
Submitted:
16 February 2024
Posted:
16 February 2024
You are already at the latest version
Abstract
Cercospora beticola Sacc. is an ascomycete pathogen that causes Cercospora leaf spot in sugar beets (Beta vulgaris L.) and other related crops. The disease symptoms are characterized by circular lesions with an ash-gray centre and dark brown to purple margins on the leaves. It can lead to significant yield losses if not effectively managed. This study aims to characterize and assess the impact of bacterial biological control agents against C. beticola. Following a dual-culture screening, 18 bacteria exhibiting over 50% inhibition were selected, with six of them demonstrating more than 80% control. Identification of the bacteria was carried out through sequencing of the16S rDNA, revealing potentially 12 species belonging to six genera including Bacillus which was represented by four species. Additionally, the bacteria underwent in-depth characterization of biochemical and molecular properties, as well as plant-growth promotion. PCR detection of genes responsible for producing antifungal metabolites revealed that 83%, 78%, 89%, and 56% of the selected bacteria possessed the bacillomycin, iturin, fengycin, and surfactin encoding genes, respectively. Infrared spectroscopy analysis confirmed the presence of lipopeptide structure in the bacteria supernatant filtrate. Subsequently, the bacteria were assessed for their effect on sugar beet plants in controlled conditions. The bacteria exhibited notable capabilities, promoting growth in both roots and shoots, resulting in significant increases in root length, weight, and shoot length. The field experiment with four bacterial candidates demonstrated good performance against C. beticola compared to difenoconazole fungicide. These bacteria played a significant role in disease control, achieving a maximum efficacy of 77.42%, slightly below the 88.51% efficacy attained by difenoconazole. Additional field trials are necessary to verify the protective and growth-promoting effects of these candidates, whether applied individually, combined in consortia, or integrated with chemical inputs in sugar beet crop production.
Keywords:
Biological control
; Cercospora beticola
; Cercospora leaf spot
; Plant-Growth Promoting Rhizobacteria
; Sugar beet
1. Introduction
Cercospora leaf spot disease of sugar beet is one of the most important foliar disease of sugar beet in humid and temperate areas [1]. It is reported to be caused by the ascomycete fungus Cercospora beticola Sacc. (Family Mycosphaerellaceae) [2]. It has been reported that this pathogen plays a significant role in sugar beet production globally. The pathogen can decrease the ton yield by as much as 50% [3,4], concurrently affecting the amount of extractable sugar by increasing impurities [5,6].
The management of the disease involves several practices, including cultural ones such as weed control, irrigation, and nitrogen fertilization. The breeding of disease-resistant varieties is also a strategy employed to control disease severity; however, they have been reported to be less productive than susceptible varieties [7]. Nonetheless, the latest varieties appear to be performing better than the previous [8,9]. Despite the ongoing debate on the use of chemicals in the control of pathogens, this treatments remains the most effective way to manage the disease, especially for susceptible varieties [10].
The excessive use of fungicides against C. beticola has led to the development of acquired resistance to various chemical molecules, including Benomyl [11], Methyl thiophanate [12], Tetraconazol, Epoxyconazole, Difenoconazole Flutriaofol [13,14], Azoxystrobin, Trifloxystrobin [15], and Picoxystrobin [16]. Furthermore, several isolates are now capable of exhibiting multiple types of resistance [17]. To address this issue, biological control using different types of microorganisms (Fungi, Bacteria, and Virus) emerges as highly promising technology [18].
Biological control stands out as a promising approach to manage fungal disease resistance, gaining increasing attention for this environmentally friendly biotechnology. Bacteria are extensively utilized as Biological Control Agents (BCAs) due to their notable effectiveness [19]. Their efficacy stems from their remarkable ability to produce a diverse range of antibiotic compounds. BCAs contribute to the reduction of fungal disease outbreaks through mechanisms such as competition for space and nutrients, induction of plant systemic resistance or the production of antifungal metabolites [19].
Lipopeptides, cyclic amphiphilic oligopeptides with a low molecular weight, are synthesized by non-ribosomal peptide synthetases. They exhibit robust antimicrobial properties, owing to their capacity to interact with cell membranes, leading to pore formation and membrane solubilization at higher concentrations. These lipopeptides are categorized into three primary families: surfactins / lichenysins, iturins / bacillomycins / mycosubtilins, and fengycins/ plipastatins. Bacteriocins, including subtilosin A, are ribosomally produced by various microorganisms and exhibit antimicrobial activity by forming membrane pores [20,21,22].
Bacterial and fungal agents have indeed demonstrated their potential as BCAs [23,24], addressing a wide spectrum of diseases and, notably, the management of C. beticola [25,26]. Furthermore, rhizospheric bacteria are recognized for their plant growth-promoting effects, stimulating overall plant growth [27,28], including the specific case of sugar beet [29,30].
In Morocco, multiple strains of C. beticola have been documented as resistant to multiple fungicides groups, as reported by El Housni et al. (2020) [14]. This study aims to develop a biological alternative to manage this pathogen in sugar beet production. We isolated, selected, and characterized several bacteria from sugar beet rhizospheric soil. These bacteria were assessed for their antagonistic potential against C. beticola, both in vitro and in vivo conditions. Furthermore, the study focuses on characterizing the isolated bacteria, taking into account their biochemical and molecular attributes, along with evaluating their impacts as plant growth-promoting rhizobacteria (PGPR).
2. Materials and Methods
2.1. Fungal Material
The ascomycete fungus used in this present study is C. beticola isolate B9-1, which exhibited cross-resistance to the three main chemical families (Methyl Benzimidazole Carbamate (DMI), Demethylation inhibitors (DMI), and Quinone Outside Inhibitor (QoI)) [14]. This strain was originally isolated from a symptomatic sugar beet leaf during the 2017 growing season in Morocco. Before the experiments, the fungus isolate was subcultured on Potato Dextrose Agar (PDA) and incubated for seven days in darkness at 25°C.
2.1. Isolation of Bacteria from Sugar Beet Rhizosphere Soil
Bacteria were isolated from the rhizosphere soil (described in this context as the soil encompassing the taproot of sugar beet at a distance of 15 cm) of healthy sugar beet (plants display no visible symptoms of C. beticola attacks, and there were no foliar lesions from other pathogens) four months after sowing in the sugar beet regions of Gharb, Doukkala and Tadla (Figure S1), where sugar beet is planted as an autumn crop sown in October and November and harvested in June and July. Eight samples were taken, with approximately 1 kg soil per sample collected from the vicinity of healthy sugar beet plant within a 15 cm diameter up to 50 cm deep, and thoroughly mixed. The mixed soil is then placed in plastic bags and transported to the laboratory. One gram of each soil sample was mixed with 9 ml of sterile distilled water. After shaking, a series of dilutions from 10-4 to 10-6 were carried out [25]. A volume of 0.1 mL of each diluted suspension was incubated on Petri dish filled with PDA medium and incubated at 28°C for at least 48 hours. The resulting bacterial colonies were subsequently transferred to a new PDA Petri dish until pure bacterial morphotype colonies were obtained (colonies with homogeneous morphology). A single colony was chosen based on differences in colour, shape and size. A solid portion of pure culture of each bacterium was then placed in micro-tubes with sterile distilled water and stored at 4°C [25]. BIOBAC® Bacillus subtilis strain Y1336 (SAOAS, Casablanca, Morocco) was used as reference.
2.2. Screening of Antagonistic Activity among Rhizosphere Bacteria
2.2.1. Dual-Culture Setup for Confrontation Testing
The inhibitory potential of 50 bacterial isolates against C. beticola isolate B9-1 was assessed using the direct confrontation method on PDA medium with 4 streaks made at equidistance from the fungal colony [21]. The inhibition rate is calculated at day 7 according to the formula: TI = [(Ct - Ci) / Ct] x 100, where TI represents the growth inhibition rate of C. beticola (%), Ct is the diameter of C. beticola without rhizospheric bacteria (control), and Ci is the diameter of C. beticola in the presence of rhizospheric bacteria.
2.2.2. Antibiotic Activity through Bacterial Supernatant Analysis
A 100 μL volume of each bacterial suspension (1 × 108 CFU/mL) was inoculated into nutrient broth medium (NB), and then incubated in a rotary shaker at 25°C for 6 days (130 rpm) in the dark. The mixture underwent centrifugation at 5500 rpm for 25 minutes, and the resulting supernatant was collected and filtered through a 0.22 µm pore size syringe filter. For the ‘Poison food technique’, the filtrate was added to a PDA medium at 40°C with a 10% filtrate concentration (50 mL filtrate/500 mL liquid PDA medium). Petri dishes containing the medium amended with the filtrate of rhizospheric bacteria were used to inoculate 7 mm diameter portions of C. beticola (obtained from pure culture on PDA medium) taken with a crock borer. The incubation period lasted 7 days at 25°C in complete darkness. Three replicates were carried out for each bacterial filtrate, and three replicates for the control (10% concentration of liquid NB medium to the PDA instead of the bacterial supernatant). The petri dishes were randomly placed at the incubator.
The rate of indirect inhibition of C. beticola growth by rhizosphere bacterial filtrates is calculated according to the following formula [31]: GI = [(Cd - Ci) / Cd] x 100
Where:
GI: growth inhibition rate of C. beticola (%)
Cd: diameter of C. beticola without rhizosphere bacteria (control)
Ci: diameter of C. beticola in the presence of rhizosphere bacteria.
Bacillus subtilis was used as a reference strain.
2.3. Fourier Transform Infrared Spectroscopy (FTIR) Analysis.
The analysis was conducted using an FTIR Spectrophotometer (Shimadzu Model 8400S, Shimadzu, Noisiel, Marne-la-Vallée, France). A homogeneous mixture was prepared by dissolving one millilitre of the biosurfactant sample was in potassium bromide pellets to create the samples (Merck, USA). An integrated plotter was used to generate IR absorption spectra. The IR spectra were acquired with a resolution of 4 cm-1, covering the wavelength range of 450-4500 cm-1. The spectral measurements represented the average of 50 scans across the instrument’s complete operating range.
2.4. DNA Extraction, PCR Amplification of 16S rDNA and Sanger Sequencing
DNA extraction from bacterial suspensions followed the protocol of Llop et al. (2000) [32]. Amplification of 16S rDNA from the genomic DNA of bacterial isolates was carried out using the following pair of primers: 27F /1492R that amplify 1450bp fragment [22]. The PCR reaction mixture was performed in a total volume of 25 μL containing PCR buffer, 2.5 μM of each primer, and 1 U of Taq DNA polymerase (Bioline, London, UK) and 2.5 μL of DNA template. The following cycling conditions were used: Initial denaturation at 96°C for 4 min, followed by 35 cycles of denaturation at 96°C for 10 s, then annealing at 52°C for 40 s and elongation at 72°C for 2 min, and finally an extension at 72 °C for 4 min in Eppendorf Thermal Cycler (Eppendorf, Hambourg, Germany).
PCR reactions that showed amplification bands of excepted fragment approximately 1.4 Kb, were sequenced using sanger dioxide method in Stab Vida laboratory in Portugal. The obtained sequences were analyzed using DNAMAN software (version 6.0, Lynnon Biosoft, Quebec, Canada), and then blasted at the NBCI website (http://blast.ncbi.nlm.nih.gov/Blast.cgi), using BLAST nucleotide search. The 16S rDNA sequences have been deposited in GenBank database, with their corresponding accession numbers.
For the phylogenetic analysis, we employed the maximum likelihood approach using the default settings in MEGA 11 [33]. To ensure statistical robustness, we conducted bootstrap analysis with 1,000 replicates.
2.5. PCR Amplifications Lipopeptide-Encoding Genes
The genes encoding bacillomycin, fengycin, iturin, and surfactin were identified using the genomic DNA isolated from the 18 selected bacterial isolates. For each PCR amplification, a total volume of 25 L of PCR mixture was used, which contained PCR buffer, 2.5 µM of each primer, 1U of Taq DNA polymerase (Invitrogen, California, USA), 2.5 L of genomic DNA. Table S1 shows the specific primers designed to detect the target genes were utilized for amplification. The PCR products were visualized on a 1.5% agarose gel stained with ethidium bromide (0.02 g/mL), observed under an ultraviolet light source, and digitally recorded.
2.6. Biochemical and Plant-Growth Promotion Tests
Different tests with three replicates were performed to detect various biochemical bacterial properties. Table S2 summaries those test with the adopted evaluation method.
2.7. Effects of Bacterial Isolates on Sugar Beet Growth in Greenhouse Conditions
The culture substrate used in this greenhouse trial consisted of 2/3 commercial peat and 1/3 sand. The substrate mixture was autoclaved twice at 121°C for 30 min [34] in hermetically sealed plastic bags. A weight of 200 g of substrate per pot was used for testing rhizospheric bacteria PGPR effect. Commercial sugar beet seeds of the Liberata variety, K.W.S. company were soaked in sterile distilled water for 3 hours to remove the coating. Subsequently, the seeds manually rubbed with a sterile glove and rinsed twice with sterile distilled water (Figure S2).
Bacteria isolated were grown as pure cultures in YEP medium at 28°C under agitation for 24 hours. Subsequently, the bacterial cultures of each isolate were then centrifuged at 5000 rpm for 20 minutes, and the supernatant was discarded. The resulting pellet was re-suspended in PBS at a concentration of 108 CFU/mL, achieved by ensuring an OD 600 nm of 0.1.
Sugar beet seeds underwent disinfection with 70% ethanol for 1 minute, followed washed by a single wash with sterile distilled water. Subsequently, they were soaked in a 5.25% sodium hypochlorite solution with a few drops of Tween 20 and then rinsed with sterile distilled water five times for 1 minute [35]. The sterilised seeds were immersed in the bacterial solution for one hour at room temperature, while control seeds were soaked in PBS alone.
Sugar beet seeds planted in pots containing sterilized substrate were placed in a controlled greenhouse with a temperature of 25°C and relative humidity of 70%. The pots were placed in a completely randomized design with three replicates. After 30 days, the sugar beet plants were assessed for the development of both vegetative and root organs, considering length and dry Wight.
No chemical or organic fertilizers were added to the sugar beet plants, and irrigation was conducted once every three days with 60 mL of tap water per pot.
2.8. Effects of Four Bacterial Isolates on Sugar Beet Growth in the Field
Four bacterial isolates (BGH 1-6, BGH 2-2, BGH1-3, BGH 2-7) displaying significant in vitro antagonism against C. beticola, were chosen for field trial. A suspension containing vegetative cells of these bacteria was utilized with a concentration of 1 x 108 CFU/ml. The slurry application rate was determined at 400 L/ha (Table S3).
The experiments were carried out in 2023 in the Gharb area of Province Sidi Slimane (34°22’12.9”N 6°09’28.4”W). The experimental design was a randomized complete block design with four replicates (Figure S3). Each field plot consisted of eight-meter rows spaced 50 cm apart and planted with the sugar beet cultivar LIBERATA KWS, known for its medium tolerance to Cercospora leaf spot.
Treatment application occurred at the end of February, ensuring a uniform incidence of 64% through a backpack sprayer delivering a 400 L/ha spray. All test plots received the same recommended herbicide and insecticide treatments. Severity was monitored over four weeks, and the results were employed to calculate the Area under the Disease Progression Curve (AUDPC) [36].
where Yi represents the severity of the ith observation, ti denotes the time in days of the observations, and n is the total number of observations.
2.9. Statistical Analysis
The data were analysed by ANOVA at a significance level of 5%, and in instances where the effect was deemed significant at P ≤ 0.001, a multiple comparison of Duncan means was executed. Principal Component Analysis (ACPM) was employed to explore the correlation between bacteria antagonism, lipopeptide-encoding genes, and hydraulic enzyme production. All analyses were conducted using the XLSTAT Pro software, version 7.5.2.
3. Results
3.1. Isolating and Identifying Bacteria from the Rhizosphere Soil of Sugar Beet
A subset comprising 18 bacterial isolates underwent various screening tests. Identification of these isolates was carried out through 16S rDNA and Sanger sequencing, as detailed in Table 1. Sequencing enabled the identification of three isolates at the species level, with a sequence identity exceeding 99% (BGH1-6 identified as Pantoea agglomerans; G1A as Pseudomonas azotoformans), and the isolate G4A matched Pantoea sp., even though the sequence length was approximately 550 bp. For the remaining 15 isolates, the percentage of sequence identity ranged between 91.12% and 97.84%, falling below the threshold for species identification [50].
The genus Bacillus dominated the isolated community with eight isolates, followed by Pantoea (four isolates), Serratia (three isolates), and Enterobacter, Kosakonia, and Pseudomonas, each represented by one isolate.
3.2. Antagonism against C. beticola in Confrontation Assay in PDA Medium and PDA Amended with Supernatant
We conducted two distinct yet complementary assays to evaluate the antagonistic impact of the 18 bacteria against C. beticola. In the confrontation assay on PDA medium, 50% of the isolates demonstrated a significant (P = 0.05) growth inhibitory effect against C. beticola compared to the control (Figure 1A). Notably, six isolates (BGH2-2, BGH2-5, BGH1-6, BGH 1-3, BGH 2-7, G3d) exhibited a growth inhibitory effect exceeding 80%.
The second assay involved applying the supernatants of the 18 bacteria on PDA medium, which was subsequently used for the cultivation of C. beticola. The inhibitory effect of the selected bacteria was statistically significant (P = 0.05) compared to the control, with four isolates (BGH1-3, G3c, BGH2-7, BGH 4-1) showing more than 25% growth inhibition (Figure 1B).
3.3. PCR Detection of Lipopeptide Genes
PCR analysis revealed the existence of genes (BamC, Itup, FenD, and Sfp) encoding for antifungal products, specifically leptopeptides (bacillomycin, iturin, fengycin, and surfactin). Table 2 summarizes the presence and absence of these genes in the bacterial isolates. The genes associated with lipopeptide production were found in varying proportions: 67%, 78%, 83%, and 89% for Sfp, Itup, BamC, and FenD genes, respectively.
Four bacteria (G1d, G1b, BGH 1-3, BGH 2-2) were identified to have all the genes essential for lipopeptide production, each exhibiting different antagonistic capabilities in direct confrontation or through filtrate against C. beticola. Among the six bacteria with an inhibition rate exceeding 80% in direct confrontation (Figue 1A), all possessed at least two lipopeptide production genes (BamC and Itup). Similarly, the four bacteria with an inhibition rate surpassing 25% by filtrate (Figure 1B) all harbored at least two genes responsible for lipopeptide production (Itup and FenD).
3.4. Biochemical and Plant-Growth Promotion Tests
The bacterial isolates underwent comprehensive characterization tests to unveil their functional traits. Table 3 shows the results of eight biochemical activities, including enzymatic assessments for cellulose, pectinase, protease, amylase, and chitinase, along with phosphate solubilization, indol acetic acid production, and hydrogen cyanide production. Among the findings, 56% of the bacteria exhibited proteolytic activity, 61% demonstrated phosphate solubilization, 33% displayed amylolytic activity, 28% showed cellulosic activity, and an equivalent percentage for pectinase activity. Additionally, 17% demonstrated chitinase activity, while 78% displayed hydrogen cyanide production (Table 3).
The filtrate from the BGH 1-3 isolate’s supernatant, belonging to Serratia genus, showed the best performance for inhibition rate in both antagonistic assays, underwent analysis using infrared (IR) spectroscopy. The observed peaks closely resembled the IR spectrum of bacterial lipopeptides. The presence of ester carbonyl groups (-C=O bond in –COOH) was indicated by the peak around 1049 cm-1 [37]. Additionally, the peak at approximately 1,058 cm-1 suggested the existence of amide moieties in proteins [38], and the peaks between 1540 and 1645 cm-1 were characteristic of the –C=O amide I vibration [39].
Contrastingly, the absorption in the 1,500–1,650 cm-1 range, typically absent in the FTIR spectra of glycolipid biosurfactants [40], was not observed. Furthermore, the peak between 1370-1470 (1463) indicated the presence of deformation and bending vibrations of –C-CH2 and -C-CH3 groups in aliphatic chains. The two peaks in the 2,850–2,950 cm-1 region supported the –CH stretching mode of CH3 and CH groups in alkyl chains [39].
These findings suggest the presence of peptide groups in the bacteria supernatant filtrate, affirming the lipopeptide structure of the produced biosurfactant (Figure S4).
3.5. Effects of Bacterial Isolates on Sugar Beet Growth in Controlled Conditions
The inoculation of the 18 bacterial isolates had a highly significant impact on sugar beet root length (F = 23.653; P ≤ 0.0001) and root dry weight (F=35.258; P < 0.0001). According to the Duncan test, three bacteria (BGH2-3, BGH 1-5, BGH2-1) formed a distinct group with the best performance, while two others (G3c and G1a) showed a somewhat adverse effect on root growth (Figure 2A-C and Table S4). Regarding root dry weight, the Duncan test revealed various groups (Figure 2C). Seven bacterial isolates (G1b, BGH1-6, G2c, G3f, BGH2-3, BGH1-5, BGH2-1) exhibited an enhanced effect exceeding 100%. Only G3d, BGH2-5, G3c, and G1a did not have a significant effect on sugar beet root dry weight gain compared to the control (Figure 2 and Table S4).
Bacterial inoculation also influenced root hair development (Table S4 and Figure 2D). Four bacterial isolates (BGH1-6, G1b, G2c, G3f) had a highly positive effect on sugar beet root hair development, while the isolate G3c had a negative impact (Table S4).
ANOVA analysis indicated a highly significant effect (F = 23.65; P < 0.0001) of rhizosphere bacteria on increasing the length of the shoots of sugar beet. The DUNCAN test identified several groups (Figure 2E). Four bacterial isolates (BGH4-1, BGH1-5, BGH2-1, BGH2-7) induced an increase in the length of the shoots by more than 50%, ranked from highest to lowest. Only G4a had a statistically insignificant effect compared to the untreated control (TNT).
Similarly, ANOVA analysis showed a highly significant effect (F = 12.40; P < 0.0001) of rhizosphere bacteria on the weight of the sugar beet above-ground part. The DUNCAN test grouped the effect of bacteria into several categories (Figure 2F). Six bacteria (BGH4-1, BGH1-5, BGH2-1, BGH2-7, G1B, BGH1-6) induced an increase of more than 50% in the weight of the vegetative part, ranked from highest to lowest.
A comprehensive principal components analysis was conducted to examine the correlation between PGPR antagonism, including both direct confrontation assays and bacterial supernatant, and the bacterial ability to produce pectinase and cellulase. It is noteworthy that bacteria displaying higher rates of direct or indirect antagonism are more prominently linked to the production of hydrolytic enzymes rather than the presence of genes associated with antifungal metabolite production (Figure S5).
3.6. Effects of Four Bacterial Isolates on Sugar Beet Growth in the Field
Four bacterial isolates were chosen for the field trial (BGH 2-7, BGH 1-3, BGH 2-2, and BGH 1-6), and the ANOVA revealed a highly significant impact of the treatment on the severity of C. beticola (F = 74.63; P < 0.0001). The Duncan test categorized the treatments into four distinct groups. The most effective treatment was Difenoconazole, recording an efficacy of 88.5%, followed by three bacteria (BGH2-7, BGH 1-3, and BGH1-6) with respective efficacies of 77.4%, 73.8%, and 76.1%. BGH2-2 exhibited the lowest efficacy, with only 61.4% (Table 4).
4. Discussion
The objective of this study was to explore the practical applications of bacterial bioinoculants with the capability to control Cercospora leaf spot disease in sugar beets while concurrently enhancing plant growth and vigor. We isolated bacteria from the rhizosphere of healthy sugar beets cultivated in different regions of Morocco. A subset of 18 bacterial strains underwent various tests, including identification through 16S rDNA sequencing, revealing the prevalence of the genus Bacillus, represented by eight isolates, followed by Pantoea (four isolates), Serratia (three isolates), and Enterobacter, Kosakonia, and Pseudomonas, each represented by one isolate. This is in line with previous studies that showed the dominance of members of the genus Bacillus in conventional agricultural soils [41,42].
However, the sequencing of 16S rDNA allowed the identification of three isolates at the species level, with a sequence identity exceeding 99% (BGH1-6 identified as Pantoea agglomerans; G1A as Pseudomonas azotoformans), and the isolate G4A matched Pantoea sp., although the sequence length was approximately 550 bp for these isolates. For the remaining 15 isolates, the percentage of sequence identity ranged between 91.12% and 97.84%, falling below the threshold for species identification proposed as 98.7% [43]. This percentage falls into the range of genus identification, which is 94.9 [43]. Out of the 18 bacterial isolates, 16 could be potential new species, including G4A (Pantoea sp.). Further investigations such as whole-genome sequencing, phylogenetic analysis, and DNA-DNA hybridization are required to determine the taxonomic position of these isolates or to describe new species.
Moreover, we conducted two antagonist tests, comprising a direct confrontation test on PDA medium and an indirect test involving the supplementation of the bacterial culture supernatant on PDA medium where C. beticola was cultivated, thereby revealing the potential of the isolated bacteria to inhibit the growth of C. beticola. A majority of the bacterial isolates exhibited a significant antagonistic effect, particularly in dual confrontation cultures against C. beticola. Isolates with an antagonism rate exceeding 80% included BGH2-2 (Serratia sp.), BGH 1-3 (Serratia sp.), BGH2-5 (Pantoea sp.), BGH1-6 (Pantoea sp.), BGH 2-7 (Bacillus sp.), G3d (Bacillus sp.). These bacteria are recognized for their capability to inhibit various pathogens [44,45,46,47,48]. Notably, while only Bacillua spp. [26,48,49,50] have been previously tested against C. beticola, our study is the first to investigate the efficacy of bacterial taxa such as Pantoea, Kosakonia, Bacillus, Serratia and Pseudomonas azotoformans, as biological control agents against C. beticola, demonstrating in vitro radial inhibition exceeding 50%.
Unlike other studies, our in vitro antagonism tests, utilizing the addition of bacterial culture supernatant, yielded inhibition rates relatively lower, not surpassing 29%. Interestingly, species affiliated with the Bacillus genus demonstrated inhibition rates exceeding 50% [25], despite PCR analysis confirming the presence of several genes associated with lipopeptide production in the tested PGPR bacteria.
Indeed, upon comparing the growth inhibition results from both direct and indirect confrontations with the presence of the lipopeptides: bacillomycin, fengycin, iturin, and surfactin encoding genes (Table S1), we found that the presence of these genes does not necessarily correlate with optimal performance in inhibiting the growth of C. beticola. This is exemplified by the bacterial isolate G1d (Bacillus sp.), which possesses all four lipopeptide-encoding genes yet demonstrates a direct antagonism of 51% and 0% in indirect antagonism test. Conversely, the top six bacterial isolates in direct confrontation all have at least the bacillomycin encoding gene, while the four performing bacteria in indirect confrontation possess at least two genes encoding for fengycin and iturin. Specific conditions may be necessary for each bacterial taxon to express these genes. The biosynthesis of surfactin and fengycin is influenced by the culture medium, with factors such as nutrient availability, sugar type [51], and the presence of oxidative [52] or diamide [53] agents playing a role in modulating surfactin production. This implies that the potential for antagonism through lipopeptide extraction via supernatant harvesting, based on only one combination of culture media and temperature in in vitro tests, might be underestimated.
Eleven bacterial isolates, displaying diverse levels of antagonism (both direct and indirect), exhibited a positive association with three lipopeptides: bacillomycin, fengycin, and iturin encoding genes, along with the capacity to produce chitinase. Some bacteria possess the ability to produce lipopeptides, recognized for their significant impact on inhibiting fungi, and chitinase, which enables antagonistic bacteria to degrade the cell walls of fungi [21].
The greenhouse experiment demonstrated that the tested isolates delivered effective growth inhibition control, with efficacy ranging from 61% to 77%. These findings align with those reported by Arzanlou et al. (2016) [25] in the context of post-infectious inoculation. Nevertheless, the chemical treatment using difenoconazole proved to be more effective, achieving an efficacy of 88.5%.
The field experiment involving the four bacterial isolates (BGH 2-7, BGH 1-3, BGH 2-2, and BGH 1-6) validated the efficacy of these candidates in reducing C. beticola damage on sugar beet plants. As a result, they could serve as potential candidates for biological control agents (BCA).
A sustainable approach involving biological solutions appears to be effective against C. beticola, particularly in preventive measures. This has been demonstrated with Pythium oligandrum, a mycoparasite used to control fungal diseases, showing a remarkable preventive efficacy of 87% [54]. Examining the possible correlation between various bacterial species [56] or fungal-bacterial antagonists [57,58] holds promise, particularly when these combinations demonstrate an additive or synergistic effects. Another area of research involves delving into the interaction between antagonistic bacteria and fungicides [59]. Moreover, the identification of factors that induce antagonist-induced systemic resistance provides opportunities to strengthen plant resistance or integrate it with other methods to control specific pathogens [60].
In addition to their role as microbial control agents for managing plant diseases, stimulating plant growth, and enhancing overall performance and yield, plant growth-promoting microbes play an important role in suppressing plant diseases [55]. They achieve this by producing inhibitory chemicals and inducing immune responses in plants against phytopathogens [55]. As biofertilizers and biopesticides, plant growth-promoting microbes are regarded as a viable and economically attractive approach for sustainable agriculture.
Bacteria belonging to Bacillus, Pantoea, Serratia and Kosakonia have been successfully tested for their plant-growth promotion effects on beet crops [30,56]. Meanwhile, others, such as Bacillus subtilis, Bacillus amyloliquefaciens, Bacillus siamensis, Kosakonia cowanii, and Serratia nematodiphila, have shown positive effects on the growth of various plants [58,59,60,61,62,63].
In this study, seven bacteria (Serratia nematodiphila BGH 2-3, Pantoea agglomerans BGH 1-5, Pantoea conspicua BGH 2-1, Bacillus subtilis G3f, Bacillus halotolerans, Bacillus amyloliquefaciens G2c) demonstrated the ability to increase root length and weight by over 100%. This increase could be attributed to the inhibition of root ethylene through the ACC deaminase effect of PGPRs [64].
On the plant shoots, only four bacterial isolates (BGH 4-1, BGH 1-5, BGH 2-1, and BGH 2-7) were able to increase the length by more than 50%. In terms of total weight, 17 out of 18 bacteria had a positive effect on increasing the total plant weight, with only nine bacteria showing the ability to increase weight by more than 50%.
Species belongings to Bacillus and Pantoea have been successfully tested for their plant-growth promotion effect on beet crops [30,56,57,58,59,60,61,62]. In this investigation, seven bacterial isolates Bacillus, Pantoea, and Serratia were able to increase root length and dry weight by more than 100%. This increase may be due to inhibition of root ethylene by the ACC deaminase effect [63].
Whereas, on the aerial plant part, only four bacteria (Bacillus and Pantoea) were able to increase the length of the shoots by more than 50%. In terms of total weight, 17 out of 18 bacterial isolates had a positive effect on increasing total plant weight, while only nine bacterial isolates were able to increase weight by more than 50%.
When comparing the impact of bacterial inoculation on the development of plant roots and shoots, we observed that the greatest increase in root length does not necessarily correlate with the increase in root dry weight (e.g., BGH 2-3 versus BGH 4-1) or an increase in root hair (G1b versus G3c). However, for the shoots, a positive proportional relationship exists between the increase in length and the increase in dry weight. One possibility is that the significance of root hair development may explain the variation in dry weight. Surprisingly, beetroot plants with a root dry weight increase of over 50% had less developed root hair than the control (e.g., BGH 4-1; BGH 1-5).
Several PGPRs can exhibit an inhibitory effect on the primary root in favor of lateral roots and root hairs [64]. PGPR-mediated alterations in root traits pave the way toward sustainable crop production, while some PGPRs positively influence root hair development, possibly through auxin-independent mechanisms [65].
Another avenue where PGPRs can contribute is in the induction of systemic resistance, aiding in the control of plant pests in general [66].
5. Conclusion
The bacterial isolates examined and evaluated in this study exhibit multiple biochemical characteristics that could lead to a plant growth-promoting effect on B. vulgaris plants. There is a high likelihood of their utility as a biological control agent against C. beticola, as confirmed by both in vitro and in vivo trials. These antagonist bacteria, derived from the soil of healthy sugar beet plants, align with the view that the selection of microorganisms as biological control agent should involve screening from local niches and application in the same environment to achieve desired benefits.
The positive outcomes of this study mark a crucial step, offering new possibilities for the development of biocontrol strategies to manage Cercospora Leaf Spot resistance against various fungicide groups. Consequently, four distinct antagonist bacterial isolates (Serratia sp. (BGH2-2), Pantea sp. (BGH1-6), Serratia sp. (BGH 1-3), and Serratia sp. (BGH 2-7), either individually or in a consortia, are proposed for controlling and preventing damages caused by C. beticola. However, there is still a need to identify synergistic combinations among different bacterial isolates and determine the best compatibilities between bacterial isolates and sugar beet varieties to optimize results in promoting cultivation and resistance against diverse adversaries. Additionally, exploring the combined use of fungicides and antagonistic bacteria in the field, particularly in addressing resistance issues, presents another avenue for future investigation.
Supplementary Materials
The following supporting information can be downloaded at Preprints.org. Table S1: Oligonucleotide sequences (in the 5′ to 3′ direction) were employed for the amplification of genes encoding lipopeptides from the DNA of bacterial isolates. Table S2: Diverse biochemical analyses were performed, encompassing both the aspect of revelation and the evaluation of activity indices. Table S3: Application of treatments with varying concentrations against C. beticola in the field experiment. Table S4: The impact of bacterial inoculation on the growth of sugar beet plants in a greenhouse experiment. Figure S1: A map showing the 6 sites that have been sampled in Morocco in three regions: G, Gharb; D, Doukkala; and T, Tadla. Figure S2: Panel a displays seeds of sugar beet with a coating, whereas Panel b illustrates seeds that have been washed to eliminate the coated reagents. Figure S3: The field trial site’s location and the experimental setup. Figure S4: Fourier Transform Infrared (FTIR) Spectroscopy used to perform qualitative and quantitative analysis the bacterial isolate BGH2-2 supernatant. Figure S4: Principal components analysis was conducted for the simultaneous assessment of hydrolytic enzyme production, bacterial antagonism, and the presence of lipopeptide encoding genes.
Author Contributions
Z.E. performed experiments, analyzed data, and wrote the manuscript draft. S.E. and N.R. analyzed data and wrote the manuscript draft. A.T. contributed to conceptualization, experimental design, co-supervision, and manuscript editing. K.E. contributed to supervision and data analysis. M.H. contributed to concepts, design, supervision, and manuscript writing. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by funds from the Moulay Ismail University, École Nationale d’Agriculture de Meknès, and University Mohammed VI Polytechnic (UM6P), which is gratefully acknowledged.
Data Availability Statement
The 16S rDNA sequences have been deposited at GenBank under the accessions: MT256072, MT256074 to MT256077, MT254751, MT254758, MT254817, MT254818, MW002221, MW002558, MW008604, MW008870, MW079843, MW079530, MW092005, MW092092, MW086541.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tan, W.; Li, K.; Liu, D.; Xing, W. Cercospora leaf spot disease of sugar beet. Plant Signal Behav 2023, 18, 2214765. [Google Scholar] [CrossRef]
- Rangel, L.I.; Spanner, R.E.; Ebert, M.K.; Pethybridge, S.J.; Stukenbrock, E.H.; de Jonge, R.; Secor, G.A.; Bolton, M.D. Cercospora beticola: The intoxicating lifestyle of the leaf spot pathogen of sugar beet. Molecular Plant Pathology 2020, 21, 1020–1041. [Google Scholar] [CrossRef] [PubMed]
- Skaracis, G.N.; Biancardi, E. Breeding for Cercospora resistance in sugarbeet. In Cercospora beticola Sacc. biology, agronomic influence and control measures in sugar beet., 2nd Edition ed.; Asher, M.J.C., Ed.; Advances in Sugar Beet Research; International Institute for Beet Research: 2000; pp. 177-195.
- Jacobsen, B.J.; Franc, G.D.; Harveson, R.M.; Hanson, L.E.; Hein, G.L. Foliar disease casused by fungi and Oomycetes. In Compendium of beet diseases and pests: The Cercospora leaf spot, 2nd Edition ed.; Harveson, R.M., Hanson, L.E., Hein, G.L., Eds.; American Phytopathological Society: 2009; pp. 7-10.
- Rossi, V.; Meriggi, P.; Biancardi, E.; Rosso, F. Effect of Cercospora leaf spot on sugarbeet growth, yield and quality. Cercospora beticola Sacc. biology, agronomic influence and control measures in sugar beet. 2000, 49-76. [CrossRef]
- Lartey, R.T.; Weiland, J.J.; Bucklin-Comiskey, S. A PCR protocol for rapid detection of Cercospora beticola in sugarbeet tissues. Journal of sugar beet research 2003, 40, 1–10. [Google Scholar] [CrossRef]
- Brown, J.K.M. Yield penalties of disease resistance in crops. Current opinion in plant biology 2002, 5, 339–344. [Google Scholar] [CrossRef]
- Gummert, A.; Ladewig, E.; Bürcky, K.; Märländer, B. Variety resistance to Cercospora leaf spot and fungicide application as tools of integrated pest management in sugar beet cultivation–A German case study. Crop Protection 2015, 72, 182–194. [Google Scholar] [CrossRef]
- Vogel, J.; Kenter, C.; Holst, C.; Marlander, B. New Generation of Resistant Sugar Beet Varieties for Advanced Integrated Management of Cercospora Leaf Spot in Central Europe. Front Plant Sci 2018, 9, 222. [Google Scholar] [CrossRef] [PubMed]
- Panella, L.; Lewellen, R.T. Broadening the genetic base of sugar beet: introgression from wild relatives. Euphytica 2007, 154, 383–400. [Google Scholar] [CrossRef]
- Davidson, R.M.; Hanson, L.E.; Franc, G.D.; Panella, L. Analysis of β-tubulin gene fragments from benzimidazole-sensitive and-tolerant Cercospora beticola. Journal of Phytopathology 2006, 154, 321–328. [Google Scholar] [CrossRef]
- Secor, G.A.; Rivera, V.V.; Khan, M.F.R.; Gudmestad, N.C. Monitoring Fungicide Sensitivity of Cercospora beticola of Sugar Beet for Disease Management Decisions. Plant Disease 2010, 94, 1272–1282. [Google Scholar] [CrossRef]
- Karaoglanidis, G.S.; Thanassoulopoulos, C.C. Cross-resistance patterns among sterol biosynthesis inhibiting fungicides (SBIs) in Cercospora beticola. European journal of plant pathology 2003, 109, 929–934. [Google Scholar] [CrossRef]
- El Housni, Z.; Ezrari, S.; Tahiri, A.; Ouijja, A. Resistance of Cercospora beticola Sacc isolates to thiophanate methyl (benzimidazole), demethylation inhibitors and quinone outside inhibitors in Morocco. EPPO Bulletin 2020, epp.12673-epp.12673. [CrossRef]
- Kirk, W.W.; Hanson, L.E.; Franc, G.D.; Stump, W.L.; Gachango, E.; Clark, G.; Stewart, J. First report of strobilurin resistance in Cercospora beticola in sugar beet (Beta vulgaris) in Michigan and Nebraska, USA. New Disease Reports 2012, 26, 3-3. [Google Scholar] [CrossRef]
- Hudec, K.; Mihók, M.; Roháčik, T.; Mišľan, Ľ. Sensitivity of Cercospora beticola to fungicides in Slovakia. Acta Fytotech. Et Zootech 2020, 23, 147–154. [Google Scholar] [CrossRef]
- Trkulja, N.; Hristov, N. Morphological and genetic diversity of Cercospora beticola isolates. In Proceedings of the International Conference on BioScience: Biotechnology and Biodiversity-Step in the Future, The Fourth Joint UNS-PSU Conference, Book of Proceedings, 2012; pp. 18-20. 2012; 18–20. [Google Scholar]
- Ahmed, M.F.A.; Mikhail, S.P.H.; Shaheen, S.I. Performance efficiency of some biocontrol agents on controlling Cercospora leaf spot disease of sugar beet plants under organic agriculture system. European Journal of Plant Pathology 2023, 167, 145–155. [Google Scholar] [CrossRef]
- Bonaterra, A.; Badosa, E.; Daranas, N.; Frances, J.; Rosello, G.; Montesinos, E. Bacteria as Biological Control Agents of Plant Diseases. Microorganisms 2022, 10. [Google Scholar] [CrossRef]
- Lahlali, R.; Aksissou, W.; Lyousfi, N.; Ezrari, S.; Blenzar, A.; Tahiri, A.; Ennahli, S.; Hrustić, J.; MacLean, D.; Amiri, S. Biocontrol activity and putative mechanism of Bacillus amyloliquefaciens (SF14 and SP10), Alcaligenes faecalis ACBC1, and Pantoea agglomerans ACBP1 against brown rot disease of fruit. Microbial Pathogenesis 2020, 139, 103914-103914. [Google Scholar] [CrossRef] [PubMed]
- Ezrari, S.; Mhidra, O.; Radouane, N.; Tahiri, A.; Polizzi, G.; Lazraq, A.; Lahlali, R. Potential role of rhizobacteria isolated from citrus rhizosphere for biological control of citrus dry root rot. Plants 2021, 10, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Radouane, N.; Adadi, H.; Ezrari, S.; Kenfaoui, J.; Belabess, Z.; Mokrini, F.; Barka, E.A.; Lahlali, R. Exploring the Bioprotective Potential of Halophilic Bacteria against Major Postharvest Fungal Pathogens of Citrus Fruit Penicillium digitatum and Penicillium italicum. Horticulturae 2023, 9, 922-922. [Google Scholar] [CrossRef]
- Thambugala, K.M.; Daranagama, D.A.; Phillips, A.J.L. Fungi vs. Fungi in Biocontrol : An Overview of Fungal Antagonists Applied Against Fungal Plant Pathogens. Frontiers in Cellular and Infection Microbiology 2020, 10, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Dimkić, I.; Janakiev, T.; Petrović, M.; Degrassi, G.; Fira, D. Plant-associated Bacillus and Pseudomonas antimicrobial activities in plant disease suppression via biological control mechanisms-A review. Physiological and Molecular Plant Pathology 2022, 117, 101754-101754. [Google Scholar] [CrossRef]
- Arzanlou, M.; Mousavi, S.; Bakhshi, M.; Khakvar, R.; Bandehagh, A. Inhibitory effects of antagonistic bacteria inhabiting the rhizosphere of the sugarbeet plants, on Cercospora beticola Sacc., the causal agent of Cercospora leaf spot disease on sugarbeet. Journal of Plant Protection Research 2016, 56, 6–14. [Google Scholar] [CrossRef]
- Dervišević, M.; Đorđević, N.; Knežević, I.; Đorđević, S. Antagonistic activity of bacterial isolates against Cercospora beticola in laboratory conditions. 2021, 2021; pp. 47-47.
- Goswami, D.; Thakker, J.N.; Dhandhukia, P.C. Portraying mechanics of plant growth promoting rhizobacteria ( PGPR ): A review. Cogent Food & Agriculture (2016) 2016, 2, 1–19. [Google Scholar] [CrossRef]
- Aeron, A.; Khare, E.; Jha, C.K.; Meena, V.S.; Mohammed, S.; Aziz, A.; Islam, M.T.; Kim, K.; Meena, S.K.; Pattanayak, A.; et al. Revisiting the plant growth - promoting rhizobacteria : lessons from the past and objectives for the future. Archives of Microbiology 2020, 202, 665–676. [Google Scholar] [CrossRef]
- KaragÖZ, H.; Cakmakci, R.; Hosseinpour, A.; Kodaz, S. Alleviation of water stress and promotion of the growth of sugar beet (Beta vulgaris L.) plants by multi-traits rhizobacteria. Applied Ecology & Environmental Research 2018, 16. [Google Scholar]
- Farhaoui, A.; Adadi, A.; Tahiri, A.; El Alami, N.; Khayi, S.; Mentag, R.; Ezrari, S.; Radouane, N.; Mokrini, F.; Belabess, Z.; Lahlali, R. Biocontrol potential of plant growth-promoting rhizobacteria (PGPR) against Sclerotiorum rolfsii diseases on sugar beet (Beta vulgaris L.). Physiological and Molecular Plant Pathology 2022, 119, 101829-101829. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. Journal of pharmaceutical analysis 2016, 6, 71–79. [Google Scholar] [CrossRef]
- Llop, P.; Bonaterra, A.; Peñalver, J.; López, M.a.M. Development of a Highly Sensitive Nested-PCR Procedure Using a Single Closed Tube for Detection of Erwinia amylovora in Asymptomatic Plant Material. Applied and Environmental Microbiology 2000, 66, 2071–2078. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Molecular Biology and Evolution 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Einloft, T.C.; Hartke, S.; de Oliveira, P.B.; Saraiva, P.S.; Dionello, R.G. Selection of rhizobacteria for biocontrol of Fusarium verticillioides on non-rhizospheric soil and maize seedlings roots. European Journal of Plant Pathology 2021, 1–16. [Google Scholar] [CrossRef]
- Bailadores Bollona, J.P.; Delgado Paredes, G.E.; Wagner, M.L.; Rojas Idrogo, C. In vitro tissue culture, preliminar phytochemical analysis, and antibacterial activity of Psittacanthus linearis (Killip) JK Macbride (Loranthaceae). Revista Colombiana de Biotecnología 2019, 21, 22–35. [Google Scholar]
- Wolf, P.F.J.; Verreet, J.A. An integrated pest management system in Germany for the control of fungal leaf diseases in sugar beet: The IPM sugar beet model. Plant disease 2002, 86, 336–344. [Google Scholar] [CrossRef] [PubMed]
- Parthipan, P.; Preetham, E.; Machuca, L.L.; Rahman, P.K.S.M.; Murugan, K.; Rajasekar, A. Biosurfactant and degradative enzymes mediated crude oil degradation by bacterium Bacillus subtilis A1. Frontiers in microbiology 2017, 8, 193-193. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.M.; Mody, K.; Mishra, A.; Jha, B. Physicochemical characterization of biosurfactant and its potential to remove oil from soil and cotton cloth. Carbohydrate polymers 2012, 89, 1110–1116. [Google Scholar] [CrossRef] [PubMed]
- Hajfarajollah, H.; Mokhtarani, B.; Noghabi, K.A. Newly antibacterial and antiadhesive lipopeptide biosurfactant secreted by a probiotic strain, Propionibacterium freudenreichii. Applied biochemistry and biotechnology 2014, 174, 2725–2740. [Google Scholar] [CrossRef] [PubMed]
- Ramani, K.; Jain, S.C.; Mandal, A.B.; Sekaran, G. Microbial induced lipoprotein biosurfactant from slaughterhouse lipid waste and its application to the removal of metal ions from aqueous solution. Colloids and Surfaces B: Biointerfaces 2012, 97, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Khmelevtsova, L.E.; Sazykin, I.S.; Azhogina, T.N.; Sazykina, M.A. Influence of Agricultural Practices on Bacterial Community of Cultivated Soils. Agriculture 2022, 12. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, L.; Chen, Q.; Wen, X.; Liao, Y. Conservation tillage increases soil bacterial diversity in the dryland of northern China. Agronomy for Sustainable Development 2016, 36. [Google Scholar] [CrossRef]
- Yarza, P.; Yilmaz, P.; Pruesse, E.; Glockner, F.O.; Ludwig, W.; Schleifer, K.H.; Whitman, W.B.; Euzeby, J.; Amann, R.; Rossello-Mora, R. Uniting the classification of cultured and uncultured bacteria and archaea using 16S rRNA gene sequences. Nat Rev Microbiol 2014, 12, 635–645. [Google Scholar] [CrossRef]
- Braun-Kiewnick, A.; Jacobsen, B.J.; Sands, D.C. Biological control of Pseudomonas syringae pv. syringae, the causal agent of basal kernel blight of barley, by antagonistic Pantoea agglomerans. Phytopathology 2000, 90, 368–375. [Google Scholar] [CrossRef]
- Schuhegger, R.; Ihring, A.; Gantner, S.; Bahnweg, G.; Knappe, C.; Vogg, G.; Hutzler, P.; Schmid, M.; Van Breusegem, F.; Eberl, L.E.O. Induction of systemic resistance in tomato by N-acyl-L-homoserine lactone-producing rhizosphere bacteria. Plant, Cell & Environment 2006, 29, 909–918. [Google Scholar] [CrossRef]
- Karimi, E.; Safaie, N.; Shams-Baksh, M.; Mahmoudi, B. Bacillus amyloliquefaciens SB14 from rhizosphere alleviates Rhizoctonia damping-off disease on sugar beet. Microbiological research 2016, 192, 221–230. [Google Scholar] [CrossRef]
- Khoa, N.Đ.; Giàu, N.Đ.N.; Tuấn, T.Q. Effects of Serratia nematodiphila CT-78 on rice bacterial leaf blight caused by Xanthomonas oryzae pv. oryzae. Biological Control 2016, 103, 1–10. [Google Scholar] [CrossRef]
- Sarhan, E.A.D. Induction of induced systemic resistance in fodder beet (Beta vulgaris L.) to Cercospora leaf spot caused by (Cercospora beticola Sacc.). Egyptian Journal of Phytopathology 2018, 46, 39–59. [Google Scholar] [CrossRef]
- Sehsah, M.D.; El-Kot, G.A.; El-Nogoumy, B.A.; Alorabi, M.; El-Shehawi, A.M.; Salama, N.H.; El-Tahan, A.M. Efficacy of Bacillus subtilis, Moringa oleifera seeds extract and potassium bicarbonate on Cercospora leaf spot on sugar beet. Saudi J Biol Sci 2022, 29, 2219–2229. [Google Scholar] [CrossRef]
- Pethybridge, S.J.; Vaghefi, N.; Kikkert, J.R. Management of Cercospora leaf spot in conventional and organic table beet production. Plant disease 2017, 101, 1642–1651. [Google Scholar] [CrossRef]
- Yaseen, Y.; Gancel, F.; Béchet, M.; Drider, D.; Jacques, P. Study of the correlation between fengycin promoter expression and its production by Bacillus subtilis under different culture conditions and the impact on surfactin production. Archives of microbiology 2017, 199, 1371–1382. [Google Scholar] [CrossRef] [PubMed]
- Mostertz, J.; Scharf, C.; Hecker, M.; Homuth, G. Transcriptome and proteome analysis of Bacillus subtilis gene expression in response to superoxide and peroxide stress. Microbiology 2004, 150, 497–512. [Google Scholar] [CrossRef]
- Nakano, S.; Küster-Schöck, E.; Grossman, A.D.; Zuber, P. Spx-dependent global transcriptional control is induced by thiol-specific oxidative stress in Bacillus subtilis. Proceedings of the National Academy of Sciences 2003, 100, 13603–13608. [Google Scholar] [CrossRef]
- El Housni, Z.; Tahiri, A.; Ezrari, S.; Radouane, N.; Ouijja, A. Occurrence of Cercospora beticola Sacc populations resistant to benzimidazole, demethylation-inhibiting, and quinone outside inhibitors fungicides in Morocco. European Journal of Plant Pathology 2023, 165, 73–83. [Google Scholar] [CrossRef]
- El-Saadony, M.T.; Saad, A.M.; Soliman, S.M.; Salem, H.M.; Ahmed, A.I.; Mahmood, M.; El-Tahan, A.M.; Ebrahim, A.A.M.; Abd El-Mageed, T.A.; Negm, S.H.; et al. Plant growth-promoting microorganisms as biocontrol agents of plant diseases: Mechanisms, challenges and future perspectives. Front Plant Sci 2022, 13, 923880. [Google Scholar] [CrossRef]
- ÇInar, V.M.; Aydın, Ü. The effects of some biofertilizers on yield, chlorophyll index and sugar content in sugar beet (Beta vulgaris var. saccharifera L.). Ege Üniversitesi Ziraat Fakültesi Dergisi 2021, 58, 163–170. [Google Scholar] [CrossRef]
- Collins, D.P.; Jacobsen, B.J. Optimizing a Bacillus subtilis isolate for biological control of sugar beet Cercospora leaf spot. Biological control 2003, 26, 153–161. [Google Scholar] [CrossRef]
- Gowtham, H.G.; Murali, M.; Singh, S.B.; Lakshmeesha, T.R.; Narasimha Murthy, K.; Amruthesh, K.N.; Niranjana, S.R. Plant growth promoting rhizobacteria Bacillus amyloliquefaciens improves plant growth and induces resistance in chilli against anthracnose disease. Biological Control 2018, 126, 209–217. [Google Scholar] [CrossRef]
- Gao, H.; Lu, C.; Wang, H.; Wang, L.; Yang, Y.; Jiang, T.; Li, S.; Xu, D.; Wu, L. Production exopolysaccharide from Kosakonia cowanii LT-1 through solid-state fermentation and its application as a plant growth promoter. Int J Biol Macromol 2020, 150, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Shoaib, A.; Ali, H.; Javaid, A.; Awan, Z.A. Contending charcoal rot disease of mungbean by employing biocontrol Ochrobactrum ciceri and zinc. Physiology and Molecular Biology of Plants 2020, 26, 1385–1397. [Google Scholar] [CrossRef] [PubMed]
- Samaras, A.; Roumeliotis, E.; Ntasiou, P.; Karaoglanidis, G. Bacillus subtilis MBI600 promotes growth of tomato plants and induces systemic resistance contributing to the control of soilborne pathogens. Plants 2021, 10, 1113-1113. [Google Scholar] [CrossRef] [PubMed]
- Muthu Narayanan, M.; Ahmad, N.; Shivanand, P.; Metali, F. The role of endophytes in combating fungal-and bacterial-induced stress in plants. Molecules 2022, 27, 6549-6549. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ovakim, D.H.; Charles, T.C.; Glick, B.R. An ACC deaminase minus mutant of Enterobacter cloacae UW4No longer promotes root elongation. Current microbiology 2000, 41, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Grover, M.; Bodhankar, S.; Sharma, A.; Sharma, P.; Singh, J.; Nain, L. PGPR mediated alterations in root traits: way toward sustainable crop production. Frontiers in Sustainable Food Systems 2021, 4, 618230-618230. [Google Scholar] [CrossRef]
- Desbrosses, G.; Contesto, C.; Varoquaux, F.; Galland, M.; Touraine, B. PGPR-Arabidopsis interactions is a useful system to study signaling pathways involved in plant developmental control. Plant Signaling & Behavior 2009, 4, 319–321. [Google Scholar] [CrossRef]
- Meena, M.; Swapnil, P.; Divyanshu, K.; Kumar, S.; Harish; Tripathi, Y.N.; Zehra, A.; Marwal, A.; Upadhyay, R.S. PGPR-mediated induction of systemic resistance and physiochemical alterations in plants against the pathogens: Current perspectives. Journal of Basic Microbiology 2020, 60, 828–861. [Google Scholar] [CrossRef]
Figure 1.
A, Percentage of growth inhibition of C. beticola on PDA medium in dual culture, involving 18 bacterial isolates with B. subtilis Y1336 as reference. B, Percentage of growth inhibition of C. beticola on PDA medium implemented by the supernatants of the 18 bacterial isolates. BSL denotes B. subtilis Y1336.
Figure 1.
A, Percentage of growth inhibition of C. beticola on PDA medium in dual culture, involving 18 bacterial isolates with B. subtilis Y1336 as reference. B, Percentage of growth inhibition of C. beticola on PDA medium implemented by the supernatants of the 18 bacterial isolates. BSL denotes B. subtilis Y1336.
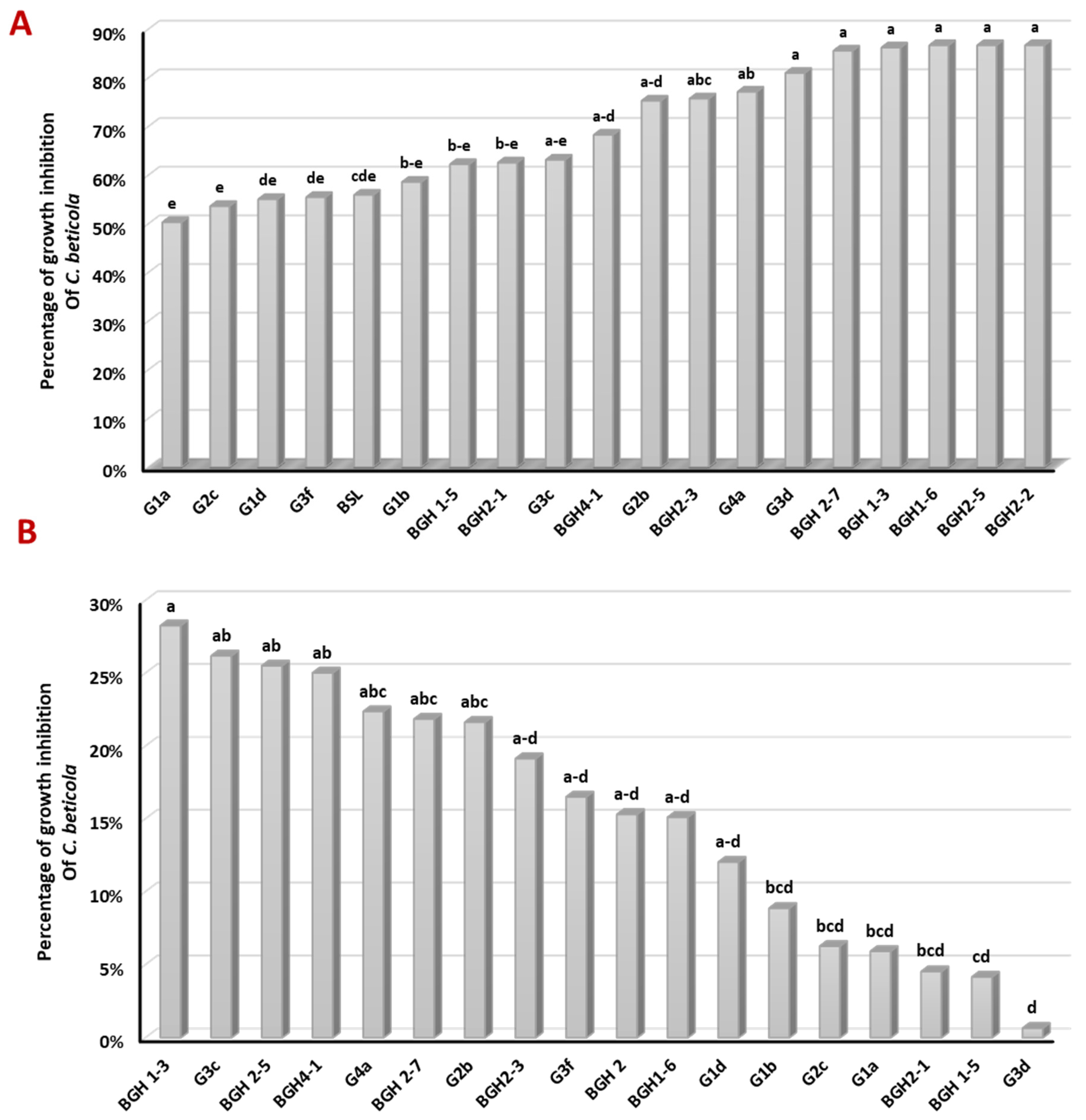
Figure 2.
(A) Impact of bacterial inoculation on the growth of sugar beet, illustrating the roots (depicted in red) and shoots (depicted in yellow) of sugar beet plants (T refers to the untreated control). (B) Influence of bacterial inoculation on the growth of sugar beet root length (TNT refers to the control). (C) Influence of bacterial inoculation on sugar beet root dry weight (TNT refers to the control). (D) Examples showcasing the impact of bacterial inoculation on root hair development (T refers to the control). (E) Impact of bacterial inoculation on sugar beet shoot length growth (TNT refers to the control). (F) Impact of bacterial inoculation on sugar beet shoot dry weight (TNT refers to the control).
Figure 2.
(A) Impact of bacterial inoculation on the growth of sugar beet, illustrating the roots (depicted in red) and shoots (depicted in yellow) of sugar beet plants (T refers to the untreated control). (B) Influence of bacterial inoculation on the growth of sugar beet root length (TNT refers to the control). (C) Influence of bacterial inoculation on sugar beet root dry weight (TNT refers to the control). (D) Examples showcasing the impact of bacterial inoculation on root hair development (T refers to the control). (E) Impact of bacterial inoculation on sugar beet shoot length growth (TNT refers to the control). (F) Impact of bacterial inoculation on sugar beet shoot dry weight (TNT refers to the control).
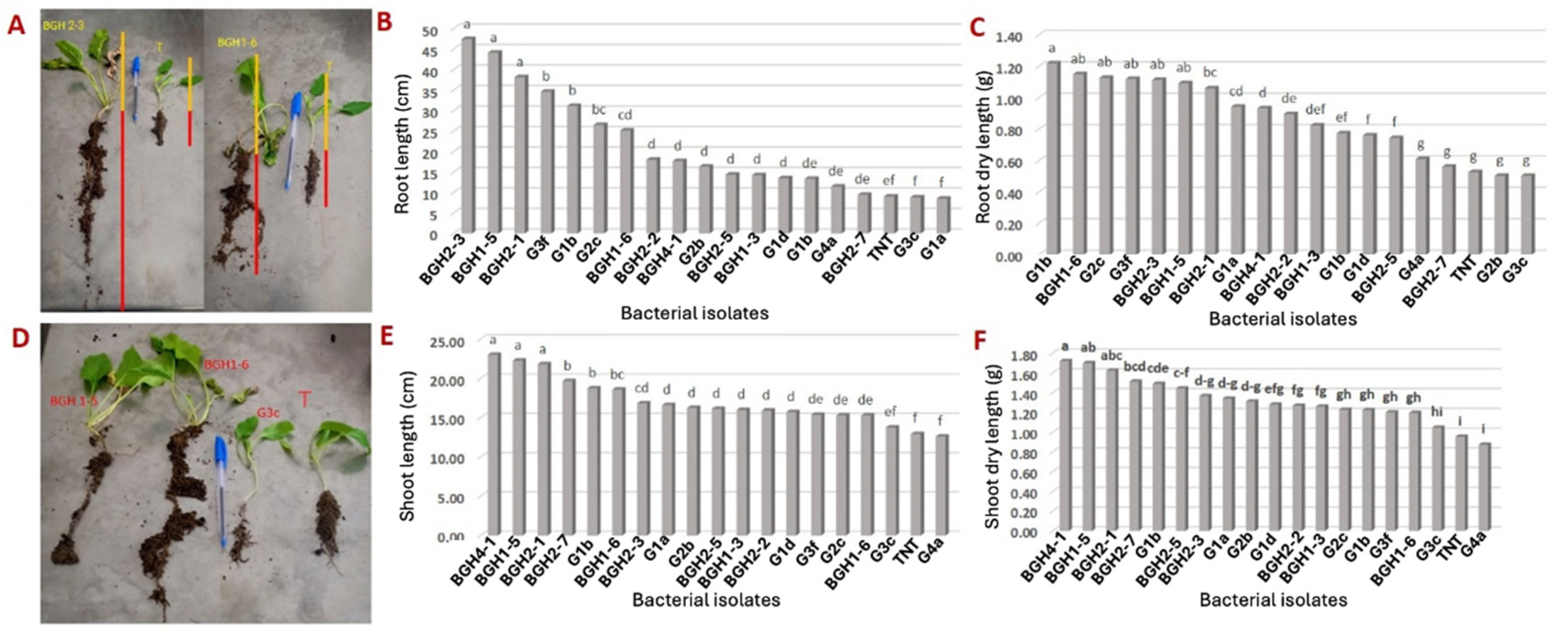

Table 1.
Bacterial sequencing based on 16S rDNA with accession number.
| Strain ID | Accession number | Length (bp) | Coverage (%) | Identity (%) | Closest taxon (Accession Number) |
|---|---|---|---|---|---|
| BGH2-7 | MT256074 | 1501 | 99 | 95.56 | Bacillus vallismortis (FJ386541) |
| BGH4-1 | MW002558 | 559 | 100 | 96.08 | Bacillus halotolerans (MT271912) |
| G1B | MT256075 | 1477 | 99 | 97.35 | Bacillus subtilis (KP876486) |
| G2B | MT256077 | 1495 | 99 | 96.45 | Bacillus halotolerans (MF417800) |
| G3C | MT256076 | 1483 | 99 | 96.46 | Bacillus amyloliquefaciens (PP125657) |
| G1D | MW002221 | 1019 | 99 | 97.84 | Bacillus subtilis (KY818937) |
| G3F | MT254817 | 1524 | 99 | 94.72 | Bacillus subtilis (OM978656) |
| G3D | MT256072 | 1500 | 98 | 95.29 | Bacillus subtilis (KY652939) |
| BGH2-3 | MW086541 | 536 | 99 | 95.86 | Enterobacter sp. (JX103562) |
| BGH1-5 | MW092092 | 1000 | 100 | 93.47 | Kosakonia cowanii MG871199 |
| BGH2-5* | MT254758 | 1513 | 98 | 94.16 | Pantoea agglomerans MZ647535 |
| BGH1-6* | MT254751 | 576 | 100 | 100 | Pantoea agglomerans (OQ202156) |
| BGH2-1 | MT254818 | 903 | 98 | 91.12 | Pantoea conspicua (MW568057) |
| G4A | MW092005 | 555 | 99 | 99.29 | Pantoea sp. (JN853255) |
| G1A | MW079530 | 539 | 100 | 99.63 | Pseudomonas azotoformans (MK883209) |
| BGH1-3 | MW079843 | 559 | 100 | 97.50 | Serratia liquefaciens (MN326772) |
| BGH2-2 | MW008870 | 301 | 99 | 97.00 | Serratia nematodiphila (MH669373) |
| G2C | MW008604 | 645 | 99 | 96.57 | Serratia nematodiphila (MN691578) |
*These isolates are closely related showing more that 99% sequence similarity.
Table 2.
PCR amplifications were performed on antimicrobial lipopeptide encoding genes extracted from the 18 bacterial strains under investigation. The symbol (+) indicates a positive amplification band of the expected size, whereas the symbol (-) denotes the absence of amplification.
Table 2.
PCR amplifications were performed on antimicrobial lipopeptide encoding genes extracted from the 18 bacterial strains under investigation. The symbol (+) indicates a positive amplification band of the expected size, whereas the symbol (-) denotes the absence of amplification.
| Bacterial isolates | Leptopetide encoding genes | |||
|---|---|---|---|---|
| BamC | Itup | fend | Sfp | |
| G1A | + | + | + | - |
| G2C | + | - | + | + |
| G1D* | + | + | + | + |
| G3F | + | + | - | + |
| G1B* | + | + | + | + |
| BGH 1-5 | + | - | + | - |
| BGH2-1 | + | + | + | + |
| G3C | - | + | + | - |
| BGH4-1 | - | + | + | + |
| G2B | - | + | + | + |
| BGH2-3 | + | - | + | - |
| G4A | + | + | + | - |
| G3D | + | - | + | + |
| BGH 2-7 | + | + | + | - |
| BGH 1-3* | + | + | + | + |
| BGH1-6 | + | + | - | - |
| BGH2-2* | + | + | + | + |
| BGH2-5 | + | + | + | - |
*Indicates the bacterial strains that demonstrated positive PCR amplifications for all examined lipopeptide encoding genes.
Table 3.
Biochemical activity assessments of the bacteria. ICL: Cellulose index; IPC: Pectinase index; IPR: Protease index; ISP: Phosphate solubilization; IAM: Amylase index; ICH: Chitinase index; AIA: Indol acetic acid production; HCN: Hydrogen cyanide production.
Table 3.
Biochemical activity assessments of the bacteria. ICL: Cellulose index; IPC: Pectinase index; IPR: Protease index; ISP: Phosphate solubilization; IAM: Amylase index; ICH: Chitinase index; AIA: Indol acetic acid production; HCN: Hydrogen cyanide production.
| Bacteria isolates | ICLa | IPCa | IPRa | ISPa | IAMa | ICHa | AIA | HCN |
|---|---|---|---|---|---|---|---|---|
| G2c | 0 | 1.37±0.08 | 4.12±0.28 | 1.02±0.16 | 1.26±0.06 | 0 | +++ | ++ |
| BGH 1-5 | 0 | 0 | 1.17±0.41 | 1.49±0.16 | 0 | 5.21±0.71 | +++ | - |
| BGH 2-1 | 0 | 0 | 0 | 0 | 0 | 1.94±0.47 | +++ | +++ |
| BGH 2-5 | 0 | 0 | 0 | 1.01±0.43 | 0 | 0 | ++ | + |
| BGH 1-6 | 1.5±0.04 | 1.98±0.79 | 0 | 1.16±0.62 | 0 | 0 | + | + |
| G3f | 0 | 0 | 3.76±0.22 | 1.37±0.08 | 0 | 0 | + | - |
| G2b | 0 | 0 | 0 | 1.40±0.12 | 0 | 0 | + | ++ |
| BGH 2-2 | 0 | 0 | 1.21±0.46 | 1.25±0.19 | 0 | 0 | + | ++ |
| BGH 1-3 | 1.61±0.09 | 1.36±0.34 | 4.12±0.30 | 0 | 1.28±0.04 | 1.53±0.01 | + | +++ |
| G3d | 1.57±0.06 | 0 | 3.09±1.45 | 0 | 0 | 0 | + | + |
| G4a | 1.54±0.01 | 1.58±0.11 | 1.82±0.10 | 0 | 1.14±0.00 | 0 | + | + |
| BGH 2-7 | 0 | 2.03±0.24 | 5.0±0.21 | 1.41±0.26 | 1.06±0.00 | 0 | + | ++ |
| G1b | 0 | 0 | 0 | 0 | 1.19±0.01 | 0 | - | - |
| BGH 2-3 | 0 | 0 | 0 | 1.55±0.01 | 0 | 0 | - | + |
| BGH 4-1 | 0 | 0 | 4.85±0.10 | 1.14±0.20 | 1.24±0.05 | 0 | - | +++ |
| G1d | 0 | 0 | 0 | 1.19±0.47 | 0 | 0 | - | + |
| G1a | 0 | 0 | 0 | 0 | 0 | 0 | - | + |
| G3c | 1.62±0.05 | 0 | 1.42±0.11 | 0 | 0 | 0 | - | - |
(0) and (−): Negatives for the trait; (+ + +), (+ +), and (+): from strong to weak response for the trait, respectively. a Values are the mean of three replicates independent assays ±standard error. Units are in mm.
Table 4.
The impact of four bacterial isolates and difenoconazole on Cercospora leaf spot in sugar beets was assessed by measuring the Area under the Disease Progression Curve (AUDPC) in both treated and control plots, along with determining the percentage of efficacy.
Table 4.
The impact of four bacterial isolates and difenoconazole on Cercospora leaf spot in sugar beets was assessed by measuring the Area under the Disease Progression Curve (AUDPC) in both treated and control plots, along with determining the percentage of efficacy.
| Treatments | Identity (closest BLAST match)a | AUDPCb | Efficiency (%) | signification |
|---|---|---|---|---|
| BGH 2-7 | Bacillus vallismortis | 10.31±0.12 | 77.42% | b |
| BGH 1-3 | Serratia liquefaciens | 11.94±0.14 | 73.86% | b |
| BGH 2-2 | Serratia nematodiphila | 17.61±0.24 | 61.45% | c |
| BGH 1-6 | Pantoea agglomerans | 10.91±0.33 | 76.10% | b |
| Difenoconazole | 5.25±0.43 | 88.51% | a | |
| Control | 45.68±0.96 | d |
a Based is the percentage of sequence identity obtained buy Blast search (Table 1). b AUDPC means the Area under the Disease Progression Curve.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



