Submitted:
02 April 2024
Posted:
03 April 2024
You are already at the latest version
Abstract
A total of 281 coleopteran species from 41 families were recorded from different sites of an abandoned cut-over peatland designated as the Carbon Measurement Supersite in Kaliningrad Oblast. This beetle assemblage is considered as a baseline (pre-impact) faunal assemblage for the further investigations during the ‘before-after’ (BA) or ‘before-after control-impact’ (BACI) study on a peatland that is planned to be rewetted. The spontaneously revegetated peatland has a less specialised beetle assemblage than at an intact raised bog. Tyrphobiontic species are completely absent from the peatland, while some tyrphophiles (5.3% of the total beetle fauna) are still found as remnants of the former raised bog communities. The predominant coenotic coleopteran group is tyrphoneutral generalists from various non-bog habitats (72.9%). The species composition is associated witho the structure vegetation of the disturbed peatland (fragmentary Sphagnum cover, lack of open habitats, and widespread birch coppice or tree stand), which does not correspond to that of a typical European raised bog. The sampled coleopteran assemblage is divided into several relative ecological groups, whose composition and peculiarities are discussed separately. Possible responses to the rewetting measurements in different coleopteran groups are predicted and briefly discussed. A complex assemblage of stenotopic peatland-specialised tyrphophiles (15 spp.) and the most abundant tyrphoneutral generalists (31 spp.) were assigned as indicators for the environmental monitoring of peatland development.
Keywords:
Coleoptera
; indicator species assemblage
; disturbed peatlands
; rewetting
; Baltic Sea region
; checklist
1. Introduction
Peatlands are a unique type of ecosystem, occupying a relatively small fraction of the Earth’s land area, but playing an important role in the global carbon cycle as huge carbon sinks [1]. On the other hand, these communities provide a wide range of structural, hydrological and vegetation diversity, especially if we consider anthropogenically modified peatlands. Although intact and disturbed peatlands often appear similar in terms of their physiognomy and ecology, they can have contrasting environmental relevance, playing a positive role in carbon accumulation and specific biodiversity conservation (natural peatlands) or being hotspots for greenhouse gas emissions and habitat loss (disturbed peatlands).
The issue of habitat transformation is often raised in the context of rewetting projects on disturbed peatlands, which aim to restore natural mire communities and their peat accumulation functions for GHG sequestration. However, peatland restoration usually does not return ecosystems to their original state, but often leads to the formation of completely new communities with unclear consequences at local and global scales [2] or to a loss of heterogeneity and consequently to a loss of species richness [3,4]. In this case, the recovery process requires monitoring of the widest possible range of ecosystem components at all stages of ecosystem development. The characteristic species or species groups of mosses and vascular plants are traditionally of great importance in these investigations when applied to the disturbed peatlands, although there are other specific groups of organisms that can have an important indicator value during the monitoring surveys.
Beetles (Coleoptera) are the most species-rich group on Earth; they have diverse and important ecosystem functions and are well represented in different freshwater and terrestrial ecosystems, as well as in different peatland habitats. Some groups of these insects can be relatively indicative of highly specific environmental conditions [5] and their changes over the course of succession within an ecosystem. The historical development and nature of individual bogs are reflected in their insect assemblages [6], and monitoring Coleoptera could be interesting in the context of environmental change. The studies of coleopteran fauna and ecology in various bog habitats and successional stages of different peatlands are not rare per se, but the knowledge of their biota is insufficent and very uneven [7]. Much of the coleopteran research has been carried out on the biodiversity of the most natural, relic and pristine bogs with focus on epigeic or water beetles e.g. [8,9,10,11,12,13,14,15,16,17,18,19]. Data on other groups of peatland beetles e.g. [20,21,22] or comprehensive studies of the entire fauna e.g. [23] are generally scarce. Faunal surveys of severely disturbed and drained peatlands with degraded bog fauna [24,25] or the fauna of rewetted peatlands after their artificial restoration [26] are underrepresented. With respect to Coleoptera, only one untouched peat bog within the present territory of Kaliningrad Oblast is relatively well studied [9,11,23,27].
In this context, obtaining data on different groups of Coleoptera on a drained peatland seems to be an interesting and relevant part of comprehensive investigations at the Rossyanka Carbon Supersite, as they could be involved into the monitoring programme on sites designated for rewetting in correlation with the other indicator groups. The results of our peatland study are promising and interesting due to the fragmentary knowledge of the coleopteran fauna of peatlands in Kaliningrad Oblast and the paucity of specialised literature.
The objectives of our study were (1) to report pre-impact data on the coleopteran taxonomic diversity in an abandoned cut-over peatland in Kaliningrad Oblast, Russia; (2) to determine the structure of the beetle assemblage in terms of species arrangement within ecological groups and peat bog affinity categories; (3) to identify indicator species and species assemblages relevant for monitoring environmental changes during further peatland rewetting.
2. Materials and Methods
The investigated territory (Figure 1 and Figure 2), the Vittgirrensky peatland (54.799˚ N, 21.658˚ E) with a total area of 122 ha is located in the Slavsk administrative district of Kaliningrad Oblast southwards the extinct village of Wittgirren (=Weißenbruch), 18 km SE of Bol’shakovo (=Groß Skaisgirren/Kreuzingen), 6 km NE of the Vysokoe (=Popelken/Markthausen). The Vittgirrensky peatland is a ditched, drained and heavily human-transformed raised bog. The edge zone of this bog was drained at the end of the 19th century and peat was mined by hand cutting [28]. In the late 1970s and 1980s, the peatland was completely drained by a combined (open and closed) drainage system. The area was intensively used for commercial peat extraction by the milling method during the period 1980–1996. Later, the Vittgirrensky peat cutting area was abandoned after the peat extraction ceased. Judging by burnt tree trunks and charcoal residues in the uppermost peat layers, the area was the subject of local surface fires. In 2021, the area was declared as a Carbon Supersite (‘Rossyanka’) to estimate and monitor greenhouse gas fluxes, study biodiversity and test rewetting technologies for fire prevention, climate change mitigate and habitat restoration.
The study area is located in a region with a temperate climate with an average annual precipitation of 750 mm and an average annual temperature of +7.8 °C. Our own measurements showed the following average monthly temperatures and average monthly liquid precipitation on the peatland area during the year 2023: +12.1°C and 7.44 in May, +16.7°C and 86.4 mm in June, +17.3°C and 81.84 mm in July, +19.2°C and 119.04 mm in August, +16.4°C and 36.0 mm in September, +8.6°C and 89.28 mm in October, and +2.2°C and 36 mm in November. The frost-free period was 180 days.
At present, the peatland is covered by regrowth communities that have developed after the destruction of the primary bog vegetation [28]. Vegetation cover is more diverse in areas with stable soil moisture (remnants of bog and wetland communities) and along the central road (ruderal plant community). The current dominant arboreal species on the Vittgirrensky peatland is birch (Betula pendula and B. pubescens), but a small admixture of Populus tremula, Salix spp. (at least 8 species), Sorbus aucuparia, and Pinus sylvestris is also present. The birch coppice and regrowth form different mosaic communities (with Calluna, Ledum, Phragmites) of mainly semi-open character with the oldest trees being about 30 years old (Figure 2). Bare-peat sites, fen-like communities (Juncus, Eriophorum, Carex) and birch thickets can be also found in the peatland. The detailed maps and comprehensive data on vegetation and soils are given in recent publications [28,29,30]. The Vittgirrensky peatland is situated within an intensively used open agricultural landscape in relative isolation from faunal interchanges with the pristine or semi-natural bog habitats for a long time. The nearest intact raised bog Bol’shoye (former Mupiau/Palmbruch, with a total area of about 600 ha) is located 7 km southwest (54,735° N, 21,615° E).
The material presented in this paper was collected by the author between April and November 2023. Adult Coleoptera were collected using the following methods and equipment:
(1) sweeping with entomological sweep nets during 7 days of field surveys (07.04.2023; 13.05.2023; 13.06.2023; 13.07.2023; 13.08.2023; 17.09.2023; 13.10.2023) in different terrestrial and aquatic biotopes of the study area;
(2) visual search of vegetation, soil, fungi, as well as under pieces of wood and bark with hand collection during 8 field survey days (07.04.2023; 13.05.2023; 13.06.2023; 13.07.2023; 13.08.2023; 17.09.2023; 13.10.2023; 17.11.2023) in different terrestrial biotopes of the study area;
(3) pitfall traps without bait (the 250 cm³ plastic containers with 9 % acetic acid as a preservative liquid, placed in lines of 10 traps with a distance between traps of about 1 m) in seven-month periods between field surveys (07.04–13.05.2023; 13.05–13.06.2023; 13.06–13.07.2023; 13.07–13.08.2023; 13.08–17.09.2023; 17.09–13.10.2023; 13.10–17.11.2023) in 3 terrestrial biotopes of the studied area (Figure 2 and Figure 3). The trap lines were placed in: (1) the N edge of the peatland (sun-exposed site with Calluna, Eriophorum, and Betula coppice), (2) the SW edge of the peatland (dense birch stand) and (3) the central part of the peatland (sun-exposed site with partially bare peat, Campylopus introflexus and Betula regrowth). Several containers were effective in attracting carrion beetles (Silphidae, Cholevinae) after the accidental capture of shrews, voles and lizards in the originally unbaited trap.
The larval stages of the beetles were ignored except for three characteristic species (Endomychus coccineus, Harmonia axyridis, and Schizotus pectinicornis). Some of the material examined is in the author’s private collection (Kaliningrad, Russia). Taxa widely distributed in Kaliningrad Oblast and identifiable in field were recorded without collecting specimen. The specimens were identified by the author using the standard European identification key [31,32,33,34]. The order used in the checklist is systematic for families and is given according to the recent classification [35]. Genera and species collected or observed in the field are arranged alphabetically (by scientific name).
A rough estimation and A.F.O.R. scale was used to measure local abundance with the following values:
1) abundant (A) – species observed or sampled by ≥10 specimens/day or species pitfall-trapped by ≥10 specimens/month;
2) frequent (F) – species observed or sampled by 5–9 specimens/day or species pitfall-trapped by ≥5–9 specimens/month;
3) occasional (O) – species observed or sampled by 2–4 specimens/day or species pitfall-trapped by 2–4 specimens/month;
4) rare (R) – species observed or sampled by single specimen/day or species pitfall-trapped by single specimen/month.
The structure of the species profile in the checklist includes: full species name (maximum local abundance estimated using the A.F.O.R. scale); the sampling method; the date or period of capture; location; ecological notes (if any); ecological category of the species. Three main sampling methods are indicated as ‘net’ (for sweeping with entomological hand net in water or through vegetation), ‘visual’ (for hand collection) or ‘pitfall’ (for pitfall traps). Approximate sampling locations are abbreviated as follow: C – central part (core peatland area, approximately restricted by corresponding quarters), E – eastern quarter of total area, N – northern, S – southern, SE – south-eastern, SW – south-western, W – western. The basic ecological category for each species was chosen from four options according to the classification of peat bog entomofauna by Spitzer and Danks [6]. The possible groups are: tyrphobiontic species (found only in bogs, not in the study area), tyrphophilous species (characteristic of bogs but not restrictid to them), tyrphoneutral species (resident in bogs and peatlands, but also in other habitats), and tyrphoxenous species (non-resident vagrants or erratics, unable to form stable perennial populations in bogs and peatlands). Inhabitants of roadside ruderal plant communities were considered tyrphoxenous.
3. Results
The examination and identification of the material from the study area (samples from 2023) resulted in a list of 281 species (41 families) was compiled. Among the Coleoptera collected, 231 species (82 %) are terrestrial in the adult stage and 50 species (18 %) are aquatic. A total of 166 coleopteran species were collected by sweep netting in water or on vegetation (144 species exclusively by this method), 104 species were sampled using terrestrial pitfall traps (83 species exclusively by this method), and 41 species were recorded by other methods (21 species registered only in this way). In total, 30 species were recorded using two or more methods.
A complete preliminary list of the beetle species found in the study area during the 2023 sampling season is given in Appendix A.
3.1. Taxonomic Structure and Abundance
In order to assess the taxonomic structure of the beetle fauna in the study area, we present our results in comparison with data from a nearly intact large (ca. 2500 ha) raised bog – Zehlau – located 50 km southwards (Figure 1), which is considered a reference site due to several entomological inventories carried out there during the last century. Skwarra [23] collected 332 coleopteran species in the Zehlau raised bog between 1920 and 1926. Främbs et al. [11] sampled 36 species of the family Carabidae in 1994 in the Zehlau bog and its nearest vicinity. Biesiadka and Мoroz [9] reported 43 species of the family Dytiscidae. The brief comparison of our results with the peatland fauna recorded by Skwarra [23], Främbs et al. [11] and Biesiadka, Мoroz [9] is possible, although it can only be done with reservations (different structural complexity and set of habitats, different studied area, different sampling methods, etc.).
The faunistic core in both peatlands is consists of the same most species-rich families (≥10 species): Cantharidae, Carabidae, Chrysomelidae, Coccinellidae, Curculionidae, Dytiscidae, Elateridae, and Staphylinidae (Table 1). The percentages of each family also show similar values and arrangements in both peatlands.
The local abundance estimated for all recorded coleopteran species shows the following result in 2023: 25 coleopteran species (8.9%) were considered as abundant, 55 species (19.6%) were estimated as frequent, 89 species (31.7%) as occasional and 112 species (39.9%) were considered as rare. The estimation of abundance is based on the frequency of occurrence of the adult stage only. As it is generally subjective in our case due to different methodological approaches and totally different bionomy of the studied objects, it should reflect the real population densities in the habitat and can be used for comparison purposes. The abundance structure does not differ in the two main ecological groups of peatland Coleoptera studied: terrestrial and aquatic beetles (amphibiontic Scirtidae with aquatic larva and terrestrial imago were included in the calculation as terrestrial). Within the aquatic beetles (all sampled with the net), the abundance composition is as follows: abundant – 4 spp. (8%), frequent – 10 spp. (20%), occasional – 16 spp. (32%), rare – 20 spp. (40%). Within terrestrial beetles (sampled by different methods), the abundance composition shows a similar arrangement: abundant – 21 spp. (9.1%), frequent – 45 spp. (19.5%), occasional – 73 spp. (31.6%), rare – 92 spp. (39.8%).
We hypothesise that the structure of abundance within coleopteran assemblage: (1) depends on the successional stage of a habitat and reflects the density of beetle populations; (2) can be effectively used for monitoring peatland Coleoptera using different simple sampling methods in both aquatic and terrestrial habitats. Such an estimate of species abundance should be sufficient to investigate the effects of human activities on coleopteran assemblages, and in particular on the rewetting of degraded peatlands.
3.2. Peat-Bog Biotope Fidelity and Habitat Structure
The ecological relevance of coleopteran species composition becomes clearer at the species level when we interpret our entomological biodiversity in the context of species distribution in basic peatland communities (Figure 2). This implies that species are grouped according to their fidelity to a particular peat bog biotope. As peat bogs are characterised by very specific and often extreme environments, many bog insect species are thought to be highly specialised and confined to them [6]. Thus, an intact bog should provide some stenotopic insect assemlages that could be considered as species groups of indicative value.
These biotope-confined species assemblages are generally identified using the Peus’ classification, which includes four categories [6,36]: (a) tyrphobionts – occuring exclusively in bogs, (b) tyrphophiles – typical of bogs, but not strictly confined to them, (c) tyrphoneutrals – distributed across different habitat types, and (d) tyrphoxenes – immigrants that cannot survive in bogs.
Tyrphobiontic species were not sampled in the study area. We suggest that this may be related to the current habitat structure and vegetation in the transformed peatland, which now differs from the typical raised bog. In the latter habitat, taxa are thoght to be strongly influenced by specific conditions – high soil moisture and Sphagnum moss dominance [6]. These two factors should apparently be combined to provide the necessary conditions for tyrphobiontic beetles, whereas in the Vittgirrensky peatland, Sphagna are found sporadically and the water table is very unstable.
Tyrphophiles were selected for each ecological group (aquatic, terrestrial, and amphibionthic). The peatland specialist species are the following 15 beetles: (1) Hydrochus elongatus and Enochrus ochropterus (aquatic acydophilous stagnobionts), (2) Contacyphon variabilis (amphibiontic beetles), (3) Scymnus suturalis and Malthodes pumilus (terrestrial dwarf shrub beetles), (4) Acupalpus brunnipes, Brachygluta haematica, Bradycellus rufcollis, Curimopsis nigrita, Ischnosoma longicorne, Platydracus fulvipes, P. latebricola and Pselaphus heisei (epigeic beetles), (5) Altica longicollis and Micrelus ericae (phytophagous beetles). These beetles constitute 5.3% of the beetle assemblage ant probably represent remnants of the original raised bog communities.
Thus, despite the lack of continuous Sphagnum cover, some bog specialists within the tyrphophilous group can survive on a disturbed peatland, apparently, due to the other ‘true’ bog plants remaining and the preservation of the residual oligotrophic peat layer. Such environments could be considered as refugia for bog-specialised insect species, which can play a role of bioindicators within different scenarios of peatland development (including bog habitat rehabilitation).
The other 266 species sampled represent elements of various secondary habitats and are mainly eurytopic generalists. Of this, 205 species (i.e. 72.9%) were considered tyrphoneutral and 61 species (21.7%) were considered tyrphoxenous. Thus, the actual fauna of the Vittgirrensky peatland consists mainly of adaptable generalists, while relict bog elements are scarcely represented, being even less species-rich than tyrphoxenous (i.e. ‘alien to petland’) species.
As the most common species are critical to the coleopteran assemblage, future comparative analyses can assess the impact of peatland restoration on the abundance of these species. The tyrphophiles (peatland specialists) as well as the most abundant tyrphoneutral generalists can be used to monitor potential environmental impacts on degraded peatland communities in case of restoration. The most abundant non-peatland specialist species recorded in 2023 and likely to be of indicative value are the following 31 tyrphoneutral species: (1) Haliplus ruficollis, Hyphydrus ovatus, Anacaena lutescens, Acilius canaliculatus, Hygrotus inaequalis, Hydroporus angustatus, and H. neglectus (aquatic environment); (2) Anthobium unicolor, Apocatops nigrita, Leistus terminatus, Nicrophorus vespilloides, Pterostichus niger, Sciodrepoides watsoni, Tachyporus dispar, Xantholinus longiventris, Amara famelica, Poecilus cupreus, Acidota crenata, Drusilla canaliculata, Oxypoda opaca, Philonthus cognatus (epigeic assemblage); (3) Altica aenescens, Lochmaea caprea, Brachysomus echinatus, Polydrusus picus, Telmatophilus typhae, Bromius obscurus, Crepidodera aurata, Orchestes jota, Polydrusus cervinus, and Sciaphilus asperatus (plant-associated beetles). Restoration measures in the Vittgirrensky peatland could have a neutral or negative effect on the abundance of these most common tyrphoneutral species.
4. Discussion
The present-day coleopteran assemblages and their species composition are primarily associated with the structure of the current vegetation cover in the Vittgirrensky peatland. Although 27 ‘true’ plant species are now common in the peatland [28], many of them are sporadic or do not form typical bog communities as dominant species [29]. Moreover, the composition, spatial heterogeneity and distribution patterns of present-day peatland communities differ from those of a typical raised bog. Table 2 compares the lists of basic plant communities on both ecosystems in a comparative manner. The comparison shows that most of the communities on a disturbed peatland, which represent the successional stages of spontaneous vegetation recovery [29], can not be transferred to the communities of a natural bog. The main characteristics of disturbed peatlands are the absence of Sphagnum carpets in the ground layer and the small area of open sites [28]. These transformations are mainly caused by a strong drainage which also leads to the loss of ‘true’ mire species and the formation of new habitats – more similar to non-bog communities (Figure 3).
All these changes result in alterations in the beetle fauna. The current coleopteran assemblage of the Vittgirrensky peatland is diverse and shows a mixture of components of different habitats and communities: edges of small-leaved forest, open habitats, secondary wetlands and ruderal communities within the agricultural landscape. Most of the coleopteran fauna of the Vittgirrensky peatland (266 spp., 94.7%) does not belong to the natural fauna of undisturbed raised bogs.
Possible rewetting of the area aims to raise the local water table and could therefore reestablish niches for species adapted to wet conditions and open water, as well as for species sensitive to significant water table fluctuations. An increase in peatland-specialised populations (15 species mentioned above) can be predicted in the case of peatland restoration, but it should be factually confirmed and registered for different taxonomic and ecological groups. Neutral effect, population decline or spatial redistribution for non-peatland specialised species (31 observable species listed above) can be assumed but is not easy to predict. Some predictions have been made in the relevant section for each ecological group.
The comparison of numbers and percentages is relatively rough, formal and its results can be interpreted in many different ways. Table 1 shows that the beetles of the family Staphylinidae are underrepresented in the materials collected in the Vittgirrensky peatland. This fact should be explained by the low representation of the small-sized dwellers of mooses and soil in our 2023 samples. The ‘better’ representation of Cantharidae, Carabidae, Curculionidae, and Elateridae in the Vittgirrensky peatland than in the nearly intact Zehlau bog could be explained by the additional number of non-specialised species from the surrounding habitats in the ditched, drained and heavily anthropogenically altered peatland compared to a pristine Sphagnum bog.
4.1. Ecological Characteristics and Peatland Habitat Affinity
The beetle assemblage has a mixed character and consists of wetland-specialised species, forest species, hydrophilous near-water species and species associated with open landscapes. The beetle assemblage of the Vittgirrensky peatland can be divided into three main ecological groups, which will be discussed separately: aquatic beetles (7 families: Hydrophilidae, Hydraenidae, Hydrochidae, Dytiscidae, Haliplidae, Gyrinidae, and Noteridae), amphibiontic beetles (single family Scirtidae with aquatic larva and terrestrial imago), and terrestrial beetles (33 families, with only a few representatives associated with peat soils, mire plants, banks of waterbodies). The latter group is further divided into subgroups (see below).
4.1.1. Aquatic Beetles
Based on Ryndevich [34], all water beetles of the geographically and climatically close Belarus can be divided into several groups according to their preference for flowing or stagnant waterbodies (reophilic, reobiontic, stagnobiontic) and their preference for water pH values (acydophilous, pH=3.3-6.8; eurytopic, and alcalophilous). The water beetle assemblage of the Vittgirrensky peatland (50 spp. in total) includes three groups of the classification: reophilic (23 spp., 46%), acydophilous stagnobionts (2 spp., 4%) and eurytopic stagnobionts (25 spp., 50%). The reophilic species (inhabitants of the both flowing and stangnant waters) included: Gyrinus substriatus, Haliplus immaculatus, H. lineatocollis, H. ruficollis, P. caesus, Noterus clavicornis, Acilius canaliculatus, A. sulcatus, Agabus bipustulatus, Dytiscus dimidiatus, Graphoderus cinereus, Graptodytes pictus, Hydaticus seminiger, Hydroporus obscurus, H. palustris, Hygrotus impressopunctatus, H. inaequalis, Hyphydrus ovatus, Ilybius ater, Laccophilus minutus, Porhydrus lineatus, Rhantus suturalis and Scarodytes halensis. The eurytopic stagnobionts (inhabitants of stagnant waters without pH preference) were included: Haliplus fulvus, Clemnius decorates, Graptodytes granularis, Hydaticus transversalis, Hydroporus angustatus, H. dorsalis, H. erythrocephalus, H. glabriusculus, H. neglectus, H. planus, Ilybius guttiger, Rhantus exsoletus, Helophorus aquaticus, H. granularis, H. flavipes, Anacaena lutescens, Cymbiodyta marginella, Enochrus coarctatus, E. testaceus, Helochares obscurus, Hydrobius fuscipes, Hydrochara caraboides, Laccobius minutus, Limnebius crinifer and Ochthebius minimus. Many species of these two groups also occur in pristine peat bogs and have been reported e.g. from the Zehlau bog [9,23], but these inhabitants are not bog specialists and could repeatedly enter the study area by air or even directly from neighbouring water bodies without leaving the water.
The most ecologically interesting group are the acydophilous stagnobionts, which prefer acidic waters and are represented in the study area by Hydrochus elongatus and Enochrus ochropterus. These two species were sampled in the southern part of the Vittgirrensky peatland in a non-drying drainage ditch, where the most diverse water assemblage was recorded in 2023. It is likely that these species only used this water body as a temporary refuge when their preferred water bodies dried up, and that the high diversity here is a consequence of the secondary concentration of flying adults.
The most abundant and widespread species of the Vittgirrensky peatland in 2023 are Haliplus ruficollis (reophilic), Hyphydrus ovatus (reophilic), and Anacaena lutescens (eurytopic stagnobiont); common and widespread are Acilius canaliculatus (reophilic), Hygrotus inaequalis (reophilic), Hydroporus angustatus (eurytopic stagnobiont), and H. neglectus (eurytopic stagnobiont).
We suggest that further monitoring of the Vittgirrensky peatland should pay more attention to indicator aquatic species (such as acydophilous stagnobionts), but changes in the water beetle assemblage may also occur in the distribution or abundance of other community members. It can be hypothesised that the restoration measures in the Vittgirrensky peatland will lead to an increase in the water level and the the number of permanent water bodies in the area. The increase of stagnobionts (and specialised acydophilous stagnobionts) occurrence and population density in such case could be predicted for the central area of the peatland. The reophilic elements would favour larger drainage ditches in the periphery of the peatland, where their diversity and abundance might increase. The general zonation and ecological stratification must take place.
4.1.2. Amphibiontic Beetles
This group of beetles is represented by five species of the single family Scirtidae: Contacyphon ochraceus, Contacyphon padi, Contacyphon variabilis, Microcara testacea, and Scirtes hemisphaericus. Three of these species are considered to be common in the study area in 2023: Contacyphon padi, Contacyphon variabilis, and Scirtes hemisphaericus. All adult scirtids are associated with shoreline habitats, and Contacyphon variabilis is a common species of peatlands and raised bogs. The likely rise in water levels and the increase in the number of permanent water bodies in the Vittgirrensky peatland after restoration should lead to an increase in the abundance and probably species richness of these beetles due to the formation of additional larval development sites, such as small persistent puddles and ditches.
4.1.3. Terrestrial Beetles
This ecological group (33 families, 226 species) is the most diverse in terms of species and bionomy and therefore the most difficult to analyse. For the purposes of analysis, we divide the sampled terrestrial beetles into three conditional ecological groups, such as plant-associated phytophagous beetles (Attelabidae, Brentidae, Byturidae, Buprestidae, Chrysomelidae, Curculionidae, Kateretidae, Megalopodidae, Nitidulidae, Phalacridae), epigeic elements (Byrrhidae, Carabidae, Staphylinidae as well as necrobiontic Dermestidae, Silphidae and Leiodidae: Cholevinae) and ‘miscellaneous’ Coleoptera (i.e. other families including representatives of mycetobionts, predators, pallinophagous vegetation feeders, etc.). The subgroups of the terrestrial beetle assemblage of the Vittgirrensky peatland are briefly and separately discussed below.
4.1.3.1. Miscellaneous Terrestrial Beetles
Many of the beetles in this subgroup are not directly associated with peatland (or even wetland) habitats in the adult or in all stages and were recorded in the study area as rare or occasional residents or short-distance migrants in a specific ‘ruderal’ community of the central roadside and shrub vegetation of peatland margins. The subgroup includes the following 17 families: Cantharidae, Cerambycidae, Coccinellidae, Cryptophagidae, Elateridae, Endomychidae, Latridiidae, Leiodidae, Lucanidae, Malachiidae, Mordellidae, Oedemeridae, Pyrochroidae, Scarabaeidae, Scraptiidae, Tenebrionidae, and Throscidae. Cantharidae, Coccinellidae and Malachiidae are predaceous or pallinophagous on vegetation. Two species of these families collected in 2023 are considered to be specific to peatlands: Scymnus suturalis and Malthodes pumilus. Other recorded representatives are eurytopic generalists and are associated with a wide range of non-specific semi-open habitats (shrubs, forest margins etc.). Members of Cerambycidae, Elateridae, Lucanidae, Mordellidae, Oedemeridae, Pyrochroidae, Scarabaeidae (Melolonthinae, Cetoniinae), Scraptiidae, Tenebrionidae and Throscidae are usually the hidden inhabitants of plant tissues (mainly wood) or soil in the larval stage and visitors of vegetation (often flowers) in the adult stage. The arboreal vegetation of the Vittgirrensky peatland (Betula, Populus, Salix, Pinus) and the presence of decayed wood determine the fauna of these beetles. Lucanidae are not considered to be part of the local beetle community, the single observed specimen of Dorcus represents a tourist (i.e. not resident) species in the peatland.
Restoration measures in the peatland, resulting in water level rise and vegetation changes in the core peatland area, are unlikely to have a major impact on the populations on most of these insects. The relatively rare species Oedemera croceicollis may be positively affected by rewetting of the area. Two registered members of the dung community (Scarabaeidae: Aphodiinae) are non-specialised and are associated with open landscapes. Mycetobiontic Leiodidae (e.g. Agathidium), Cryptophagidae, Endomychidae and Latridiidae are associated with decaying plant material and various fungi. Also for this ecological group of Coleoptera no negative effects of the restoration measures in the Vittgirrensky peatland are predicted. Apparently, many species of this group have a high dispersal and re-colonisation ability and can easily invade suitable habitats from the outside. Several additional representatives of Monotomidae, Nitidulidae, Ciidae and Scolytinae not recorded in 2023 are expected to be found in the periphery of the peatland during targeted searches in the closed canopy birch forest.
It is noteworthy that one of the species recorded in the Vittgirrensky peatland (along the central road) in 2023, Harmonia axyridis, is a recent (first decade of the XXI century) active invader in the territory of Kaliningrad Oblast. This species is now common in towns and cities, and probably arrived in the Vittgirrensky peatland by transport after the resumption of human activity in the area in recent years.
4.1.3.2. Epigeic Beetles
The epigeic beetles include representatives of 5 families: Byrrhidae, Carabidae, Dermestidae, Silphidae and Staphylinidae. The composition of this group is nominal, because the family Staphylinidae is very diverse in its bionomy and includes mycetobionts, predators occurring on soil, fungi and vegetation, scavengers, inquilines of ant nests, etc. Perhaps the most important unifying feature of the five families mentioned above is the method of collection: the beetles of this group are effectively sampled using pitfall traps. The majority of the sampled epigeic beetles are not bog or petland specialists and can be characterised as open habitat species (e.g. Amara famelica, Cymindis vaporariorum, Drusilla canaliculata, Poecilus cupreus, Stenus solutus), forest species (e.g. Anthobium unicolor, Carabus nemoralis, Leistus terminatus, Pterostichus niger, Sepedophilus testaceus, Xantholinus longiventris) or necrophagous species (e.g. Nicrophorus vespilloides). Two of the most characteristic bog species in the Baltic Region, Agonum ericeti and Pterostichus rhaeticus [11,37], were not found in the Vittgirrensky peatland.
The most abundant and widespread epigeic species of the Vittgirrensky peatland in 2023 are: Acupalpus brunnipes, Anthobium unicolor, Apocatops nigrita, Leistus terminatus, Nicrophorus vespilloides, Pterostichus niger, Sciodrepoides watsoni, Tachyporus dispar, and Xantholinus longiventris. Common and abundant are Amara famelica, Poecilus cupreus, Acidota crenata, Drusilla canaliculata, Oxypoda opaca, Philonthus cognatus and Platydracus latebricola. Proteinus brachypterus and Stenus solutus are also widespread and probably ubiquitous in the area, but are considered to be occasional or rare due to the small number of recorded specimens.
The epigeic beetle assemblage of the study area is diverse and includes a mixture of fauna from different habitats. The following epigeic coleoptera can be considered as peatland specialists: Acupalpus brunnipes, Bradycellus rufcollis, Curimopsis nigrita, Platydracus fulvipes and P. latebricola. In addition, species such as Brachygluta haematica, Ischnosoma longicorne, Pselaphus heisei are more characteristic of peatlands and prefer similar habitats, but are not strictly stenotopic (e.g. P. heisei also occurs on forest edges). A number of species are associated with different types of riparian zones, inhabiting the margins of drainage ditches or Phragmites-dominated communities: Bembidion quadrimaculatum, Dyschirius globosus, Hygronoma dimidiata, Lesteva longoelytrata, Pterostichus anthracinus, Stenus spp. Representatives of Dermestidae, Silphidae and Leiodidae (Cholevinae) are almost exclusively members of necrophagous communities and are therefore not true peatland specialists. Restoration measures may negatively affect populations of the sun-adapted species of open habitats and dry conditions (Cymindis vaporariorum, Cicindela campestris), but will favour the hydrophilous beetle assemblage of shores and wetland communities, e.g. associated with moss cover.
It should be noted that wetland specialised species in the Vittgirrensky peatland are much more numerous and abundant among epigeic beetles than among aquatic ones. It can be assumed that the degradation of the aquatic coleopteran community in the study area is deeper, while the terrestrial coleopteran communities have time to recover in the period after the end of peat extraction. The natural water bodies of the raised bog have almost completely disappeared, whereas several microhabitats of terrestrial epigeic beetles remain in small patches or partially.
4.1.3.3. Plant-Associated Phytophagous Beetles
All representatives of 9 families were considered as plant-associated phytophagous beetles: Attelabidae, Brentidae, Byturidae, Chrysomelidae, Curculionidae, Kateretidae, Megalopodidae, Nitidulidae, and Phalacridae. In addition, three species associated with living vegetation from Buprestidae (Trachys minutus), Cerambycidae (Agapanthia villosoviridescens) and Cryptophagidae (Telmatophilus typhae) are considered phytophagous and are included in this subgroup, which consists of 73 species from 12 families. Among these beetles, 8 species were considered abundant (Brachypterus urticae, Altica aenescens, Altica longicollis, Lochmaea caprea, Neocrepidodera transversa, Barypeithes pellucidus, Brachysomus echinatus and Polydrusus picus), 11 species were considered frequent (Telmatophilus typhae, Batophila rubi, Bromius obscurus, Cassida rubiginosa, Crepidodera aurata, Psylliodes chalcomerus, Orchestes jota, Phyllobius pomaceus, Polydrusus cervinus, Rhinocyllus conicus, Sciaphilus asperatus and Tanymecus palliatus), 23 species are occasional and 31 species are rare. Most of the abundant and frequent species are associated by beetles associated with Betula and Salix (Altica aenescens, Lochmaea caprea, Polydrusus picus, Crepidodera aurata, Orchestes jota, etc.) or ruderal plants of the central road such as Urtica, Chamaenerion, Rubus, Cirsium, and Carduus (Brachypterus urticae, Neocrepidodera transversa etc.). Only one ‘abundant’ species, Altica longicollis, is associated with the peatland (communities with Calluna) and only one frequent species, Telmatophilus typhae, indicates the wetland area with Typha community.
From the complied list of phytophagous beetles, the following two species can be considered as peatland specialists in the study area: Altica longicollis and Micrelus ericae (both trophically related to the peatland stenobiont Calluna vulgaris). Several other species are often found in different types of wetlands, although they tend to be eurytopic and depend mainly on the distribution of fodder plants. They are Altica aenescens, Betulapion simile, Orchestes jota, Phyllobius argentatus, Polydrusus cervinus (fodder plant: Betula), Archarius salicivorus, Byctiscus betulae, Crepidodera aurata, Cryptocephalus decemmaculatus, Cryptocephalus ocellatus, Pachybrachis hieroglyphicus, Polydrusus picus, Temnocerus coeruleus, Trachys minutus (fodder plant: Salix), Lochmaea caprea, Rhamphus pulicarius (fodder plants: Salix and Betula), Byctiscus populi, Chrysomela populi, Zeugophora subspinosa (fodder plant: Populus tremula), Brassicogethes viridescens, Phyllotreta atra, Phyllotreta striolata (olygophagous on Cruciferae), Telmatophilus typhae (fodder plant: Typha latifolia), Nanophyes marmoratus (fodder plant: Lythrum salicaria), Phyllobius pyri (polyphagous on arboreal plants), Phalacrus championi (fodder plant: Cyperaceae), Oulema obscura (fodder plant: Poaceae), and Agelastica alni (fodder plant: Alnus). The distribution of these beetles should generally coincide with the distribution of the fodder plants in the Vittgirrensky peatland and can be predicted using vegetation maps.
It is interesting to note that representatives of Kateretes (Kateretidae), Donaciinae (Chrysomelidae), Limnobaris and Erirhinini (Curculionidae), which are characteristic of wetland and associated with diverse plant communities, including their forage plants Typha, Phragmites, Scirpus, Carex, Sparganium etc., were not sampled in the study area. This fact may be related to the strong seasonal fluctuations of the water table in the area and drying up of the water bodies. These beetles were expected to be found on the northern edge of the peatland. Colonisation of the central area by these beetles after rewetting of the peatland is also likely.
4.2. Note
Several collection methods that are effective in wetlands were not used in our surveys (e.g. light trapping and sieving of substrates). Pitfall trapping was only carried out at three sampling sites. The resulting species list has a preliminary character and reflects the beetle assemblage of mainly central, western and southern areas of the Vittgirrensky peatland, and is rather incomplete with respect to small-sized soil dwellers. The possible inter-annual variation of the beetle assemblage cannot be estimated from the single six-month survey. Taking into account that both spatial and temporal factors influence the invertebrate faunistic diversity of peatlands, further studies in the Vittgirrensky peatland may add a number of additional species to the list of inhabitants and correct the estimates of species abundance.
Long-term monitoring and data collection of Coleoptera is recommended and may be of interest in any case of the possible management of the Vittgirrensky peatland, i.e. rewetting by damming ditches or leaving the situation unchanged. In spite of all the difficulties mentioned, the data obtained in 2023 provide baseline information for comparison and monitoring of environmental changes in case of rewetting of the Vittgirrensky peatland and can be used in before-after (BA) or even in before-after control-impact (BACI) studies.
5. Conclusions
After more than 20 years of abandonment and spontaneous re-establishment of vegetation cover on a cut-over peatland (Vittgirrensky) in the Kaliningrad region, a total of 281 coleopteran species from 41 families were recorded here from different sites. This beetle assemblage is considered to be a pre-impact fauna to be further investigated during the ‘before-after’ (BA) or ‘before-after control-impact’ (BACI) study on a peatland which is planned to be rewetted.
Compared to an intact raised bog, the Vittgirrensky peatland has a less specialised beetle assemblage, showing an intermixed fauna of small-leaved forest edges, open meadows, wetlands and ruderal communities of the agricultural landscape. Tyrphobiontic species (bog specialists) are completely absent from the peatland, while tyrphophiles (typical of bogs) still constitute 5.3% (15 spp.) of the beetle fauna, probably representing remnants of the former raised bog communities. The major part of the today’s peatland coleopteran assemblage are tyrphoneutral (205 spp., 72.9%), which belong to adaptable generalists from various non-bog habitats.
The current coleopteran species composition of the abandoned disturbed peatland is associated with the structure of its current vegetation cover, which is characterised by a fragmentary distribution of Sphagnum carpets, lack of the open habitats, and widespread birch coppice or tree stands. Most of today’s peatland communities do not correspond to those of a typical European raised bog.
The heterogeneity of vegetation and soil cover determines the diversity of coleopteran ecological groups in the peatland, with such groups as: terrestrial (226 spp., including plant-associated phytophages, epigeic, ‘miscellaneous’), aquatic (50 spp., including reophilic species, eurytopic stagnobionts, acydophilous stagnobionts) and amphibiontic (5 spp.).
The rewetting measures will most likely have a positive effect on the stenotopic peatland-specialised group of tyrphophiles (15 spp.) and – but neutral or negative (populations decrease or spatial redistribution) – on a group of the most abundant tyrphoneutral generalists (31 spp.). These two coleopteran aggregates of 46 species can be considered as an indicator complex (Figure 4) used to monitor environmental changes during further peatland development in terms of different scenarios.
Author Contributions
V.A. and M.N.: conceptualization and project administration; V.A.: methodology, investigation, original draft preparation; M.N.: formal analysis; V.A., M.N. and T.N-D.: review and editing; V.A. and T.N-D.: visualization; M.N. and T.N-D.: funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
The study of the second and third authors was support by the state assignment of the Ministry of Science and Higher Education of the Russian Federation, theme No. FZWM-2024-0015.
Data Availability Statement
All relevant data sets in this study are described in the manuscript.
Acknowledgments
The authors thank two anonymous reviewers for their valuable suggestions, which improved the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A
Preliminary checklist of beetles (Coleoptera) of the Vittgirrensky peatland.
1. Gyrinidae Latreille, 1810 (1 species)
1.1. Gyrinus substriatus Stephens, 1828 [O]: net; 13.05.2023; C, E; tyrphoneutral.
2. Haliplidae Aubé, 1836 (5 species)
2.1. Haliplus fulvus (Fabricius, 1801) [O]: net; 13.05, 13.06, 13.08.2023; S; tyrphoneutral.
2.2. Haliplus immaculatus Gerhardt, 1877 [O]: net; 07.04, 13.05, 13.06.2023; S; tyrphoneutral.
2.3. Haliplus lineatocollis (Marsham, 1802) [R]: net; 13.05.2023; C; tyrphoneutral.
2.4. Haliplus ruficollis (DeGeer, 1774) [A]: net; 13.05, 13.06, 13.07, 13.08, 17.09, 13.10.2023; C, E, S; tyrphoneutral.
2.5. Peltodytes caesus (Duftschmid, 1805) [O]: net; 13.05, 13.06, 13.07.2023; S; tyrphoneutral.
3. Noteridae Thomson, 1860 (1 species)
3.1. Noterus clavicornis (DeGeer, 1774) [F]: net; 07.04, 13.05, 13.06.2023; S; tyrphoneutral.
4. Dytiscidae Leach, 1815 (28 species)
4.1. Acilius canaliculatus (Nicolai, 1822) [F]: net; 07.04, 13.05, 13.06, 13.07, 13.08, 17.09.2023; C, S, W; tyrphoneutral.
4.2. Acilius sulcatus (Linnaeus, 1758) [O]: net; 13.05, 13.06, 13.08, 17.09, 13.10.2023; E, S; tyrphoneutral.
4.3. Agabus bipustulatus (Linnaeus, 1767) [R]: net; 13.05, 13.06.2023; N, S; tyrphoneutral.
4.4. Clemnius decoratus (Gyllenhal, 1810) [O]: net; 13.05.2023; S, W; tyrphoneutral.
4.5. Dytiscus dimidiatus Bergsträsser, 1778 [R]: net; 13.05, 13.06, 13.08.2023; S, W; tyrphoneutral.
4.6. Graphoderus cinereus (Linnaeus, 1758) [O]: net; 13.05.2023; SW; tyrphoneutral.
4.7. Graptodytes granularis (Linnaeus, 1767) [O]: net; 13.05, 13.06, 13.10.2023; S; tyrphoneutral.
4.8. Graptodytes pictus (Fabricius, 1787) [F]: net; 13.07, 17.09.2023; C, S; tyrphoneutral.
4.9. Hydaticus seminiger (DeGeer, 1774) [O]: net; 13.05.2023; SW; tyrphoneutral.
4.10. Hydaticus transversalis (Pontoppidan, 1763) [R]: net; 13.06, 13.08.2023; S; tyrphoneutral.
4.11. Hydroporus angustatus Sturm, 1835 [F]: net; 07.04, 13.05, 13.06, 17.09, 13.10.2023; C, S, W; tyrphoneutral.
4.12. Hydroporus dorsalis (Fabricius, 1787) [R]: net; 07.04, 13.07, 17.09, 13.10.2023; S, SE; tyrphoneutral.
4.13. Hydroporus erythrocephalus (Linnaeus, 1758) [O]: net; 13.05, 13.06.2023; S, SW; tyrphoneutral.
4.14. Hydroporus glabriusculus Aubé, 1838 [O]: net; 13.07.2023; S; tyrphoneutral.
4.15. Hydroporus neglectus Schaum, 1845 [F]: net; 07.04, 13.05, 13.10.2023; C, N, S, W; tyrphoneutral.
4.16. Hydroporus obscurus Sturm, 1835 [R]: net; 17.09.2023; S, SW; tyrphoneutral.
4.17. Hydroporus palustris (Linnaeus, 1761) [F]: net; 07.04, 13.05, 13.07, 13.08, 17.09.2023; S, W; tyrphoneutral.
4.18. Hydroporus planus (Fabricius, 1782) [R]: net; 13.06, 13.07, 13.08.2023; S, SE; tyrphoneutral.
4.19. Hygrotus impressopunctatus (Schaller, 1783) [F]: net; 07.04, 13.05.2023; S, SW; tyrphoneutral.
4.20. Hygrotus inaequalis (Fabricius, 1777) [F]: net; 07.04, 13.05, 13.07, 13.08, 17.09.2023; C, S, SW, W; tyrphoneutral.
4.21. Hyphydrus ovatus (Linnaeus, 1761) [A]: net; 13.05, 13.06, 13.07, 13.08, 17.09, 13.10.2023; C, S, W; tyrphoneutral.
4.22. Ilybius ater (DeGeer, 1774) [R]: net; 13.05, 13.07, 13.08.2023; S; tyrphoneutral.
4.23. Ilybius guttiger (Gyllenhal, 1808) [R]: net; 13.05.2023; N; tyrphoneutral.
4.24. Laccophilus minutus (Linnaeus, 1758) [O]: net; 07.04, 13.07, 13.08, 17.09, 13.10.2023; S; tyrphoneutral.
4.25. Porhydrus lineatus (Fabricius, 1775) [A]: net; 13.07, 13.08, 17.09.2023; S; tyrphoneutral.
4.26. Rhantus exsoletus (Forster, 1771) [R]: net; 07.04.2023; S; tyrphoneutral.
4.27. Rhantus suturalis (MacLeay, 1825) [R]: net; 13.05.2023; S; tyrphoneutral.
4.28. Scarodytes halensis (Fabricius, 1787) [R]: net; 13.08, 17.09.2023; S; tyrphoneutral.
5. Carabidae Latreille, 1802 (30 species)
5.1. Acupalpus brunnipes (Sturm, 1825) [A]: pitfall; 07.04-13.05, 13.05-13.06, 13.06-13.07, 13.10-17.11.2023; C, N; tyrphophilous.
5.2. Acupalpus meridianus (Linnaeus, 1761) [R]: pitfall; 13.05-13.06.2023; N; tyrphoneutral.
5.3. Amara famelica Zimmermann, 1832 [F]: pitfall; 07.04-13.05, 13.05-13.06, 13.06-13.07, 13.07-13.08, 13.08-17.09, 13.10-17.11.2023; N, SW; tyrphoneutral.
5.4. Badister bullatus (Schrank, 1798) [R]: pitfall; 13.05-13.06, 13.07-13.08.2023; SW; tyrphoxenous.
5.5. Bembidion lampros (Herbst, 1784) [F]: pitfall; 07.04-13.05.2023; N; tyrphoneutral.
5.6. Bembidion quadrimaculatum (Linnaeus, 1761) [O]: pitfall; 07.04-13.05, 13.08-17.09, 17.09-13.10.2023; C, N; tyrphoneutral.
5.7. Bradycellus rufcollis (Stephens, 1828) [R]: pitfall; 17.09-13.10.2023; C, N; tyrphophilous.
5.8. Carabus nemoralis Muller, 1764 [R]: pitfall; 13.07-13.08, 17.09-13.10.2023; SW; tyrphoxenous.
5.9. Cicindela campestris Linnaeus, 1758 [O]: pitfall, visual; 07.04-13.05, 13.05.2023; E, N, SW, W; tyrphoneutral.
5.10. Cymindis vaporariorum (Linnaeus, 1758) [O]: pitfall; 07.04-13.05, 13.07-13.08.2023; C; tyrphoneutral.
5.11. Demetrias monostigma Samouelle, 1819 [O]: net; 13.06.2023; SW, on Phragmítes; tyrphoneutral.
5.12. Dyschirius globosus (Herbst, 1784) [R]: pitfall; 13.06-13.07, 17.09-13.10.2023; C, N; tyrphoneutral.
5.13. Harpalus tardus (Panzer, 1796) [R]: pitfall; 13.06-13.07.2023; C; tyrphoxenous.
5.14. Harpalus fuliginosus (Duftschmid, 1812) [R]: pitfall; 13.07-13.08.2023; SW; tyrphoxenous.
5.15. Leistus ferrugineus (Linnaeus, 1758) [F]: pitfall; 13.06-13.07, 17.09-13.10.2023; C; tyrphoneutral.
5.16. Leistus rufomarginatus (Duftschmid, 1812) [F]: pitfall; 13.05-13.06, 17.09-13.10, 17.09-13.10.2023, C, SW; tyrphoneutral.
5.17. Leistus terminatus (Hellwig, 1793) [A]: pitfall; 13.05-13.06, 13.06-13.07, 13.07-13.08, 13.08-17.09, 17.09-13.10.2023; N, SW; tyrphoneutral.
5.18. Loricera pilicornis (Fabricius, 1775) [O]: pitfall; 13.05-13.06.2023; SW; tyrphoneutral.
5.19. Microlestes maurus (Sturm, 1827) [R]: pitfall; 07.04-13.05.2023; C; tyrphoneutral.
5.20. Microlestes minutulus (Goeze, 1777) [R]: pitfall; 17.09-13.10.2023; N; tyrphoneutral.
5.21. Nebria brevicollis (Fabricius, 1792) [O]: pitfall; 17.09-13.10.2023; N; tyrphoxenous.
5.22. Notiophilus palustris (Duftschmid, 1812) [O]: pitfall; 07.04-13.05, 13.06-13.07, 13.08-17.09.2023; C, N, SW; tyrphoneutral.
5.23. Olisthopus rotundatus (Paykull, 1790) [R]: pitfall; 13.07-13.08.2023; C; tyrphoneutral.
5.24. Ophonus rufibarbis (Fabricius, 1792) [R]: pitfall; 13.07-13.08.2023; N; tyrphoxenous.
5.25. Philorhizus sigma (Rossi, 1790) [R]: pitfall; 13.05-13.06.2023; N; tyrphoneutral.
5.26. Poecilus cupreus (Linnaeus, 1758) [F]: pitfall; 07.04-13.05, 13.05-13.06, 13.08-17.09.2023; C, N, SW; tyrphoneutral.
5.27. Pseudoophonus rufpes (DeGeer, 1774) [R]: pitfall; 13.05-13.06.2023; C; tyrphoxenous.
5.28. Pterostichus anthracinus (Illiger, 1798) [O]: pitfall; 07.04-13.05, 13.05-13.06, 17.09-13.10, 13.10-17.11.2023; C, N; tyrphoneutral.
5.29. Pterostichus crenatus (Duftschmid, 1812) [R]: pitfall; 13.08-17.09.2023; C; tyrphoneutral.
5.30. Pterostichus niger (Schaller, 1783) [A]: pitfall; 13.05-13.06, 13.07-13.08, 13.08-17.09, 17.09-13.10.2023; C, N, SW; tyrphoneutral.
6. Scirtidae Fleming, 1821 (5 species)
6.1. Contacyphon ochraceus (Stephens, 1830) [O]: net; 13.06, 13.07.2023; C; tyrphoneutral.
6.2. Contacyphon padi (Linnaeus, 1758) [F]: net; 13.06, 13.08, 13.10.2023; C, S, SW; tyrphoneutral.
6.3. Contacyphon variabilis (Thunberg, 1785) [F]: net, pitfall; 13.06, 13.07, 13.06-13.07.2023; C, S, SW; tyrphophilous.
6.4. Microcara testacea (Linnaeus, 1767) [O]: net, pitfall; 13.06, 13.06-13.07.2023; S, SW; tyrphoneutral.
6.5. Scirtes hemisphaericus (Linnaeus, 1758) [F]: net; 13.06, 13.07.2023; S, SW; tyrphoneutral.
7. Helophoridae Leach, 1815 (3 species)
7.1. Helophorus aquaticus (Linnaeus, 1758) [F]: net; 13.06.2023; S; tyrphoneutral.
7.2. Helophorus granularis (Linnaeus, 1761) [O]: net; 07.04, 13.05, 13.10.2023; N, E, S; tyrphoneutral.
7.3. Helophorus flavipes Fabricius, 1792 [F]: net; 13.06, 13.07.2023; S; tyrphoneutral.
8. Hydrochidae Thomson, 1859 (1 species)
8.1. Hydrochus elongatus (Schaller, 1783) [R]: net; 13.08.2023; S; tyrphophilous.
9. Hydrophilidae Latreille, 1802 (9 species)
9.1. Anacaena lutescens (Stephens, 1829) [A]: net; 07.04, 13.05, 13.08, 17.09.2023; C, N, S; tyrphoneutral.
9.2. Cymbiodyta marginella (Fabricius, 1792) [R]: net; 13.10.2023; E; ; tyrphoneutral.
9.3. Enochrus coarctatus (Gredler, 1863) [O]: net; 13.05.2023; C, N, S, W; tyrphoneutral.
9.4. Enochrus ochropterus (Marsham, 1802) [O]: net; 13.06.2023; S; tyrphophilous.
9.5. Enochrus testaceus (Fabricius, 1801) [R]: net; 13.05.2023; S; tyrphoneutral.
9.6. Helochares obscurus (Müller, 1776) [O]: net; 13.05, 13.06.2023; S, W; tyrphoneutral.
9.7. Hydrobius fuscipes (Linnaeus, 1758) [R]: net; 13.05.2023; E; tyrphoneutral.
9.8. Hydrochara caraboides (Linnaeus, 1758) [R]: net; 13.05.2023; E; tyrphoxenous.
9.9. Laccobius minutus (Linnaeus, 1758) [R]: net; 17.09, 13.10.2023; N, S; tyrphoneutral.
10. Hydraenidae Mulsant, 1844 (2 species)
10.1. Limnebius crinifer Rey, 1885 [R]: net; 13.05.2023; S; tyrphoneutral.
10.2. Ochthebius minimus (Fabricius, 1792) [R]: net; 13.05.2023; S; tyrphoneutral.
11. Leiodidae Fleming, 1821 (4 species)
11.1. Agathidium plagiatum (Gyllenhal, 1810) [R]: pitfall; 13.06-13.07.2023; N; tyrphoneutral.
11.2. Apocatops nigrita (Erichson, 1837) [A]: pitfall; 07.04-13.05, 13.05-13.06, 13.06-13.07, 13.07-13.08.2023; N, SW; tyrphoneutral.
11.3. Catops fuscus (Panzer 1794) [R]: pitfall; 13.10-17.11.2023; N; tyrphoneutral.
11.4. Sciodrepoides watsoni (Spence, 1813) [A]: pitfall; 13.06-13.07, 13.07-13.08, 17.09-13.10.2023; N, SW; tyrphoneutral.
12. Silphidae Latreille, 1807 (5 species)
12.1. Nicrophorus humator (Gleditsch, 1767) [R]: pitfall; 13.06-13.07.2023; C; tyrphoxenous.
12.2. Nicrophorus vespillo (Linnaeus, 1758) [O]: pitfall; 13.06-13.07, 13.08-17.09.2023; C, N; tyrphoneutral.
12.3. Nicrophorus vespilloides Herbst, 1783 [A]: pitfall; 13.06-13.07, 13.07-13.08, 13.08-17.09.2023; C, N, SW; tyrphoneutral.
12.4. Oiceoptoma thoracica (Linnaeus, 1758) [O]: pitfall; 13.07-13.08, 13.08-17.09.2023; N, SW; tyrphoneutral.
12.5. Phosphuga atrata (Linnaeus, 1758) [R]: pitfall; 07.04-13.05.2023; N; tyrphoxenous.
13. Staphylinidae Latreille, 1802 (45 species)
13.1. Acidota crenata (Fabricius, 1793) [F]: pitfall; 07.04-13.05, 17.09-13.10, 13.10-17.11.2023; C, N; tyrphoneutral.
13.2. Aleochara curtula (Goeze, 1777) [R]: visual; 13.05.2023; E; tyrphoneutral.
13.3. Anthobium unicolor (Marsham, 1802) [A]: pitfall, visual; 07.04-13.05, 13.05-13.06, 13.06-13.07, 13.10, 17.09-13.10, 13.10-17.11, 17.11.2023; C, N, S, SW; tyrphoneutral.
13.4. Bolitochara pulchra (Gravenhorst, 1806) [O]: pitfall; 17.09-13.10.2023; C; tyrphoneutral.
13.5. Brachygluta fossulata (Reichenbach, 1816) [R]: pitfall; 13.05-13.06.2023; SW; tyrphoneutral.
13.6. Brachygluta haematica (Reichenbach, 1816) [O]: pitfall; 13.05-13.06, 13.06-13.07, 13.08-17.09.2023; N, SW; tyrphophilous.
13.7. Carphacis striatus (Olivier, 1795) [R]: visual; 13.07.2023; SW, on rotten Boletaceae; tyrphoneutral.
13.8. Drusilla canaliculata (Fabricius, 1787) [F]: pitfall; 13.05-13.06, 13.06-13.07, 13.08-17.09, 17.09-13.10.2023; C, N, SW; tyrphoneutral.
13.9. Euconnus claviger (Müller & Kunze, 1822) [O]: pitfall; 07.04-13.05.2023; C; tyrphoneutral.
13.10. Euconnus wetterhallii (Gyllenhal, 1813) [O]: pitfall; 07.04-13.05, 13.06-13.07.2023; C, SW; tyrphoneutral.
13.11. Eusphalerum minutum (Fabricius, 1792) [R]: visual; 13.06.2023; W; tyrphoneutral.
13.12. Gyrophaena joyi Wendeler, 1924 [F]: visual; 13.08.2023; SW, on Basidiomycota; tyrphoneutral.
13.13. Habrocerus capillaricornis (Gravenhorst, 1806) [R]: visual; 17.11.2023; S, in rotten wood; tyrphoneutral.
13.14. Hygronoma dimidiata (Gravenhorst, 1806) [R]: visual; 13.08.2023; SW, ditch shore; tyrphoneutral.
13.15. Ischnosoma longicorne (Mäklin, 1847) [F]: pitfall; 07.04-13.05, 13.05-13.06, 13.08.-17.09, 13.10-17.11.2023; C, N; tyrphophilous.
13.16. Lesteva longoelytrata (Goeze, 1777) [O]: visual, pitfall; 13.05, 13.05-13.06.2023; E, SW, W, near open water; tyrphoneutral.
13.17. Lordithon thoracicus (Fabricius, 1777) [R]: pitfall; 13.08-17.09.2023; C; tyrphoxenous.
13.18. Mycetoporus lepidus (Gravenhorst, 1806) [O]: pitfall; 07.04-13.05, 13.07-13.08, 13.10-17.11.2023; C, N; tyrphoneutral.
13.19. Ocypus brunnipes (Fabricius, 1781) [R]: pitfall; 17.09-13.10.2023; C; tyrphoxenous.
13.20. Olophrum piceum (Gyllenhal, 1810) [F]: pitfall; 07.04-13.05, 13.10-17.11.2023; C, N; tyrphoneutral.
13.21. Omalium rivulare (Paykull, 1789) [R]: visual; 17.11.2023; S; tyrphoneutral.
13.22. Oxypoda opaca (Gravenhorst, 1802) [F]: pitfall, visual; 13.07, 13.06-13.07, 13.08, 13.08-17.09, 13.10-17.11.2023; C, N, SW; tyrphoneutral.
13.23. Oxyporus rufus (Linnaeus, 1758) [O]: visual; 13.07.2023; SW, on rotten Boletaceae; tyrphoneutral.
13.24. Parabolitobius inclinans (Gravenhorst, 1806) [F]: pitfall; 07.04-13.05.2023; C; tyrphoneutral.
13.25. Philonthus cognatus Stephens, 1832 [F]: pitfall, visual; 07.04-13.05, 13.05-13.06, 13.08, 13.07-13.08.2023; C, E, N; tyrphoneutral.
13.26. Philonthus decorus (Gravenhorst, 1802) [R]: pitfall; 17.09-13.10.2023; SW; tyrphoneutral.
13.27. Philonthus splendens (Fabricius, 1793) [R]: pitfall; 13.07-13.08.2023; SW; tyrphoneutral.
13.28. Platydracus fulvipes (Scopoli, 1763) [O]: pitfall; 13.05-13.06, 13.06-13.07, 13.07-13.08.2023; N, SW; tyrphophilous.
13.29. Platydracus latebricola (Gravenhorst, 1806) [F]: pitfall; 13.05-13.06, 13.06-13.07, 13.07-13.08.2023; N, SW; tyrphophilous.
13.30. Proteinus brachypterus (Fabricius, 1792) [O]: pitfall; 07.04, 13.06-13.07, 17.11.2023; S, SW, W, on dung and rotten mushrooms; tyrphoneutral.
13.31. Pselaphus heisei Herbst, 1792 [R]: pitfall; 07.04-13.05.2023; C; tyrphophilous.
13.32. Sepedophilus testaceus (Fabricius, 1793) [O]: pitfall; 13.06-13.07, 13.07-13.08, 17.09-13.10.2023; C, N; tyrphoneutral.
13.33. Stenichnus scutellaris (Müller & Kunze, 1822) [R]: pitfall; 13.05-13.06, 13.06-13.07.2023; C, SW; tyrphoneutral.
13.34. Stenus bimaculatus Gyllenhal, 1810 [R]: visual; 13.05.2023; E, near open water; tyrphoneutral.
13.35. Stenus clavicornis (Scopoli, 1763) [R]: pitfall; 13.08-17.09.2023; N; tyrphoneutral.
13.36. Stenus solutus Erichson, 1840 [R]: pitfall, visual; 07.04-13.05, 13.05, 13.05-13.06, 17.09-13.10, 13.10, 13.10-17.11.2023; C, N, SW; tyrphoneutral.
13.37. Tachinus corticinus Gravenhorst, 1802 [R]: net, visual; 13.06, 13.08.2023; C, E; tyrphoneutral.
13.38. Tachyporus dispar (Paykull, 1789) [A]: pitfall; 13.10-17.11.2023; C, N; tyrphoneutral.
13.39. Tachyporus hypnorum (Fabricius, 1775) [F]: pitfall, visual; 07.04-13.05, 13.10, 13.10-17.11.2023; C, N; tyrphoneutral.
13.40. Tachyporus nitidulus (Fabricius, 1781) [R]: pitfall; 17.09-13.10.2023; C, N; tyrphoneutral.
13.41. Xantholinus longiventris Heer, 1839 [A]: pitfall; 07.04-13.05, 13.05-13.06, 13.06-13.07, 13.07-13.08, 13.08-17.09, 17.09-13.10, 13.10-17.11.2023; C, N, SW; tyrphoneutral.
13.42. Zyras collaris (Paykull, 1800) [O]: pitfall; 13.08-17.09.2023; C, N; tyrphoneutral.
13.43. Zyras lugens (Gravenhorst, 1802) [F]: pitfall; 07.04-13.05, 13.07-13.08.2023; C; tyrphoneutral.
14. Lucanidae Latreille, 1804 (1 species)
14.1. Dorcus parallelepipedus (Linnaeus, 1758) [R]: visual; 13.06.2023; N, on central road; tyrphoxenous.
15. Scarabaeidae Latreille, 1802 (6 species)
15.1. Agrilinus rufus (Moll, 1782) [O]: visual; 13.08.2023; E, in dung; tyrphoxenous.
15.2. Aphodius fimetarius (Linnaeus, 1758) [R]: visual; 13.08.2023; E, in dung; tyrphoxenous.
15.3. Melolontha melolontha (Linnaeus, 1758) [O]: visual; 13.05, 13.06.2023; S, on Betula; tyrphoneutral.
15.4. Oxythyrea funesta (Poda, 1761) [R]: visual; 13.07.2023; N; tyrphoxenous.
15.5. Serica brunnea (Linnaeus, 1758) [F]: pitfall; 13.06-13.07, 13.07-13.08.2023; N, SW; tyrphoneutral.
15.6. Trichius fasciatus (Linnaeus, 1758) [R]: visual; 13.07.2023; N; tyrphoxenous.
16. Buprestidae Leach, 1815 (1 species)
16.1. Trachys minutus (Linnaeus, 1758) [O]: visual, net; 13.06, 13.07, 13.08.2023; C, S, on Salix; tyrphoneutral.
17. Byrrhidae Latreille, 1804 (2 species)
17.1. Curimopsis nigrita (Palm, 1934) [A]: pitfall; 07.04-13.05, 13.05-13.06, 13.06-13.07.2023; C, N; tyrphophilous.
17.2. Cytilus sericeus (Forster, 1771) [R]: pitfall; 13.06-13.07.2023; SW; tyrphoxenous.
18. Throscidae Laporte, 1840 (2 species)
18.1. Aulonothroscus brevicollis (Bonvouloir, 1859) [R]: pitfall; 17.09-13.10.2023; C; tyrphoneutral.
18.2. Trixagus dermestoides (Linnaeus, 1767) [R]: pitfall; 13.05-13.06, 13.06-13.07, 13.07-13.08.2023; C, SW; tyrphoneutral.
19. Elateridae Leach, 1815 (11 species)
19.1. Adrastus pallens (Fabricius, 1792) [F]: net; 13.07.2023; S; tyrphoneutral.
19.2. Agriotes ustulatus (Schaller, 1783) [R]: pitfall; 13.05-13.06.2023; N, SW; tyrphoneutral.
19.3. Agrypnus murinus (Linnaeus, 1758) [R]: net; 13.05, 13.06.2023; C, S; tyrphoneutral.
19.4. Ampedus balteatus (Linnaeus, 1758) [F]: net, visual, pitfall; 13.05, 13.06, 13.06-13.07.2023; C, E, N, S, SW; tyrphoneutral.
19.5. Ampedus pomorum (Herbst. 1784) [O]: net, visual; 13.05, 13.05-13, 13.06-13.07.2023; C, N; tyrphoneutral.
19.6. Ampedus sanguineus (Linnaeus, 1758) [R]: visual; 13.06.2023; S; tyrphoneutral.
19.7. Cidnopus aeruginosus (Olivier, 1790) [F]: net; 13.05.2023; C, N, S, W; tyrphoneutral.
19.8. Dalopius marginatus (Linnaeus, 1758) [R]: net; 13.05-13.06, 13.06-13.07.2023; N, SW; tyrphoneutral.
19.9. Hemicrepidius niger (Linnaeus, 1758) [O]: net; 13.06.2023; S; tyrphoxenous.
19.10. Melanotus villosus (Geoffroy, 1785) [R]: net; 13.05.2023; E; tyrphoneutral.
19.11. Sericus brunneus (Linnaeus, 1758) [O]: net; 13.05.2023; C, N; tyrphoneutral.
20. Cantharidae Imhoff, 1856 (1815) (13 species)
20.1. Cantharis flavilabris Fallén, 1807 [F]: net; 13.06, 13.07.2023; S, SW; tyrphoneutral.
20.2. Cantharis livida Linnaeus, 1758 [O]: net; 13.06.2023; C, along central road; tyrphoxenous.
20.3. Cantharis nigricans (Müller, 1776) [O]: net; 13.06.2023; C; tyrphoneutral.
20.4. Cantharis pallida Goeze, 1777 [O]: net; 13.06.2023; W; tyrphoneutral.
20.5. Cantharis pellucida Fabricius, 1792 [O]: net; 13.06.2023; C; tyrphoneutral.
20.6. Cantharis rufa Linnaeus, 1758 [O]: net; 13.06.2023; SW; tyrphoxenous.
20.7. Malthodes marginatus (Latreille, 1806) [R]: net; 13.06, 13.07.2023; S, SW; tyrphoneutral.
20.8. Malthodes minimus (Linnaeus, 1758) [R]: net; 13.06.2023; SW; tyrphoneutral.
20.9. Malthodes pumilus (Brébisson, 1835) [O]: net; 13.06.2023; SW, on Calluna; tyrphophilous.
20.10. Rhagonycha fulva (Scopoli, 1763) [A]: net; 13.07.2023; C, N, S; tyrphoxenous.
20.11. Rhagonycha lignosa (Müller, 1764) [O]: net; 13.06.2023; SW; tyrphoneutral.
20.12. Rhagonycha nigriventris Motschulsky, 1860 [R]: net; 13.05.2023; C; tyrphoneutral.
20.13. Rhagonycha testacea (Linnaeus, 1758) [O]: net; 13.06.2023; SW; tyrphoxenous.
21. Dermestidae Latreille, 1804 (1 species)
21.1. Dermestes murinus Linnaeus, 1758 [F]: pitfall; 13.06-13.07, 13.08-17.09.2023; C, N; tyrphoneutral.
22. Latridiidae Erichson, 1842 (1 species)
22.1. Melanophthalma transversalis (Gyllenhal, 1827) [O]: net; 13.06, 13.08.2023; C, along central road; tyrphoxenous.
23. Endomychidae Leach, 1815 (1 species)
23.1. Endomychus coccineus (Linnaeus, 1758) [O]: visual, net; 07.04, 13.06.2023; S, E, larvae on a birch stump with Stereum hirsutum; tyrphoneutral.
24. Coccinellidae Latreille, 1807 (10 species)
24.1. Chilocorus renipustulatus (Scriba, 1791) [O]: net; 13.07.2023; S, SW; tyrphoneutral.
24.2. Coccinella hieroglyphica Linnaeus, 1758 [O]: pitfall; 13.06-13.07.2023; N; tyrphoneutral.
24.3. Coccinella septempunctata Linnaeus, 1758 [F]: net, pitfall; 13.06, 13.07, 17.09-13.10.2023; C, N; tyrphoneutral.
24.4. Halyzia sedecimguttata (Linnaeus, 1758) [R]: net; 13.07.2023; SW; tyrphoneutral.
24.5. Harmonia axyridis (Pallas, 1773) [F]: net, visual; 13.06 [larva], 13.07, 13.08.2023; C, along road; tyrphoxenous.
24.6. Propylea quatuordecimpunctata (Linnaeus, 1758) [F]: net; 13.05, 13.06, 13.07, 13.08.2023; C, E, N, SW, W; tyrphoneutral.
24.7. Psyllobora vigintiduopunctata (Linnaeus, 1758) [F]: net; 13.05, 13.07, 17.09.2023; C, S, SW; tyrphoneutral.
24.8. Scymnus suturalis Thunberg, 1795 [O]: net, pitfall; 13.05-13.06, 13.06, 13.06-13.07, 13.07, 13.08, 13.08-17.09, 17.09-13.10.2023; C, N, SW; tyrphophilous.
24.9. Stethorus punctillum (Weise, 1891) [R]: net; 13.08.2023; C; tyrphoxenous.
24.10. Tytthaspis sedecimpunctata (Linnaeus, 1761) [O]: net; 13.07, 13.08.2023; C; tyrphoneutral.
25. Mordellidae Latreille, 1802 (3 species)
25.1. Mordella aculeata Linnaeus, 1758 [F]: net, pitfall; 13.06-13.07, 13.08, 13.08-17.09.2023; C, N; tyrphoneutral.
25.2. Mordellistena koelleri Ermisch, 1956 [R]: net; 13.06.2023; C; tyrphoneutral.
25.3. Variimorda villosa (Schrank, 1781) [F]: net; 13.07.2023; C, along central road on Apiaceae; tyrphoneutral.
26. Tenebrionidae Latreille, 1802 (2 species)
26.1. Isomira murina (Linnaeus, 1758) [R]: net; 13.06.2023; C; tyrphoneutral.
26.2. Lagria hirta (Linnaeus, 1758) [A]: net; 13.07.2023; C, N, S; tyrphoneutral.
27. Oedemeridae Latreille, 1810 (4 species)
27.1. Anogcodes melanurus (Fabricius, 1787) [R]: net; 13.07.2023; N; tyrphoxenous.
27.2. Oedemera croceicollis (Gyllenhal, 1827) [O]: net; 13.06.2023; SW; tyrphoneutral.
27.3. Oedemera lurida (Marsham, 1802) [F]: net; 13.07.2023; C, N, S; tyrphoneutral.
27.4. Oedemera virescens (Linnaeus, 1767) [R]: net; 13.05.2023; S; tyrphoxenous.
28. Pyrochroidae Latreille, 1806 (1 species)
28.1. Schizotus pectinicornis (Linnaeus, 1758) [O]: visual; 13.05 (adult), 17.11.2023 (larva); S, larvae under bark of birch stump; tyrphoneutral.
29. Scraptiidae Mulsant, 1856/Gistel, 1856 (1 species)
29.1. Anaspis frontalis (Linnaeus, 1758) [O]: net; 13.06.2023; C, S; tyrphoneutral.
30. Byturidae Jacquelin du Val, 1858 (1 species)
30.1. Byturus ochraceus (Scriba, 1790) [O]: net; 13.05.2023; E; tyrphoxenous.
31. Melyridae Leach, 1815 (3 species)
31.1. Cordylepherus viridis (Fabricius, 1787) [O]: net; 13.06, 13.07.2023; C, S; tyrphoneutral.
31.2. Dasytes niger (Linnaeus, 1761) [R]: pitfall; 13.07-13.08.2023; N; tyrphoneutral.
31.3. Dasytes plumbeus (Müller, 1776) [F]: pitfall, net; 13.06-13.07, 13.07; C, N, SW; tyrphoneutral.
32. Cryptophagidae Kirby, 1837 (2 species)
32.1. Cryptophagus lycoperdi (Scopoli, 1763) [A]: visual, pitfall; 13.08, 13.08-17.09.2023; N, SW, W, associated with Scleroderma citrinum; tyrphoneutral.
32.2. Telmatophilus typhae (Fallén, 1802) [F]: net; 13.07.2023; C, S, associated with Typha latifolia; tyrphoneutral.
33. Phalacridae Leach, 1815 (1 species)
33.1. Phalacrus championi Guillebeau, 1892 [R]: net; 13.06, 13.07.2023; C, W; tyrphoneutral.
34. Kateretidae Kirby, 1837 (1 species)
34.1. Brachypterus urticae (Fabricius, 1792) [A]: net; 13.06, 13.07.2023; C, on Urtica dioica; tyrphoxenous.
35. Nitidulidae Latreille, 1802 (4 species)
35.1. Brassicogethes aeneus (Fabricius, 1775) [O]: net; 13.06.2023; S, along central road; tyrphoneutral.
35.2. Brassicogethes viridescens (Fabricius, 1787) [O]: net; 13.07.2023; C, along central road; tyrphoneutral.
35.3. Meligethes flavimanus Stephens, 1830 [R]: pitfall; 13.06-13.07.2023; N; tyrphoxenous.
35.4. Pocadius ferrugineus (Fabricius, 1775) [O]: pitfall, visual; 13.06-13.07, 13.07.2023; SW; tyrphoneutral.
36. Cerambycidae Latreille, 1802 (8 species)
36.1. Agapanthia villosoviridescens (DeGeer, 1775) [O]: net; 13.06, 13.07.2023; S; tyrphoxenous.
36.2. Lamia textor (Linnaeus, 1758) [R]: visual; 13.07.2023; N; tyrphoneutral.
36.3. Leptura quadrifasciata Linnaeus, 1758 [O]: visual, net; 13.07.2023; C, S, along central road on Apiaceae; tyrphoneutral.
36.4. Stenurella melanura (Linnaeus, 1758) [O]: visual; 13.07.2023; N, along central road on Apiaceae; tyrphoneutral.
36.5. Stenurella nigra (Linnaeus, 1758) [O]: visual; 13.06.2023; S, along central road on Apiaceae; tyrphoneutral.
36.6. Stictoleptura maculicornis (De Geer, 1775) [R]: net; 13.07.2023; C, along central road on Apiaceae; tyrphoneutral.
36.7. Stictoleptura rubra (Linnaeus, 1758) [O]: visual; 13.07, 13.08.2023; N, S; tyrphoneutral.
36.8. Strangalia attenuata (Linnaeus, 1758) [R]: net; 13.07.2023; C, along central road on Apiaceae; tyrphoneutral.
37. Megalopodidae Latreille, 1802 (1 species)
37.1. Zeugophora subspinosa (Fabricius, 1781) [R]: net; 13.06.2023; E, on Populus tremulae; tyrphoneutral.
38. Chrysomelidae Latreille, 1802 (24 species)
38.1. Agelastica alni (Linnaeus, 1758) [O]: net, pitafall; 13.05, 13.05-13.06.2023; N; tyrphoneutral.
38.2. Altica aenescens Weise, 1888 [A]: visual, net; 13.05, 13.06, 13.07, 13.08.2023; C, E, N, W, on leaves of Betula pubescens; tyrphoneutral.
38.3. Altica chamaenerii (Lindberg, 1926) [O]: net, pitfall; 13.05, 13.08-17.09.2023; C, N, on leaves of Chamaenerion angustifolium; tyrphoneutral.
38.4. Altica longicollis (Allard, 1860) [A]: pitfall; 07.04-13.05, 13.05-13.06, 13.06-13.07, 13.07-13.08.2023; C, N, in Calluna-communities; tyrphophilous.
38.5. Batophila rubi (Paykull, 1799) [F]: net; 13.05, 13.06, 13.06-13.07, 13.07.2023; C, N, along road on Rubus idaeus; tyrphoxenous.
38.6. Bromius obscurus (Linnaeus, 1758) [F]: visual, net; 13.05.2023; C, N, along central road on Chamaenerion angustifolium; tyrphoneutral.
38.7. Cassida denticollis Suffrian, 1844 [O]: net; 13.07.2023; S, along central road on Achillea millefolium; tyrphoxenous.
38.8. Cassida rubiginosa Müller, 1776 [F]: net; 13.05, 13.08, 17.09.2023; C, N, S, on Cirsium arvense; tyrphoxenous.
38.9. Cassida viridis Linnaeus, 1758 [R]: pitfall; 13.05-13.06.2023; N; tyrphoneutral.
38.10. Chrysomela populi Linnaeus, 1758 [R]: net; 13.05.2023; S, on regrowth of Populus tremulae; tyrphoneutral.
38.11. Crepidodera aurata (Marsham, 1802) [F]: net; 13.05, 13.07.2023; C, S, on leaves of Salix sp.; tyrphoneutral.
38.12. Cryptocephalus decemmaculatus (Linnaeus, 1758) [O]: net; 13.06.2023; C, on leaves of Salix sp.; tyrphoneutral
38.13. Cryptocephalus ocellatus Drapiez, 1819 [O]: net; 13.06, 13.07.2023; C, on leaves of Salix sp.; tyrphoneutral
38.14. Cryptocephalus pusillus Fabricius, 1777 [R]: pitfall, net; 13.06-13.07, 13.07.2023; C, N.; tyrphoneutral
38.15. Galeruca tanaceti (Linnaeus, 1758) [R]: net; 13.08.2023; N, central road; tyrphoxenous.
38.16. Lochmaea caprea (Linnaeus, 1758) [A]: visual, net, pitfall; 13.05, 13.05-13.06, 13.06, 13.06-13.07, 13.08, 13.08-17.09.2023; C, N, S, SW, on Salix and Betula; tyrphoneutral.
38.17. Longitarsus jacobaeae (Waterhouse, 1858) [R]: net; 13.08.2023; C, along central road; tyrphoxenous.
38.18. Neocrepidodera transversa (Marsham, 1802) [A]: net; 13.06, 13.07, 13.08.2023; C, N, S, on Cirsium arvense; tyrphoxenous.
38.19. Oulema obscura (Stephens, 1831) [R]: net; 13.05, 13.07.2023; C, N, central road; tyrphoneutral.
38.20. Oulema melanopus (Linnaeus, 1758) [R]: net; 13.07.2023; N; tyrphoxenous.
38.21. Pachybrachis hieroglyphicus (Laicharting, 1781) [R]: net; 13.06.2023; C, on leaves of Salix sp.; tyrphoneutral.
38.22. Phyllotreta atra (Fabricius, 1775) [R]: net; 13.07, 17.09.2023; C, SW; tyrphoneutral.
38.23. Phyllotreta striolata (Illiger, 1803) [R]: net; 13.07, 13.08.2023; C, SW; tyrphoneutral.
38.24. Psylliodes chalcomerus (Illiger, 1807) [F]: net, visual; 13.05, 13.07.2023; C, N, along road on Carduus; tyrphoxenous.
39. Attelabidae Billberg, 1820 (3 species)
39.1. Byctiscus betulae (Linnaeus, 1758) [R]: net; 13.05.2023; C, on Salix sp. ; tyrphoneutral.
39.2. Byctiscus populi (Linnaeus, 1758) [O]: net; 13.06.2023; E, on regrowth of Populus trmulae; tyrphoneutral.
39.3. Temnocerus coeruleus (Fabricius, 1798) [O]: net; 13.06.2023; C, along road on Salix sp.; tyrphoneutral.
40. Brentidae Billberg, 1820 (6 species)
40.1. Betulapion simile (Kirby, 1811) [R]: net; 13.06.2023; S, on Betula sp.; tyrphoneutral.
40.2. Ceratapion onopordi (Kirby, 1808) [R]: net; 13.05, 13.10.2023; C, along road on Carduus crispus; tyrphoxenous.
40.3. Eutrichapion ervi (Kirby, 1808) [R]: net; 13.06.2023; C, along central road on Lathyrus pratensis; tyrphoxenous.
40.4. Nanophyes marmoratus (Goeze, 1777) [O]: net; 13.07.2023; C, on Lythrum salicaria near open water; tyrphoneutral.
40.5. Oxystoma cerdo (Gerstaecker, 1854) [R]: net; 13.05, 13.07.2023; C, along road on Vicia cracca; tyrphoxenous.
40.6. Protapion fulvipes (Geoffroy, 1785) [O]: net; 13.06.2023; C, along road on Trifolium repens; tyrphoxenous.
41. Curculionidae Latreille, 1802 (30 species)
41.1. Anthonomus rubi (Herbst, 1795) [O]: net; 13.06, 13.08.2023; C, along road on Rubus idaeus; tyrphoxenous.
41.2. Archarius salicivorus (Paykull, 1792) [R]: net; 13.06.2023; C, along central road on Salix sp.; tyrphoneutral.
41.3. Barypeithes pellucidus (Boheman, 1834) [A]: pitfall; 13.06-13.07.2023; SW; tyrphoxenous.
41.4. Brachysomus echinatus (Bonsdorff, 1785) [A]: net; 13.05-13.06, 13.06-13.07.2023; SW; tyrphoneutral.
41.5. Calosirus terminatus (Herbst, 1795) [O]: net; 13.07, 13.10.2023; C, N, along road; tyrphoxenous.
41.6. Ceutorhynchus obstrictus (Marsham, 1802) [O]: net; 13.07.2023; N, along road; tyrphoxenous.
41.7. Cleonis pigra (Scopoli, 1763) [O]: visual, pitfall; 13.05, 13.05-13.06.2023; N; tyrphoxenous.
41.8. Cryptorhynchus lapathi (Linnaeus, 1758) [R]: net; 13.06.2023; C; tyrphoneutral.
41.9. Hypera arator (Linnaeus, 1758) [R]: pitfall; 07.04-13.05.2023; N; tyrphoxenous.
41.10. Hypera nigrirostris (Fabricius, 1775) [R]: pitfall; 13.06-13.07.2023; SW; tyrphoxenous.
41.11. Hypera postica (Gyllenhal, 1813) [R]: net; 13.08.2023; S; tyrphoxenous.
41.12. Larinus turbinatus Gyllenhal, 1835 [R]: net; 13.06.2023; N, along road on Carduus; tyrphoxenous.
41.13. Micrelus ericae (Gyllenhal, 1813) [R]: pitfall; 13.06-13.07.2023; N, in Calluna community; tyrphophilous.
41.14. Nedyus quadrimaculatus (Linnaeus, 1758) [O]: net; 13.05.2023; S, on Urtica dioica; tyrphoxenous.
41.15. Orchestes jota (Fabricius, 1787) [F]: net; 13.05, 13.06, 13.08.2023; C, N, S, W, on Betula; tyrphoneutral.
41.16. Otiorhynchus ovatus (Linnaeus, 1758) [O]: pitfall; 13.07-13.08, 17.09-13.10.2023; C, N; tyrphoxenous.
41.17. Otiorhynchus tristis (Scopoli, 1763) [R]: net; 13.07.2023; N; tyrphoxenous.
41.18. Phyllobius argentatus (Linnaeus, 1758) [R]: net; 13.06.2023; C; tyrphoneutral.
41.19. Phyllobius pomaceus Gyllenhal, 1834 [F]: net; 13.06.2023; C, S, along road on Urtica dioica; tyrphoxenous.
41.20. Phyllobius pyri (Linnaeus, 1758) [R]: net; 13.05.2023; S; tyrphoneutral.
41.21. Polydrusus cervinus (Linnaeus, 1758) [F]: net, pitfall; 13.05, 13.05-13.06, 13.06, 13.06-13.07, 13.07.2023; C, E, N, S, SW, on Betula; tyrphoneutral.
41.22. Polydrusus picus (Fabricius, 1792) [A]: net, visual; 13.06, 13.07.2023; C, S, SW, on Salix sp.; tyrphoneutral.
41.23. Rhamphus pulicarius (Herbst, 1795) [O]: net, visual; 13.05, 13.06.2023; C, SW, on Salix sp.; tyrphoneutral.
41.24. Rhinocyllus conicus (Frölich, 1792) [F]: net; 13.05, 13.06, 13.07.2023; C, N, along road on Carduus; tyrphoxenous.
41.25. Rhinusa linariae (Panzer, 1793) [R]: net; 13.07.2023; C, along road on Linaria vulgaris; tyrphoxenous.
41.26. Rutidosoma graminosum (Gistel, 1857) [R]: net; 13.06.2023; SW; tyrphoneutral.
41.27. Sciaphilus asperatus (Bonsdorff, 1785) [F]: pitfall; 13.06-13.07, 13.07-13.08.2023; SW; tyrphoneutral.
41.28. Sitona hispidulus (Fabricius, 1777) [O]: net; 13.08.2023; S; tyrphoneutral.
41.29. Sitona lineatus (Linnaeus, 1758) [R]: pitfall; 07.04-13.05.2023; N; tyrphoxenous.
41.30. Tanymecus palliatus (Fabricius, 1787) [F]: net; 13.06.2023; C, along central road; tyrphoxenous.
References
- Frolking, S.; Talbot, J.; Jones, M.; Treat, C.C.; Kauffman, J.B.; Tuittila, E.S.; Roulet, N.T. Peatlands in the Earth's 21st century climate system. Environmental Reviews 2011, 19, 371–396. [CrossRef]
- Kreyling, J.; Tanneberger, F.; Jansen, F.; Linden, van der S.; Aggenbach, C.; Blüml, V.; Couwenberg, J.; Emsens, W-J.; Joosten, H.; Klimkowska, A.; Kotowski, W.; Kozub, L.; Lennartz, B.; Liczner, Y.; Liu, H.; Michaelis, D.; Oehmke, C.; Parakenings, K.; Pleyl, E.; Poyda, A.; Raabe, S.; Röhl, M.; Rücker, K.; Schneider, A.; Schrautzer, J.; Schröder, C.; Schug, F.; Seeber, E.; Thiel, F.; Thiele, S.; Tiemeyer, B.; Timmermann, T.; Urich, T.; Diggelen, van R.; Vegelin, K.; Verbruggen, E.; Wilmking, M.; Wrage-Mönnig, N.; Wołejko, L.; Zak, D.; Jurasinski, G. Rewetting does not return drained fen peatlands to their old selves. Nature communications 2021, 12 (5693). [CrossRef]
- Duinen, van G.-J.; Brock, A.M.T.; Kuper, J.T.; Leuven, R.S.E.W.; Peeters, T.M.J.; Roelofs, J.G.M.; Velde, van der G.; Verberk, W.C.E.P.; Esselink, H. Do restoration measures rehabilitate fauna diversity in raised bogs? A comparative study on aquatic macroinvertebrates. Wetlands Ecology and Management 2003, 11, 447–459. [CrossRef]
- Verberk, W.C.E.P.; Kuper, J.T.; Duinen, van G.-J.; Esselink, H. Changes in macroinvertebrate richness and diversity following large scale rewetting measures in a heterogeneous bog landscape. Proceedings of the Netherlands Entomological Society meeting 2006, 17, 27–36.
- Watts, C.H.; Clarkson, B.R.; Cave, V.M.; Thornburrow, D.; Thorpe, S. Invertebrate communities in a modified isolated raised bog compared to an intact raised bog in New Zealand. Mires and Peat 2020, 26 (20), 1–12. [CrossRef]
- Spitzer, K.; Danks, H.V. Insect biodiversity of boreal peat bogs. Annual Review of Entomology 2006, 51, 137–161. [CrossRef]
- Buczyńska, E.; Buczyński, P. Aquatic insects of man-made habitats: environmental factors determining the distribution of caddisflies (Trichoptera), dragonflies (Odonata), and beetles (Coleoptera) in acidic peat pools. Journal of Insect Science 2019, 19 (1), 1–15. [CrossRef]
- Hejkal, J. Carabids (Coleoptera, Carabidae) of the peat bog Soos in N-W Bohemia: a faunistical and ecological study. Folia Musei Rerum Naturalium Bohemiae Occidentalis, Plzeň, Zoologica 1990, 32, 3–55.
- Biesiadka, E.; Мoroz, M.D. A preliminary description of water beetles (Coleoptera) of the Zehlau peatbog. In Flora and Fauna of the Zehlau Peatbog; KSU: Kaliningrad, Russia, 1996; pp. 38–40.
- Spitzer, K.; Bezděk, A.; Jaroš, J. Ecological succession of a relict Central European peat bog and variability of its insect biodiversity. Journal of Insect Conservation 1999, 3, 97–106. [CrossRef]
- Främbs, H.; Dormann, W.; Mossakowski, D. Spatial distribution of carabid beetles on Zehlau bog. Baltic Journal of Coleopterology 2002, 1 (2), 7–13.
- Boháč, J.; Bezděk, A. Staphylinid beetles (Coleoptera, Staphylinidae) recorded by pitfall and light trapping in Mrtvý Luh peat bog. Silva Gabreta 2004, 10, 141–150.
- Dapkus, D.; Tamutis, V. Assemblages of beetles (Coleoptera) in a peatbog and surrounding pine forest. Baltic Journal of Coleopterology 2008, 8 (1), 31–40.
- Sushko, G.G. Beetles (Coleoptera) of Raised Bogs in North-Western Belarus (Belarusian Land O’Lakes). Baltic Journal of Coleopterology 2007, 7 (2), 207–214.
- Sushko, G.G. Spatial distribution of epigeic beetles (Insecta, Coleoptera) in the “Yelnia” peat bog. Baltic Journal of Coleopterology 2014, 14 (2), 151–161.
- Sushko, G.G. Rove beetles (Coleoptera: Staphylinidae) of a large pristine peat bog in Belarus Lake District. Fragmenta Faunistica 2018, 61 (2), 99–104. [CrossRef]
- Sushko, G.G. Key factors affecting the diversity of Sphagnum cover inhabitants with the focus on ground beetle assemblages in Central-Eastern European peat bogs. Community Ecology 2019, 20 (1), 45–52. [CrossRef]
- Sushko, G.G. A comparison of ground beetle assemblages (Coleoptera: Carabidae) between birch forests of continuous and isolated peat bog habitats in Belarusian Lake District. Proceedings 2021, 68. [CrossRef]
- Prokin, A.A.; Sazhnev, A.S.; Philippov, D.A. Water beetles (Insecta: Coleoptera) of some peatlands in the North Caucasus. Nature Conservation Research 2019, 4 (2), 57–66. [CrossRef]
- Sushko, G.G. Diversity and species composition of beetles in the herb-shrub layer of a large isolated raised bog in Belarus. Mires and Peat 2017, 19 (10), 1–14. [CrossRef]
- Sushko, G.G. Leaf beetles (Coleoptera, Chrysomelidae) in the pristine peat bog in Belarus: biodiversity and spatial distribution. North-Western Journal of Zoology 2020, 16 (2), 211–215.
- Sushko, G.G. Peat bog weevil beetles (Insecta, Coleoptera) of Belarusian Lake District. Bulletin of VDU 2020, 3 (108), 22–28. (in Belorussian).
- Skwarra, E. Die Käferfauna des Zehlaubruches. Schriften der Physikalisch-ökonomischen Gesellschaft zu Königsberg i. Pr. 1929, 66 (2), 181–274.
- Aleksandrowicz, O.R. Changes in the carabid fauna of Polesie peat-bog due to drainage, ploughing and agricultural development. In How to protect or what we know about Carabid Beetles, Szyszko, J. et al., Eds.; Agricultural University Press: Warsaw, Poland, 2002; pp. 171–184.
- Novikova, Yu.I.; Sushko, G.G. Biodiversity variability of the ground beetle assemblages (Сoleoptera, Сarabidae) of a peat bog during anthropogenic transformation of their habitats. Journal of the Belarusian State University. Ecology 2023, 1, 12–19. (in Russian). [CrossRef]
- Zoch, L.; Budig, S.; Reich, M. Sphagnum cultivation sites as habitat for beetles (Coleoptera) and the efect of vegetation structure on species occurrence and abundance. Journal of Insect Conservation 2023. [CrossRef]
- Emden, van F. Coleopterenlarven aus dem Zehlaubruch. Schriften der Physikalisch-ökonomischen Gesellschaft zu Königsberg i. Pr. 1929, 66 (2), 275–286.
- Antsiferova, O.A.; Napreenko, M.G.; Napreenko-Dorokhova, T.V. Transformation of soils and mire community reestablishment potential in disturbed abandoned peatland: a case study from the Kaliningrad Region, Russia. Land 2023, 12 (1880). [CrossRef]
- Napreenko, M.; Danchenkov, A.; Napreenko-Dorokhova, T.; Samerkhanova, A. Vegetation mapping in drained peatlands for the carbon research objectives: a case study from Kaliningrad Region. Mires and Peat 2023, 29 (19), 1–25. [CrossRef]
- Napreenko, M.G.; Napreenko-Dorokhova, T.V.; Karelina, V.I.; Pelens, E.D. Species composition and habitat ecology of sphagna: inventory and monitoring programme on the carbon polygon ‘Rossyanka’ (Kaliningrad region, Russia). Natural and medical sciences 2022, 1, 73–87. (in Russian).
- Freude, H.; Harde, K.W.; Lohse, G.A. Die Käfer Mitteleuropas (Bd. 1-15); Goecke & Evers: Krefeld, Germany, 1965–1989.
- Freude, H.; Harde, K.W.; Lohse, G.A., Klausnitzer, B. Die Käfer Mitteleuropas (Bd. 2: Carabidae); Elsevier GmbH-Spectrum: München, Germany, 2004; pp. 1-521.
- Bieńkowski, A.O. Leaf-beetles (Coleoptera: Chrysomelidae) of the Eastern Europe. New key to subfamilies, genera, and species. Mikron-print: Moscow, Russia, 2004; pp. 1-278.
- Ryndevich, S.K. The fauna and ecology of the Belarusian water beetles. Technoprint: Minsk, Belarus, 2004; pp. 1–272. (in Russian).
- Lawrence, J.F. Classification (families & subfamilies). In Handbook of Zoology–Coleoptera, Beetles, Beutel, R.g., Leschen, R.A.B., Eds.; de Gruyter: Berlin, Germany, 2016; Volume 1, pp. 13–22. [CrossRef]
- Peus, F. Beitrage zur Kenntnis der Tierwelt nordwestdeutscher Hochmoore. Eine ökologische Studie. Insecten, Spinnentiere, Wirbeltiere. Zeitschrift für Morphologie und Ökologie der Tiere 1928, 12 (3/4), 533-683.
- Mossakowski, D.; Främbs, H.; Lakomy, W. The carabid and staphylinid fauna of raised bogs. A comparison of Northwest Germany and the Baltic region. Baltic Journal of Coleopterology 2003, 3 (2), 137–144.
Figure 1.
Location of the Vittgirrensky peatland (the ‘Rossyanka’ Carbon Measurement Supersite) and the Zehlau raised bog (mentioned in the text for comparison) in Kaliningrad Oblast.
Figure 1.
Location of the Vittgirrensky peatland (the ‘Rossyanka’ Carbon Measurement Supersite) and the Zehlau raised bog (mentioned in the text for comparison) in Kaliningrad Oblast.
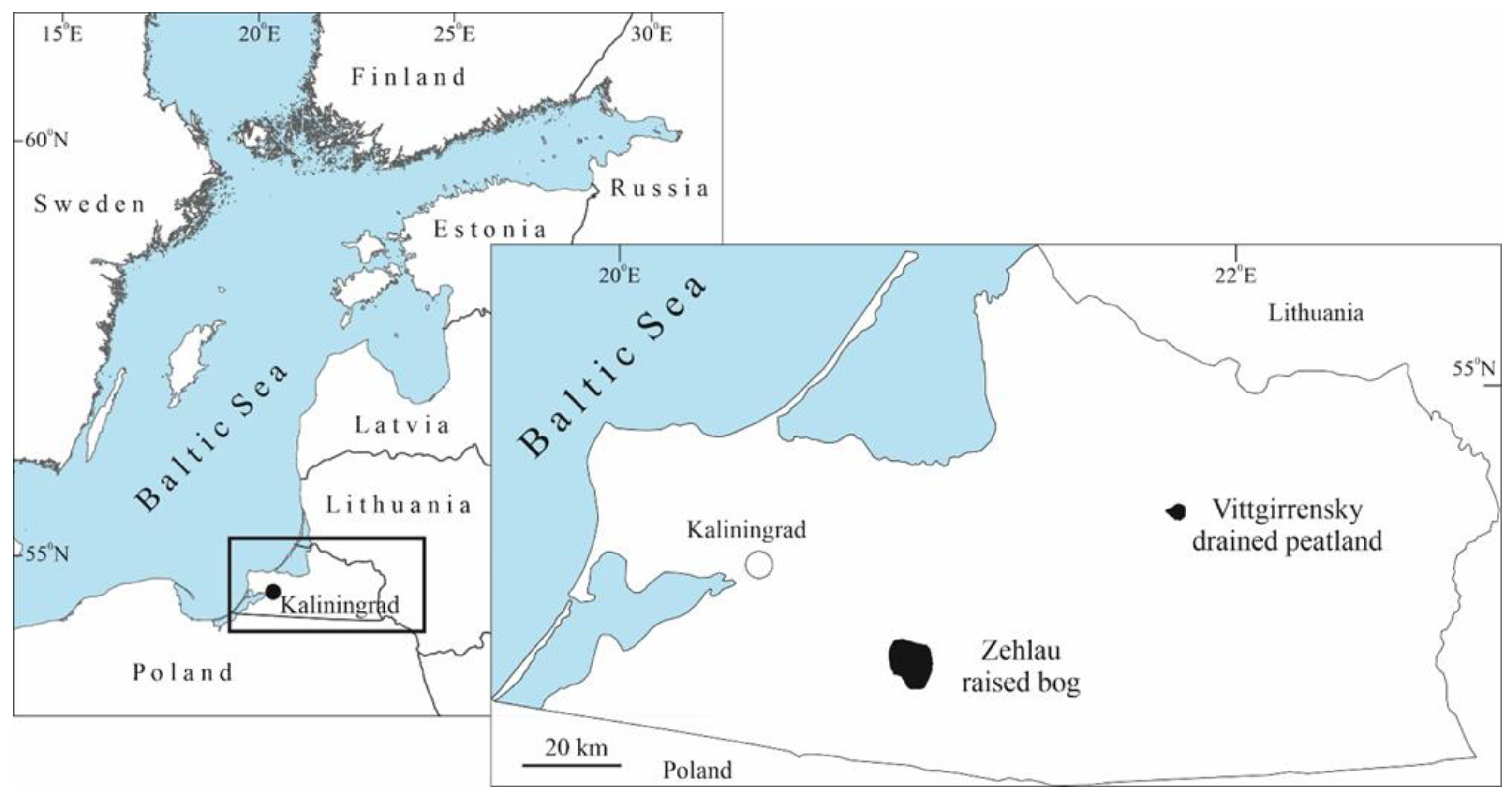
Figure 2.
Vegetation cover in the Vittgirrensky Peatland [after 28,29] and location of the sampling points for Coleoptera in 2023: 1 – Dry shrublands, 2 – Wet shrublands, 3 – Dry birch stand, 4 – Fen-like communities (Juncus), 5 – Birch coppice, 6 – Reed beds, 7 – pitfall traps set in lines, 8 – Bare-peat sites, 9 – Dense closed-canopy stand, 10 – Fen-like communities (Eriophorum/Carex), 11 – Wet forest (birch and aspen), 12 – Hydrophilic communities in ditches, 13 – dirt road, 14 – places of sweeping with net in water.
Figure 2.
Vegetation cover in the Vittgirrensky Peatland [after 28,29] and location of the sampling points for Coleoptera in 2023: 1 – Dry shrublands, 2 – Wet shrublands, 3 – Dry birch stand, 4 – Fen-like communities (Juncus), 5 – Birch coppice, 6 – Reed beds, 7 – pitfall traps set in lines, 8 – Bare-peat sites, 9 – Dense closed-canopy stand, 10 – Fen-like communities (Eriophorum/Carex), 11 – Wet forest (birch and aspen), 12 – Hydrophilic communities in ditches, 13 – dirt road, 14 – places of sweeping with net in water.
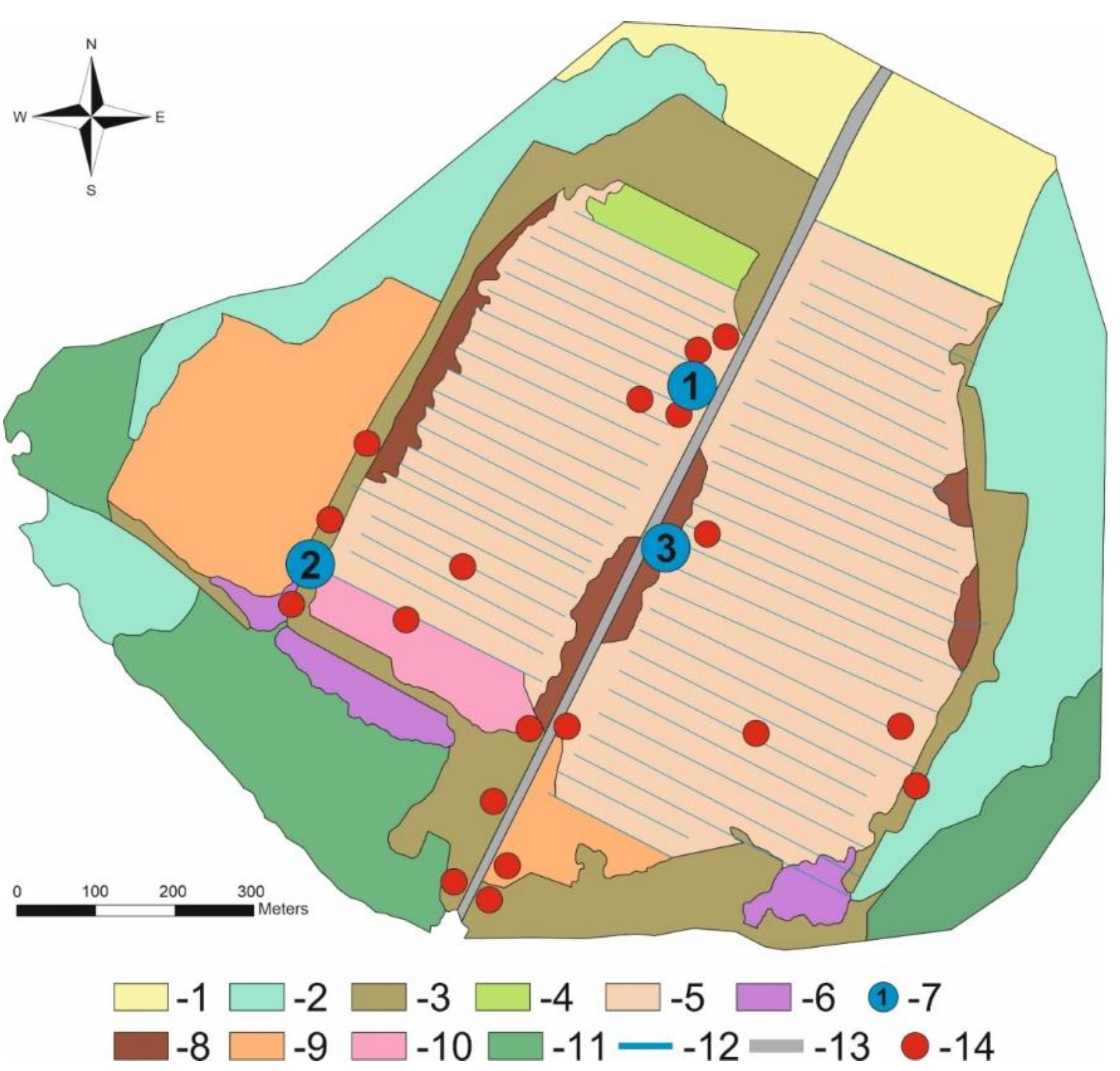
Figure 3.
The main typical habitats of the Vittgirrensky peatland: A – Birch coppice with Calluna (site of the pitfall line 1), habitat for Micrelus ericae, Curimopsis nigrita, Altica longicollis; B – Open bare peat (site of the pitfall line 3), habitat for Cymindis vaporariorum, Parabolitobius inclinans, Pselaphus heisei, Curimopsis nigrita; C – Dry birch stand (site of pitfall line 2), habitat for Sciaphilus asperatus, Barypeithes pellucidus, Brachysomus echinatus; D – Phragmites-dominated birch coppice, habitat for Malthodes pumilus, Scymnus suturalis; E – The drainage ditch drying up in summer, sampling place of Dytiscus dimidiatus, Graphoderus cinereus, Hydaticus seminiger; F – non-drying drainage ditch, sampling place of Hydrochus elongatus, Enochrus ochropterus.
Figure 3.
The main typical habitats of the Vittgirrensky peatland: A – Birch coppice with Calluna (site of the pitfall line 1), habitat for Micrelus ericae, Curimopsis nigrita, Altica longicollis; B – Open bare peat (site of the pitfall line 3), habitat for Cymindis vaporariorum, Parabolitobius inclinans, Pselaphus heisei, Curimopsis nigrita; C – Dry birch stand (site of pitfall line 2), habitat for Sciaphilus asperatus, Barypeithes pellucidus, Brachysomus echinatus; D – Phragmites-dominated birch coppice, habitat for Malthodes pumilus, Scymnus suturalis; E – The drainage ditch drying up in summer, sampling place of Dytiscus dimidiatus, Graphoderus cinereus, Hydaticus seminiger; F – non-drying drainage ditch, sampling place of Hydrochus elongatus, Enochrus ochropterus.

Figure 4.
Some of the potential indicator species (tyrphophiles and abundant tyrphoneutrals) that can be used, among others, to monitor environmental impacts in the Vittgirrensky peatland: A – Altica aenescens; B – Orchestes jota; C – Platydracus fulvipes; D – Scymnus suturalis; E – Lochmaea caprea; F – Polydrusus cervinus.
Figure 4.
Some of the potential indicator species (tyrphophiles and abundant tyrphoneutrals) that can be used, among others, to monitor environmental impacts in the Vittgirrensky peatland: A – Altica aenescens; B – Orchestes jota; C – Platydracus fulvipes; D – Scymnus suturalis; E – Lochmaea caprea; F – Polydrusus cervinus.


Table 1.
The most species-rich families (≥10 species) in the Vittgirrensky peatland (own results) and the Zehlau bog [9,11,23].
| No | Family | Vittgirrensky | Zehlau | ||
|---|---|---|---|---|---|
| Number of species | % | Number of species | % | ||
| 1 | Cantharidae | 13 | 4.6 | 10 | 2.6 |
| 2 | Carabidae | 30 | 10.7 | 36 | 9.3 |
| 3 | Chrysomelidae | 24 | 8.5 | 41 | 10.6 |
| 4 | Coccinellidae | 10 | 3.6 | 20 | 5.2 |
| 5 | Curculionidae | 30 | 10.7 | 38 | 9.8 |
| 6 | Dytiscidae | 28 | 10.0 | 43 | 11.1 |
| 7 | Elateridae | 11 | 3.9 | 11 | 2.8 |
| 8 | Staphylinidae | 43 | 15.3 | 85 | 22.0 |
| Coleoptera in total | 281 | 100 | 387 | 100 | |
Table 2.
Comparison of basic plant communities on an idealised Central European raised bog and in the Vittgirrensky peatland.
Table 2.
Comparison of basic plant communities on an idealised Central European raised bog and in the Vittgirrensky peatland.
| Typical Central European raised bog [after 6] | Vittgirrensky peatland [28] |
|---|---|
| (1) Central bog pool | Hydrophilic communities in ditches (!) – only separate fragments and only for a limited period of time |
| (2) Pine elfin forest | Absent |
| (3) Open dead pine forest and treeless formation with high water level | Absent |
| (4) Birches (Betula spp.) and pines (Pinus spp.) on inner lagg (moat) margin | Birch coppice (!) – only separate small fragments with Sphagna |
| (5) Treeless lagg | Fen-like communities (!) – only for a limited period of time |
| (6) Border communities between lagg and surrounding forest | Absent |
| (7) Surrounding forest | - Wet forest (birch and aspen) - Dry birch stand - Dense closed-canopy stand |
| Absent | Birch coppice (most part without Sphagna) |
| Absent | Dry shrublands |
| Absent | Wet shrublands |
| Absent | Reed beds |
| Absent | Bare-peat sites |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



