
Submitted:
10 February 2024
Posted:
12 February 2024
You are already at the latest version
Abstract
The criteria of “Distinctness, Uniformity and Stability” as well as a high "overall quality index” are used to register the Italian modern varieties to the national register. Differently, local conservation varieties can be certified under different EU Directives having, as overall objective, the preservation of biodiversity and the containment of genetic erosion. In recent years, products derived from an-cient grains are perceived to be healthier and more sustainable by consumer, especially in Italy, with consequent higher market prices. Ancient tetraploid wheat varieties registered in the national reg-ister of conservation varieties are 28, 24 of which are Sicilian. They are supposed to have a wide genetic variability compared to modern ones, making them vulnerable of fraud because they are difficult to trace. It is therefore important to have tools able to discriminate the autochthonous Si-cilian varieties. This can be done by gluten proteins composition, which also gives information on the technological properties of derived products. Fifty-one accessions belonging to 22 ancient va-rieties of Sicilian tetraploid (mostly durum) wheat were analyzed. Although a wide intra-accession and intra-varietal variability was assessed, the gliadin pattern of bulks of seeds belonging to each variety was discriminatory. Moreover, differences in technological attitudes were found between landraces. This paves the way to use gluten protein patterns for traceability allowing local farmers and producers to valorize their products and assure consumers about the transparency of the entire supply chain.
Keywords:
durum wheat
; landraces
; traceability
; gliadin
; glutein subunits
1. Introduction
Wheat is one of the most cultivated crops in the world and provides a major source of nutrition globally. About 95% of the wheat grown worldwide is hexaploid bread wheat (Triticum aestivum L., 2n = 6x = 42, genome AABBDD) whilst the remaining 5% is mainly represented by durum wheat (Triticum turgidum ssp. durum, 2n = 4x = 28, genome AABB) [1]. The latter is well adapted to semiarid environments and is mostly grown in the Mediterranean regions and North America.
After Canada, the second biggest producer of durum wheat in the world is Italy with 3.9 million tons (average data of the period 2019-2023) and more than half of the production comes from southern regions [2,3]. The primary role of Italy in the cultivation of durum wheat is partly attributable to the use of semolina to produce traditional foods of Mediterranean diet. In particular, pasta is a highly popular durum wheat-based food product because of its convenience, versatility, sensorial and nutritional values [4]. According to the Italian law it can only be made with semolina. The economic importance of the pasta industry has fueled the intense breeding work carried out in Italy since the beginning of the 20th century. The main durum wheat breeding programs were focused on the release of élite varieties with higher grain yield and improved protein composition, strictly related to dough and pasta quality traits [5,6,7].
The spread of the modern varieties caused a drastic reduction of the cultivation of landraces and old cultivars which are characterized by lower yield and technological performance, general late maturity coupled with higher stature. For this reason, they were grown for years in marginal areas and their conservation was delegated to public research institutes and custodian farmers. However, in the latter years, there has been an increasing interest in old and landrace wheats, collectively known as “ancient wheats”, especially in Italy, because the consumers perceive their products more healthy, “natural” and sustainable, compared to the modern ones. To date, literature showed conflicting results about the superior healthy properties of old varieties and landraces. Several studies demonstrated that some of these accessions have a higher content of phenols [8,9,10], higher concentration of minerals [11] and prebiotic carbohydrates [12] compared to élite varieties. Other studies supposed that their superior beneficial effects were not due to their characteristic to be “old”, but they are more related to genotype and growth conditions [13,14,15,16,17]. To prove the healthy properties of ancient varieties, further investigations are required, with standard analysis methods on multiple genotypes of old and modern wheat grown in replicate multi-site field trials [18].
Nevertheless, the old cultivars and landraces represent a precious resource of genetic variability for traits associated with tolerance to (a)biotic stress and adaptation to different pedoclimatic conditions and low-input farming systems [19,20,21,22,23]. These favorable alleles or QTLs can be introgressed in élite varieties.
The landraces and historical varieties are populations that do not fall within the regulatory standards that today define modern varieties. In Italy, since 2009, these varieties “which are naturally adapted to the local and regional conditions and threatened by genetic erosion” can be registered in the National Catalogue of agricultural species (Legislative Decree No. 149 of 2009, now repealed and replaced by Legislative Decree No 20 of 2021). This decree has the general objective of safeguarding biodiversity and limiting genetic erosion in agriculture. Nowadays 28 varieties of tetraploid wheat are registered in the National catalogue as conservation varieties and 24 of these are Sicilian.
In Sicily, the agriculture sector is of fundamental importance for the island economy and for the development of rural communities. Tetraploid wheat, including both Triticum turgidum ssp. durum, ssp turgidum and ssp. turanicum, this latter also known as Khorasan wheat, represents one of the most important crops in terms of cultivated area (272,405 ha) [2], and some landraces are still grown in the mountain areas, constituting an important resource to low input farming in marginal areas [24]. Although the old varieties are still produced, conserved, and exchanged among local farmers, the preservation of the grain purity is often difficult during harvesting and post-harvesting procedures. Indeed, the lack of traceability along the supply chain (from seed to end-product) does not guarantee the production and marketing of certified grain, increasing the risk of fraud and putting farmers' economic profits at risk.
In this context, many efforts are made to preserve old varieties (i.e., Sicilian landraces) in purity and their traditional end-products, with the aim of avoiding commercial frauds. Different strategies have been used in the development of traceability methods. Fiore et al. [25] used Cluster Analyses and Principal Component Analysis (PCA) obtained by single nucleotide polymorphisms (SNP) markers, together with agro-morphological, phenological and quality-related traits to classify and evaluate Sicilian ancient wheat germplasm. The same authors demonstrated that the SNP panel represents an efficient tool for the genetic traceability of old wheat varieties and can help to elude commercial frauds sustaining the economic profits for the farmers. Fiore et al. [26] used the same strategy with a wider germplasm collection, including 75 accessions of Sicilian landraces, achieving the same results. Similarly, Taranto et al. [27] dissected the genetic variation patterns of two large germplasm collections of “Timilia” and “Russello” using SNP genotyping. Other molecular markers, such as RFLPs and SSRs, were successfully used for the characterization of durum wheat germplasm collections allowing the variation and resources of landraces more accessible for the exploitation [28,29,30,31,32,33,34]. More analyses were carried out by markers based on morphological descriptors, storage protein composition, digestibility of starch, and concentration of secondary metabolites permitting the discrimination between new élite varieties and landraces [8,35,36,37,38,39,40].
Among biochemical markers, prolamins constitute an efficient system for the traceability of ancient wheat varieties due to their polymorphic nature [41,42]. Prolamins are storage proteins characterized by high proline and glutamine contents and distinguished in polymeric alcohol-insoluble (glutenins) and monomeric alcohol-soluble (gliadins) fractions. Both fractions constitute the gluten, a visco-elastic network responsible for the rheological and technological quality of the dough. Glutenins confer strength and elasticity to the dough [43,44,45] and are formed by polymers stabilised by intra- and inter-chain disulphide bonds. The reduction of these bonds releases the individual glutenin subunits which have been classified in high molecular weight (HMW-GS) and low molecular weight (LMW-GS) glutenin subunits, on the basis of their molecular weight in Sodium Dodecyl Sulphate-PolyAcrylamide Gel Electrophoresis (SDS-PAGE) (reviewed by [46,47]). Gliadins confer extensibility and viscosity to the dough and have been classified in three groups, known as α/β-, γ- and ω-gliadins, based on their decreasing mobility in electrophoresis at acid pH [48,49]. In particular, durum wheat technological quality is mostly due to the presence of the B-type low molecular weight glutenin subunits, with many subunits encoded by the Glu-B3 locus. Two major group of LMW-GS, designed LMW-1 and LMW-2, associated with g-42 and g-45 gliadins, were identified and associated with poor and good technological quality, respectively. In addition, both group of subunits include components controlled not only at the Glu-B3 locus, but also at Glu-A3 and Glu-B2 loci [50].In both group variants differing for the presence or absence of subunits have also been identified [51].Although it is definitively clear that technological quality is due to LMW-2 rather than to g-gliadins, it is still not clear yet if the association between the presence of this allelic form and quality is due to structural or quantitative differences [52].
Visioli et al. [53] used HMW-GS as markers to analyze the genetic purity of grain and flours, marketed and labelled as monovarietal. This study was carried out on four different tetraploid wheat Sicilian landraces (Timilia, Russello, and Margherito, that are durum wheats, and Perciasacchi that is a Khorasan wheat) and highlighted numerous cross contaminations by the other local varieties of durum and bread wheat grown on the same farm. They indicated that HMW-GS analysis was a useful marker to trace the varietal correspondence and the genetic pureness of grains.
In this work, the biochemical profile of prolamins protein was used to trace the varietal correspondence and the gluten quality of 51 accessions derived from 22 Sicilian landraces and old durum/tetraploid wheat varieties obtained by different custodian farmers in two different growing seasons.
2. Results
2.1. Analysis of Gluten Protein
The electrophoretic separations of the gluten components present in the Sicilian varieties were analyzed with the aim to evaluate the presence of heterogeneity in each accession. The A-PAGE and SDS-PAGE analyses of the 40 T. durum, 2 T. turgidum and 9 T. turanicum accession, listed in Table S1, were carried out along with five varieties used as controls. To characterize the presence of intra-accession variability, ten spikes (two kernels for each) and a bulk sample were studied for each accession. Both electrophoretic analyses showed heterogeneity (Table S2) thus indicating the presence of variability. The gliadin patterns showed greater differences than the glutenin ones and showed heterogeneity on 44 accessions of 51 studied (Table S2). Instead, five accessions (Bivona, Ciciredda, Gioia and 2 accessions of Perciasacchi) had a homogeneous profile for both gliadins and glutenins. The analysis of bulk samples confirmed the heterogeneity among accessions of the same variety both for gliadin and glutenin profiles (Figures S1-S5).
To obtain a prolamin profile representative of each accession (here and after called “accession reference pattern”), all the kernels from ten spikes were pulled together. The electrophoretic analyses (Figure S1-S5) detected inter-accession polymorphisms except for Scorsonera, Faricello e Castiglione Glabro that were uniform. Of three accessions of Timilia Reste Bianche, one presented a different profile for gliadins (TRB-4) while Timilia Reste Nere showed inter-accession heterogeneity for both gliadins and LMW glutenin subunits (LMW-GS). Ruscia (synonimous of Russello Ibleo) showed two different profiles for gliadins and LMW-GS. Four accessions of Bidì were analyzed and one of them (BID-1) showed different profiles for gliadins, HMW-GS and LMW-GS. The main differences were probably due to the mixture of two landraces differing at GluB1 (20x+20y and 13x+16y), GliA1 and GliA2 loci. Nine accessions of Perciasacchi (Khorasan wheat) showed 2 different composition profiles for both gliadins and glutenins. In particular, PER-3; PER-6, PER-8 and PER-13 had the tipically composition of HMW-GS (Bx6+By8) while the PER-9 present the subunit Bx20+By20. The other accessions present a mixed profile of the previous ones. Moreover, the morphological analysis of these materials (data unpublished) revealed that two main types of Perciasacchi are grown by custodian farmers in Sicily, differing in terms of flowering time and beck morphology, although the most common seems the type showing Bx6+By8 HMW-GS.
To obtain a representative profile of each landrace, a pool of kernels from all studied accessions (here and after called landrace reference patterns) were analyzed (Figure 1). The electrophoresis analyses permitted to distinguish a unique storage protein composition for each landrace for both gliadins and glutenins. Timila Reste Bianche and Timilia Reste Nere showed the same gliadins pattern, but different C-type LMW-GS. The only exceptions in which gliadins and glutenins failed to distinguish the unique profiles were for the landraces Bidì, Senatore Cappelli and Capeiti 8 that showed a similar profile either for gliadins or glutenins.
In Table 1 the different gluten allelic compositions in studied accessions are listed. Since LMW-GS attribution to a specific allelic form is susceptible to errors due to several protein bands with similar electrophoretic mobilities, we decided to attribute to the LMW-1 or LMW-2 types on the basis of the presence, in this latter, of the slowest moving protein band, named 42K LMW-GS that characterizes this allelic form, whereas its absence identifies the LMW-1 types [52].
2.2. Measurement of the Polymeric Glutenin
UPPs are the large unextractable polymers, which are related to dough properties. The recent breeding programs led to an improvement in the technological properties of wheat varieties, so it is expected to find a higher %UPP in all modern wheats.
SE-HPLC analysis was performed to quantify the amount and size of glutenin polymer in the grain of Sicilian tetraploid landraces Bidì, Castiglione Glabro, Gioia, Perciasacchi, Russello, Ruscia, Timilia Reste Bianche, Timilia Reste Nere and two control durum wheat varieties, an old one, Senatore Cappelli and a modern one, Saragolla.
Differences among wheat landraces/varieties and growing seasons were found for %UPP and Glu/Gli ratio (Table 2). In detail, as expected, the modern cultivar Saragolla had a higher %UPP. During 2020 all genotypes analyzed showed an increase in the content of unextractable polymeric protein compared to the 2019 season except for the old cultivar Senatore Cappelli which showed a decrease of 21% and the conservation varieties Bidì, Castiglione Glabro, Gioia and Timilia Reste Bianche in which the %UPP did not change. In both growing seasons, Ruscia showed the lowest content of %UPPs, whereas Russello the highest among Sicilian landraces. The %UPP value of the latter one was close to modern variety Saragolla. Instead, Perciasacchi had the largest difference in %UPP (>65%) between the growing season 2019 and 2020.
The ratio Glu/Gli was influenced by the growing season differently than the %UPPs (Figure 2). The conservation varieties Bidì and Gioia showed the lowest ratio of Glu/Gli during 2019. On the other way, during the season 2020 Bidì showed the highest value, even more than the modern variety Saragolla. Castiglione Glabro, Timilia Reste Bianche and Timilia Reste Nere showed a worsening in the Glu/Gli ratio in the 2020 season compared to the previous one. Finally, Saragolla confirmed good technological quality during the season 2020 while Senatore Cappelli was the worst.
The percentage of sums of squares accounted for the effect of the growing season was 12% and 2% for %UPP and Glu/Gli respectively. The greatest contribution to the difference in Glu/Gli ratio was given by the interaction of variety x year. Instead, the varietal effect contributed to 52% and 12% for %UPP and Glu/Gli ratio respectively (Figure 2).
3. Discussion
There has been an increasing interest in wheat landraces in the latter years, especially in Italy where tetraploid wheats, particularly durum wheat, have a long tradition of growing and breeding. Sicily is the Region that mostly contributes with 24 out of the 28 landraces recorded in the National Register of Conservation Varieties. This germplasm is agronomically and nutritionally interesting and represents a valuable resource to preserve cereal genetic diversity. Products based on wheat landraces, so called “ancient Sicilian grains”, are present on the shelves of large Italian retailers. However, the correspondence with variety is not guaranteed due to a lack of traceability along the supply chain. It is thus important that such landraces are deeply and clearly characterized and traceable.
Within this context, our work aimed to characterized 51 accessions belonging to 22 Sicilian tetraploid wheat landraces and to assess the intra- and inter-accession variability. Since each cultivar has a specific prolamin composition, electrophoretic patterns of both glutenins and gliadins were used as characterization criterion. Several studies have demonstrated that the electrophoretic profile of prolamins is specific for each cultivar and is not influenced by growing conditions, kernel protein content or incomplete seed maturation [54,55,56].
Analyses of individual spikes allowed to detect heterogeneity in the same accession, mostly revealed by gliadins pattern. Indeed, 85% of studied accessions differed by allelic composition in at least one Gli locus. This was expected as gliadins are one of the most polymorphic proteins in nature, with 147 alleles in tetraploid wheat identified to date [54]. Cases of intra-varietal non-uniformity of common wheat cultivars have been previously described [56,57,58,59]. This variability could be explained as the landraces were obtained by crossing different wheat genotypes that were characterized by different allelic compositions at the Gli loci [60]. and were not selected for this trait. For this reason, the different ecotypes that constitute a landrace may contribute to its adaptivity, and thereby to the known high plasticity of the local varieties.
The accessions reference patterns showed different composition of HMW-GS in Gioia, Russello and Perciasacchi landraces. For Gioia, it seems to be a contamination because the different bands were faint and present only in one accession (Figure S1). Perciasacchi accessions showed the presence of the Bx20+By20 or the Bx6+By8 subunits. The first HMW-GS composition was already described by Visioli et al. [53] in the pure seeds provided by the CREA’s seed bank. In this study, the authors analyzed the commercial flour labelled as monovarietal and found contaminations regarding the HMW-GS subunits. Most of the samples are characterized by a different profile with the presence of subunits Bx6+By8. Our data confirmed that the form showing Bx6+By8 HMW-GS is the most common in Sicilian Perciasacchi landraces grown by custodian farmers and showed how the institutional seed banks often do not reflect the genetic variation of the actually grown materials.
In the same study [53], the analysis of the flours obtained by Russello showed the HMW-GS composition Bx13+By16 and Bx6+By8. The presence of Bx6+By8 was explained by the authors as a contamination with Ruscia. In this paper, the accessions of the same landrace (Russello) showed polymorphism with the presence at the loci Glu-A1 (null or 2*), Glu-B1 (Bx6+By8 or Bx13+By16 or both) gluten subunits. In our study, all accessions of Ruscia highlighted a different composition at Glu-B1 (Bx20+By20) compared to those described by Visioli et al. [53] (Bx6+By8), while the same composition (Bx20+By20) was observed for Bidì (Margherito). In agreement with our results, another work confirmed the presence of the Bx13+By16 subunits for Russello while the same authors described a different HMW-GS (Bx20+By20 vs Bx6+By8 subunits) for Timilia Reste Bianche [35].
The landrace reference patterns showed a different composition in prolamins profiles for all ancient wheats, confirming that the electrophoretic analysis of gliadins and glutenins is a valid method for the traceability of these materials. The only exception was for Bidì, Senatore Cappelli and Capeiti 8 which showed the same prolamin composition. The similarity of gliadins and glutenins profiles is explained by the common origin shared by these varieties. In particular, Bidì and Senatore Cappelli are very similar morphologically and they were reported to be selected respectively by the Tunisian landraces Mahmoudi and Jean Retifah; Capeiti 8 was obtained by the cross Eiti 6 x Senatore Cappelli [5,34,61]. However, De Cillis [62] reported that both Bidì and Senatore Cappelli were selected from the same Tunisian landrace named Bidì and recently some other studies supported this hypothesis [26,27,63]. To trace these landraces is necessary a different method, such as SNP genotyping that was useful to revealing genetic diversity in a large population of two Sicilian landraces, Russello and Timilia [27].
Besides, in this paper, analysis of storage proteins allowed to compare dough quality among landraces in two different growing seasons. Gluten polymers contribute to the technological quality of the wheat by influencing the rheological properties of the dough and this can be predicted by the amount of %UPP and Glu/Gli [50,64]. The amount of insoluble polymeric proteins (%UPP) was higher for all accessions analyzed during the 2020 season due to favorable growing conditions except for Senatore Cappelli which showed a decrease in these proteins. Furthermore, the low %UPP found in this variety was predictable due to the presence of Bx20+By20 HMW-GS correlated to a low technological quality [65]. Instead, during the 2019 growing season Bidi showed a similar % UPP to the modern variety Saragolla, and during the 2020 growing season the higher Glu/Gli ratio, despite having the same HMW composition of Senatore Cappelli. Differences in %UPP between landraces with the same allelic composition was due to differences in storage protein subunit amount and could be explained by difference in gluten strength [35]. Therefore, the 2019 season was characterized by adverse growing conditions and the Russello variety showed to possess a technological quality close to the modern variety Saragolla, although when the environmental conditions are favorable (as in 2020), the Saragolla values are higher, as expected. Instead, Perciasacchi is the genotype that suffered the most from the adverse weather conditions and had the largest difference in %UPP between the two-growing season.
4. Materials and Methods
4.1. Plant Material and Experimental Conditions
The collection of Sicilian grains included in this study are 19 Triticum turgidum subsp. durum (durum wheat), two Triticum turgidum subsp. turgidum and one -Triticum turgidum subsp. turanicum (khorasan wheat) for a total of 51 accessions (Table S1); two historical cultivars (Senatore Cappelli and Trinakria) and three modern durum wheat cultivars (Iride, Core and Saragolla) were used as controls. The cultivation of these materials was carried out in Sicily in two different locations: in 2019 (Palermo, 38°06′54.36″ N; 13°21′02.88″ E. 46 m a.s.l.) and in 2020 (Vicari, Palermo province, 37°49'29"28 N., 13°34'1"20 E., 640 m a.s.l.) under conventional agricultural regime.
Initially, in gliadin and glutenin gels, the extract from two seeds from each of ten spikes, along with a bulk of seeds obtained from each accession were run. Subsequently, to compare accessions with each other, gels were prepared using the bulk samples of all accessions from all varieties. Finally, the bulk samples of all accessions from each variety were mixed to obtain a representative bulk of the varieties, which were then used to produce the final gels.).
4.2. Extraction of Gluten Protein
Two seeds from each sample-spike and five seeds for each sample-bulk of the different wheat landraces and commercial varieties were crushed in a mortal with a pestle until a fine powder was obtained. The procedure for extracting gluten protein following the sequential extraction protocol. The gliadins were extracted from 15 mg of whole flour in 1.5M dimethyl formamide (1:10, w/v) with continuous mixing for 1h at room temperature, followed by centrifugation at 14,000g for 10 min. The pellet was stored at -20 °C and subsequently used for the extraction of the glutenin subunits while the supernatant containing the gliadins fraction was collected. The gliadin loading dye (50% saccharose (w/v) and 0.02% (w/v) dimethyl violet) was added to each sample. The tubes were then vortex mixed, centrifuged for 2 minutes at 14,000g and 10 μL of the supernatant were used to load the gels.
Extraction of glutenin fraction was conducted as described by Ibba et al. [66] with minor modifications. Briefly, the pellet obtained from the first dimethyl formamide treatment was washed two times with 750 μL of 50% 1-propanol and, after centrifugation, the supernatant was discarded to remove any remaining gliadins from the pellet. A volume of 100 μL of a solution with dithiothreitol (DTT) at 1.5%(w/v) formed with 50 μL of 1-propanol at 50% (v/v) and 50 μL of Tris-HCl 0.08M pH 8.0, was then added to the pellet. The tubes were mixed in a vortex and incubated for 30 minutes at 65°C for the extraction of glutenin fraction. After the centrifugation at 10,000g for 2 minutes, 100 μL of a solution with vinylpyridine at 1.4 % (v/v) formed with 50 μL of propanol at 50% (v/v) and 50 μL Tris-HCl 0.08M pH 8.0 was added to the tubes which were then mixed with a vortex, incubated for 15 minutes at 65 ºC, and centrifuged for 2 minutes at 14,000g for alkylation of glutenin subunits. The supernatant containing the glutenin fraction was recovered and transferred to new tubes containing the loading solution 2X (Tris-HCl pH 6.8, 2% SDS, 40% glycerol and 0.02% bromophenol). The samples were mixed in a vortex, incubated for 5 minutes at 90 ºC, and then centrifuged for 2 minutes at 14,000g, 10 μL of the supernatant were loaded on polyacrylamide gels.
4.3. Electrophoretic Separation
A-PAGE was performed using polyacrylamide gels (8 × 13 cm) with 8% of T value (total acrylamide concentration) and 1.25% of C value (cross-linker concentration) containing 40 mM aluminum lactate, 260 mM lactic acid, 0.002 g of ascorbic acid. After electrophoretic separation at 25 mA, the gels were stained with a Commassie R-250 solution fixed in 5% ethanol and 12% trichloroacetic and de-stained in deionized water.
SDS-PAGE was performed using separating gel with 15% T value, and 1.3% of C value. The main gel was prepared using 0.36 M Tris-HCl, pH 8.8 and 0.1% SDS. Gels were run at 12.5 mA per gel for 20 hours and stained with brilliant blue G-colloidal solution fixed in 20% methanol and 10% phosphoric acid and de-stained in deionized water.
The two biotypes of the Italian durum wheat cultivar Lira 42 and Lira 45 were used as standards of g-gliadins g- 42 and g-45, as well as of LMW-1 and LMW-2 types. As HMW-GS standards, the Italian durum wheat cultivars Core (7+8), Iride (7+8), Saragolla (6+8) and Senatore Cappelli (20x+20y) were used (data not shown)
4.4. Analysis Of Unextractable Polymeric Proteins (%UPP) by SE-HPLC
SE-HPLC was used to measure the percentage of non-extractable polymer proteins (%UPP). The extraction of the SDS-soluble fraction and the SDS-insoluble fraction was performed as reported by Gagliardi et al. [67], except for the sonication, that was performed with the probe type sonicator SONICS Vibracell model VC 50T (power 50 W, frequency 20 KHz) for 15s. Briefly, soluble fraction was first extracted from semolina using SDS-phosphate buffer. The remaining pellet was resuspended in the same buffer and sonicated to obtain the UPP portion of semolina protein. The HPLC equipment used was a HPLC Knauer Smartline system equipped with two Smartline 1000 pumps, one HT300L autosampler, one Smartline 2600 UV detector and a TSKgel SuperSW3000 column (300 mm × 4,6 mm). The flow rate was 0.35 mL/min and the detection was at 214 nm. Every sample was analyzed in two replicates. The area under the first peak of the soluble fraction was called P1s while the one under the other peaks was named P2s. The areas of the insoluble fraction were named respectively P1ns and P2ns (Figure S6). The %UPP and the Glu/Gli ratio has been determined as:
4.5. Statistical Analysis
The %UPP and the Glu/Gli values of tetraploid varieties cultivated in the same year were analysed by one-way analysis of variance (ANOVA) and pairwise analysis was carried out using the Tukey Honestly Significant Difference test (Tukey test) (p <0.05). The comparison of these values between the two years was analysed with Student t-test (p<0.05). For the calculation of the Sum of Squares a two-way ANOVA was performed.
All statistical processing was done with R [68].
5. Conclusions
Most of the Italian conservation varieties are Sicilian, where agriculture represents one of the most important economic sectors, fueled also by the business of products obtained from ancient wheats, especially tetraploids, among which durum and Khorasan wheats play the major role. Given that the production is not high, and the demand is increasing, so much that it is possible to find products based on Sicilian tetraploid wheats in the large retail market, it is important to avoid the risk of fraud, for the benefit of both consumers and producers.
In this work, the biochemical profile of prolamins protein permitted to trace the varietal correspondence and the gluten quality of 51 accessions derived from 22 Sicilian landraces and old durum/tetraploid wheat varieties obtained by different custodian farmers in two different growing seasons. This work confirmed that the prolamin composition is a useful tool to trace varietal correspondence of ancient Sicilian landraces and that it is possible to use %UPP to predict technological quality.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1-S5: electrophoresis separation of glutens proteins; Figure S6: examples of SE-HPLC chromatograms used for %UPP calculation; Table S1. List of wheat samples characterized by gluten protein; Table S2. High molecular weight (Glu-HMW), low molecular weight (Glu-LMW), and gliadins (Gli) pattern of Sicilian wheat landraces. The protein profile is defined as heterogenous when at least one sample analysed has a different pattern.
Author Contributions
Conceptualization, S.M.; methodology, M.B., Sa.P. and Si.P.; resources, B.M., G.R. and S.M.; formal analysis, M.B., Sa.P. and Si.P.; data curation, B.F., C.D., C.M., M.B., M.G., Sa.P. and Si.P.; writing—original draft preparation, M.B., Sa.P. and S.M.; writing—review and editing, all authors; Supervisor, S.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research has been funded by the project CA.VA.SI.F.D. (Characterization of Sicilian autochthonous varieties of durum wheat) of the Department of Agriculture of Sicilian Region and MIPAAF on Fund for the protection of biodiversity of agricultural and food interest (L.N. 194/2015, Art. 10).
Data Availability Statement
Not applicable.
Acknowledgments
The authors wish to acknowledge Prof. Domenico Lafiandra for the very useful advises and results discussion; moreover, the custodian farmers are acknowledged for providing the seeds used in this work
Conflicts of Interest
The authors declare no conflict of interest.
References
- Shewry, P. R. Wheat. J. Exp. Bot 2009, 60, 1537–1553. [Google Scholar] [CrossRef] [PubMed]
- ISTAT, Istituto Nazionale di Statistica. Available online: http://dati.istat.it//Index.aspx?QueryId=64761. (accessed on 2 February 2024).
- Xynias, I. N.; Mylonas, I.; Korpetis, E. G.; Ninou, E.; Tsaballa, A.; Avdikos, I. D.; Mavromatis, A. G. Durum wheat breeding in the Mediterranean region: Current status and future prospects. Agronomy 2020, 10, 432. [Google Scholar] [CrossRef]
- Sicignano, A.; Di Monaco, R.; Masi, P.; Cavella, S. From raw material to dish: Pasta quality step by step. J. Sci. Food Agric 2015, 95, 2579–2587. [Google Scholar] [CrossRef] [PubMed]
- De Vita, P.; Nicosia, O. L. D.; Nigro, F.; Platani, C.; Riefolo, C.; Di Fonzo, N.; Cattivelli, L. Breeding progress in morpho-physiological, agronomical and qualitative traits of durum wheat cultivars released in Italy during the 20th century. Eur. J. Agron. 2007, 26, 39–53. [Google Scholar] [CrossRef]
- Malalgoda, M.; Ohm, J. B.; Manthey, F. A.; Elias, E. M.; Simsek, S. Quality characteristics and protein composition of durum wheat cultivars released in the last 50 years. Cereal Chem. 2019, 96, 508–519. [Google Scholar] [CrossRef]
- Royo, C; Dreisigacker, S.; Ammar, K.; Villegas, D. Agronomic performance of durum wheat landraces and modern cultivars and its association with genotypic variation in vernalization response (Vrn-1) and photoperiod sensitivity (Ppd-1) genes. Eur. J. Agron 2020, 120, 126129. [Google Scholar] [CrossRef]
- Lo Bianco, M. L.; Siracusa, L.; Dattilo, S.; Venora, G.; Ruberto, G. Phenolic Fingerprint of Sicilian Modern Cultivars and Durum Wheat Landraces: A Tool to Assess Biodiversity. Cereal Chem 2017, 94, 1045–1051. [Google Scholar] [CrossRef]
- Di Loreto, A.; Bosi, S.; Montero, L.; Bregola, V.; Marotti, I.; Sferrazza, R. E.; Dinelli, G.; Herrero, M.; Cifuentes, A. Determination of phenolic compounds in ancient and modern durum wheat genotypes. Electrophoresis 2018, 39, 2001–2010. [Google Scholar] [CrossRef]
- Menga, V.; Giovanniello, V.; Savino, M.; Gallo, A.; Colecchia, S.A.; De Simone, V.; Zingale, S.; Ficco, D.B.M. Comparative analysis of qualitative and bioactive compounds of whole and refined flours in durum wheat grains with different year of release and yield potential. Plants 2023, 12, 1350. [Google Scholar] [CrossRef]
- Ficco, D. B. M.; Riefolo, C.; Nicastro, G.; De Simone, V.; Di Gesù, A. M.; Beleggia, R.; Platani, C.; Cattivelli, L.; De Vita, P. Phytate and mineral elements concentration in a collection of Italian durum wheat cultivars. Field Crops Res. 2009, 111, 235–242. [Google Scholar] [CrossRef]
- Ficco, D. B M.; Prandi, B.; Amaretti, A.; Anfelli, I.; Leonardi, A.; Raimondi, S.; Pecchioni, N.; De Vita, P.; Faccini, A.; Sforza, S.; Rossi, M. Comparison of gluten peptides and potential prebiotic carbohydrates in old and modern Triticum turgidum ssp. genotypes. Food Res. Int. 2019, 120, 568–576. [CrossRef] [PubMed]
- Shewry, P. R.; Hey, S. Do “ancient” wheat species differ from modern bread wheat in their contents of bioactive components? J. Cereal Sci. 2015, 65, 236–243. [Google Scholar] [CrossRef]
- Ziegler, J.U.; Steiner, D.; Longin, C.F.H.; Würschum, T.; Schweiggert, R.; Carle, R. Wheat and the irritable bowel syndrome – FODMAP levels of modern and ancient species and their retention during bread baking. J. Funct. Foods 2016, 25, 257–266. [Google Scholar] [CrossRef]
- Dinu, M.; Whittaker, A.; Pagliai, G.; Benedettelli, S.; Sofi, F. Ancient wheat species and human health: Biochemical and clinical implications. J. Nutr. Biochem. 2018, 52, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Shewry, P. R. Do ancient types of wheat have health benefits compared with modern bread wheat? J. Cereal Sci. 2018, 79, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Palombieri, S.; Bonarrigo, M.; Cammerata, A.; Quagliata, G.; Astolfi, S.; Lafiandra, D.; Sestili, F.; Masci, S. Characterization of Triticum turgidum sspp. durum, turanicum, and polonicum grown in Central Italy in relation to technological and nutritional aspects. Front. Plant Sci. 2023, 14, 1269212. [Google Scholar] [CrossRef]
- Shewry, P. R.; Hassall, K. L.; Grausgruber, H.; Andersson, A. M.; Lampi, A. M.; Piironen, V.; Rakszegi, M.; Ward, J.L; Lovegrove, A. Do modern types of wheat have lower quality for human health? Nutr. Bull. 2020, 45, 362–373. [Google Scholar]
- Pecetti, L.; Boggini, G.; Gorham, J. Performance of durum wheat landraces in a Mediterranean environment (eastern Sicily). Euphytica 1994, 80, 191–199. [Google Scholar] [CrossRef]
- Lopes, M.S.; El-Basyoni, I.; Baenziger, P.S.; Singh, S.; Royo, C.; Ozbek, K.; Aktas, H.; Ozer, E.; Ozdemir, F.; Manickavelu, A.; Ban, T.; Vikram, P. Exploiting genetic diversity from landraces in wheat breeding for adaptation to climate change. J. Exp. Bot. 2015, 66, 3477–3486. [Google Scholar] [CrossRef]
- Ruisi, P.; Frangipane, B.; Amato, G.; Frenda, AS.; Plaia, A.; Giambalvo, D.; Saia, S. Nitrogen uptake and nitrogen fertilizer recovery in old and modern wheat genotypes grown in the presence or absence of interspecific competition. Front. Plant Sci. 2015, 6, 185. [Google Scholar] [CrossRef]
- Sharma, S.; Schulthess, A.W.; Bassi, F.M.; Badaeva, E.D.; Neumann, K.; Graner, A.; Özkan, H.; Werner, P.; Knüpffer, H.; Kilian, B. Introducing beneficial alleles from plant genetic resources into the wheat germplasm. Biology 2021, 10, 982. [Google Scholar] [CrossRef]
- Nazco, R., Villegas, D., Ammar, K., Peña, R. J., Moragues, M., & Royo, C. (2012). Can Mediterranean durum wheat landraces contribute to improved grain quality attributes in modern cultivars? Euphytica, 185, 1–17. [CrossRef]
- Scavo, A.; Pandino, G.; Restuccia, A.; Caruso, P.; Lombardo, S.; Mauromicale, G. Allelopathy in Durum Wheat Landraces as Affected by Genotype and Plant Part. Plants 2022, 11, 1021. [Google Scholar] [CrossRef]
- Fiore, M.C.; Mercati, F.; Spina, A.; Blangiforti, S.; Venora, G.; Dell’Acqua, M.; Lupini, A.; Preiti, G.; Monti, M.; Pè, M.E.; Sunseri, F. High-Throughput Genotype, Morphology, and Quality Traits Evaluation for the Assessment of Genetic Diversity of Wheat Landraces from Sicily. Plants 2019, 8, 116. [Google Scholar] [CrossRef]
- Fiore, M.C.; Blangiforti, S.; Preiti, G.; Spina, A.; Bosi, S.; Marotti, I.; Mauceri, A.; Puccio, G.; Sunseri, F.; Mercati, F. Elucidating the Genetic Relationships on the Original Old Sicilian Triticum Spp. Collection by SNP Genotyping. Int. J. Mol. Sci. 2022, 23, 13378. [Google Scholar] [CrossRef]
- Taranto, F.; Di Serio, E.; Miazzi, M. M.; Pavan, S.; Saia, S.; De Vita, P.; D’Agostino, N. Intra-and Inter-Population Genetic Diversity of “Russello” and “Timilia” Landraces from Sicily: A Proxy towards the Identification of Favorable Alleles in Durum Wheat." Agronomy 2022, 12, 1326.
- Laidò, G.; Mangini, G.; Taranto, F.; Gadaleta, A.; Blanco, A.; Cattivelli, L.; Marone, D.; Mastrangelo, A.M; Papa, R.; De Vita, P. Genetic diversity and population structure of tetraploid wheats (Triticum turgidum L.) estimated by SSR, DArT and pedigree data. PLoS ONE 2013, 8, e67280. [Google Scholar] [CrossRef]
- Sahri, A.; Chentoufi, L.; Arbaoui, M.; Ardisson, M.; Belqadi, L.; Birouk, A.; Roumet, P.; Muller, M.H. Towards a comprehensive characterization of durum wheat landraces in Moroccan traditional agrosystems: Analysing genetic diversity in the light of geography, farmers’ taxonomy and tetraploid wheat domestication history. BMC Evol. Biol. 2014, 14, 264. [Google Scholar] [CrossRef] [PubMed]
- Moragues, M.; Moralejo, M.; Sorrells, M.E.; Royo, C. Dispersal of durum wheat [Triticum turgidum L. ssp. turgidum convar.durum (Desf.) MacKey] landraces across the Mediterranean basin assessed by AFLPs and microsatellites. Genet. Resour. CropEvol. 2007, 54, 1133–1144. [Google Scholar] [CrossRef]
- Figliuolo, G.; Mazzeo, M.; Greco, I. Temporal variation of diversity in Italian durum wheat germplasm. Genet. Resour. Crop Evol. 2007, 54, 615–626. [Google Scholar] [CrossRef]
- Ruiz, M.; Giraldo, P.; Royo, C.; Villegas, D.; Aranzana, M.J.; Carrillo, J.M. Diversity and Genetic Structure of a Collection of Spanish Durum Wheat Landraces. Crop Sci. 2012, 52, 2262–2275. [Google Scholar] [CrossRef]
- Maccaferri, M.; Sanguineti, M.C.; Donini, P.; Tuberosa, R. Microsatellite analysis reveals a progressive widening of the genetic basis in the elite durum wheat germplasm. Theor. Appl. Genet. 2003, 107, 783–797. [Google Scholar] [CrossRef]
- Sicilia, A.; Anastasi, U.; Bizzini, M.; Montemagno, S.; Nicotra, C.; Blangiforti, S.; Spina, A.; Cosentino, S.L.; Lo Piero, A.R. Genetic and Morpho-Agronomic Characterization of Sicilian Tetraploid Wheat Germplasm. Plants 2022, 11, 130. [Google Scholar] [CrossRef]
- De Santis, M.A.; Giuliani, M.M.; Giuzio, L.; De Vita, P.; Lovegrove, A.; Shewry, P.R.; Flagella, Z. Differences in gluten protein composition between old and modern durum wheat genotypes in relation to 20th century breeding in Italy. Eur. J. Agron. 2017, 87, 19–29. [Google Scholar] [CrossRef]
- Di Francesco, A.; Saletti, R.; Cunsolo, V.; Svensson, B.; Muccilli, V.; De Vita, P.; Foti, S. Dataset of the metabolic and CM-like protein fractions in old and modern wheat Italian genotypes. Data Brief 2019, 27, 104730. [Google Scholar] [CrossRef]
- Di Francesco, A.; Saletti, R.; Cunsolo, V.; Svensson, B.; Muccilli, V.; De Vita, P.; Foti, S. Qualitative proteomic comparison of metabolic and CM-like protein fractions in old and modern wheat Italian genotypes by a shotgun approach. J. Proteom. 2020, 211, 103530. [Google Scholar] [CrossRef]
- Di Francesco, A.; Saletti, R.; Cunsolo, V.; Svensson, B.; Muccilli, V.; De Vita, P.; Foti, S. Quantitative label-free comparison of the metabolic protein fraction in old and modern italian wheat genotypes by a shotgun approach. Molecules 2021, 26, 2596. [Google Scholar] [CrossRef]
- Pandolfo, A.; Messina, B.; Russo, G. Evaluation of Glycemic Index of Six Different Samples of Commercial and Experimental Pasta Differing in Wheat Varieties and Production Processes. Foods 2021, 10, 2221. [Google Scholar] [CrossRef]
- Blangiforti, S.; Gallo, G.; Porfiri, O. Morpho-agronomic and molecular assessment of a collection of Sicilian wheats [Triticum aestivum L.; Triticum durum Desf.]. Italus Hortus 2006, 13, 364–369. [Google Scholar]
- Rombouts, I.; Lagrain, B.; Brunnbauer, M.; Delcour, J. A.; Koehler, P. Improved identification of wheat gluten proteins through alkylation of cysteine residues and peptide-based mass spectrometry. Sci. Rep. 2013, 3, 1–11. [Google Scholar] [CrossRef]
- Shewry, P. R.; Halford, N. G.; Lafiandra, D. Genetics of wheat gluten proteins. Adv. Genet. 2003, 49, 111–184. [Google Scholar]
- Shewry, P. R.; Halford, N. G.; Tatham, A. S.; Popineau, Y.; Lafiandra, D.; Belton, P. S. The high molecular weight subunits of wheat glutenin and their role in determining wheat processing properties. Adv Food Nutr Res. 2003, 45:219-302. [CrossRef] [PubMed]
- Wieser, H. Chemistry of gluten proteins. Food Microbiol. 2007, 24, 115–119. [Google Scholar] [CrossRef]
- Wrigley, C. W.; Békés, F.; Bushuk, W. Gluten: A balance of gliadin and glutenin. Gliadin and glutenin: The unique balance of wheat quality 2006, 3-32.
- Shewry, P. R.; Lafiandra, D. Wheat glutenin polymers 1. structure, assembly and properties. J. Cereal Sci 2022, 103486.
- Lafiandra, D.; Shewry, P. R. Wheat Glutenin polymers 2, the role of wheat glutenin subunits in polymer formation and dough quality. J. Cereal Sci 2022, 103487. [Google Scholar] [CrossRef]
- Woychik, J. H.; Boundy, J. A.; Dimler, R. J. Starch gel electrophoresis of wheat gluten proteins with concentrated urea. Arch Biochem Biophys 1961, 94, 477–482. [Google Scholar] [CrossRef]
- Kasarda, D.D; Adalsteins, A.E; Laird, N.F. Gamma-gliadins with alpha-type structure coded on chromosome 6B of the wheat (Triticum aestivum L.) cultivar "Chinese Spring". In: Lasztity R and Bekes F (eds) Proc 3rd Int Workshop on Gluten Proteins 1993, Budapest, Hungary, pp. 20–29.
- Ruiz, M.; Giraldo, P. The influence of allelic variability of prolamins on gluten quality in durum wheat: An overview. J. Cereal Sci. 101 (2021): 103304.
- Carrillo J.M., Vazquez, J.F. Orellana J. Relationship Between Gluten Strength and Glutenin Proteins in Durum Wheat Cultivars Plant Breeding 104, 325-333 (1990)J.
- D’Ovidio, R.; Masci, S. The low-molecular-weight glutenin subunits of wheat gluten. J. Cereal Sci. 2004, 39. [Google Scholar] [CrossRef]
- Visioli, G.; Giannelli, G.; Agrimonti, C.; Spina, A.; Pasini, G. Traceability of Sicilian durum wheat landraces and historical varieties by high molecular weight glutenins footprint. Agronomy 2021, 11, 143. [Google Scholar] [CrossRef]
- Payne, P.I.; Holt, L.M.; Lawrence, G.J.; Law, C.N. The genetics of gliadin and glutenin, the major storage proteins in the wheat endosperm. Plant Food Hum. Nutr. 1982, 31, 229–241. [Google Scholar]
- Metakovsky, E. V. Wheat storage proteins: Genes, inheritance, variability, mutations, phylogeny, seed production, flour quality. Lambert Acad. Pabl. 2015 (in Russian, English summary).
- Metakovsky, E.; Melnik, V.; Rodriguez-Quijano, M.; Upelniek, V.; Carrillo, J. M. A catalog of gliadin alleles: Polymorphism of 20th-century common wheat germplasm. Crop J. 2018, 6, 628–641. [Google Scholar] [CrossRef]
- Pogna, N.E.; Peruffo, A.D.B.; Boggini, G.; Corbellini, M. Analysis of wheat varieties by gliadin electrophoregrams. II. Nature, origin and quality of biotypes present in six Italian common wheat varieties. Genetica Agraria, 1982 36:143-154.
- Lawrence, G.J.; Moss, H.J.; Shepherd, K.W.; Wrigley, C.W. Dough quality of biotypes of eleven Australian wheat cultivars that differ in high-molecular-weight glutenin subunit composition. J. Cereal Sci. 1987, 6, 99–101. [Google Scholar] [CrossRef]
- Sontag-Strohm, T.; Payne, P.I.; Salovaara, H. Effect of allelic variation of glutenin subunits and gliadins on baking quality in the progeny of two biotypes of bread wheat cv. Ulla. J. Cereal Sci. 1996, 24, 115–124. [Google Scholar] [CrossRef]
- Metakovsky, E. V.; Branlard, G. P.; Graybosch, R. A. Gliadins of common wheat: Polymorphism and genetics. Gliadin and glutenin: The unique balance of wheat quality 2006, 35-84.
- De Cillis, U. I Frumenti Siciliani; Tipografia Zuccarello & Izzi: Catania, Italy, 1942; ISBN 88-7751-229-6. [Google Scholar]
- De Cillis, E. I Grani d’Italia. Roma: Tipografia della Camera dei Deputati, 1927173.
- Marzario, S.; Logozzo, G.; David, J. L.; Zeuli, P. S.; Gioia, T. Molecular Genotyping (SSR) and Agronomic Phenotyping for Utilization of Durum Wheat (Triticum durum Desf.) Ex Situ Collection from Southern Italy: A Combined Approach Including Pedigreed Varieties. Genes 2018, 9, 10. [Google Scholar] [CrossRef]
- Gupta, R.B.; Popineau, Y.; Lefebvre, J.; Cornec, M.; Lawrence, G.J.; MacRitchie, F. Biochemical basis of flour properties in bread wheats. II. Changes in polymeric protein formation and dough/gluten properties associated with the loss of low Mr or high Mr glutenin subunits. J. Cereal Sci. 1995, 21, 103–116. [Google Scholar] [CrossRef]
- Santagati, V.D.; Sestili, F.; Lafiandra, D.; D’Ovidio, R.; Rogniaux, H.; Masci, S. Characterization of durum wheat high molecular weight glutenin subunits Bx20 and By20 sequences by a molecular and proteomic approach. J. Mass Spectrom. 2016, 51, 512–517. [Google Scholar] [CrossRef] [PubMed]
- Ibba, M. I.; Kiszonas, A. M.; Guzmán, C.; Morris, C. F. Definition of the low molecular weight glutenin subunit gene family members in a set of standard bread wheat (Triticum aestivum L.) varieties . J. Cereal Sci. 2017, 74, 263–271. [Google Scholar] [CrossRef]
- Gagliardi, A.; Carucci, F.; Masci, S.; Flagella, Z.; Gatta, G.; Giuliani, M.M. Effects of genotype, growing season and nitrogen level on gluten protein assembly of durum wheat grown under Mediterranean conditions. Agronomy 2020, 10, 755. [Google Scholar] [CrossRef]
- R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing 2022, Vienna, Austria. URL https://www.R-project.org/.
Figure 1.
Electrophoretic separation of glutens proteins of the varietal bulks. (a,c) A-PAGE analysis of gliadins fractions. (b,d) SDS-PAGE analysis of glutenins fraction. FRA, Francesa; SAM, Sammartinara; TRB, Timilia reste bianche; TRN, Timilia reste nere; PER, Perciasacchi; BIA, Biancuccia; GIU, Giustalisa; MAR,Martinella; URR, Urria; RUS, Russello; RUI, Ruscia; BIV, Bivona; CIC, Ciciredda; BID, Bidì; CAPP, Senatore Cappelli; FAR, Faricello; CAP, Capeìti; TRI, Tripolino; TRK, Trinakria; CAS, Castiglione Glabro; VAL, Vallelunga; SCA, Scavuzza; GIO, Gioia; SCO, Scorsonera. The arrows indicate the g-42 associated with the presence of LMW-1 in the varieties Faricello e Tripolino. All the other varieties show allelic forms of LMW-2 type (a,d).
Figure 1.
Electrophoretic separation of glutens proteins of the varietal bulks. (a,c) A-PAGE analysis of gliadins fractions. (b,d) SDS-PAGE analysis of glutenins fraction. FRA, Francesa; SAM, Sammartinara; TRB, Timilia reste bianche; TRN, Timilia reste nere; PER, Perciasacchi; BIA, Biancuccia; GIU, Giustalisa; MAR,Martinella; URR, Urria; RUS, Russello; RUI, Ruscia; BIV, Bivona; CIC, Ciciredda; BID, Bidì; CAPP, Senatore Cappelli; FAR, Faricello; CAP, Capeìti; TRI, Tripolino; TRK, Trinakria; CAS, Castiglione Glabro; VAL, Vallelunga; SCA, Scavuzza; GIO, Gioia; SCO, Scorsonera. The arrows indicate the g-42 associated with the presence of LMW-1 in the varieties Faricello e Tripolino. All the other varieties show allelic forms of LMW-2 type (a,d).
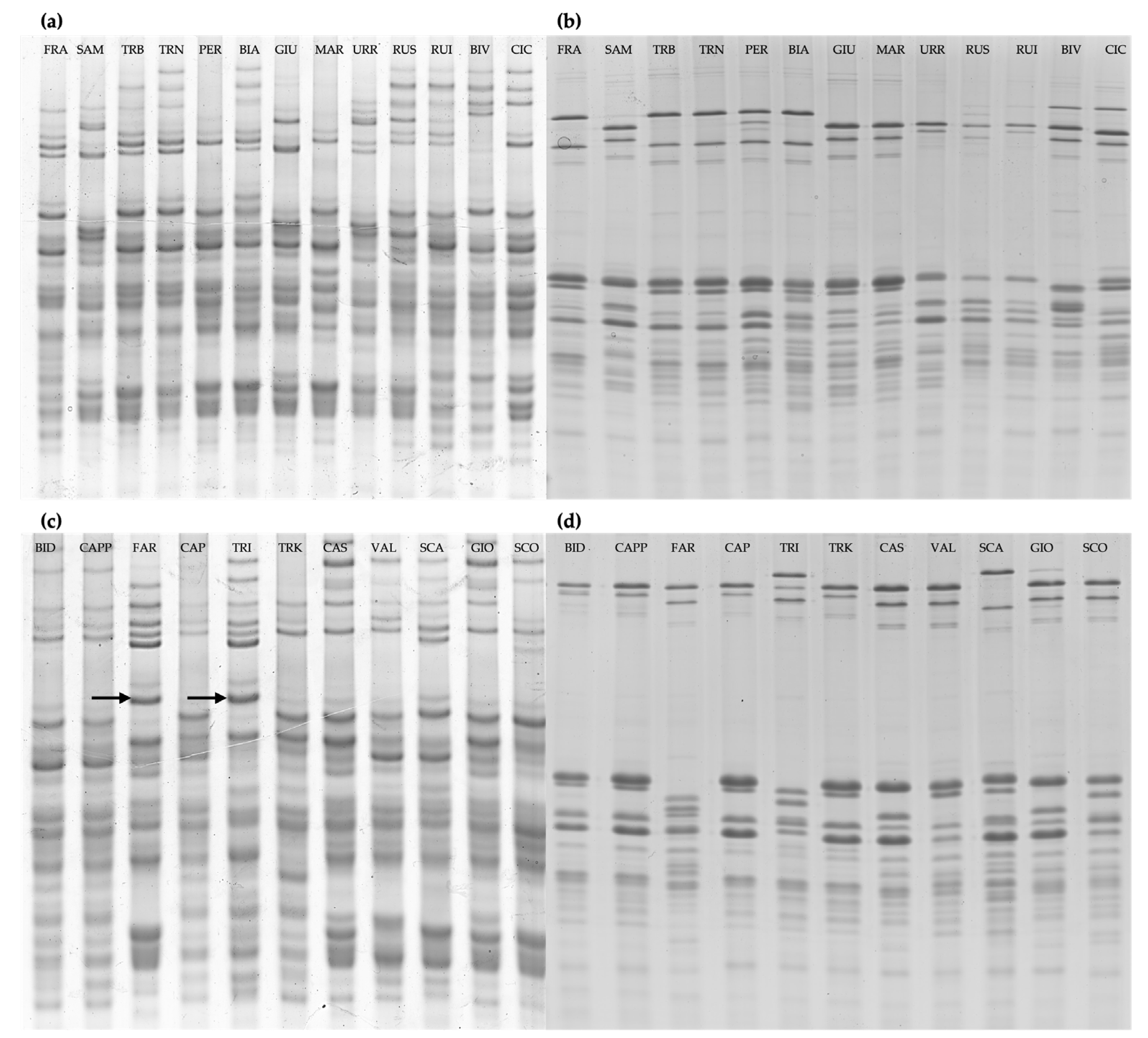
Figure 2.
Percentage of the total sums of squares for the effects of variety, year, interaction between variety and year and residuals.
Figure 2.
Percentage of the total sums of squares for the effects of variety, year, interaction between variety and year and residuals.
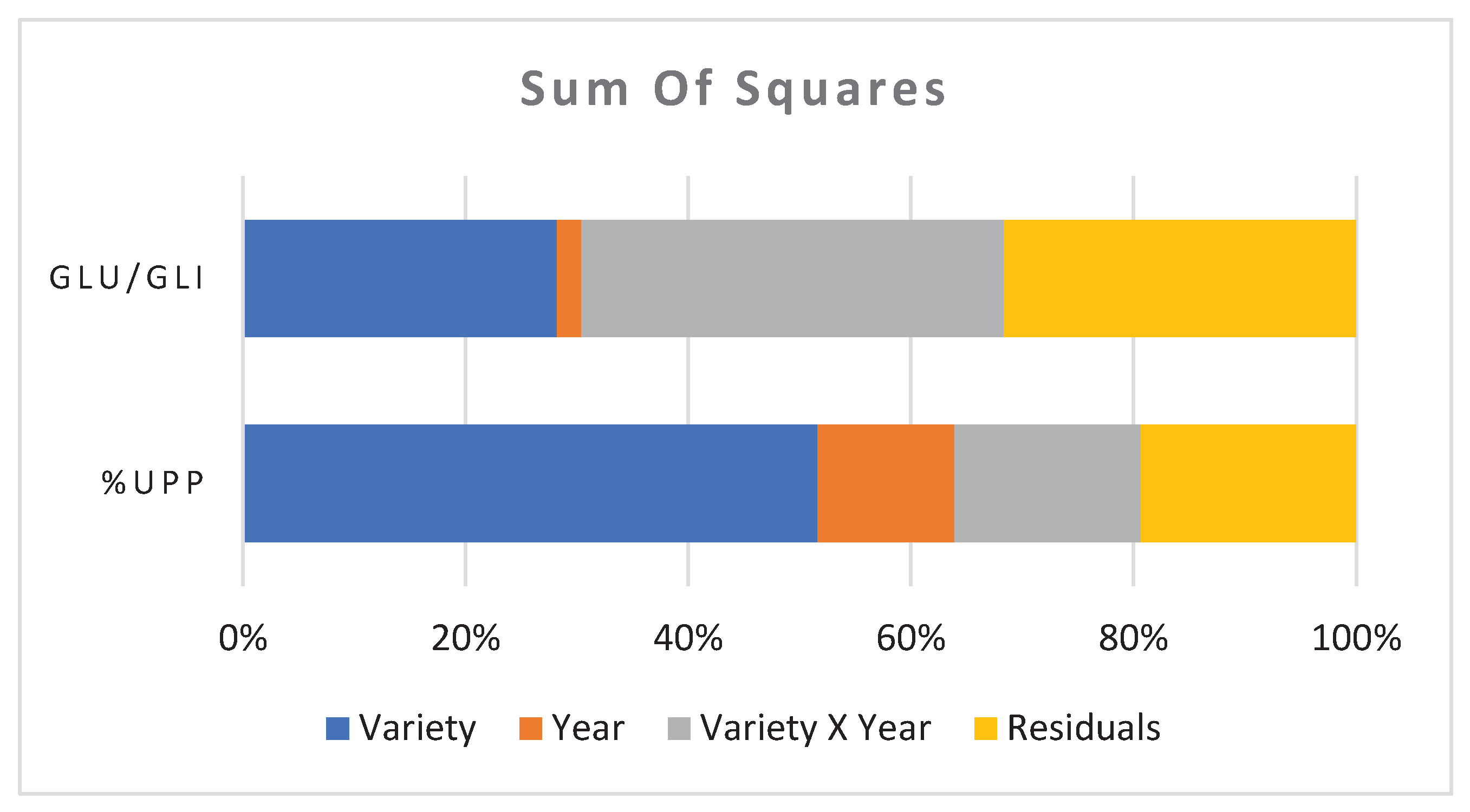

Table 1.
Details of the allelic composition of gluten genes of Sicilian landraces. HMW-GS coded by Glu-A1 and Glu-B1 loci are listed in sequence.
Table 1.
Details of the allelic composition of gluten genes of Sicilian landraces. HMW-GS coded by Glu-A1 and Glu-B1 loci are listed in sequence.
| ID_CREA | Conservation Varieties | HMW-GS | LMW-GS Type | g-gliadins |
|---|---|---|---|---|
| FD-BIA-1 | Biancuccia | null,6+8 | 2 | 42 |
| FD-BID-1 | Bidì | null,20x+20y | 2 | 45 |
| FD-BID-2 | Bidì | null,20x+20y | 2 | 45 |
| FD-BID-3 | Bidì | null,20x+20y | 2 | 45 |
| FD-BID-5 | Bidì | null,20x+20y | 2 | 45 |
| FD-BIV-1 | Bivona | 2*,13+16 | 1 | 44 |
| FD-CAP-1 | Capeiti 8 | null,20+20 | 2 | 45 |
| FD-CAS-2 | Castiglione Glabro | null,13+16 | 1 | 45 |
| FD-CAS-3 | Castiglione Glabro | null,13+16 | 1 | 45 |
| FD-CIC-1 | Ciciredda | 2*,32+33 | 2 | 45 |
| FD-FAR-1 | Faricello | null,13+16 | 1 | 42 |
| FD-FAR-2 | Faricello | null,13+16 | 1 | 42 |
| FD-FRA-1 | Francesa | null,6+8 | 2 | 45 |
| FD-GIO-1 | Gioia | null,6+8/13+16 | 1 | 45 |
| FD-GIO-2 | Gioia | null,13+16 | 1 | 45 |
| FD-GIU-1 | Giustalisa | null,13+16 | 2 | 47 |
| FD-MAR-1 | Martinella | null,13+16 | 2 | 45 |
| FD-PER-1 | Perciasacchi | null,6+8/20x+20y | 2 | 45 |
| FD-PER-2 | Perciasacchi | null,6+8/20x+20y | 2 | 45 |
| FD-PER-3 | Perciasacchi | null,6+8 | 2 | 45 |
| FD-PER-4 | Perciasacchi | null,6+8/20x+20y | 2 | 45 |
| FD-PER-6 | Perciasacchi | null,6+8 | 2 | 45 |
| FD-PER-7 | Perciasacchi | null,6+8/20+20 | 2 | 45 |
| FD-PER-8 | Perciasacchi | null,6+8 | 2 | 45 |
| FD-PER-9 | Perciasacchi | null,20x+20y | 2 | 45 |
| FD-PER-13 | Perciasacchi | null,6+8 | 2 | 45 |
| FD-RUI-1 | Ruscia | null,20x+20y | 2 | 45 |
| FD-RUI-3 | Ruscia | null,20x+20y | 2 | 45 |
| FD-RUI-4 | Ruscia | null,20x+20y | 2 | 45 |
| FD-RUS-1 | Russello | 2*,13+16/6+8 | 1 | 45 |
| FD-RUS-3 | Russello | 2*,13+16/6+8 | 1 | 45 |
| FD-RUS-4 | Russello | null,13+16/6+8 | 1 | 45 |
| FD-RUS-6 | Russello | null,13+16 | 1 | 45 |
| FD-SAM-1 | Sammartinara | null,13+16 | 2 | 47 |
| FD SCA-1 | Scavuzza | null,6+8 | 2 | 44 |
| FD-SCO-1 | Scorsonera | null,20 | 2 | 45 |
| FD-SCO-2 | Scorsonera | null,20 | 2 | 45 |
| FD-TRB-1 | Timilia R.B. | null,6+8 | 2 | 44 |
| FD-TRB-2 | Timilia R.B. | null,6+8 | 2 | 44 |
| FD-TRB-4 | Timilia R.B. | null,6+8 | 2 | 44 |
| FD-TRI-1 | Tripolino | null,6+8/13+16 | 1 | 42 |
| FD-TRN-1 | Timilia R.N. | null,6+8 | 2 | 44 |
| FD-TRN-3 | Timilia R.N. | null,6+8 | 2 | 44 |
| FD-TRN-6 | Timilia R.N. | null,6+8 | 2 | 44 |
| FD-TRN-7 | Timilia R.N. | null,6+8 | 2 | 44 |
| FD-TRN-8 | Timilia R.N. | null,6+8 | 2 | 44 |
| FD-TRN-9 | Timilia R.N. | null,6+8 | 2 | 44 |
| FD-TRN-11 | Timilia R.N. | null,6+8 | 2 | 44 |
| FD-TRN-12 | Timilia R.N. | null,6+8 | 2 | 44 |
| FD-URR-1 | Urria | null,20x+20y | 2 | 47 |
| FD-VAL-2 | Vallelunga pubescente | null,13+16 | 2 | 45 |
Table 2.
Content of %UPP and Glu/Gli ratio of different landraces related to the two growing seasons.
Table 2.
Content of %UPP and Glu/Gli ratio of different landraces related to the two growing seasons.
| %UPP | Glu/Gli | ||||
|---|---|---|---|---|---|
| Name | Type | 2019 | 2020 | 2019 | 2020 |
| Bidì | Landrace | 33.9 ± 1.0 ab | 38.8 ± 2.6 bc | 0.480 ± 0.021 c | 0.734 ± 0.040 a* |
| Castiglione Glabro | Landrace | 29 ± 1.2 abc | 28.1 ± 0.3 de | 0.639 ± 0.042 ab* | 0.412 ± 0.026 d |
| Gioia | Landrace | 29.4 ± 1.1 abc | 31.2 ± 0.8 de | 0.465 ± 0.037 c | 0.556 ± 0.004 bcd |
| Perciasacchi | Landrace | 25.9 ± 1.7 c | 38.9 ± 3.0 c* | 0.586 ± 0.024 bc | 0.505 ± 0.031 cd |
| Russello | Landrace | 36.9 ± 1.2 a | 44.2 ± 2.0 b* | 0.622 ± 0.018 ab | 0.573 ± 0.029 bc |
| Ruscia | Landrace | 15.1 ± 0.5 d | 25.2 ± 1.3 e* | 0.560 ± 0.016 bc | 0.492 ± 0.056 cd |
| Saragolla | Modern Cultivar | 30.8 ± 3.4 abc | 50.7 ± 2.2 a* | 0.580 ± 0.031 bc | 0.664 ± 0.017 ab* |
| Senatore Cappelli | Old Cultivar | 28.3 ± 1.6 bc* | 22.4 ± 1.9 e | 0.723 ± 0.017 a* | 0.559 ± 0.014 bcd |
| Timilia Reste Bianche | Landrace | 30.1 ± 0.7 abc | 30.3 ± 1.0 de | 0.530 ± 0.022 bc* | 0.424 ± 0.021 d |
| Timilia Reste Nere | Landrace | 27.2 ± 1.4 bc | 33.5 ± 1.1 d* | 0.519 ± 0.031 bc* | 0.428 ± 0.018 d |
* For each landrace, means within columns followed by different lower-case letters refer to the statistical analysis performed using one-way analysis of variance (ANOVA) with post-hoc Tukey HSD test (p<0.05). ** For each sample, means within raw followed by asterisks indicate a significant difference in in the two different years. The statistical analysis was performed using the Student's T test (p<0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



