
Submitted:
06 February 2024
Posted:
06 February 2024
You are already at the latest version
Abstract
Lactobacilli are considered an inexhaustible source of potentially bioactive substances; indeed, several products from their metabolism are known to have immunomodulatory and anti-inflammatory activity. Recently, we demonstrated that cell-free supernatants (CFS) obtained from Lactobacillus (L.) acidophilus, L. plantarum, L. rhamnosus and L. reuteri can impair Candida pathogenic potential in an in vitro model of epithelial vaginal infection. This effect could be ascribed to a direct effect of living lactobacilli on Candida virulence and to the production of metabolites which are able to impair fungal virulence. In the present work, stemming from those data, we deepened our knowledge on the CFS from these 4 lactobacilli by performing a metabolomic analysis to better characterize their composition. By using an untargeted metabolomic approach, we detected consistent differences in the metabolites produced by these four different lactobacilli. Interestingly, L. rhamnosus showed the most peculiar metabolic profile. Specifically, after a hierarchical clustering analysis in positive and negative ionization mode, L. rhamnosus showed a specific area of significant overexpressed metabolites that strongly differed from the same area in other lactobacilli. In this area, inosine was identified among the overexpressed metabolites of L. rhamnosus. This molecule has been described to have antioxidant, anti-inflammatory, anti-infective and neuroprotective properties. The biological significance of its overproduction by L. rhamnosus might be important in its probiotic and/or postbiotic activity.
Keywords:
L. rhamnosus (L. RHA) metabolome
; inosine
; untargeted metabolomics
1. Introduction
Among probiotics, lactobacilli are beneficial microbes for human health, when administered in adequate quantity [1,2]. As all the probiotics, lactobacilli have effects on microbial pathogens and on the host. Specifically, lactobacilli compete with pathogens for nutrients and binding to receptors and they also produce antimicrobial molecules to inhibit the growth of pathogens. Their beneficial effects on the host include improvement of the epithelial barrier function (through the enhanced production of mucus and of tight junction proteins that help to prevent the passage of the pathogens to the blood), the modulation of dendritic cells and T cells activity (immunomodulatory effects), and the regulation of the production and secretion of several neurotransmitters [1,2]. In addition, lactobacilli help to prevent and manage several pathological conditions, such as allergic diseases, cancer, hypercholesterolemia, irritable bowel syndrome, diarrhea, lactose intolerance and inflammatory bowel disease [1,3]. One of the main roles played by probiotic lactobacilli is to help the recovery of the eubiosis state in the host. However, the way how this goal is achieved is partly unknown. In particular, the precise role of the metabolites produced by specific bacteria during their life cycle and their impact on the environment where they proliferate is yet to be elucidated. In addition, it must be considered that the use of living bacteria in vulnerable people is linked to possible safety concerns; also, to maintain bacterial viability is a challenging task [4]. Interestingly, new scientific evidence points out that the health benefits granted by lactobacilli are not necessarily related to viable bacteria. Indeed, also their metabolites or bacterial components, collectively indicated as postbiotics, may be the driving force behind health promotion. Postbiotics have been shown to have several biological activities (antimicrobial, antioxidant, anti-inflammatory, anti-proliferative, and immunomodulatory). Moreover, numerous studies have suggested the significant potential of postbiotics for disease treatment [5]. The metabolites produced by lactobacilli, especially some antimicrobial molecules already identified, can inhibit the growth of pathogens [6,7]. In addition, during the interaction with the host and other microorganisms that dwell in the same host niche, the metabolites produced by “beneficial microbes” such as lactobacilli may exert a significant impact to counteract the infection process [8,9]. Similarly to the living bacteria, the metabolites produced by probiotics have been demonstrated to have many beneficial effects on the host, such as the improvement of barrier function, (stimulating the enhanced production of tight junctions’ proteins and mucous), the promotion of changes in the microbiota composition, immunomodulatory and anti-inflammatory activities [8,10]. Since postbiotics are made up of inactivated microbial cells and/or cell components, their employment is characterized by higher levels of stability and safety for the user. Consequently, interest is increasing on their possible therapeutic employment, also because they can be considered an inexhaustible source of possible new bioactive substances [11]. We recently showed that cell-free supernatants (CFS) obtained from Lactobacillus (L.) rhamnosus (L. RHA), L. acidophilus (L. AC), L. plantarum (L. PLA) and L. reuteri (L. REU) could impair Candida parapsilosis (C. parapsilosis) pathogenic potential in an in vitro model of epithelial vaginal infection [12]. This effect could be ascribed to the direct effect of lactobacilli on Candida virulence, and to the production of metabolites by lactobacilli which are able to weaken C. parapsilosis virulence [12]. Moreover, it has been recently shown that L. RHA could impair C. albicans pathogenicity in a model of intestinal epithelial infection [9]. Therefore, by improving our knowledge on the metabolome of beneficial microorganisms that can act within specific host niches, novel important information would become available on the mechanisms they use to interact with the resident microbiota and with the host cells. For this reason, here an untargeted metabolomics approach was applied to compare the metabolome of four different lactobacilli often used as probiotics: L. RHA, L. AC, L. REU and L. PLA. Our data show that such metabolomes are significantly different, resulting in an increased production of some specific metabolites, such as inosine, from L. RHA. Since inosine can exert antioxidant, anti-inflammatory and neuroprotective effects [13], other than displaying relevant properties in the prokaryotic metabolisms, our data suggest that the overproduction of inosine by L. RHA could have a positive impact on the host and even on its resident microbiota.
2. Materials and Methods
Lactobacillus strains and growth conditions:
Four different Lactobacillus (L.) strains were employed in this study: L. acidophilus ATCC 314, L. reuteri DSM 17938, L. rhamnosus ATCC 7469 and L. plantarum ATCC 8014. The Lactobacillus colonies were inoculated in 5 ml of MRS liquid medium (De Man, Rogosa and Sharpe, Oxoid LTD, England) and incubated for 24 h at 37°C, under agitation and in anaerobic conditions. After incubation, bacteria were centrifuged, washed twice with PBS, counted, and resuspended at 1x108/ml in 5 ml of MRS broth and incubated for 24 h at 37 °C under agitation in anaerobic conditions. After incubation the cell-free supernatant (CFS) were preprepared as detailed below.
Preparation of cell-free supernatants (CFS) from Lactobacillus strains:
The cell free supernatants (CFS) of lactobacilli were obtained by centrifugation of the bacterial suspensions carried out at 4,000 rpm, at 4 °C for 15 minutes. The supernatants were then collected and filtered with 0.22 μm syringe filters (Corning Incorporated, Germany). The potential bacterial contamination of CFS was excluded incubating 1 ml of each CFS at 37 °C and checking the turbidity (from 24h to 72h). The pH of each CFS was measured by a pH meter (Hanna Instrument, Italy), returning an average pH = 4, as previously described [12]. The control samples consisted of sterile MRS medium (blank). The CFS obtained were finally stored at -80 °C until their use.
Liquid Chromatography – Electrospray/High Resolution Mass Spectrometry (HPLC-ESI/HRMS)
The CFS from lactobacilli, which had been stored at -80°C, were thawed and centrifuged at 14,000 rpm for 10 minutes. Subsequently, the CFS were transferred to Amicon-Ultra 0.5 tubes, centrifuged at 14,000 rpm for 15 minutes and then transferred into the autosampler vials pending analysis. The Quality Control pool samples (QC) were prepared by mixing equal volumes of each cohort supernatant and used to minimize technical data variance [14].
The analyses were performed using an Ultimate 3000 HPLC connected to a QExactive High Resolution Mass spectrometer via a HESI-II electrospray ionization source (Thermo Scientific), controlled by Xcalibur software (Thermo Scientific, v. 29 build 2926). A 10 µL volume of sample solution was injected onto a Hypersil Gold C18 100 x 2.1 mm ID 1.9µm ps column (Thermo Scientific) kept at 30 °C and separation was performed at 0.4 mL/min flow with a gradient elution scheme using methanol (B) and 0.1% formic acid in water (A). The mobile phase composition was kept at 2% B for 1 minute after injection then linearly raised to 42% B in 60 minutes and further on to 98% B in 5 minutes. Methanol was kept at 98% up to minute 74.9 then lowered to 2% at minute 75. The total runtime was 90 minutes. ESI source was operated in both positive and negative ionization mode. Capillary temperature was set at 320 °C; the following nitrogen flows (arbitrary units) were used to assist the ionization: Sheath Gas 45, Aux Gas 25 (at 290 °C), Sweep Gas 2. The capillary voltage was set to 3.8 kV (3.4 kV for negative ionization) and S-Lens RF level was set at 45 (arbitrary units).
A Data-Dependent Acquisition (DDA) strategy was used to acquire MS2 fragmentation spectra of the Top 5 singly charged precursor ions revealed in Full Scan MS experiments. Positive and Negative ionization DDA experiments were performed in separate analyses. Full MS spectra were obtained from m/z 100 to 1500 at 70000 FWHM resolving power using an automatic gain control (AGC) of 3 x 106 and a maximum Injection Time (max IT) of 250 ms. Fragmentation Spectra (MS2) acquisition was performed at 17500 FWHM, with 2 x 105 AGC target and 120 ms max IT. The isolation window for precursor ion selection was set at 1.0 Th and HCD normalized collision energy (NCE) was stepped at 20, 50 and 80. Fragmented precursors were dynamically excluded for 6 seconds. Inosine standard was purchased from Sigma-Aldrich. The data are from triplicates samples from 3 different experiments.
Compounds discoverer data analysis:
Raw files (triplicates samples from 3 different experiments) were processed by Compound Discoverer (CD) 3.3.2.31 (Copyright 2014-2023 Thermo Fisher Scientific Inc.) using a slightly modified processing workflow template for Untargeted Metabolomics with Statistics Detect Unknowns with ID Using Local Databases. The core of the workflow consisted of Spectra selection from raw files (Retention Time limited from 0.2 to 75 min), Retention Time Alignment (ChromAlign) with respect to a QC sample file, Compound Detection and Grouping with RT tolerance of 0.3 min and 5 ppm mass deviation. Then Gap Filling, SERRF QC Correction and Background removal were performed along with Compound Annotation using Predicted Composition and different types of databases (mzCloud, Metabolika, Human Metabolome Database, Chemspider, BioCyc) [15]. The so detected compounds were used for differential analysis of sample groups (Nested Design; Generated Ratios: lactobacilli PLA/AC, REU/AC, RHA/AC, REU/PLA, RHA/PLA and RHA/REU).
3. Results
Here, an untargeted metabolomics approach was used to compare the metabolome from four different species of lactobacilli, often used as probiotics: L. rhamnosus (L. RHA), L. acidophilus (L. AC), L. plantarum (L. PLA) and L. reuteri (L. REU). The Principal Component Analysis (PCA)-2 showed that the metabolome differed between L. RHA and the other species, as well as between L. AC and the other lactobacilli. Conversely, the metabolome of L. PLA and L. REU were more similar, according to the analysis carried out both in positive and in negative ionization mode (Figure 1).
A hierarchical clustering analysis in positive and negative ionization mode, carried out to compare the four metabolomes, revealed distinct cluster of metabolites overexpressed in the CFS of the different lactobacilli. Once again, L. PLA and L. REU showed a more similar metabolome profiles; differently, L. RHA and L. AC showed a more peculiar metabolome profile (Figure 2). Specifically, L. RHA revealed an area of significantly overexpressed metabolites (p value <0.01; Log2 fold change=2) that strongly differed from the same areas from L. AC, L. PLA and L. REU (Figure 2, see red line). Therefore, we performed a more detailed analysis of the metabolites included in this area of L. RHA. Within this area of overexpressed compounds, we considered only those with the best identification profile, according to: Predicted Compositions, mzCloud Search, mzVault Search, Metabolika Search, ChemSpider Search and MassList Search.
According to the results of the analysis, inosine returned the best identification profile, as shown in Table 1.
Table 2 shows the identified pathways that included inosine. For each pathway, the mapped and matched compounds and the total compounds in the pathway are shown.
Interestingly, this molecule has also a well-known biological role. Indeed, inosine is a key intracellular energy substrate for nucleotide synthesis by salvage pathways and it possesses cell protective activity and cell repair properties [16].
To increase the identification confidence over inosine, from probable to possibly confirmed structure [17], an inosine reference standard solution was used to confirm the [M+H]+ molecular ion mass-to-charge ratio, along with its fragmentation spectrum and retention time (Figure 3 and Figure 4).
Once confirmed the identification, inosine from L. RHA CFS was quantified using a set of calibration samples obtained by adding proper amount of inosine to MRS covering from 1 to 50 µg/ml concentration range. Inosine of the L. RHA sample was quantified in the range of 5-8 µg/ml.
4. Discussion
Lactobacilli are beneficial microbes, and they are often used as probiotics. The concept of probiotics has been evolving with the currently accepted definition of “living microorganisms that can benefit the host when consumed in sufficient quantities” [18]. Although this definition implies that microorganisms must be viable to be beneficial, increasing evidence suggests that also microbial products provide benefits to the host [19,20]. Indeed, the so-called postbiotics, also known as metabolites, biogenic or cell-free-supernatants (CFS), are defined as “soluble factors secreted by living bacteria or released by bacterial lysis”, and their role in providing health benefits to the host has been reported [21]. Hence, today it is demonstrated that the beneficial effects of lactobacilli are based either on living bacteria (the “probiotics”), or on their metabolites/cell lysates (the “postbiotics”). Here, we describe the assessment of the metabolomic profile of four different lactobacilli, currently used as safe probiotics: L. RHA, L. AC, L. PLA and L. REU [22]. An untargeted metabolomic approach has been employed to compare the differences in metabolites production in the CFS from the different lactobacilli under the same culture conditions. Specifically, the hierarchical clustering analysis of the compounds release by the four lactobacilli showed for L. RHA a specific area of significantly overexpressed metabolites, that strongly differ from the same area of L. AC, L. PLA and L. REU. It has been shown that CFS from L. RHA strain SCB0119 altered the transcription profiles of several genes involved in fatty acid degradation, ion transport, and the biosynthesis of amino acids in E. coli, as well as fatty acid degradation, protein synthesis, DNA replication, and ATP hydrolysis in Staphylococcus aureus, which are important for bacterial survival and growth [23]. In addition, L. RHA colonization of the epithelial cells has been demonstrated to be responsible for the drastic changes in the metabolic environment, forcing metabolic adaptation in C. albicans and reducing fungal virulence [9]. Furthermore, antimicrobial properties of L. RHA have been described also against Listeria monocytogenes [24] and Salmonella spp. [25,26]. Therefore, according to all the above-mentioned literature data, reporting antimicrobial and antifungal activity of L. RHA [27,28,29], a detailed investigation has been carried out to get more information on the metabolites overexpressed by this species. Among the upregulated compounds with the best identification profile in the CFS obtained by L. RHA, we have identified inosine as one of the molecules strongly overproduced. Inosine is a non-canonical nucleotide, mainly occurring in the form of a monophosphate. It base pairs with deoxythymidine, deoxyadenosine and deoxyguanosine [30]. Among the possible role of such unconventional nucleotide, it has been reported that the incorporation of inosine in place of guanine modulates translational events [31]. Several studies carried out in various neuronal cell types have identified a growth-promoting activity of inosine, comparable to that induced by canonical neurotrophic factors such as brain-derived neurotrophic factor (BDNF) or nerve growth factor (NGF) [32,33]. Benowitz and colleagues have shown that inosine promotes axon outgrowth in a rat model of corticospinal tract injury [34]. Furthermore, inosine has been demonstrated to modulate several biological processes through the adenosine receptors, such as the enhancement of neurite outgrowth in depressive disorders [35]. Because of its antioxidant, anti-inflammatory, pro-axogenic and neuroprotective functions, inosine is also employed as a therapeutic supplement, and it is prescribed in cases of nerve injury, inflammation and oxidative stress [13,36]. In addition, several drugs used in the treatment of autoimmune and inflammatory diseases (such as adenosine kinase inhibitors) exert their beneficial effects by releasing adenosine [37]. Since the latter is readily degraded to inosine in the extracellular space, it is conceivable a direct involvement of inosine in the anti-inflammatory effects of these adenosine-releasing agents [37]. Inosine has also immunomodulatory effects by contributing to the efficacy of Isoprinosine (inosine pranobex), a synthetic agent formed by inosine combined with the immunostimulant dimepranol acedoben (acetamidobenzoic acid and dimethylaminoisopropanol). Even though many of the biological actions of inosine (particularly in the context of microbial infections) have yet to be described, this molecule is already employed for the treatment of acute respiratory viral infections, genital warts, herpes simplex infections, hepatitis B and subacute sclerosing panencephalitis [13]. Inosine is used also for the treatment of sepsis in infections, and it has been shown to reduce systemic inflammation, organ damage, tissue dysoxia, and vascular dysfunction, resulting in improved survival in a mouse model of septic shock [38].
5. Conclusions
In conclusion, here we show that L. RHA overproduce inosine during its life cycle, and this might have a significant impact when administered in vivo. The data shown in the present manuscript have been generated by an in vitro experimental system, supplemented by an extremely thorough in silico metabolomic analysis. For this reason, future studies are warranted to translate these very interesting preliminary results in ex-vivo systems and to contextualize them in infection models.
Author Contributions
L.S., A.A., D.P. and E.P. have contributed significantly to this work including conceptualization, original draft, review, and editing. M.C., S.K. and F.D.S. contributed to final review and editing. L.S., N.P., F.R., D.P. and G.C. were responsible for methodology, laboratory investigation and research. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Not applicable.
Acknowledgments
The authors acknowledge the “Fondazione Cassa di Risparmio di Modena” for funding the HPLC-ESI/HRMS QExactive system at the Centro Interdipartimentale Grandi Strumenti (CIGS) of the University of Modena and Reggio Emilia.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Latif, A.; Shehzad, A.; Niazi, S.; Zahid, A.; Ashraf, W.; Iqbal, M.W.; Rehman, A.; Riaz, T.; Aadil, R.M.; Khan, I.M.; et al. Probiotics: Mechanism of Action, Health Benefits and Their Application in Food Industries. Front. Microbiol. 2023, 14, 1216674. [CrossRef]
- Ayivi, R.D.; Gyawali, R.; Krastanov, A.; Aljaloud, S.O.; Worku, M.; Tahergorabi, R.; Silva, R.C.D.; Ibrahim, S.A. Lactic Acid Bacteria: Food Safety and Human Health Applications. Dairy 2020, 1, 202–232. [CrossRef]
- Tang, C.; Lu, Z. Health Promoting Activities of Probiotics. J Food Biochem 2019, 43. [CrossRef]
- Huang, R.; Wu, F.; Zhou, Q.; Wei, W.; Yue, J.; Xiao, B.; Luo, Z. Lactobacillus and Intestinal Diseases: Mechanisms of Action and Clinical Applications. Microbiological Research 2022, 260, 127019. [CrossRef]
- Liang, B.; Xing, D. The Current and Future Perspectives of Postbiotics. Probiotics & Antimicro. Prot. 2023, 15, 1626–1643. [CrossRef]
- Alvarez-Sieiro, P.; Montalbán-López, M.; Mu, D.; Kuipers, O.P. Bacteriocins of Lactic Acid Bacteria: Extending the Family. Appl Microbiol Biotechnol 2016, 100, 2939–2951. [CrossRef]
- Murphy, E.F.; Clarke, S.F.; Marques, T.M.; Hill, C.; Stanton, C.; Ross, R.P.; O’Doherty, R.M.; Shanahan, F.; Cotter, P.D. Antimicrobials: Strategies for Targeting Obesity and Metabolic Health? Gut Microbes 2013, 4, 48–53. [CrossRef]
- Mosca, A.; Abreu Y Abreu, A.T.; Gwee, K.A.; Ianiro, G.; Tack, J.; Nguyen, T.V.H.; Hill, C. The Clinical Evidence for Postbiotics as Microbial Therapeutics. Gut Microbes 2022, 14, 2117508. [CrossRef]
- Alonso-Roman, R.; Last, A.; Mirhakkak, M.H.; Sprague, J.L.; Möller, L.; Großmann, P.; Graf, K.; Gratz, R.; Mogavero, S.; Vylkova, S.; et al. Lactobacillus rhamnosus Colonisation Antagonizes Candida albicans by Forcing Metabolic Adaptations That Compromise Pathogenicity. Nat Commun 2022, 13, 3192. [CrossRef]
- Thoda, C.; Touraki, M. Immunomodulatory Properties of Probiotics and Their Derived Bioactive Compounds. Applied Sciences 2023, 13, 4726. [CrossRef]
- Abdul Hakim, B.N.; Xuan, N.J.; Oslan, S.N.H. A Comprehensive Review of Bioactive Compounds from Lactic Acid Bacteria: Potential Functions as Functional Food in Dietetics and the Food Industry. Foods 2023, 12, 2850. [CrossRef]
- Spaggiari, L.; Sala, A.; Ardizzoni, A.; De Seta, F.; Singh, D.K.; Gacser, A.; Blasi, E.; Pericolini, E. Lactobacillus acidophilus, L. plantarum, L. rhamnosus, and L. reuteri Cell-Free Supernatants Inhibit Candida parapsilosis Pathogenic Potential upon Infection of Vaginal Epithelial Cells Monolayer and in a Transwell Coculture System In Vitro. Microbiol Spectr 2022, 10, e02696-21. [CrossRef]
- Doyle, C.; Cristofaro, V.; Sullivan, M.P.; Adam, R.M. Inosine - a Multifunctional Treatment for Complications of Neurologic Injury. Cell Physiol Biochem 2018, 49, 2293–2303. [CrossRef]
- Fan, S.; Kind, T.; Cajka, T.; Hazen, S.L.; Tang, W.H.W.; Kaddurah-Daouk, R.; Irvin, M.R.; Arnett, D.K.; Barupal, D.K.; Fiehn, O. Systematic Error Removal Using Random Forest for Normalizing Large-Scale Untargeted Lipidomics Data. Anal. Chem. 2019, 91, 3590–3596. [CrossRef]
- Züllig, T.; Zandl-Lang, M.; Trötzmüller, M.; Hartler, J.; Plecko, B.; Köfeler, H.C. A Metabolomics Workflow for Analyzing Complex Biological Samples Using a Combined Method of Untargeted and Target-List Based Approaches. Metabolites 2020, 10, 342. [CrossRef]
- Shafy, A.; Molinié, V.; Cortes-Morichetti, M.; Hupertan, V.; Lila, N.; Chachques, J.C. Comparison of the Effects of Adenosine, Inosine, and Their Combination as an Adjunct to Reperfusion in the Treatment of Acute Myocardial Infarction. ISRN Cardiology 2012, 2012, 1–9. [CrossRef]
- Schymanski, E.L.; Jeon, J.; Gulde, R.; Fenner, K.; Ruff, M.; Singer, H.P.; Hollender, J. Identifying Small Molecules via High Resolution Mass Spectrometry: Communicating Confidence. Environ. Sci. Technol. 2014, 48, 2097–2098. [CrossRef]
- Sanders, M.E.; Merenstein, D.J.; Reid, G.; Gibson, G.R.; Rastall, R.A. Probiotics and Prebiotics in Intestinal Health and Disease: From Biology to the Clinic. Nat Rev Gastroenterol Hepatol 2019, 16, 605–616. [CrossRef]
- De Almada, C.N.; De Almada, C.N.; De Souza Sant’Ana, A. Paraprobiotics as Potential Agents for Improving Animal Health. In Probiotics and Prebiotics in Animal Health and Food Safety; Di Gioia, D., Biavati, B., Eds.; Springer International Publishing: Cham, 2018; pp. 247–268 ISBN 978-3-319-71948-1.
- Vallejo-Cordoba, B.; Castro-López, C.; García, H.S.; González-Córdova, A.F.; Hernández-Mendoza, A. Postbiotics and Paraprobiotics: A Review of Current Evidence and Emerging Trends. In Advances in Food and Nutrition Research; Elsevier, 2020; Vol. 94, pp. 1–34 ISBN 978-0-12-820218-0.
- Martín, R.; Langella, P. Emerging Health Concepts in the Probiotics Field: Streamlining the Definitions. Front. Microbiol. 2019, 10, 1047. [CrossRef]
- Zhao, X.; Zhong, X.; Liu, X.; Wang, X.; Gao, X. Therapeutic and Improving Function of Lactobacilli in the Prevention and Treatment of Cardiovascular-Related Diseases: A Novel Perspective From Gut Microbiota. Front. Nutr. 2021, 8, 693412. [CrossRef]
- Peng, H.; Zhou, G.; Yang, X.-M.; Chen, G.-J.; Chen, H.-B.; Liao, Z.-L.; Zhong, Q.-P.; Wang, L.; Fang, X.; Wang, J. Transcriptomic Analysis Revealed Antimicrobial Mechanisms of Lactobacillus rhamnosus SCB0119 against Escherichia coli and Staphylococcus aureus. Int J Mol Sci 2022, 23, 15159. [CrossRef]
- Iglesias, M.B.; Viñas, I.; Colás-Medà, P.; Collazo, C.; Serrano, J.C.E.; Abadias, M. Adhesion and Invasion of Listeria monocytogenes and Interaction with Lactobacillus rhamnosus GG after Habituation on Fresh-Cut Pear. Journal of Functional Foods 2017, 34, 453–460. [CrossRef]
- Muyyarikkandy, M.S.; Amalaradjou, M. Lactobacillus Bulgaricus, Lactobacillus rhamnosus and Lactobacillus paracasei Attenuate Salmonella enteritidis, Salmonella Heidelberg and Salmonella typhimurium Colonization and Virulence Gene Expression In Vitro. IJMS 2017, 18, 2381. [CrossRef]
- Burkholder, K.M.; Bhunia, A.K. Salmonella enterica Serovar typhimurium Adhesion and Cytotoxicity during Epithelial Cell Stress Is Reduced by Lactobacillus rhamnosus GG. Gut Pathog 2009, 1, 14. [CrossRef]
- Coman, M.M.; Verdenelli, M.C.; Cecchini, C.; Silvi, S.; Orpianesi, C.; Boyko, N.; Cresci, A. In Vitro Evaluation of Antimicrobial Activity of Lactobacillus rhamnosus IMC 501 ® , Lactobacillus paracasei IMC 502 ® and SYNBIO ® against Pathogens. J Appl Microbiol 2014, 117, 518–527. [CrossRef]
- Song, Y.-G.; Lee, S.-H. Inhibitory Effects of Lactobacillus rhamnosus and Lactobacillus casei on Candida Biofilm of Denture Surface. Archives of Oral Biology 2017, 76, 1–6. [CrossRef]
- Chew, S.Y.; Cheah, Y.K.; Seow, H.F.; Sandai, D.; Than, L.T.L. Probiotic Lactobacillus rhamnosus GR -1 and Lactobacillus reuteri RC -14 Exhibit Strong Antifungal Effects against Vulvovaginal Candidiasis-causing Candida glabrata Isolates. J Appl Microbiol 2015, 118, 1180–1190. [CrossRef]
- Alseth, I.; Dalhus, B.; Bjørås, M. Inosine in DNA and RNA. Curr Opin Genet Dev 2014, 26, 116–123. [CrossRef]
- Licht, K.; Hartl, M.; Amman, F.; Anrather, D.; Janisiw, M.P.; Jantsch, M.F. Inosine Induces Context-Dependent Recoding and Translational Stalling. Nucleic Acids Res 2019, 47, 3–14. [CrossRef]
- Benowitz, L.I.; Jing, Y.; Tabibiazar, R.; Jo, S.A.; Petrausch, B.; Stuermer, C.A.; Rosenberg, P.A.; Irwin, N. Axon Outgrowth Is Regulated by an Intracellular Purine-Sensitive Mechanism in Retinal Ganglion Cells. J Biol Chem 1998, 273, 29626–29634. [CrossRef]
- Irwin, N.; Li, Y.-M.; O’Toole, J.E.; Benowitz, L.I. Mst3b, a Purine-Sensitive Ste20-like Protein Kinase, Regulates Axon Outgrowth. Proc Natl Acad Sci U S A 2006, 103, 18320–18325. [CrossRef]
- Benowitz, L.I.; Goldberg, D.E.; Madsen, J.R.; Soni, D.; Irwin, N. Inosine Stimulates Extensive Axon Collateral Growth in the Rat Corticospinal Tract after Injury. Proc Natl Acad Sci U S A 1999, 96, 13486–13490. [CrossRef]
- Muto, J.; Lee, H.; Lee, H.; Uwaya, A.; Park, J.; Nakajima, S.; Nagata, K.; Ohno, M.; Ohsawa, I.; Mikami, T. Oral Administration of Inosine Produces Antidepressant-like Effects in Mice. Sci Rep 2014, 4, 4199. [CrossRef]
- Haskó, G.; Sitkovsky, M.V.; Szabó, C. Immunomodulatory and Neuroprotective Effects of Inosine. Trends Pharmacol Sci 2004, 25, 152–157. [CrossRef]
- Haskó, G.; Kuhel, D.G.; Németh, Z.H.; Mabley, J.G.; Stachlewitz, R.F.; Virág, L.; Lohinai, Z.; Southan, G.J.; Salzman, A.L.; Szabó, C. Inosine Inhibits Inflammatory Cytokine Production by a Posttranscriptional Mechanism and Protects Against Endotoxin-Induced Shock. The Journal of Immunology 2000, 164, 1013–1019. [CrossRef]
- Liaudet, L.; Mabley, J.G.; Soriano, F.G.; Pacher, P.; Marton, A.; Haskó, G.; Szabó, C. Inosine Reduces Systemic Inflammation and Improves Survival in Septic Shock Induced by Cecal Ligation and Puncture. Am J Respir Crit Care Med 2001, 164, 1213–1220. [CrossRef]
Figure 1.
Principal Component Analysis (PCA)-2 of the metabolome from L. RHA, L. AC, L. PLA and L. REU analyzing in both positive and negative ionization mode. Data are from triplicate samples from 3 different experiments.
Figure 1.
Principal Component Analysis (PCA)-2 of the metabolome from L. RHA, L. AC, L. PLA and L. REU analyzing in both positive and negative ionization mode. Data are from triplicate samples from 3 different experiments.
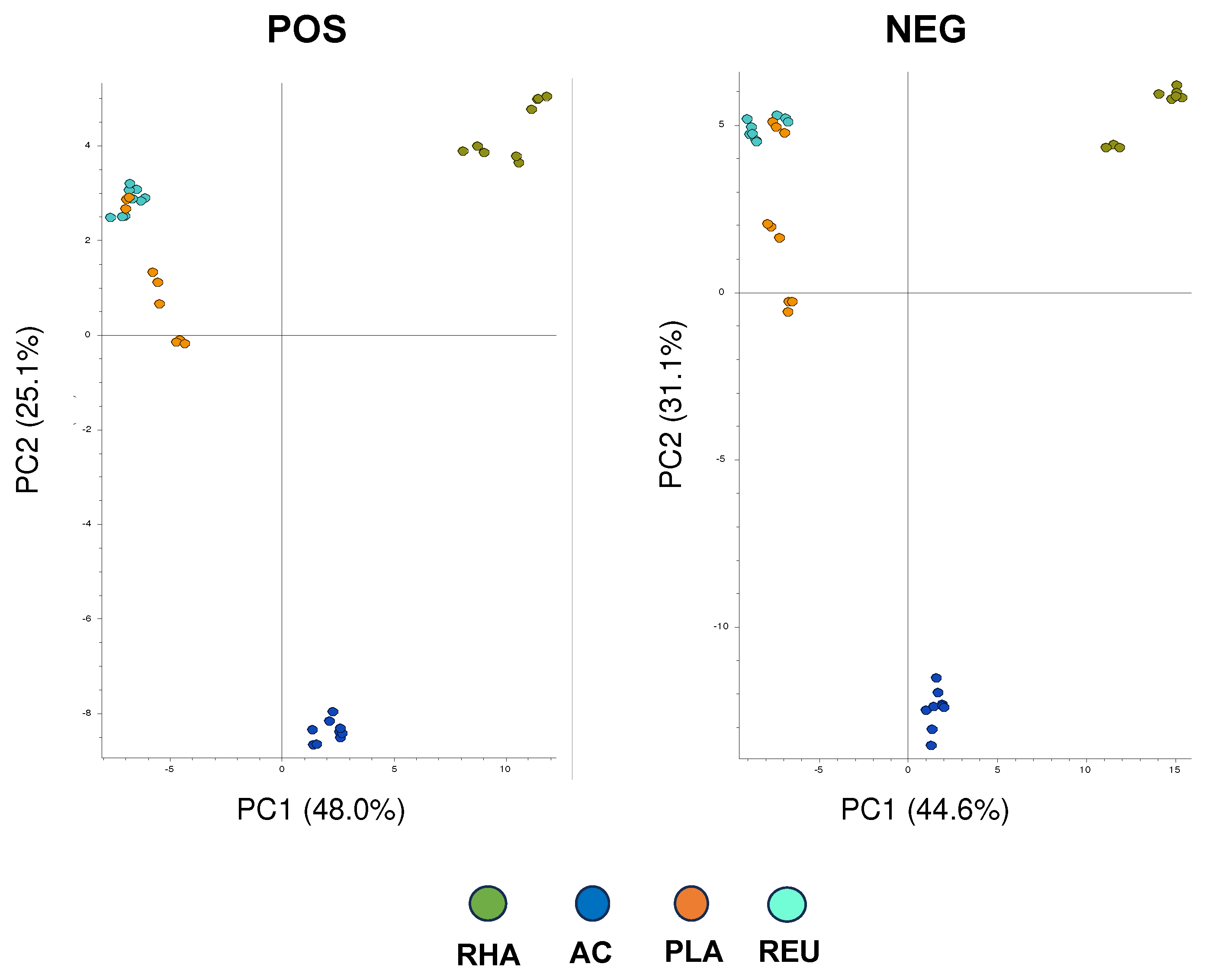
Figure 2.
Hierarchical clustering analysis in positive and negative ionization mode, carried out to compare the metabolome from L. RHA, L. AC, L. PLA and L. REU according to Compound Discoverer (CD) 3.3.2.31 analysis. Red line highlights the distinct cluster of metabolites overexpressed in the CFS from L. RHA. Data are from triplicate samples from 3 different experiments.
Figure 2.
Hierarchical clustering analysis in positive and negative ionization mode, carried out to compare the metabolome from L. RHA, L. AC, L. PLA and L. REU according to Compound Discoverer (CD) 3.3.2.31 analysis. Red line highlights the distinct cluster of metabolites overexpressed in the CFS from L. RHA. Data are from triplicate samples from 3 different experiments.
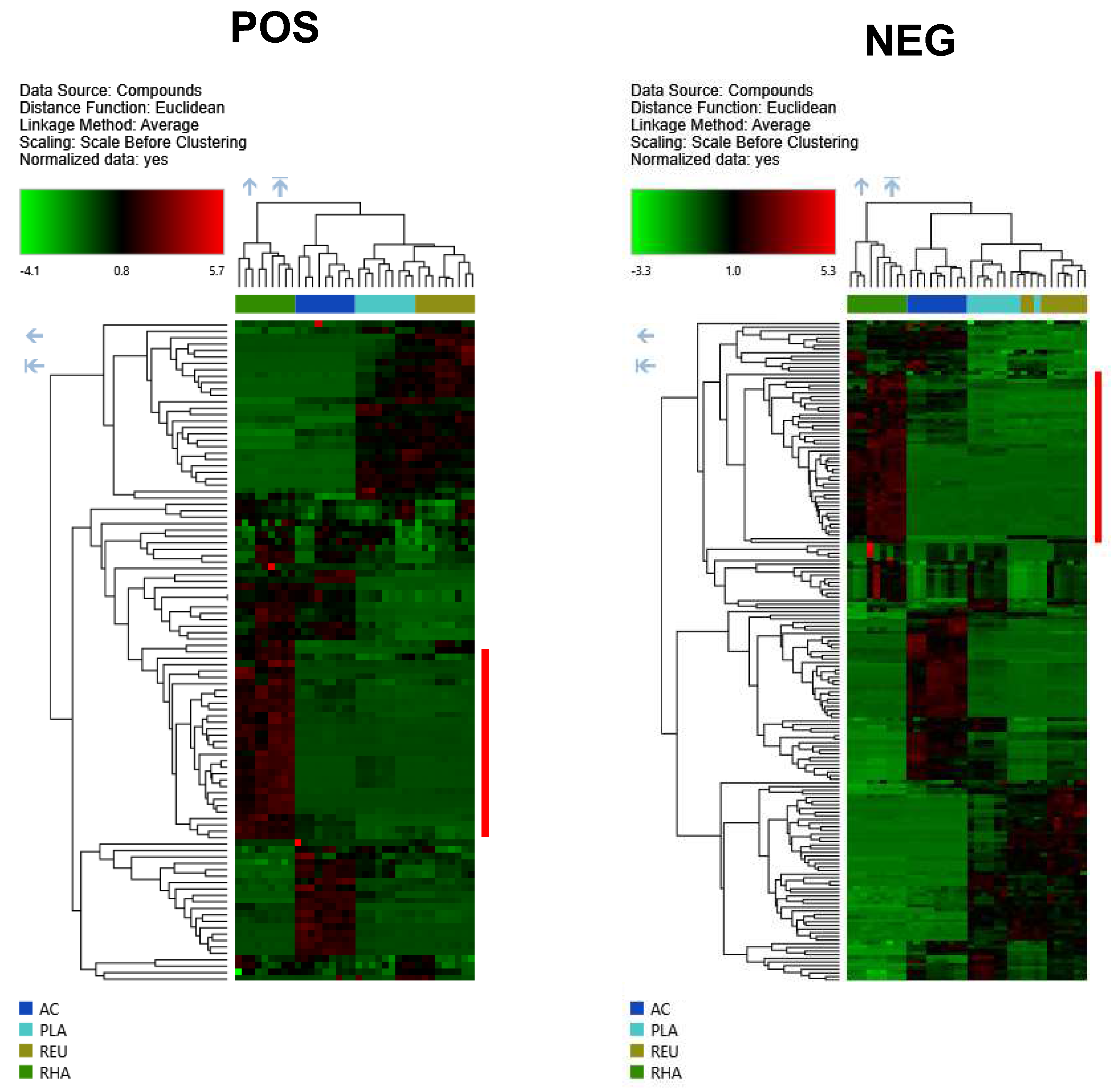
Figure 3.
Extracted ion chromatogram of inosine (C10H12N4O5) theoretical [M+H]+ molecular ion at m/z=269.08805 (± 5ppm) in (a) L. RHA CFS, (b) L. RHA CFS spiked with inosine and (c) inosine standard solution.
Figure 3.
Extracted ion chromatogram of inosine (C10H12N4O5) theoretical [M+H]+ molecular ion at m/z=269.08805 (± 5ppm) in (a) L. RHA CFS, (b) L. RHA CFS spiked with inosine and (c) inosine standard solution.
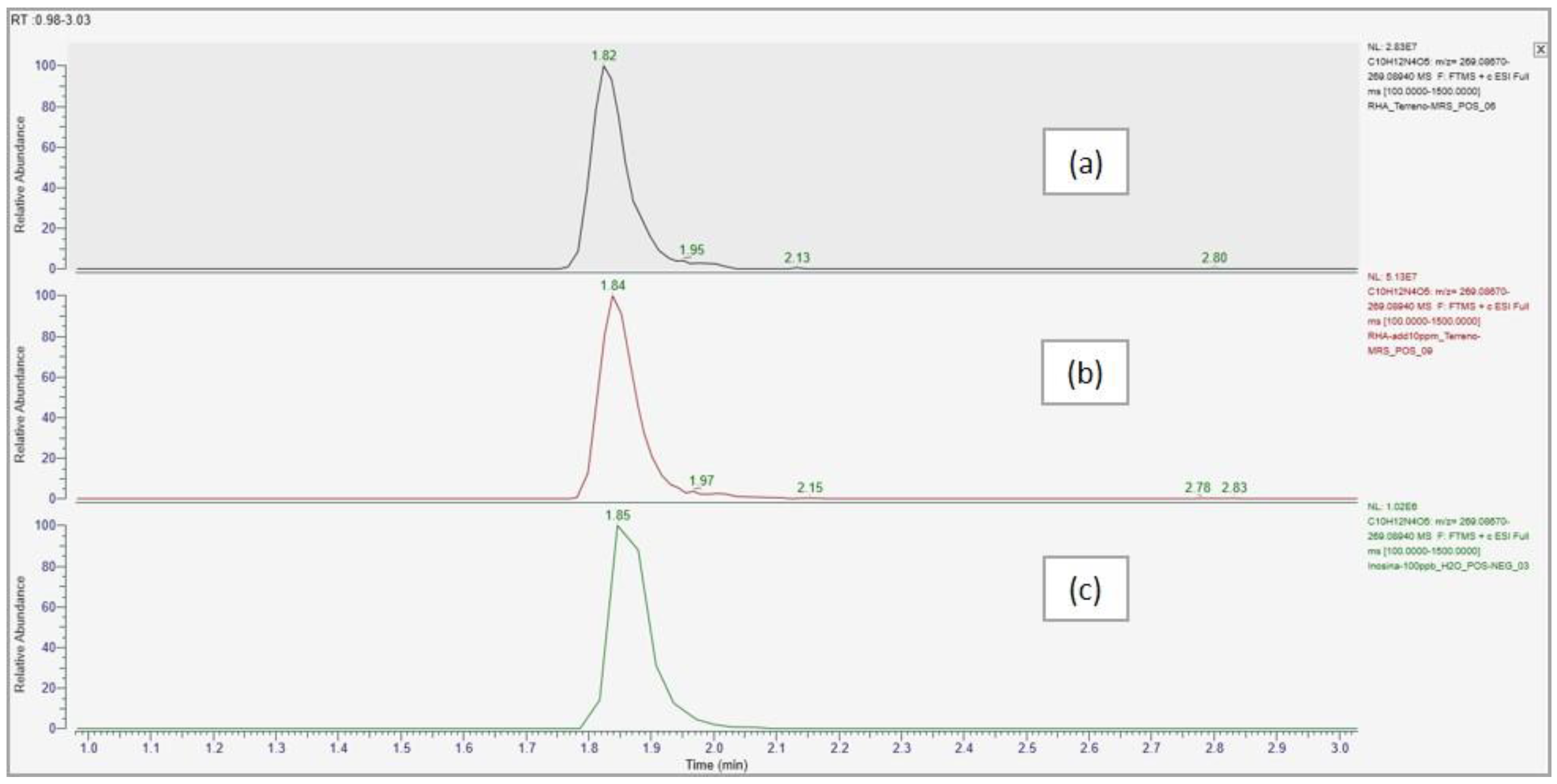
Figure 4.
Comparison of HCD fragmentation spectra of m/z=269.08805 precursor ion at 1.83 min in (a) inosine standard solution and (b) L. RHA CFS.
Figure 4.
Comparison of HCD fragmentation spectra of m/z=269.08805 precursor ion at 1.83 min in (a) inosine standard solution and (b) L. RHA CFS.
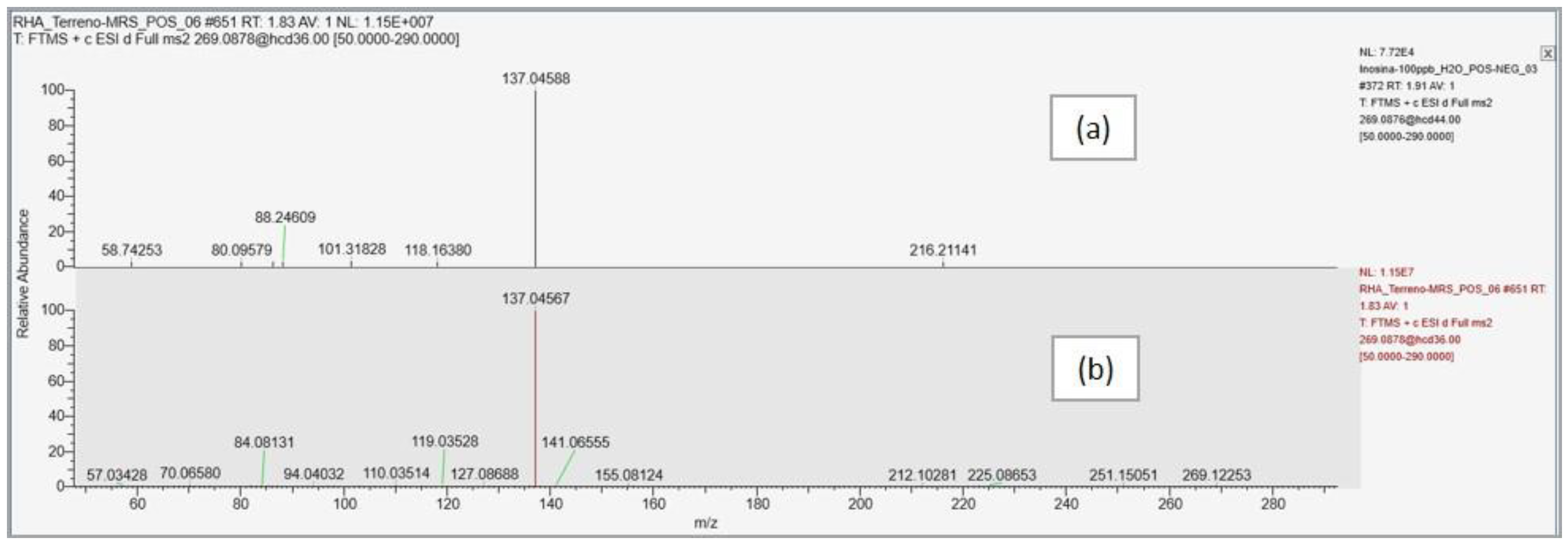

Table 1.
Inosine identification profile according to Compound Discoverer (CD) 3.3.2.31 analysis. Data are from triplicate samples from 3 different experiments.
Table 1.
Inosine identification profile according to Compound Discoverer (CD) 3.3.2.31 analysis. Data are from triplicate samples from 3 different experiments.
| Compound name | Predicted composition | mzCloud Search |
mzValue Search |
Metabolika Search | ChemSpider Search |
MassList Search |
|---|---|---|---|---|---|---|
| Inosine | Full Match | Full Match | Full Match | Full Match | Partial Match | Partial Match |
Table 2.
Inosine identified pathways according to Compound Discoverer (CD) 3.3.2.31 analysis. Data are from triplicate samples from 3 different experiments.
Table 2.
Inosine identified pathways according to Compound Discoverer (CD) 3.3.2.31 analysis. Data are from triplicate samples from 3 different experiments.
| Compound name | Formula | n° identified pathways |
Pathways | Mapped compounds |
Matched compounds |
Compounds in pathways |
|---|---|---|---|---|---|---|
| Inosine | C10H12N4O5 | 1 | Superpathway of purine nucleotide salvage | 14 | 10 | 54 |
| 2 | Purine nucleotides degradation II (aerobic) | 12 | 8 | 27 | ||
| 3 | Purine nucleotides degradation I (plants) | 10 | 7 | 23 | ||
| 4 | Superpathway of purine degradation in plants | 10 | 7 | 34 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



