Submitted:
07 February 2024
Posted:
08 February 2024
You are already at the latest version
Abstract
Extracellular vesicles (EVs), a diverse group of cell-derived exocytosed particles, are pivotal in mediating intercellular communication due to their ability to selectively transfer biomolecules to specific cell types. EVs, composed of proteins, nucleic acids, and lipids, are taken up by cells to affect a variety of signaling cascades. Research in the field has primarily focused on stem-cell derived EVs, with a particular focus on mesenchymal stem cells, for their potential therapeutic benefits. Recently, tissue specific EVs or cell type specific extracellular vesicles (CTS-EVs), have garnered attention for their unique biogenesis and molecular composition because they enable highly targeted cell-specific communication. Various studies have outlined the roles that CTS-EVs play in signaling for physiological function and maintenance of homeostasis, including immune modulation, tissue regeneration, and organ development. These properties are also exploited for disease propagation, such as in cancer, neurological disorders, infectious diseases, autoimmune conditions, and more. The insights gained from analyzing CTS-EVs in different biological roles not only enhance our understanding of intercellular signaling and disease pathogenesis, but also open new avenues for innovative diagnostic biomarkers and therapeutic targets for a wide spectrum of medical conditions. This review comprehensively outlines current understanding of CTS-EV origins, function within normal physiology, and implications in diseased states.
Keywords:
cell type specific extracellular vesicles
; extracellular vesicles
; exosomes
; intercellular communication
; diagnostic biomarkers
; disease pathogensis
1. Introduction
Extracellular vesicles (EVs) are a diverse group of lipid bilayer-encased particles that are characterized by their heterogeneity in size (ranging from 30 to 1000 nm), origin (secreted by nearly all cell types), and composition [1,2]. These endogenously produced particles have inherent capacity to convey a wide range of bioactive molecules, comprising of proteins, nucleic acids, and lipids. EV cargo is absorbed and utilized to elicit a cellular signaling response, and thus can play a crucial role in intra and intercellular communications, angiogenesis, cancer progression, homeostasis, immune, and inflammatory responses [3,4,5,6,7]. There are multiple classifications of EVs based on their origin, size, and functionality. Three major sub-types of EVs are apoptotic bodies (1000-5000 nm), microvesicles (MVs) or ectosomes (100-1500 nm), and exosomes (30-100 nm) [1,8,9,10]. EVs are found in all body fluids, including plasma, urine, breast milk, saliva and tears [11]. Their widely available biodistribution not only signifies their role in normal physiological activities, but also their potential to affect pathological states [12].The field of EV detection, purification, and analysis is still in its infancy and the best techniques are still being debated [13].
The field of EVs is gaining great attention because of EVs’ unique role in intercellular communication. Predominant research in this field has concentrated on therapeutic applications of EVs derived from mesenchymal stem cells (EV-MSCs) which have demonstrated a significant effect across diverse diseases including [14,15], neurological disorders [3], cardiovascular disorders [16], respiratory disorders [17], metabolic disorders [18], ophthalmic conditions [19] and various cancers [20,21]. However, more recently, cell-type specific EVs (CTS-EVs) have also begun to be recognized as pivotal entities in the realm of intercellular communication, contributing significantly to our understanding of cellular interactions in both physiological and pathological contexts. CTS-EVs are essential for orchestrating a range of homeostatic cellular activities including growth, proliferation, and differentiation, and their composition changes in response to cellular stress and pathological status. Their tissue-specific origins allow them to transport specific biomolecules from their parent cells and enables them to selectively influence target cells and the surrounding microenvironment, thereby facilitating specific cellular functions and signaling pathways critical for maintaining homeostasis, differentiation, immune modulation, and tissue regeneration [22]. Additionally, this specificity and ability to be extracted from biofluids enables CTS-EVs to serve as valuable biomarkers of disease [23]. Dysregulation of CTS-EV mediated intercellular communication has been implicated in various diseases, including cancer, neurological disorders, autoimmune conditions, cardiovascular diseases, and more (Figure 1) [24]. Therefore, understanding the mechanisms and functions of CTS-EVs can provide insights into disease pathogenesis and potentially lead to the identification of both diagnostic and therapeutic targets. In the present review we will highlight the role of CTS-EVs in both physiological and pathological conditions.
2. EV Origins and Intercellular Communication
2.1. Biogenesis and Release
CTS-EVs are produced through intricate biogenesis pathways unique to their cell of origin. These vesicles originate from endosomal compartments or plasma membranes depending on the type of EV [25]. Apoptotic bodies are the largest EVs, roughly 1 to 5 μm in diameter, making them in the size range of smaller cells [26]. Apoptotic bodies, as the name implies, are only formed during programmed cell death [27]. Microvesicles (MVs) are a subtype of EVs that range in size from 100 to 1000 nm [9,10]. They are formed and released via outward budding and fission of the plasma membrane (blebbing), though their role in intercellular communication is less well defined compared to other EV classifications [25]. Exosomes, with size in the 30-150 nm range [3,28], are formed within endosomal compartments. The process begins via endocytosis of the cell’s plasma membrane with internalized cargoes to form an early endosome [29]. These early endosomes mature into multivesicular bodies (MVBs), rich with many intraluminal vesicles (ILVs). ILV formation relies on reorganization via tetraspanins and recruitment via endosomal sorting complexes required for transport (ESCRTs) [25,30,31]. These ILVs contain a variety of cargo loaded from the Golgi apparatus. The MVB is then transported to the plasma membrane along the cytoskeletal and microtubule network. Once the MVB docks with the plasma membrane, the exocytosis process occurs and the various intraluminal vesicles, now called exosomes, are released into the extracellular space [29]. They either remain in the extracellular space or release throughout wider vasculature, with widely available biodistribution being observed in most organs [32]. Most research on CTS-EVs is largely attributed to the activities of cell type specific exosomes.
2.2. Molecular Composition
The content of CTS-EVs varies with respect to their cell type, physiological condition, and disease status [25]. Proteomic and genomic analyses reveal that CTS-EVs contain a wide array of bioactive molecules, including a number of cell-specific nucleic acids, proteins, lipids, and glycans [33]. miRNAs are one of the most abundant nucleic acids in EVs, and they play a variety of roles in mediated cellular communication [34]. For example, miR-21 and 26a may play a role in cardiac hypertrophy remodeling while miR-233 and 23a have been described as regulators of macrophage function in hypoxia and inflammation [35,36]. Other nucleic acid species carried by EVs include ribosomal RNA (rRNA), long non-coding RNA (lncRNA), transfer RNA (tRNA), and small nuclear RNA (snRNA), all of which have different influences on physiological processes [34].
In terms of proteins, CTS-EVs carry both proteins generalized to all EVs with regards to their biogenesis/release as well as effector functions for their microenvironment. Among these effector proteins include transport proteins, such as annexins and Rab [37]. Donor cells may release exosomes contain MHC Class I and II proteins as part of immune surveillance, while cancer cells may do the same for immune avoidance [38]. Cancer cell EVs can also release antiapoptotic factors such as thioredoxine, and peroxidase for similar reasons [39]. Some CTS-EVs carry metabolic enzymes, including fatty acid synthase, G3P-dehydrogenase, ATPase, and more [37]. Other EVs have been shown to transport receptors such as tumor necrosis factor (TNF)-α receptors, transforming growth factor (TGF)-β receptors, and transferring receptors [34]. Lipids not only have a structural role in CTS-EV membranes, but also contribute to regulation of release and communication. EVs are enriched in cholesterol, sphingomyelin, glycosphingolipids, phosphatidylserine, and other fatty acids [40]. Membrane lipids have been observed to interact directly with receptor molecules or act as second messenger molecules, thus allowing EVs to have multiple mechanisms to participate in cell signaling [41]. These bioactive lipids from EVs can be internalized into recipient cells, with an example study showing that accumulations of fatty acids internalized by exosomes creating a prostaglandin-dependent biological response [34,42].
Finally, glycans are typically found as part of glycoproteins and glycolipids. These complex molecules are crucial for exosome biogenesis, release, uptake, and interaction with recipient cells [43]. The specific types of glycans present in exosomes can serve as biomarkers for certain diseases or conditions, and they can influence the biological functions of the exosomes [43,44].
2.3. CTS-EVs in Intercellular Communication
EVs have been observed to interact in cell signaling in two primary methods: direct interaction and membrane fusion. In direct interaction, a surface ligand on an EV binds with a target surface bound receptor, which initiates a downstream cellular signaling cascade [29]. This can directly impact cellular functions and may additionally trigger endocytosis of the EV. In the second method, EVs fuse with the target cells plasma membrane directly and release their contents into the cytoplasm [29]. Families of SNARE and Rab proteins modulate this fusion, ensuring a cell only accepts certain EVs [45]. Lysosomes and other phagosomes break apart the lipid bilayer of the EV to expose the internal contents [46]. The various RNAs and proteins carried by the EV are released, which release can affect gene expression and modulation of intracellular signaling pathways. CTS-EVs exhibit a remarkable ability to target specific cell types with individualized effects, a feature that is crucial for their role in selective intercellular communication (Figure 1). And due to the sheer volume of EVs released, they play vital roles in maintaining the overall balance of homeostasis in the body. Examples of this will be outlined in sections below.
3. CTS-EVs in Physiology and Pathology
3.1. Normal Physiological Function
In normal physiological processes, CTS-EVs play a fundamental role in maintaining normal physiological functions through their involvement in various aspects of cellular communication and homeostasis. EVs are capable of traversing various barriers within the body (including the blood-brain barrier between the bloodstream and brain parenchyma) to facilitate intercellular communication and transport biomolecules between different physiological compartments. The cell-specific nature of their cargo ensures that their actions are precise and targeted, thereby facilitating the fine-tuning of changes in response to tissue status [47].
CTS-EVs have emerged as crucial modulators of the immune system [48]. They possess the ability to convey signals that either activate or suppress immune responses, thereby playing a pivotal role in immune regulation. All immune cell types engaged in inflammation can produce EVs, and these EVs serve multiple functions in inflammatory processes [49,50]. EVs carry bioactive lipids, enzymes, and even molecules known as eicosanoids, which guide the movement of cells towards a specific location (chemotactic effects) [51]. Eicosanoids are signaling molecules that create chemical gradients in the body’s tissues. Cells, particularly immune cells, respond to these gradients and migrate towards the source of the eicosanoid signal. This chemotactic response is crucial in processes such as inflammation, where such signaling directs immune cells to sites of injury or infection. Through the transfer of these chemotactic agents, EVs facilitate critical interactions and responses in the body’s immune and healing processes.
CTS EVs are also shown to be pivotal in innate and adaptive immunity, including but not limited to inflammation, antigen presentation, and the maturation and activation of both B cells and T cells. Studies on exosomes isolated from human thymic epithelial cells (TECs), which are present in thymic tissue in both mice and humans, were shown to carry tissue-restricted antigens (TRAs) such as myelin basic protein and desmoglein 3. This finding suggested that exosomes derived from thymic epithelia could play a crucial role in the thymocyte selection process. By disseminating self-antigens, TEC exosomes potentially enhance the exposure of maturing thymocytes to a wider range of TRAs, aiding in the critical processes of positive and negative selection within the thymus and thus proper functioning of central tolerance mechanisms to prevent immune reactions [49,52]. CTS-EVs are also known to transport growth factors and signaling molecules that promote cell proliferation and differentiation, aiding in tissue regeneration and maintenance [53,54]. Dermal-related CTS-EVs, including those from dermal fibroblasts and keratinocytes, contribute to extracellular matrix (ECM) deposition, and tissue remodeling [55,56]. Moreover, blood-derived EVs, such as those from platelet-rich plasma, umbilical cord blood, and endothelial cells, can regulate fibroblasts and contribute to angiogenesis in cutaneous tissue [57,58].
EVs are essential for the development and maintenance of the central nervous system (CNS) and are secreted from all types of neural cells including neurons, astrocytes, microglia, and oligodendrocytes. These vesicles play critical roles in neuronal communication, synaptic function, and the regulation of neurogenesis [59,60,61]. Early research in this field revealed a correlation between enhanced neuronal activity and an uptick in the release of extracellular vesicles, a process thought to be crucial for the effective maintenance of synapses. CTS-EVs facilitate the exchange of proteins, miRNAs, and other signaling molecules between cells, contributing to the dynamic regulation of the neural environment, both in the CNS and peripheral nerves such as in the cornea [6,62]. This exchange helps maintain synaptic plasticity, which underlies learning and memory. Lopez et al found that CTS-EVs from human neural rosettes and pluripotent stem cells (PSCs) play a critical role in human CNS development [63]. These EVs carry neuronal and glial cellular components necessary for brain development, and play a role in influencing PSC morphology [64]. In addition to synaptic pruning, the regional development of the CNS is significantly influenced by intercellular communication. Sonic hedgehog (Shh) signaling, a key regulator of cortical development, exemplifies this. Tanaka and colleagues observed that the embryonic fluid flow in developing mice causes the release of EVs in a leftward direction. These EVs, carrying Shh, contribute to the lateralization of the CNS, highlighting their role in developmental processes [65].
CTS-EVs are vital in regulating metabolic processes, including insulin sensitivity and lipid metabolism [66,67]. Studies reveal that EVs from adipose tissues modulate insulin signaling pathways, impacting glucose homeostasis and peripheral insulin sensitivity [68,69]. These vesicles are also important in lipid metabolism, transporting lipids and lipid-related enzymes and receptors, primarily in liver and adipose tissues, where they influence lipid storage and breakdown [70]. Additionally, EVs facilitate metabolic communication between organs like the liver, adipose tissue, and skeletal muscle, essential for maintaining energy balance and metabolic stability [71]. These insights underscore the importance of EVs in metabolic regulation and their growing relevance in metabolic disease research and treatment. Additionally, in current cell-based and tissue engineering approaches, including co-culture systems, there is growing evidence that the differentiation of various cell types, including neurons, is remarkably improved with the presence of specific surrounding cells. Neuronal differentiation benefits from the presence of cells like meningeal cells or astrocytes [64,72]. However, there is also evidence indicating that the secretion of soluble factors alone, without the direct inclusion of the supporting cells, may be sufficient to augment the differentiation and maturation processes. These secreted factors, including EVs, can improve key aspects of cellular differentiation by delivering a range of bioactive molecules, such as proteins, lipids, and various types of RNA [73,74].
In addition to CTS EVs roles in normal physiological processes, CTS-EVs can either exacerbate disease progression or function as potent (or impotent) modulators in various disease states. Table 1 systematically catalogs the current understanding of CTS-EVs’ involvement across a diverse array of diseases and disorders, categorizing them according to disease type for ease of reference. Subsequent sections of this review follows the sectional numbering provided in Table 1.
3.2. Cancer
Within the context of cancer, EVs from both tumor and non-tumor sources have been shown to play a role in tumor migration, immunosuppression, and angiogenesis. Cancer CTS-EVs can contain metalloproteinases (MMPs) which degrade ECM, allowing for local invasion of tumor cells [75]. Furthermore, EVs from cancer cells can cause fibroblast dedifferentiation into cancer-associated fibroblasts, which can lead to metastasis [76]. It has been shown that tumor-derived EVs can carry immunosuppressive cargo that prevents immune cells from attacking circulating tumor cells. One such “cargo” is the protein programmed death ligand 1 (PD-L1). When this receptor binds with T cells, it may induce immunosuppression [77]. Natural killer (NK) cells are also affected by a similar mechanism, where EVs expressing ligands of NKG2D lead to the immunosuppression of NK cells [78]. Tumor-derived EVs can also stimulate macrophages to release the factors that promote angiogenesis, further increasing the tumor’s vascular permeability [79]. EVs from cancer stem cells (CSCs) are linked to an increase in epithelial-mesenchymal transition (EMT) status, which can enhance the aggressiveness of the cancer. For instance, miR-21, which has been involved in many antiapoptotic pathways, is a common component of tumor-derived EVs [80]. In vitro studies also demonstrated that when lung cancer cells are treated with CSC-derived EVs, the lung cancer cells become more invasive [81].
Due to the dramatic impact of EVs on cancer progression, it is important to study the composition of cancer-associated EVs to help diagnose and treat cancer. In certain cancers, including melanoma, glioblastoma, prostate cancer, breast cancer, and more, there is a higher concentration of all circulating EVs in plasma for cancer patients versus healthy donors [82,83,84,85,86]. However, the true cause for the increased EV count is still controversial. There is some evidence that blood serum EV levels drop after the primary tumor mass is surgically removed [82,87]. In addition, there is evidence that cancer can be detected not just by counting the number of EVs in a sample, but also by identifying specific biomarkers within these EVs found in liquid biopsies, like blood and urine [82,88]. Tumor-specific EV biomarkers may provide a more precise method for cancer diagnosis than tissue biopsies. Several studies report that analyzing the contents of EVs by way of liquid biopsies can be a successful alternative to more invasive tissue biopsies [82,88,89,90]. Some of the biomarkers found in EVs include miR-1246 (breast cancer), PSA (prostate cancer), and a highly upregulated in liver cancer lncRNA (liver cancer) [82]. EVs that are known to target cancer can be modified to incorporate strategies to combat cancer [91]. For example, lung cancer cell-derived EVs can be loaded with an oncolytic adenovirus combined with chemotherapeutic drugs [92]. EVs derived from immune cells have also been shown to be helpful for cancer therapy [93]. EVs that target cancer cells can also be used to deliver gene therapy—namely the CRISPR/Cas9 system—and can be effective for normally hard-to-transfect cells and cancer cells [94]. M1 macrophage-derived EVs combined with PD-L1, a checkpoint inhibitor, also enhanced cancer therapy [95]. The diagnostic and therapeutic use of EVs has been explored for many different cancers, including breast cancer, lung cancer, lung cancer, prostate cancer, leukemia, ovarian cancer, and glioblastoma. Examples are given below.
For breast cancer diagnosis, specifically, there are several potential biomarkers. Connexin-46 (Cx-46), a protein that is important for gap junction channels [96], is shown to be present in high concentrations in inflammatory breast cancer (IBC) derived-EVs [97]. Inflammatory carcinoma of the breast is an often missed diagnosis and is confused with mastitis, so having a liquid biopsy based cancer detection could be transformational [91,98]. For detecting whether a breast cancer is estrogen-receptor based or not, 241 unique serum-derived EV (sEV) proteins were profiled [90]. sEV protein biomarkers can also be used for breast cancer prognosis; higher levels of developmental endothelial locus-1 (Del-1) from plasma-derived sEVs was used as a prognostic indicator [99,100]. Clinical trials have been conducted to use EVs in diagnostics, including a study from 2014 to use tumor derived exosomes as a diagnostic and prognostic marker [101]. To target triple-negative breast cancer, engineered EVs were loaded with Adriamycin, a chemotherapeutic, and a modified version of miR-159 [102]. EVs tend to act as fertilizers for the tissue in which they were sourced from—thus leading to a very large range of options in terms of cancer therapy [79]. A handful of circulating nucleic acids can be used for diagnostic and prognosis of lung cancer [103]. One such example is a clinical trial using ctDNA from pulmonary exosomes to identify benign vs malignant pulmonary nodules [104]. EVs can also be used to detect lung cancer treatment effectiveness, as EVs extracted from a non-small cell lung cancer cell line that were treated with gefitinib were found to reduce cisplatin’s antitumor effectiveness [105,106]. Cisplatin usually increases autophagy of cells; however, it was found to have an antagonistic relationship with gefitinib [105]. Several EV-based biomarkers have been found for prostate cancer, including a test for TMPRSS2:ERG in urine-derived EVs, which mimics the results of prostate biopsies. There are additional EV-based genes that can be used as biomarkers including PCA3, ERG, BIRC5, and TMPRRS2 [107]. Several drug resistance biomarkers were also found for prostate cancer prognosis [88]. For detecting castrate-resistant prostate cancer, a form of prostate cancer that persists after androgen depletion therapy, the presence of androgen receptor AR-Variant 7 in plasma-derived EVs was of special interest [108]. In one of the more common forms of leukemia, acute lymphoblastic leukemia (ALL), leukemic cells released EVs containing miR-43a-5p to bone marrow [109]—which caused inhibition of osteogenesis, leading to leukemia progression [110]. In acute myeloid leukemia (AML), a panel of miRNAs from serum EVs can be used to distinguish leukemic mice from controls [108,111]. For the treatment of chronic lymphocytic leukemia (CLL), a study used B cells that were targeted by EVs tagged with a viral envelope protein (gp350) and transferred gp350 and CD40L to the patients’ cells, leading to an anti-CLL immune response [112]. Other proteins can be transferred to patients’ cells, such as pp65 [113], or chemotherapeutics like fludarabine [112,114]. In ovarian cancer, there are a handful of potential protein biomarkers for diagnosis. A strong CD24 presence also coincided with epCAM, a known marker of cancer, in EVs from an ovarian cancer cell line, which confirmed that the strong expression of CD24 was indeed tumor-derived [115]. Keseru et al claims that the mitochondrial DNA (mtDNA) copy number found in plasma derived EVs was significantly greater in all stages (except stage 1) of ovarian cancer [116], which could prove to be a valuable tool in detecting early-stage ovarian cancer [117]. A specially-made EV engineering system named Immune Derived Exosome Mimetics (IDEM) containing doxorubicin was able to be produced in large quantities and was effective in drug delivery for treatment of ovarian cancer cells [118]. There are other EV-based therapeutic approaches for ovarian cancer, such as ExoSTING, that are being developed [117]. EVs in urine have shown promise for enhancing glioblastoma identification in an assay called “liquid gold.” A study concluded that EV concentration in urine can be correlated to glioblastoma prognosis, prompting further investigation towards the diagnostic capabilities of EVs [119]. Current clinical trials are ongoing, including one demonstrating detection by next generation sequencing of molecular abnormalities present in exosomes from glioblastomas to non-invasively identify tumor subtypes [120].
3.3. Neurological Disorders
Traditionally, communication within the brain’s neural network has been attributed primarily to synaptic vesicles moving from pre- to post-synaptic neurons [121]. However, recent studies show that CTS-EVs released from neurons, astrocytes, oligodendrocytes, and microglia represent an emerging mode of intraneuronal communication [122]. These CTS-EVs transfer proteins and genomic materials between neurons, playing a role in both normal neuronal functions and the propagation of pathological proteins in neurodegenerative diseases. CTS-EVs can offer a non-invasive approach to diagnosing and monitoring neurological disorders, as well as a potential therapeutic tool for treating these conditions. Because EVs can pass through the blood brain barrier, if loaded with therapeutics, they can also transport therapeutics through the blood brain barrier [122]. Alzheimer’s disease (AD) is characterized by memory, cognitive functioning, and behavioral impairments because of abnormal neurofibrillary tangles and neurotic plaques. AD is known to induce β-amyloid plaque sheaths in the neural membrane. Neuron-derived CTS-EVs are known to transport amyloid-β peptides and tau proteins, which are key in the disease’s progression [123]. These vesicles assist in spreading these neurotoxic agents, leading to synaptic dysfunction and widespread neuronal damage [124]. Due to the nanoparticle size of EVs, biolabeling the particles with microRNA markers allows their location to be visualized [125]. Specifically, miR-9-5p, miR-598, and miR-125b are shown to be highly stable and proliferative, which allows for identification in patients [125,126,127,128]. You et al identified a specific protein, integrin β1, that was found to be elevated in CTS-EVs from astrocytes associated with brain β-amyloid and tau load [129].
Parkinson’s disease (PD) is a neurodegenerative disease characterized by tremors, rigidity, and akinesia. In PD, the dopaminergic neurons of the substantia nigra are lost or reduced [130,131]. EVs can propagate PD pathogenesis by interacting and transporting α-synuclein, a protein whose misfolding and aggregation are hallmark features of the disease. It has been demonstrated that plasma EVs containing α-synuclein can be used as a minimally invasive biomarker of PD [132]. Additionally, EVs can be shown to deliver dopamine across the blood brain barrier, interacting with appropriate receptors and reducing the negative effects of dopamine deficiency [133]. Multiple Sclerosis (MS), a central nervous system autoimmune disease, is characterized by inflammation and immune dysfunction caused by demyelination of the CNS. CTS-EVs originating from distinct cell types are implicated in MS pathology, encapsulating specific biomarkers, including myelin-derived proteins such as myelin basic protein (MBP), proteolipid protein (PLP), and myelin oligodendrocyte glycoprotein (MOG). These biomarkers are indicative of the autoimmune response targeting myelin components in MS [134]. The investigation of CTS-EVs allows for a meticulous dissection of the molecular nuances, offering a targeted understanding of immune dysregulation in MS. Moreover, the bioactive cargo within these EVs, consisting of anti-inflammatory cytokines and neurotrophic factors, present tailored opportunities for therapeutic modulation, aiming at specific cellular pathways contributing to MS pathogenesis [134,135]. Oligodendrocyte derived EVs (OD-EVs) have proved to be useful markers of cerebrovascular accidents (strokes). Oligodendrocytes provide an established role in remyelination in post ischemic injury and influence neuronal survival [136,137]. Fröhlich et al examined the effects of OD-EVs in an oligodendrocyte/neuron co-culture stressed with ischemic injury, and saw that cells treated with OD-EVs had significantly higher metabolic activity compared to control cells [138]. OD-EVs appeared to contain cargo including enzymes, such as catalase and superoxide dismutase 1, that provided antioxidant benefit [138]. Broadly, knowledge of EVs derived from OD-EVs and EVs in the context of stroke remain limited with more extensive research efforts needed [139].
Traumatic brain injury (TBI) is difficult to assess and identify because no known biomarkers are readily accessible and imaging techniques lack the specificity to visualize specific affected brain injuries. EVs can pass through the blood brain barrier which can enable them to be engineered to deliver molecular tracers, thus potentially illuminating injured areas within the brain without the need for high resolution scanning neurotechnology. EVs are also implicated with neuronal – glial cell communication and reduced neuroinflammation when derived from the neuronal cells of the CNS [140].
Mental disorders encompass a wide range of neurological diseases. Depression, for example, is thought to be caused by neuroinflammation and hormone imbalance but the effect of CTS-EVs in depression has not been thoroughly investigated. A study showed that exosomes, derived from patients with major depressive disorder, when injected into healthy mice caused the mice to exhibit depressive-like behavior. This study found the major difference in these exosomes compared with exosomes from healthy persons was hsa-miR-139-5p upregulation [141]. Another study investigated the expression levels of microRNA-26a (miR-26a) in EVs in the hippocampus of depressed rats [142]. The study uncovers that the expression of miR-26a is significantly reduced in the hippocampus of rats with depression. This discovery positions miR-26a as a potential diagnostic marker for depression, highlighting its importance in identifying the condition. In amyotrophic lateral sclerosis (ALS), EVs are associated with TDP-43 fragments and SOD1 mutations. While TDP-43 pathology correlates with disease severity, suggesting potential use as a biomarker, associations with SOD1 mutations remain unclear, casting doubt on their utility for biomarker monitoring [143]. Misfolded proteins associated with EVs represent a promising avenue for biomarker discovery in prion diseases, as they may allow the detection of aberrant isoforms. Studies in EV isolation techniques have allowed identification of intercellular communication mechanisms through CTS-EVs that contribute to infection in prion disease [144,145]. Transport of misfolded proteins also extends to Huntington’s disease, where the mutant huntingtin protein is exported via vesicles. Detection of mutant huntingtin in EVs in plasma provides a less invasive way to track disease progression [146]. Changes in the viscoelasticity of EVs induced by mutant huntingtin offer a new approach for rapid identification of misfolded proteins.
3.4. Ophthalmic Conditions
CTS-EVs have been studied for their potential role as biomarkers or in the broader context of diagnostics in various diseases of the eye. One example is the use of CTS-EVs derived from the retinal pigment epithelium (RPE) as potential biomarkers for age-related macular degeneration (AMD). Klingeborn et al discusses the isolation of retinal EV biomarkers from blood using targeted immunocapture, which represents a promising approach for early AMD biomarker development [147]. The RPE, being a critical location for disease-associated changes in AMD, sheds EVs that enter the systemic circulation and thus could be harnessed for blood-based biomarker discovery. Further research, as mentioned by Kang et al, explores EV proteins in the aqueous humor as potential biomarkers for neovascular AMD, comparing the protein profiles in the aqueous humor from individuals with the disease to unaffected individuals [148]. These studies suggest that EVs, and particularly exosomes, could serve as valuable biomarkers for diagnosing and monitoring the progression of AMD. Furthermore, EVs derived from the urine and retinal tissue of diabetic retinopathy (DR) patients have shown distinct protein profiles when compared to those from healthy individuals. This study found that junction plakoglobin (JUP), a protein associated with cell-cell junctions, was present in EVs from DR patients but not in controls [149]. This suggests that JUP could potentially serve as a biomarker for DR, offering opportunities for earlier diagnosis and better prognosis. Another study explored the impact of plasma-derived EVs on angiogenesis and the progression of proliferative diabetic retinopathy (PDR), with a focus on the role of microRNA-30b. This research suggests that the mechanisms influenced by plasma-derived EVs may regulate angiogenesis in PDR via a mechanism dependent on miR-30b [150].
In the context of glaucoma, recent studies have been investigating the role of CTS-EVs in intraocular pressure regulation and optic nerve damage [151]. Research demonstrates that CTS-EVs play a role in the pathology of glaucoma, particularly in the regulation of intraocular pressure and signaling within the ocular system. By comparing the proteome of EVs isolated from trabecular meshwork (TM) cells of both glaucomatous and non-glaucomatous human donors, it was found that EVs from glaucomatous TM cells had a different protein profile, with a reduced abundance of key extracellular matrix proteins, suggesting a mechanism for ECM regulation in glaucoma and potentially revealing targets for future therapies [152].
Uveal melanoma (UM) is the most common intraocular cancer, which is also known to spread to the liver, causing incurable metastases. It was found by Piquet et al that uveal melanoma cells, when compared to healthy melanocytes, release significantly more EVs that also exhibit hepatotropism [153]. These EVs also induced angiogenesis and fibrosis on hepatic stellate cells and endothelial cells, which was not found with EVs that came from melanocytes. Melanoma EVs have been found to increase metastatic UM cell attachment to the liver through reprogrammed hepatic stellate cells by modifying the extracellular matrix [153].
With regards to Dry Eye Syndrome (DED), Andrew et al explored the association between inflammatory-related tear film microRNAs (miRNAs)-derived EVs and EVs in non-Sjogren’s Syndrome DED [154]. Tear samples from DED and non-DED individuals were analyzed for EVs and miRNAs. The findings revealed EVs in both groups’ tear films, with RNA-Seq identifying 126 differentially expressed miRNAs. Nine of these miRNAs, linked to inflammation, were significantly upregulated in DED patients. The research suggested that miRNAs in tear film EVs are involved in DED’s inflammatory pathways. Another study investigated the role of EVs in the epithelial-to-mesenchymal transition (EMT) of lens epithelial cells under oxidative stress, a process related to posterior capsule opacification (PCO), a common complication after cataract surgery [155]. The study found that EVs and reactive oxygen species (ROS) were involved in this transition. The results suggested that EVs participate in ROS-induced lens EMT, indicating that EVs could be a potential target for treating PCO.
In relation to uveitis, a study conducted in 2023 focused on profiling CTS-EVs in uveitis and scleritis to discover biomarkers and explore mechanisms. This comprehensive analysis involved proteomics of small and large EV subpopulations from patients with various uveitis types and posterior scleritis. The study identified over 3,000 proteins and found that EV proteomic profiles correlated more with disease than plasma proteomic profiles [156]. It highlighted potential pathogenic mechanisms and identified biomarker panels for these diseases, providing insights for diagnosis and potential therapeutic targets.
The human cornea and its optical transparency are crucial for vision [157]. It has been proposed that EVs are actively secreted in corneal epithelial defects to enable communication between stroma and epithelium, suppressing scar formation and maintaining surface clarity [5,157]. Both corneal fibroblast and MSC-derived EVs have been specifically identified able to regulate the inflammatory response and modulate nerve regeneration through transport of signaling proteins such as TGF-β [158,159,160,161]. Another study found corneal epithelial-EVs to have the ability to influence differentiation of conjunctival and corneal epithelial cells, further participating in injury remodeling [158,162]. In another study, the differential impacts of exosomes derived from non-diabetic (N) and diabetic (DM) human limbal epithelial cells (LECs) on limbal stromal cells (LSCs) were analyzed, uncovering critical insights into corneal health and diabetic complications. A key finding of this study was that exosomes from N LECs significantly enhance wound healing and cell proliferation in LSCs, unlike those from DM LECs. This effect was attributed to the distinct miRNA and protein compositions of the exosomes, suggesting a crucial role of cellular communication mechanisms in diabetic corneal conditions [163]. Another study from the same group highlighted the role of miR-146a in corneal wound healing, showing it is upregulated in the diabetic limbus and reduces pro-inflammatory responses in LECs [164]. Overexpression of miR-146a decreased inflammation-related genes and mediators, particularly in diabetic conditions. This suggests miR-146a’s potential as a therapeutic target for managing diabetic corneal complications. Research concerning corneal CTS-EVs and their applications are still in early stages, but show great potential for impacting disease management in the future.
3.5. Infectious Disease:
In the context of infectious diseases, EVs play complex roles. They participate in immune system processes and have been implicated in both the progression and protection against diseases. In infections, EVs can carry viral and host components, modulating the immune response. Proteomic analysis of EVs from patients with infections like SARS-CoV-2 has provided insights into their role in disease progression and potential as biomarkers. These findings point to a critical role for EVs in understanding and managing infectious diseases, with implications for developing new therapeutic and diagnostic tools. Elettra et al conducted a proteomic analysis of circulating exosomes in patients with SARS-CoV-2 to explore the host response to the virus [165]. It revealed that these exosomes carry molecules related to immune response, inflammation, coagulation, and complement pathways, which are crucial in COVID-19 pathogenesis and organ damage. Notably, the research identified potential biomarkers for disease severity and diagnostic accuracy. It also discovered SARS-CoV-2 RNA within exosomes, suggesting a possible mechanism for viral spread [165]. This study provided significant insights into the role of exosomes in inflammation, coagulation, and immune modulation during COVID-19 infection and highlighted their potential as biomarkers.
Furthermore, the research conducted by Masatoshi et al on Hepatitis B Virus (HBV) illustrated that apart from producing infectious virions, HBV also generates various non-infectious particles, including EVs [166]. These EVs, particularly exosomes, play a crucial role in cell-to-cell communication. They are taken up by monocyte/macrophages, primarily leading to the upregulation of PD-L1, which is involved in immune response suppression. Importantly, the study showed that the characteristics and effects of these EVs are modified by nucleotide reverse transcriptase inhibitors (NRTIs), suggesting a significant role in HBV pathogenesis and the clinical response in patients treated with NRTIs.
In the context of HIV, EVs have both pro-viral and anti-viral effects [167]. For example, HIV-1-infected EVs have been demonstrated to carry gp120, a key envelope protein of HIV, which enhances viral entry in lymphoid tissues [168]. They additionally increase disease progression through the use of T-cell immunoglobulin and mucin domain-containing 4 (TIM4), which can facilitate the entry of HIV-1 into host cells. EVs containing surface TIM4 binds with the virus, trafficking of HIV-1 to immune cells is hastened, resulting in increased infectivity and disease progression [169]. With regards to antiviral properties from the host, EVs released by CD8 T cells can carry ligands that have anti-HIV properties [170]. In addition, it has been reported that EVs trigger innate immunity to restrict HIV. When Toll-like receptor 3 is activated in human brain endothelial cells, they release EVs containing interferons and other antiviral factors which can prevent HIV infection [171].
3.6. Autoimmune Diseases:
CTS-EVs have become a focal point in the scientific exploration of autoimmune conditions. Generally, immunosuppression is the pathway followed to treat many autoimmune diseases (AD), though after some time, the complexity of the human immune system counters its efforts [172]. Many of today’s steroidal and non-steroidal therapies have dull results, due to their inability to offer highly specific effects. EV’s on the other hand, have highly configurable packages and can deliver in a targeted fashion [172,173].
Rheumatoid Arthritis is an autoimmune disease leading to a chronic inflammatory state primarily affecting the synovial tissues encapsulating joints. The underlying cause is multifactorial, involving genetic predispositions including human leukocyte antigen (HLA) alleles and environmental triggers [174]. EVs from regulatory T cells (Tregs), characterized by anti-inflammatory microRNAs, may offer a protective influence by suppressing immune responses. Conversely, EVs released by activated immune cells, carrying pro-inflammatory cytokines, could worsen joint inflammation. Biomarkers such as miR-155 and miR-146a in EVs from Tregs may signify immunoregulatory potential, while EVs containing citrullinated proteins or inflammatory mediators may mark heightened autoimmunity [174,175,176]. Additionally, EVs from mesenchymal stem cells may harbor reparative factors like Transforming Growth Factor-β (TGF-β), presenting a positive influence, while those carrying matrix metalloproteinases might contribute to tissue degradation [175,176,177,178].
The exploration of CTS-EVs in systemic lupus erythematosus (SLE) reveals a unique molecular signature indicative of the underlying immune dysregulation. These EVs carry specific biomarkers, including antinuclear antibodies (ANA), anti-double-stranded DNA antibodies (anti-dsDNA), and anti-Smith antibodies, pinpointing cellular targets within SLE [179]. Additionally, cytokines carried by CTS-EVs, such as interleukin-6 (IL-6) and tumor necrosis factor-alpha (TNF-α), contribute to the inflammatory environment associated with SLE [180]. Focusing on biomarkers from implicated cell types, such as B cells and dendritic cells, offers a targeted understanding of SLE pathogenesis. Leveraging these specific biomarkers holds promise for precise diagnostic assays and informs potential therapeutic strategies aimed at modulating the identified molecular pathways contributing to immune dysregulation in SLE [179,180].
In Sjogren’s syndrome, a disorder affecting exocrine glands, the scrutiny of CTS-EVs as diagnostic and therapeutic entities is anchored in their distinct molecular composition [172,181]. Originating from salivary and lacrimal gland cells, these EVs encapsulate hallmark biomarkers, including anti-Ro (SSA) and anti-La (SSB) antibodies, indicative of autoimmune responses [181]. Focusing on CTS-EVs enables targeted exploration of molecular intricacies tied to glandular dysfunction and local inflammation. Additionally, the bioactive cargo within these EVs, containing cytokines like interleukin-17 (IL-17) and interferon-gamma (IFN-γ), provides insight into the pro-inflammatory environment characterizing Sjogren’s syndrome [181,182]. This nuanced investigation facilitates a refined understanding of the disease and lays the groundwork for personalized diagnostics and therapeutic interventions.
In Irritable Bowel Syndrome (IBS), a complex gastrointestinal disorder, the exploration of CTS-EV’s as diagnostic and therapeutic agents involves a meticulous analysis of diverse biomarkers encapsulated within these vesicles [183]. Derived from various cell types in the gastrointestinal tract, these EVs carry biomarkers such as serotonin, tryptase, and inflammatory cytokines, shedding light on altered neurotransmission, mast cell activation, and immune dysregulation inherent to IBS pathology [183,184,185]. The specificity of investigating CTS-EVs enables a targeted examination of the molecular signatures associated with distinct cellular contributors to IBS. Furthermore, the bioactive cargo within these EVs, encompassing neuropeptides, pro-inflammatory mediators, and additional markers like calprotectin and fecal lactoferrin, offers insights into the neuroimmune interactions contributing to symptoms such as altered gut motility and visceral hypersensitivity in IBS [186]. This intricate investigation of cell type-specific-EVs not only deepens our understanding of IBS at the molecular level but also opens avenues for personalized diagnostics and therapeutic strategies by leveraging a comprehensive array of biomarkers. In Myasthenia Gravis (MG), a neuromuscular junction autoimmune disorder, the investigation of CTS-EVs unveils a nuanced molecular landscape. These EVs, originating from diverse neuromuscular junction cell types, encapsulate biomarkers like acetylcholine receptor (AChR) antibodies and muscle-specific kinase (MuSK), offering insights into the immunological responses underlying MG [187,188]. The targeted study of CTS-EVs enables a focused examination of molecular signatures associated with neuromuscular dysfunction. Additionally, the bioactive cargo within these EVs, comprising miRNAs and cytokines, provides a comprehensive view of the immune modulation and inflammatory processes in MG [188]. The emerging research on CTS-EVs in MG holds promise for innovative diagnostics and personalized therapeutic strategies, enhancing our understanding and management of this complex autoimmune disorder.
3.7. Cardiovascular Diseases
Nearly one-third of deaths globally are caused by cardiovascular diseases (CVDs), which constitute a major cause of mortality [189]. According to a number of recent studies, cells associated with the cardiovascular system, including cardiomyocytes, endotheliocytes, fibroblasts, platelets, smooth muscle cells (SMCs), leucocytes, monocytes, and macrophages, can generate EVs [190]. Under physiological conditions, EVs have vital roles in maintaining proper heart structure and function. Conversely, in pathological circumstances, EVs alter their composition and aid in the formation of CVDs [191]. It has been demonstrated that the main sources of EVs in individuals with stable coronary artery disease (CAD) are endotheliocytes and platelets. miR-92a-3p is packaged into endothelial EVs under atherosclerotic circumstances, which in turn controls angiogenesis in recipient endotheliocytes through a process that is reliant on thrombospondin-1 [192]. Through the miR-939-mediated nitric oxide signaling pathway, coronary serum EVs isolated from individuals with myocardial ischemia (MI) induce angiogenesis [193]. In individuals with stable CAD, increased expression of miR-126 and miR-199a in EVs but not plasma has been linked to a decrease in major adverse cardiovascular events [194]. In MI, in response to hypoxia and inflammation, cardiomyocytes release EVs enriched in miRNAs linked to endothelium proliferation and differentiation, such as miR-143 and miR-222 [195]. miR-15-b could also play a mechanistic role in the dysregulation of cardiac and during release from cardiac EVs following ischemic damage [196]. MI has been linked to a circulating EV protein network that also plays a role in the inflammatory response [197]. Furthermore, EVs are essential for the remodeling of the myocardium following a MI, indicating a great deal of potential for therapies involving the upregulation or downregulation of miR-containing EVs [191].
In individuals with atrial fibrillation, miR-21-3p has been detected in EVs and plasma with links to abnormal cardiac tissue expansion. It has been demonstrated that miR-199a-3p in extracellular matrix-derived EVs stimulates the formation of heart tissue and influences atrial electrical activity by upregulating Gata-binding 4 (Gata4) acetylation and suppressing the expression of homeodomain-only protein X (HOPX) [198]. In the development of peripartum cardiomyopathy, endotheliocyte-secreted EVs loaded with miR-146a function as both biomarker and messenger [199]. A 16 kDa N-terminal prolactin fragment (16 K PRL) stimulates endotheliocytes to produce more miR-146a and facilitates the production of EVs laden with miR-146a. Maladaptive hypertrophy is caused by miR-146a, which also decreases the expression of small ubiquitin-like protein modifier 1 (SUMO1) and causes cardiac dysfunction [200]. In summary, CTS EVs play a crucial role in both the maintenance of cardiovascular health and the progression of cardiovascular diseases. CTS EVs, originating from various cell types within the cardiovascular system, are key in regulating heart function and structure under normal conditions. Conversely, in diseased states, they contribute to the development and progression of CVDs through mechanisms involving specific microRNAs and proteins. This dual role of EVs underscores their importance in cardiovascular research and highlights their potential as targets for therapeutic intervention in heart-related diseases.
3.8. Hematologic Disorders
In physiological states, CTS EVs secreted from blood cells, especially platelet-derived EVs, have garnered considerable scientific interest. Their abundance in the bloodstream and pivotal roles in processes such as wound healing, angiogenesis, thrombosis, and atherosclerosis make them a significant focus of study [201,202,203,204]. Isolation of these EVs, which require further study, could aid understanding and treatment for a variety of clotting disorders and thrombocytopenias [205]. Red blood cell-derived EVs (RBC-EVs) have also attracted attention due to their potential role in erythropoiesis, iron homeostasis, and hemolysis. Expelled RBC-EVs contain hemoglobin, which can be taken up by macrophages and recycled, as well as other proteins and lipids that may affect cellular functioning [206]. Harnessing these EVs could aid in replacing hemoglobin in anemias, such as sickle cell anemia. White blood cell-derived EVs (WBC-EVs), including those from neutrophils, monocytes, and lymphocytes, have been implicated in various immune responses, such as phagocytosis, antigen presentation, and cytokine production [50]. WBC-EVs can also transfer antigens and adjuvants to dendritic cells, leading to the activation of T cells and the initiation of adaptive immunity in those who have immunodeficiencies [207].
3.9. Reproductive Disorders
It has been reported that EVs are participating in conception and implantation processes by cross talk with sperm and embryo cells and leads to normal and healthy birth. EVs mediated pathways in reproduction can be explored as an alternative to invasive amniocentesis and chorionic villi sampling [208]. In polycystic ovarian syndrome (PCOS), Sang et al identified miRNAs within CTS-EVs of human follicular fluids that were decreased in PCOS compared with healthy patients [209]. These miRNAs were previously identified to be associated with key roles in the etiology of PCOS in a previous genome. Additionally, CTS-EVs from endometrial stromal cells from women with endometriosis compared to healthy patients demonstrated different profiles of EV miRNA cargo [208,210]. Among these miRNAs, miR21 is already known for a role in angiogenesis, which is associated with pathology of endometriosis.
3.10. Metabolic Diseases
EVs have been implicated in the regulation of metabolic homeostasis. A meta-analysis conducted by Li et al revealed elevated levels of a variety CTS-EVs originating from platelets, endothelial cells, and monocytes in individuals with type 2 diabetes, with cargo including annexin V, CD31, CD104, and CD62E [211,212]. Kobayashi et al found that circulating CTS EVs numbers were higher in men, and even more so in individuals with impaired glucose tolerance. These EVs, particularly from hypertrophic adipocytes, correlated strongly with insulin resistance and triglyceride levels. Additionally, EVs were associated with β-cell function and metabolic parameters, including lipid and glucose metabolism. The presence of specific markers on some EVs suggests their origin from adipocytes and hepatocytes, reinforcing their potential as metabolic biomarkers and their role in monitoring human metabolic health [213]. Increased CTS-EV quantity additionally correlated with hypertension and increased body mass index (BMI). In another study, patients with obesity (BMI 30+ kg/m²) were found to exhibit a plasma EV concentration approximately ten times higher than individuals of normal weight [214,215,216,217]. These EVs consist of about 20% exosomes and 80% microvesicles. This rise in plasma EV levels is also observed in overweight patients (BMI 25-30 kg/m²) [215]. Notably, subjecting patients to a reduced-calorie diet, a regimen of diet and exercise, or weight reduction following sleeve gastrectomy surgery led to a decline in their plasma EV levels [214,215]. Campello et al found that although there was a decrease in these levels after post-sleeve gastrectomy, they did not match the levels observed in healthy individuals [218].
3.11. Liver Disease
Recently, it has been documented that CTS-EVs regulates liver cells proliferation, differentiation, and metabolism as they carry and transfer several liver-specific proteins and miRNAs [219,220]. CTS-EVs harboring different types of RNAs that can regulate functions in other hepatic stellate cells (HSCs) which are crucial in liver fibrosis in conditions like nonalcoholic fatty liver disease (NAFLD) with their activation being a key factor. Davide and et al explored the role of CTS EVs released by fat-laden hepatocytes in triggering this activation. The CTS EVs, containing miRNAs like miR-128-3p, were found to be internalized by HSCs, leading to their activation, increased pro-fibrogenic gene expression, proliferation, and migration. These effects were primarily due to the suppression of PPAR-γ expression in HSCs by the miRNAs in EVs. These findings suggested that hepatocyte-derived EVs play a significant role in liver fibrosis, offering potential new anti-fibrotic targets for NAFLD and other related diseases [221]. In another study, focusing on insulin resistance, HepG2 liver-derived cells were used to investigate the cleavage of the insulin receptor (IR), highlighting the significant role of exosomes [222]. The research found that in these liver-derived cells, elevated glucose levels accelerate the cleavage of the IR. This process is facilitated by calpain 2, which is secreted into the extracellular space in association with exosomes, leading to the cleavage of the IRβ subunit [222]. This finding was crucial as it links liver metabolism to impaired insulin signaling, emphasizing the role of exosomes in metabolic pathways associated with insulin resistance and the potential impact of treatments like metformin. In hepatocellular carcinoma, a cancer with only a 17% 5 year survival rate, EVs have shown oncogenic behavior by transporting miR-21 and miR-223 which participates in tumor growth and metastasis [219,223,224,225]. Finding specific CTS-EV targets for hepatocellular carcinoma can provide potential targets for diagnosis, prevention, and future treatment.
3.12. Renal Diseases
An increasing amount of research has been done to establish the role of CTS-EVs in kidney physiology and pathology. Chronic kidney disease (CKD) follows a slow, progressive course and with biopsy as the gold standard diagnostic tool to determine etiology. Evaluation of urinary small EVs may serve as a “liquid” biopsy [226,227]. As an example, Barutta et al found urinary extracellular vesicles from patients with diabetic nephropathy showed enrichment of miR-130a and miR-145 and a reduction of miR-155 and miR-424 [228]. Another study found increased levels of miR-200b with a decrease in miR-29c in urinary CTS-EVs isolated from CKD patients compared with healthy individuals [229]. CTS-EVs additionally have therapeutic potential, as pre-clinical studies have utilized MSC-EVs to spur kidney regeneration [226]. Eirin et al treated localized tubular cells a model of renal artery stenosis with MSC-EVs carrying IL-10 and found improved renal inflammation and oxygenation [230]. Renal cell carcinoma (RCC) derived CTS-EVs have also been found to influence tumor microenvironment for promotion of cancer progression [226]. Several markers, including metalloproteinase 9, nephrilysin, ceruloplasmin, podocalyxin, and more have been identified to have different profiles within urinary EVs in RCC compared to healthy patients [231]. This niche tumor microenvironment also provides a crucial step in potential metastasis formation. Long non-coding RNA (lncRNA) have been implicated in particular, with Jin et al finding that RCC CTS-EVs promote cancer cell migration and metastasis by carrying a lncRNA called lung adenocarcinoma transcript 1 (MALAT1), which is also associated with other cancer metastasis [232,233].
3.13. Respiratory Diseases
CTS-EVs have been observed to have roles in many respiratory diseases. In acute lung injury (ARI) and acute respiratory distress syndrome, mouse models have shown an increase of EVs filled with inflammatory molecules that were sent to alveolar macrophages after the initial insult [234,235]. The EV profiles of both infectious and non-infectious lung injury showed an increased in CTS-EVs containing cytokines such as IL-6, TNF-α, TLR2, TLR6, and IL-10 that were dispersed to alveolar macrophages [235]. Detection of these CTS-EVs could help predict likelihood of progression in potential ARDS cases. Asthma is a chronic process featuring chronic inflammation and airway hyperresponsiveness. Lung CTS-EVs containing miRNA, such as miR-145, have been identified as key mediators in smooth muscle function for asthma induced bronchoconstriction [234,236]. Though asthma is considered incurable with current medication, CTS-EVs from MSCs could be used as a potential therapeutic option. These EVs have been found to suppress pro-inflammatory responses and reduce pulmonary fibrosis related remodeling in inflammatory lung diseases animal models, with the added benefit of a respected safety profile [237]. COPD is another chronic inflammatory lung process, resulting in exacerbations that can lead to poor oxygenation and bacterial infection. CTS-EVs have been thought to be implicated in the pathophysiology, with a study finding an increase in exosome levels in COPD patients, with those exosomes being observed to mediate leukotriene conversion in affected tissues [234,238,239,240]. Specifically, increased concentrations of EV-miR-21 were found in bronchial epithelial cells of COPD patients [234,241]. EV-miR-21 is hypothesized to directly control autophagy functions of myofibroblasts, with autophagy dysregulation being involved in COPD progression [241,242]. The specificity of EV-miR-21 creates the possibility of a clinically relevant biomarker for COPD.
3.14. Dermatological Diseases
Recent studies have indicated that EVs play key immunomodulatory roles in skin disorders such as psoriasis, atopic dermatitis, melanoma, and more. Psoriasis is a chronic inflammatory dermatologic disease driven by keratinocytes and infiltration of immune cells [243]. Studies have demonstrated platelet and endothelial cell CTS-EVs were increased in psoriasis patients, and were additionally correlated with disease severity [244,245,246,247]. These CTS-EVs were found to be reduced by anti-TNF-α treatment, a mainstay of severe psoriasis treatment [244,248]. One of the most common skin inflammatory skin disorders is atopic dermatitis (AD), or eczema, and these patients are highly suspectable to bacterial skin infections that in turn drive disease exacerbation [244,249]. Zhou et al observed that S. epidermidis EVs significantly attenuated inflammatory cell inflammation and IgE levels in AD mice models [250]. Others have found that S. aureus EVs could exacerbate AD by carrying mediators that increase expression of inflammatory cytokines such as IL-6 [251]. Thus, these EVs could represent potential therapeutic targets when managing AD exacerbations. Skin cancer is one of the most important diagnoses in the field of dermatology, with squamous cell carcinoma (SCC) of the skin being one such example. Li et al recently reported that cutaneous SCC cells an increased number of EVs compared to normal epidermal keratinocytes [252]. Moreover, tumor xenograft analysis via RNA sequencing revealed that impaired EV-production of cutaneous SCC cells impaired phenotype transformation and inhibited tumor growth [252]. These findings could serve as both a diagnostic biomarker and therapeutic target for SCC management in the future.
3.14. Challenges and Emerging Trends
In the field of using of CTS-EVs as diagnostic tools and to understand their roles in normal physiology, several challenges are intertwined with emerging trends. One of the primary challenges is achieving high specificity and sensitivity in diagnosis, because accurately identifying EVs from specific cells amid a diverse population in body fluids is complex. Additionally, the lack of standardized protocols for EV isolation and analysis introduces variability, complicating their interpretation as biomarkers. A further significant hurdle is the limited understanding of the roles of these EVs in normal physiological processes, making it difficult to differentiate between physiological and pathological EV signatures. Moreover, current technological limitations in detailed EV analysis present challenges, especially given their small size and complex cargo.
However, these challenges are being met with innovative responses. Advanced isolation techniques, such as microfluidics, bead-capture flow cytometry, magnetic based bead absorption strategies, and nanoparticle-based methods, among others, are being developed to improve the purity and yield of specific EVs. The use of high-throughput sequencing and proteomics is enhancing the characterization of EVs, providing deeper insights into their diagnostic potential. Artificial Intelligence and machine learning are increasingly used to analyze complex EV datasets, aiding in the identification of disease-specific diagnostic patterns. The integration of cell type-specific EVs into personalized medicine is gaining traction, promising more tailored diagnostic and prognostic strategies. Additionally, there is a growing focus on functional studies of EVs in normal physiological contexts. Such studies are crucial for distinguishing the roles of CTS-EVs in health versus disease. Together, these advancements are paving the way for the effective utilization of CTS-EVs in both understanding normal physiological processes and developing novel diagnostic approaches.
4. Conclusion
As covered in this review, research in the area of CTS-EVs has demonstrated their great importance in cellular signaling and homeostasis through a variety of physiological systems. CTS-EVs are also implicated in propagating diseased states, such as cancer, neurodegenerative disorders, infectious organisms and more. Thorough identification of the involved CTS-EVs is allowing for better understanding the pathophysiology behind a variety of human diseases. Moreover, these CTS-EVs have promising potential as diagnostic markers, with their unique molecular signatures and carried content that circulate in a variety of body fluids allowing for non-invasive testing. Understanding the specific proliferative or non-proliferative effect EV cargo may have on a disease state additionally opens up the possibility of targeted therapeutics, with EVs themselves serving as fitting non-invasive delivery mechanisms.
Author Contributions
S.A contributed to conception, design, and manuscript writing. EJ contributed to the conception and design of the study, revise the manuscript, and provided the final approval of the manuscript. A.R.D contributed to conception and final approval of manuscript. H.M contributed to image design and manuscript writing. D.T contributed to design and writing of manuscript. A.R, M.C, C.J, and M.Si contributed equally to manuscript writing. M.So, E.K, and A.M contributed equally in manuscript writing. All authors have read and agreed to the published version of the manuscript.
Funding
The project described was supported by the National Center for Advancing Translational Sciences, National Institutes of Health, through Grant KL2TR002002. The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIH. R01 EY024349 (ARD); unrestricted grant to the Department and Physician–Scientist Award both from Research to Prevent Blindness and UG3/UH3 EY031809 from NEI; Department of Defense, under grant VR170180.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors want to thank Lauren Kalinoski for revising the schematic figure.
Conflicts of Interest
The authors declare no competing financial or nonfinancial conflict of interest.
References
- Massoumi, H.; Amin, S.; Soleimani, M.; Momenaei, B.; Ashraf, M.J.; Guaiquil, V.H.; Hematti, P.; Rosenblatt, M.I.; Djalilian, A.R.; Jalilian, E. Extracellular-Vesicle-Based Therapeutics in Neuro-Ophthalmic Disorders. Int. J. Mol. Sci. 2023, 24. [CrossRef]
- Murphy, D.E.; de Jong, O.G.; Brouwer, M.; Wood, M.J.; Lavieu, G.; Schiffelers, R.M.; Vader, P. Extracellular Vesicle-Based Therapeutics: Natural versus Engineered Targeting and Trafficking. Exp. Mol. Med. 2019, 51, 1–12. [CrossRef]
- Jalilian, E.; Bigit, B.; Amin, S.; Massoumi, H.; Katz, E.; Guaiquil, V.H.; Rosenblatt, M.; Djalilian, A.R. Exosomes from 3D Cultures of Bone Marrow Mesenchymal Stem Cell (BM-MSCs) Have Higher Neuro-Regenerative Potential than Those Generated from 2D Conditions. Invest. Ophthalmol. Vis. Sci. 2022, 63, 1209-A0209-1209-A0209.
- Jalilian, E.; Massoumi, H.; Bigit, B.; Amin, S.; Katz, E.A.; Guaiquil, V.H.; Anwar, K.N.; Hematti, P.; Rosenblatt, M.I.; Djalilian, A.R. Bone Marrow Mesenchymal Stromal Cells in a 3D System Produce Higher Concentration of Extracellular Vesicles (EVs) with Increased Complexity and Enhanced Neuronal Growth Properties. Stem Cell Res. Ther. 2022, 13, 425. [CrossRef]
- Amin, S.; Jalilian, E.; Katz, E.; Frank, C.; Yazdanpanah, G.; Guaiquil, V.H.; Rosenblatt, M.I.; Djalilian, A.R. The Limbal Niche and Regenerative Strategies. Vision 2021, 5, 43. [CrossRef]
- Jalilian, E.; Putra, I.; Katz, E.; Yazdanpanah, G.; Guaiquil, V.H.; Shen, X.; Anwar, K.; An, S.; Rosenblatt, M.; Djalilian, A.R. Interactions between Mesenchymal Stem Cells (MSCs) and Trigeminal Ganglion (TGs) Improve Neurite Growth and Elongation. Invest. Ophthalmol. Vis. Sci. 2021, 62, 892.
- Massoumi, H.; Katz, E.; Nguyen, T.T.; Zhou, Q.; Jazayerli, C.; Anwar, K.; Ashraf, M.; Soleimani, M.; Guaiquil, V.H.; Rosenblatt, M.; et al. Extracellular Vesicles (EVs) from 3D Cultured Human Bone Marrow Mesenchymal Stem Cells (hBM-MSC) Demonstrated Increased Complexity and Neurite Elongation in an in-Vivo Corneal Injury Model. Invest. Ophthalmol. Vis. Sci. 2023, 64, 2350.
- Carnino, J.M.; Lee, H.; Jin, Y. Isolation and Characterization of Extracellular Vesicles from Broncho-Alveolar Lavage Fluid: A Review and Comparison of Different Methods. Respir Res 2019, 20, 240. [CrossRef]
- Thery, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J Extracell Vesicles 2018, 7, 153575. [CrossRef]
- Consortium, E.-T. EV-TRACK: Transparent Reporting and Centralizing Knowledge in Extracellular Vesicle Research. Nat Methods 2017, 14, 228–232. [CrossRef]
- Boukouris, S.; Mathivanan, S. Exosomes in Bodily Fluids Are a Highly Stable Resource of Disease Biomarkers. Proteomics Clin. Appl. 2015, 9, 358–367. [CrossRef]
- Xu, R.; Rai, A.; Chen, M.; Suwakulsiri, W.; Greening, D.W.; Simpson, R.J. Extracellular Vesicles in Cancer — Implications for Future Improvements in Cancer Care. Nat. Rev. Clin. Oncol. 2018, 15, 617–638. [CrossRef]
- Akbar, A.; Malekian, F.; Baghban, N.; Kodam, S.P.; Ullah, M. Methodologies to Isolate and Purify Clinical Grade Extracellular Vesicles for Medical Applications. Cells 2022, 11, 186. [CrossRef]
- Mesenchymal Stem Cell-Derived Extracellular Vesicles for Immunomodulation and Regeneration: A next Generation Therapeutic Tool? | Cell Death & Disease Available online: https://www.nature.com/articles/s41419-022-05034-x (accessed on 26 January 2024).
- Fuloria, S.; Subramaniyan, V.; Dahiya, R.; Dahiya, S.; Sudhakar, K.; Kumari, U.; Sathasivam, K.; Meenakshi, D.U.; Wu, Y.S.; Sekar, M.; et al. Mesenchymal Stem Cell-Derived Extracellular Vesicles: Regenerative Potential and Challenges. Biology 2021, 10, 172. [CrossRef]
- Mesenchymal Stromal Cell Exosomes in Cardiac Repair - PMC Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC9885380/ (accessed on 30 January 2024).
- Zargar, M.J.; Kaviani, S.; Vasei, M.; Soufi Zomorrod, M.; Heidari Keshel, S.; Soleimani, M. Therapeutic Role of Mesenchymal Stem Cell-Derived Exosomes in Respiratory Disease. Stem Cell Res. Ther. 2022, 13, 194. [CrossRef]
- Yang, M.; Chen, J.; Chen, L. The Roles of Mesenchymal Stem Cell-Derived Exosomes in Diabetes Mellitus and Its Related Complications. Front. Endocrinol. 2022, 13, 1027686. [CrossRef]
- Wu, K.Y.; Ahmad, H.; Lin, G.; Carbonneau, M.; Tran, S.D. Mesenchymal Stem Cell-Derived Exosomes in Ophthalmology: A Comprehensive Review. Pharmaceutics 2023, 15, 1167. [CrossRef]
- Samsonraj, R.M.; Raghunath, M.; Nurcombe, V.; Hui, J.H.; van Wijnen, A.J.; Cool, S.M. Concise Review: Multifaceted Characterization of Human Mesenchymal Stem Cells for Use in Regenerative Medicine. Stem Cells Transl. Med. 2017, 6, 2173–2185. [CrossRef]
- Mesenchymal Stem Cell-Derived Exosomes in Cancer Therapy Resistance: Recent Advances and Therapeutic Potential | Molecular Cancer | Full Text Available online: https://molecular-cancer.biomedcentral.com/articles/10.1186/s12943-022-01650-5 (accessed on 30 January 2024).
- Fafián-Labora, J.A.; O’Loghlen, A. Classical and Nonclassical Intercellular Communication in Senescence and Ageing. Trends Cell Biol. 2020, 30, 628–639. [CrossRef]
- Kosaka, N.; Kogure, A.; Yamamoto, T.; Urabe, F.; Usuba, W.; Prieto-Vila, M.; Ochiya, T. Exploiting the Message from Cancer: The Diagnostic Value of Extracellular Vesicles for Clinical Applications. Exp. Mol. Med. 2019, 51, 1–9. [CrossRef]
- Liu, W.; Jin, M.; Chen, Q.; Li, Q.; Xing, X.; Luo, Y.; Sun, X. Insight into Extracellular Vesicles in Vascular Diseases: Intercellular Communication Role and Clinical Application Potential. Cell Commun. Signal. 2023, 21, 310. [CrossRef]
- Abels, E.R.; Breakefield, X.O. Introduction to Extracellular Vesicles: Biogenesis, RNA Cargo Selection, Content, Release, and Uptake. Cell. Mol. Neurobiol. 2016, 36, 301–312. [CrossRef]
- Hristov, M.; Erl, W.; Linder, S.; Weber, P.C. Apoptotic Bodies from Endothelial Cells Enhance the Number and Initiate the Differentiation of Human Endothelial Progenitor Cells in Vitro. Blood 2004, 104, 2761–2766. [CrossRef]
- Kerr, J.F.; Wyllie, A.H.; Currie, A.R. Apoptosis: A Basic Biological Phenomenon with Wide-Ranging Implications in Tissue Kinetics. Br. J. Cancer 1972, 26, 239–257. [CrossRef]
- Akers, J.C.; Gonda, D.; Kim, R.; Carter, B.S.; Chen, C.C. Biogenesis of Extracellular Vesicles (EV): Exosomes, Microvesicles, Retrovirus-like Vesicles, and Apoptotic Bodies. J. Neurooncol. 2013, 113, 1–11. [CrossRef]
- Gurung, S.; Perocheau, D.; Touramanidou, L.; Baruteau, J. The Exosome Journey: From Biogenesis to Uptake and Intracellular Signalling. Cell Commun. Signal. 2021, 19, 47. [CrossRef]
- Pols, M.S.; Klumperman, J. Trafficking and Function of the Tetraspanin CD63. Exp. Cell Res. 2009, 315, 1584–1592. [CrossRef]
- Molecular Mechanism of Multivesicular Body Biogenesis by ESCRT Complexes - PubMed Available online: https://pubmed.ncbi.nlm.nih.gov/20305637/ (accessed on 20 January 2024).
- Morishita, M.; Takahashi, Y.; Nishikawa, M.; Takakura, Y. Pharmacokinetics of Exosomes—An Important Factor for Elucidating the Biological Roles of Exosomes and for the Development of Exosome-Based Therapeutics. J. Pharm. Sci. 2017, 106, 2265–2269. [CrossRef]
- Kim, D.-K.; Kang, B.; Kim, O.Y.; Choi, D.-S.; Lee, J.; Kim, S.R.; Go, G.; Yoon, Y.J.; Kim, J.H.; Jang, S.C.; et al. EVpedia: An Integrated Database of High-Throughput Data for Systemic Analyses of Extracellular Vesicles. J. Extracell. Vesicles 2013, 2. [CrossRef]
- Zhang, Y.; Liu, Y.; Liu, H.; Tang, W.H. Exosomes: Biogenesis, Biologic Function and Clinical Potential. Cell Biosci. 2019, 9, 19. [CrossRef]
- Waldenström, A.; Ronquist, G. Role of Exosomes in Myocardial Remodeling. Circ. Res. 2014, 114, 315–324. [CrossRef]
- Li, Y.; Tan, J.; Miao, Y.; Zhang, Q. MicroRNA in Extracellular Vesicles Regulates Inflammation through Macrophages under Hypoxia. Cell Death Discov. 2021, 7, 1–12. [CrossRef]
- Vlassov, A.V.; Magdaleno, S.; Setterquist, R.; Conrad, R. Exosomes: Current Knowledge of Their Composition, Biological Functions, and Diagnostic and Therapeutic Potentials. Biochim. Biophys. Acta BBA - Gen. Subj. 2012, 1820, 940–948. [CrossRef]
- Mashouri, L.; Yousefi, H.; Aref, A.R.; Ahadi, A. mohammad; Molaei, F.; Alahari, S.K. Exosomes: Composition, Biogenesis, and Mechanisms in Cancer Metastasis and Drug Resistance. Mol. Cancer 2019, 18, 75. [CrossRef]
- G, N.; Gundogan, B.; Tan, A.; Farhatnia, Y.; Wu, W.; Rajadas, J.; Seifalian, A.M. Exosomes as Immunotheranostic Nanoparticles. Clin. Ther. 2014, 36, 820–829. [CrossRef]
- Skotland, T.; Hessvik, N.P.; Sandvig, K.; Llorente, A. Exosomal Lipid Composition and the Role of Ether Lipids and Phosphoinositides in Exosome Biology. J. Lipid Res. 2019, 60, 9–18. [CrossRef]
- Sunshine, H.; Iruela-Arispe, M.L. Membrane Lipids and Cell Signaling. Curr. Opin. Lipidol. 2017, 28, 408–413. [CrossRef]
- Subra, C.; Grand, D.; Laulagnier, K.; Stella, A.; Lambeau, G.; Paillasse, M.; Medina, P.D.; Monsarrat, B.; Perret, B.; Silvente-Poirot, S.; et al. Exosomes Account for Vesicle-Mediated Transcellular Transport of Activatable Phospholipases and Prostaglandins [S]. J. Lipid Res. 2010, 51, 2105–2120. [CrossRef]
- Williams, C.; Royo, F.; Aizpurua-Olaizola, O.; Pazos, R.; Boons, G.-J.; Reichardt, N.-C.; Falcon-Perez, J.M. Glycosylation of Extracellular Vesicles: Current Knowledge, Tools and Clinical Perspectives. J. Extracell. Vesicles 2018, 7, 1442985. [CrossRef]
- Yokose, T.; Kabe, Y.; Matsuda, A.; Kitago, M.; Matsuda, S.; Hirai, M.; Nakagawa, T.; Masugi, Y.; Hishiki, T.; Nakamura, Y.; et al. O-Glycan-Altered Extracellular Vesicles: A Specific Serum Marker Elevated in Pancreatic Cancer. Cancers 2020, 12, 2469. [CrossRef]
- Chernomordik, L.V.; Melikyan, G.B.; Chizmadzhev, Y.A. Biomembrane Fusion: A New Concept Derived from Model Studies Using Two Interacting Planar Lipid Bilayers. Biochim. Biophys. Acta BBA - Rev. Biomembr. 1987, 906, 309–352. [CrossRef]
- Hu, Y.-B.; Dammer, E.B.; Ren, R.-J.; Wang, G. The Endosomal-Lysosomal System: From Acidification and Cargo Sorting to Neurodegeneration. Transl. Neurodegener. 2015, 4, 18. [CrossRef]
- Raposo, G.; Stoorvogel, W. Extracellular Vesicles: Exosomes, Microvesicles, and Friends. J. Cell Biol. 2013, 200, 373–383. [CrossRef]
- Zhou, X.; Xie, F.; Wang, L.; Zhang, L.; Zhang, S.; Fang, M.; Zhou, F. The Function and Clinical Application of Extracellular Vesicles in Innate Immune Regulation. Cell. Mol. Immunol. 2020, 17, 323–334. [CrossRef]
- Saliba, D.G.; Céspedes-Donoso, P.F.; Bálint, Š.; Compeer, E.B.; Korobchevskaya, K.; Valvo, S.; Mayya, V.; Kvalvaag, A.; Peng, Y.; Dong, T.; et al. Composition and Structure of Synaptic Ectosomes Exporting Antigen Receptor Linked to Functional CD40 Ligand from Helper T Cells. eLife 2019, 8, e47528. [CrossRef]
- Buzas, E.I. The Roles of Extracellular Vesicles in the Immune System. Nat Rev Immunol 2023, 23, 236–250. [CrossRef]
- Boilard, E. Thematic Review Series: Exosomes and Microvesicles: Lipids as Key Components of Their Biogenesis and Functions Extracellular Vesicles and Their Content in Bioactive Lipid Mediators: More than a Sack of microRNA. J. Lipid Res. 2018, 59, 2037–2046. [CrossRef]
- Skogberg, G.; Lundberg, V.; Berglund, M.; Gudmundsdottir, J.; Telemo, E.; Lindgren, S.; Ekwall, O. Human Thymic Epithelial Primary Cells Produce Exosomes Carrying Tissue-restricted Antigens. Immunol. Cell Biol. 2015, 93, 727–734. [CrossRef]
- Zhou, X.; Brown, B.A.; Siegel, A.P.; El Masry, M.S.; Zeng, X.; Song, W.; Das, A.; Khandelwal, P.; Clark, A.; Singh, K.; et al. Exosome-Mediated Crosstalk between Keratinocytes and Macrophages in Cutaneous Wound Healing. ACS Nano 2020, 14, 12732–12748. [CrossRef]
- Guo, S.; DiPietro, L.A. Factors Affecting Wound Healing. J. Dent. Res. 2010, 89, 219–229. [CrossRef]
- Han, X.; Wu, P.; Li, L.; Sahal, H.M.; Ji, C.; Zhang, J.; Wang, Y.; Wang, Q.; Qian, H.; Shi, H.; et al. Exosomes Derived from Autologous Dermal Fibroblasts Promote Diabetic Cutaneous Wound Healing through the Akt/β-Catenin Pathway. Cell Cycle 2021, 20, 616–622. [CrossRef]
- Glady, A.; Vandebroek, A.; Yasui, M. Human Keratinocyte-Derived Extracellular Vesicles Activate the MAPKinase Pathway and Promote Cell Migration and Proliferation in Vitro. Inflamm. Regen. 2021, 41, 4. [CrossRef]
- Guo, S.-C.; Tao, S.-C.; Yin, W.-J.; Qi, X.; Yuan, T.; Zhang, C.-Q. Exosomes Derived from Platelet-Rich Plasma Promote the Re-Epithelization of Chronic Cutaneous Wounds via Activation of YAP in a Diabetic Rat Model. Theranostics 2017, 7, 81–96. [CrossRef]
- Xu, Y.; Lin, Z.; He, L.; Qu, Y.; Liu, O.; Han, Y.; Xu, C.; Duan, D. Platelet Rich Plasma-Derived Exosomal USP15 Promotes Cutaneous Wound Healing via Deubiquitinating EIF4A1; In Review, 2021.
- Schnatz, A.; Müller, C.; Brahmer, A.; Krämer-Albers, E. Extracellular Vesicles in Neural Cell Interaction and CNS Homeostasis. FASEB BioAdvances 2021, 3, 577–592. [CrossRef]
- Holm, M.M.; Kaiser, J.; Schwab, M.E. Extracellular Vesicles: Multimodal Envoys in Neural Maintenance and Repair. Trends Neurosci. 2018, 41, 360–372. [CrossRef]
- Ma, Y.; Li, C.; Huang, Y.; Wang, Y.; Xia, X.; Zheng, J.C. Exosomes Released from Neural Progenitor Cells and Induced Neural Progenitor Cells Regulate Neurogenesis through miR-21a. Cell Commun. Signal. 2019, 17, 96. [CrossRef]
- Zappulli, V.; Friis, K.P.; Fitzpatrick, Z.; Maguire, C.A.; Breakefield, X.O. Extracellular Vesicles and Intercellular Communication within the Nervous System. J. Clin. Invest. 2016, 126, 1198–1207. [CrossRef]
- Herrera Lopez, M.; Bertone Arolfo, M.; Remedi, M.; Gastaldi, L.; Wilson, C.; Guendulain, G.; Ceschin, D.; Cardozo Gizzi, A.; Caceres, A.; Moyano, A.L. Human Neural Rosettes Secrete Bioactive Extracellular Vesicles Enriched in Neuronal and Glial Cellular Component 2024.
- Jalilian, E.; Shin, S.R. Novel Model of Cortical-Meningeal Organoid Co-Culture System Improves Human Cortical Brain Organoid Cytoarchitecture. Sci. Rep. 2023, 13, 7809. [CrossRef]
- Cruz, L.; Romero, J.A.A.; Iglesia, R.P.; Lopes, M.H. Extracellular Vesicles: Decoding a New Language for Cellular Communication in Early Embryonic Development. Front. Cell Dev. Biol. 2018, 6, 94. [CrossRef]
- Huang-Doran, I.; Zhang, C.-Y.; Vidal-Puig, A. Extracellular Vesicles: Novel Mediators of Cell Communication In Metabolic Disease. Trends Endocrinol. Metab. 2017, 28, 3–18. [CrossRef]
- Kranendonk, M.E.G.; Visseren, F.L.J.; Van Herwaarden, J.A.; Nolte-’t Hoen, E.N.M.; De Jager, W.; Wauben, M.H.M.; Kalkhoven, E. Effect of Extracellular Vesicles of Human Adipose Tissue on Insulin Signaling in Liver and Muscle Cells: Adipose Tissue Vesicles and Insulin Signaling. Obesity 2014, 22, 2216–2223. [CrossRef]
- Kranendonk, M.E.G.; Visseren, F.L.J.; Van Balkom, B.W.M.; Nolte-’t Hoen, E.N.M.; Van Herwaarden, J.A.; De Jager, W.; Schipper, H.S.; Brenkman, A.B.; Verhaar, M.C.; Wauben, M.H.M.; et al. Human Adipocyte Extracellular Vesicles in Reciprocal Signaling between Adipocytes and Macrophages. Obesity 2014, 22, 1296–1308. [CrossRef]
- Gesmundo, I.; Pardini, B.; Gargantini, E.; Gamba, G.; Birolo, G.; Fanciulli, A.; Banfi, D.; Congiusta, N.; Favaro, E.; Deregibus, M.C.; et al. Adipocyte-Derived Extracellular Vesicles Regulate Survival and Function of Pancreatic β Cells. JCI Insight 2021, 6, e141962. [CrossRef]
- Zaborowski, M.P.; Balaj, L.; Breakefield, X.O.; Lai, C.P. Extracellular Vesicles: Composition, Biological Relevance, and Methods of Study. BioScience 2015, 65, 783–797. [CrossRef]
- Liu, W.; Liu, T.; Zhao, Q.; Ma, J.; Jiang, J.; Shi, H. Adipose Tissue-Derived Extracellular Vesicles: A Promising Biomarker and Therapeutic Strategy for Metabolic Disorders. Stem Cells Int. 2023, 2023, 1–16. [CrossRef]
- Bylicky, M.A.; Mueller, G.P.; Day, R.M. Mechanisms of Endogenous Neuroprotective Effects of Astrocytes in Brain Injury. Oxid. Med. Cell. Longev. 2018, 2018, 6501031. [CrossRef]
- Bahram Sangani, N.; Gomes, A.R.; Curfs, L.M.G.; Reutelingsperger, C.P. The Role of Extracellular Vesicles during CNS Development. Prog. Neurobiol. 2021, 205, 102124. [CrossRef]
- Kaminska, A.; Radoszkiewicz, K.; Rybkowska, P.; Wedzinska, A.; Sarnowska, A. Interaction of Neural Stem Cells (NSCs) and Mesenchymal Stem Cells (MSCs) as a Promising Approach in Brain Study and Nerve Regeneration. Cells 2022, 11, 1464. [CrossRef]
- Oncogenic H-Ras Reprograms Madin-Darby Canine Kidney (MDCK) Cell-Derived Exosomal Proteins Following Epithelial-Mesenchymal Transition* - Molecular & Cellular Proteomics Available online: https://www.mcponline.org/article/S1535-9476(20)32528-7/fulltext (accessed on 18 January 2024).
- Hu, T.; Hu, J. Melanoma-Derived Exosomes Induce Reprogramming Fibroblasts into Cancer-Associated Fibroblasts via Gm26809 Delivery. Cell Cycle 2019, 18, 3085–3094. [CrossRef]
- Zhou, H.; Zhu, L.; Song, J.; Wang, G.; Li, P.; Li, W.; Luo, P.; Sun, X.; Wu, J.; Liu, Y.; et al. Liquid Biopsy at the Frontier of Detection, Prognosis and Progression Monitoring in Colorectal Cancer. Mol. Cancer 2022, 21, 86. [CrossRef]
- Clayton, A.; Mitchell, J.P.; Court, J.; Linnane, S.; Mason, M.D.; Tabi, Z. Human Tumor-Derived Exosomes Down-Modulate NKG2D Expression1. J. Immunol. 2008, 180, 7249–7258. [CrossRef]
- Guo, S.; Huang, J.; Li, G.; Chen, W.; Li, Z.; Lei, J. The Role of Extracellular Vesicles in Circulating Tumor Cell-Mediated Distant Metastasis. Mol. Cancer 2023, 22. [CrossRef]
- Brown, T.J.; James, V. The Role of Extracellular Vesicles in the Development of a Cancer Stem Cell Microenvironment Niche and Potential Therapeutic Targets: A Systematic Review. Cancers 2021, 13, 2435. [CrossRef]
- Wang, L.; He, J.; Hu, H.; Tu, L.; Sun, Z.; Liu, Y.; Luo, F. Lung CSC-derived Exosomal miR-210-3p Contributes to a Pro-metastatic Phenotype in Lung Cancer by Targeting FGFRL1. J. Cell. Mol. Med. 2020, 24, 6324–6339. [CrossRef]
- Irmer, B.; Chandrabalan, S.; Maas, L.; Bleckmann, A.; Menck, K. Extracellular Vesicles in Liquid Biopsies as Biomarkers for Solid Tumors. Cancers 2023, 15, 1307. [CrossRef]
- Alegre, E.; Zubiri, L.; Perez-Gracia, J.L.; González-Cao, M.; Soria, L.; Martín-Algarra, S.; González, A. Circulating Melanoma Exosomes as Diagnostic and Prognosis Biomarkers. Clin. Chim. Acta Int. J. Clin. Chem. 2016, 454, 28–32. [CrossRef]
- Osti, D.; Del Bene, M.; Rappa, G.; Santos, M.; Matafora, V.; Richichi, C.; Faletti, S.; Beznoussenko, G.V.; Mironov, A.; Bachi, A.; et al. Clinical Significance of Extracellular Vesicles in Plasma from Glioblastoma Patients. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2019, 25, 266–276. [CrossRef]
- Duijvesz, D.; Versluis, C.Y.L.; van der Fels, C.A.M.; Vredenbregt-van den Berg, M.S.; Leivo, J.; Peltola, M.T.; Bangma, C.H.; Pettersson, K.S.I.; Jenster, G. Immuno-Based Detection of Extracellular Vesicles in Urine as Diagnostic Marker for Prostate Cancer. Int. J. Cancer 2015, 137, 2869–2878. [CrossRef]
- König, L.; Kasimir-Bauer, S.; Bittner, A.-K.; Hoffmann, O.; Wagner, B.; Santos Manvailer, L.F.; Kimmig, R.; Horn, P.A.; Rebmann, V. Elevated Levels of Extracellular Vesicles Are Associated with Therapy Failure and Disease Progression in Breast Cancer Patients Undergoing Neoadjuvant Chemotherapy. Oncoimmunology 2017, 7, e1376153. [CrossRef]
- Ghoroghi, S.; Mary, B.; Asokan, N.; Goetz, J.G.; Hyenne, V. Tumor Extracellular Vesicles Drive Metastasis (It’s a Long Way from Home). FASEB BioAdvances 2021, 3, 930–943. [CrossRef]
- Ramirez-Garrastacho, M.; Bajo-Santos, C.; Line, A.; Martens-Uzunova, E.S.; de la Fuente, J.M.; Moros, M.; Soekmadji, C.; Tasken, K.A.; Llorente, A. Extracellular Vesicles as a Source of Prostate Cancer Biomarkers in Liquid Biopsies: A Decade of Research. Br. J. Cancer 2022, 126, 331–350. [CrossRef]
- Chen, L. Use of a Liquid Biopsy Signature as Blood Biomarker for Early Detection and Monitoring Early-Onset Gastric Cancer; clinicaltrials.gov, 2023.
- Moon, P.-G.; Lee, J.-E.; Cho, Y.-E.; Lee, S.J.; Chae, Y.S.; Jung, J.H.; Kim, I.-S.; Park, H.Y.; Baek, M.-C. Fibronectin on Circulating Extracellular Vesicles as a Liquid Biopsy to Detect Breast Cancer. Oncotarget 2016, 7, 40189–40199. [CrossRef]
- Zhang, X.; Wang, C.; Yu, J.; Bu, J.; Ai, F.; Wang, Y.; Lin, J.; Zhu, X. Extracellular Vesicles in the Treatment and Diagnosis of Breast Cancer: A Status Update. Front. Endocrinol. 2023, 14, 1202493. [CrossRef]
- Garofalo, M.; Villa, A.; Rizzi, N.; Kuryk, L.; Rinner, B.; Cerullo, V.; Yliperttula, M.; Mazzaferro, V.; Ciana, P. Extracellular Vesicles Enhance the Targeted Delivery of Immunogenic Oncolytic Adenovirus and Paclitaxel in Immunocompetent Mice. J. Control. Release Off. J. Control. Release Soc. 2019, 294, 165–175. [CrossRef]
- Wu, M.; Wang, M.; Jia, H.; Wu, P. Extracellular Vesicles: Emerging Anti-Cancer Drugs and Advanced Functionalization Platforms for Cancer Therapy. Drug Deliv. 29, 2513–253. [CrossRef]
- Zhu, X.; Gao, M.; Yang, Y.; Li, W.; Bao, J.; Li, Y. The CRISPR/Cas9 System Delivered by Extracellular Vesicles. Pharmaceutics 2023, 15, 984. [CrossRef]
- Wu, M.; Wang, M.; Jia, H.; Wu, P. Extracellular Vesicles: Emerging Anti-Cancer Drugs and Advanced Functionalization Platforms for Cancer Therapy. Drug Deliv. 2022, 29, 2513–2538. [CrossRef]
- Chen, L.; Su, D.; Li, S.; Guan, L.; Shi, C.; Li, D.; Hu, S.; Ma, X. The Connexin 46 Mutant (V44M) Impairs Gap Junction Function Causing Congenital Cataract. J. Genet. 2017, 96, 969–976. [CrossRef]
- Acuña, R.A.; Varas-Godoy, M.; Berthoud, V.M.; Alfaro, I.E.; Retamal, M.A. Connexin-46 Contained in Extracellular Vesicles Enhance Malignancy Features in Breast Cancer Cells. Biomolecules 2020, 10, 676. [CrossRef]
- Faldoni, F.L.C.; Villacis, R.A.R.; Canto, L.M.; Fonseca-Alves, C.E.; Cury, S.S.; Larsen, S.J.; Aagaard, M.M.; Souza, C.P.; Scapulatempo-Neto, C.; Osório, C.A.B.T.; et al. Inflammatory Breast Cancer: Clinical Implications of Genomic Alterations and Mutational Profiling. Cancers 2020, 12, 2816. [CrossRef]
- Lee, Y.; Ni, J.; Beretov, J.; Wasinger, V.C.; Graham, P.; Li, Y. Recent Advances of Small Extracellular Vesicle Biomarkers in Breast Cancer Diagnosis and Prognosis. Mol. Cancer 2023, 22, 33. [CrossRef]
- Moon, P.-G.; Lee, J.-E.; Cho, Y.-E.; Lee, S.J.; Jung, J.H.; Chae, Y.S.; Bae, H.-I.; Kim, Y.-B.; Kim, I.-S.; Park, H.Y.; et al. Identification of Developmental Endothelial Locus-1 on Circulating Extracellular Vesicles as a Novel Biomarker for Early Breast Cancer Detection. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2016, 22, 1757–1766. [CrossRef]
- Eubanks, S.E. A Pilot Study of Tumor-Derived Exosomes as Diagnostic and Prognostic Markers in Breast Cancer Patients Receiving Neoadjuvant Chemotherapy; clinicaltrials.gov, 2017.
- Armstrong, J.P.; Holme, M.N.; Stevens, M.M. Re-Engineering Extracellular Vesicles as Smart Nanoscale Therapeutics. ACS Nano 2017, 11, 69–83. [CrossRef]
- Müller, S.; Janke, F.; Dietz, S.; Sültmann, H. Circulating MicroRNAs as Potential Biomarkers for Lung Cancer. Recent Results Cancer Res. Fortschritte Krebsforsch. Progres Dans Rech. Sur Cancer 2020, 215, 299–318. [CrossRef]
- 104. Sun, J. Clinical Study of ctDNA and Exosome Combined Detection to Identify Benign and Malignant Pulmonary Nodules; clinicaltrials.gov, 2019.
- Li, X.-Q.; Liu, J.-T.; Fan, L.-L.; Liu, Y.; Cheng, L.; Wang, F.; Yu, H.-Q.; Gao, J.; Wei, W.; Wang, H.; et al. Exosomes Derived from Gefitinib-Treated EGFR-Mutant Lung Cancer Cells Alter Cisplatin Sensitivity via up-Regulating Autophagy. Oncotarget 2016, 7, 24585–24595. [CrossRef]
- Saviana, M.; Romano, G.; Le, P.; Acunzo, M.; Nana-Sinkam, P. Extracellular Vesicles in Lung Cancer Metastasis and Their Clinical Applications. Cancers 2021, 13, 5633. [CrossRef]
- Urine Exosomes for Non-Invasive Assessment of Gene Expression and Mutations of Prostate Cancer | PLOS ONE Available online: https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0154507 (accessed on 18 January 2024).
- Del Re, M.; Crucitta, S.; Sbrana, A.; Rofi, E.; Paolieri, F.; Gianfilippo, G.; Galli, L.; Falcone, A.; Morganti, R.; Porta, C.; et al. Androgen Receptor (AR) Splice Variant 7 and Full-Length AR Expression Is Associated with Clinical Outcome: A Translational Study in Patients with Castrate-Resistant Prostate Cancer. BJU Int. 2019, 124, 693–700. [CrossRef]
- Yuan, T.; Shi, C.; Xu, W.; Yang, H.-L.; Xia, B.; Tian, C. Extracellular Vesicles Derived from T-Cell Acute Lymphoblastic Leukemia Inhibit Osteogenic Differentiation of Bone Marrow Mesenchymal Stem Cells via miR-34a-5p. Endocr. J. 2021, 68, 1197–1208. [CrossRef]
- Izadirad, M.; Huang, Z.; Jafari, F.; Hamidieh, A.A.; Gharehbaghian, A.; Li, Y.-D.; Jafari, L.; Chen, Z.-S. Extracellular Vesicles in Acute Leukemia: A Mesmerizing Journey With a Focus on Transferred microRNAs. Front. Cell Dev. Biol. 2021, 9, 766371. [CrossRef]
- Hornick, N.I.; Huan, J.; Doron, B.; Goloviznina, N.A.; Lapidus, J.; Chang, B.H.; Kurre, P. Serum Exosome MicroRNA as a Minimally-Invasive Early Biomarker of AML. Sci. Rep. 2015, 5, 11295. [CrossRef]
- Ruiss, R.; Jochum, S.; Mocikat, R.; Hammerschmidt, W.; Zeidler, R. EBV-Gp350 Confers B-Cell Tropism to Tailored Exosomes and Is a Neo-Antigen in Normal and Malignant B Cells—A New Option for the Treatment of B-CLL. PLoS ONE 2011, 6, e25294. [CrossRef]
- Gärtner, K.; Luckner, M.; Wanner, G.; Zeidler, R. Engineering Extracellular Vesicles as Novel Treatment Options: Exploiting Herpesviral Immunity in CLL. J. Extracell. Vesicles 2019, 8, 1573051. [CrossRef]
- Dubois, K.; Tannoury, M.; Bauvois, B.; Susin, S.A.; Garnier, D. Extracellular Vesicles in Chronic Lymphocytic Leukemia: Tumor Microenvironment Messengers as a Basis for New Targeted Therapies? Cancers 2023, 15, 2307. [CrossRef]
- Runz, S.; Keller, S.; Rupp, C.; Stoeck, A.; Issa, Y.; Koensgen, D.; Mustea, A.; Sehouli, J.; Kristiansen, G.; Altevogt, P. Malignant Ascites-Derived Exosomes of Ovarian Carcinoma Patients Contain CD24 and EpCAM. Gynecol. Oncol. 2007, 107, 563–571. [CrossRef]
- Keserű, J.S.; Soltész, B.; Lukács, J.; Márton, É.; Szilágyi-Bónizs, M.; Penyige, A.; Póka, R.; Nagy, B. Detection of Cell-Free, Exosomal and Whole Blood Mitochondrial DNA Copy Number in Plasma or Whole Blood of Patients with Serous Epithelial Ovarian Cancer. J. Biotechnol. 2019, 298, 76–81. [CrossRef]
- McAlarnen, L.A.; Gupta, P.; Singh, R.; Pradeep, S.; Chaluvally-Raghavan, P. Extracellular Vesicle Contents as Non-Invasive Biomarkers in Ovarian Malignancies. Mol. Ther. Oncolytics 2022, 26, 347–359. [CrossRef]
- Pisano, S.; Pierini, I.; Gu, J.; Gazze, A.; Francis, L.W.; Gonzalez, D.; Conlan, R.S.; Corradetti, B. Immune (Cell) Derived Exosome Mimetics (IDEM) as a Treatment for Ovarian Cancer. Front. Cell Dev. Biol. 2020, 8, 553576. [CrossRef]
- Hallal, S.M.; Tűzesi, Á.; Sida, L.A.; Xian, E.; Madani, D.; Muralidharan, K.; Shivalingam, B.; Buckland, M.E.; Satgunaseelan, L.; Alexander, K.L. Glioblastoma Biomarkers in Urinary Extracellular Vesicles Reveal the Potential for a ‘Liquid Gold’ Biopsy. Br. J. Cancer 2024. [CrossRef]
- University Hospital, Limoges Clinical Relevance of Detecting Molecular Abnormalities in Glial Tumor Exosomes; clinicaltrials.gov, 2023.
- Lovinger, D.M. Communication Networks in the Brain. Alcohol Res. Health 2008, 31, 196–214.
- Cheng, L.; Hill, A.F. Therapeutically Harnessing Extracellular Vesicles. Nat. Rev. Drug Discov. 2022, 21, 379–399. [CrossRef]
- Pérez, M.; Avila, J.; Hernández, F. Propagation of Tau via Extracellular Vesicles. Front. Neurosci. 2019, 13. [CrossRef]
- Gomes, P.; Tzouanou, F.; Skolariki, K.; Vamvaka-Iakovou, A.; Noguera-Ortiz, C.; Tsirtsaki, K.; Waites, C.L.; Vlamos, P.; Sousa, N.; Costa-Silva, B.; et al. Extracellular Vesicles and Alzheimer’s Disease in the Novel Era of Precision Medicine: Implications for Disease Progression, Diagnosis and Treatment. Exp. Neurol. 2022, 358, 114183. [CrossRef]
- Si, Q.; Wu, L.; Pang, D.; Jiang, P. Exosomes in Brain Diseases: Pathogenesis and Therapeutic Targets. MedComm 2023, 4, e287. [CrossRef]
- Xian, X.; Cai, L.-L.; Li, Y.; Wang, R.-C.; Xu, Y.-H.; Chen, Y.-J.; Xie, Y.-H.; Zhu, X.-L.; Li, Y.-F. Neuron Secrete Exosomes Containing miR-9-5p to Promote Polarization of M1 Microglia in Depression. J. Nanobiotechnology 2022, 20, 122. [CrossRef]
- Riancho, J.; Vázquez-Higuera, J.L.; Pozueta, A.; Lage, C.; Kazimierczak, M.; Bravo, M.; Calero, M.; Gonalezález, A.; Rodríguez, E.; Lleó, A.; et al. MicroRNA Profile in Patients with Alzheimer’s Disease: Analysis of miR-9-5p and miR-598 in Raw and Exosome Enriched Cerebrospinal Fluid Samples. J. Alzheimers Dis. JAD 2017, 57, 483–491. [CrossRef]
- Wang, X.; Zhou, Y.; Gao, Q.; Ping, D.; Wang, Y.; Wu, W.; Lin, X.; Fang, Y.; Zhang, J.; Shao, A. The Role of Exosomal microRNAs and Oxidative Stress in Neurodegenerative Diseases. Oxid. Med. Cell. Longev. 2020, 2020, 3232869. [CrossRef]
- You, Y.; Muraoka, S.; Jedrychowski, M.P.; Hu, J.; McQuade, A.K.; Young-Pearse, T.; Aslebagh, R.; Shaffer, S.A.; Gygi, S.P.; Blurton-Jones, M.; et al. Human Neural Cell Type-Specific Extracellular Vesicle Proteome Defines Disease-Related Molecules Associated with Activated Astrocytes in Alzheimer’s Disease Brain. J Extracell Vesicles 2022, 11, e12183. [CrossRef]
- 130. John van Geest Centre for Brain Repair, Department of Clinical Neurosciences, University of Cambridge, UK; Kouli, A.; Torsney, K.M.; Department of Medicine for the Elderly, Cambridge University Hospitals NHS Foundation Trust, UK; Clinical Gerontology Unit, Department of Public Health and Primary Care, University of Cambridge, UK; Kuan, W.-L.; John van Geest Centre for Brain Repair, Department of Clinical Neurosciences, University of Cambridge, UK Parkinson’s Disease: Etiology, Neuropathology, and Pathogenesis. In Parkinson’s Disease: Pathogenesis and Clinical Aspects; John Van Geest Centre for Brain Repair, Department of Clinical Neurosciences, University of Cambridge, UK, Stoker, T.B., Greenland, J.C., John Van Geest Centre for Brain Repair, Department of Clinical Neurosciences, University of Cambridge, UK, Eds.; Codon Publications, 2018; pp. 3–26 ISBN 978-0-9944381-6-4.
- Yu, H.; Sun, T.; An, J.; Wen, L.; Liu, F.; Bu, Z.; Cui, Y.; Feng, J. Potential Roles of Exosomes in Parkinson’s Disease: From Pathogenesis, Diagnosis, and Treatment to Prognosis. Front. Cell Dev. Biol. 2020, 8, 86. [CrossRef]
- Li, Z.; Liu, F.; He, X.; Yang, X.; Shan, F.; Feng, J. Exosomes Derived from Mesenchymal Stem Cells Attenuate Inflammation and Demyelination of the Central Nervous System in EAE Rats by Regulating the Polarization of Microglia. Int. Immunopharmacol. 2019, 67, 268–280. [CrossRef]
- Pinnell, J.R.; Cui, M.; Tieu, K. Exosomes in Parkinson Disease. J. Neurochem. 2021, 157, 413–428. [CrossRef]
- Agliardi, C.; Guerini, F.R.; Zanzottera, M.; Bolognesi, E.; Picciolini, S.; Caputo, D.; Rovaris, M.; Pasanisi, M.B.; Clerici, M. Myelin Basic Protein in Oligodendrocyte-Derived Extracellular Vesicles as a Diagnostic and Prognostic Biomarker in Multiple Sclerosis: A Pilot Study. Int. J. Mol. Sci. 2023, 24, 894. [CrossRef]
- Karnas, E.; Dudek, P.; Zuba-Surma, E.K. Stem Cell- Derived Extracellular Vesicles as New Tools in Regenerative Medicine - Immunomodulatory Role and Future Perspectives. Front. Immunol. 2023, 14, 1120175. [CrossRef]
- Ollen-Bittle, N.; Roseborough, A.D.; Wang, W.; Wu, J.D.; Whitehead, S.N. Mechanisms and Biomarker Potential of Extracellular Vesicles in Stroke. Biology 2022, 11, 1231. [CrossRef]
- Kawata, K.; Mitsuhashi, M.; Aldret, R. A Preliminary Report on Brain-Derived Extracellular Vesicle as Novel Blood Biomarkers for Sport-Related Concussions. Front. Neurol. 2018, 9, 239. [CrossRef]
- Fröhlich, D.; Kuo, W.P.; Frühbeis, C.; Sun, J.-J.; Zehendner, C.M.; Luhmann, H.J.; Pinto, S.; Toedling, J.; Trotter, J.; Krämer-Albers, E.-M. Multifaceted Effects of Oligodendroglial Exosomes on Neurons: Impact on Neuronal Firing Rate, Signal Transduction and Gene Regulation. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2014, 369, 20130510. [CrossRef]
- Ollen-Bittle, N.; Roseborough, A.D.; Wang, W.; Wu, J.D.; Whitehead, S.N. Mechanisms and Biomarker Potential of Extracellular Vesicles in Stroke. Biology 2022, 11, 1231. [CrossRef]
- Khan, N.A.; Asim, M.; El-Menyar, A.; Biswas, K.H.; Rizoli, S.; Al-Thani, H. The Evolving Role of Extracellular Vesicles (Exosomes) as Biomarkers in Traumatic Brain Injury: Clinical Perspectives and Therapeutic Implications. Front. Aging Neurosci. 2022, 14, 933434. [CrossRef]
- Wei, Z.-X.; Xie, G.-J.; Mao, X.; Zou, X.-P.; Liao, Y.-J.; Liu, Q.-S.; Wang, H.; Cheng, Y. Exosomes from Patients with Major Depression Cause Depressive-like Behaviors in Mice with Involvement of miR-139-5p-Regulated Neurogenesis. Neuropsychopharmacology 2020, 45, 1050–1058. [CrossRef]
- Lafourcade, C.A.; Fernández, A.; Ramírez, J.P.; Corvalán, K.; Carrasco, M.Á.; Iturriaga, A.; Bátiz, L.F.; Luarte, A.; Wyneken, U. A Role for Mir-26a in Stress: A Potential sEV Biomarker and Modulator of Excitatory Neurotransmission. Cells 2020, 9, 1364. [CrossRef]
- Couch, Y. Challenges Associated with Using Extracellular Vesicles as Biomarkers in Neurodegenerative Disease. Expert Rev. Mol. Diagn. 2023, 23, 1091–1105. [CrossRef]
- Khadka, A.; Spiers, J.G.; Cheng, L.; Hill, A.F. Extracellular Vesicles with Diagnostic and Therapeutic Potential for Prion Diseases. Cell Tissue Res. 2023, 392, 247–267. [CrossRef]
- Ugalde, C.L.; Finkelstein, D.I.; Lawson, V.A.; Hill, A.F. Pathogenic Mechanisms of Prion Protein, Amyloid-β and A-synuclein Misfolding: The Prion Concept and Neurotoxicity of Protein Oligomers. J. Neurochem. 2016, 139, 162–180. [CrossRef]
- Ananbeh, H.; Vodicka, P.; Kupcova Skalnikova, H. Emerging Roles of Exosomes in Huntington’s Disease. Int. J. Mol. Sci. 2021, 22, 4085. [CrossRef]
- Klingeborn, M.; Skiba, N.P.; Stamer, W.D.; Bowes Rickman, C. Isolation of Retinal Exosome Biomarkers from Blood by Targeted Immunocapture. Adv. Exp. Med. Biol. 2019, 1185, 21–25. [CrossRef]
- Kang, G.-Y.; Bang, J.Y.; Choi, A.J.; Yoon, J.; Lee, W.-C.; Choi, S.; Yoon, S.; Kim, H.C.; Baek, J.-H.; Park, H.S.; et al. Exosomal Proteins in the Aqueous Humor as Novel Biomarkers in Patients with Neovascular Age-Related Macular Degeneration. J. Proteome Res. 2014, 13, 581–595. [CrossRef]
- Mighty, J.; Rubio-Navarro, A.; Shi, C.; Zhou, J.; Flores-Bellver, M.; Heissel, S.; Onwumere, O.; Einbond, L.; Gharbaran, R.; Casper, D.S.; et al. Extracellular Vesicles of Human Diabetic Retinopathy Retinal Tissue and Urine of Diabetic Retinopathy Patients Are Enriched for the Junction Plakoglo Bin Protein. Front. Endocrinol. 2022, 13, 1077644. [CrossRef]
- Wang, P.; Li, C.; Deng, Y.; Yu, Q.; Meng, X.; Jiang, T.; Wang, Q.; Fu, Y. Effect of Plasma-Derived Extracellular Vesicles on Angiogenesis and the Ensuing Proliferative Diabetic Retinopathy through a miR-30b-Dependent Mechanism. Diabetol. Metab. Syndr. 2022, 14, 188. [CrossRef]
- Park, M.; Shin, H.A.; Duong, V.-A.; Lee, H.; Lew, H. The Role of Extracellular Vesicles in Optic Nerve Injury: Neuroprotection and Mitochondrial Homeostasis. Cells 2022, 11, 3720. [CrossRef]
- McDonnell, F.S.; Riddick, B.J.; Roberts, H.; Skiba, N.; Stamer, W.D. Comparison of the Extracellular Vesicle Proteome between Glaucoma and Non-Glaucoma Trabecular Meshwork Cells. Front. Ophthalmol. 2023, 3, 1257737. [CrossRef]
- Cells | Free Full-Text | Extracellular Vesicles from Ocular Melanoma Have Pro-Fibrotic and Pro-Angiogenic Properties on the Tumor Microenvironment Available online: https://www.mdpi.com/2073-4409/11/23/3828 (accessed on 16 January 2024).
- Pucker, A.D.; Ngo, W.; Postnikoff, C.K.; Fortinberry, H.; Nichols, J.J. Tear Film miRNAs and Their Association With Human Dry Eye Disease. Curr. Eye Res. 2022, 47, 1479–1487. [CrossRef]
- Wang, R.; Li, J.; Zhang, X.; Zhang, X.; Zhang, X.; Zhu, Y.; Chen, C.; Liu, Z.; Wu, X.; Wang, D.; et al. Extracellular Vesicles Promote Epithelial-to-Mesenchymal Transition of Lens Epithelial Cells under Oxidative Stress. Exp. Cell Res. 2021, 398, 112362. [CrossRef]
- Wu, L.; Zhou, L.; An, J.; Shao, X.; Zhang, H.; Wang, C.; Zhao, G.; Chen, S.; Cui, X.; Zhang, X.; et al. Comprehensive Profiling of Extracellular Vesicles in Uveitis and Scleritis Enables Biomarker Discovery and Mechanism Exploration. J. Transl. Med. 2023, 21, 388. [CrossRef]
- McKay, T.B.; Yeung, V.; Hutcheon, A.E.K.; Guo, X.; Zieske, J.D.; Ciolino, J.B. Extracellular Vesicles in the Cornea: Insights from Other Tissues. Anal. Cell. Pathol. 2021, 2021, 1–12. [CrossRef]
- Yeung, V.; Boychev, N.; Farhat, W.; Ntentakis, D.P.; Hutcheon, A.E.K.; Ross, A.E.; Ciolino, J.B. Extracellular Vesicles in Corneal Fibrosis/Scarring. Int. J. Mol. Sci. 2022, 23, 5921. [CrossRef]
- Hindman, H.B.; DeMagistris, M.; Callan, C.; McDaniel, T.; Bubel, T.; Huxlin, K.R. Impact of Topical Anti-Fibrotics on Corneal Nerve Regeneration in Vivo. Exp. Eye Res. 2019, 181, 49–60. [CrossRef]
- Katz, E.; Guaiquil, V.H.; Ivakhnitskaia, E.; Lara, D.; Anwar, K.; Jalilian, E.; Rosenblatt, M.; Djalilian, A.R. Exosomes as a Novel Multitarget Approach to Promote Growth of Corneal Sensory Neurons. Invest. Ophthalmol. Vis. Sci. 2021, 62, 749.
- Soleimani, M.; Mirshahi, R.; Cheraqpour, K.; Baharnoori, S.M.; Massoumi, H.; Chow, C.; Shahjahan, S.; Momenaei, B.; Ashraf, M.J.; Koganti, R.; et al. Intrastromal versus Subconjunctival Injection of Mesenchymal Stem/Stromal Cells for Promoting Corneal Repair. Ocul. Surf. 2023, 30, 187–195. [CrossRef]
- Ramos, T.; Parekh, M.; Kaye, S.B.; Ahmad, S. Epithelial Cell-Derived Extracellular Vesicles Trigger the Differentiation of Two Epithelial Cell Lines. Int. J. Mol. Sci. 2022, 23, 1718. [CrossRef]
- Verma, N.; Khare, D.; Poe, A.J.; Amador, C.; Ghiam, S.; Fealy, A.; Ebrahimi, S.; Shadrokh, O.; Song, X.-Y.; Santiskulvong, C.; et al. MicroRNA and Protein Cargos of Human Limbal Epithelial Cell-Derived Exosomes and Their Regulatory Roles in Limbal Stromal Cells of Diabetic and Non-Diabetic Corneas. Cells 2023, 12, 2524. [CrossRef]
- Poe, A.J.; Shah, R.; Khare, D.; Kulkarni, M.; Phan, H.; Ghiam, S.; Punj, V.; Ljubimov, A.V.; Saghizadeh, M. Regulatory Role of miR-146a in Corneal Epithelial Wound Healing via Its Inflammatory Targets in Human Diabetic Cornea. Ocul. Surf. 2022, 25, 92–100. [CrossRef]
- Barberis, E.; Vanella, V.V.; Falasca, M.; Caneapero, V.; Cappellano, G.; Raineri, D.; Ghirimoldi, M.; De Giorgis, V.; Puricelli, C.; Vaschetto, R.; et al. Circulating Exosomes Are Strongly Involved in SARS-CoV-2 Infection. Front. Mol. Biosci. 2021, 8, 632290. [CrossRef]
- Kakizaki, M.; Yamamoto, Y.; Yabuta, S.; Kurosaki, N.; Kagawa, T.; Kotani, A. The Immunological Function of Extracellular Vesicles in Hepatitis B Virus-Infected Hepatocytes. PloS One 2018, 13, e0205886. [CrossRef]
- Lee, J.-H. The Complex Role of Extracellular Vesicles in HIV Infection. BMB Rep. 2023, 56, 335–340. [CrossRef]
- Arakelyan, A.; Fitzgerald, W.; Zicari, S.; Vanpouille, C.; Margolis, L. Extracellular Vesicles Carry HIV Env and Facilitate Hiv Infection of Human Lymphoid Tissue. Sci. Rep. 2017, 7, 1695. [CrossRef]
- Sims, B.; Farrow, A.L.; Williams, S.D.; Bansal, A.; Krendelchtchikov, A.; Gu, L.; Matthews, Q.L. Role of TIM-4 in Exosome-Dependent Entry of HIV-1 into Human Immune Cells. Int. J. Nanomedicine 2017, 12, 4823–4833. [CrossRef]
- Tumne, A.; Prasad, V.S.; Chen, Y.; Stolz, D.B.; Saha, K.; Ratner, D.M.; Ding, M.; Watkins, S.C.; Gupta, P. Noncytotoxic Suppression of Human Immunodeficiency Virus Type 1 Transcription by Exosomes Secreted from CD8+ T Cells. J. Virol. 2009, 83, 4354–4364. [CrossRef]
- Sun, L.; Wang, X.; Zhou, Y.; Zhou, R.-H.; Ho, W.-Z.; Li, J.-L. Exosomes Contribute to the Transmission of Anti-HIV Activity from TLR3-Activated Brain Microvascular Endothelial Cells to Macrophages. Antiviral Res. 2016, 134, 167–171. [CrossRef]
- Haghighitalab, A.; Dominici, M.; Matin, M.M.; Shekari, F.; Ebrahimi Warkiani, M.; Lim, R.; Ahmadiankia, N.; Mirahmadi, M.; Bahrami, A.R.; Bidkhori, H.R. Extracellular Vesicles and Their Cells of Origin: Open Issues in Autoimmune Diseases. Front. Immunol. 2023, 14, 1090416. [CrossRef]
- Tang, T.-T.; Wang, B.; Lv, L.-L.; Liu, B.-C. Extracellular Vesicle-Based Nanotherapeutics: Emerging Frontiers in Anti-Inflammatory Therapy. Theranostics 2020, 10, 8111–8129. [CrossRef]
- Schioppo, T.; Ubiali, T.; Ingegnoli, F.; Bollati, V.; Caporali, R. The Role of Extracellular Vesicles in Rheumatoid Arthritis: A Systematic Review. Clin. Rheumatol. 2021, 40, 3481–3497. [CrossRef]
- Testa, U.; Pelosi, E.; Castelli, G.; Labbaye, C. miR-146 and miR-155: Two Key Modulators of Immune Response and Tumor Development. Non-Coding RNA 2017, 3, 22. [CrossRef]
- Ciesielski, O.; Biesiekierska, M.; Panthu, B.; Soszyński, M.; Pirola, L.; Balcerczyk, A. Citrullination in the Pathology of Inflammatory and Autoimmune Disorders: Recent Advances and Future Perspectives. Cell. Mol. Life Sci. 2022, 79, 94. [CrossRef]
- González-González, A.; García-Sánchez, D.; Dotta, M.; Rodríguez-Rey, J.C.; Pérez-Campo, F.M. Mesenchymal Stem Cells Secretome: The Cornerstone of Cell-Free Regenerative Medicine. World J. Stem Cells 2020, 12, 1529–1552. [CrossRef]
- Shimoda, M.; Khokha, R. Metalloproteinases in Extracellular Vesicles. Biochim. Biophys. Acta BBA - Mol. Cell Res. 2017, 1864, 1989–2000. [CrossRef]
- Admou, B.; Eddehbi, F.; Elmoumou, L.; Elmojadili, S.; Salami, A.; Oujidi, M.; Brahim, I.; Hazime, R. Anti-Double Stranded DNA Antibodies: A Rational Diagnostic Approach in Limited-Resource Settings. Pract. Lab. Med. 2022, 31, e00285. [CrossRef]
- Ohl, K.; Tenbrock, K. Inflammatory Cytokines in Systemic Lupus Erythematosus. J. Biomed. Biotechnol. 2011, 2011, 1–14. [CrossRef]
- Huang, Y.; Li, R.; Ye, S.; Lin, S.; Yin, G.; Xie, Q. Recent Advances in the Use of Exosomes in Sjögren’s Syndrome. Front. Immunol. 2020, 11, 1509. [CrossRef]
- Roescher, N.; Tak, P.; Illei, G. Cytokines in Sjögren’s Syndrome. Oral Dis. 2009, 15, 519–526. [CrossRef]
- Soares, R.L. Irritable Bowel Syndrome: A Clinical Review. World J. Gastroenterol. 2014, 20, 12144. [CrossRef]
- Uranga, J.A.; Martínez, V.; Abalo, R. Mast Cell Regulation and Irritable Bowel Syndrome: Effects of Food Components with Potential Nutraceutical Use. Molecules 2020, 25, 4314. [CrossRef]
- Weaver, K.R.; Mustapic, M.; Kapogiannis, D.; Henderson, W.A. Neuronal-enriched Extracellular Vesicles in Individuals with IBS: A Pilot Study of COMT and BDNF. Neurogastroenterol. Motil. 2022, 34, e14257. [CrossRef]
- Langhorst, J.; Elsenbruch, S.; Koelzer, J.; Rueffer, A.; Michalsen, A.; Dobos, G.J. Noninvasive Markers in the Assessment of Intestinal Inflammation in Inflammatory Bowel Diseases: Performance of Fecal Lactoferrin, Calprotectin, and PMN-Elastase, CRP, and Clinical Indices. Am. J. Gastroenterol. 2008, 103, 162–169. [CrossRef]
- Maggio, S.; Ceccaroli, P.; Polidori, E.; Cioccoloni, A.; Stocchi, V.; Guescini, M. Signal Exchange through Extracellular Vesicles in Neuromuscular Junction Establishment and Maintenance: From Physiology to Pathology. Int. J. Mol. Sci. 2019, 20, 2804. [CrossRef]
- Sabre, L.; Punga, T.; Punga, A.R. Circulating miRNAs as Potential Biomarkers in Myasthenia Gravis: Tools for Personalized Medicine. Front. Immunol. 2020, 11, 213. [CrossRef]
- Benjamin, E.J.; Blaha, M.J.; Chiuve, S.E.; Cushman, M.; Das, S.R.; Deo, R.; De Ferranti, S.D.; Floyd, J.; Fornage, M.; Gillespie, C.; et al. Heart Disease and Stroke Statistics—2017 Update: A Report From the American Heart Association. Circulation 2017, 135. [CrossRef]
- Sarlon-Bartoli, G.; Bennis, Y.; Lacroix, R.; Piercecchi-Marti, M.D.; Bartoli, M.A.; Arnaud, L.; Mancini, J.; Boudes, A.; Sarlon, E.; Thevenin, B.; et al. Plasmatic Level of Leukocyte-Derived Microparticles Is Associated With Unstable Plaque in Asymptomatic Patients With High-Grade Carotid Stenosis. J. Am. Coll. Cardiol. 2013, 62, 1436–1441. [CrossRef]
- Fu, S.; Zhang, Y.; Li, Y.; Luo, L.; Zhao, Y.; Yao, Y. Extracellular Vesicles in Cardiovascular Diseases. Cell Death Discov. 2020, 6, 68. [CrossRef]
- Liu, Y.; Li, Q.; Hosen, M.R.; Zietzer, A.; Flender, A.; Levermann, P.; Schmitz, T.; Frühwald, D.; Goody, P.; Nickenig, G.; et al. Atherosclerotic Conditions Promote the Packaging of Functional MicroRNA-92a-3p Into Endothelial Microvesicles. Circ. Res. 2019, 124, 575–587. [CrossRef]
- Li, H.; Liao, Y.; Gao, L.; Zhuang, T.; Huang, Z.; Zhu, H.; Ge, J. Coronary Serum Exosomes Derived from Patients with Myocardial Ischemia Regulate Angiogenesis through the miR-939-Mediated Nitric Oxide Signaling Pathway. Theranostics 2018, 8, 2079–2093. [CrossRef]
- Jansen, F.; Yang, X.; Proebsting, S.; Hoelscher, M.; Przybilla, D.; Baumann, K.; Schmitz, T.; Dolf, A.; Endl, E.; Franklin, B.S.; et al. MicroRNA Expression in Circulating Microvesicles Predicts Cardiovascular Events in Patients With Coronary Artery Disease. J. Am. Heart Assoc. 2014, 3, e001249. [CrossRef]
- Ribeiro-Rodrigues, T.M.; Laundos, T.L.; Pereira-Carvalho, R.; Batista-Almeida, D.; Pereira, R.; Coelho-Santos, V.; Silva, A.P.; Fernandes, R.; Zuzarte, M.; Enguita, F.J.; et al. Exosomes Secreted by Cardiomyocytes Subjected to Ischaemia Promote Cardiac Angiogenesis. Cardiovasc. Res. 2017, 113, 1338–1350. [CrossRef]
- Pantaleão, L.C.; Loche, E.; Fernandez-Twinn, D.S.; Dearden, L.; Córdova-Casanova, A.; Osmond, C.; Salonen, M.; Kajantie, E.; Niu, Y.; de Almeida-Faria, J.; et al. Programming of Cardiac Metabolism by miR-15b-5p, a miRNA Released in Cardiac Extracellular Vesicles Following Ischemia-Reperfusion Injury. Mol. Metab. 2024, 101875. [CrossRef]
- Vélez, P.; Parguiña, A.; Ocaranza-Sánchez, R.; Grigorian-Shamagian, L.; Rosa, I.; Alonso-Orgaz, S.; Cuesta, F.D.L.; Guitián, E.; Moreu, J.; Barderas, M.; et al. Identification of a Circulating Microvesicle Protein Network Involved in ST-Elevation Myocardial Infarction. Thromb. Haemost. 2014, 112, 716–726. [CrossRef]
- An, M.; Kwon, K.; Park, J.; Ryu, D.-R.; Shin, J.-A.; Lee Kang, J.; Choi, J.H.; Park, E.-M.; Lee, K.E.; Woo, M.; et al. Extracellular Matrix-Derived Extracellular Vesicles Promote Cardiomyocyte Growth and Electrical Activity in Engineered Cardiac Atria. Biomaterials 2017, 146, 49–59. [CrossRef]
- Halkein, J.; Tabruyn, S.P.; Ricke-Hoch, M.; Haghikia, A.; Nguyen, N.-Q.-N.; Scherr, M.; Castermans, K.; Malvaux, L.; Lambert, V.; Thiry, M.; et al. MicroRNA-146a Is a Therapeutic Target and Biomarker for Peripartum Cardiomyopathy. J. Clin. Invest. 2013, 123, 2143–2154. [CrossRef]
- Oh, J.G.; Watanabe, S.; Lee, A.; Gorski, P.A.; Lee, P.; Jeong, D.; Liang, L.; Liang, Y.; Baccarini, A.; Sahoo, S.; et al. miR-146a Suppresses SUMO1 Expression and Induces Cardiac Dysfunction in Maladaptive Hypertrophy. Circ. Res. 2018, 123, 673–685. [CrossRef]
- Eustes, A.S.; Dayal, S. The Role of Platelet-Derived Extracellular Vesicles in Immune-Mediated Thrombosis. Int. J. Mol. Sci. 2022, 23, 7837. [CrossRef]
- Bordin, A.; Chirivì, M.; Pagano, F.; Milan, M.; Iuliano, M.; Scaccia, E.; Fortunato, O.; Mangino, G.; Dhori, X.; De Marinis, E.; et al. Human Platelet Lysate-derived Extracellular Vesicles Enhance Angiogenesis through miR-126. Cell Prolif. 2022, 55, e13312. [CrossRef]
- Johnson, J.; Law, S.Q.K.; Shojaee, M.; Hall, A.S.; Bhuiyan, S.; Lim, M.B.L.; Silva, A.; Kong, K.J.W.; Schoppet, M.; Blyth, C.; et al. First-in-human Clinical Trial of Allogeneic, Platelet-derived Extracellular Vesicles as a Potential Therapeutic for Delayed Wound Healing. J. Extracell. Vesicles 2023, 12, 12332. [CrossRef]
- Gardin, C.; Ferroni, L.; Leo, S.; Tremoli, E.; Zavan, B. Platelet-Derived Exosomes in Atherosclerosis. Int. J. Mol. Sci. 2022, 23, 12546. [CrossRef]
- He, Y.; Wu, Q. The Effect of Extracellular Vesicles on Thrombosis. J Cardiovasc. Transl. Res. 2023, 16, 682–697. [CrossRef]
- Thangaraju, K.; Neerukonda, S.N.; Katneni, U.; Buehler, P.W. Extracellular Vesicles from Red Blood Cells and Their Evolving Roles in Health, Coagulopathy and Therapy. Int. J. Mol. Sci. 2020, 22, 153. [CrossRef]
- Georgatzakou, H.T.; Fortis, S.P.; Papageorgiou, E.G.; Antonelou, M.H.; Kriebardis, A.G. Blood Cell-Derived Microvesicles in Hematological Diseases and Beyond. Biomolecules 2022, 12, 803. [CrossRef]
- Simon, C.; Greening, D.W.; Bolumar, D.; Balaguer, N.; Salamonsen, L.A.; Vilella, F. Extracellular Vesicles in Human Reproduction in Health and Disease. Endocr. Rev. 2018, 39, 292–332. [CrossRef]
- Sang, Q.; Yao, Z.; Wang, H.; Feng, R.; Wang, H.; Zhao, X.; Xing, Q.; Jin, L.; He, L.; Wu, L.; et al. Identification of microRNAs in Human Follicular Fluid: Characterization of microRNAs That Govern Steroidogenesis in Vitro and Are Associated with Polycystic Ovary Syndrome in Vivo. J. Clin. Endocrinol. Metab. 2013, 98, 3068–3079. [CrossRef]
- Harp, D.; Driss, A.; Mehrabi, S.; Chowdhury, I.; Xu, W.; Liu, D.; Garcia-Barrio, M.; Taylor, R.N.; Gold, B.; Jefferson, S.; et al. Exosomes Derived from Endometriotic Stromal Cells Have Enhanced Angiogenic Effects in Vitro. Cell Tissue Res. 2016, 365, 187–196. [CrossRef]
- Berezin, A.E.; Kremzer, A.A.; Berezina, T.A.; Martovitskaya, Y.V. The Pattern of Circulating Microparticles in Patients with Diabetes Mellitus with Asymptomatic Atherosclerosis. Acta Clin. Belg. 2016, 71, 38–45. [CrossRef]
- Li, S.; Wei, J.; Zhang, C.; Li, X.; Meng, W.; Mo, X.; Zhang, Q.; Liu, Q.; Ren, K.; Du, R.; et al. Cell-Derived Microparticles in Patients with Type 2 Diabetes Mellitus: A Systematic Review and Meta-Analysis. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2016, 39, 2439–2450. [CrossRef]
- Kobayashi, Y.; Eguchi, A.; Tempaku, M.; Honda, T.; Togashi, K.; Iwasa, M.; Hasegawa, H.; Takei, Y.; Sumida, Y.; Taguchi, O. Circulating Extracellular Vesicles Are Associated with Lipid and Insulin Metabolism. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E574–E582. [CrossRef]
- Eguchi, A.; Lazic, M.; Armando, A.M.; Phillips, S.A.; Katebian, R.; Maraka, S.; Quehenberger, O.; Sears, D.D.; Feldstein, A.E. Circulating Adipocyte-Derived Extracellular Vesicles Are Novel Markers of Metabolic Stress. J. Mol. Med. Berl. Ger. 2016, 94, 1241–1253. [CrossRef]
- Murakami, T.; Horigome, H.; Tanaka, K.; Nakata, Y.; Ohkawara, K.; Katayama, Y.; Matsui, A. Impact of Weight Reduction on Production of Platelet-Derived Microparticles and Fibrinolytic Parameters in Obesity. Thromb. Res. 2007, 119, 45–53. [CrossRef]
- Pardo, F.; Villalobos-Labra, R.; Sobrevia, B.; Toledo, F.; Sobrevia, L. Extracellular Vesicles in Obesity and Diabetes Mellitus. Mol. Aspects Med. 2018, 60, 81–91. [CrossRef]
- Stepanian, A.; Bourguignat, L.; Hennou, S.; Coupaye, M.; Hajage, D.; Salomon, L.; Alessi, M.-C.; Msika, S.; de Prost, D. Microparticle Increase in Severe Obesity: Not Related to Metabolic Syndrome and Unchanged after Massive Weight Loss. Obes. Silver Spring Md 2013, 21, 2236–2243. [CrossRef]
- Campello, E.; Zabeo, E.; Radu, C.M.; Spiezia, L.; Foletto, M.; Prevedello, L.; Gavasso, S.; Bulato, C.; Vettor, R.; Simioni, P. Dynamics of Circulating Microparticles in Obesity after Weight Loss. Intern. Emerg. Med. 2016, 11, 695–702. [CrossRef]
- Liu, G.; Yin, X.-M. The Role of Extracellular Vesicles in Liver Pathogenesis. Am. J. Pathol. 2022, 192, 1358–1367. [CrossRef]
- Azparren-Angulo, M.; Royo, F.; Gonzalez, E.; Liebana, M.; Brotons, B.; Berganza, J.; Goñi-de-Cerio, F.; Manicardi, N.; Abad-Jordà, L.; Gracia-Sancho, J.; et al. Extracellular Vesicles in Hepatology: Physiological Role, Involvement in Pathogenesis, and Therapeutic Opportunities. Pharmacol. Ther. 2021, 218, 107683. [CrossRef]
- Povero, D.; Panera, N.; Eguchi, A.; Johnson, C.D.; Papouchado, B.G.; de Araujo Horcel, L.; Pinatel, E.M.; Alisi, A.; Nobili, V.; Feldstein, A.E. Lipid-Induced Hepatocyte-Derived Extracellular Vesicles Regulate Hepatic Stellate Cells via MicroRNA Targeting Peroxisome Proliferator-Activated Receptor-γ. Cell. Mol. Gastroenterol. Hepatol. 2015, 1, 646-663.e4. [CrossRef]
- Yuasa, T.; Amo-Shiinoki, K.; Ishikura, S.; Takahara, M.; Matsuoka, T.; Kaneto, H.; Kuroda, A.; Matsuhisa, M.; Hashida, S. Sequential Cleavage of Insulin Receptor by Calpain 2 and γ-Secretase Impairs Insulin Signalling. Diabetologia 2016, 59, 2711–2721. [CrossRef]
- Zhou, Y.; Ren, H.; Dai, B.; Li, J.; Shang, L.; Huang, J.; Shi, X. Hepatocellular Carcinoma-Derived Exosomal miRNA-21 Contributes to Tumor Progression by Converting Hepatocyte Stellate Cells to Cancer-Associated Fibroblasts. J. Exp. Clin. Cancer Res. 2018, 37, 324. [CrossRef]
- Gu, J.; Xu, H.; Chen, Y.; Li, N.; Hou, X. MiR-223 as a Regulator and Therapeutic Target in Liver Diseases. Front. Immunol. 2022, 13, 860661. [CrossRef]
- Liver Cancer: Your Chances for Recovery (Prognosis) | Saint Luke’s Health System Available online: https://www.saintlukeskc.org/health-library/liver-cancer-your-chances-recovery-prognosis (accessed on 23 January 2024).
- Grange, C.; Bussolati, B. Extracellular Vesicles in Kidney Disease. Nat. Rev. Nephrol. 2022, 18, 499–513. [CrossRef]
- Korecka, K.; Gawin, M.; Pastuszka, A.; Partyka, M.; Koszutski, T.; Pietrowska, M.; Hyla-Klekot, L. Proteomics of Urinary Small Extracellular Vesicles in Early Diagnosis of Kidney Diseases in Children-Expectations and Limitations. PROTEOMICS n/a, 2300168. [CrossRef]
- Barutta, F.; Tricarico, M.; Corbelli, A.; Annaratone, L.; Pinach, S.; Grimaldi, S.; Bruno, G.; Cimino, D.; Taverna, D.; Deregibus, M.C.; et al. Urinary Exosomal microRNAs in Incipient Diabetic Nephropathy. PloS One 2013, 8, e73798. [CrossRef]
- Lv, L.-L.; Cao, Y.-H.; Ni, H.-F.; Xu, M.; Liu, D.; Liu, H.; Chen, P.-S.; Liu, B.-C. MicroRNA-29c in Urinary Exosome/Microvesicle as a Biomarker of Renal Fibrosis. Am. J. Physiol.-Ren. Physiol. 2013, 305, F1220–F1227. [CrossRef]
- Eirin, A.; Zhu, X.-Y.; Puranik, A.S.; Tang, H.; McGurren, K.A.; van Wijnen, A.J.; Lerman, A.; Lerman, L.O. Mesenchymal Stem Cell-Derived Extracellular Vesicles Attenuate Kidney Inflammation. Kidney Int. 2017, 92, 114–124. [CrossRef]
- Raimondo, F.; Morosi, L.; Corbetta, S.; Chinello, C.; Brambilla, P.; Mina, P.D.; Villa, A.; Albo, G.; Battaglia, C.; Bosari, S.; et al. Differential Protein Profiling of Renal Cell Carcinoma Urinary Exosomes. Mol. Biosyst. 2013, 9, 1220–1233. [CrossRef]
- Takeda, M.; Akamatsu, S.; Kita, Y.; Goto, T.; Kobayashi, T. The Roles of Extracellular Vesicles in the Progression of Renal Cell Carcinoma and Their Potential for Future Clinical Application. Nanomaterials 2023, 13, 1611. [CrossRef]
- Jin, C.; Shi, L.; Li, K.; Liu, W.; Qiu, Y.; Zhao, Y.; Zhao, B.; Li, Z.; Li, Y.; Zhu, Q. Mechanism of Tumor-Derived Extracellular Vesicles in Regulating Renal Cell Carcinoma Progression by the Delivery of MALAT1. Oncol. Rep. 2021, 46, 187. [CrossRef]
- Holtzman, J.; Lee, H. Emerging Role of Extracellular Vesicles in the Respiratory System. Exp. Mol. Med. 2020, 52, 887–895. [CrossRef]
- Lee, H.; Zhang, D.; Laskin, D.L.; Jin, Y. Functional Evidence of Pulmonary Extracellular Vesicles in Infectious and Noninfectious Lung Inflammation. J. Immunol. Baltim. Md 1950 2018, 201, 1500–1509. [CrossRef]
- Nagano, T.; Katsurada, M.; Dokuni, R.; Hazama, D.; Kiriu, T.; Umezawa, K.; Kobayashi, K.; Nishimura, Y. Crucial Role of Extracellular Vesicles in Bronchial Asthma. Int. J. Mol. Sci. 2019, 20, 2589. [CrossRef]
- Fujita, Y.; Kadota, T.; Araya, J.; Ochiya, T.; Kuwano, K. Clinical Application of Mesenchymal Stem Cell-Derived Extracellular Vesicle-Based Therapeutics for Inflammatory Lung Diseases. J. Clin. Med. 2018, 7, 355. [CrossRef]
- O’Farrell, H.E.; Yang, I.A. Extracellular Vesicles in Chronic Obstructive Pulmonary Disease (COPD). J. Thorac. Dis. 2019, 11, S2141–S2154. [CrossRef]
- Hough, K.P.; Chanda, D.; Duncan, S.R.; Thannickal, V.J.; Deshane, J.S. Exosomes in Immunoregulation of Chronic Lung Diseases. Allergy 2017, 72, 534–544. [CrossRef]
- Carnino, J.M.; Lee, H.; Jin, Y. Isolation and Characterization of Extracellular Vesicles from Broncho-Alveolar Lavage Fluid: A Review and Comparison of Different Methods. Respir. Res. 2019, 20, 240. [CrossRef]
- Fujita, Y.; Araya, J.; Ito, S.; Kobayashi, K.; Kosaka, N.; Yoshioka, Y.; Kadota, T.; Hara, H.; Kuwano, K.; Ochiya, T. Suppression of Autophagy by Extracellular Vesicles Promotes Myofibroblast Differentiation in COPD Pathogenesis. J. Extracell. Vesicles 2015, 4, 28388. [CrossRef]
- Tan, W.S.D.; Shen, H.-M.; Wong, W.S.F. Dysregulated Autophagy in COPD: A Pathogenic Process to Be Deciphered. Pharmacol. Res. 2019, 144, 1–7. [CrossRef]
- Perera, G.K.; Di Meglio, P.; Nestle, F.O. Psoriasis. Annu. Rev. Pathol. 2012, 7, 385–422. [CrossRef]
- Shao, S.; Fang, H.; Li, Q.; Wang, G. Extracellular Vesicles in Inflammatory Skin Disorders: From Pathophysiology to Treatment. Theranostics 2020, 10, 9937–9955. [CrossRef]
- Tamagawa-Mineoka, R.; Katoh, N.; Kishimoto, S. Platelet Activation in Patients with Psoriasis: Increased Plasma Levels of Platelet-Derived Microparticles and Soluble P-Selectin. J. Am. Acad. Dermatol. 2010, 62, 621–626. [CrossRef]
- Takeshita, J.; Mohler, E.R.; Krishnamoorthy, P.; Moore, J.; Rogers, W.T.; Zhang, L.; Gelfand, J.M.; Mehta, N.N. Endothelial Cell-, Platelet-, and Monocyte/Macrophage-Derived Microparticles Are Elevated in Psoriasis beyond Cardiometabolic Risk Factors. J. Am. Heart Assoc. 2014, 3, e000507. [CrossRef]
- Martínez-Sales, V.; Vila, V.; Ricart, J.M.; Vayá, A.; Todolí, J.; Nńñez, C.; Contreras, T.; Ballester, C.; Reganon, E. Increased Circulating Endothelial Cells and Microparticles in Patients with Psoriasis. Clin. Hemorheol. Microcirc. 2015, 60, 283–290. [CrossRef]
- Ovcina-Kurtovic, N.; Kasumagic-Halilovic, E. Serum Levels of Tumor Necrosis Factor - Alpha in Patients With Psoriasis. Mater. Socio-Medica 2022, 34, 40–43. [CrossRef]
- Dainichi, T.; Kitoh, A.; Otsuka, A.; Nakajima, S.; Nomura, T.; Kaplan, D.H.; Kabashima, K. The Epithelial Immune Microenvironment (EIME) in Atopic Dermatitis and Psoriasis. Nat. Immunol. 2018, 19, 1286–1298. [CrossRef]
- Zhou, H.; Tan, X.; Chen, G.; Liu, X.; Feng, A.; Liu, Z.; Liu, W. Extracellular Vesicles of Commensal Skin Microbiota Alleviate Cutaneous Inflammation in Atopic Dermatitis Mouse Model by Re-Establishing Skin Homeostasis. J. Invest. Dermatol. 2023, S0022-202X(23)00169-0. [CrossRef]
- Hong, S.-W.; Kim, M.-R.; Lee, E.-Y.; Kim, J.H.; Kim, Y.-S.; Jeon, S.G.; Yang, J.-M.; Lee, B.-J.; Pyun, B.-Y.; Gho, Y.S.; et al. Extracellular Vesicles Derived from Staphylococcus Aureus Induce Atopic Dermatitis-like Skin Inflammation. Allergy 2011, 66, 351–359. [CrossRef]
- Li, C.; Sun, C.; Lohcharoenkal, W.; Ali, M.M.; Xing, P.; Zheng, W.; Görgens, A.; Gustafsson, M.O.; EL Andaloussi, S.; Sonkoly, E.; et al. Cutaneous Squamous Cell Carcinoma-Derived Extracellular Vesicles Exert an Oncogenic Role by Activating Cancer-Associated Fibroblasts. Cell Death Discov. 2023, 9, 1–12. [CrossRef]
Figure 1.
Schematic of CTS-EVs and their role in different physiological and pathological processes. From right to left, the process of microvesicle and exosome biogenesis and exocytosis from their cell of origin is detailed. CTS-EV responses have a variety of diagnostic and therapeutic implications in a variety of disease types as shown here.
Figure 1.
Schematic of CTS-EVs and their role in different physiological and pathological processes. From right to left, the process of microvesicle and exosome biogenesis and exocytosis from their cell of origin is detailed. CTS-EV responses have a variety of diagnostic and therapeutic implications in a variety of disease types as shown here.
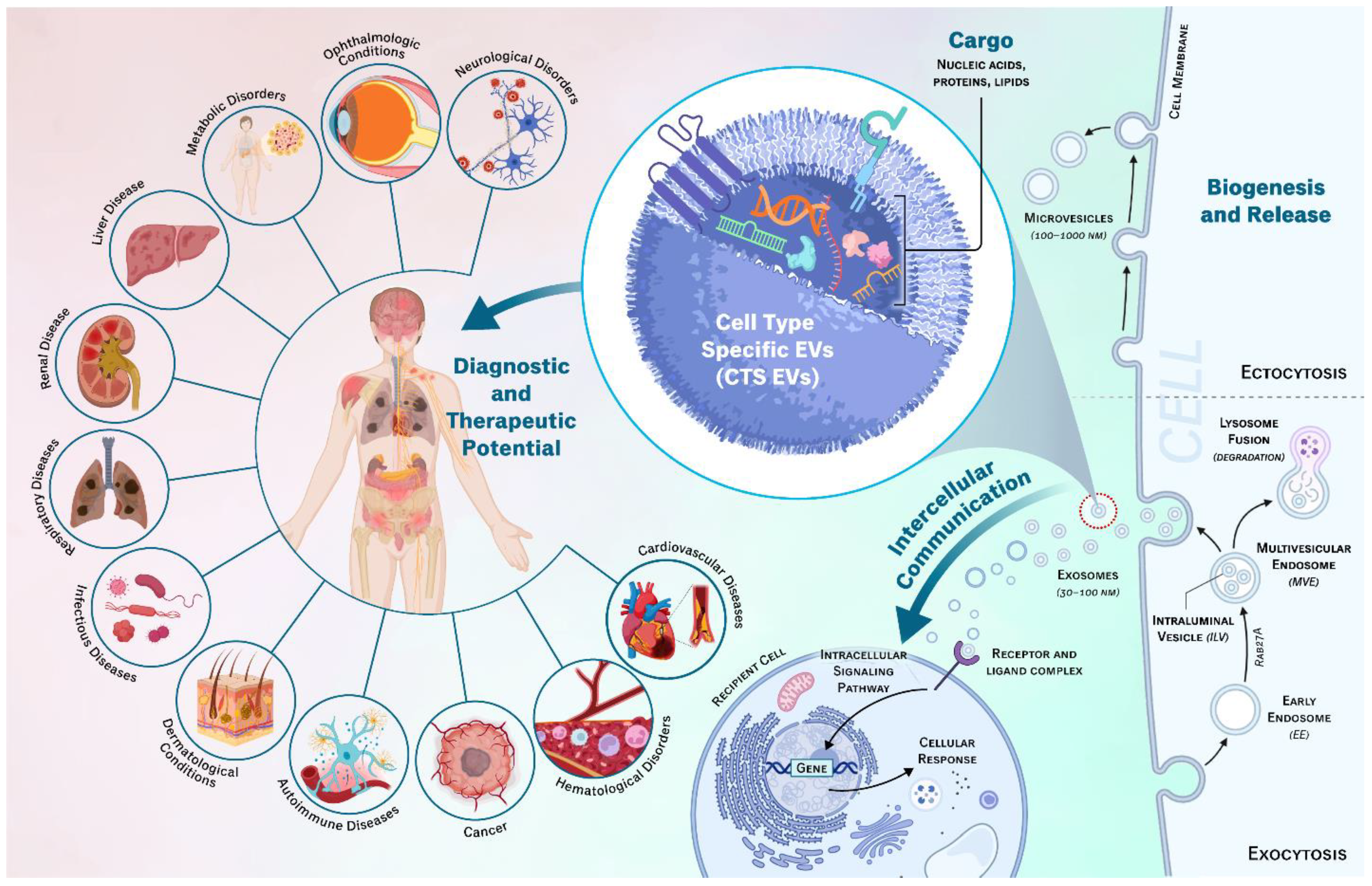

Table 1.
Overview of Diseases and Disorders Associated with CTS-Derived EVs.
| Disease Type | Specific Diseases | References | |
|---|---|---|---|
| 3.2 | Cancer | Breast, Lung, Prostate, Leukemia, Ovarian, Glioblastoma, Pancreatic, Colorectal, Bladder, Oropharyngeal, Gastric | [75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112,113,114,115,116,117,118,119,120] |
| 3.3 | Neurological Disorders | Alzheimer’s, Parkinson, Multiple Sclerosis, Stroke, Epilepsy, Depression, Traumatic Brain Injury (TBI), Multiple sclerosis, Huntington, ALS, Prion disease | [121,122,123,124,125,126,127,128,129,130,131,132,133,134,135,136,137,138,139,140,141,142,143,144,145,146] |
| 3.4 | Ophthalmologic Conditions | Age-Related Macular Degeneration (AMD), Diabetic Retinopathy, Glaucoma, Uveitis, Retinal dystrophies, Dry Eye Disease, Corneal trauma, Posterior Capsular Opacification, Ocular Melanoma | [1,5,6,7,147,148,149,150,151,152,153,154,155,156,157,158,159,160,161,162,163,164] |
| 3.5 | Infectious Diseases | Human Immunodeficiency Virus (HIV), Hepatitis Viruses, COVID-19, Tuberculosis | [50,165,166,167,168,169,170,171] |
| 3.6 | Autoimmune Diseases | Rheumatoid Arthritis (RA), Systemic Lupus Erythematosus (SLE), Sjogren’s Syndrome, Inflammatory Bowel Diseases (IBD), Myasthenia Gravis, Autoimmune Thyroid Diseases, Celiac Disease, Systemic Sclerosis (Scleroderma), Antiphospholipid Syndrome | [172,173,174,175,176,177,178,179,180,181,182,183,184,185,186,187,188] |
| 3.7 | Cardiovascular diseases | Myocardial infarction, Coronary artery disease, Atrial fibrillation, Cardiomyopathy | [189,190,191,192,193,194,195,196,197,198,199,200] |
| 3.8 | Hematological Disorders | Thrombosis, Sickle Cell anemia, Hemophilia, Anemia, Thrombocytopenia, Immunodeficiency | [50,201,202,203,204,205,206,207] |
| 3.9 | Reproductive Disorders | Endometriosis, Infertility, Genetic Testing, Polycystic Ovarian Syndrome (PCOS) | [208,209,210] |
| 3.10 | Metabolic Disorders | Diabetes, Metabolic syndrome, Obesity | [211,212,213,214,215,216,217,218] |
| Liver Disease | Hepatocellular carcinoma, Liver fibrosis and cirrhosis | [219,220,221,222,223,224,225] | |
| 3.11 | Renal Diseases | Chronic kidney disease, Renal artery stenosis, Renal cell Carcinoma | [226,227,228,229,230,231,232,233] |
| 3.12 | Respiratory Diseases | Acute Lung Injury (ARI), Acute Respiratory Distress Syndrome (ARDS), Chronic obstructive pulmonary disease (COPD), Asthma | [234,235,236,237,238,239,240,241,242] |
| 3.13 | Dermatological Conditions | Psoriasis, Atopic dermatitis, Skin Cancer | [243,244,245,246,247,248,249,250,251,252] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



