Subchronic Cadmium Exposure. Int. J. Mol. Sci. 2024, 25, 3829. [Google Scholar] [CrossRef]
- Satarug, S.; Garrett, S.H.; Somji, S.; Sens, M.A.; Sens, D.A. Zinc, zinc transporters, and cadmium cytotoxicity in a cell culture model of human urothelium. Toxics 2021, 9, 94. [Google Scholar] [CrossRef] [PubMed]
- Nagamatsu, S.; Nishito, Y.; Yuasa, H.; Yamamoto, N.; Komori, T.; Suzuki, T.; Yasui, H.; Kambe, T. Sophisticated expression responses of ZNT1 and MT in response to changes in the expression of ZIPs. Sci. Rep. 2022, 12, 7334. [Google Scholar] [CrossRef] [PubMed]
- Verzelloni, P.; Urbano, T.; Wise, L.A.; Vinceti, M.; Filippini, T. Cadmium exposure and cardiovascular disease risk: A systematic review and dose-response meta-analysis. Environ. Pollut. 2024, 345, 123462. [Google Scholar] [CrossRef]
- Chen, S.; Shen, R.; Shen, J.; Lyu, L.; Wei, T. Association of blood cadmium with all-cause and cause-specific mortality in patients with hypertension. Front. Public Health 2023, 11, 1106732. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, D.; Shi, F.; Wang, F.; Liu, X.; Wen, H.; Mubarik, S.; Yu, C. Association of serum 25(OH)D, cadmium, CRP with all-cause, cause-specific mortality: A prospective cohort study. Front. Nutr. 2022, 9, 803985. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, X.; Ma, Z.; Dang, Y.; Yang, Y.; Cao, S.; Ouyang, C.; Shi, X.; Pan, J.; Hu, X. Associations of urinary and blood cadmium concentrations with all-cause mortality in US adults with chronic kidney disease: A prospective cohort study. Environ. Sci. Pollut. Res. Int. 2023, 30, 61659–61671. [Google Scholar] [CrossRef] [PubMed]
- Tägt, J.; Helte, E.; Donat-Vargas, C.; Larsson, S.C.; Michaëlsson, K.; Wolk, A.; Vahter, M.; Kippler, M.; Åkesson, A. Long-term cadmium exposure and fractures, cardiovascular disease, and mortality in a prospective cohort of women. Environ. Int. 2022, 161, 107114. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.H.; Wang, C.W.; Wu, D.W.; Lee, W.H.; Chen, Y.C.; Li, C.H.; Tsai, C.C.; Lin, W.Y.; Chen, S.C.; Hung, C.H.; et al. Association of heavy metals with overall mortality in a Taiwanese population. Nutrients 2021, 13, 2070. [Google Scholar] [CrossRef] [PubMed]
- Chiu, L.C.; Lee, C.S.; Hsu, P.C.; Li, H.H.; Chan, T.M.; Hsiao, C.C.; Kuo, S.C.; Ko, H.W.; Lin, S.M.; Wang, C.H.; et al. Urinary cadmium concentration is associated with the severity and clinical outcomes of COVID-19: a bicenter observational cohort study. Environ. Health 2024, 23, 29. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Deng, Y.P.; Xu. J.; Zhu, F.M.; He, Q.Y.; Tang, M.M., Liu, Y.; Yang, J.; Liu, H.Y., Fu, L.; et al. Association of blood cadmium concentration with chronic obstructive pulmonary disease progression: a prospective cohort study. Respir. Res. 2024, 25, 91. [Google Scholar] [CrossRef] [PubMed]
- Nogawa, K.; Yamada, Y.; Honda, R.; Tsuritani, I.; Kobayashi, E.; Ishizaki, M. Copper and zinc levels in serum and urine of cadmium-exposed people with special reference to renal tubular damage. Environ. Res. 1984, 33, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, M.; Kobayashi, E.; Suwazono, Y.; Uetani, M.; Oishi, M.; Inaba, T.; Kido, T.; Shaikh, Z.A.; Nogawa, K. Excretion of urinary cadmium, copper, and zinc in cadmium-exposed and non-exposed subjects, with special reference to urinary excretion of beta2-microglobulin and metallothionein. Biol. Trace Elem. Res. 2005, 108, 17–31. [Google Scholar] [CrossRef]
- Satarug, S.; Nishijo, M.; Ujjin, P.; Moore, M.R. Chronic exposure to low-level cadmium induced zinc-copper dysregulation. J. Trace Elem. Med. Biol. 2018, 46, 32–38. [Google Scholar] [CrossRef]
- Eom, S.Y.; Yim, D.H.; Huang, M.; Park, C.H.; Kim, G.B.; Yu, S.D.; Choi, B.S.; Park, J.D.; Kim, Y.D.; Kim, H. Copper-zinc imbalance induces kidney tubule damage and oxidative stress in a population exposed to chronic environmental cadmium. Int. Arch. Occup. Environ. Health 2020, 93, 337–344. [Google Scholar] [CrossRef]
- Ortega-Romero, M.; Rojas-Lima, E.; Rubio-Gutiérrez, J.C.; Aztatzi-Aguilar, O.G.; Narváez-Morales, J.; Esparza-García, M.; Barrera-Hernández, Á.; Mejia, M.Á.; Mendez-Hernández, P.; Medeiros, M.; et al. Associations among environmental exposure to trace elements and biomarkers of early kidney damage in the pediatric population. Biometals 2024. [Google Scholar] [CrossRef] [PubMed]
- Satarug, S.; Ujjin, P.; Vanavanitkun, Y.; Baker, J.R.; Moore, M.R. Influence of body iron store status and cigarette smoking on cadmium body burden of healthy Thai women and men. Toxicol. Lett. 2004, 148, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, M.E.; Wong, L.Y.; Osterloh, J.D. Smoking status and urine cadmium above levels associated with subclinical renal effects in U.S. adults without chronic kidney disease. Int. J. Hyg. Environ. Health 2011, 214, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Quraishi, S.M.; Adams, S.V.; Shafer, M.; Meliker, J.R.; Li, W.; Luo, J.; Neuhouser, M.L.; Newcomb, P.A. Urinary cadmium and estimated dietary cadmium in the Women's Health Initiative. J. Expo. Sci. Environ. Epidemiol. 2016, 26, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Foreman, K.J.; Marquez, N.; Dolgert, A.; Fukutaki, K.; Fullman, N.; McGaughey, M.; Pletcher, M.A.; Smith, A.E.; Tang, K.; Yuan, C.W.; et al. Forecasting life expectancy, years of life lost, and all-cause and cause-specific mortality for 250 causes of death: Reference and alternative scenarios for 2016-40 for 195 countries and territories. Lancet 2018, 392, 2052–2090. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.S.; Caffrey, J.L.; Lin, J.W.; Bayliss, D.; Faramawi, M.F.; Bateson, T.F.; Sonawane, B. Increased risk of cancer mortality associated with cadmium exposures in older Americans with low zinc intake. J. Toxicol. Environ. Health A 2013, 76, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.S.; Caffrey, J.L.; Chang, M.H.; Dowling, N.; Lin, J.W. Cigarette smoking, cadmium exposure, and zinc intake on obstructive lung disorder. Respir. Res. 2010, 11, 53. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.S.; Ho, W.C.; Caffrey, J.L.; Sonawane, B. Low serum zinc is associated with elevated risk of cadmium nephrotoxicity. Environ. Res. 2014, 134, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhang, H.; Yang, Y.; Cao, Y.; Liang, D. Association between dietary zinc intake and increased renal function in US adults. Biol. Trace Elem. Res. 2023. [CrossRef]
- Aoshima, K. Epidemiology of renal tubular dysfunction in the inhabitants of a cadmium-polluted area in the Jinzu river basin in Toyama Prefecture. Tohoku J. Exp. Med. 1987, 152, 151–172. [Google Scholar] [CrossRef]
- Nambunmee, K.; Honda, R.; Nishijo, M.; Swaddiwudhipong, W.; Nakagawa, H.; Ruangyuttikarn, W. Bone resorption acceleration and calcium reabsorption impairment in a Thai population with high cadmium exposure. Toxicol. Mech. Methods 2010, 20, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Nishijo, M.; Nambunmee, K.; Suvagandha, D.; Swaddiwudhipong, W.; Ruangyuttikarn, W.; Nishino, Y. Gender-specific impact of cadmium exposure on bone metabolism in older people living in a cadmium-polluted area in Thailand. Int. J. Environ. Res. Public Health 2017, 14, 401. [Google Scholar] [CrossRef] [PubMed]
- Nambunmee, K.; Nishijo, M.; Swaddiwudhipong, W.; Ruangyuttikarn, W. Bone fracture risk and renal dysfunction in a highly cadmium exposed Thai population. J. Res. Health Sci. 2018, 18, e00419. [Google Scholar] [PubMed]
- Ellis, J.K.; Athersuch, T.J.; Thomas, L.D.; Teichert, F.; Pérez-Trujillo, M.; Svendsen, C.; Spurgeon, D.J.; Singh, R.; Järup, L.; Bundy, J.G.; et al. Metabolic profiling detects early effects of environmental and lifestyle exposure to cadmium in a human population. BMC Med. 2012, 10, 61. [Google Scholar] [CrossRef] [PubMed]
- Suvagandha, D.; Nishijo, M.; Swaddiwudhipong, W.; Honda, R.; Ohse, M.; Kuhara, T.; Nakagawa, H.; Ruangyuttikarn, W. A biomarker found in cadmium exposed residents of Thailand by metabolome analysis. Int. J. Environ. Res. Public Health 2014, 11, 3661–3677. [Google Scholar] [CrossRef] [PubMed]
- Ran, D.; Zhou, D.; Liu, G. ; Ma. Y.; Ali, W.; Yu, R.; Wang, Q.; Zhao, H.; Zhu, J.; Zou, H.; et al. Reactive oxygen species control osteoblast apoptosis through SIRT1/PGC-1α/P53Lys382 signaling, mediating the onset of Cd-induced osteoporosis. J. Agric. Food Chem. 2023. [Google Scholar] [CrossRef]
- Aydemir, T.B.; Troche, C.; Kim, J.; Kim, M.H.; Teran, O.Y.; Leeuwenburgh, C.; Cousins, R.J. Aging amplifies multiple phenotypic defects in mice with zinc transporter Zip14 (Slc39a14) deletion. Exp. Gerontol. 2016, 85, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, S.; Tsukamoto, M.; Saito, M.; Hojyo, S.; Fukada, T.; Takami, M.; Furuichi, T. Disruption of the mouse Slc39a14 gene encoding zinc transporter ZIP14 is associated with decreased bone mass, likely caused by enhanced bone resorption. FEBS Open Bio 2018, 8, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Ou, L.; Wang, H.; Wu, Z.; Wang, P.; Yang, L.; Li, X.; Sun, K.; Zhu, X.; Zhang, R. Effects of cadmium on osteoblast cell line: Exportin 1 accumulation, p-JNK activation, DNA damage and cell apoptosis. Ecotoxicol. Environ. Saf. 2021, 208, 111668. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Zhuo, L.; Ran, D.; Ma, Y.; Luo, T.; Zhao, H.; Song, R.; Zou, H.; Zhu, J.; Gu, J.; et al. Cadmium induces apoptosis via generating reactive oxygen species to activate mitochondrial p53 pathway in primary rat osteoblasts. Toxicol. 2020, 446, 152611. [Google Scholar] [CrossRef]
- Seo, H.J.; Cho, Y.E.; Kim, T.; Shin, H.I.; Kwun, I.S. Zinc may increase bone formation through stimulating cell proliferation, alkaline phosphatase activity and collagen synthesis in osteoblastic MC3T3-E1 cells. Nutr. Res. Pract. 2010, 4, 356–361. [Google Scholar] [CrossRef]
- Tian, J.; Li, Z.; Wang, L.; Qiu, D.; Zhang, X.; Xin, X.; Cai, Z.; Lei, B. Metabolic signatures for safety assessment of low-level cadmium exposure on human osteoblast-like cells. Ecotoxicol. Environ. Saf. 2021, 207, 111257. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhu, G.; Gu, S.; Jin, T.; Shao, C. Effects of cadmium on osteoblasts and osteoclasts in vitro. Environ. Toxicol. Pharmacol. 2009, 28, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Amin, N.; Clark, C.C.T.; Taghizadeh, M.; Djafarnejad, S. Zinc supplements and bone health: The role of the RANKL-RANK axis as a therapeutic target. J. Trace Elem. Med. Biol. 2020, 57, 126417. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Dai, N.; Wang, Y.; Xu, C.; Zhao, H.; Xia, P.; Gu, J.; Liu, X.; Bian, J.; Yuan, Y.; et al. Role of autophagy in cadmium-induced apoptosis of primary rat osteoblasts. Sci. Rep. 2016, 6, 20404. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Shen, H.; Zhu, J.; Zhu, Y.; He, Y.; Li, Z.; Lu, H. Geniposide attenuates cadmium-induced oxidative stress injury via Nrf2 signaling in osteoblasts. Mol. Med. Rep. 2019, 20, 1499–1508. [Google Scholar] [CrossRef] [PubMed]
- Hadley, K.B.; Newman, S.M.; Hunt, J.R. Dietary zinc reduces osteoclast resorption activities and increases markers of osteoblast differentiation, matrix maturation, and mineralization in the long bones of growing rats. J. Nutr. Biochem. 2010, 21, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Nagata, M.; Lönnerdal, B. Role of zinc in cellular zinc trafficking and mineralization in a murine osteoblast-like cell line. J. Nutr. Biochem. 2011, 22, 172–178. [Google Scholar] [CrossRef]
Figure 1.
The tubulo-glomerular effects of cadmium. Phase 1, a subtle effect of Cd is indicated by urinary excretion of KIM1. Phase 2, an intensified effect of Cd is indicated by urinary excretion of CdMT and NAG and a fall of eGFR. Phase 3, overt effects Cd on glomerular and tubular functions are indicated by a fall of eGFR to 60 mL/min/1.73m2, and a rise of urinary β2M excretion to 300 µg/g creatinine. Abbreviation: ALB, albumin, β2M, β2-microglobulin; CdMT, cadmium-metallothionine complexes; KIM1, kidney injury molecule 1; NAG, N-acetyl-β-D-glucosaminidase.
Figure 1.
The tubulo-glomerular effects of cadmium. Phase 1, a subtle effect of Cd is indicated by urinary excretion of KIM1. Phase 2, an intensified effect of Cd is indicated by urinary excretion of CdMT and NAG and a fall of eGFR. Phase 3, overt effects Cd on glomerular and tubular functions are indicated by a fall of eGFR to 60 mL/min/1.73m2, and a rise of urinary β2M excretion to 300 µg/g creatinine. Abbreviation: ALB, albumin, β2M, β2-microglobulin; CdMT, cadmium-metallothionine complexes; KIM1, kidney injury molecule 1; NAG, N-acetyl-β-D-glucosaminidase.
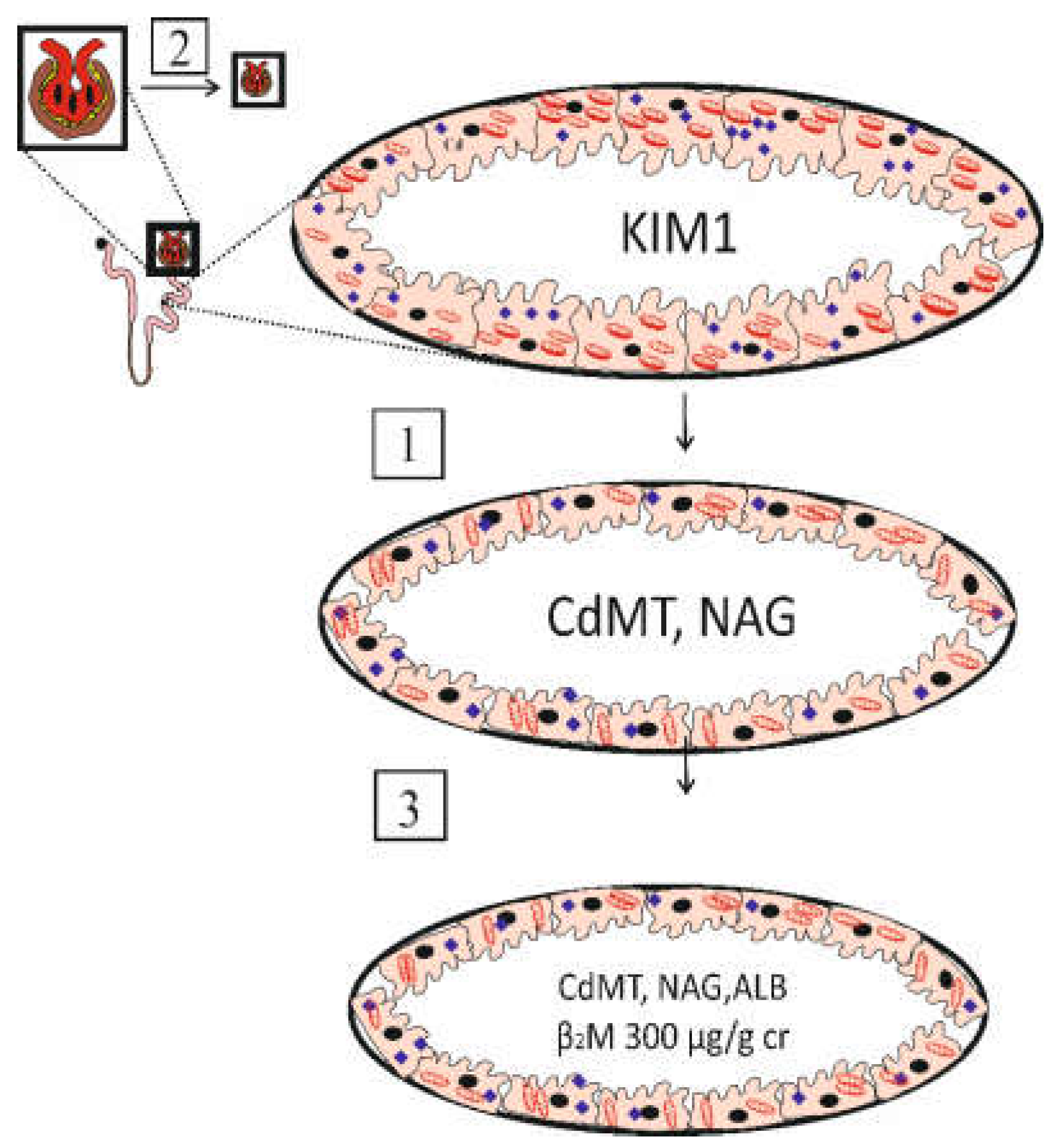
Figure 2.
Cellular homeostasis of zinc and its antioxidative function. ZIP14, ZIP8, DMT1 mediate Zn entry into cells. ZnT1 mediates Zn extrusion. The antioxidative function of Zn is through MT, SOD1, and heme biosynthesis, which supplies a substrate precursor for producing bilirubin. Abbreviations: GLUT, glucose transporter; SOD, superoxide dismutase; δ-ALA, delta-aminolevulinic acid; ALAD, aminolevulinic acid dehydratase; PPXI, protoporphyrin XI; ER, endoplasmic reticulum; CPR, cytochrome P450 reductase; HO-1, heme oxygenase-1; HO-2, heme oxygenase-2, BVR, biliverdin reductase; CdRE, cadmium response element; MRE, metal response element; MTF-1, metal-response element binding transcription factor-1.
Figure 2.
Cellular homeostasis of zinc and its antioxidative function. ZIP14, ZIP8, DMT1 mediate Zn entry into cells. ZnT1 mediates Zn extrusion. The antioxidative function of Zn is through MT, SOD1, and heme biosynthesis, which supplies a substrate precursor for producing bilirubin. Abbreviations: GLUT, glucose transporter; SOD, superoxide dismutase; δ-ALA, delta-aminolevulinic acid; ALAD, aminolevulinic acid dehydratase; PPXI, protoporphyrin XI; ER, endoplasmic reticulum; CPR, cytochrome P450 reductase; HO-1, heme oxygenase-1; HO-2, heme oxygenase-2, BVR, biliverdin reductase; CdRE, cadmium response element; MRE, metal response element; MTF-1, metal-response element binding transcription factor-1.
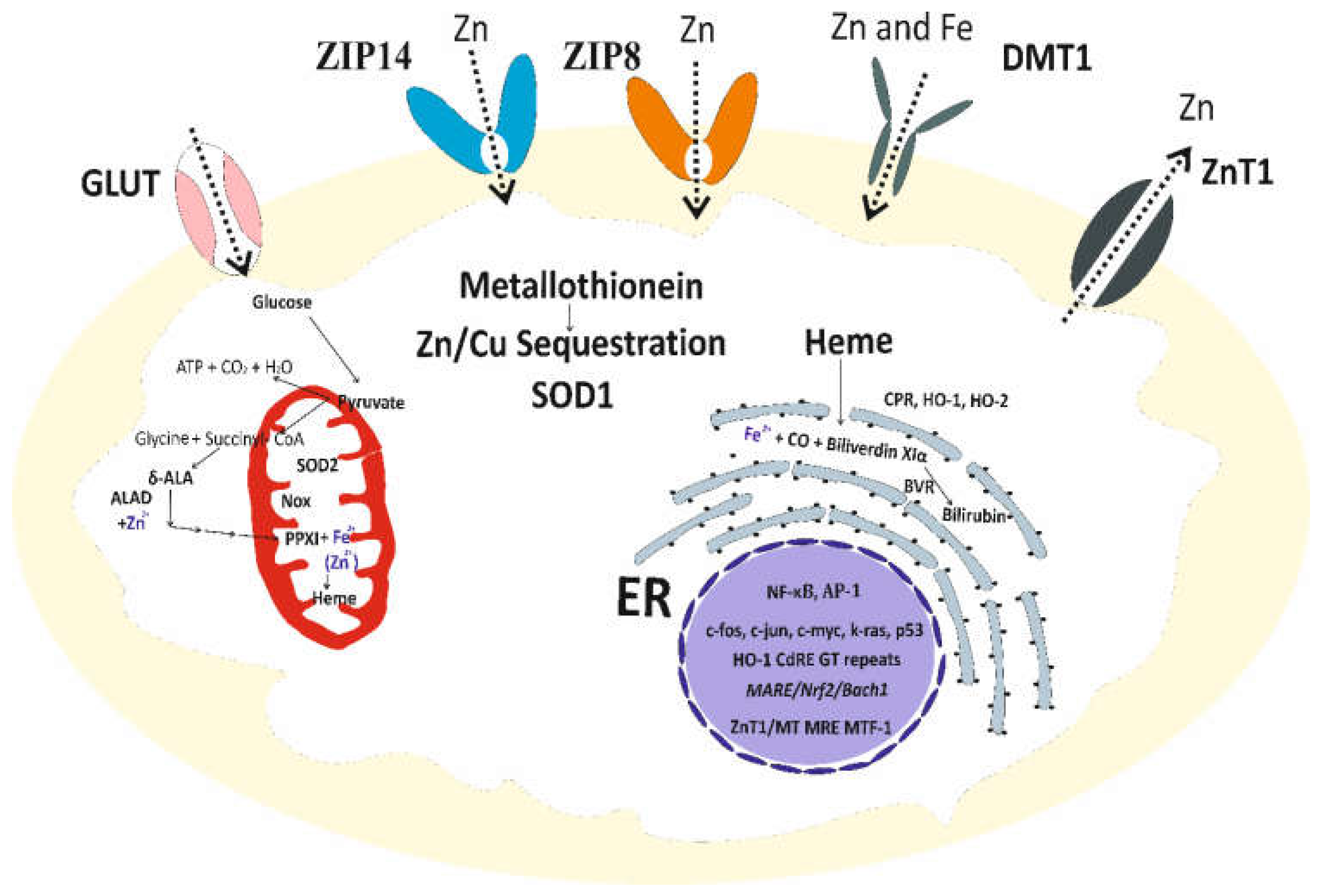
Figure 3.
Bone resorption induced by cadmium (upper row) and maintenance of bone mass by zinc and (lower row). Effects of Cd and Zn are through the RANKL-RANK axis. Abbreviations: Cd, cadmium; Zn, zinc; RANKL, receptor activator of nuclear factor kappa-Β ligand; RANK, receptor activator of nuclear factor kappa-Β.
Figure 3.
Bone resorption induced by cadmium (upper row) and maintenance of bone mass by zinc and (lower row). Effects of Cd and Zn are through the RANKL-RANK axis. Abbreviations: Cd, cadmium; Zn, zinc; RANKL, receptor activator of nuclear factor kappa-Β ligand; RANK, receptor activator of nuclear factor kappa-Β.
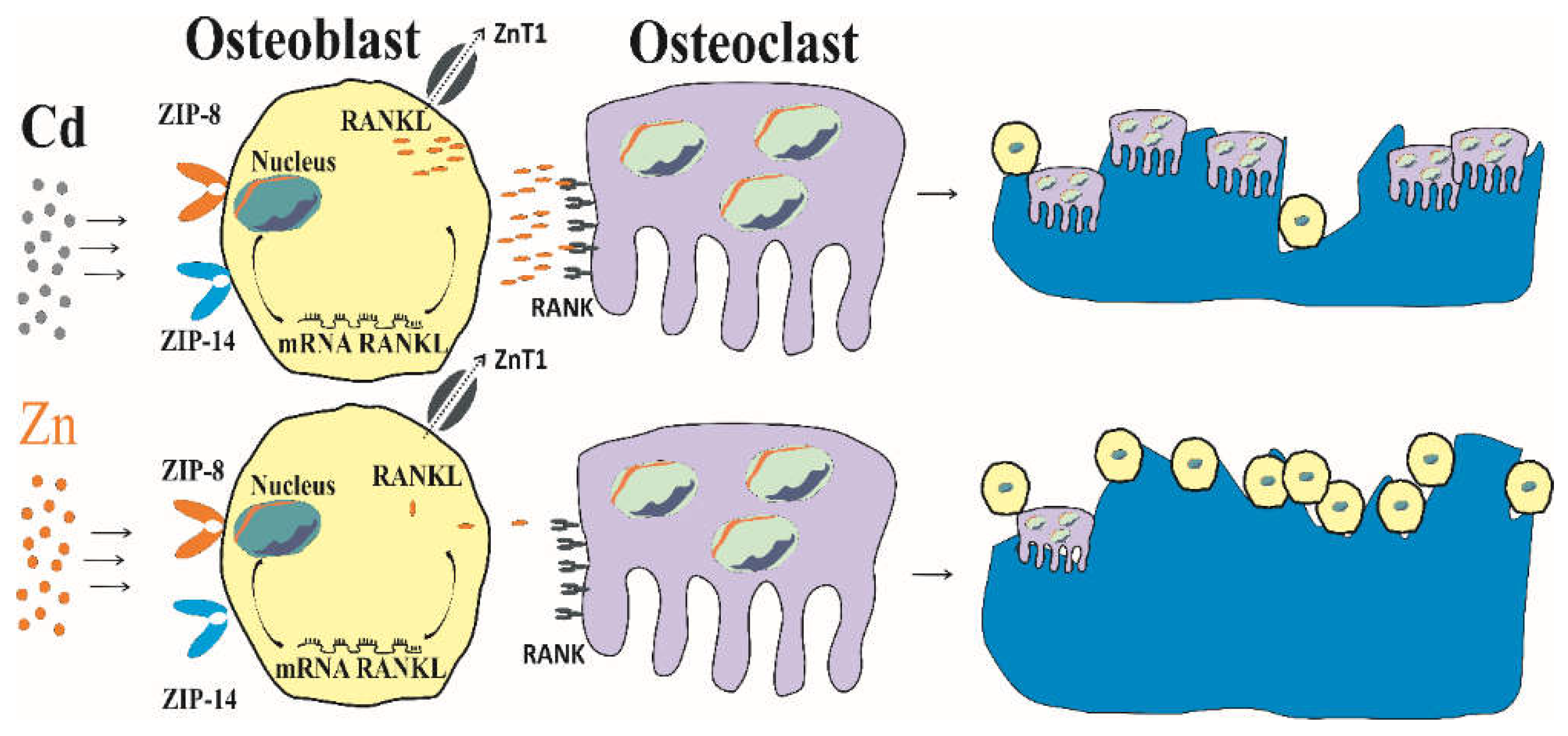

Table 1.
Recommended zinc intake levels versus tolerable intake levels of cadmium.
| Nutrient/ Contaminant |
Recommended Zinc Intake Level versus Tolerable Cadmium Intake Level | Reference |
|---|---|---|
| Zinc (atomic weight 65.4) | An adult female (60 kg): 8 mg/day, or 0.13 mg per kg of body weight per day. An adult male (70 kg): 11 mg/day, or 0.16 mg per kg of body weight per day. Infants: 2–3 mg/day. Children: 5–9 mg/day. Pregnant women: 11–12 mg/day. Lactating women: 12–13 mg/day. |
The U.S. National Academy of Sciences Institute of Medicine (IOM) [8] |
| Cadmium (atomic A weight 112.4) | A tolerable lifetime intake of 2 g or an intake of 0.83 μg per kg body weight per day (58 µg per day for a 70-kg person). a Assumption: Absorption rate at 3-7%; urinary excretion of β2-microglobulin (β2M) levels ≥ 300 µg/g creatinine indicated kidney toxicity with a threshold level of 5.24 μg Cd/g creatinine. |
JECFA [38] |
| Cadmium | BMDL or the NOAEL equivalent value of Cd exposure was 0.0104 µg/L of filtrate (≈ 0.01−0.02 µg/g creatinine). Assumption: Cd accumulation level producing a 5% decline in the eGFR represented an upper limit of a permissible Cd exposure level. |
Satarug et al., 2022 [39] |
| Cadmium | A tolerable intake of 0.36 μg/kg body weight per day (25.2 µg per day for a 70-kg person) Assumption: Urinary β2M excretion levels ≥ 300 µg/g creatinine indicated kidney toxicity with a threshold level of 1 μg Cd/g creatinine |
EFSA [40] |
| Cadmium | A tolerable intake level of 0.28 μg per kg body weight per day (16.8 µg per day for a 60- kg person). Assumption: Urinary β2M excretion levels ≥ 300 µg/g creatinine to indicate kidney toxicity with a threshold level of 3.07 μg Cd/g creatinine. |
Qing et al., 2021 [42] |
| Cadmium | A tolerable intake level of 0.35 μg Cd per kg body weight per day, assuming a bone toxicity threshold level of 0.5 μg/g creatinine. | Leconte et al., 2021 [42] |
| Cadmium | By reverse dosimetry PBPK model, a tolerable intake levels range between 0.21 and 0.36 μg per kg body weight per day, assuming a similar bone and kidney toxicity threshold level of 0.5 μg/g creatinine. | Schaefer et al., 2023 [43] |
PBPK, physiologically based pharmacokinetics; BMDL, benchmark dose limit; NOAEL, no-observed-adverse-effect level. a On a molar basis, a tolerable lifetime intake of Cd at 2 g is half of total Zn in the body of 2 g. A life time intake of 1 g of Cd was linked to a 49% increase in the likelihood of dying from kidney failure [44].
Table 2.
Specialized transport proteins for absorption of metal nutrients and cadmium.
| Metal | Transporter | Localization | Description |
|---|---|---|---|
| Zn | SLC39A4 (ZIP4) |
Apical membrane |
Two-zinc binding sites in histidine-rich motif [57]. Transport dietary Zn into enterocytes. Functional loss due to mutation causes zinc deficiency [58]. |
| Zn | SLC30A5 (ZnT5) variant B |
Apical membrane |
Transport Zn into enterocytes [59]. Zn supplementation depressed ZIP4 and ZnT5B protein levels in the ileum [60]. |
| Zn | SLC30A1 (ZnT1) | Basolateral membrane | Transport Zn from the cytoplasm to the extracellular medium or intracellular vesicles [61,62]. Histidine-rich motif confers Zn selectivity over Cd. This Cd discrimination is unique to the mammalian ZnT1 [62]. |
| Fe, Mn, Cd |
SLC39A14 (ZIP14) | Basolateral membrane |
Trafficking Zn to tight junctions, especially in the jejunum for the maintenance of the intestinal barrier [63,64]. |
| Fe, Cd Cu, Zn |
SLC11A2 (DMT1) |
Apical membrane |
The same high affinity for Cd as it does for Fe (Km 0.5~1 μM); High abundance in the duodenum [65-68]. |
| Fe, Zn, Co | SLC40A1 (FPN1) |
Basolateral membrane | Transport Fe, Zn and Co, but not of Cu, Cd, or Mn from basolateral membrane into the portal blood [69,70]. |
| Cu, Fe, Zn | SLC31A1 (CTR1) |
Apical membrane, Early endosome | Transport dietary Cu into enterocytes [61,71]. |
| Cu, Cd | ATP7A | Trans-Golgi network, Cytosol Basolateral membrane |
Transport Cu into the portal blood and the ATP7A mutations are associated with Menkes disease [61]. May contribute to Cd absorption [72]. |
| Ca, Cd | TRPV5, TRPV6 | Apical membrane |
Transport Ca2+ into enterocytes [73] and may provide Cd entry routes into enterocytes [74,75]. |
| Ca | Calbindin-D9k | Cytoplasm | Transport Ca to basolateral membrane and extrusion of Ca into portal blood [76-78]. Expression in the ileum is induced by 1,25-dihydroxycholecalciferol [77]. |
SLC, solute-linked carrier; ZIP4, Zrt- and Irt-related protein 4; ZIP14, Zrt- and Irt-related protein14; ZnT1; zinc transport 1; DMT1, divalent metal transporter 1; FPN1, ferroportin 1; CTR1, copper transporter 1; ATP7A, copper transporting ATPases (Cu-ATPases); TRPV5, transient receptor potential vanilloid5; TRPV6, transient receptor potential vanilloid6.
Table 3.
Changes in ZnT and ZIP transcript levels in UROtsa cells after exposure to cadmium.
| Transporters | Transcripts per 1000 β-Actin | ||||
|---|---|---|---|---|---|
| Batch I, 0 µM Cd2+ |
Batch II, 0 µM Cd2+ |
1 µM Cd2+ | 2 µM Cd2+ | 4 µM Cd2+ | |
| SLC30A | |||||
| ZnT1 | 181 ± 23 | 365 ± 38 | 3007 ± 465 | 1434 ± 146 | 1216 ± 153 *** |
| ZnT2 | 0.01 ± 0.001 | 0.06 ± 0.01 | 73 ± 15 | 16 ± 1.9 | 11 ± 1.5 *** |
| ZnT3 | 0.03 ± 0.007 | 0.15 ± 0.01 | 0.24 ± 0.05 | 0.10 ± 0.02 | 0.10 ± 0.02 * |
| ZnT4 | 1.6 ± 0.26 | 11.4 ± 0.8 | 10 ± 1 | 8.7 ± 1 | 6.2 ± 0.5 ** |
| ZnT5 | 150 ± 19 | 510 ± 30 | 1038 ± 132 | 495 ± 54 | 568 ± 91 ** |
| ZnT6 | 4.5 ± 0.15 | 65 ± 8 | 77 ± 6 | 63 ± 13 | 57 ± 12 |
| ZnT7 | 734 ± 28 | 758 ± 76 | 1007 ± 136 | 706 ± 44 | 488 ± 63 * |
| ZnT10 | 0.04 ± 0.005 | 1.1 ± 0.2 | 2.4 ± 0.2 | 1.7 ± 0.2 | 1.1 ± 0.1 *** |
| SLC39A | |||||
| ZIP1 | 19.5 ± 2.0 | 82 ± 9 | 99 ± 15 | 55 ± 10 | 59 ± 12 * |
| ZIP2 | 0.02 ± 0.004 | 1.2 ± 0.1 | 0.8 ± 0.2 | 0.4 ± 0.1 | 0.2 ± 0.03 *** |
| ZIP3A | 9.1 ± 0.4 | 19 ± 1 | 23 ± 2.3 | 17 ± 1.7 | 14 ± 1.3 * |
| ZIP3B | 0.48 ± 0.05 | 4.1 ± 0.2 | 6.2 ± 0.7 | 4.4 ± 0.2 | 4.2 ± 0.4 * |
| ZIP4 | 0.18 ± 0.04 | 0.06 ± 0.01 | 0.06 ± 0.01 | 0.04 ± 0.01 | 0.06 ± 0.01 |
| ZIP5 | 0.01 ± 0.003 | 0.01 ± 0.001 | 0.01 ± 0.003 | 0.01 ± 0.002 | 0.01 ± 0.002 |
| ZIP6 | 18.3 ± 2.6 | 92 ± 8 | 133 ± 12 | 80 ± 9 | 75 ± 10 ** |
| ZIP7 | 121 ± 9.5 | 204 ± 25 | 342 ± 69 | 149 ± 32 | 94 ± 21 *** |
| ZIP8 | 0.09 ± 0.01 | 2.1 ± 0.2 | 2.6 ± 0.3 | 2.0 ± 0.2 | 2.7 ± 0.4 |
| ZIP10 | 5 ± 0.3 | 54 ± 4 | 30 ± 8 | 14 ± 3 | 14 ± 3 *** |
| ZIP14 | 83.4 ± 10.5 | 146 ± 19 | 218 ± 24 | 158 ± 26 | 128 ± 19 * |
Number of transcripts for each transporter was expressed per 1000 transcripts of the β-actin gene [114]. *p = 0.01–0.05; **p = 0.001; ***p ≤ 0.001.
Table 4.
Effects of lifetime exposure to environmental cadmium on mortality and disease risks.
| Study Population | Cadmium Exposure and Effects Observed | Reference |
|---|---|---|
| United States NHANES, 1999–2012, n = 12,208 with hypertension Mean follow-up 10.8 years |
Among those with hypertension, respective HR (95% CI) for all-cause, CVD and Alzheimer’s disease mortality were 1.85 (1.59, 2.14), 1.76 (1.33, 2.34), and 3.41 (1.54, 7.51), comparing blood Cd levels ≥ 0.80 versus ≤ 0.25 μg/L. HR (95% CI) for CVD mortality among non-smokers who had hypertension was 2.12 (1.36, 3.30) |
Chen et al., 2023 [117] |
| United States NHANES, 2001–2010, n = 2945 with diabetes Mean follow-up period, 9.1 years |
Among those with diabetes, HR (95% CI) for all-cause mortality was 1.49 (1.06, 2.09) at urinary Cd levels > 0.60 μg/L. HR (95% CI) for all-cause mortality was 1.65 (1.24, 2.19) at serum CRP levels > 0.49 mg/dL. HR (95% CI) for cancer mortality was 3.25 (1.82, 5.80) at serum CRP levels > 0.49 mg/dL. |
Liu et al., 2022 [118] |
| United States NHANES, 1999–2014, n = 1825 with CKD Mean follow-up period, 6.8 years |
Among those with CKD, HR (95% CI) for all-cause mortality was 1.75 (1.28, 2.39) at urinary Cd levels ≥ 0.60 μg/g creatinine. HR (95%CI) for death from any causes was 1.59 (1.17, 2.15) at blood Cd levels ≥ 0.70 μg/L. |
Zhang et al., 2023 [119] |
| Sweden n = 4024 women, aged 56-85 years Mean follow-up 10.5 years |
Respective HR (95% CI) for all-cause mortality and any fracture were 1.38 (1.10, 1.74) and 1.20 (1.01,1.43), comparing the top tertile of urinary Cd (median 0.54 µg/g creatinine) with the bottom tertial (median urinary Cd of 0.20 µg/g creatinine). | Tägt et al., 2022 [120] |
| Taiwan n = 2497 (1001 males, 1496 females), Median follow-up 3.5 years. |
HR (95%CI) for all-cause mortality was 1.35 (1.09, 1.68) per 1 μg/L increment of urinary Cd. HR (95%CI) for all-cause mortality was 1.35 (1.15, 1.58) per 1 μg/dL increment of urinary copper. |
Liu et al., 2021 [121] |
| Taiwan n = 252 with a severe COVID-19, n = 322 with non-severe COVID-19 June 2022− July 2023 |
Among COVID-19 infected subjects, OR (95%CI) for severe symptoms was 5.35 (1.12, 25.6) at urinary Cd levels > 2.05 μg/g creatinine. | Chiu et al., 2024 [122] |
| China n = 196 COPD patients Enrolment: September 2020-June 2022, Follow-up 2 years |
Per every 1 µg/L increase in blood Cd, respective OR (95% CI) for acute exacerbation, hospitalization and mortality were 2.24 (1.17, 4.29), 1.28 (1.13, 1.56) and 1.69 (1.06, 2.70). After adjustment for potential confounders, respective OR (95%CI) for acute exacerbation and mortality were 2.26 (1.03, 4.24) and 2.12 (1.13, 7.11) per 1 µg/L-increment of blood Cd. |
Sun et al., 2024 [123] |
HR, hazard ratio; OR, odds ratio; CI, confidence interval; CRP, c-reactive protein; NHANES, National Health and Nutrition Examination Survey
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



