
Submitted:
04 March 2024
Posted:
04 March 2024
You are already at the latest version
Abstract
This study aimed to investigate the venom sac extracts (VSE) of the European hornet (EH) Vespa crabro (Linnaeus, 1758) (Hymenoptera: Vespidae), focusing on the differences between stinging females, gynes (G) and workers (W), at the protein level. Using a quantitative “Sequential Window Acquisition of all Theoretical Fragment Ion Mass Spectra” (SWATH-MS) analysis, we identified and quantified a total of 240 proteins. Notably, within the group, 45.8 % (n = 110) showed significant differential expression between VSE-G and VSE-W. In this set, 57.3 % (n = 63) were upregulated and 42.7 % (n = 47) downregulated in the G. Additionally, the 200 quantified proteins from the class Insecta belong to 16 different species, six of them to the Hymenoptera/Apidae lineage, comprising seven proteins with known potential allergenicity. Thus, phospholipase A1 (Vesp v 1), phospholipase A1 verutoxin 2b (VT-2b), hyaluronidase A (Vesp v 2A), hyaluronidase B (Vesp v 2B), and venom allergen 5 (Vesp v 5) were significantly downregulated in the G, and vitellogenin (Vesp v 6) was upregulated. Overall, 46 % of the VSE proteins showed differential expression, with a majority being upregulated in G. Data are available via ProteomeXchange with identifier PXD047955. These findings shed light on the proteomic differences in VSE between EH castes, potentially contributing to our understanding of their behavior and offering insights for allergy research.
Keywords:
Vespa crabro
; European hornet
; wasps
; venom sac
; castes
; gynes
; workers
; proteomics
; SWATH-MS analysis
; venomics.
1. Introduction
A wide array of venomous creatures can be found across various phyla, showcasing an extraordinary variety of taxa, toxins, targets, clinical effects, and outcomes [1]. In fact, venomous organisms have captivated the scientific community due to their remarkable chemical adaptations and diverse biological functions [2,3]. Proteomics, the large-scale study of proteins within a biological sample, has emerged as a powerful tool to decipher the intricate molecular components of venoms. By combining advanced analytical techniques, such as liquid chromatography-mass spectrometry (LC-MS), with bioinformatics approaches, researchers can obtain detailed insights into the venom's proteinaceous constituents [4,5,6]. This in-depth understanding of the venom proteome allows the identification, characterization, and functional annotation of venom proteins, enabling a holistic understanding of the venom's biological activities and potential applications in various fields.
The Vespa crabro (Linnaeus, 1758) (Hymenoptera: Vespidae), is a hornet, commonly known as the European hornet (EH). It is one of the largest species of hornet, measuring up to 35 mm in length, and is found in Europe, Asia and North America [7]. EHs are social insects that live in large colonies. They build their nests in tree hollows, cavities in buildings, or other protected areas. The nest is made of a paper-like material created with wood fibers which are chewed up and mixed with saliva. As predators, EHs feed on their brood with a variety of insects, including other bees and wasps, flies, and caterpillars. Adults are also attracted to sweet substances and may be seen feeding on ripe fruit or sugary liquids. While EH can be beneficial for controlling other insect populations, they can also be a nuisance or a danger to humans -and other animals. Their sting is painful, and some people sensitized against their venom may develop an allergic reaction after suffering from a sting [8,9,10,11,12,13,14,15,16,17,18,19,20].
Research has been conducted to identify and characterize the bioactive components present in the venom of wasps in general and EH in particular. In general, wasp venoms are composed of a diverse array of peptides, proteins, and enzymes that play crucial roles in prey capture, defence, and communication within the colony [21,22]. Studies have shown that EH venom components like crabrolin and mastoparans exhibit antimicrobial properties against various pathogens, including bacteria and fungi [23,24,25,26]. Furthermore, the venom of EH contains enzymes, such as hyaluronidase, which play a role in facilitating the spread of venom within the prey or victim's tissues [27]. Moreover, the immunological properties of EH venom compounds have been investigated. Allergenic components, such as antigen 5 molecules, have been identified and characterized, shedding light on the structural basis of antigenic cross-reactivity among different hymenopteran venoms [28,29,30]. By investigating the venom compounds of different wasps, researchers aim to gain insights into the molecular mechanisms underlying its venomous properties and explore potential applications in various fields, including pharmacology and biotechnology [31,32,33,34].
Currently, research on the venom composition of social insects, particularly wasps, has gained attention due to the multifaceted functions of venom beyond prey capture and defence. However, these studies have been limited by their inability to quantify the complete abundance of venom components accurately. Moreover, investigations into the caste-specific differences in venom composition have been scarce [35], despite the behavioural and physiological variations observed among female castes in wasps colonies.
To address these knowledge gaps and build upon the existing body of research, our study applies a quantitative Sequential Window Acquisition of all Theoretical Fragment Ion Mass Spectra (SWATH-MS) approach to comprehensively analyze and compare the venom sac extract (VSE) proteome of gyne (G) and worker (W) EH females. SWATH-MS is a mass spectrometry-based technique that allows for comprehensive and quantitative proteomic analysis [36,37]. It is a data-independent acquisition method that offers advantages over traditional mass spectrometry quantitative methods, such as selected reaction monitoring or parallel reaction monitoring. In the present work, by quantifying the abundance of proteins from venom sac extracts (VSE) of gynes (VSE-G) and workers (VSE-W), we aim to uncover potential caste-specific differences within the EH social structure.
2. Results
2.1. Source of Insects: the Nest and Colony Composition
A nest of the European hornet, Vespa crabro, in its native habitat in Galicia (NW Spain) was obtained (Figure 1). The nest was affixed to the wall within a firewood storage at a height of 2 m above the ground, distinguishing: a very friable outer layer of paper-like material, typically in varying shades of grey and brown, that is wrapped around, except for a small opening at the bottom and an internal chamber, with eight combs. The insect adult population (IAP) found (gyne, male, queen and worker) was as follows: n = 168 individuals (58, 42, 1 and 67).
2.2. Venom Sac, Extract Protein Quantitation and SDS-PAGE.
The venom sac (VS) was carefully removed from both gynes (designated as VS-G) and workers (designated as VS-W) stinging apparatus. After being cleaned, a distinct contrast in the external structure of castes VS was observed (as shown in Figure 2). Specifically, the VS-W appeared elongated and exhibited greater transparency compared to the more rounded and opaque appearance of the VS-G.
The protein of the venom sac extracts (VSE) of gynes (VSE-G) and workers (VSE-W) was quantified (Figure 3a), and no differences (Mann-Whitney test p > 0.99) were found between G (186.9 ± 29 g/sac) and W (188.8 ± 89 g/sac). Figure 3b illustrates the electrophoretic separation of VSE-G and VSE-W. The analysis unveiled a diverse composition of venom components, spanning molecular masses from 10 to 250 kDa. Notably, the VSE-G profile displayed a higher concentration of proteins within the 25-250 kDa mass range compared to the VSE-W, while no variations were detected within the low-molecular mass range (10-25 kDa). Evident and distinct bands corresponding to known potential allergens of EH (Vesp c 2, two isoforms of Vesp c 2 and Vesp c 5) were pinpointed based on their molecular masses in VSE-W, with these bands being less pronounced in VSE-G.
2.3. Proteomic Quantitative Analysis of Total Proteins in the Venom Sac Extract
To delve deeper into the distinctions within VSE content between gynes (G) and workers (W), we conducted LC-MS/MS mass spectrometry, utilizing the SWATH-MS quantification approach to quantified proteins with varying expression. Across the four biological replicates, a cumulative count of 240 proteins were both identified and quantified (detailed in Table 1), encompassing 11 distinct classes and 33 different species. As anticipated, a significant majority of the identified proteins, constituting 83.3% (n = 200), were categorized under the class Insecta.
Among the 240 identified proteins, 45.8 % (n = 110) showed significant differential expression between VSE-G and VSE-W samples (FC ≥ 1.5 and p ≤ 0.05). Among them, 57.3% (n = 63) were upregulated and 42.7% (n = 47) downregulated in the VSE-G. A volcano plot (Figure 4) was employed to represent the global quantification of the VSE proteins in the female castes and indicate the dysregulated proteins between the groups. The SWATH-MS most-likely ratio (MLR) normalized areas (per protein and sample) and the fold change and p-values (t-test) are available in Supplementary File S1.
Among the 63 proteins upregulated in the VSE-G, the 4 most overexpressed are Glycogen debranching enzyme (FC = 40.1; p = 0.004), muscle LIM protein MIp84B isoform X2 (FC = 34.6; p = 0.016), myosin light chain alkali (FC = 30.3; p = 0.039), and vitellogenin (FC = 17.8; p = 0.0005). Regarding de 4 most downregutaled proteins in the VSE-G, we observed a decreased expression of venom allergen 5 (FC = 38.2; p = 0.006), phospholipase A1 (FC = 19.2; p = 0.0003), calmodulin (FC = 19.2; p = 0.025), and alpha-glucosidase (FC = 15.0; p < 0.0001).
Figure 5 shows the most common gene ontology (GO) terms of the differentially expressed proteins in EH female castes, allowing for an assessment of the biological processes (Figure 5A) in which they are involved, and a quick comparison of proteins functions at the molecular level (Figure 5B). The results revealed the involvement of the VSE-G downregulated proteins in the regulation of carbohydrate metabolic process, lipid metabolic process, defense response, glucose metabolic process, and proteolysis, as well as the participation of the VSE-W overexpressed proteins in the glycogen metabolic process, protein phosphorylation, tricarboxylic acid cycle, and cell differentiation. Regarding the molecular function, the results revealed the involvement of the VSE-G upregulated proteins in ATP binding, metal ion binding, calcium ion binding, magnesium ion binding, and pyridoxal phosphate binding. On the other hand, the VSE-W upregulated protein are involved in NAD binding, lipid binding, hyalurononglucasaminidase activity, and phospholipase A1 activity. A detailed list with GO terms is available in Supplementary File S2.
2.4. Proteomic Quantitative Analysis from Dysregulated Proteins of the Class Insecta.
The 200 quantified proteins from the class Insecta belong to 16 different species (Table 1), six of them to the Hymenoptera/Apidae lineage. Among them, we were able to quantified seven proteins with known potential as allergens (showed as blue spots in Figure 4). As shown in Figure 6, phospholipase A1 (Vesp v 1), phospholipase A1 verutoxin 2b (VT-2b), hyaluronidase A (Vesp v 2A), hyaluronidase B (Vesp v 2B), and venom allergen 5 (Vesp v 5) were significantly downregulated in the VSE-G, and vitellogenin (Ves v 6) was upregulated (Mann-Whitney test, p < 0.05).
In general, 46% (n = 92) of the proteins from class Insecta showed significant differential expression between VSE-G and VSE-W samples (FC ≥ 1.5 and p ≤ 0.05); the 65% (n = 60) were upregulated and 35 % (n = 32) downregulated in the VSE-G (Supplementary Table S1 and Table S2, respectively). From the Hymenoptera/Apidae lineage, a total of 44, 43, 36, 29 and 28 proteins from B. terrestris, A. mellifera, A. cerana, D. novaeangliae and F. varia, respectively, were quantified in the VSE of EH female castes.
From the D. novaeangliae (Family Halictidae) we could quantified 29 proteins, showing 48.3 % (n = 14) differential expression between the EH female castes, eleven were upregulated in the VSE-G and three were downregulated in the VSE-G. All the proteins quantified from D. novaeangliae are available in STRING, the protein-protein interaction network tool (accessed on june 16 of 2023). In this way, a map of the interaction of proteins and their main biological function was obtained (Figure 7). We performed k-means clustering analysis and obtained three clusters, with 8, 10 and 11 proteins. The three downregulated proteins in VSE-G all belong to the cluster 3 (Figure 7A and 7D), being the upregulated proteins mainly in the cluster 1 (6 proteins) and 2 (4 proteins). Complementary, we carried out a search for biological processes using the functional enrichment analysis (FunRich) tool. The results revealed the involvement of the upregulated proteins in the generation of precursor metabolites and energy and the actomyosin structure organization; while the downregulated proteins are involved in the glicerol-3-phosphate metabolic process.
2.5. Proteomic Quantitative Analysis from Dysregulated Proteins of the Other Classes.
A total of 40 proteins from the other classes were quantified in the VSE-G and VSE-W. Among them, 45 % (n = 18) showed significant differential expression between VSE-G and VSE-W samples (FC ≥ 1.5 and p ≤ 0.05); the 16.7 % (n = 3) were upregulated and 83.3 % (n = 15) downregulated in the VSE-G (Table 2). A total of 17 (42.5 %) proteins belonged to the mimic poison frog R. imitator, among them, the 47 % (n = 8) were downregulated in the VSE-G.
3. Discussion
The selective sampling approach by collecting specimens exclusively from a single colony, offers a unique advantage, ensuring a cohesive genetic and environmental context for the study. Moreover, the integration of cutting-edge analytical techniques, by the application of SWATH-MS, with its capacity for quantitative analysis of the complete proteomic profile, allow to explore the differences in venom composition across castes with unprecedented precision. We showed for the first time differences in the external morphological and in the protein-banding pattern between venom sacs (VS) and venom sac extracts (VSE) from female castes of the EH. These differences were similar to those found in Vespa velutina [35]; thus, this two hornet species exhibing a great degree of conservation between venom sac of female castes. Previous work has shown similar results for Vespa orientalis (an egg-shaped venom sac, 3-5 mm length and 2-3 mm at maximum width) [38]. In contrast, other authors indicated venom sacs of Vespa velutina to be approximately 1 mm in length, white, and transparent [39]. The electrophoretic separation of VSE highlights differences in their profiles, suggesting functional specialization and adaptation to their distinct roles within the colony. Gynes appear to possess a higher diversity or abundance of proteins in the 25-250 kDa mass range, potentially related to reproductive and colony-establishment functions. In contrast, workers display clear and distinct bands of known allergenic proteins, indicating a focus on defense and deterring potential threats. These findings underscore the caste-specific roles and ecological interactions of the EH, shedding light on the complex nature of their venom composition. Further research is needed in more hornet species to identify and characterize specific venom components, enhancing our understanding of the functional significance and mechanisms behind these differences.
Since the EH protein database is not well annotated, protein and peptide identification were based on homology of sequence, and was carried out using a customized database including Vespa + Vespa velutina + Apis mellifera + poison + toxins (Available online: https://www.uniprot.org/ (accesed on 2 February 2023)). A total of 240 proteins were quantified in the VSE-G and VSE-W, belonging to 11 different classes, 14 ordes, 17 families and 33 species, highligting: Insecta (n=200); Amphibia (n=17); γ-Proteobacteria (n=8); and Arachnida (n=5).
Proteins belonging to class Insecta were quantified by SWATH-MS in the VSE-G and VSE-W. F. varia (n=28 proteins), aka yellow marmalade bee is a highly eusocial aggressive stingless bee distributed in several areas of Brazil. Workers are completely sterile, with a programmed cell death during pupal development, resulting in non-functional ovaries. A. cerana (n=36 proteins), the Asian honeybee, is a species native to South, Southeast and East Asia; territory where 22 species of hornets are present. Hornets and their native honeybees prey are the main characters of a coevolutionary arms race that is made evident by the conspicuous number of reciprocal adaptations evolved by both animals. D.novaeangliae (n=29 proteins), the sweat bee, is a soil-dwelling solitary species found in north-eastern USA and Canada, and forages for pollen only on pickerel weed (P. cordata). Prior research indicates the possibility that the entire existing Hymenoptera lineage could have originated from a shared ancestor with venomous traits. [40].
Proteins were quantified by SWATH-MS from R. imitator (n=17), the mimic poison frog, naturally distributed in the north-central region of eastern Peru. R. imitator mimics not one but three other Ranitomeya species of highly toxic poison frogs (R. fantastica, R.variabilis, and R. ventrimaculata). This represents the only known example of mimetic radiation in amphibians. Also, this species shows pair-bonding and biparental care, being the only known monogamous amphibian in the wild confirmed by paternity analysis [41]. Additional investigation is required to determine if these proteins pose a potential threat to mammals following EH stings,
Proteins from γ-proteobacteria (Klebsiella sp. RIT-PI-d, Cronobacter sakazakii, Citrobacter freundii, Escherichia coli) and Bacilli (Lactobacillus apis , Lactococcus lactis) were identified by SWATH-MS in the VSE of EH. The gut bacteria found in various species and regions within the Vespa genus exhibited significant similarity in their compositions at both the phylum and class levels [42]. In fact, Lactobacillus sp [43] which are the main species in the gut of honeybees are also present in EH. Klebsiella sp, [44] was also previously reported in Vespa sp individuals. Further research is imperative to ascertain the origin of these microorganisms, some of which are typically part of hornet gut flora or could potentially appear in the analyzed extracts due to cross-contamination during the sac extraction process. Two proteins from T. mercedesae (Arachnida) were quantified. Tropilaelaps mites are originally parasites of the giant honey bees (A. dorsata, A. laboriosa, and A. breviligula). Amongst the four species of Tropilaelaps, T. clareae and T. mercedesae are successfully adapted to A. mellifera. T. mercedesae, has recently expanded its geographic range to areas of South Korea and China [45].
The invasive species Vespa velutina, commonly known as the Asian hornet, has garnered significant attention both in the media and the medical community due to its expanding presence and the potential health implications associated with its venom [46,47,48,49,50]. Media coverage has often emphasized the negative impact of Vespa velutina on honeybee populations and its potentially aggressive behaviour towards humans, amplifying public awareness. From an allergological point of view, a comparison between Vespa velutina [35] and EH highlights the difference at the level of phospholipase A1 (Vesp v 1), where two distinct bands are identified [designated as Vesp v 1 (a) and (b)]. It is notable that in Vespa velutina the Vesp v 1 (a) band is the most predominant, while in EH the most abundant band is Vesp c 1 (b). In addition, VSE-G have been observed to have higher numbers of Vesp c 2 and Vesp c 1 compared to Vespa velutina gynes.
While this study has provided valuable insights into the proteomic differences within the venom sac extracts (VSE) of European hornets (EH) Vespa crabro castes, there are certain limitations that warrant consideration. One limitation lies in the fact that the analysis focused on homogenized venom sac extracts, potentially overlooking variations that may arise from different extraction methods [51]. Future research could explore alternative techniques, such as direct filtration of venom sac content without homogenization, to elucidate potential differences in protein profiles and their implications. Furthermore, the functional implications of the identified differentially expressed proteins remain to be fully elucidated. Future investigations could delve into the specific roles of these proteins in envenomation processes, potentially uncovering novel mechanisms underlying caste-specific venom activity. To fully comprehend the allergological implications of these distinctions, future research should employ IgE immunoblot analyses to evaluate the specific immunoreactivity of these proteins and their potential contribution to allergic responses [52,53,54,55,56]. By comprehensively cataloging and comparing the proteomic signatures of various hymenopteran species, we could discern patterns, establish correlations with clinical outcomes of stings, and ultimately enhance our capacity to predict and manage the medical consequences of hymenopteran envenomations. An optimal approach for advancing our understanding of hymenopteran envenomation and its implications would involve comprehensive proteomic analyses encompassing various hymenopteran species within a specific geographical region. This holistic approach aligns with the broader One Health perspective, integrating entomological, medical, and ecological dimensions to address the multifaceted challenges posed by stinging insects.
4. Materials and Methods
4.1. Source of Insects
4.1.1. Study Area
Field research aimed at collecting the Vespa crabro insects was conducted in the Galician municipality of Pazos de Borbén. Galicia is located in the northwest of Europe on the Iberian Peninsula (Figure 8). The municipal term of Pazos de Borbén has an extension of 50 km2 and is located, between the 42° 14' 40" North latitude, and 4º 47' 30" and 4º 54' 40" latitude West. The climate is typified as humid oceanic with a tendency to summer aridity (average annual temperature of 13 ºC, and an average annual precipitation of 1,994 mm.). Rock substrate is composed of granite of two mica. The most abundant soils are the brown earths and the rankers. Considering the flora, the native forest is mainly composed by Quercus robur. and accompanied by Quercus pyrenaica, Castanea sativa, Laurus nobilis, Fagus sylvatica); with a herbaceous layer formed by: Teucrium scorodonia, Viola riviniana. Luzula forsteri or Luzula sylvatica.
Riparian forest, bordering the river courses (Borbén, Asemonde, Pozo Negro, Pequeno and Alvedosa), is dominated by Alnus glutinosa, and accompanied by Betula pubescens subsp. celtibérica, various species of Salix sp. and even Q. robur. Large patches of Ulex europaeus. Rubus sp.. Pteridium aquilinum and Cytisus striatus. among other. Ascending at height, other species appear, such as Genista florida. Cytisus scoparius or Cytisus grandiflorus. Reforestation productive forest is formed by E. globulus, P. pinaster and mixed masses of the two species.
4.1.2. Insect Localization and Identification
Eucalyptus logging points were identified within the study area and strategically selected as potential hotspots for vespid activity (Figure 9).
4.1.3. Identification. Tracking and Locating Nest of Target Species
The identification of Vespa crabro individuals was conducted based on their external morphological characteristics. Easily distinguished from V. velutina by coloration (Figure 9b): in particular, the coloration of the second abdomen tergum (which has the posterior half almost entirely yellow, with the part clearly tridentate apically), and that of the fifth and sixth tergs (which are almost entirely yellow coloring. Upon successful identification of Vespa crabro individuals at the eucalyptus cutting points, tracking was initiated. Individual hornets were followed to trace their flight paths and possible nesting locations.
4.1.4. Nest removal
The nest of the European hornet, Vespa crabro was collected on 5 December 2022 during the night (Figure 10). A complete apiary suit designed to offer full body protection was utilized during the Vespa crabro nest collection activity. This suit includes a one-piece set of pants and a jacket, a hat, a veil, and a pair of long-sleeve leather gloves. For the removal of the nest, firstly, its lower opening was covered to contain the insects inside. Subsequently, with the help of a spatula, its adherent parts were separated from the wall. It was then placed in carton box for 24 hours at a temperature of −18 °C in a freezer, until it is taken to the laboratory.
4.1.5. Nest Colony Composition and Determining Sex
The count of mature individuals was conducted manually to determine insect adult population (IAP). Subsequently, the collected insects were sorted based on the categories of sex, reproductive individuals and castes: gyne (G), male (M), queen (Q) and worker (W). Female and male hornets exhibit sexual dimorphism. Female hornets have 12 antennal segments (including the scape and pedicel), while male hornets have a relatively long, curly-endded antennae with 13 segments. Female hornets have 6 gastral segments and are equipped with a stinger (modified ovipositor used for defense). On the other hand, male hornets have 7 exposed gastral segments and lack a stinger. Males have a bilobate apex of the last sternite, which appears sharp in females. The distinction between workers and gynes (future queens) was established based on the size, the gynes being larguer than the workers. Size: workers, 15-25 mm.; queens, 25-35 mm.
4.2. Venom Sac Extract Obtention
The venom sacs (VS) were collected from frozen insects by carefully extracting the stinger apparatus from the hornet's abdomen using forceps. Subsequently, the VS were separated from the stingers. Three VS were combined to create four pools of worker (VS-W) and gyne (VS-G), which were then homogenized in phosphate-buffered saline (PBS). Any remaining tissues were eliminated by centrifugation at 3000 rpm for 3 minutes. The resulting venom sac extract from workers (VSE-W) and gynes (VSE-G), was transferred to new tubes and stored at -20 °C, until further analysis.
4.3. Venom Sac Extract Protein Quantitation
The protein content of the VSE-W and VSE-G was quantified using the Bradford method from Bio-Rad, Hercules, CA, USA.
4.4. Sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE).
Analysis of the VSE proteins was conducted through 12% SDS–PAGE (sodium dodecyl sulfate polyacrylamide gel electrophoresis) under reducing conditions, followed by staining with Sypro Ruby (Bio-Rad, Hercules, CA, USA). Finally, the protein bands were visualized using the ChemiDoc MP imaging system (Bio-Rad, Hercules, CA, USA).
4.5. Sample Preparation for Mass Spectrometric Analysis and Quantification Using Sequential Window Acquisition of All Theoretical Mass Spectra (SWATH-MS)
A total of 24 μg of protein was concentrated within a single band on an SDS–PAGE gel and subjected to manual digestion. Subsequently, the resulting peptides were dissolved in 0.1% formic acid for further analysis following established procedures [35]. To conduct quantitative proteomic analysis, a hybrid quadrupole-TOF mass spectrometer 6600+ (SCIEX, Framingham, MA, USA) coupled with a micro-liquid chromatography (LC) system Ekspert nLC425 (Eksigen, Dublin, CA, USA) was utilized. Data acquisition was performed using a combination of ProteinPilot v.5.0.1, PeakView v.2.2, MarkerView, and SWATH Acquisition MicroApp v.2.0 software package (SCIEX, Framingham, MA, USA). A customizer database containing Vespa + Vespa velutina + Apis mellifera + venom + toxins Uniprot databases [Available online: https://www.uniprot.org/ (accessed on 2 February 2023)] was employed. The peptide mixtures from sample pools were chromatographed for 40 minutes using data-dependent acquisition in positive ion mode to generate the MS/MS spectral libraries, as described in previous studies [57-59]. The false discovery rate was set at 1%, with a confidence score above 99%. This spectral library was then used to create the spectral window acquisition for SWATH-MS analysis. Subsequently, 4 μL from each sample were individually analyzed. The SWATH-MS method involves acquiring data in a cycle of 100-flight mass spectrometry (TOF MS/M) windows and quantitative analyses were supported by extracted ion chromatograms at both the MS1 and MS2 levels. Proteins with more than 10 peptides and seven transitions were selected for quantification, while shared or modified peptides were excluded. To compare the two Vespa crabro castes groups (VSE-G and SW), a Student’s t-test analysis was performed based on the averaged area sums of all the transitions derived for each protein. The resulting p values were used to indicate the significance of differences between the two groups. Proteins with a p value less than 0.05, a 1.5-fold increase or 0.66-fold decrease were considered differentially expressed.
4.6. Protein Functional Analysis
The quantified proteins were subjected to functional analysis for biological information related to molecular functions, biological processes, cellular components and protein families. We searched them against the protein databases UniProtKB (Available online: http://www.uniprot.org (accesed on 10 February 2023)). The uniprot codes of the proteins from D. novaeangliae were analyzed using the STRING: Functional protein association network (Available online: http://www.string-db.org (accessed on 14 June 2023)) to develop interaction maps, cluster analysis and analyze biological processes.
4.7. Statistical Analysis
Graphical representations comparing the SWATH-normalized area of expressed proteins in the VSE-G and VSE-W were created using box plots, presenting the median, whiskers (spanning from minimum to maximum values), and black dots indicating individual sample values. Additionally, a Volcano plot was generated by plotting the log2 fold change of the identified proteins against their corresponding adjusted -log10 p-value. A protein is considered up- or downregulated when the fold change (FC) is ≥ 1.5, and a p-value of ≤ 0.05 is considered statistically significant in all tests. The graphical analyses were performed using GraphPad Prism software (GraphPad Software, San Diego, CA, USA).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Supplementary File S1: Excel file with all quantified proteins with the individual values of SWATH normalized areas per protein and sample, and the FC and p value of the t-test; Supplementary File S2: Excel file download from UniProtKB with the GO terms for the biological process, the molecular function, protein family as well the cellular component of the 240 quantified proteins in the venom sac extracts from Vespa crabro; Table S1: Proteins from class Insecta with upregulated expression in Vespa crabro gyne venom sac extracts; Table S2: Proteins from class Insecta with downregulated expression in Vespa crabro gyne venom sac extracts.
Author Contributions
Conceptualization, C.V. and X.F.; methodology, M.A.-S.; software, S.B.B.; formal analysis, M.A.-S.; investigation, X.F.; resources, C.V.; data curation, M.A.-S. and X.F.; writing—original draft preparation, M.A.-S. and X.F.; writing—review and editing, M.A.-S., X.F. and C.V.; supervision, C.V.; project administration, M.A.-S.; funding acquisition, C.V. All authors have read and agreed to the published version of the manuscript. M.A.-S. and X.F. contributed equally to this work. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by grants from the Carlos III Institute of Health (Instituto de Salud Carlos III-ISCIII/PI19/01023/ (Co-funded by the European Union), and the Network for Research on Chronicity, Primary Care, and Health Promotion (Instituto de Salud Carlos IIIISCIII/RD21/0016/0022/ (Co-funded by the European Union). M.A.-S. was supported by the Galician Innovation Agency-Competitive Benchmark Groups (GAIN-GRC/IN607A/2021/02/Xunta de Galicia). X.F. was supported by Spanish Network for Addictive Disorders (Instituto de Salud Carlos III-ISCIII/RD16/0017/0018/ (Co-funded by the European Union).
Institutional Review Board Statement
The study protocol was approved by the “Comité Ético de Investigación Clínica (CEIC) de Galicia” (protocol code 2018/622 on 19 December 2018).
Informed Consent Statement
Not applicable.
Data Availability Statement
The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE partner repository with the dataset identifier PXD047955.
Acknowledgments
X.F. would like to acknowledge Dani Slizt for their field support and expertise in locating and removing the Vespa crabro nest.
Conflicts of Interest
The authors declare no conflict of interest. The funders were not involved in designing the study, collecting, analyzing, or interpreting data, writing the manuscript, or deciding to publish the results.
References
- White, J. Venomous animals: Clinical toxinology. In: Luch A, editor. Molecular, Clinical and Environmental Toxicology. Birkhäuser Verlag AG, 2010. p. 233–291. [CrossRef]
- Morsy, M.A.; Gupta, S.; Dora, C.P.; et al. Venoms classification and therapeutic uses: A narrative review. Eur. Rev. Med. Pharmacol. Sci. 2023, 27, 1633–1653. [Google Scholar] [CrossRef] [PubMed]
- Nelsen, D.R.; Nisani, Z.; Cooper, A.M.; Fox, G.A.; Gren, E.C.; Corbit, A.G.; Hayes, W.K. Poisons, toxungens, and venoms: Redefining and classifying toxic biological secretions and the organisms that employ them. Biol. Rev. Camb. Philos. Soc. 2014, 89, 450–465. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Aziz, T.M.; Soares, A.G.; Stockand, J.D. Advances in venomics: Modern separation techniques and mass spectrometry. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2020, 1160, 122352. [Google Scholar] [CrossRef]
- Calvete, J.J. Venomics: Integrative venom proteomics and beyond. Biochem. J. 2017, 474, 611–634. [Google Scholar] [CrossRef]
- Kaas, Q.; Craik, D.J. Bioinformatics-Aided Venomics. Toxins 2015, 7, 2159–2187. [Google Scholar] [CrossRef] [PubMed]
- Bass, A.; Needham, K.; Bennett, A.M.R. First record of Vespa crabro Linnaeus (Hymenoptera: Vespidae) in western North America with a review of recorded species of Vespa Linnaeus in Canada. Zootaxa 2022, 5154, 305–318. [Google Scholar] [CrossRef]
- Stetsun, H.A.; Matushkina, N.A. Sting morphology of the European hornet, Vespa crabro L., (Hymenoptera: Vespidae) re-examined. Entomol. Sci. 2020, 23, 416–429. [Google Scholar] [CrossRef]
- Erzen, R.; Koren, A.; Selb, J.; Bajrovic, N.; Lalek, N.; Kopac, P.; Zidarn, M.; Korosec, P.; Kosnik, M. Clinical, serological and basophil response to a wasp sting in patients with European hornet sting anaphylaxis. Clin. Exp. Allergy 2021, 51, 1641–1644. [Google Scholar] [CrossRef]
- Jacobson, R.S.; Hoffman, D.R. The European hornet (Vespa crabro) an emerging sting problem in the eastern U.S. J. Allergy Clin. Immunolog. 1985, 75, 121. [Google Scholar] [CrossRef]
- Kosnik, M.; Korosec, P.; Silar, M.; Music, E.; Erzen, R. Wasp venom is appropriate for immunotherapy of patients with allergic reaction to the European hornet sting. Croat. Med. J. 2002, 43, 25–27. [Google Scholar]
- Macchia, D.; Cortellini, G.; Mauro, M.; Meucci, E.; Quercia, O.; Manfredi, M.; Massolo, A.; Valentini, M.; Severino, M.; Passalacqua, G. Vespa crabro immunotherapy versus Vespula-venom immunotherapy in Vespa crabro allergy: A comparison study in field re-stings. World Allergy Organ. J. 2018, 11. [Google Scholar] [CrossRef]
- Blank, S.; Bazon, M.L.; Grosch, J.; Schmidt-Weber, C.B.; Brochetto-Braga, M.R.; Bilò, M.B.; Jakob, T. Antigen 5 Allergens of Hymenoptera Venoms and Their Role in Diagnosis and Therapy of Venom Allergy. Curr Allergy Asthma Rep. 2020, 20, 58. [Google Scholar] [CrossRef] [PubMed]
- Panzani, R.; Blanca, M.; Sánchez, F.; Juarez, C. Sensitivity to European wasps in a group of allergic patients in Marseille: Preliminary results. J. Investig. Allergol. Clin. Immunol. 1994, 4, 42–46. [Google Scholar]
- Severino, M.G.; Caruso, B.; Bonadonna, P.; Labardi, D.; Macchia, D.; Campi, P.; Passalacqua, G. Cross reactivity between European hornet and yellow jacket venoms. Eur. Ann. Allergy Clin. Immunol. 2010, 42, 141–145. [Google Scholar] [CrossRef]
- Korantzopoulos, P.; Kountouris, E.; Voukelatou, M.; Charaktsis, I.; Dimitroula, V.; Siogas, K. Acute myocardial infarction after a European hornet sting--a case report. Angiology 2006, 57, 383–386. [Google Scholar] [CrossRef] [PubMed]
- Lubrano, D.; Helias, P.; Bach, P.; Leclercq, M.; Fillet, G.; Lecomte, J.; Damas, J. [Case of fatal poisoning with hyperfibrinolysis by multiple wasp stings (Vespa crabro L.)]. Rev. Med. Liege 1985, 40, 844–846. [Google Scholar]
- Okutucu, S.; Sabanov, C.; Abdulhayoğlu, E.; Aksu, N.M.; Erbil, B.; Aytemir, K.; Ozkutlu, H. A rare cause of atrial fibrillation: A European hornet sting. Anadolu. Kardiyol. Derg. 2011, 11, 559–560. [Google Scholar] [CrossRef]
- Saggese, S. A new fatal case of Vespa crabro sting. Nuovi Ann. Ig. Microbiol. 1956, 7, 306–308. [Google Scholar] [PubMed]
- Antonicelli, L.; Bilò, M.B.; Napoli, G.; Farabollini, B.; Bonifazi, F. European hornet (Vespa crabro) sting: A new risk factor for life-threatening reaction in hymenoptera allergic patients? Eur. Ann. Allergy Clin. Immunol. 2003, 35, 199–203. [Google Scholar]
- Lee, S.H.; Baek, J.H.; Yoon, K.A. Differential Properties of Venom Peptides and Proteins in Solitary vs. Social Hunting Wasps. Toxins 2016, 8, 32. [Google Scholar] [CrossRef]
- Dashevsky, D.; Baumann, K.; Undheim, E.A.B.; Nouwens, A.; Ikonomopoulou, M.P.; Schmidt, J.O.; Ge, L.; Kwok, H.F.; Rodriguez, J.; Fry, B.G. Functional and Proteomic Insights into Aculeata Venoms. Toxins 2023, 15, 224. [Google Scholar] [CrossRef]
- Krishnakumari, V.; Nagaraj, R. Antimicrobial and hemolytic activities of crabrolin, a 13-residue peptide from the venom of the European hornet, Vespa crabro, and its analogs. J. Pept. Res. 1997, 50, 88–93. [Google Scholar] [CrossRef]
- Argiolas, A.; Pisano, J.J. Isolation and characterization of two new peptides, mastoparan C and crabrolin, from the venom of the European hornet, Vespa crabro. J. Biol. Chem. 1984, 259, 10106–10111. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, L.; Wu, Y.; Wang, L.; Ma, C.; Xi, X.; Bininda-Emonds, O.R.P.; Shaw, C.; Chen, T.; Zhou, M. Evaluation of the bioactivity of a mastoparan peptide from wasp venom and of its analogues designed through targeted engineering. Int. J. Biol. Sci. 2018, 14, 599–607. [Google Scholar] [CrossRef]
- Yoon, K.A.; Kim, K.; Nguyen, P.; Seo, J.B.; Park, Y.H.; Kim, K.; Seo, H.; Koh, Y.; Lee, S.H. Comparative bioactivities of mastoparans from social hornets Vespa crabro and Vespa analis. J. Asia-Pac Entomol. 2015, 18, 825–829. [Google Scholar] [CrossRef]
- Allalouf, D.; Ber, A.; Ishay, J. Hyaluronidase activity of extracts of venom sacs of a number of vespinae (Hymenoptera). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1972, 43, 119–123. [Google Scholar] [CrossRef]
- Hoffman, D.R.; Jacobson, R.S.; Zerboni, R. Allergens in hymenoptera venom. XIX. Allergy to Vespa crabro, the European hornet. Int. Arch. Allergy Appl. Immunol. 1987, 84, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, D.R. Allergens in Hymenoptera venom XXV: The amino acid sequences of antigen 5 molecules and the structural basis of antigenic cross-reactivity. J. Allergy Clin. Immunol. 1993, 92, 707–716. [Google Scholar] [CrossRef]
- Yoon, K.A.; Kim, K.; Nguyen, P.; Seo, J.B.; Park, Y.H.; Kim, K.G.; Seo, H.-Y.; Koh, Y.H.; Lee, S.H. Comparative functional venomics of social hornets Vespa crabro and Vespa analis. J. Asia-Pac Entomol. 2015, 18, 815–823. [Google Scholar] [CrossRef]
- Mueller, U.; Elliott, W.; Reisman, R.; Ishay, J.; Walsh, S.; Steger, R.; Wypych, J.; Arbesman, C. Comparison of biochemical and immunologic properties of venoms from four hornet species. J. Allergy Clin. Immunol. 1981, 67, 290–298. [Google Scholar] [CrossRef]
- Turillazzi, S.; Bruschini, C.; Lambardi, D.; Francese, S.; Spadolini, I.; Mastrobuoni, G. Comparison of the medium molecular weight venom fractions from five species of common social wasps by MALDI-TOF spectra profiling. J. Mass Spectrom. 2007, 42, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.A.; Kim, K.; Kim, W.J.; Bang, W.Y.; Ahn, N.H.; Bae, C.H.; Yeo, J.H.; Lee, S.H. Characterization of Venom Components and Their Phylogenetic Properties in Some Aculeate Bumblebees and Wasps. Toxins 2020, 12, 47. [Google Scholar] [CrossRef]
- Yoon, K.A.; Kim, W.J.; Lee, S.H. Expression profiles of venom components in some social hymenopteran species over different post-capture periods. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2022, 253, 109247. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Sampedro, M.; Feás, X.; Bravo, S.B.; Chantada-Vázquez, M.P.; Vidal, C. Proteomics of Vespa velutina nigrithorax Venom Sac Queens and Workers: A Quantitative SWATH-MS Analysis. Toxins 2023, 15, 266. [Google Scholar] [CrossRef] [PubMed]
- Rotello, R.J.; Veenstra, T.D. Mass Spectrometry Techniques: Principles and Practices for Quantitative Proteomics. Curr. Protein Pept. Sci. 2021, 22, 121–133. [Google Scholar] [CrossRef]
- Ludwig, C.; Gillet, L.; Rosenberger, G.; Amon, S.; Collins, B.C.; Aebersold, R. Data-independent acquisition-based SWATH-MS for quantitative proteomics: A tutorial. Mol. Syst. Biol. 2018, 14. [Google Scholar] [CrossRef]
- Ishay, J.; Shved, A.; Gitter, S. Lethality of venom sac extracts of the oriental hornet (Vespa orientalis) as related to ontogenesis. Toxicon. 1977, 15, 307–315. [Google Scholar] [CrossRef]
- Le, T.N.; Da Silva, D.; Colas, C.; Darrouzet, E.; Baril, P.; Leseurre, L.; Maunit, B. Asian hornet Vespa velutina nigrithorax venom: Evaluation and identification of the bioactive compound responsible for human keratinocyte protection against oxidative stress. Toxicon 2020, 176, 1–9. [Google Scholar] [CrossRef]
- Koludarov, I.; Velasque, M.; Timm, T.; Greve, C.; Ben Hamadou, A.; Kumar Gupta, D.; Lochnit, G.; el Heinzinger, M.; Vilcinskas, A.; Gloag, R.; et al. A common venomous ancestor? Prevalent bee venom genes evolved before the aculeate stinger while few major toxins are bee-specific. bioRxiv 2022. [Google Scholar] [CrossRef]
- Brown, J.L.; Morales, V.; Summers, K. A key ecological trait drove the evolution of biparental care and monogamy in an amphibian. Am. Nat. 2010, 175, 436–446. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, F.; Wang, X.L.; Wang, P.H.; Ma, S.L.; Yang, Y.; Ye, W.-G.; Diao, K.-Y.; Dai, P.L. Midgut Bacterial Communities of Vespa velutina Lepeletier (Hymenoptera: Vespidae). Front. Ecol. Evol. 2022, 10, 934054. [Google Scholar] [CrossRef]
- Suenami, S.; Konishi, N.M.; Miyazaki, R. Community analysis of gut microbiota in hornets, the largest eusocial wasps, Vespa mandarinia and V. simillima. Sci. Rep. 2019, 9, 9830. [Google Scholar] [CrossRef]
- Cini, A.; Meriggi, N.; Bacci, G.; Cappa, F.; Vitali, F.; Cavalieri, D.; Cervo, R. Gut microbial composition in different castes and developmental stages of the invasive hornet Vespa velutina nigrithorax. Sci. Total Environ. 2020, 745, 140873. [Google Scholar] [CrossRef]
- Chantawannakul, P.; Ramsey, S.; vanEngelsdorp, D.; Khongphinitbunjong, K.; Phokasem, P. Tropilaelaps mite: An emerging threat to European honey bee. Curr. Opin. Insect Sci. 2018, 26, 69–75. [Google Scholar] [CrossRef]
- Feás, X.; Vidal, C.; Remesar, S. What We Know about Sting-Related Deaths? Human Fatalities Caused by Hornet, Wasp and Bee Stings in Europe (1994–2016). Biology 2022, 11, 282. [Google Scholar] [CrossRef]
- Feás, X. Human Fatalities Caused by Hornet, Wasp and Bee Stings in Spain: Epidemiology at State and Sub-State Level from 1999 to 2018. Biology 2021, 10, 73. [Google Scholar] [CrossRef]
- Vidal, C. The Asian wasp Vespa velutina nigrithorax: Entomological and allergological characteristics. Clin. Exp. Allergy 2022, 52, 489–498. [Google Scholar] [CrossRef]
- Feás, X.; Vázquez-Tato, M.P.; Seijas, J.A.; Pratima GNikalje, A.; Fraga-López, F. Extraction and Physicochemical Characterization of Chitin Derived from the Asian Hornet, Vespa velutina Lepeletier 1836 (Hym.: Vespidae). Molecules 2020, 25, 384. [Google Scholar] [CrossRef] [PubMed]
- Feás Sánchez, X.; Charles, R.J. Notes on the nest architecture and colony composition in winter of the yellow-legged Asian Hornet, Vespa velutina Lepeletier 1836 (Hym.: Vespidae), in its introduced habitat in Galicia (NW Spain). Insects 2019, 10, 237. [Google Scholar] [CrossRef] [PubMed]
- Feás, X.; Vidal, C.; Vázquez-Tato, M.P.; Seijas, J.A. Asian Hornet, Vespa velutina Lepeletier 1836 (Hym.: Vespidae), Venom Obtention Based on an Electric Stimulation Protocol. Molecules 2021, 27, 138. [Google Scholar] [CrossRef] [PubMed]
- Vidal, C.; Armisén, M.; Monsalve, R.; Gómez-Rial, J.; González-Fernández, T.; Carballada, F.; Lombardero, M.; González-Quintela, A. Vesp v 5 and glycosylated Vesp v 1 are relevant allergens in Vespa velutina nigrithorax anaphylaxis. Clin. Exp. Allergy 2020, 50, 1424–1427. [Google Scholar] [CrossRef] [PubMed]
- Vidal, C.; Armisén, M.; Monsalve, R.; González-Vidal, T.; Lojo, S.; López-Freire, S.; Méndez, P.; Rodríguez, V.; Romero, L.; Galán, A.; et al. Anaphylaxis to Vespa velutina nigrithorax: Pattern of Sensitization for an Emerging Problem in Western Countries.J. Investig. Allergol. Clin. Immunol. 2021, 31, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Vázquez, V.; Gómez-Rial, J.; Monsalve, R.I.; Vidal, C. Consistency of Determination of sIgE and the Basophil Activation Test in Vespa velutina nigrithorax Allergy. J. Investig. Allergol. Clin. Immunol. 2022, 32, 143–145. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Vázquez, V.; Armisén, M.; Gómez-Rial, J.; Lamas-Vázquez, B.; Vidal, C. Immunotherapy with Vespula venom for Vespa velutina nigrithorax anaphylaxis: Preliminary clinical and immunological results. Clin. Exp. Allergy 2022, 52, 345–347. [Google Scholar] [CrossRef]
- Ruiz-Leon, B.; Serrano, P.; Vidal, C.; Moreno-Aguilar, C. Management of Double Sensitization to Vespids in Europe. Toxins 2022, 14, 126. [Google Scholar] [CrossRef] [PubMed]
- Peñas-Martínez, J.; Barrachina, M.N.; Cuenca-Zamora, E.J.; Luengo-Gil, G.; Bravo, S.B.; Caparrós-Pérez, E.; Teruel-Montoya, R.; Eliseo-Blanco, J.; Vicente, V.; García, Á.; et al. Qualitative and quantitative comparison of plasma exosomes from neonates and adults. Int. J. Mol. Sci. 2021, 22, 1926. [Google Scholar] [CrossRef]
- Camino, T.; Lago-Baameiro, N.; Sueiro, A.; Bravo, S.B.; Couto, I.; Santos, F.F.; Teruel-Montoya, R.; Eliseo-Blanco, J.; Vicente, V.; García, Á.; et al. Brown Adipose Tissue Sheds Extracellular Vesicles That Carry Potential Biomarkers of Metabolic and Thermogenesis Activity Which Are Affected by High Fat Diet Intervention. Int. J. Mol. Sci. 2022, 23, 10826. [Google Scholar] [CrossRef]
- Chantada-Vázquez, M.D.P.; Conde-Amboage, M.; Graña-López, L.; Vázquez-Estévez, S.; Bravo, S.B.; Núñez, C. Circulating Proteins Associated with Response and Resistance to Neoadjuvant Chemotherapy in HER2-Positive Breast Cancer. Cancers 2022, 14, 1087. [Google Scholar] [CrossRef]
Figure 1.
The sampled European Hornet Vespa crabro (Linnaeus, 1758) (Hymenoptera: Vespidae) nest after being removed; (a) Lateral view; (b) Lateral view after peeling away the paper envelope showing the internal chamber with the combs (n=8); (c) From top to bottom: queen, gyne, worker and male. Scales bars: a = 4 cm; b = 3 cm; c = 0.5 cm. Photo Author: X. Feás.
Figure 1.
The sampled European Hornet Vespa crabro (Linnaeus, 1758) (Hymenoptera: Vespidae) nest after being removed; (a) Lateral view; (b) Lateral view after peeling away the paper envelope showing the internal chamber with the combs (n=8); (c) From top to bottom: queen, gyne, worker and male. Scales bars: a = 4 cm; b = 3 cm; c = 0.5 cm. Photo Author: X. Feás.

Figure 2.
European hornet Vespa crabro (Linnaeus, 1758) (Hymenoptera: Vespidae) stinging apparatus (up) and venom sac (down) of (a) gynes and (b) workers. Scale in mm. Photo Author: M. Alonso-Sampedro.
Figure 2.
European hornet Vespa crabro (Linnaeus, 1758) (Hymenoptera: Vespidae) stinging apparatus (up) and venom sac (down) of (a) gynes and (b) workers. Scale in mm. Photo Author: M. Alonso-Sampedro.

Figure 3.
Analysis of the venom sac extracts (VSE) of gynes (VSE-G) and workers (VSE-W) of European hornet Vespa crabro (Linnaeus, 1758) (Hymenoptera: Vespidae); (a) Box plot with median, whiskers min. to max., and black dots for the individual values of total proteins in micrograms (μg) per venom sac (ns, no statistical significance Mann-Whitney test); (b) Sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE), arrows indicate the bands corresponding to the known potential allergens according to their molecular mass: Vesp c 5, venom allergen 5 (23 kDa); Vesp c 1, phospholipase A1 (34 kDa); and Vesp c 2, hyaluronidase (40 kDa). kDa, kilodalton; ST, precision plus protein standard.
Figure 3.
Analysis of the venom sac extracts (VSE) of gynes (VSE-G) and workers (VSE-W) of European hornet Vespa crabro (Linnaeus, 1758) (Hymenoptera: Vespidae); (a) Box plot with median, whiskers min. to max., and black dots for the individual values of total proteins in micrograms (μg) per venom sac (ns, no statistical significance Mann-Whitney test); (b) Sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE), arrows indicate the bands corresponding to the known potential allergens according to their molecular mass: Vesp c 5, venom allergen 5 (23 kDa); Vesp c 1, phospholipase A1 (34 kDa); and Vesp c 2, hyaluronidase (40 kDa). kDa, kilodalton; ST, precision plus protein standard.
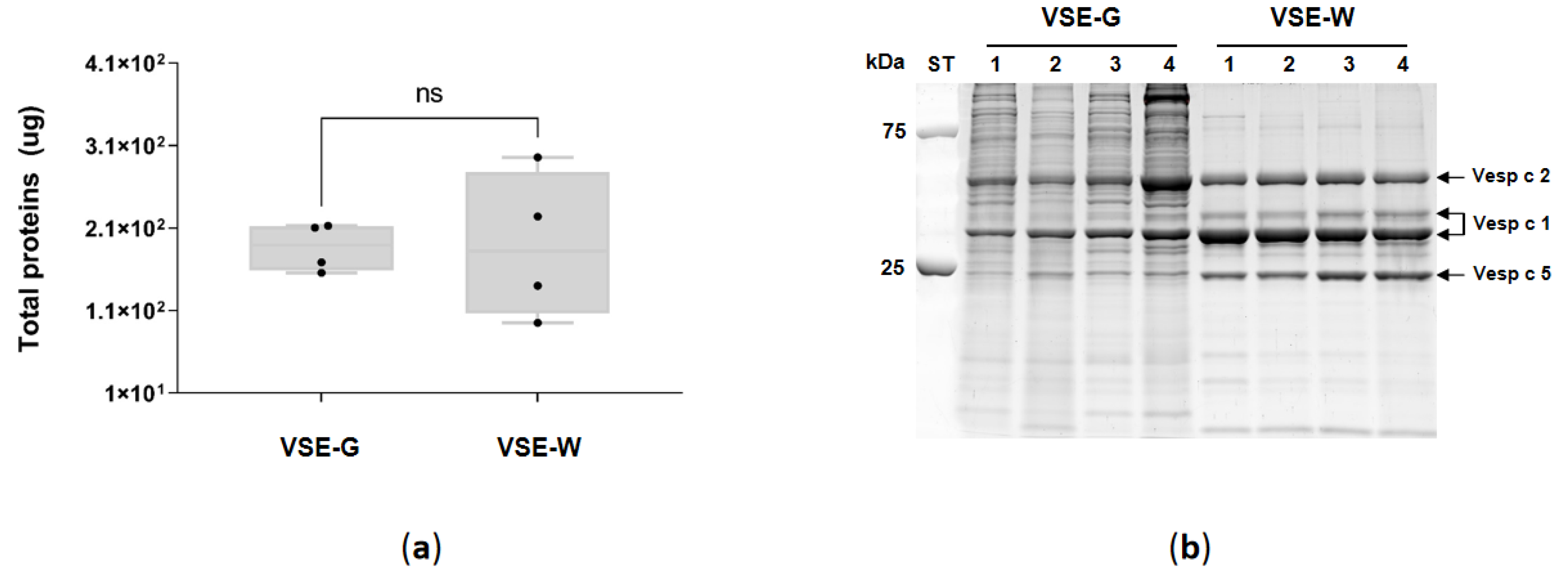
Figure 4.
Volcano plot of the venom sac extract (VSE) quantitative proteomics data of the European hornet Vespa crabro (Linnaeus, 1758) (Hymenoptera: Vespidae) female castes (gynes and workers). VSE proteins (shown as spots) are ranked according to their statistical p-value (y-axis) as –log10 and their relative abundance ratio as log2 fold change (x-axis). Off-centered spots are those that vary the most between the venom sacs of gynes (G) and workers (W). The cut-offs for significant changes are fold change ≥ 1.5 (dashed line red and green) and p ≤ 0.05 (t-test). Green spots show the upregulated proteins in VSE-G, red spots show the downregulated proteins in VSE-G, and black spots show the non-dysregulated proteins between the two groups. The known potential allergens are showed as blue spots: Vesp v 5, venom allergen 5; Vesp v 1, phospholipase A1; Vesp v 2A, hyaluronidase A; Vesp v 2B, hyaluronidase B; VT-2b, phospholipase A1 verotoxin-2b; Ves v 6, dipeptidylpeptidase IV; Ves v 6, vitellogenin. VSE-G: venom sac extracts of gynes; VSE-W: venom sac extracts of workers. The nomenclature of proteins is based on software identification.
Figure 4.
Volcano plot of the venom sac extract (VSE) quantitative proteomics data of the European hornet Vespa crabro (Linnaeus, 1758) (Hymenoptera: Vespidae) female castes (gynes and workers). VSE proteins (shown as spots) are ranked according to their statistical p-value (y-axis) as –log10 and their relative abundance ratio as log2 fold change (x-axis). Off-centered spots are those that vary the most between the venom sacs of gynes (G) and workers (W). The cut-offs for significant changes are fold change ≥ 1.5 (dashed line red and green) and p ≤ 0.05 (t-test). Green spots show the upregulated proteins in VSE-G, red spots show the downregulated proteins in VSE-G, and black spots show the non-dysregulated proteins between the two groups. The known potential allergens are showed as blue spots: Vesp v 5, venom allergen 5; Vesp v 1, phospholipase A1; Vesp v 2A, hyaluronidase A; Vesp v 2B, hyaluronidase B; VT-2b, phospholipase A1 verotoxin-2b; Ves v 6, dipeptidylpeptidase IV; Ves v 6, vitellogenin. VSE-G: venom sac extracts of gynes; VSE-W: venom sac extracts of workers. The nomenclature of proteins is based on software identification.
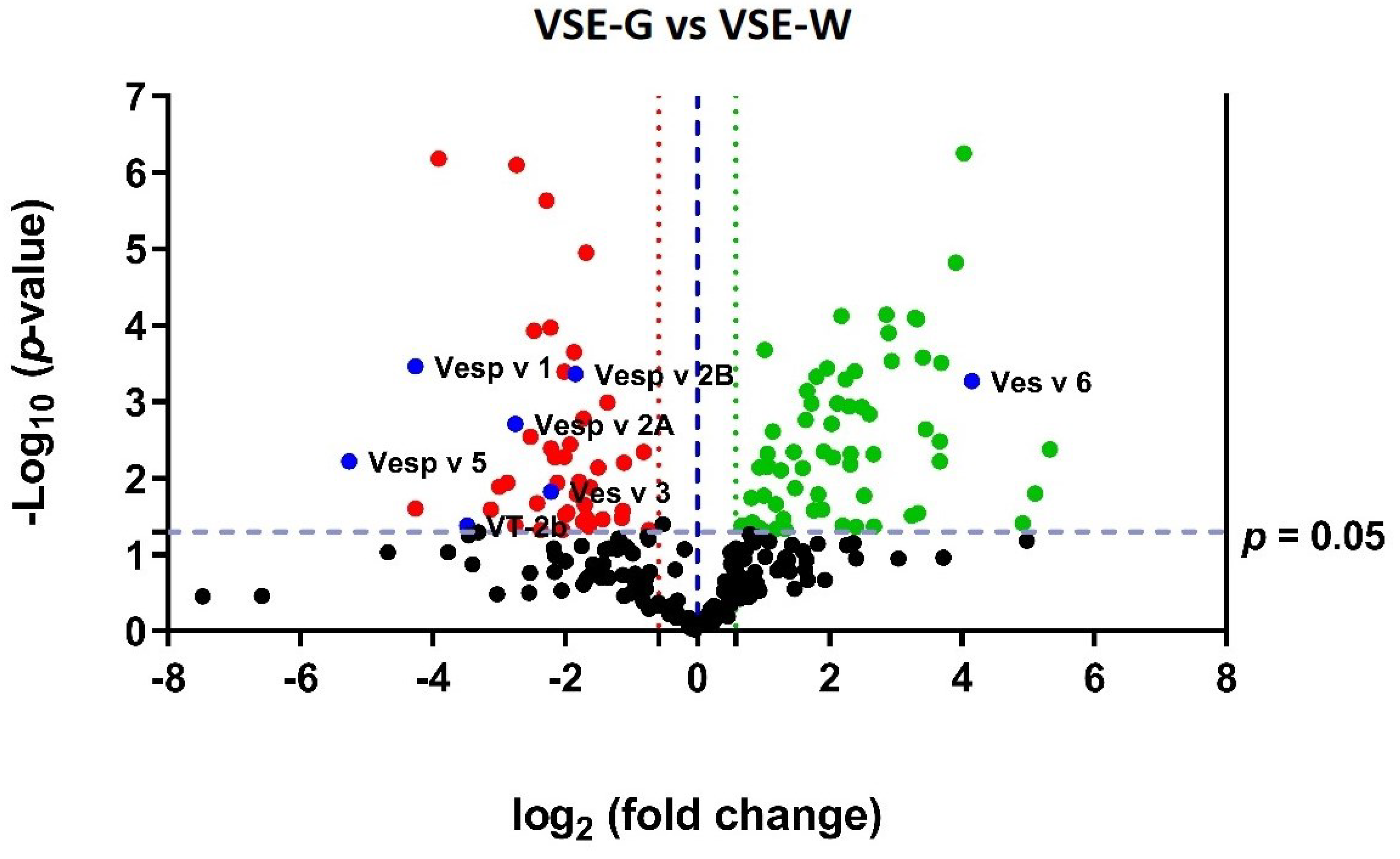
Figure 5.
(A) Biological processes and (B) molecular functions mainly related to the differentially expressed proteins in the venom sac extracts (VSE) of European hornet Vespa crabro (Linnaeus, 1758) (Hymenoptera: Vespidae) gynes (G) and workers (W). The histograms represent the main categories for each gene ontology (GO) term in which differentially expressed proteins were involved (p < 0.05). The y-axis shows the number of individual proteins in each GO term, and the x-axis shows the GO term. Red, VSE-G downregulated; green, VSE-G upregulated. The nomenclature of proteins is based on software identification.
Figure 5.
(A) Biological processes and (B) molecular functions mainly related to the differentially expressed proteins in the venom sac extracts (VSE) of European hornet Vespa crabro (Linnaeus, 1758) (Hymenoptera: Vespidae) gynes (G) and workers (W). The histograms represent the main categories for each gene ontology (GO) term in which differentially expressed proteins were involved (p < 0.05). The y-axis shows the number of individual proteins in each GO term, and the x-axis shows the GO term. Red, VSE-G downregulated; green, VSE-G upregulated. The nomenclature of proteins is based on software identification.
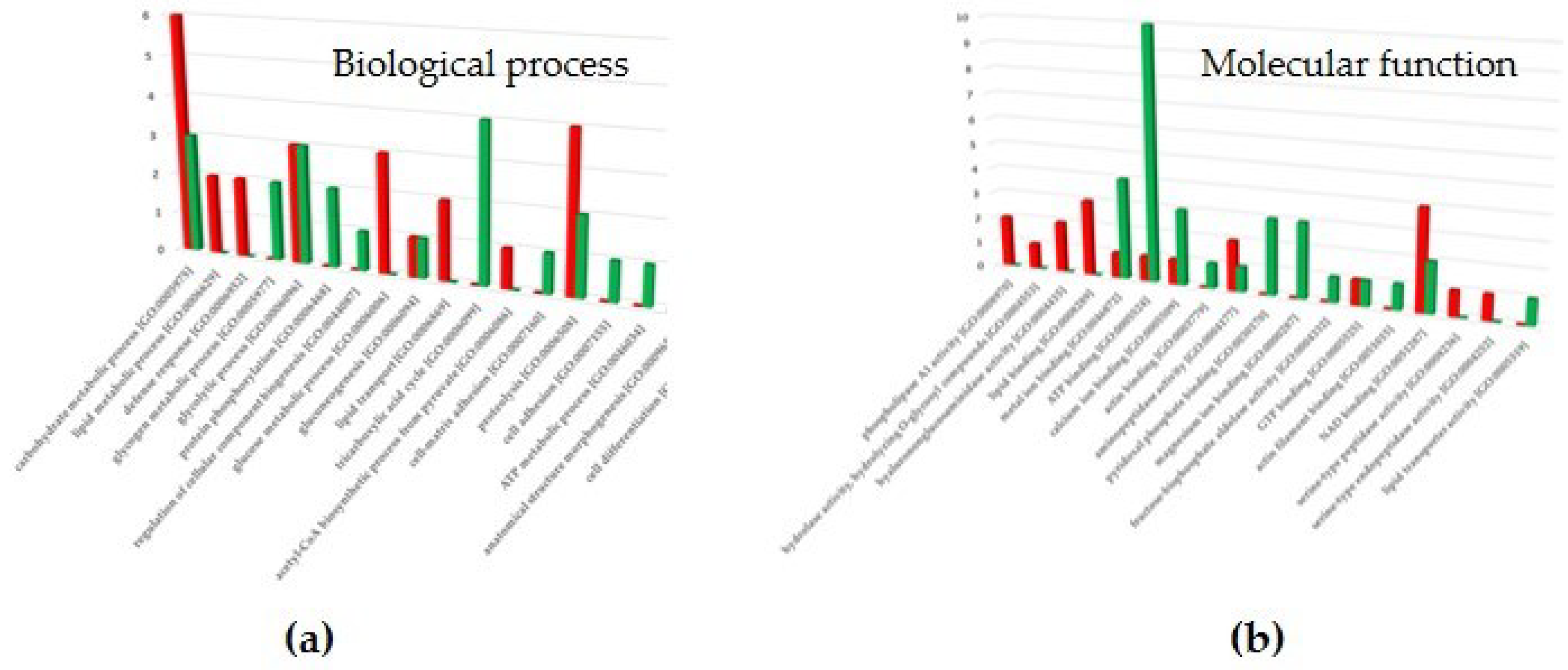
Figure 6.
Representation of the SWATH-MS normalized area of the known potential allergens in the venom sac extract (VSE) of the European hornet (EH) Vespa crabro (Linnaeus, 1758) (Hymenoptera: Vespidae) gynes (G) and workers (W). Box plot with median, whiskers min to max, and black dots for the individual values of the normalized area. Statistical differences by Mann-Whitney test, * p < 0.05. VSE-G: venom sac extracts of gynes; VSE-W: venom sac extracts of workers. The nomenclature of proteins is based on software identification.
Figure 6.
Representation of the SWATH-MS normalized area of the known potential allergens in the venom sac extract (VSE) of the European hornet (EH) Vespa crabro (Linnaeus, 1758) (Hymenoptera: Vespidae) gynes (G) and workers (W). Box plot with median, whiskers min to max, and black dots for the individual values of the normalized area. Statistical differences by Mann-Whitney test, * p < 0.05. VSE-G: venom sac extracts of gynes; VSE-W: venom sac extracts of workers. The nomenclature of proteins is based on software identification.
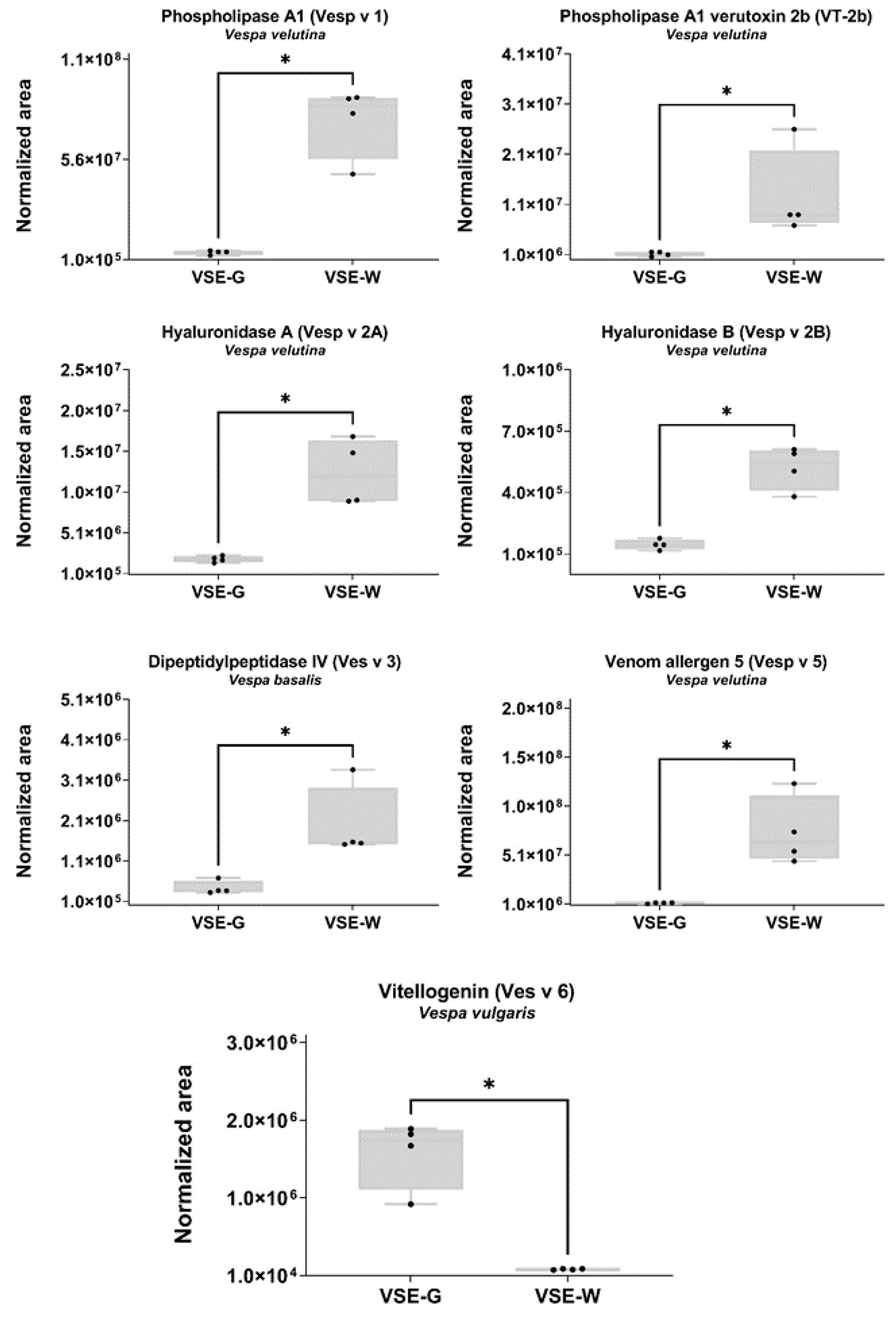
Figure 7.
STRING protein-protein interaction (PPI) network analyses of the 29 proteins quantified from Dufourea novaeanliae in the venom sac extract (VSE) of the European hornet Vespa crabro (Linnaeus, 1758) (Hymenoptera: Vespidae) gynes and workers. (A) Network clustered using k-means clustering: Cluster 1 (green bubbles) with 8 proteins; Cluster 2 (blue bubbles) with 10 proteins; and Cluster 3 (red bubbles) with 11 proteins. The main biological process of proteins in cluster 1 (B), cluster 2 (C), and cluster 3 (D). The colored proteins in the clusters are involved in the indicated biological process. Classification of proteins was based on Gene Ontology (GO) biological processes. All the biological processes that are referred to present a false Discovery rate (FDR) < 0.05. VSE-G: venom sac extracts of gynes; VSE-W: venom sac extracts of workers. The nomenclature of proteins is based on software identification.
Figure 7.
STRING protein-protein interaction (PPI) network analyses of the 29 proteins quantified from Dufourea novaeanliae in the venom sac extract (VSE) of the European hornet Vespa crabro (Linnaeus, 1758) (Hymenoptera: Vespidae) gynes and workers. (A) Network clustered using k-means clustering: Cluster 1 (green bubbles) with 8 proteins; Cluster 2 (blue bubbles) with 10 proteins; and Cluster 3 (red bubbles) with 11 proteins. The main biological process of proteins in cluster 1 (B), cluster 2 (C), and cluster 3 (D). The colored proteins in the clusters are involved in the indicated biological process. Classification of proteins was based on Gene Ontology (GO) biological processes. All the biological processes that are referred to present a false Discovery rate (FDR) < 0.05. VSE-G: venom sac extracts of gynes; VSE-W: venom sac extracts of workers. The nomenclature of proteins is based on software identification.
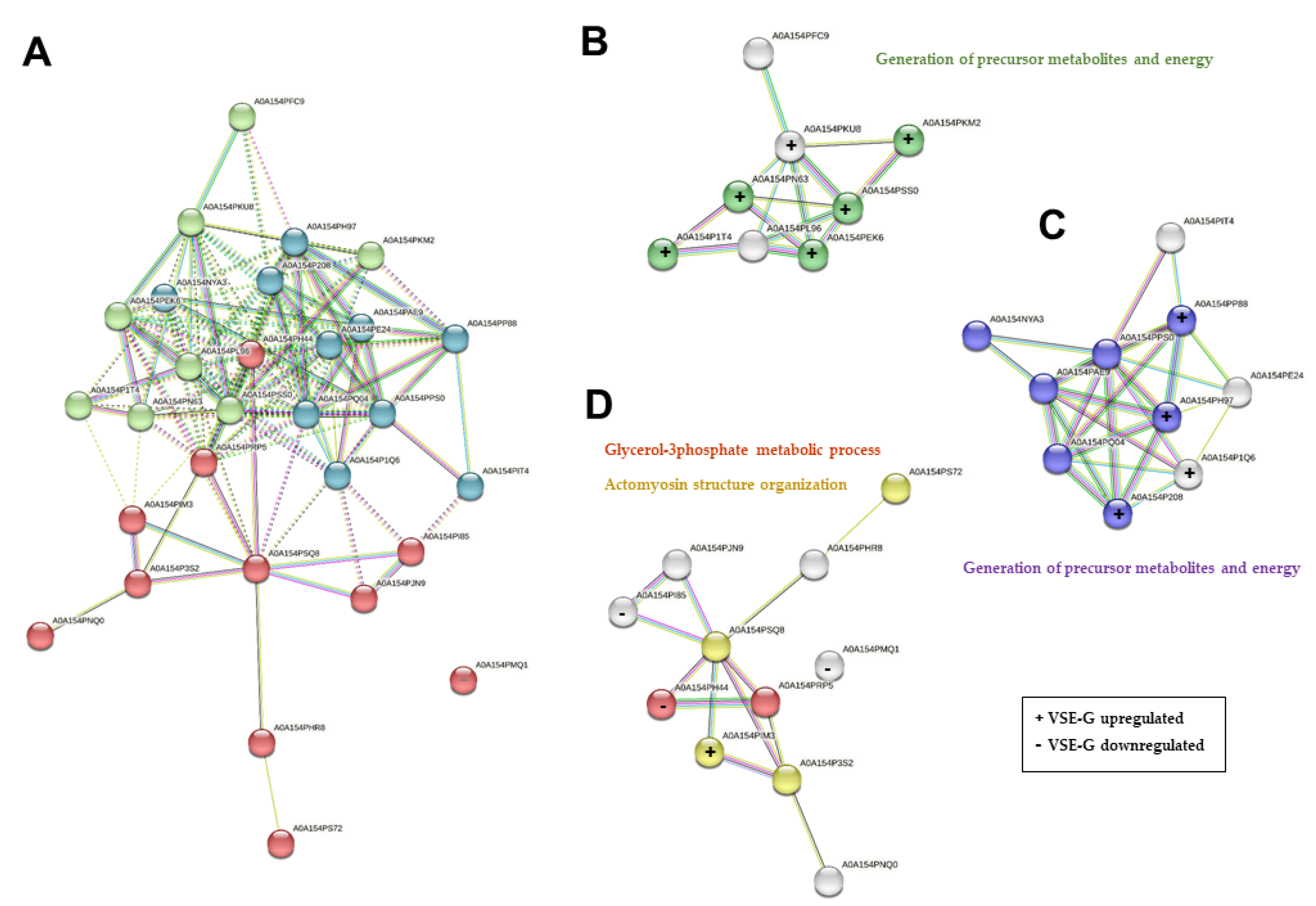
Figure 8.
The nest of the European hornet Vespa crabro (Linnaeus, 1758) (Hymenoptera: Vespidae) was collected in Pazos de Borbén (coordinates: 42.3451657, -8.4453559), depicted in blue on the hypsometric map of Galicia.
Figure 8.
The nest of the European hornet Vespa crabro (Linnaeus, 1758) (Hymenoptera: Vespidae) was collected in Pazos de Borbén (coordinates: 42.3451657, -8.4453559), depicted in blue on the hypsometric map of Galicia.
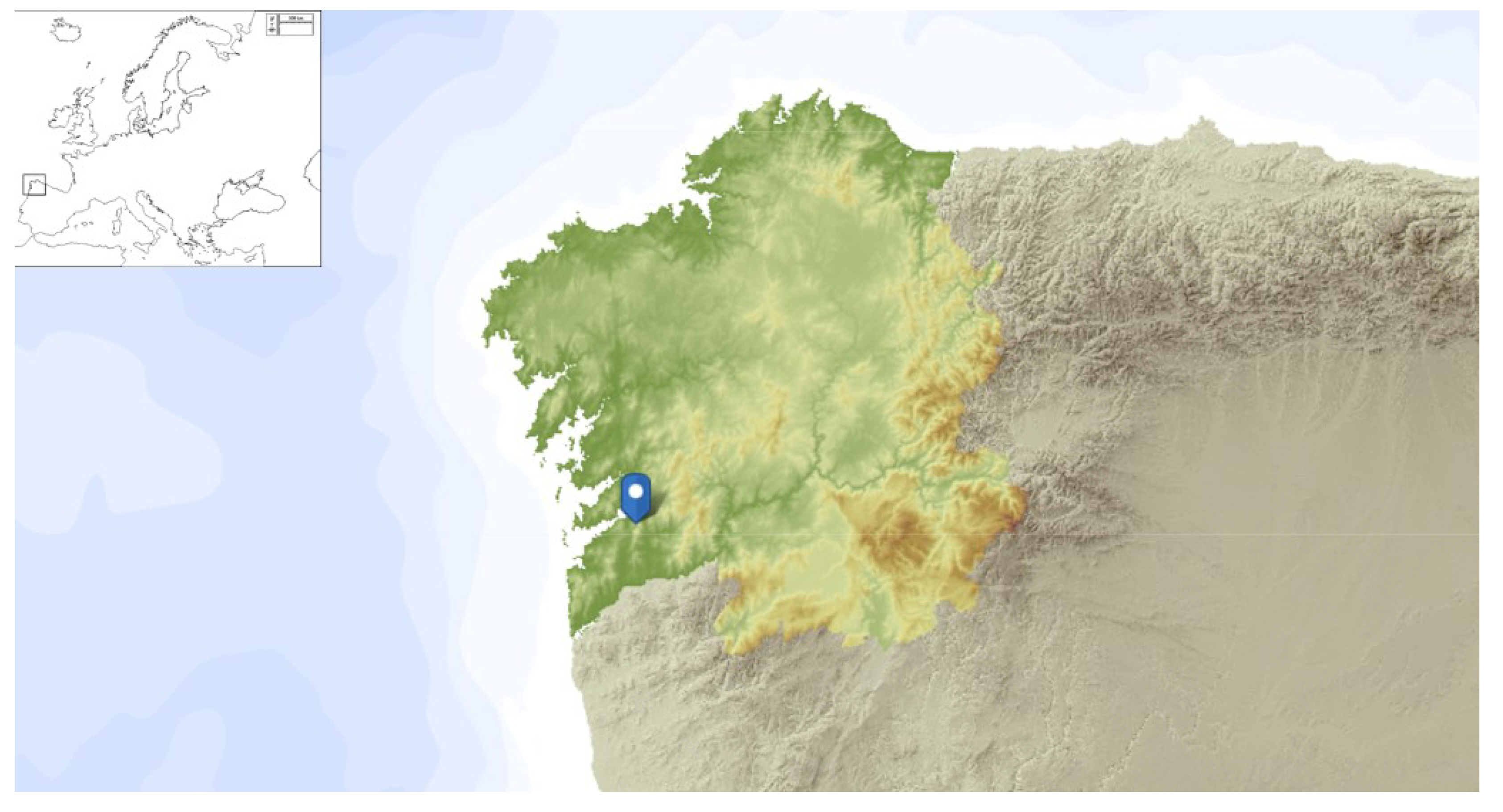
Figure 9.
(a) An Eucalyptus logging point; (b) Detail of a specimen of European hornet Vespa crabro (Linnaeus, 1758) (left) and Vespa velutina (Lepeletier, 1836) (right) on cut eucalyptus trunks.. Photo Author: X. Feás.
Figure 9.
(a) An Eucalyptus logging point; (b) Detail of a specimen of European hornet Vespa crabro (Linnaeus, 1758) (left) and Vespa velutina (Lepeletier, 1836) (right) on cut eucalyptus trunks.. Photo Author: X. Feás.

Figure 10.
The nest of the European Hornet Vespa crabro (Linnaeus, 1758) (Hymenoptera: Vespidae) found and subsequently removed (a) A side view; (b) a bottom view showing its opening. Photo Author: D. Slizt.
Figure 10.
The nest of the European Hornet Vespa crabro (Linnaeus, 1758) (Hymenoptera: Vespidae) found and subsequently removed (a) A side view; (b) a bottom view showing its opening. Photo Author: D. Slizt.


Table 1.
Classification of the 240 proteins (pp) found in the European Hornet Vespa crabro (Linnaeus, 1758) (Hymenoptera: Vespidae) venom sac extracts by “Sequential Window Acquisition of all Theoretical Fragment Ion Mass Spectra” (SWATH-MS) according to the species, family, order and class to which they belong.
Table 1.
Classification of the 240 proteins (pp) found in the European Hornet Vespa crabro (Linnaeus, 1758) (Hymenoptera: Vespidae) venom sac extracts by “Sequential Window Acquisition of all Theoretical Fragment Ion Mass Spectra” (SWATH-MS) according to the species, family, order and class to which they belong.
| Class | Order | Family | Scientific name | Common name | nº | Group |
|---|---|---|---|---|---|---|
| Actinopterygii | Cyprinodontiformes | Nothobranchiidae | Nothobranchius furzeri (Jubb, 1971) | African Turquoise killifish | 1 | NOTFU |
| Perciformes | Siganidae | Siganus canaliculatus (Park, 1797) | White-spotted spinefoot | 1 | SIGCA | |
| Amphibia | Anura | Dendrobatidae | Ranitomeya imitator (Schulte, 1986) | Mimic poison frog | 17 | 9NEOB |
| Arachnida | Mesostigmata | Laelapidae | Tropilaelaps mercedesae (Delfinado & Baker, 1961) | Tropilaelaps mite | 2 | 9ACAR |
| Scorpiones | Buthidae | Centruroides hentzi (Banks, 1940) | Hentz striped scorpion | 2 | 9SCOR | |
| Caraboctonidae | Hadrurus spadix (Stahnke, 1940) | Black back scorpion | 1 | |||
| Bacilli | Lactobacillales | Lactobacillaceae | Lactobacillus apis (Killer et al. 2014) | Lactic acid bacteria | 1 | 9LACO |
| Streptococcaceae | Lactococcus lactis (Lister 1873) Schleifer et al. 1986 | Lactic acid bacteria | 1 | 9LACT | ||
| Clitellata | Rhynchobdellida | Glossiphoniidae | Helobdella robusta (Shankland, Bissen & Weisblat, 1992) | Californian leech | 1 | HELRO |
| Insecta | Coleoptera | Ptilodactylidae | Anchycteis velutina (Horn, 1880) | Toe-winged beetle | 1 | |
| Hymenoptera | Apidae | Apis cerana (Fabricius, 1793) | Asian honey bee | 31 | APICE | |
| Apis cerana subsp. cerana (Fabricius, 1793) | Asian honey bee | 4 | APICC | |||
| Apis cerana subsp. indica (Fabricius, 1793) | Indian honey bee | 1 | ||||
| Apis mellifera (Linnaeus, 1758) | European honey bee | 41 | APIME | |||
| Apis mellifera ligustica (Spinola, 1806) | Italian honey bee | 2 | APILI | |||
| Bombus festivus(Smith, 1861) | Bumblebee of Sichuan | 1 | ||||
| Bombus humilis(Illiger, 1806) | Brown-banded carder bumblebee | 1 | BOMHU | |||
| Bombus terrestris (Linnaeus, 1758) | Buff-tailed bumblebee | 44 | BOMTE | |||
| Frieseomelitta varia(Lepeletier, 1836) | Yellow marmalade bee | 28 | 9HYME | |||
| Halictidae | Agapostemon leunculus (Vachal, 1903) | Sweat bee | 1 | 9COLE | ||
| Dufourea novaeangliae (Robertson, 1897) | Sweat bee | 29 | DUFNO | |||
| Vespidae | Vespa basalis (Smith, 1852) | Black-bellied hornet | 1 | VESBA | ||
| Vespa crabro (Linnaeus, 1758) | European hornet | 1 | VESCR | |||
| Vespa magnifica (Smith, 1852) | Magnifica hornet | 1 | VESMG | |||
| Vespa mandarinia (Smith, 1852) | Northern giant hornet | 1 | ||||
| Vespa mandarinia subsp. Japonica (Smith, 1852) | Northern giant hornet | 1 | VESMA | |||
| Vespa simillima subsp. xanthoptera (Cameron, 1903) | Yellow hornet | 4 | VESXA | |||
| Vespa velutina (Lepeletier, 1836) | Yellow-legged Asian hornet | 6 | VESVE | |||
| Vespula vulgaris(Linnaeus, 1758) | Common wasp | 1 | VESVU | |||
| Mammalia | Primates | Hominidae | Homo sapiens (Linnaeus, 1758) | Human | 2 | HUMAN |
| Microsporidia | Nosematida | Nosematidae | Nosema apis (Zander 1909) | Nosema | 1 | 9MICR |
| Pisoniviricetes | Picornavirales | Iflaviridae | Deformed wing virus | Deformed wing virus | 1 | 9VIRU |
| Reptilia | Squamata | Viperidae | Crotalus adamanteus (Palisot de Beauvois, 1799) | Eastern diamondback rattlesnake | 1 | CROAD |
| γ-proteobacteria | Enterobacterales | Enterobacteriaceae | Klebsiella sp. RIT-PI-d | Klebsiella | 1 | 9ENTR |
| Cronobacter sakazakii (Farmer et al. 1980) Iversen et al. 2007. | Cronobacter | 1 | CROSK | |||
| Citrobacter freundii (Braak 1928) Werkman & Gillen 1932 | Citrobacter | 1 | CITFR | |||
| Escherichia coli (Migula 1895)Castellani & Chalmers 1919 | coli | 5 | ECOLX |
Table 2.
Proteins from other classes with dysregulated expression in the European hornet Vespa crabro (Linnaeus, 1758) (Hymenoptera: Vespidae) gyne and worker venom sac extracts.
Table 2.
Proteins from other classes with dysregulated expression in the European hornet Vespa crabro (Linnaeus, 1758) (Hymenoptera: Vespidae) gyne and worker venom sac extracts.
| Family/Protein | Uniprot code | Specie | P Value (t test) | FC (VSE-G/VSE-W) |
|---|---|---|---|---|
| Aldo-keto reductase/alcohol dehydrogenase (NADP(+)) | J3SBS4 | adamanteus | < 0.001 | 16.364 |
| Aldehyde dehydrogenase/Multifunctional fusion protein | A0A1W7RAT0 | H. spadix | 0.007 | 4.969 |
| Heat shock protein/Heat shock 70 kDa protein | A0A2I9LP41 | C. hentzi | 0.004 | 2.081 |
| Intermediate filament/hypothetical protein | A0A822AXZ5 | R. imitator | 0.027 | 0.458 |
| Transferrin/Serotransferrin | A0A822GLX5 | R. imitator | 0.011 | 0.289 |
| Apolipoprotein A1/A4/E/hypothetical protein | A0A822ID78 | R. imitator | 0.004 | 0.216 |
| A0A821UX93 | imitator | 0.006 | 0.465 | |
| Peptidase/Proteasome subunit alpha type | A0A821TAG2 | R. imitator | < 0.001 | 0.313 |
| Globin/hypothetical protein | A0A822FRT2 | R. imitator | 0.004 | 0.265 |
| Glyceraldehyde-3-phosphate dehydrogenase | A0A0A5SYN4 | C. freundii | 0.002 | 0.174 |
| A0A821LTU2 | R. imitator | 0.011 | 0.231 | |
| Lpp/Major outer membrane lipoprotein Lpp | A0A0L0AQN9 | Klebsiella sp. | 0.048 | 0.243 |
| Thioredoxin /Thioredoxin | W8TGE9 | E. coli | < 0.001 | 0.214 |
| No family described/Enolase | K4WKC0 | E. coli | 0.005 | 0.247 |
| No family described/TetR family transcriptional regulator | A0A1C4ADY6 | L. apis | < 0.001 | 0.180 |
| No family described/IgG H chain | S6B291 | H. sapiens | < 0.001 | 0.247 |
| No family described/Ig-like domain-containing protein | Q8TCD0 | H. sapiens | 0.007 | 0.353 |
| No family described/hypothetical protein | A0A822HYL0 | R. imitator | 0.005 | 0.571 |
VSE-G, venom sac extract of gynes; VSE-W, venom sac extracts of workers; FC (VSE-G/VSE-W), fold change of gyne/worker; FC < 1, downregulated protein in VSE-G; FC > 1, upregulated protein in VSE-G; Statistical differences by t Test, p < 0.05. The nomenclature of proteins is based on software identification.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



