
Submitted:
22 April 2024
Posted:
24 April 2024
You are already at the latest version
Abstract
Plant responses to different light and water availability are variable among species and their re-spective phenotypic plasticity, and the combination between these two abiotic factors can alleviate or intensify stressful effects. This study aimed to evaluate the impacts of exposure time of Cedrela fissilis Vell. seedlings to different water and light availability and the interaction of these factors. Seedlings were submitted to combinations of three shading levels – SH (0, 30 and 70%) and three water regimes based on the water holding capacity (WHC) in the substrate, constituting nine cul-tivation conditions: T1 – 0% SH + 40% WHC; T2 – 0% SH + 70% WHC; T3 – 0% SH + 100% WHC; T4 – 30% SH + 40% WHC; T5 – 30% SH + 70% WHC; T6 – 30% SH and 100% WHC; T7 – 70% SH + 40% WHC; T8 – 70% SH +70% WHC; T9 – 70% SH + 100% WHC. C. fissilis seedlings respond to water deficit, here represented by 40% WHC, regardless of exposure time, and when cultivated in full sun, the stressful effects were enhanced, acting in a synergistic manner. The condition that provided better gas exchange performance and greater total dry mass accumulation for C. fissilis seedlings was 30% shading combined with 100% WHC. C. fissilis seedlings have physiological plasticity and resilience to survive under different water and light conditions.
Keywords:
cedro
; abiotic stress
; phenotypic plasticity
; photosynthetic metabolism
1. Introduction
Currently, there is greater need for practices aimed at maintaining natural resources and planted forests in order to meet the objectives of sustainable development. It is important to understand the interactions between environmental changes and plant species to implement strategies that drive a future agenda for the agroforestry system, boosting forest recovery and restoration [1,2].
In their initial development phase, especially when they are in the seedling phase, plants are highly responsive to the abiotic factors of the environment in which they are inserted or produced [3]. These factors fluctuate in a marked way due to global climate changes that have occurred in recent years [4]. Decreased precipitation, changes in incident light, increased evapotranspiration and increased air temperature are some factors that can influence plant growth, development and performance.
Afforestation and reforestation of areas have increased and consequently seedling production, and therefore the cultivation conditions must be known to obtain good quality seedlings. To achieve this objective, knowledge of the water and light requirements of each species and its adjustment potential through plasticity is essential. Through adjustments in photosynthetic biochemistry, leaf anatomy and morphology, plants have the ability to acclimatize to different light and water availability, which can guarantee their survival and improve their growth [5,6].
Light is essential for plant growth and development, which regulates its physiology throughout its cycle; however, it is necessary to understand the ecological classification of each species as either too little or too much light exposure can lead to a stressful cultivation condition, especially at the initial phase. As water availability, especially water deficit, is also a limiting factor for the establishment and development of forest species, plants respond to low soil water content through a complex chain of mechanisms that constitute their adaptation strategies [7].
Each species responds in a different way when submitted to different light and water availability, and depending on the interaction between these two factors, it can alleviate stressful effects [8,9]. For example, some species tolerate water deficits under high solar radiation, others are more adapted to shady and dry environments, while others do not tolerate full sunlight or water deficits. These responses are important, as they define species with desirable characteristics for forest restoration and classify those that need shading to develop in open areas and even species that require shading and regular irrigation if used in reforestation. The usefulness of a given species is determined by its tolerance to stressors at the initial stages of its life cycle [10].
Cedrela fissilis Vell. (Meliaceae) is among native species with potential for restoration of degraded areas and enrichment plantations, which prefers clayey, well-drained soils [11,12]. This species has high ecological and economic value and has been gaining prominence in the national and international forestry sector, being recommended for landscaping, urban afforestation, among others. These characteristics led to predatory logging, which resulted in the species currently being classified as vulnerable to extinction [13].
However, there are divergences regarding the ecological classification of this species as some authors classify it as a pioneer species [14], while others classify it as an initial secondary species [15], in which case, excess shade can be stressful for C. fissilis, and understanding its mechanism when submitted to different light levels and which of these conditions can be stressful or can mitigate the deficit is important.
The present study aimed to understand the effects and impacts of exposure time to different water and light availability and the interaction of these factors on gas exchange and the initial growth of Cedrela fissilis Vell seedlings. Given the above, the following questions were proposed: (i) is water deficit stressful for C. fissilis seedlings, regardless of exposure time? (ii) as a pioneer species, can excess shade harm the development of C. fissilis seedlings? (iii) does shading accentuate the effects of water deficit on the photosynthetic and growth responses of C. fissilis? (iv) does this species show resilience and plasticity to adapt to different light and water conditions?
2. Results
2.1. Visual Aspects
C. fissilis seedlings were responsive to different shading levels and water regimes, in which those grown in 30% and 70% SH with 40% WHC presented better visual appearance than those grown in 0% SH in the same water regime (Figure 1). Furthermore, it is noteworthy that as water availability increased under 0 and 30% SH, seedlings showed greater height and number of leaves, while seedlings under 70% SH had greater uniformity regardless of water regime.
2.2. Gas Exchange and Chlorophyll Index
The highest A values were observed in C. fissilis seedlings grown under 0% SH + 100% WHC and 30% SH + 100% WHC (8.0 and 8.8 μmol CO2 m-2 s-1, respectively). On the other hand, the lowest A values (< 2.0 μmol CO2 m-2 s-1) occurred in seedlings with 40% WHC, regardless of shading level. However, it was observed that from 45 days onwards, A values under 0% SH and 100% WHC increased (Figure 2A and Table 1).
Regarding gs at 15 days, the highest values occurred in seedlings under conditions of 30% and 70% SH + 100% WHC (0.13 and 0.12 mol H2O m-2 s-1), and the lowest were related to seedlings grown in 0% and 30% SH + 40% WHC (0.02 and 0.03 mol H2O m-2 s-1). Reduction in gs in all cultivation conditions at 30 and 45 days was observed, with resumption after 45 days. At 60 days of evaluation, the highest value occurred in seedlings grown in 30% SH + 100% WHC, whereas the lowest values were observed in seedlings grown in 70% SH + 40% WHC in 30% SH + 70% WHC (Figure 2B).
At 15 and 60 days, the highest A/Ci values were observed in seedlings under 0% SH + 100% WHC in full sun. The lowest values occurred in seedlings grown in 70% SH + 40% WHC. At 30 and 45 days, the highest values were observed in seedlings with 30% SH + 70% WHC and 70% SH + 100% WHC. On the other hand, at the same times, seedlings grown in 0% SH + 40% WHC, 70% + 40% WHC and 30% SH + 40% WHC showed lower values (Figure 2C).
The highest E value occurred at 15 days in seedlings grown in 0% SH + 100% WHC, at 30 and 45 days, the same seedlings were those with the lowest values. The lowest value observed at 15 days occurred in seedlings grown in 0% SH + 40% WHC, while at 30, 45 and 60 days, increase in E values was observed. In general, seedlings corresponding to cultivation conditions of 30% SH + 40% WHC showed lower values at all assessment times (Figure 2D).
For WUE, the factors under study were significant in isolation. Among cultivation conditions, the highest WUE value was observed in seedlings grown in 0% SH + 100% WHC (Figure 3A). Regarding evaluation times, A/E values were lower at 15 and 60 days and higher at 30 and 45 days (Figure 3B).
The chlorophyll index (CI) was influenced by the interaction between factors under study, in which the highest values were observed in seedlings at 70% SH + 70% WHC at all evaluation times. At 30 and 45 days, seedlings grown in 70% SH had the highest CI values, and the lowest values were observed in seedlings grown in 0% SH, regardless of WHC, at the same time. Seedlings at 0% SH + 40% WHC were those with the lowest CI values throughout the evaluation times (Figure 3C).
2.3. Chlorophyll a Fluorescence
The effective quantum yield of photochemical energy conversion in FSII (Fv/F0) was higher in seedlings grown under 70% SH + 100% WHC, followed by seedlings grown in 70% SH + 40% WHC and 70% SH + 70% WHC environments. The lowest values were observed for seedlings under 0% SH + 40% WHC and 0% SH + 70% WHC (Figure 4A).
Regarding the potential quantum efficiency of FSII Fv/Fm, seedlings under 70% SH were those that presented the highest values regardless of WHC, and the lowest values were found in seedlings under 0% SH + 40% WHC (Figure 4C). Regarding evaluation times, the highest Fv/F0 and Fv/Fm values were 2.67 and 0.683 at 32 and 37 days, respectively (Figure 4B and 4D).
2.4. Relative Leaf Water Content and Growth
WHC, LA, RL and TDM values were influenced by the interaction between cultivation conditions and evaluation times. Regarding relative water content (RWC), the highest values observed at 15 days occurred in seedlings grown in 0% SH + 100% WHC and the lowest in those grown in 0% SH + 40% WHC. At 60 days, the highest values were in seedlings under 0% SH + 70% WHC, 30% SH + 70% WHC, 70% SH + 40 WHC and 70% SH + 100% WHC, while the lowest value was observed in cedar seedlings grown in 0% SH + 40% WHC (Figure 5A).
At 15 and 60 days, C. fissilis seedlings submitted to the cultivation conditions of 70% SH + 70% WHC and 70% SH + 100% WHC showed the highest leaf area values. On the other hand, at 15 days, the lowest values were observed for seedlings under 0% SH + 40% WHC and 0% SH + 100% WHC and in 30% SH + 40% WHC, and at 60 days, values were observed at 0 %SH + 40% WHC (Figure 5B).
The highest root length values at 15 days were observed for seedlings submitted to 30 and 70% SH, both with 70% WHC, the other cultivation conditions at the same time did not differ from each other, corresponding to the lowest values. At 60 days, seedlings under 70% SH + 100% WHC had the highest values, while seedlings under 0% SH + 100% WHC showed the lowest RL values (Figure 5C).
Regarding total dry mass (TDM), the highest values were observed for seedlings at 0% SH + 70 and 100% WHC and in those at 30% SH + 100% WHC at 15 days. However, in the same period, seedlings under 40% WHC had lower values, regardless of shading level. At 60 days, the highest and lowest TDM values occurred in seedlings grown in 30% SH + 100% WHC and 0% SH + 40% WHC, respectively (Figure 5D).
2.5. Phenotypic Plasticity
Seedlings grown in 30% SH corresponded to the highest phenotypic plasticity index values for A and TDM. The highest PPI values for Fv/Fm and LA were observed for seedlings grown in full sun. Seedlings grown in the 70% SH environment presented lower PPI values for A, Fv/Fm, LA and TDM (Table 2).
3. Discussion
C. fissilis seedlings are sensitive to water deficit, here represented by 40% WHC, regardless of exposure time, as they showedreduced photosynthetic metabolism and growth, and when cultivated in full sun, these stressful effects were enhanced, acting in a synergistic way. However, it was observed that shading, especially 70% SH, mitigates the stressful effect of water deficit.
Changes in the soil water regime can influence the morphological and physiological responses of plants [16,17]. Reductions in A, gs and E values in seedlings grown in 40% WHC were observed, regardless of shading levels. However, the full sun environment further accentuated stress due to water deficit, because under this cultivation condition, in addition to the fact that soil water status is lower, there is greater evaporation of water from the substrate and exposure to higher temperatures, a fact contrary to the shaded environment, which reduces the evapotranspiration intensity, contributing to the maintenance of RWC in cells and their turgidity. Similarly, [18] observed that 30% and 70% shading in the cultivation of Campomanesia xanthocarpa (Mart.) O. Berg minimized the negative effects of water deficit on gas exchange, and the authors associate this response to the fact that shading allowed less water loss through the substrate, which favored stomatal opening for longer time and resulted in greater CO2 assimilation.
One of the first responses to stress due to water scarcity is stomatal closure, which is mediated by a complex signaling network involving abscisic acid [19], as a mechanism to prevent excessive water loss through leaf transpiration and maintain water potential [7,20,21], a fact observed in seedlings at 30 days under 0% SH + 40% WHC, which reduced gs. Likewise, [7] found that C. fissilis showed greater transpiration rate control as a mechanism to reduce water loss.
At the same time that stomatal closure acts in water regulation, this mechanism limits the efficiency of the biochemical step of photosynthesis in C. fissilis seedlings. According to [22], this reduction is due to the fact that there is a decrease in mesophyll conductance and CO2 fixation in the chloroplast due to the lower carboxylation efficiency. On the other hand, although there is reduction in gs after 30 days of exposure to stress, there is an increase in conductance values in seedlings under 30% SH + 40% WHC and 0% SH + 40% WHC, indicating that as a survival mechanism, plants regulate the stomatal openings over time so that there is less water loss without harming the entry of CO2. From 45 days onwards, gs and A values increased, suggesting that the species is adjusting to this cultivation condition. Therefore, it is emphasized that the species presents adjustment strategies that ensure its phenotypic plasticity and potential for ecological resilience, especially after 45 days.
Another factor that may have contributed to the 0% SH environment to accentuate the reduction in photosynthetic metabolism, especially in seedlings under 40% WHC, was the increase in leaf temperature that occurred in this environment. Studies have reported that high temperatures directly impact the photosynthetic enzymes involved in carbon assimilation, being harmful to the proper functioning of photosynthetic processes [23]. As shading increases, reduction in leaf temperature is observed, demonstrating that shading mitigated the negative effects caused by the increase in temperature.
The lower water availability, represented here by 40% WHC, combined with the highest light intensity (0% SH) reduced the chlorophyll index and photochemical activities in the photosystem II of C. fissilis seedlings in all evaluation periods. Excessive light energy induces photoinhibition of photosystem II (PSII) and causes photodamage to the photosynthetic system [24], also associated with reduction in ATP as a result of the lower H+ production due to water deficit [25], reducing photosynthetic carbon gain. The full sun condition associated with lower water availability caused decrease in these factors, indicating that there was a decline in the PSII function and that photosynthetic units and electron transfer processes linked to the membrane were affected under this stress [26].
Under the same cultivation conditions mentioned above, seedlings presented lower leaf area values, which, although considered a strategy found by plants to minimize water loss through evapotranspiration, reduces the photosynthetically active area. Furthermore, this response is associated with reduction in turgor, since under the same cultivation condition, lower WHC was also observed.
Conversely, the full sun environment (0% SH) combined with high water availability (100% WHC) provided C. fissilis seedlings with higher A, A/Ci, E and RWC values in a short period of exposure to stress, reinforcing that if the species is cultivated at this shading level, it is necessary to increase the water regime to meet energy costs and alleviate the negative effect of light stress.
The fact that the 0% SH + 100% WHC condition provided higher A/Ci values can be justified by studies that report that plants grown under high irradiance present increase in Rubisco content and activity to optimally use more absorbed light energy [27,28,29].
However, as the exposure time to 0% SH + 100% WHC increased, there was a decrease in A, gs, A/Ci and E values up to 45 days of cultivation under these conditions. Between 45 and 60 days, the values of these variables increased, suggesting that the 0% SH condition associated with greater water availability was initially beneficial for seedlings, but excess light caused stress; however, seedlings adapted to this condition over time, recovering their gas exchange values.
What may have helped with this response was the fact that under this cultivation condition (0% SH + 100% WHC), seedlings presented higher WUE values. In general, WUE was higher at 38 days, coinciding with the period in which gas exchange decreased for C. fissilis seedlings grown in full sun with high water availability, WUE is an important mechanism of physiological adaptation [30], and the increase in this factor led to better use of available resources. WUE reflects the proportion between the water used in plant metabolism and the water lost through transpiration [31]. The species' lower sensitivity to exposure to full sun in the first evaluation can be attributed to the short exposure time (15 days), which may have been insufficient to cause stress responses, but over time, the photosynthetic metabolism reduced, especially under 40% WHC. However, it can also be attributed to the physiological and morphological properties of its leaves, which are well adapted to greater irradiance. Pioneer or early secondary species tend to have greater capacity to increase water use efficiency under more energetic luminosity [32].
The increse in the chlorophyll index in shaded seedlings, especially under 70% SH, regardless of water regime, can be considered a compensatory mechanism. This is because the increase in chlorophyll content per area unit increases the absorption of photosynthetically active radiation to increase the efficiency of CO2 assimilation [32,33,34,35].
Most plants under shade invest more in the synthesis and maintenance of structures that capture light than those in full sun conditions [36]. Plants with greater tolerance to reduced light are plastic, as they can control light capture variables, such as leaf size and chlorophyll content. In this sense, higher leaf area values for C. fissilis in 70% shading were also observed. Increasing leaf area is a strategy of the species to optimize the use of light, increasing the surface for capturing photons to maintain photosynthetic processes [37,38].
The increase in the shading level resulted in higher of Fv/F0 and Fv/Fm values in C. fissilis seedlings, regardless of water regimes, which may be related to greater potential for use and optimization of available light energy [39]. Furthermore, under the 0% SH condition, there was higher leaf temperature, which increase can cause physiological disturbances in plants, for example, photosystem I and photosystem II are negatively affected at high temperatures, decreasing Fv/ F0 and Fv/Fm values [40]. However, shading can improve the light energy use rate [41] and in this case can mitigate the damage caused to the photosynthetic apparatus, providing conditions that would reduce the leaf temperature of seedlings. Plant plasticity helps to improve the use of photosynthetic energy under intense light, and C. fissilis seedlings grown in 0% SH showed higher PPI for Fv/Fm, as a mechanism for adjusting the photosynthetic apparatus in response to unfavorable conditions.
C. fissilis seedlings that presented the highest total dry mass values were those grown in moderate shading (30% shading) combined with 100% WHC. This condition provided seedlings with adjustments in gas exchange throughout the evaluation periods for carrying out the photosynthetic process such as the regulation of the ideal stomatal opening, providing the entry of CO2, as well as higher transpiration levels and Rubisco carboxylation activity.
C. fissilis is considered a species with high light demand for growth [42]. Even though this species is classified as pioneer or initial secondary, excess light under full sun conditions, after 15 days of exposure, caused reduction in the photosynthetic metabolism of seedlings, which reflected in less investment in seedling growth, an accentuated response under 40% WHC. Although plants increase A values over evaluation times, reductions that occurred before seedlings adapted to the condition caused plants to invest their energy in survival to the detriment of growth. In the present study, it was found that the best seedling performance occurred under moderate shading combined with greater water availability.
Moderate shading associated with greater water availability resulted in greater production of photoassimilates for investment in greater dry mass in C. fissilis. [7] found that the survival and growth of C. fissilis were associated with adequate water availability (70% WHC and 100% WHC). This is because low water availability directly affects photosynthesis and consequently organic matter accumulation [43], as observed in this study with C. fissilis.
C. fissilis seedlings showed higher phenotypic plasticity rates for A, demonstrating that under different light conditions, this species adjusts in order to maintain its photosynthetic activity and survival. Some studies have reported that leaf characteristics are highly plastic, although phenotypic plasticity measures are strongly correlated with the context and are not comparable between different studies, gradients and species [44,45]. In the full sun environment, C. fissilis showed greater morphological adjustments in the leaf area and physiological adjustments in Fv/Fm, reinforcing that this cultivation condition was stressful, especially when associated with 40% WHC, a result confirmed by the highest PPI values.
Seedlings showed few adjustments in the 70% shading condition, which leads us to believe that for the factors evaluated under this environmental condition, plants did not need many adjustments to guarantee their survival. However, 70% shading did not correspond to the condition in which seedlings best expressed their potential.
Based on our results, it is possible to verify that the species presents plasticity to contrasting light environments, but that in the seedling phase, its best morphophysiological performance is under moderate shading, reinforcing its classification as initial secondary. On the other hand, if the cultivation is in full sun, for example in open areas, the ideal would be to use water regime with greater availability to supply its photosynthetic metabolism, while if implemented in areas with low water status or subject to temporary drought, shaded cultivation is a strategy to mitigate stressful effects.
4. Material and Methods
4.1. Experiment Location and Seedling Production
The experiment was conducted at the Federal University of Grande Dourados - UFGD, implemented in different light conditions of the Faculty of Agricultural Sciences (22º11'43.7"S and 54º56'08.5"W, 452 m a.s.l.), in the city of Dourados – MS, Brazil.
C. fissilis seedlings were produced from ripe fruits collected in a remaining Cerrado area (Authorization for Access and Shipment of Genetic Heritage Component Sample No. 010220/2015-1 – CNPq/CGEN/MMA), in the state of Mato Grosso do Sul. The collection of fruits and seeds was carried out during periods of dispersal, and processing was manually performed, so that seeds were extracted and selected.
Sowing took place on 08/02/2021 in tubes with volume of 280 cm3 filled with substrate consisting of Distroferric Red Latosol + commercial substrate (Bioplant®) (1:1, v/v). Tubes were placed on benches in a nursery environment protected with black nylon screen (Sombrite®) with 30% shading. Full emergence occurred 90 days after sowing (DAS).
Transplanting into pots was carried out when seedlings reached average height of 8.0 cm. Seedlings were grown in 8 kg plastic pots filled with substrate composed of Distroferric Red Latosol + coarse sand (3:1, v/v), with two plants each.
4.2. Shading Level and Water Regimes
After transplanting, seedlings were submitted to acclimatization for 45 days at each shading level – SH (0, 30 and 70%). After this period, seedlings were submitted to three water regimes based on the water holding capacity (WHC) in the substrate, constituting nine cultivation conditions: T1 – 0% SH + 40% WHC; T2 – 0% SH + 70% WHC; T3 – 0% SH + 100% WHC; T4 – 30% SH + 40% WHC; T5 – 30% SH + 70% WHC; T6 – 30% SH and 100% WHC; T7 – 70% SH + 40% WHC; T8 – 70% SH +70% WHC; T9 – 70% SH + 100% WHC.
In the different light conditions, a structure with top and side cover made of transparent plastic was inserted to protect against possible precipitations in the experiment. Shading levels of 30 and 70% were obtained through the use of black nylon screens (Sombrite®), in which photosynthetically active radiation and leaf temperature values were recorded on a portable photosynthesis meter LCIPro - SD ADC BioScientific Ltd. (IRGA) and can be found in Table 3.
The water holding capacity of the substrate was determined according to methodology of [46], in which the 100% WHC was obtained through the water content retained after draining, and the others were calculated using the simple rule of three as a function of weight. Pots were weighed on a precision scale (0.001 g) (Figure 6) and individual irrigation was carried out daily, adding enough water to reach the pre-established weight for each cultivation condition. The substrate of each pot was covered with plastic to prevent water evaporation from the soil.
4.3. Experimental Design
The experimental design used was completely randomized in a factorial scheme with 9 cultivation conditions (WHC and shading combination) x 4 evaluation times, with four replicates. The experimental unit consisted of one pot with two seedlings each.
4.4. Evaluations
Gas exchange, chlorophyll a fluorescence and chlorophyll index characteristics were evaluated at 15, 30, 45 and 60 days of cultivation, while those of relative water content and growth were evaluated at 15 and 60 days.
Gas exchange, chlorophyll a fluorescence and chlorophyll index determination was carried out between 8 and 11 am, on the same fully expanded and previously marked leaves, so that all measurements were always carried out on the same plants.
a) Gas exchange: the net photosynthetic rate - A (µmol CO2 m-2 s-1), stomatal conductance - gs (mol H2O m-2 s-1), internal CO2 concentration - Ci (mol CO2 m-2 air-1) and transpiration rate - E (mmol H2O m-2 s-1) were quantified with portable photosynthesis meter (LCIPro - SD ADC BioScientific Ltd). From these data, the Rubisco carboxylation efficiencies - A/Ci (µmol mol CO2 m-2 s-1) and water use efficiency WUE - A/E (µmol CO2/mmol H2O) were calculated.
b) Chlorophyll a fluorescence: the potential quantum efficiency of FSII - Fv/Fm and effective quantum yield of photochemical energy conversion in FSII - Fv/F0 were determined using portable fluorometer model OS-30p (Opti-Sciences Chlorophyll Fluorometer, Hudson, USA). Leaves were submitted to a period of 30 minutes of dark adaptation with the aid of adapter clips, so that all reaction centers in this leaf region reached the “open” condition, that is, complete oxidation of the electron transport photosynthetic system [47].
c) Chlorophyll index - CI: obtained using portable chlorophyll meter (model SPAD-502, Minolta Camera Co. Ltd.) and carried out on the median limbs of the same leaves as the gas exchange and fluorescence assessments.
d) Relative water content of leaves - RWC: calculated using the methodology of [48]. Growth: root length (RL) was measured with graduated ruler and leaf area - LA (cm2) was evaluated using the ImageJ software. The materials were placed in an oven with forced air circulation at 60 ± 5oC for 72 h, and weighed on decimal precision scale (0.0001 g).
f) Phenotypic plasticity index (PPI): the PPI for A, Fv/Fm, LA and TDM was calculated from data obtained after 60 days of cultivation according to methodology proposed by [49]. PPI was calculated considering the highest and lowest values of these characteristics, calculating between the highest and lowest value between water regimes for each shading level.
4.5. Data Analysis
Data were submitted to analysis of variance (ANOVA) and when significant by the F test (p ≤ 0.05), means were compared by the Scott-Knott test (p ≤ 0.05) for the different cultivation conditions. Data analyzed at 15 and 60 days were compared by the F test (p ≤ 0.05) and at 15, 30, 45 and 60 days by regression analysis (p ≤ 0.05), using the SISVAR 5.3 statistical software [53]. PPI results were presented in a descriptive manner, without applying statistical analysis.
5. Conclusions
The availability of 40% water holding capacity was detrimental to gas exchange, regardless of exposure time and consequently affected the growth of C. fissilis, especially when combined with full sun environment.
Seedlings grown in 70% shade presented strategies such as adaptation mechanisms to survive, demonstrating that this species has physiological plasticity, and this condition helped to mitigate the stressful effect of water deficit in C. fissilis.
The condition that provided better gas exchange performance and greater total dry mass accumulation for C. fissilis seedlings was 30% shading combined with 100% WHC. C. fissilis seedlings showed physiological plasticity and resilience to survive under different water and light conditions.
Author Contributions
Conceptualization, S.P.Q.S. and C.C.S.; methodology, J.M.S., J.A.L., T.D., A.S.D.; formal analysis, J.M.S., A.S.D. and R.S.B.; investigation, J.M.S., J.A.L., R.S.B. and T.D. resources, S.d.P.Q.S.; data curation, C.C.S. and S.P.Q.S.; writing—original draft preparation, J.M.S.; writing—review and editing, C.C.S. and S.P.Q.S.; supervision, C.C.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding
Institutional Review Board Statement
Not aplicable
Data Availability Statement
The data presented in this study are available in the graphs and tables provided in the manuscript.
Acknowledgments
The National Council for Scientific and Technological Development (CNPq) and Coordination for the Improvement of Higher Education Personnel (CAPES), for the granting of scholarships, and the Foundation for Support to the Development of Education, Science and Technology of the State of Mato Grosso do Sul (FUNDECT) for financial support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lima, V.P.; Lima, R.A.F. de; Joner, F.; Siddique, I.; Raes, N.; Steege, H.T. Climate change threatens native potential agroforestry plant species in Brazil. Sci. Rep. 2022, 12, 2267. [Google Scholar] [CrossRef] [PubMed]
- Vourlitis, G.L.; Pinto Jr., O. B.; Dalmagro, H.J.; Arruda, P.E.Z.; Lobo, F.A.; Nogueira J.S. Tree growth responses to climate variation in upland and seasonally flooded forests and woodlands of the Cerrado-Pantanal transition of Brazil. Forest. Ecol. Manag. 2022, 505, 119917. [Google Scholar] [CrossRef]
- Wei, C.; Wang, Q.; Han, H.; Gan, X. Low soil moisture improved shading tolerance by regulating leaf functional traits in Tetracentron sinense Oliv. Seedlings. Glob. Ecol. Conserv. 2023, 48, e02757. [Google Scholar] [CrossRef]
- Salmona, Y.B.; Matricardi, E.A.T.; Skole, D.L.; Silva, J.F.A.; Coelho Filho, O.A.; Pedlowski, M.A.; Sampaio, J.M.; Castrillón, L.C.R.; Brandão, R.A.; Silva, A.L.; et al. A Worrying Future for River Flows in the Brazilian Cerrado Provoked by Land Use and Climate Changes. Sustainability 2023, 15, 4251. [Google Scholar] [CrossRef]
- Paradiso, R.; Proietti, S. Light-Quality Manipulation to Control Plant Growth and Photomorphogenesis in Greenhouse Horticulture: The State of the Art and the Opportunities of Modern LED Systems. J. Plant Growth Regul. 2022, 41, 742–780. [Google Scholar] [CrossRef]
- Cerqueira, W.M.; Scalon, S.P.Q.; Santos, C.C.; Santiago, E.F.; Almeida, J.L.C.S.; Figueiredo, V.M.A.; Linné, J.A.; Silverio, J.M. Ecophysiological mechanisms and growth of Inga vera Willd. under different water and light availability. Braz. J. Biol. 2023, 83, e275378. [Google Scholar] [CrossRef]
- Griebeler, A.M.; Araujo, M.M.; Barbosa, F.M.; Kettenhuber, P.L.; Nhantumbo, L.S.; Berghetti, A.L.P.; Denardi, L. Morphophysiological responses of forest seedling species subjected to different water regimes. J. For. Res. 2021, 32, 2099–2110. [Google Scholar] [CrossRef]
- Parkash, V.; Singh, S. A Review on potential plant-based water stress indicators for vegetable crops. Sustainability 2020, 12, 3945. [Google Scholar] [CrossRef]
- Bottero, A.; Forrester, D.I.; Cailleret, M.; Kohnle, U.; Gessler, A.; Michel, D.; Bose, A.K.; Bauhus, J.; Bugmann, H.; Cuntz, M; et al. Growth resistance and resilience of mixed silver fir and Norway spruce forests in central Europe: Contrasting responses to mild and severe droughts. Glob. Change Biol. 2021, 27, 4403–4419. [Google Scholar] [CrossRef]
- Croce, J.; Badano, E.I.; Trigo, C.B.; Martinez-Galvez, F.; Tálamo, A. Experimental approaches to select tree species for forest restoration: effects of light, water availability and interspecific competition in degraded areas. J. For. Res. 2022, v. 33, 1197–1207. [Google Scholar] [CrossRef]
- Pereira, G.A.; Barbosa, A.C.M.C.; Torbenson, M.C.A.; Stahle, D.W.; Granato-Souza, D.; Santos, R. M.; Barbosa, J.P.D. The Climate Response of Cedrela fissilis Annual Ring Width in the Rio São Francisco Basin, Brazil. Tree-Ring Res. 2018, 74, 162–171. [Google Scholar] [CrossRef]
- Prado, M.C.N.; Giuliani, G.K.F.; Ghiotto, T.C.; Carmo, J.B.; Guereiro, J.C.; Prado, E. P.; Pogetto, M.H.F.A.S.; Masson, M.V.; Tavares, W.S.; Wilcken, C.F.; et al. Detection and estimation of Mastigimas anjosi (Hemiptera: Calophyidae) populations on Cedrela fissilis trees. R. Soc. Open. Sci. 2022, 9, 211340. [Google Scholar] [CrossRef] [PubMed]
- Krainovic, P.M.; Resende, A.F.; Amazonas, N.T.; Almeida, C.T.; Almeida, D.R.A.; Silva, C.C.; Andrade, H.S.F.; Rodrigues, R.R.; Brancalion, P.H.S. Potential native timber production in tropical forest restoration plantations. Perspect. Ecol. Conserv. 2023, 21, 294–301. [Google Scholar] [CrossRef]
- Santos, T.R.S.; Santos, J.A.S.; Pereira, E.G.; Garcia, Q.S. Revegetation of an area impacted by iron ore tailings: evaluating fertilization alternatives in native pioneer and secondary trees. Environ. Sci. Pollut. Res. Int. 2023, 30, 3760–3773. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, L.O.; Dresch, D.M.; Scalon, L.; Scalon, S.P.Q. Ecophysiological strategies of Cedrela fissilis Vell. seedlings under conditions of flooding and light availability. J. Sustain. For. 2021, 41, 783–798. [Google Scholar] [CrossRef]
- Rodriguez, D.R.O.; Sánchez-Salguero, R.; Hevia, A.; Bovi, R.C.; Ferreira, M. J.; Speer, J.H.; Tomazello-Filho, M. Does climate change alter the nutrient trends of Cedrela fissilis Vell. trees in the southern Brazilian Amazon? Ecol. Process. 2023, 12, 58. [Google Scholar] [CrossRef]
- Leonel, L.V.; Reis, F.O.; Figueiredo, F.A.M.M.A.; Ferraz, T.M.; Maia Júnior, S.O.; Silva, P.C.; Andrade, J.R. Light intensity and hydrogel soil amendment differentially affect growth and photosynthesis of successional tree species. J. For. Res. 2023, 34, 257–268 ttps://doiorg/101007/s11676. [Google Scholar] [CrossRef]
- Bartieres, E.M.M.; Dresch, D.M.; Reis, L.C.; Pereira, Z.V.; Mussury, R.M.; Scalon, S.P.Q. Shading minimizes the effects of water deficit in Campomanesia xanthocarpa (Mart.) O. Berg seedlings. Braz. J. Biol. 2023, 83, e244718. [Google Scholar] [CrossRef]
- Mukherjee, A.; Dwivedi, S.; Bhagavatuba, L.; Datta, S. Integration of light and ABA signaling pathways to combat drought stress in plants. Plant Cell Rep. 2023, 42, 829–841. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought Stress Impacts on Plants and Different Approaches to Alleviate Its Adverse Effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef]
- Puértolas, J.; Dodd, I.C. Evaluating soil evaporation and transpiration responses to alternate partial rootzone drying to minimise water losses. Plant Soil 2022, 480, 473–489. [Google Scholar] [CrossRef]
- Zhu, L.; Li, H.; Thorpe, M.R.; Hocart, C.H.; Song, X. Stomatal and mesophyll conductance are dominant limitations to photosynthesis in response to heat stress during severe drought in a temperate and a tropical tree species. Trees 2021, 35, 1613–1626. [Google Scholar] [CrossRef]
- Moore, C.E.; Meacham-Hensold, K.; Lemonnier, P.; Stattery, R.A.; Benjamin, C.; Bernacchi, C.J.; Lawson, T.; Cavanagh, A. P. The effect of increasing temperature on crop photosynthesis: from enzymes to ecosystems. J. Exp. Bot. 2021, 72, 2822–2844. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xie, S.; Yan, S.; Xu, W.; Chen, J. Light energy partitioning and photoprotection from excess light energy in shade-tolerant plant Amorphophallus xiei under steady-state and fluctuating high light. Acta Physiol. Plant 2021, 43, 125. [Google Scholar] [CrossRef]
- Razi, K.; Muneer, S. Drought stress-induced physiological mechanisms, signaling pathways and molecular response of chloroplasts in common vegetable crops. Crit. Rev. Biotechnol. 2021, 41, 669–691. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ma, J.; Wang, Y.; Xu, S.; Jiang, L.; Zhang, L.; Hou, W. Effects of exogenous NO on the growth and photosynthetic fluorescence characteristics of ryegrass seedlings under B[a]P stress. Acta Bot. Croat. 2023, 82, 71–79. [Google Scholar] [CrossRef]
- Eckert, D.; Jensen, A.M.; Gu, L. The maximum carboxylation rate of Rubisco affects CO2 refixation in temperate broadleaved forest trees. Plant Physiol. Biochem. 2020, 155, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Cummins, P. L. The coevolution of rubisco, photorespiration, and carbon concentrating mechanisms in higher plants. Front. Plant. Sci. 2021, 12, 662425. [Google Scholar] [CrossRef]
- Ye, X.; Gao, Z.; Xu, K.; Li, B.; Ren, T.; Li, X.; Cong, R.; Lu, Z.; Cakmak, I.; Lu, J. Photosynthetic plasticity aggravates the susceptibility of magnesium-deficient leaf to high light in rapeseed plants: the importance of Rubisco and mesophyll conductance. Plant J. 2023, 117, 483–497. [Google Scholar] [CrossRef]
- Zhao, W.; Liu, L.; Shen, Q.; Yang, J.; Han, X.; Tian, F.; Wu, J. Effects of water stress on photosynthesis, yield, and water use efficiency in winter wheat. Water 2020, 12, 2127. [Google Scholar] [CrossRef]
- Fonseca, C.L.S.; Silva, D. M.; Gasparini, T.A.S.; Cuzzuol, G.R.F. Photosynthesis of plants of shade-tolerant ecotype of Paubrasilia echinata are more tolerance to drought than the sun-tolerant ecotype. Plant Stress 2023, 8, 100157. [Google Scholar] [CrossRef]
- Angadi, S.V. Umesh, M.R.; Begna, S.; Gowda, P. Light interception, agronomic performance, and nutritive quality of annual forage legumes as affected by shade. Field Crops Res. 2022, 275, 108358. [Google Scholar] [CrossRef]
- Genesio, L.; Bassi, R.; Miglietta, F. Plants with less chlorophyll: A global change perspective. Glo. Change Biol. 2020, 27, 959–967. [Google Scholar] [CrossRef]
- Slattery, R.A.; Ort, D.R. Perspectives on improving light distribution and light use efficiency in crop canopies. Plant Physiol 2021, 185, 34–48. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhang, J.; Zhang, Z.; Sui, Y.; Zhu, H.; Shi, Y.; Sui, X.; Li, Y.; Jiang, X. Insights into the changes in leaf functional traits of Aralia elata grown under different shading treatments. Plant Growth Regul. 2023, 101, 679–691. [Google Scholar] [CrossRef]
- Vetoshkina, D.; Balashov, N.; Ivanov, B.; Ashikhmin, A.; Borisova-Mubarakshina, M. Light harvesting regulation: A versatile network of key components operating under various stress conditions in higher plants. Plant Physiol. Biochem. 2023, 194, 576–588. [Google Scholar] [CrossRef]
- Xu, R.; Wang, L.; Zhang, J.; Zhou, J.; Cheng, S.; Tigabu, M.; Ma, X.; Wu, P.; Li, M. Growth Rate and Leaf Functional Traits of Four Broad-Leaved Species Underplanted in Chinese Fir Plantations with Different Tree Density Levels. Forests 2022, 13, 308. [Google Scholar] [CrossRef]
- Luo, D.; Huang, G.; Zhang, Q.; Zhou, G.; Peng, S.; Li, Y. Plasticity of mesophyll cell density and cell wall thickness and composition play a pivotal role in regulating plant growth and photosynthesis under shading in rapeseed. Ann. Bot. 2023, 132, 963–978. [Google Scholar] [CrossRef]
- Yu, X.F.; Ming, X.Y.; Xiong, M.; Zhang, C.; Yue, L.J.; Yang, L.; Fan, C.Y. Partial shade improved the photosynthetic capacity and polysaccharide accumulation of the medicinal plant Bletilla ochracea Schltr. Photosynthetica 2022, 60, 168–178 1032615/ps2021064. [Google Scholar] [CrossRef]
- Yan, Z.; Ma, T.; Guo, S.; Liu, R.; Li, M. Leaf anatomy, photosynthesis and chlorophyll fluorescence of lettuce as influenced by arbuscular mycorrhizal fungi under high temperature stress. Sci. Hortic. 2021, 280, 109933. [Google Scholar] [CrossRef]
- Roeber, V.M.; Bajaj, I.; Rohde, M.; Schmulling, T.; Cortleven, A. Light acts as a stressor and influences abiotic and biotic stress responses in plants. Plant Cell Environ. 2020, 44, 645–664. [Google Scholar] [CrossRef]
- Sanches, M.C.; Marzinek, J.; Bragiola, N.G.; Nascimento, A.R.T. Morpho-physiological responses in Cedrela fissilis Vell. submitted to changes in natural light conditions: implications for biomass accumulation. Trees 2017, 31, 215–227. [Google Scholar] [CrossRef]
- Xingyang, S.; Guangsheng, Z.; Qijing, H.; Huailin, Z. Stomatal limitations to photosynthesis and their critical Water conditions in different growth stages of maize under water stress. Agric. Water Manag. 2020, 241, 106330. [Google Scholar] [CrossRef]
- Granata, M.U.; Bracco, F.; Catoni, R. C. Phenotypic plasticity of two invasive alien plant species inside a deciduous forest in a strict nature reserve in Italy. J. Sustain. For. 2019, 39, 346–364. [Google Scholar] [CrossRef]
- Olguin, F.Y.; Moretti, A.P.; Pinazo, M.; Gortari, F.; Bahima, J.V.; Graciano, C. Morphological and physiological plasticity in seedlings of Araucaria angustifolia and Cabralea canjerana is related to plant establishment performance in the rainforest. For. Ecol. Manag. 2020, 460, 117867. [Google Scholar] [CrossRef]
- Souza, C.C.; Oliveira, F.A.; Silva, I.F.; Amorim Neto, M.S. Avaliação de métodos de determinação de água disponível e manejo da irrigação em terra roxa sob cultivo de algodoeiro herbáceo. Rev. Bras. Eng. Agric. 2000, 4, 338–342. [Google Scholar] [CrossRef]
- Bolhàr-Nordenkampf, H.R.; Long, S.P.; Baker, N.R. Fluorescência da clorofila como sonda da competência fotossintética de folhas no campo: uma revisão do instrumento atual. Funct. Ecol. 1989, 3, 497–514. [Google Scholar] [CrossRef]
- Slavick, B. Methods of Studying Plant Water Relations. Springer- Verlag, New York, 1979. ISBN 978-3-642-65834-1.
- Valladares, F.; Sanchez-Gomez, D.; Zavala, M.A. Quantitative estimation of phenotypic plasticity: Bridging the gap between the evolutionary concept and its ecological applications. J. Ecol. 2006, 94, 1103–1116. [Google Scholar] [CrossRef]
Figure 1.
Visual aspects of Cedrela fissilis Vell. seedlings of cultivation under different water and light availability.
Figure 1.
Visual aspects of Cedrela fissilis Vell. seedlings of cultivation under different water and light availability.

Figure 2.
Net photosynthetic rate - A (A), stomatal conductance - gs (B), Rubisco carboxylation efficiencies - A/Ci (C) and transpiration rate - E (D) of Cedrela fissilis Vell. seedlings evaluated at 15, 30, 45 and 60 days of cultivation under different water and light availability. T1 – 0% SH + 40% WHC; T2 – 0% SH + 70% WHC; T3 – 0% SH + 100% WHC; T4 – 30% SH + 40% WHC; T5 – 30% SH + 70% WHC; T6 – 30% SH + 100% WHC; T7 – 70% SH + 40% WHC; T8 – 70% SH +70% WHC; T9 – 70% SH + 100% WHC. Equal letters between markers of different colors in each evaluation period do not differ statistically according to the Scott-Knott test (p ≤ 0.05) for cultivation conditions.
Figure 2.
Net photosynthetic rate - A (A), stomatal conductance - gs (B), Rubisco carboxylation efficiencies - A/Ci (C) and transpiration rate - E (D) of Cedrela fissilis Vell. seedlings evaluated at 15, 30, 45 and 60 days of cultivation under different water and light availability. T1 – 0% SH + 40% WHC; T2 – 0% SH + 70% WHC; T3 – 0% SH + 100% WHC; T4 – 30% SH + 40% WHC; T5 – 30% SH + 70% WHC; T6 – 30% SH + 100% WHC; T7 – 70% SH + 40% WHC; T8 – 70% SH +70% WHC; T9 – 70% SH + 100% WHC. Equal letters between markers of different colors in each evaluation period do not differ statistically according to the Scott-Knott test (p ≤ 0.05) for cultivation conditions.
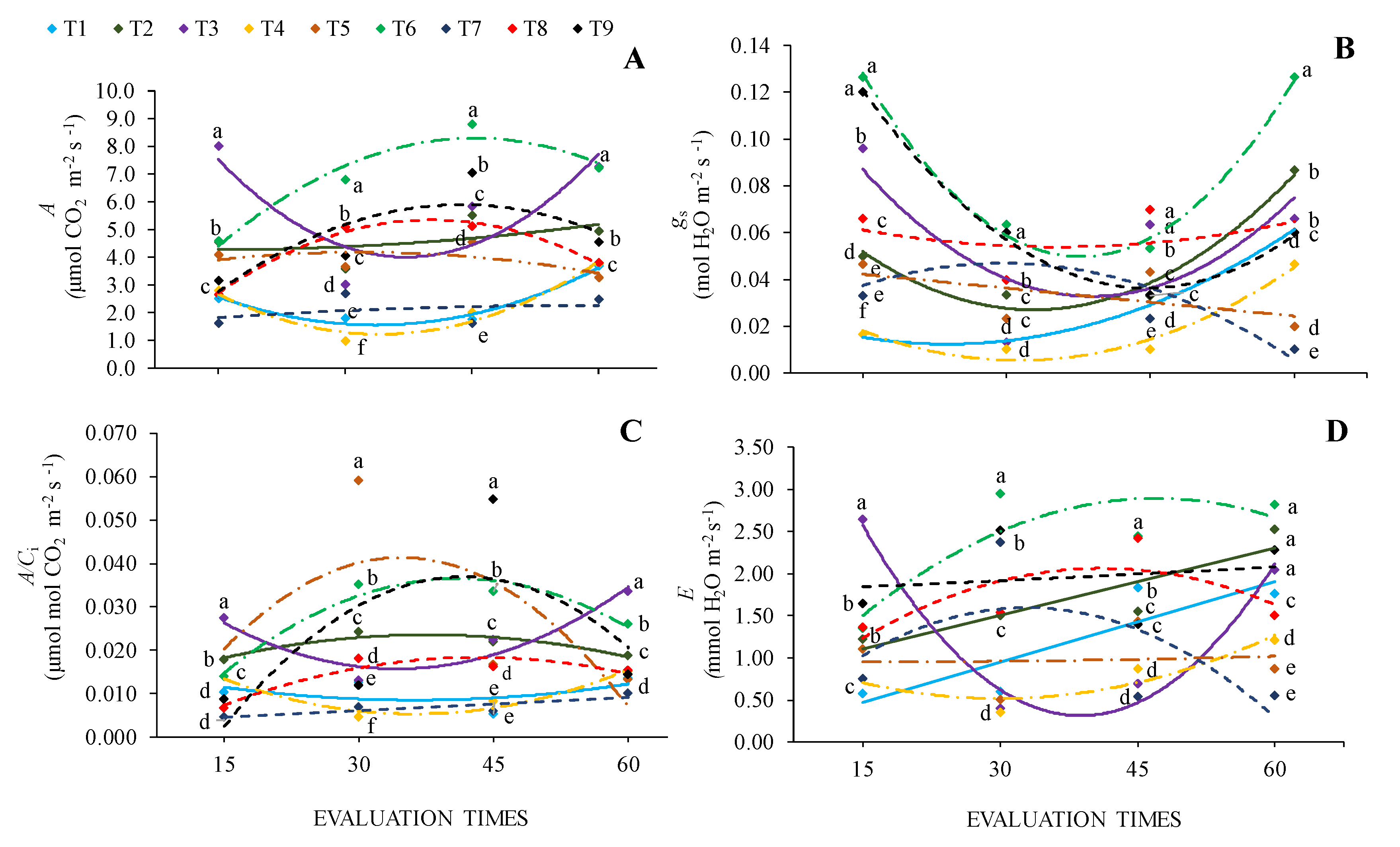
Figure 3.
Water use efficiency - WUE (A, B) and chlorophyll index -CI (C) of Cedrela fissilis Vell. under different availability of water and light evaluated at 15, 30, 45 and 60 days of cultivation. T1 – 0% SH + 40% WHC; T2 – 0% SH + 70% WHC; T3 – 0% SH + 100% WHC; T4 – 30% SH + 40% WHC; T5 – 30% SH + 70% WHC; T6 – 30% SH + 100% WHC; T7 – 70% SH + 40% WHC; T8 – 70% SH +70% WHC; T9 – 70% SH + 100% WHC. Equal letters between markers of different colors in each evaluation period do not differ statistically according to the Scott-Knott (p ≤ 0.05) test for cultivation conditions.
Figure 3.
Water use efficiency - WUE (A, B) and chlorophyll index -CI (C) of Cedrela fissilis Vell. under different availability of water and light evaluated at 15, 30, 45 and 60 days of cultivation. T1 – 0% SH + 40% WHC; T2 – 0% SH + 70% WHC; T3 – 0% SH + 100% WHC; T4 – 30% SH + 40% WHC; T5 – 30% SH + 70% WHC; T6 – 30% SH + 100% WHC; T7 – 70% SH + 40% WHC; T8 – 70% SH +70% WHC; T9 – 70% SH + 100% WHC. Equal letters between markers of different colors in each evaluation period do not differ statistically according to the Scott-Knott (p ≤ 0.05) test for cultivation conditions.
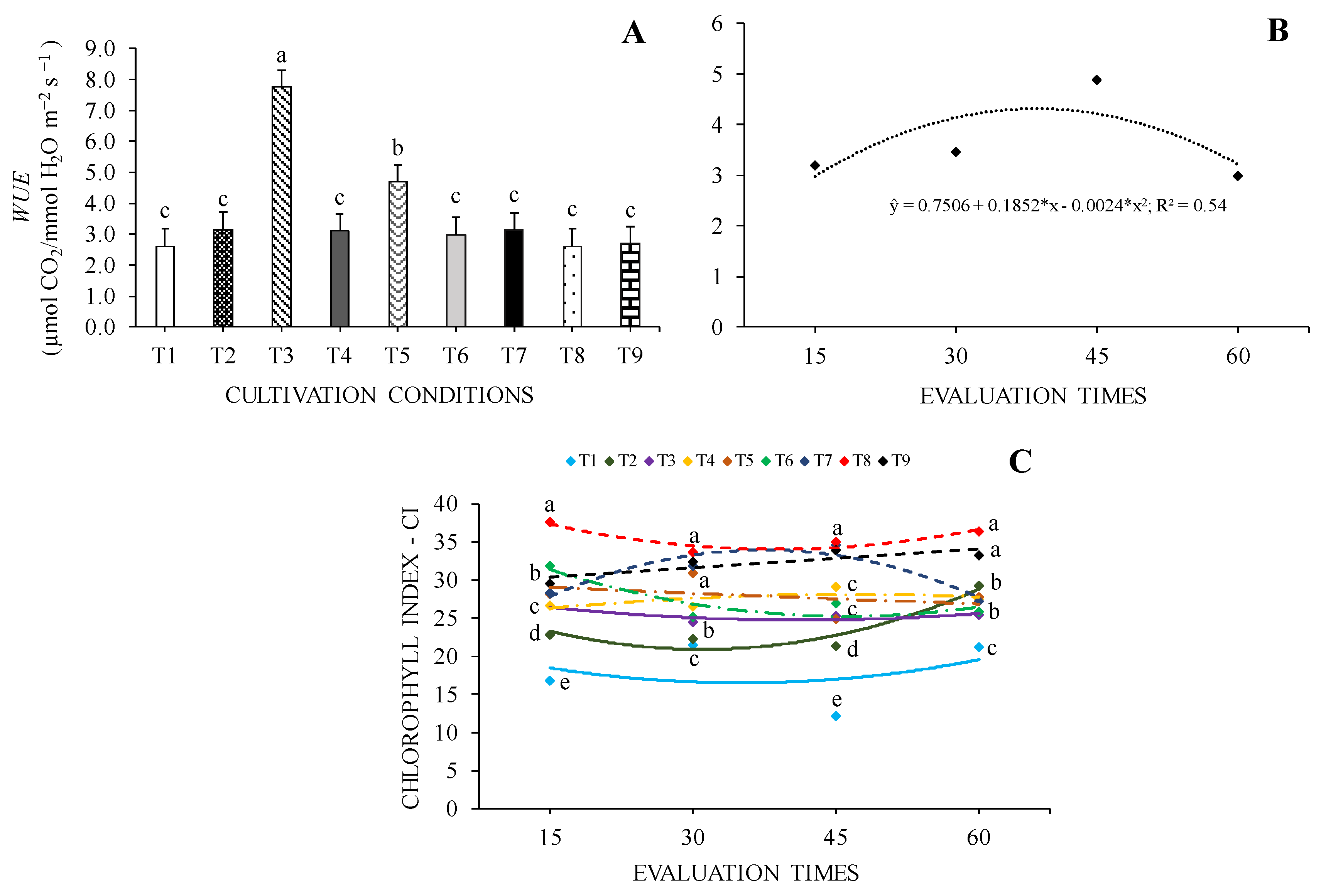
Figure 4.
Effective quantum yield of photochemical energy conversion in FSII - Fv/F0 (A, B) and potential quantum efficiency of FSII - Fv/Fm (C, D) of Cedrela fissilis Vell. under different availability of water and light evaluated at 15, 30, 45 and 60 days of cultivation. T1 – 0% SH + 40% WHC; T2 – 0% SH + 70% WHC; T3 – 0% SH + 100% WHC; T4 – 30% SH + 40% WHC; T5 – 30% SH + 70% WHC; T6 – 30% SH + 100% WHC; T7 – 70% SH + 40% WHC; T8 – 70% SH +70% WHC; T9 – 70% SH + 100% WHC. Equal letters between markers of different colors in each evaluation period do not differ statistically according to the Scott-Knott test (p ≤ 0.05) for cultivation conditions.
Figure 4.
Effective quantum yield of photochemical energy conversion in FSII - Fv/F0 (A, B) and potential quantum efficiency of FSII - Fv/Fm (C, D) of Cedrela fissilis Vell. under different availability of water and light evaluated at 15, 30, 45 and 60 days of cultivation. T1 – 0% SH + 40% WHC; T2 – 0% SH + 70% WHC; T3 – 0% SH + 100% WHC; T4 – 30% SH + 40% WHC; T5 – 30% SH + 70% WHC; T6 – 30% SH + 100% WHC; T7 – 70% SH + 40% WHC; T8 – 70% SH +70% WHC; T9 – 70% SH + 100% WHC. Equal letters between markers of different colors in each evaluation period do not differ statistically according to the Scott-Knott test (p ≤ 0.05) for cultivation conditions.
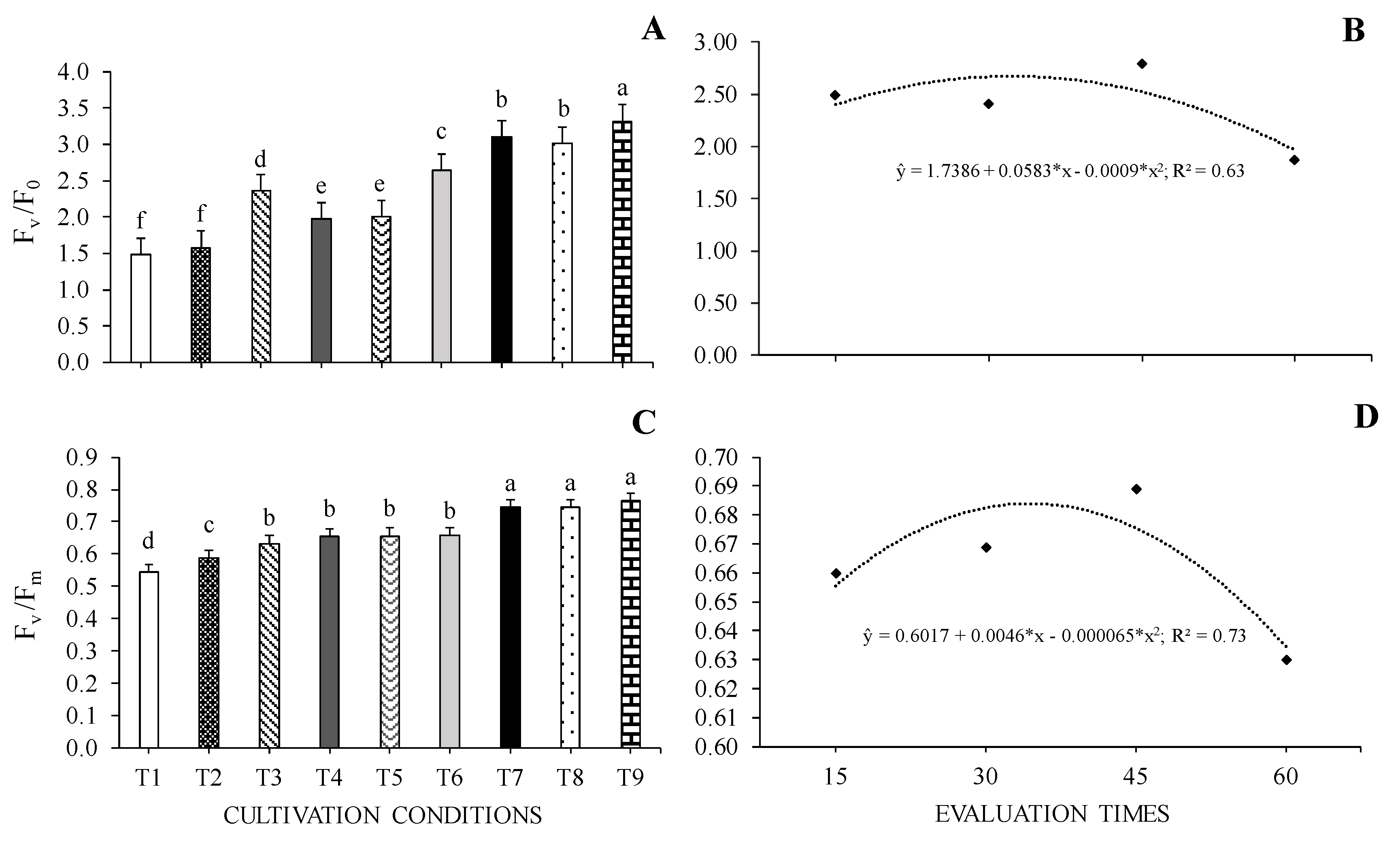
Figure 5.
Relative water content of leaves – RWC (A), leaf area - LA (B), root length - RL and total dry mass – TDM of Cedrela fissilis Vell. under different availability of water and light evaluated at 15 and 60 days of cultivation. T1 – 0% SH + 40% WHC; T2 – 0% SH + 70% WHC; T3 – 0% SH + 100% WHC; T4 – 30% SH + 40% WHC; T5 – 30% SH + 70% WHC; T6 – 30% SH + 100% WHC; T7 – 70% SH + 40% WHC; T8 – 70% SH +70% WHC; T9 – 70% SH + 100% WHC. Lowercase letters compare the effect of cultivation conditions at each evaluation time using the Scott-Knott test (p ≤ 0.05) and uppercase letters compare the effect of evaluation times within each cultivation condition using the F test (p ≤ 0.05).
Figure 5.
Relative water content of leaves – RWC (A), leaf area - LA (B), root length - RL and total dry mass – TDM of Cedrela fissilis Vell. under different availability of water and light evaluated at 15 and 60 days of cultivation. T1 – 0% SH + 40% WHC; T2 – 0% SH + 70% WHC; T3 – 0% SH + 100% WHC; T4 – 30% SH + 40% WHC; T5 – 30% SH + 70% WHC; T6 – 30% SH + 100% WHC; T7 – 70% SH + 40% WHC; T8 – 70% SH +70% WHC; T9 – 70% SH + 100% WHC. Lowercase letters compare the effect of cultivation conditions at each evaluation time using the Scott-Knott test (p ≤ 0.05) and uppercase letters compare the effect of evaluation times within each cultivation condition using the F test (p ≤ 0.05).
Figure 6.
Maintenance of the WHC, with the aid of scales on Cedrella fissilis Vell seedlings (A). Pots with plastic protection used to reduce soil water evapotranspiration (B).
Figure 6.
Maintenance of the WHC, with the aid of scales on Cedrella fissilis Vell seedlings (A). Pots with plastic protection used to reduce soil water evapotranspiration (B).


Table 1.
Regression equations of the effect of evaluation time for photosynthetic rate (A), stomatal conductance – (gs), Rubisco carboxylation efficiencies (A/Ci), transpiration rate – (E) and chlorophyll index (CI) in each cultivation condition.
Table 1.
Regression equations of the effect of evaluation time for photosynthetic rate (A), stomatal conductance – (gs), Rubisco carboxylation efficiencies (A/Ci), transpiration rate – (E) and chlorophyll index (CI) in each cultivation condition.
| CHARACTERISTICS EVALUATED | ||||
|---|---|---|---|---|
| A (µmol CO2 m-2 s-1) | gs (mol H2O m-2 s-1) | A/Ci (µmol mol CO2 m-2 s-1) | ||
| T1 | ŷ = 4.8925 - 0.1979*x + 0.0029*x2; R2; = 0.96 |
ŷ = 0.0333 – 0.0017*x + 0.000037*x2; R2; = 0.97 |
No adjustment | |
| T2 | No adjustment | ŷ = 0.1108 – 0. 0051*x + 0.000078*x2 R2; = 0.96 |
ŷ = 0.0091 + 0.00076*x – 0.000010*x2; R2; = 0.88 |
|
| T3 | ŷ = 13.8925 - 0.5310*x + 0.0071*x2; R2; = 0.70 |
No adjustment | ŷ = 0.0492 – 0.00195*x + 0.000028*x2; R2; = 0.89 |
|
| T4 | ŷ = 5.825 - 0.2688x + 0.0039*x2; R2; = 0.94 |
ŷ = 0.0525 – 0.0030*x + 0.000048*x2; R2; = 0.95 |
ŷ = 0.0289 – 0.0013*x + 0.000018*x2; R2; = 0.93 |
|
| T5 | No adjustment | No adjustment | No adjustment | |
| T6 | ŷ= -0.3825 + 0.3827*x - 0.0042*x2; R2; = 0.93 |
ŷ = 0.2658 – 0.0114*x + 0.000152*x2; R2; = 0.99 |
ŷ = -0.0152 + 0.0024*x – 0.000029*x2; R2; = 0.98 |
|
| T7 | No adjustment | ŷ = 0.0083 + 0.0026*x – 0.000044*x2; R2; = 0.72 |
ŷ = 0.0032 + 0.000099*x; R2; = 0.73 | |
| T8 | ŷ= -1.3425 + 0.3626*x - 0.0041*x2; R2; = 0.98 |
No adjustment | ŷ = -0.0072 + 0.0011*x – 0.000014*x2; R2; = 0.86 |
|
| T9 | ŷ = -1.3275 + 0.3303*x - 0.0037x2; R2; = 0.65 |
ŷ = 0.2283 – 0.0086*x + 0.000096*x2; R2; = 0.99 |
No adjustment | |
| E (mmol H2O m-2 s-1) | CI (Chlorophyll index) | |||
| T1 | ŷ = -0.0033 + 0.0318*x; R2; = 0.78 | No adjustment | ||
| T2 | ŷ = 0.7100 + 0.0264*x; R2; = 0.80 | ŷ = 29.8666 – 0.5777*x + 0.0093*x2; R2; = 0.88 | ||
| T3 | ŷ = 6.330 – 0.3102*x + 0.0040*x2; R2; = 0.96 | No adjustment | ||
| T4 | ŷ = 1.2658 – 0.0499*x + 0.00083*x2; R2; = 0.83 | No adjustment | ||
| T5 | No adjustment | No adjustment | ||
| T6 T7 |
ŷ = -0.1050 + 0.1275*x – 0.00135*x2; R2; = 0.71 | ŷ = 38.6916 – 0.5836*x + 0.0063*x2; R2; = 0.76 | ||
| No adjustment | ŷ = 16.8666 + 0.9144*x – 0.0122*x2; R2; = 0.87 | |||
| T8 | No adjustment | ŷ = 43.0416 – 0.4765*x + 0.0061*x2; R2; = 0.79 | ||
| T9 | No adjustment | ŷ = 29.200 + 0.0824*x; R2; = 0.69 | ||
Table 2.
Phenotypic plasticity index (PPI) for photosynthetic rate (A), potential quantum efficiency of FSII (Fv/Fm), leaf area (LA) and total dry mass (TDM) of Cedrela fissilis Vell. plants grown in different shading levels and water availabilities at 60 days.
Table 2.
Phenotypic plasticity index (PPI) for photosynthetic rate (A), potential quantum efficiency of FSII (Fv/Fm), leaf area (LA) and total dry mass (TDM) of Cedrela fissilis Vell. plants grown in different shading levels and water availabilities at 60 days.
|
Shading level |
PPI (0.00 to 1.00) | |||
|---|---|---|---|---|
| A | Fv/Fm | LA | TDM | |
| 0% | 0.49 | 0.16 | 0.88 | 0.42 |
| 30% | 0.54 | 0.04 | 0.70 | 0.48 |
| 70% | 0.45 | 0.02 | 0.30 | 0.38 |
Table 3.
Average photosynthetically active radiation and leaf temperature values at each shading level at different evaluation times in the cultivation of Cedrela fissilis Vell seedlings.
Table 3.
Average photosynthetically active radiation and leaf temperature values at each shading level at different evaluation times in the cultivation of Cedrela fissilis Vell seedlings.
| Evaluation Period | Shading Level | PAR | Leaf T. (°C) |
|---|---|---|---|
| Days | (μmol m-2 s-1) | (°C) | |
|
15 |
0% 30% 70% |
1159 750 58 |
38.0 33.0 32.0 |
|
30 |
0% 30% 70% |
2038 714 140 |
33.0 31.7 32.0 |
|
45 |
0% 30% 70% |
1934 688 79 |
42.0 36.8 36.1 |
|
60 |
0% 30% 70% |
1212 960 87 |
40.3 38.2 31.5l |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



