
Submitted:
10 January 2024
Posted:
11 January 2024
You are already at the latest version
Abstract
RNA-viruses of the genera Ambivirus, Mitovirus, Sclerotimonavirus, and Partitivirus were found in a single isolate of Fusarium graminearum. The genomes of the mitovirus, sclerotimonavirus, and partitivirus were assigned to previously described viruses, whereas the ambivirus genome putatively represents a new species, named Fusarium graminearum ambivirus 1 (FgAV1). To investigate the effect of mycoviruses on the fungal phenotype, spontaneous loss of mycoviruses during meiosis and transmission of mycoviruses into a new strain by anastomosis were used to obtain isogenic F. graminearum strains with and without mycoviruses. Notable effects observed in mycovirus-harboring strains were (i) suppression of the synthesis of trichothecene mycotoxins and their precursor trichodiene, (ii) suppression of the synthesis of the defense compound aurofusarin, (iii) stimulation of the emission of 2-methyl-1-butanol and 3-methyl-1-butanol, and (iv) increased attractiveness of fungal mycelia for fungivorous collembolans. Increased attractiveness of mycovirus-infected filamentous fungi to animal predators opens new perspectives on the ecological implications of infection of fungi with viruses.
Keywords:
mycovirus
; Fusarium graminearum
; trichothecenes
; deoxynivalenol
; aurofusarin
; VOC
; Collembola
; fungivory
; food preference
; Folsomia candida
1. Introduction
Gibberella ear rot, caused by the pathogen Fusarium graminearum, is a widely spread disease of maize causing severe yield losses and grain quality decline [1]. Contamination of grain with mycotoxins of Fusarium spp. threatens food and feed safety [2,3,4]. Ears infected with F. graminearum often contain deoxynivalenol (DON) [5] or nivalenol (NIV) [6] and their acetylated derivatives, which act as virulence factors [7] by suppressing plant defense responses [8]. Furthermore, the pathogen produces the estrogenic metabolite zearalenone [9], which protects the fungus against mycoparasites [10], and the bis-naphthopyrone pigment aurofusarin [11], which protects the fungus against predators [12].
Viruses infecting fungi (mycoviruses) have been described in most fungal taxa [13,14]. The genome of mycoviruses typically consists of single-stranded or double-stranded RNA, less often of DNA. Despite a high level of genomic and structural diversity, most viruses can be taxonomically classified based on conserved motifs of their RNA-dependent RNA polymerase (RdRp) [15]. Mycoviruses are transmitted by cell fusion (hyphal anastomosis) or sporulation. Dissemination of mycoviruses by fungivores has been documented in a single case so far [16].
The most extensively studied effect of mycoviruses on fungal phenotype is the suppression of aggressiveness in plant pathogens, termed hypovirulence [17,18], which gained attention due to its potential for biological control of plant diseases [19,20]. F. graminearum is an economically important pathogen and producer of mycotoxins with significant impact on food safety. Examples of mycoviruses inducing hypovirulence in F. graminearum are the dsRNA virus FgV1 [21], F. gramineaum virus china-9 (FgV-ch9) [22], and F. graminearum hypovirus 2 (FgHV2) [23]. The cause of the suppression of aggressiveness in F. graminearum by mycoviruses is unknown. Studies in fungi other than Fusarium spp. showed that mycoviruses may also increase the aggressiveness of plant pathogens, which is called hypervirulence [14,24].
Few studies have so far investigated the effect of mycoviruses on mycotoxin production. The first mycovirus found in F. graminearum was reported to cause a 60-fold decrease in the production of trichothecenes [21]. The claim was perpetuated in literature, though the "60-fold decrease" was due to a calculation error (see Section 3.4.), and suppression of trichothecene synthesis was not proven. Twelve years later, the same laboratory showed that FgV1 suppressed agmatine-induced synthesis of trichothecenes in Fusarium gramineaum, while three other mycoviruses did not substantially affect trichothecene synthesis [25]. By contrary, two studies reported stimulation of mycotoxin synthesis by mycoviruses. According to the first study, transfection of Aspergillus ochraceus with a partitivirus from a different strain stimulated the production of ochratoxin A up to 3-fold [26]. Mycotoxin levels were normalized to fungal biomass, determined as weight of mycelia harvested from liquid media. The second study reported that mycoviral infection triggered the synthesis of tenuazonic acid (TnA) in Magnaporthe oryzae [27]. The finding was surprising because M. oryzae, which is the causal agent of rice blast disease, was not known to produce TnA. TnA amount in fungal cultures were not normalized to fungal biomass, but the intensities of other HPLC signals in extracts of cultures with and without the mycovirus were comparable, and TnA in strains without mycoviruses was undetectable, corroborating a casual relationship between mycotoxin infection and TnA production.
Except for a report on dissemination of a mycovirus by a mycophagous predator [16], however limited to a single system and not yet independently reproduced, nothing is known about the effect of mycoviruses on interactions of fungi with their predators. It is plausible to hypothesize that selection favors mycoviruses with the ability to render their hosts less attractive to predators, except for predators transmitting mycoviruses.
In this study, we report on a F. graminearum strain infected with four mycoviruses; we characterize a new member of the genus ambivirus; and we investigated the effect of mycoviruses on the radial growth of fungal colonies, production of volatile and nonvolatile secondary metabolites, and the attractiveness of F. graminearum to the fungivorous arthropod Folsomia candida.
2. Results
2.1. Characterization of F. graminearum 266MV carrying mycoviruses and its descendants that lost mycoviruses by mitotic segregation
Strain F. graminearum 266.1 was isolated from a maize ear from South Germany. The strain was putatively identified as a member of the F. graminearum species-complex based on the morphology of macroconidia and the deep red color of colonies on agar plates. The taxonomic assignment was supported by melting profiles of amplified fragments of genes encoding for the second largest subunit of RNA polymerase II (RPB2) and translation elongation factor 1 alpha (TEF-1α) [28] and by sequencing a 503 bp fragment of the TEF-1α gene according to O’Donnell et al. [29]. The sequence was deposited in NCBI under accession number OP035498. The strain was later found to carry mycoviruses (see below), and therefore, it was renamed as F. graminearum 266MV.
Agarose electrophoresis of total nucleic acids extracted from strain 266MV revealed the presence of an extrachromosomal element of approximately 2.5 kb (Figure 1, lane 4). Treatment with DNase I completely removed genomic DNA, but it left small RNAs and the 2.5 kb fragment unaffected, indicating that the 2.5 kb fragment consisted of RNA (Figure 1, lane 5).
We hypothesized that the 2.5 kb RNA fragment originated from one or more mycoviruses. To generate a mycovirus-free strain and investigate the effect of putative mycoviruses on the host, we attempted to cure the strain by treatments with cycloheximide and ribavirin, however with no success (data not shown). Therefore, we grew strain 266MV on PDA, purified about 200 single-spore isolates, extracted their nucleic acids, and screened the extracts by agarose electrophoresis for the loss of the extrachromosomal element. Indeed, one strain lost the extrachromosomal element visible in agarose electrophoresis (Figure 2C), and it was designated as F. graminearum 266.
On PDA plates, strain 266MV exhibited an irregular growth pattern and dark red pigmentation, while strain 266 formed fluffy colonies with smooth margin. The color of colonies of strain 266 was less intense, similar to most F. graminearum strains in our collection (Figure 2A,B).
2.2. Confirmation of the presence of mycoviruses by RNA sequencing and electron microscopy
Sequencing of dsRNA extracted from strain 266MV revealed the presence of 5 viral RNAs, constituting the genomes of 4 distinct mycoviruses. The MiSeq run generated a total of 4,090,354 reads. After quality trimming, 3,692,466 reads were left. Normalized reads were mapped against Fusarium graminearum PH-1 genome/transcriptome accessions (chromosomes 1 to 4, 13401 predicted transcripts and mitochondrion sequences; BioProject PRJNA243 and NC_009493), and the remaining 17,641 reads were de novo assembled into contigs. Complete genome sequences were determined by RACE and deposited in NCBI GenBank (Table 1). The BLAST alignment against the virus reference databases revealed 5 hits with various degrees of identity to genomes of mycoviruses (Table 1). Two sequences were assigned to RNA 1 and RNA 2 of Fusarium poae virus 1 (FpV1), with a high percentage of identity to the nuclear core sequences from the primary host species. Icosahedral particles about 35-40 nm in diameter, visible by electron microscopy of mycelial extracts, confirmed the infection with FpV1 (Figure 3A). This suggests horizontal transmission of the mycovirus between F. poae and F. graminearum. The genomic segments of FpV1 were not further characterized, given their high similarity to reference sequences.
Two sequences showed up to 90% identity to Soybean leaf-associated negative-stranded RNA virus and Cryphonectria parasitica ambivirus 1, and they were tentatively assigned to Fusarium graminearum negative-stranded RNA virus 2 (FgSV2) and Fusarium graminearum ambivirus 1 (FgAV1), according to the guidelines of the International Committee on Taxonomy of Viruses (ICTV). A sequence of 2,606 nt showed 96% identity with a Plasmopara viticola lesion-associated mitovirus 13. The genomes of these three mycoviruses has been characterized in detail and is presented in section 2.3.
RT-PCR with virus-specific primers (Section 5.2., Appendix Table A1) detected all 5 segments in RNA-extracts from strain 266MV yet failed to detect them in strain 266.
2.3. Characterization of mycoviral genomes
2.3.1. Fusarium graminearum ambivirus 1 (FgAV1)
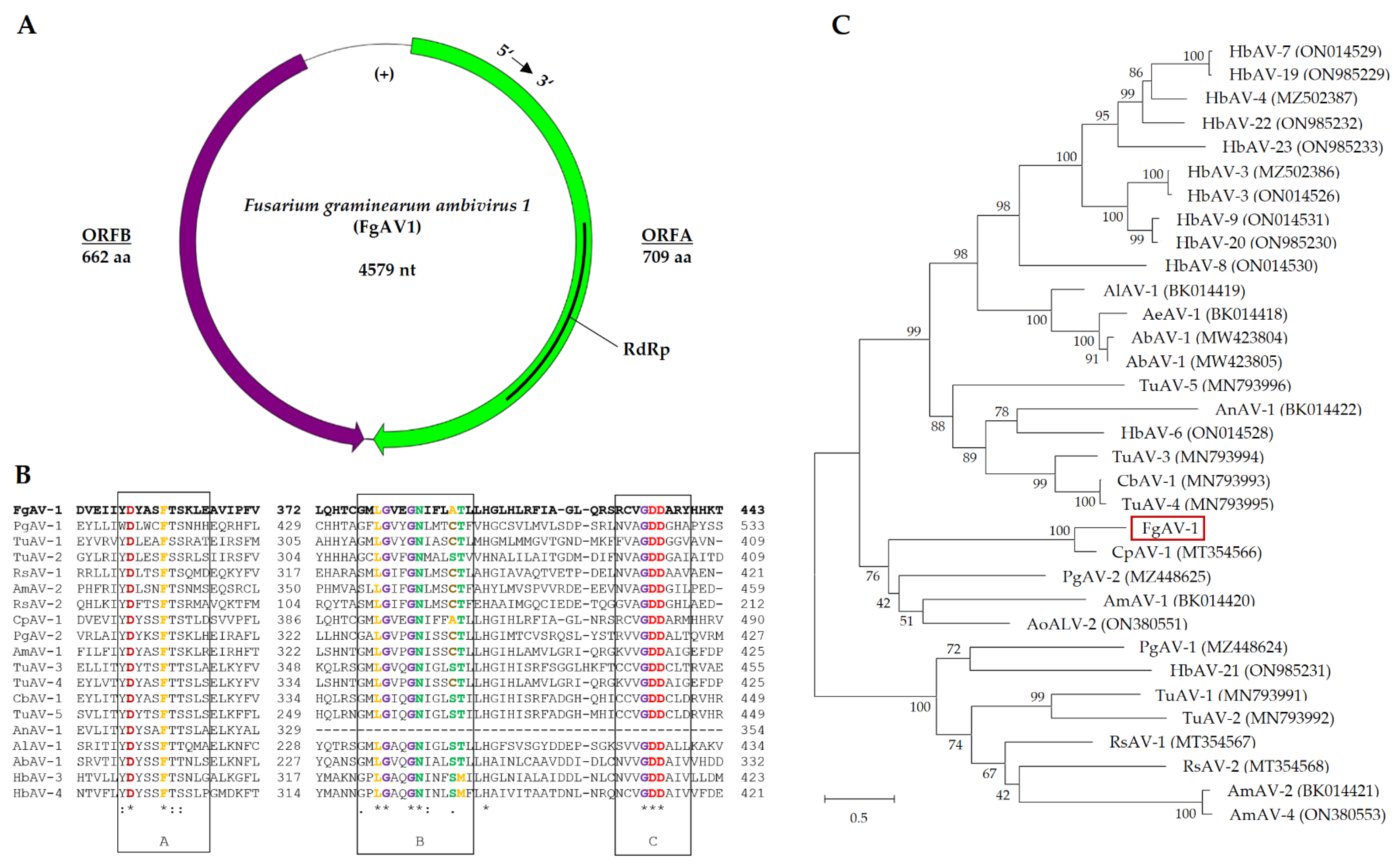
The genome of Fusarium graminearum ambivirus 1 (FgAV1) with a length of 4,579 nt exhibited a circular ssRNA structure with two open reading frames in opposite directions: ORF A in 5’-3’ orientation and ORF B in 3’-5’ orientation (Figure 4), as the typical feature of ambiviruses. ORF A encodes a protein of 709 amino acids (aa), predicted to be the RNA-dependent RNA polymerase (RdRp), while ORF B codes for a protein of 662 aa, with unknown function. FgAV1 shares low sequence similarity with the sequences in public databases, with highest similarity (73 %) to Cryphonectria parasitica ambivirus 1 (MT354566). The genome structure and phylogenetic analysis of the putative RdRp protein are shown in Figure 4.
The replication of ambiviruses has only been recently unraveled. In their seminal paper, Forgia et al. [30] showed that ambiviral genomes replicate by a rolling-circle mechanism, which has so far been known mainly from much smaller replicons of viroids. In contrast to viroids, replication of ambiviruses is catalyzed by their own RdRp. Concatenated genomes produced by rolling-circle replication are dissected by the action of two antiparallel hammerhead ribozymes. Predicted secondary structures of the ribozymes of FgAV1 and their cleavage sites, kindly provided by Dr. Beatriz Navarro and Dr. Francesco Di Serio (Istituto per la Protezione Sostenibile delle Piante, CNR, Bari, Italy) are shown in Appendix Figure A2.
Figure 4.
Genome organization of Fusarium graminearum ambivirus 1 (FgAV1). (A) Schematic representation of the circular genome. Open reading frames A and B are represented by purple and green colors, respectively. (B) Alignment of RdRp sequences; conserved motifs according to [31] are framed in black boxes. Amino acid sequences were selected from the accessions used for the generation of the dendrogram in part C (Appendix Table A2). The alignment was created with Clustal Omega [32]. Asterisks (*) mark positions with fully conserved amino acid residues, colons (:) residues with highly similar physicochemical properties that score > 0.5 in PAM 250 [33], and periods (.) residue scoring 0.0 to 0.5 in PAM 250. (C) Dendrogram of putative RdRp sequence of FgAV1, other ambiviruses, and ambivirus-like sequences. The dendrogram was constructed using the Neighbor-Joining method [34] implemented in MEGA X 10.1.8 [35]. Bootstrap values in % (1000 replicas) are shown. The tree is drawn to scale, with branch lengths corresponding to evolutionary distances, which were estimated using a JTT matrix-based method [36]. The scale is in substitutions per site.
Figure 4.
Genome organization of Fusarium graminearum ambivirus 1 (FgAV1). (A) Schematic representation of the circular genome. Open reading frames A and B are represented by purple and green colors, respectively. (B) Alignment of RdRp sequences; conserved motifs according to [31] are framed in black boxes. Amino acid sequences were selected from the accessions used for the generation of the dendrogram in part C (Appendix Table A2). The alignment was created with Clustal Omega [32]. Asterisks (*) mark positions with fully conserved amino acid residues, colons (:) residues with highly similar physicochemical properties that score > 0.5 in PAM 250 [33], and periods (.) residue scoring 0.0 to 0.5 in PAM 250. (C) Dendrogram of putative RdRp sequence of FgAV1, other ambiviruses, and ambivirus-like sequences. The dendrogram was constructed using the Neighbor-Joining method [34] implemented in MEGA X 10.1.8 [35]. Bootstrap values in % (1000 replicas) are shown. The tree is drawn to scale, with branch lengths corresponding to evolutionary distances, which were estimated using a JTT matrix-based method [36]. The scale is in substitutions per site.
2.3.2. Fusarium graminearum negative-stranded RNA virus 2 (FgNSRV2)
The genome sequence assigned to Fusarium graminearum negative-stranded RNA virus 2 is 8,939 nt long and contains five putative ORFs (Figure 5). The 3’- and 5´- UTRs are 127 and 5 nt long, respectively. The fourth and largest ORF encodes a putative protein of 1,941 aa with a RdRp domain (pfam00946), an mRNA-capping region (pfam14318), and a methyltransferase domain (cl27811). Phylogenetic analysis of ORF 4 (Figure 5) placed FgNSRV2 in the order Mononegavirales, family Mymonaviridae, genus Sclerotimonavirus. The ORF 1 of FgNSRV2 encodes a 274 aa long protein that contains three conserved regions, which were putatively identified as a helix-rich domain (TIGR04523), an ATPase (COG1196), and a phosphatidylglycerophosphatase domain (PRK03918) according to NCBI Conserved Domain Database (CDD). Comparative sequence analysis with BLASTp showed that ORF 1 shared high homology (>75%) with proteins encoded by Soybean leaf-associated negative-stranded RNA virus 1 (ALM62222) and Fusarium graminearum negative-stranded RNA virus 1 (UNG44333), which belong to the family Mymonaviridae. No conserved domain was found in ORF 2 (388 aa), ORF 3 (56 aa), nor ORF 5 (195 aa). However, ORF2 exhibits a high level of sequence similarity to other members of the order Mymonaviruses from Fusarium spp., including ATP75709 of F. graminearum, UNG44330 of F. asiaticum, and UWK02084 of F. proliferatum.
The sequence of FgNSRV2 contains a conserved gene-junction section, which is commonly found and regarded as a characteristic feature in mononegaviruses [38,39]. An alignment of the gene-junction sequence of FgNSRV2 with sequences from other known Mononegaviruses is presented in Figure 6. So far, only a few exceptions with 6 rather than 5 genome ORFs were reported in Mononegaviruses [39]. One gene-junction section per ORF was reported for SsNSRV-1, whereas 4 junctions were found in FgNSRV1 and FgNSRV2.
2.3.3. Fusarium graminearum mitovirus 2 (FgMV2)
The genome of Fusarium graminearum mitovirus 2 (FgMV2) has a length of 2,634 nt (Figure 7). The 5’-UTR and 3’-UTR sequences are 194 nt and 169 nt long. A single ORF, which encodes a protein of 757 aa (L-Protein), was found using the mitochondrial genetic code. In the same way as described for a mycovirus of Sclerotinia sclerotiorum [41], FgMV2 can be assigned to Class II mitoviruses. The genome of Plasmopara viticola lesion associated mitovirus 13 (MN539774) is the most similar to FgMV2, sharing 96.3 % similarity (Appendix Table A2). Investigation of secondary structures with RNAfold version 2.6.3 [42] revealed stem-loop structures characteristic of mitoviruses [43], which are believed to protect ssRNA genomes from enzymatic degradation (Appendix Figure A3).
2.4. Transfection of mycoviruses from strain 266MV into mycovirus-free strain and the effect of mycoviruses on radial growth and sporulation
Transfection of mycoviruses from strain 266MV to strain 668 (Table 3) was carried out by coincubation of the strains on agar plates (Appendix Figure A1). The selection of transfectants was facilitated by a resistance marker carried by the acceptor strain (Section 5.4.) Two transfected strains were verified by the presence of viral dsRNA fragments in agarose electrophoreses. In case of FpV1, the transmission was also verified by the visualization of viral particles by electron microscopy (Figure 3B).
Strain 266MV harbored four mycoviruses (Table 1). To find out which mycoviruses have been transfected into strain 668MV and also which mycoviruses were still present in the cured strain 266, we tested all strains used in this study for the presence of each mycovirus by RT PCR (Section 4.2.) with mycovirus-specific primers (Table A1, Table 2). Differences among the strains in mycoviral load allowed us to correlate phenotypic features with mycoviral infection.
Radial growth of the strain was measured at 25 °C (Section 4.5.). Strain 266MV exhibited an irregular growth pattern with altered pigmentation (Figure 2B), but no effect of mycoviral infection on the radial growth rate was observed (Figure 8A). In contrast, the radial growth rate of strain 668MV was reduced by about 28% as compared to strain 668. Colonies of the transfected strain 668MV were deeply red, similar to colonies of strain 266MV (Figure 2A,B).
The capacity of all four strains to produce conidia in liquid SNA media varied to a large extent, as can be seen in Figure 8B,D, though parallel cultures were grown under identical conditions. No differences in the sporulation between strains 266 and 266MV, nor between 668 and 668MV were observed.
2.5. Effect of mycoviral infection on the production of secondary metabolites
2.5.1. Production of trichothecenes and aurofusarin
To investigate the effect of infection with mycoviruses on the capability of F. graminearum to produce trichothecenes and aurofusarin, strains 266, 266MV, 668, and 668MV were cultivated on rice kernels. Nine replicates of strains 266 and 266MV and 8 replicates of strains 668 and 668MV were processed. Concentration of aurofusarin, DON, 3ADON, and 15ADON were determined by HPLC-MS/MS (Section 5.6.), and fungal DNA was quantified by qPCR (Section 5.3.). The concentrations of secondary metabolites were then normalized to fungal DNA to compensate for differences in fungal biomass accumulated by strains infected with mycoviruses and mycovirus-free strains.
In five out of six combinations of strain pairs and trichothecenes, lower amounts of trichothecenes were found in cultures infected with mycoviruses (Figure 9). The suppression of trichothecene synthesis was substantial and statistically highly significant, except for 15ADON in strain pair 668/668MV.
The accumulation of aurofusarin was suppressed in strain 668MV as compared to 668, but it remained unchanged in strain 266MV as compared to 266. Of note, cultures of strain 668 accumulated more aurofusarin (2.2 to 4.5 g/kg) than cultures of strain 266 (1.0 to 3.5 g/kg). This can likely be explained by the disruption of zearalenone synthesis in strain 668 (Section 4.1), which increased the pool of acetyl-CoA available for the synthesis of aurofusarin.
2.5.2. Effect of mycoviruses on the production of volatile compounds
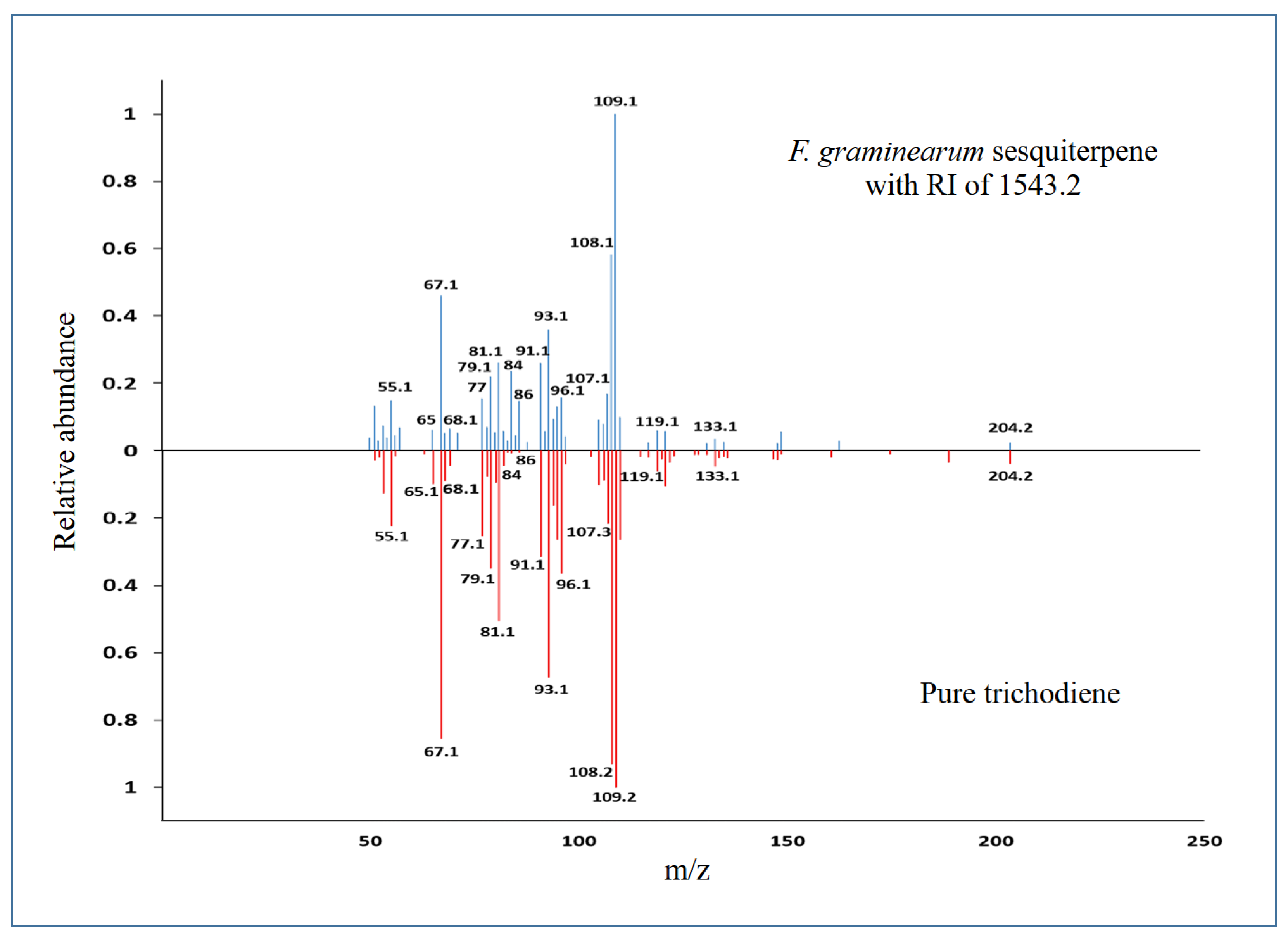
Volatile organic compounds (VOC) emitted by fungal cultures on PDA were analyzed by GC-MS (Section 5.7.). Only metabolites detected in both genotypes were quantified. In both pairs of strains, infection with mycoviruses strongly stimulated the emission of two isomeric methyl-butanols, and it strongly suppressed the emission of the trichothecene precursor trichodiene (Figure 10). The identity of trichodiene was confirmed by comparing the retention index (RI) and fragmentation pattern with an authentic standard (Appendix Figure A4).
2.6. Effect of mycoviral infection on food preference in fungivorous arthropods
To assess whether infection of F. graminearum with mycovirus affected the feeding behavior of fungivores, we carried out a dual-choice food preference assay with the collembolan Folsomia candida. In strain 266, at all times more springtails fed on cultures infected with mycoviruses than on uninfected cultures (Figure 11A). In strain 668, springtails have not distinguished between infected and uninfected mycelia in the first half of the experiment, but more animals fed on mycelia infected with mycoviruses in the second half of the experiment (Figure 11B).
Statistical significance of springtails' preference for infected mycelia was estimated using cumulative binomial probabilities (Section 4.8). The preference of springtails for strain 266MV over 266 was statistically significant (p ≤ 0.03) at all time points except at 6 h and 8 h. Over the entire experiment, the preference of F. candida for 266MV over 266 was highly significant with p < 0.0001. In contrast, the preference of springtails for strain 668MV over 668 was not significant (p > 0.05) at any single time point, but over the duration of the entire experiment, the springtails preferred feeding on strain 668MV over 668 with a p value of 0.028.
3. Discussion
3.1. Generation of isogenic strains differing in the presence of mycoviruses
Pairs of isogenic fungal strains differing in the presence of mycoviruses are pivotal in studies of the effect of mycoviruses on fungal phenotype. Such strains can be obtained by curing strains harboring mycoviruses or transmission into mycovirus-free strains. Our attempt to cure mycoviruses of strain 266MV by ribavirin and cycloheximide failed (results not shown), and indeed varying success rates of this procedure have been reported [44]. An attempt to cure mycoviruses by mitotic segregation was more successful, resulting in successful cure of two mycoviruses (Table 2). A second pair of isogenic strains differing in the load of mycoviruses was constructed by transmitting mycoviruses from strain 266MV to 668 (Table 2). We have not succeeded in obtaining F. graminearum strains harboring single mycoviruses, as pioneered by Lee at al. [25]. #### Still, comparison of the effect of mycoviruses on fungal phenotype in the two isogenic strains allowed for the tentative assignment of phenotypic effects to infection with specific mycoviruses. Assignment to single virus isolates could be inferred by comparison of the combinations reported in Table 2, however caution should be taken due to potential different response of the Fusarium genotypes to the same virus and/or to virus interactions.
A cautionary remark concerns the pair 688/688MV. Strains 688 and 688MV may not be strictly isogenic because formation of heterokaryons and parasexual recombination might have taken place during the anastomosis. The same limitation holds for the results obtained with strains harboring single mycoviruses [25]. These strains were obtained by protoplast fusion, which, too, may cause the formation of heterokaryons followed by parasexual recombination. Strictly speaking, rigorous verification would require multiple pairs of strains obtained independently, regardless of whether the strains were obtained by anastomosis or protoplast fusion. This situation is comparable to the use of gene disruption mutants in the elucidation of gene function. While keeping these restrictions in mind, comparative analysis of fungal strains harboring different sets of mycoviruses is a powerful strategy to untangle the effects of mycoviruses on the fungal phenotype.
3.2. Effect of mycoviral infection on fungal growth and sporulation
Colonies of strains 266 and 266MV differed in growth pattern and pigmentation (Figure 2A,B), but they exhibited the same radial growth rate (Figure 8A). Colonies of 668MV and 668 looked similar in the first passage. We propose that this behavior can be explained by a low virus titer in freshly transfected cultures, as reported for the transfection of FgV-ch9 [22]. After several passages, colonies of 668MV turned dark red, and their margin became irregular, similarly to 266MV. The radial growth of 668MV was strongly reduced (Figure 8C). From these results and the comparison among the strains (Table 2), we could infer that the reduction of growth rates of strain 668MV was caused by mycovirus FpV1. An alternative explanation, which holds for all inferences of this kind in the following sections, is that the genotypes 266 and 668 respond differently to the presence of the same mycovirus. If this is the case, the growth reduction in strain 668MV may have been caused by mycovirus FgV1. In any case, neither FgNSRV1 nor FgMV2 were the cause of the growth rate reduction in strain 668MV, as they were not transmitted to the 668 wild-type strain.
Reduced sporulation [45,46] and deformation of conidia [47] due to infection of F. graminearum with mycoviruses has been reported previously. Visual inspection of cultures on agar indicated that conidia production may have been reduced in 266MV and 668MV, but the effect was not statistically supported.
3.3. Effect of mycoviral infection on the accumulation of aurofusarin
The observation of stronger pigmentation of strains carrying mycoviruses (e.g., cf. Figure 2A and 2B) suggested that mycoviral infection stimulated the synthesis of red pigments. The red pigment aurofusarin is the key defense compound of F. graminearum protecting the fungus from predators [12]. Although the colonies of strains infected with mycoviruses appeared darker than uninfected strains, aurofusarin levels in infected cultures did not increase. In strain 668MV, the accumulation of aurofusarin was actually reduced, and in strain 266MV it remained unchanged (Figure 9D, 9H). The comparison among the strains (Table 2) suggests that the mycovirus responsible for the reduction of aurofusarin synthesis in 668MV was FpV1.
Could other pigments be responsible for the stronger coloration of mycovirus-infected colonies? Red pigments of F. graminearum other than aurofusarin are rubrofusarin and carotenoids. Rubrofusarin is a precursor of aurofusarin; its overproduction would therefore cause an increase in the production of aurofusarin. Carotenoids do not appear to contribute substantially to the red pigmentation of F. graminearum because strains without aurofusarin form whitish colonies with a faint yellow-orange tone [48]. Dominant carotenoids of F. graminearum are orange neurosporaxanthin and rose-red torulene [49]. We cannot exclude that the increased pigmentation of 266MV and 688MV was caused by an overproduction of torulene, but we think that it is unlikely. Examination of fungal colonies in this work (Figure 2A,B) and published photos (Figure 2B in [22], Figure 2A in [45], and Figure 3A in [47]) suggests another explanation. Uninfected colonies form abundant aerial mycelia and are fluffy, while colonies of infected strains are slimy with few aerial mycelia. We believe that this explains why colonies with mycoviruses appear darker.
3.4. Effect of mycoviral infection on the production of trichothecenes in F. graminearum
The accumulation of DON and both acetylated derivatives of DON in strain 266 was strongly suppressed by mycoviral infection (Figure 9). In strain 668, the accumulation of DON and 3ADON but not 15ADON was suppressed by mycoviral infection. Suppression of the synthesis of trichothecenes in F. graminearum has been reported in 2002 for the first time [21]; in the abstract, the authors wrote that fungal strains with a mycovirus (which was later named FgV1) exhibited "decreased (60-fold) production of trichothecene mycotoxins". According to their results, mycovirus-free cultures contained 78 to 84 mg/kg of DON, while cultures infected with the mycovirus contained 3 to 7 mg/kg. This is a 16-fold difference rather than 60-fold. More importantly, DON concentrations were not normalized to fungal biomass, though the authors noted that the growth of strains with the mycovirus was reduced. Thus, lower concentrations of DON found in cultures with mycovirus may have resulted solely from the reduction of fungal biomass. Remarkably, both errors remained unnoticed for two decades, and the claim has been perpetuated in literature till today. For instance, a recent review [50] claims that fungal viruses "...demonstrated a substantial reduction in trichothecene production", citing another review [51] instead of a primary source, and the only source cited by [51] in support of the claim is [21]. This is just one among countless cases; many publications even quote the "60-fold decrease of trichothecenes" verbatim. To put a stop to this sloppy citation practice, we have remarked this mistake firmly.
Twelve years after their discovery of the mycovirus FgV1, the authors showed that it indeed suppresses trichothecene synthesis in F. graminearum [25]. Using protoplast fusion, they generated four strains of F. graminearum, each harboring a single mycovirus, and showed that three other mycoviruses have not affected trichothecene synthesis substantially (though they observed a slight increase in normalized trichothecene levels in one of these cultures that was statistically significant).
In our work, suppression of trichothecene synthesis by mycoviruses was observed in both pairs 266/266MV and 688/688MV. The only mycovirus that was absent in both 266 and 688 and present in both 266MV and 688MV was FgAV1. Therefore, the most likely cause of the suppression of trichothecene synthesis was the ambivirus FgAV1. FgAV1 is a circular ssRNA virus with a genome length of 4.6 kb (Section 2.3.1.), while FgV1 is a dsRNA virus with a genome length of 7.5 kbp [25]. Thus, different mycoviruses are apparently able to suppress trichothecene synthesis in F. graminearum.
3.5. Effect of mycoviral infection on the emission of volatile metabolites
Mycoviruses suppressed the emission of sesquiterpene trichodiene in both fungal genotypes (Figure 9), which was expected because trichodiene is a precursor of trichothecenes [2,4].
In both genotypes, mycoviral infection strongly stimulated the emission of 2-methyl-1-butanol and 3-methyl-1-butanol (Figure 10). Considering the distribution of mycoviruses among the strains (Table 2), an analogous reasoning regarding the cause of the suppression of trichothecene synthesis (Section 3.4.) suggests that the most plausible cause of the increased production of 2- and 3-methyl-1-butanol was the infection with the ambivirus FgAV1.
3.6. Effect of mycoviruses on the attractiveness of fungal mycelia for collembolans and its possible causes
Collembolans preferred strain 266MV over 266 at all time points (Figure 11), and the preference was highly significant over the entire experiment (Section 2.5.). Instead, the number of collembolans feeding on 668MV exceeded those on 668 only in the second half of the experiment. At 5 h and 6 h, the proportion of animals feeding on 668MV was high, yet not statistically significant due to a low number of animals. Over the entire experiment, the preference for strain 668MV over 668 was significant (Section 2.5.).
These observations bring the following question: can these effects be accounted for by modulation of the synthesis of secondary metabolites? In strain 668, mycoviruses suppressed the accumulation of aurofusarin (Figure 9H), which is the major (and possibly the only, see [12]) metabolite protecting F. graminearum from fungivory. Thus, suppression of aurofusarin synthesis likely contributed to the preference of collembolans for strain 668MV over 668. It cannot be the only reason, however, because aurofusarin levels in 266 and 266MV were comparable (Figure 9D).
Has the reduction of DON content contributed to the enhanced attractiveness of mycovirus-infected strains for collembolans? A large body of literature has been devoted to feeding preferences of collembolans, but few have addressed the role of trichothecenes directly. In most of these studies, grains colonized with Fusarium were used as a food source, which obscured the cause of observed effects (e.g., [52,53]). We are aware of only two reports on the effect of DON on collembolans under controlled conditions. B. Ulber et al. (unpublished results quoted in [54]) found that collembolans fed with high concentrations of DON did not show any symptoms, and disruption of trichothecene synthesis in F. graminearum has not affected the food preference of F. candida (Suppl. Figure 5 in [12]). Both reports disproved the hypothesis that reduced content of trichothecenes B accounted for the increase attractiveness of infected strains to collembolans. Recurring exposure of collembolans to trichothecenes B might have led to resistance. Selection for resistance to aurofusarin might have been prevented by vast amounts accumulating in mycelia, which denied partially resistant mutants of a fitness advantage [12].
Fungal VOCs are known to trigger olfactory responses in collembolans and modulate their food preference [55]. Our results revealed two groups of VOCs able to explain the effect of mycoviruses on the attractiveness of F. graminearum for collembolans. The first of them is trichodiene. We excluded the role of trichothecenes B produced by F. graminearum, but trichodiene is also a precursor of trichothecenes A and macrocyclic trichothecenes, which are substantially more toxic than trichothecene B. It is conceivable that the ability of collembolans to avoid mycelia that emit trichodiene was selected by the toxicity of highly toxic trichothecenes produced by fungi other than Fusarium spp. The hypothesis could be easily tested by an olfactometric experiment.
The second candidate for the cause of the increased attractiveness of infected mycelia for collembolans are isomeric methylbutanols (Figure 10). Low concentrations of 2-methyl-1-butanol saturated the antennal receptors of Orchesella cincta and Tomocerus flavescens [56] and attracted collembolan Onychiurus armatus [57]. 3-methyl-1-butanol has not been tested in these studies, though the production of this VOC by fungi investigated in these studies was explicitly mentioned; we assume that the compound was not commercially available at that time. The hypothesis that 2-methyl-1-butanol and/or 3-methyl-1-butanol contributed to the attractiveness of mycovirus-infected strains to F. candida can be easily tested in an olfactometric experiment.
Does stimulation of VOC emission by mycoviruses possess an ecological function? Plant viruses modulate the attractiveness of their hosts to animal vectors, increasing their chance of propagation [58,59]. Dissemination of mycoviruses by fungivores has been demonstrated in a single case [16]. We suggest that the hypothesis that stimulation of the emission of volatile attractants contributes to the dissemination of mycoviruses can be tested by monitoring the spread of mycoviruses in soil microcosmoses exposed to different levels of fungivory.
4. Materials and Methods
4.1. Fungal strains and cultures
The strains of Fusarium graminearum used are listed in Table 3. F. graminearum 266MV, deposited in DSMZ culture collection (Braunschweig, Germany) as F. graminearum DSM 116490, was isolated in 2018 from a naturally infected maize ear grown in Pocking (Bavaria, Germany) with a disease rating of 5 % according to a modified symptom scale [60] as previously described [61]. F. graminearum 668 (DSM 113709), kindly provided by Wilhelm Schäfer (University of Hamburg, Hamburg, Germany), is a zearalenone-deficient mutant (△ZEN) generated from wildtype strain F. graminearum 1003 by disrupting the polyketide synthase gene PKS4 [62]. The strain carries a hygromycin B (HPH) resistance gene, which served as a resistance marker in transmission experiments. Strain 266 (DSM 116489) is a descendant of 266MV (DSM 116490) that spontaneously lost mycoviruses (for details, see Section 3.1). Strain 668MV (DSM 116488) is a transfectant of 668 (DSM 113709) harboring mycoviruses from 266MV (DSM 116490). All strains were grown on potato dextrose agar (PDA, Carl Roth, Karlsruhe, Germany) at 25 °C in the dark.

Table 3.
Strains of Fusarium graminearum.
| Strain | Short label | Properties |
|---|---|---|
| F. graminearum DSM 116490* | 266MV | Natural isolate harboring mycoviruses |
| F. graminearum DSM 116489 | 266 | Descendant of 266MV that lost two mycoviruses |
| F. graminearum DSM 113709 | 668 | Hygromycin-resistant strain without mycoviruses |
| F. graminearum DSM 116488 | 668MV | Transfectant of 668 harboring two mycoviruses from 266MV |
* DSMZ-German Collection of Microorganisms and Cell Cultures GmbH, Inhoffenstraße 7 B, 38124 Braunschweig, Germany.
4.2. RNA extraction, sequencing, RACE, RT-PCR, and phylogenetic analysis
Mycelium was scrubbed from the surface of a PDA plate, snap-frozen in liquid nitrogen, and ground in a ceramic mortar. Total RNA was extracted from frozen mycelium using TRIzol (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. The quality of RNA extracts was assessed by electrophoresis in nondenaturing 2.2 % agarose gels [63].
The cDNA was synthetized with random octamer primers and Maxima H Minus Reverse Transcriptase (Thermo Fischer Scientific, Waltham, MA, USA), followed by second-strand synthesis using the NEB Next Ultra II Non-Directional RNA Second Strand Module (New England Biolabs, Ipswich, MA, USA). Following on column purification with a NucleoSpin Gel and PCR Clean-up kit (Macherey-Nagel, Dueren, Germany), an Illumina Nextera XT DNA library was prepared according to manufacturer´s recommendations and quantified (Qubit, Thermo Fischer Scientific), and the size of DNA fragments was assessed with a 2100 Bioanalyzer system and a High Sensitivity DNA Assay Kit (Agilent, Darmstadt, Germany). After normalization, the samples were sequenced on an Illumina MiSeq instrument as paired-end reads (2 x 300 bp). Contigs were de novo assembled and further analyzed in GeneiousPrime (Biomatters, Auckland, New Zealand) for virus discovery by BLAST alignment and taxonomic assignment.
Complete genome sequences of the identified viruses were obtained by RACE according to [64]. 5' and 3' RACEs (rapid amplification of cDNA ends) were performed for FpV1, FgSV1 and FgMV2. For this purpose, cDNA was synthesized using a virus specific primer, tailed with A, C, G or T, and the products were amplified using a virus-specific primer and a primer matching the respective tail. Specifically, 4 µl of dsRNA solution, 1 µl of the respective primer (10 μM; Eurofins Genomics, Hamburg, Germany), and 7 µl of H2O were incubated at 95 °C for 3 min and then immediately transferred to liquid nitrogen. A pre-warmed (50 °C) RT-mix consisting of 4 µl 5X RT buffer, 1 µl dNTPs (10mM each; Carl Roth, Karlsruhe, Germany), 0.25 µl Maxima H Minus Reverse Transcriptase, and 0.5 µl Ribolock RNase Inhibitor, all from Thermo Fisher Scientific (Darmstadt, Germany). 4.25 µl H2O were added and the reaction was incubated for 60 min at 50 °C, for 15 min at 55 °C, for 15 min at 60 °C, and for 5 min at 85 °C. After purification using SureClean Plus system (Bioline, Luckenwalde, Germany), cDNA was tailed with A, C, G and T. To this end, 3 µl of purified cDNA were incubated with 4 µl of 5X reaction buffer, 1 µl terminal deoxynucleotidyl transferase (Thermo Fisher Scientific, Darmstadt, Germany), 1 µl of the respective dNTP (100 mM), and 11 µl of H2O for 30 min at 37 °C, followed by 10 min at 70 °C. The subsequent PCR reaction, consisted of 3 µl tailed cDNA, 15 µl of 2X Phusion Flash High-Fidelity PCR Master Mix (Thermo Fisher Scientific, Darmstadt, Germany), 1.5 µl of virus- and tail-specific primer (10 μM each) and 9 µl water. PCR program started at 98 °C for 15 s, followed by 34 cycles of 98 °C for 5 s, primer annealing (55 to 59 °C) for 5 s, 72 °C for 15 s, and ended with a final elongation of 5 min. Half of the sample was loaded onto a 1 % agarose gel, purified using NucleoSpin Gel and PCR Clean-up Kit (Macherey-Nagel, Düren, Germany) in 20 µl final volume following manufactorer's instructions and either sequenced directly or cloned into a standard cloning vector (CloneJET PCR Cloning Kit, Thermo Fisher Scientific, Darmstadt, Germany). If a single fragment was clearly visible on the gel, the second half of the sample that was not loaded onto the agarose gel was purified using SureClean Plus system (Bioline, Luckenwalde, Germany) in 20 µl final volume and sequenced directly.
For the reverse transcription of specific fragments of mycoviral genomes, 2 µl of total RNA were digested with 2 U/µl DNAseI (New England Biolabs, Frankfurt am Main, Germany) in DNase reaction buffer (10 mM Tris-HCl, 2.5 mM MgCl2, 0.5 mM CaCl2, pH 7.6), the quality of dsRNA was assessed by agarose gel electrophoresis (Section 2.2). RNA was reverse transcribed using primers specific for viral sequences (Appendix Table A1) as follows. 3 µl of dsRNA extract were incubated together with 1 µl of the appropriate sense and antisense primer (10 μM each; Eurofins Genomics, Hamburg, Germany) for 5 min at 95 °C and then immediately cooled on ice. This is followed by the addition of 4 µl 5X RT buffer (Thermo Fisher Scientific), 0.5 µl dNTPs (10mM each; Carl Roth), 1 µl RevertAid Reverse Transcriptase (20 U/µl) (Thermo Fisher Scientific), and 9.5 µl H2O and an incubation of 45 min at 42 °C. For the subsequent PCR, a reaction mix consisting of 2 µl cDNA, 5 µl 2X Phusion Flash High-Fidelity PCR Master Mix (Thermo Fisher Scientific, Darmstadt, Germany), 1 µl of sense and antisense primer each (10 μM each; salt-free; Eurofins Genomics, Hamburg, Germany) and 1 µl H2O, started at 98 °C for 15 s, followed by 34 cycles of 98 °C for 5 s, primer annealing (55 to 59 °C) for 5 s, 72 °C for 15 s, and ended with a final elongation of 5 min. Subsequently, the sample was loaded onto a 1 % agarose gel and selected fragments were purified using Macherey-Nagel NucleoSpin Gel and PCR Clean-up Kit (Macherey-Nagel, Düren, Germany) and sequenced.
Following translation into amino acids, the complete gene for RNA-dependent RNA polymerase (RdRp) was used to determine the evolutionary relationships and genetic distances to reference genomes in the NCBI database and construct phylogenetic dendrograms [65,66]. Sequence alignments were created and Neighbor-Joining dendrograms [34] were constructed with MEGA 10.1.8 [35].
4.3. DNA extraction, DNase treatment, and quantification of fungal biomass
In rice cultures, fungal DNA was used as a proxy for biomass. Mycelia harvested from a liquid culture (for DNA standards) and entire rice cultures consisting of rice grains and fungal mycelium (samples) were freeze-dried and ground to fine powder in a reciprocal mill (MM400, Retsch, Haan, Germany). DNA was extracted using a cetyltrimethylammonium-based protocol [67]. To differentiate between DNA and RNA bands in agarose electrophoresis, nucleic acid extracts were treated with 2 U/µl DNaAse I (New England Biolabs, Frankfurt am Main, Germany) in DNase reaction buffer (10 mM Tris-HCl, 2.5 mM MgCl2, 0.5 mM CaCl2, pH 7.6) for 1 h at 37 °C.
Fungal biomass in grains was determined by quantitative PCR (qPCR) as described [67] using two replicates of densely spaced standards (see Figure A2 in [60]). The biomass of fungal colonies growing on agar was determined directly, as described in Section 4.7.
4.4. Transmission of mycoviruses between F. graminearum strains
Transmission of viral RNAs from F. graminearum 266MV to F. graminearum 668 was achieved by hyphal anastomosis during co-cultivation according to Darissa et al. [22]. Briefly, an agar block of 6 mm diameter overgrown with mycelium of strain 266MV was placed on a PDA plate and incubated for 5 days at 25° C in the dark at room temperature. A second agar block with virus-free hygromycin-resistant strain 668 was positioned at a distance of 1 cm. After 7 days of incubation in the dark, several agar blocks were cut out from the area were the fungal colonies met, transferred to PDA plates containing 50 µg/ml of hygromycin B1 (Carl Roth GmbH, Karlsruhe, Germany), and incubated for 5 days. Single spores were harvested by rinsing the cultures with sterile tap water and plating dilutions of the spore suspensions onto agar plates. This workflow is visualized in Figure S1.
Single-spore isolated were assigned to the 266 or 668 genotype by PCR with the primer pair FGPKS4f1 and FGPKS4r1 (Appendix Table A1), which encloses the area of PKS4 that was disrupted in the acceptor strain 668. PCR was carried out using the conditions used for the amplification of TEF-1α [29]. The presence of viral dsRNAs in the acceptor strain was also detected by agarose electrophoresis and confirmed by reverse transcription PCR (see Section 4.2.).
4.5. Determination of radial growth rate and sporulation
Radial growth rate was determined on potato dextrose agar (PDA) plates (Carl Roth, Karlsruhe, Germany). Round agar blocks covered with mycelium with a diameter of 5 mm were placed onto agar plates and incubated in the dark at 25 °C for 48 h. Eight plates were used for each estimate. From each plate, 4 measurements of the distance from the agar block to the tips of mycelial hyphae were taken at a 90° angle.
In order to assess the effect of infection with mycovirus on sporulation, infected and uninfected strains were cultivated in 100 ml of nutrient-deprived broth (SNA), containing 9.9 mM KNO3, 7.3 mM KH2PO4, 0.3 mM MgS04 x 7H2O, 6.7 mM KCl, 1.1 mM glucose, and 0.6 mM sucrose) [68] for 7 days at 25 °C with shaking at 130 rpm. Spores were collected from culture supernatant by centrifugation at 4,500 rcf for 10 min, suspended in 1 ml of sterile tap water, and counted in a Thoma chamber. Macroconidia were counted with a Thoma chamber. Spore concentrations below the limit of quantification (5 x 104 spores/ml) were replaced with limit of quantification, analogously to left-censored data in analytical chemistry.
4.6. Quantification of trichothecenes and aurofusarin
Rice cultures were prepared as previously described [69] and grown for 21 d at 25 °C. The extraction and HPLC-MS/MS analysis of trichothecenes were previously described [69]. Aurofusarin was extracted separately, using 1 g of freeze-dried and ground culture material and 10 mL methanol, and analyzed by HPLC-MS/MS essentially as described by Xu at al. [12]. MS/MS transitions, limits of detection (LODs), and limits of quantification (LOQs) are listed in Appendix Table A1. To compensate for differences in fungal biomass, concentrations of metabolites were normalized to fungal DNA determined by qPCR (Section 4.3.).
4.7. Analysis of volatile compounds
Petri dishes (35 mm diameter; Sarstedt AG & Co. KG, Nuembrecht, Germany) were filled with 4 mL of PDA, inoculated with F. graminearum strains, and cultivated at 25 °C in the dark. After seven days, the lid was removed, and the plate was transferred into a top-open headspace glass vessel (50 mm diameter, 115 mm height) with an inlet and an outlet for airflow at the opposite sides (openings of 9 mm diameter). The inlets were closed with screw caps, and the upper opening was sealed with polyester oven bag foil (Confresco Frischhalteprodukte GmbH & Co. KG, Minden, Germany). The glass vessel was then further incubated for 24 h at 25 °C in darkness. After incubation, the vessels were attached to an air inflow adjusted to 320 ml purified air/min, and the air was sucked out at a rate of 300 ml/min with a Laboport vacuum pump (KNF Neuberger GmbH, Freiburg, Germany). Before entering the headspace, the air was filtered through two hydrocarbon/moisture traps (Agilent Technologies, Model HT200-4). Headspace sampling was carried out for 2 h in darkness at 24 °C and 45% relative humidity. Volatiles were collected with Porapak Q filters (VCT-1/4-3-POR-Q, www.ars-fla.com) placed in the outflow opening and eluted with 150 µL dichloromethane. Porapak filters were rinsed with 1 mL dichloromethane before each sampling. Empty Petri dishes and Petri dishes filled with PDA were analyzed similarly to identify background volatiles. Six replicates of each culture were analyzed.
200 ng of tetralin standard (20 ng/µL; Sigma-Aldrich Chemie GmbH, Taufkirchen, Germany) were added to each sample and 40 µL of the spiked sample were transferred to a new glass vial with a glass insert for GC-MS analysis, which was carried out on an Agilent Technologies 7890 GC System with Agilent 5977B GC/MSD. An HP5-MS analytical column was used (30 m × 0.25 mm inner diameter, 0.25 μm film thickness, Agilent Technologies, Santa Clara, CA, USA). 2 µL of the eluate were injected in splitless mode at 220 °C and 18.84 psi. The oven temperature was held at 40 °C for 3 min, raised to 320 °C at a rate of 8 °C min−1, and held at this temperature for 8 min. Helium was used as a carrier gas at a constant flux of 1.5 ml min−1.
Chromatograms were analyzed using Enhanced ChemStation, MSD ChemStation F.01.03.2357 (Agilent, Darmstadt, Germany). VOC identities were confirmed by comparing mass spectra and retention indices (RI) with the values listed by NIST (National Institute of Standards and Technology, USA, www.webbook.nist.gov). The identity of trichodiene was confirmed by the comparison of retention index and fragmentation spectrum with authentic standard (+)-trichodiene (HPC Standards, Cunnersdorf, Germany). Only signals occurring in all strains were quantified. For relative quantification, peak areas were normalized with the area of the internal standard tetralin.
Intensities of signals that were found in both F. graminearum genotypes were normalized to fungal biomass as follows. Fungal mycelium was separated from agar by placing agar overgrown with mycelia into a beaker with water and heating the mixture in a microwave until agar melted and dissolved in water. The mycelium was then separated by filtration, freeze-dried, and weighed. Volatile levels were normalized to the mycelial weight. The significance of differences between means was assessed by Student t-test or Wilcoxon rank-sum test. Samples that have not emitted any detectable VOCs were excluded from the analysis.
4.8. Food preference assay
The springtail Folsomia candida was reared in Petri dishes on a layer of plaster of Paris and activated charcoal (13:1, w/w) at 20 °C in darkness. Collembolans were fed weekly with dry baker's yeast (Lucullus Food Service GmbH & Co. KG, Trittau, Germany) ad libitum. The plate was aerated during feeding. Tap water was added when needed to maintain humidity levels. Before being used in experiments, collembolans were transferred to a new plate with pure plaster of Paris and starved for 48 h.
Food preference assays with collembolans were performed in plastic Petri dishes (9 cm diameter; Sarstedt AG & Co. KG, Nuembrecht, Germany) on plaster. All trials were carried out with one animal per plate to avoid crowding effects in three repetitions with ten replicates each. The animals were placed in the middle of the arena and had to choose between the mycelium of infected and healthy strains of F. graminearum, presented as agar plugs (9 mm diameter) overgrown with fungal mycelium. The results were evaluated by counting animals that fed on infected or healthy mycelium or that have been located in the periphery (‘no choice’). Counting was repeated once per hour for 8 hours. During the experiment, the animals were kept in the dark at 21 °C.
P-value was estimated as a cumulative binomial probability that the same or a larger number of animals would feed on cultures with mycoviruses under the conditions of the experiment if the animals had chosen the food source randomly.
4.9. Transmission electron microscopy
Fresh mycelium was scrubbed from the surface of a PDA plate of the infected and uninfected cultures, snap-frozen in liquid nitrogen, and ground in a ceramic mortar. Mycelium powder was resuspended in 1 ml of a 0.1 M sodium-phosphate buffer (2 % PVP, 0.2 % sodium sulfite, 0.05 % sodium azide) and incubated for 30 min at 4 °C in a rotary shaker at 400 rpm (Thermomixer comfort; Eppendorf SE, Hamburg, Germany). Then, 800 µl of chloroform/isoamyl alcohol (24:1, v/v) were added to the mixture and the samples were centrifuged at 10.000 x g for 10 min (Centrifuge 5415D; Eppendorf SE, Hamburg, Germany). Then, 600 µl of the supernatant were transferred to a new 2 ml tube with 800 µl chloroform/isoamylalkohol (24:1, v/v) and the centrifugation step was repeated. After that, 600 µl of the supernatant were transferred to a new 1.5 ml Eppendorf tube, and 200 µl of 30 % (w/v) PEG 6000 and 100 µl 5 M NaCl were added. The samples were mixed by inverting the tube 7 times. Precipitation was performed for 1 h at 4 °C in a rotary shaker at 400 rpm. Afterwards, the sample was centrifuged at 10,000 x g and 4 °C for 15 min. The supernatant was discarded, and the pellet was dissolved in 20 µl of 0.1 M sodium-phosphate buffer. All samples were stored at 4-7 °C until transmission electron microscopy analysis.
Electron microscopy was performed at the facilities of the institute for Epidemiology and Pathogen Diagnostics of the Julius Kühn-Institute in Braunschweig. Briefly, a copper grid (G400EM-C3, 400 lines/inch square mesh; Plano, Wetzlar, Germany) was coated with a 0.5 % Pioloform in chloroform solution and sputtered with carbon (Mikrotechnik Dr. Hert, München, Germany). Mycelium extract was applied on the surface of Parafilm, and a coated grid was placed on the top of the extract. Virions were allowed to adsorb on the grid for 5 min, the grid was washed with distilled water, drained with filter paper, and stained with a 1 % uranyl acetate solution (Riedel de Haën, Seelze, Germany). Transmission electron microscopy was performed with a Tecnai G2 Spirit TWIN microscope (Hillsboro, Oregon, USA) system. Photos were taken with a CCD camera, model WA-Veleta (Olympus Life Science Solutions, Hamburg, Germany).
Author Contributions
Conceptualization, S.S. and P.K.; methodology, S.S., M.S., M.A., D.K., P.M., H.R., K.R., M.R. and P.K.; software, S.S:, M.S., M.A., H.R., D.K. and P.M.; validation, S.S, M.S., M.A., H.R., D.K. and P.M.; formal analysis, S.S., M.S., M.A. and P.K. ; investigation, S.S., M.S., M.A., M.B., D.K., P.M., H.R. and K.R.; resources, S.S., M.R. and P.K.; data curation, S.S., M.S. and M.A.; writing—original draft preparation, S.S.; writing—review and editing, all authors; visualization, S.S. and K.R.; supervision, M.R. and P.K.; project administration, P.K.; funding acquisition, M.R., P.K., and S.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data are provided in the paper and its appendices. Nucleotide sequences have been deposited in NCBI, Acc. Nos. are listed in Appendix Table A2. All strains are available from DSMZ-German Collection of Microorganisms and Cell Cultures GmbH, Inhoffenstraße 7 B, 38124 Braunschweig, Germany.
Acknowledgments
The authors would like to thank Luciana Macis, Nahid Najafi Hajiwar, Inga Hilbrich, Ruth Pilot, Jonas Wattenrott, Heike Rollwage, Sabine Schuhmann and Jessica Ponath for technical support, Wilhelm Schäfer (University of Hamburg) for fungal strains, and Marilia Bueno da Silva and Kiran Parajuli for help with some experiments. We thank Beatriz Navarro and Francesco Di Serio (Istituto per la Protezione Sostenibile delle Piante(CNR), Unità Organizzativa di Bari, Bari, Italy) for the prediction of RNA secondary structure.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Table A1.
Primers.
| name | sequence (5’-3’) | fragment length [bp] | target | source |
| FP-F1N | TATCGTCTCCACGAATCTCC | 786 | FpV RNA1 | [70] |
| FP-R1N | CTCGAATTGTTTCGAGACCG | |||
| FgSV1_F2 | CTCACACGTTTTTCGCAAGTT | 697 | FgSV1 | this study |
| FgSV1_R2 | GGCATCATTGGTAATCCTCCTA | |||
| Mito76/88uniF | GACARCCTATGGGAGCGTA | 670 | FgMV2 | this study |
| Mito76R20 | GCTTCAGTTTCTAGCACAGCT | |||
| FGPKS4f1 | CATGGTCCAAGTGCTACTGT | 800 | FGPKS4 | this study |
| FGPKS4r1 | CCACTAACGCTCATACGCTT | |||
| FgAV1_F1 | GCATCAAATGTAGCATAAAGATTGC | 783 | FgAV1 | this study |
| FgAV1_R1 | GCTCCTGGAGTATCTAAGGATGAG | |||
| EF1αF | ATGGGTAAGGARGACAAGAC | 690 | TEF-1α | [29] |
| EF1αR | GGARGTACCAGTRATCATGTT | |||
| Fg16NF | ACAGATGACAAGATTCAGGCACA | 280 | - | [71] |
| Fg16NR | TTCTTTGACATCTGTTCAACCCA | |||
| FpV_RNA1_cDNA_5endas | CGGATACTGTGGCGTTAGTGG | 560+X | FpV RNA1 | this study |
| FpV_RNA1_PCR_5endas | CGGGAGTAGCAGTGACGACG | 431+X | FpV RNA1 | this study |
| FpV_RNA1_cDNA_3ends | CAAACTCGTCGCACAGCTCTG | 422+X | FpV RNA1 | this study |
| FpV_RNA1_PCR_3ends | GTTTAACGATGATCGCGCCG | 284+X | FpV RNA1 | this study |
| FpV_RNA2_cDNA_5endas | CAGAGTGAGACAGTAGCCAGCG | 459+X | FpV RNA2 | this study |
| FpV_RNA2_PCR_5endas | CATAATCACCGGAGTCAAGTTCG | 319+X | FpV RNA2 | this study |
| FpV_RNA2_cDNA_3ends | CAATGATCGCTCCACTGCTGC | 574+X | FpV RNA2 | this study |
| FpV_RNA2_PCR_3ends | CTTACTCCCGAACTCCACCACG | 399+X | FpV RNA2 | this study |
| FgSV1_5end_cDNA_s | CTTTGTCCTGTTGCTTTTGAGC | 517+X | FgSV1 | this study |
| FgSV1_5end_PCR_s | CTTCCTTGACCGTGACATCCG | 334+X | FgSV1 | this study |
| FgSV1_3end_cDNA_as | CCGAGACAGAGATGCAGATGGTC | 516+X | FgSV1 | this study |
| FgSV1_3end_PCR_as | GAATTGATTATGCGAACCACCG | 328+X | FgSV1 | this study |
| Mito_5end_cDNA_as | GAATCAAAGGTTGAGTTAATAGGCG | 461+X | FgMV2 | this study |
| Mito_5end_PCR_as | GTTACCCACAAGACGACCATCAG | 293+X | FgMV2 | this study |
| Mito_3end_cDNA_s | CACACAATGGATGTTCTCATGCC | 743+X | FgMV2 | this study |
| Mito_3end_PCR1_s | GTACGCTAAGAATAGGAATGCCG | 544+X | FgMV2 | this study |
| Mito_3end_PCR2_s | GCCAATAAAACCGGCAATTTG | 498+X | FgMV2 | this study |
| Poly-A16 | CCTCGGGCAGTCCAAAAAAAAAAAAAAAA | - | respective tailed cDNA | this study |
| Poly-G15 | CTCAAACAGTCACGGGGGGGGGGGGGGG | - | respective tailed cDNA | this study |
| Poly-C14 | ATCCTGCAGGCGCGCCCCCCCCCCCCCC | - | respective tailed cDNA | this study |
| Poly-T18 | CCTCGGGCAGTCCTTTTTTTTTTTTTTTTTT | - | respective tailed cDNA | this study |
Table A2.
References for analysis of mitoviral RDRP motifs by multiple sequence alignment.
| Abbreviation | Full virus species name | GenBank accession number |
| RsAV1 | Rhizoctonia solani ambivirus 1 | QMP84024 |
| RsAV2 | Rhizoctonia solani ambivirus 2 | QMP84026 |
| AmAV2 | Armillaria mellea ambi-like virus 2 | DAD54839 |
| TuAV1 | Tulasnella ambivirus 1 | QPB44664 |
| HbAV1 | Heterobasidion ambi-like virus 1 | UHK02572 |
| PgAV1 | Phlebiopsis gigantea ambi-like virus 1 | UJT31805 |
| CpAV1 | Cryphonectria parasitica ambivirus 1 | QMP84022 |
| PgAV2 | Phlebiopsis gigantea ambi-like virus 2 | UJT31806 |
| AmAV1 | Armillaria mellea ambi-like virus 1 | DAD54837 |
| AbAV1 | Armillaria borealis ambi-like virus 1 | QUD20357 |
| AlAV1 | Armillaria luteobubalina ambi-like virus 1 | DAD54835 |
| HbAV3 | Heterobasidion ambi-like virus 3 | UHK02576 |
| TuAV5 | Tulasnella ambivirus 5 | QPB44674 |
| AnAV1 | Armillaria novae-zelandiae ambi-like virus 1 | DAD54841 |
| TuAV3 | Tulasnella ambivirus 3 | QPB44670 |
| TuAV4 | Tulasnella ambivirus 4 | QPB44672 |
| CbAV1 | Ceratobasidium ambivirus 1 | QPB44668 |
| CRV | Chimay rhabdovirus | AVM86063 |
| SsNSRV-1 | Sclerotinia sclerotiorum negative-stranded RNA virus 1 | YP009094317 |
| SsNSRV-3 | Sclerotinia sclerotiorum negative-stranded RNA virus 3 | YP009129259 |
| SsNSRV-9 | Sclerotinia sclerotiorum negative-stranded RNA virus 9 | QUE49142 |
| BcNSRV-3 | Botrytis cinerea negative-stranded RNA virus 3 | QJT73696 |
| CpSM1 | Cryphonectria parasitica sclerotimonavirus 1 | QMP84020 |
| SlNSRV-2 | Soybean leaf-associated negative-stranded RNA virus 2 | ALM62227 |
| AtNSRV1 | Alternaria tenuissima negative-stranded RNA virus 1 | QDB75013 |
| PvLAMNV2 | Plasmopara viticola lesion associated mononega virus 2 | QHD64785 |
| SlNSRV1 | Soybean leaf-associated negative-stranded RNA virus 1 | ALM62220 |
| FgNSRV1 | Fusarium graminearum negative-stranded RNA virus 1 | ATP75709 |
| BcNSRV4 | Botrytis cinerea negative-stranded RNA virus 4 | QJT73698 |
| Narnavirus sp. | Narnavirus sp. | QDH90729 |
| PvLAMV | Plasmopara viticola lesion associated mymonavirus 1 | QHD64779 |
| PcNSRV1 | Penicillium cairnsense negative-stranded RNA virus 1 | QDB75012 |
| SsNSRV7 | Sclerotinia sclerotiorum negative-stranded RNA virus 7 | AWY11040 |
| BcMV1 | Botrytis cinerea mymonavirus 1 | AXS76906 |
| CdNSRV1 | Coniothyrium diplodiella negative-stranded RNA virus 1 | QDB75015 |
| SlNSRV1 | Soybean leaf-associated negative-stranded RNA virus 1 | ALM62222 |
| FgNSRV1 | Fusarium graminearum negative-stranded RNA virus 1 | UNG44333 |
| FgNSRV1 | Fusarium graminearum negative-stranded RNA virus 1 | ATP75707 |
| FaNSRV1 | Fusarium asiaticum negative-stranded RNA virus 1 | UNG44330 |
| FmMV1 | Fusarium mangiferae mitovirus 1 | UBZ25880 |
| SlMV1 | Soybean leaf-associated mitovirus 1 | ALM62241 |
| BdMV1 | Botryosphaeria dothidea mitovirus 1 | QMU24933 |
| StMV1 | Setosphaeria turcica mitovirus 1 | AZT88625 |
| FsMV2 | Fusarium sambucinum mitovirus 2 | BCP96873 |
| CpMV1 | Cryphonectria parasitica mitovirus 1-NB631 | NP660174 |
| GaNV1 | Grapevine associated narnavirus-1 | CEZ26304 |
| SsMV7 | Sclerotinia sclerotiorum mitovirus 7-A | AWY10969 |
| BsMV1 | Buergenerula spartinae mitovirus 1 | AHY03257 |
| FaMV1 | Fusarium asiaticum mitovirus 1 | UNG44312 |
| PvMV13 | Plasmopara viticola lesion associated mitovirus 13 | QIR30236 |
| NpMV1 | Neofusicoccum parvum mitovirus 1 | QDB74992 |
| RsMV2 | Rhizoctonia solani mitovirus 2 | ALD89121 |
| FuMV1 | Fushun mitovirus 1 | UHM27612 |
| EnMV13 | Erysiphe necator associated mitovirus 13 | QKI79961 |
| BcMV1 | Botrytis cinerea mitovirus 1 | CEZ26300 |
| OpMV3 | Ophiostoma mitovirus 3b | CAJ32468 |
| MlSV | Mito-like spartanusvirus | QOW97240 |
Table A3.
MS/MS transitions, limits of detection (LODs), and limits of quantification (LOQs) for trichothecenes and aurofusarin.
Table A3.
MS/MS transitions, limits of detection (LODs), and limits of quantification (LOQs) for trichothecenes and aurofusarin.
| Toxin | Source | Polarity | Parent ion | Frag. [V] |
Collision [V] | Product ion | LOD [mg/kg] |
LOQ [mg/kg] |
| NIV | Romer Labs, Butzbach, Germany | Neg. | 357.1 | 95 | 10 10 |
311.1 281.1 |
0.02 | 0.08 |
| DON | Romer Labs, Butzbach, Germany | Pos. | 297.1 | 100 | 4 64 |
249.1 91.2 |
0.006 | 0.018 |
| 3-ADON | LGC Standards, Wesel, Germany | Pos. | 339.2 | 100 | 8 8 |
231.1 203.0 |
0.02 | 0.06 |
| 15-ADON | Sigma Aldrich, Steinheim am Albuch, Germany | Pos. | 339.2 | 90 | 10 10 10 |
321.1 261.0 137.0 |
0.04 | 0.13 |
| Aurofusarin | Bioviotica Naturstoffe GmbH, Dransfeld, Germany | Pos. | 571.0 | 195 | 21 21 |
556.0 541.0 |
7.8 | 26 |
Figure A1.
Scheme of the workflow for the transfection of mycoviruses. Hygromycin-resistant F. graminearum strain 668 was transfected with viruses from the wildtype isolate 266MV by hyphal anastomosis. Transfected colonies were selected on PDA with hygromycin B1 at 50 µg/ml. The bottom-view of the Petri dishes is shown.
Figure A1.
Scheme of the workflow for the transfection of mycoviruses. Hygromycin-resistant F. graminearum strain 668 was transfected with viruses from the wildtype isolate 266MV by hyphal anastomosis. Transfected colonies were selected on PDA with hygromycin B1 at 50 µg/ml. The bottom-view of the Petri dishes is shown.
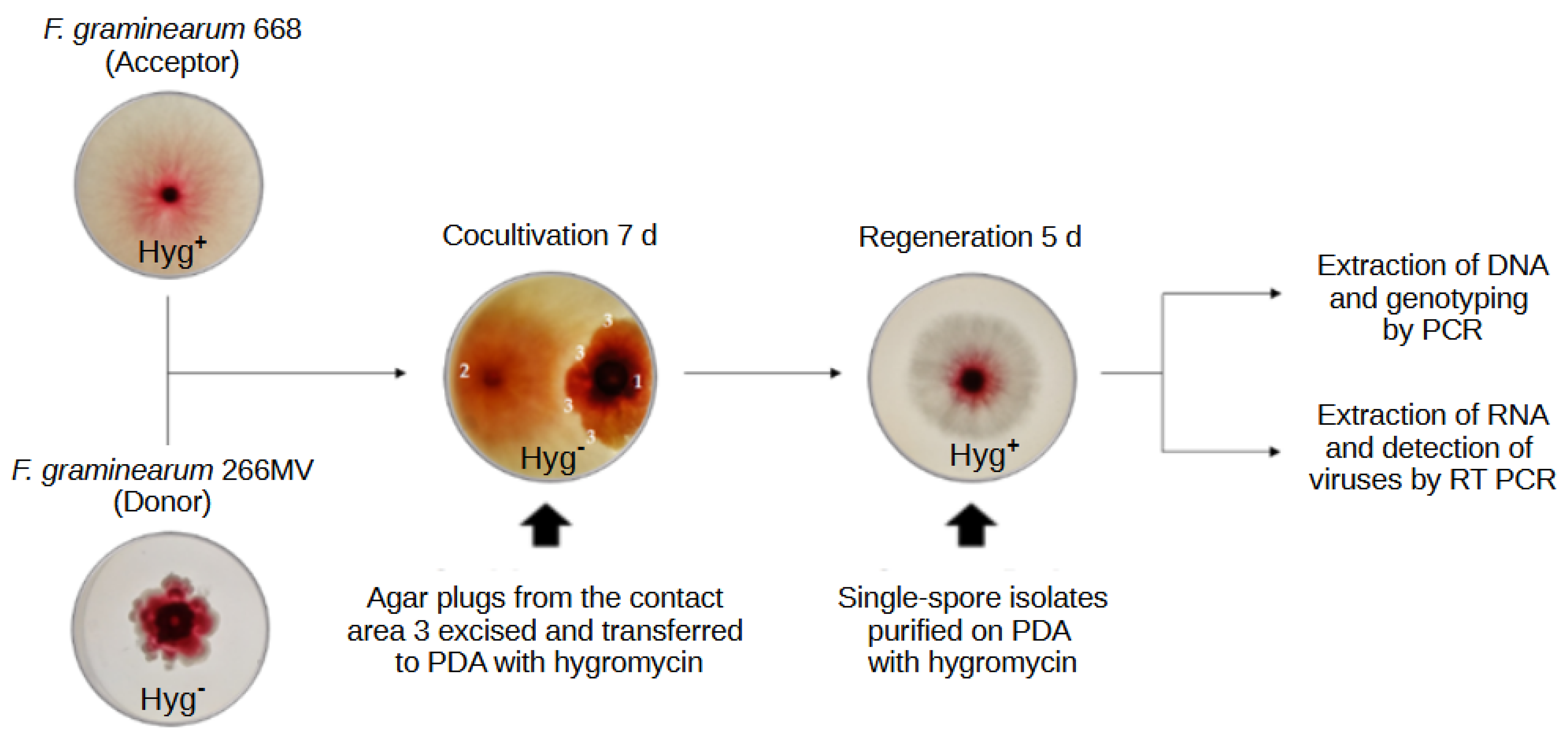
Figure A2.
Predicted secondary structure of proposed ribozymes located on the (A) positive and (B) negative strand of the FgAV1. Helices I and II are printed in blue, helix III in green, and 13 core nucleotides required for RNA cleavage are printed in red. The self-cleavage sites are marked with blue arrows.
Figure A2.
Predicted secondary structure of proposed ribozymes located on the (A) positive and (B) negative strand of the FgAV1. Helices I and II are printed in blue, helix III in green, and 13 core nucleotides required for RNA cleavage are printed in red. The self-cleavage sites are marked with blue arrows.
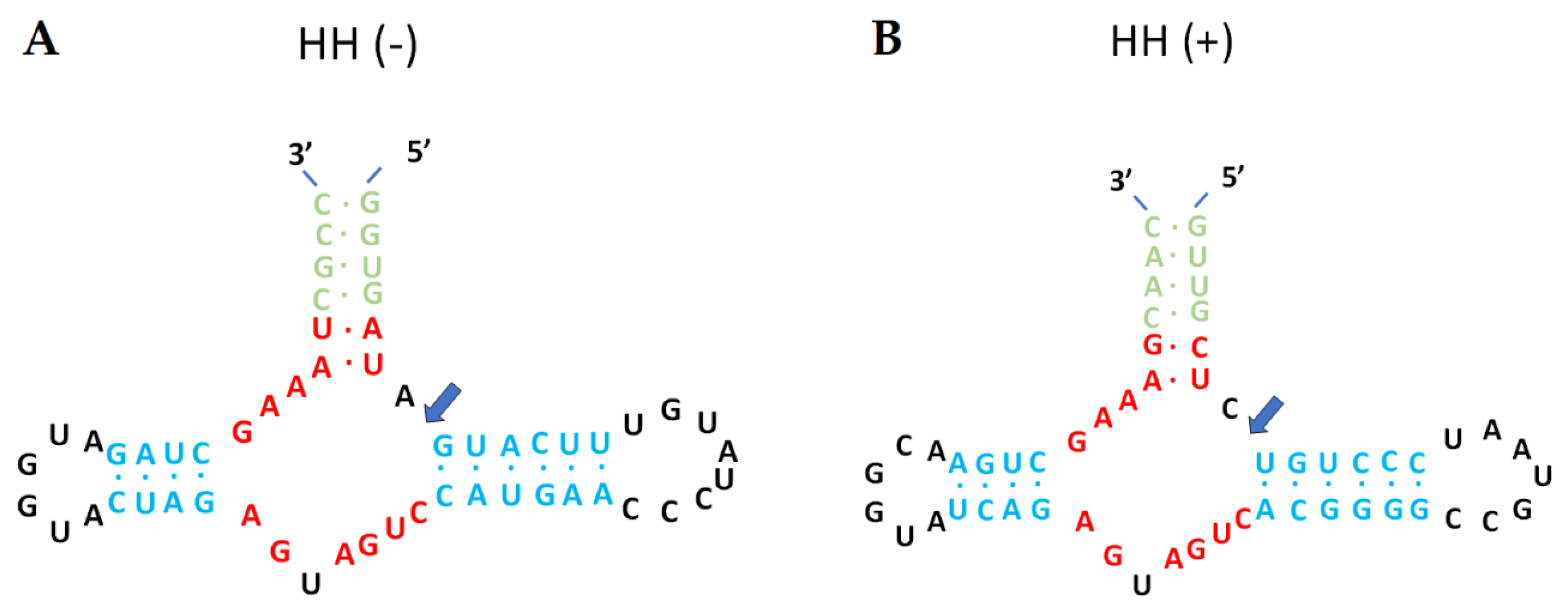
Figure A3.
Minimum free energy (MFE) prediction of the secondary structure of the FgMV2 RNA genome at the terminal regions. (A) Predicted structure of the 5’ end. (B) Energy released by folding three loops and a stem of the 5’ end. (C) Predicted structure of the and 3’ end. (D) Energy released by folding two loops at the 5’ end. The prediction was generated with RNAfold V2.6.3 [42].
Figure A3.
Minimum free energy (MFE) prediction of the secondary structure of the FgMV2 RNA genome at the terminal regions. (A) Predicted structure of the 5’ end. (B) Energy released by folding three loops and a stem of the 5’ end. (C) Predicted structure of the and 3’ end. (D) Energy released by folding two loops at the 5’ end. The prediction was generated with RNAfold V2.6.3 [42].
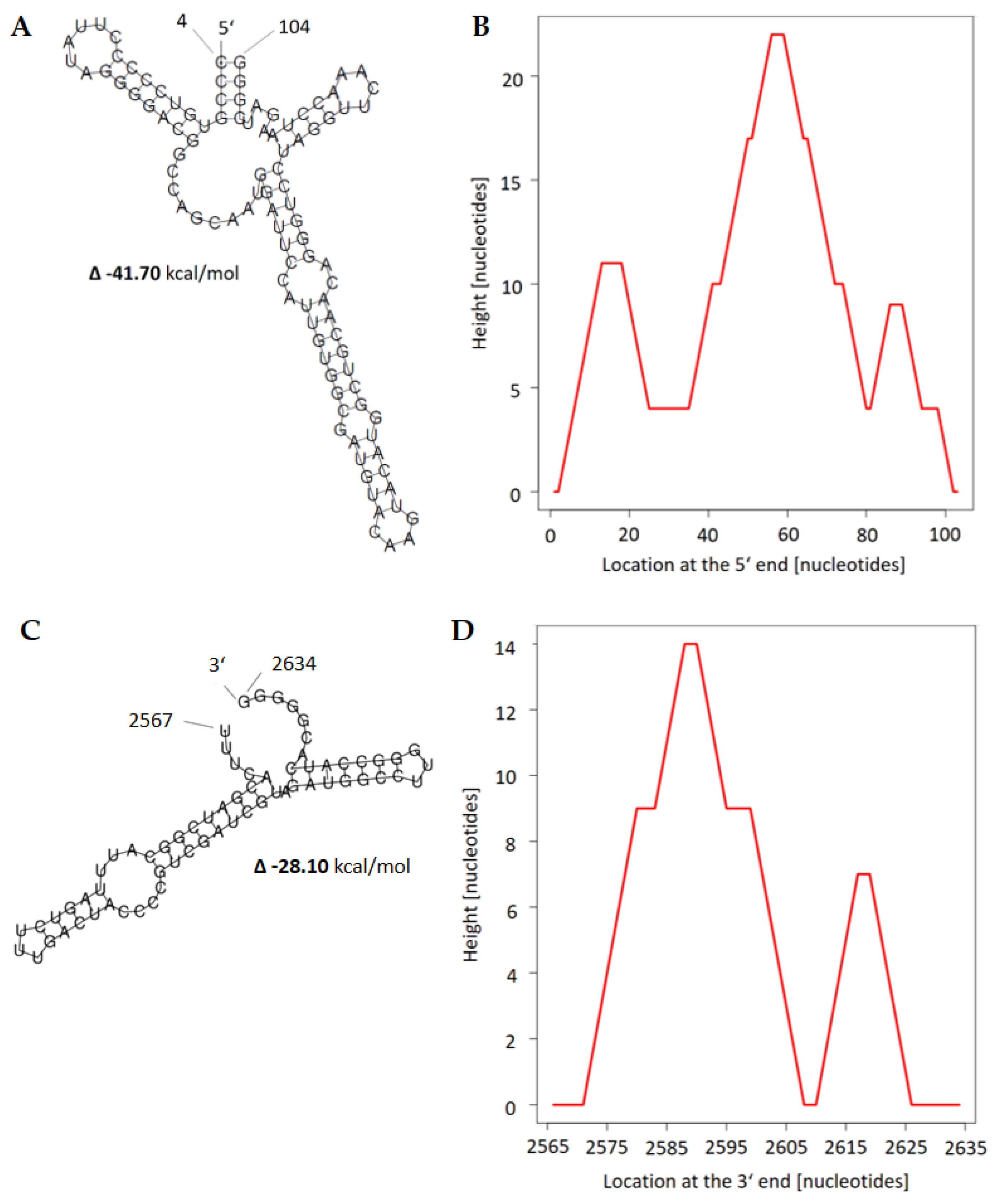
Figure A4.
Comparison of MS fragmentation patterns of F. graminearum sesquiterpene with RI = 1543.2 (above) and (+)-trichodiene standard (below). Mass-to-charge ratios are given for coincidental major peaks in clusters.
Figure A4.
Comparison of MS fragmentation patterns of F. graminearum sesquiterpene with RI = 1543.2 (above) and (+)-trichodiene standard (below). Mass-to-charge ratios are given for coincidental major peaks in clusters.
References
- Munkvold, G.P. Epidemiology of Fusarium diseases and their mycotoxins in maize ears. Eur. J. Plant Pathol. 2003, 109, 705–713. [Google Scholar] [CrossRef]
- Desjardins, A.E. Fusarium Mycotoxins: Chemistry, Genetics and Biology; American Phytopathological Society, St. Paul. MN, USA, 2006; ISBN 0-89-54-335-6.
- Ferrigo, D.; Raiola, A.; Causin, R. Fusarium toxins in cereals: Occurrence, legislation, factors promoting the appearance and their management. Molecules 2016, 21, 627. [Google Scholar] [CrossRef]
- Foroud, N.A.; Baines, D.; Gagkaeva, T.Y.; Thakor, N.; Badea, A.; Steiner, B.; Bürstmayr, M.; Bürstmayr, H. Trichothecenes in cereal grains – an update. Toxins 2019, 11, 634. [Google Scholar] [CrossRef]
- Yoshizawa, T. Thirty-five years of research on deoxynivalenol, a trichothecene mycotoxin: with special reference to its discovery and co-occurrence with nivalenol in Japan. Food Safety 2013, 1, 2013002. [Google Scholar] [CrossRef]
- Tatsuno, T.; Saito, M.; Enomoto, M.; Tsunoda, H. Nivalenol, a toxic principle of Fusarium nivale. Chem. Pharm. Bull. 1968, 16, 2519–2520. [Google Scholar] [CrossRef] [PubMed]
- Proctor, R.H.; Hohn, T.M.; McCormick, S.P. reduced virulence of Gibberella zeae caused by disruption of a trichothecene toxin biosynthetic gene. Mol Plant Microbe Interact 1995, 8, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Jansen, C.; von Wettstein, D.; Schäfer, W.; Kogel, K.-H.; Felk, A.; Maier, F.J. Infection patterns in barley and wheat spikes inoculated with wild-type and trichodiene synthase gene disrupted Fusarium graminearum. Proc. Natl. Acad. Sci. U.S.A. 2005, 102, 16892–16897. [Google Scholar] [CrossRef] [PubMed]
- Urry, W.H.; Wehrmeister, H.H.; Hodge, E.B.; Hidy, P.H. The structure of zearalenone. Tetrahedron Lett. 1966, 7, 3109–3114. [Google Scholar] [CrossRef]
- Utermark, J.; Karlovsky, P. Role of zearalenone lactonase in protection of Gliocladium roseum from fungitoxic effects of the mycotoxin zearalenone. Appl. Environ. Microbiol. 2007, 73, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Ashley, J.N.; Hobbs, B.C.; Raistrick, H. Studies in the biochemistry of micro-organisms: the crystalline colouring matters of Fusarium culmorum. Biochem. J. 1937, 31, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Vinas, M.; Alsarrag, A.; Su, L.; Pfohl, K.; Rohlfs, M.; Schäfer, W.; Chen, W.; Karlovsky, P. Bis-naphthopyrone pigments protect filamentous ascomycetes from a wide range of predators. Nat. Commun. 2019, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ghabrial, S.A.; Castón, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. Virology 2015, 479–480, 356–368.
- Hough, B.; Steenkamp, E.; Wingfield, B.; Read, D. Fungal viruses unveiled: a comprehensive review of mycoviruses. Viruses 2023, 15, 1202. [Google Scholar] [CrossRef] [PubMed]
- te Velthuis, A. J. W.; Common and unique features of viral RNA-dependent polymerases. Cell. Mol. Life Sci., 2014, 71: 4403-20.
- Liu, S.; Xie, J.; Cheng, J.; Li, B.; Chen, T.; Fu, Y.; Li, G.; Wang, M.; Jin, H.; Wan, H.; et al. Fungal DNA virus infects a mycophagous insect and utilizes it as a transmission vector. Proc. Natl. Acad. Sci. U. S. A. 2016, 113, 12803–12808. [Google Scholar] [CrossRef] [PubMed]
- Nuss, D.L. Hypovirulence: Mycoviruses at the fungal-plant interface. Nat. Rev. Microbiol. 2005, 3, 632–642. [Google Scholar] [CrossRef] [PubMed]
- Hillman, B.I.; Annisa, A.; Suzuki, N. Viruses of plant-interacting fungi. Adv. Virus Res. 2018, 100, 99–116. [Google Scholar] [PubMed]
- Pearson, M.N.; Beever, R.E.; Boine, B.; Arthur, K. Mycoviruses of filamentous fungi and their relevance to plant pathology. Mol. Plant Pathol. 2009, 10, 115–128. [Google Scholar] [CrossRef] [PubMed]
- García-Pedrajas, M.D.; Cañizares, M.C.; Sarmiento-Villamil, J.L.; Jacquat, A.G.; Dambolena, J.S. Mycoviruses in biological control: from basic research to field implementation. Phytopathology 2019, 109, 1828–1839. [Google Scholar] [CrossRef]
- Chu, Y.; Jeon, J.; Yea, S.; Kim, Y.; Yun, S.; Lee, Y.; Kim, K. Double-stranded RNA mycovirus from Fusarium graminearum. Appl. Environ. Microbiol. 2002, 68, 2529–2534. [Google Scholar] [CrossRef]
- Darissa, O.; Adam, G.; Schäfer, W. A dsRNA mycovirus causes hypovirulence of Fusarium graminearum to wheat and maize. Eur. J. Plant Path. 2012, 134, 181–189. [Google Scholar] [CrossRef]
- Li, P.; Zhang, H.; Chen, X.; Qiu, D.; Guo, L. Molecular characterization of a novel hypovirus from the plant pathogenic fungus Fusarium graminearum. Virology 2015, 481, 151–160. [Google Scholar] [CrossRef]
- Kotta-Loizou, I.; Coutts, R.H.A. Studies on the virome of the entomopathogenic fungus Beauveria bassiana reveal novel dsRNA elements and mild hypervirulence. PLoS Pathog. 2017, 13, 1–19. [Google Scholar] [CrossRef]
- Lee, K.-M.; Cho, W.K.; Yu, J.; Son, M.; Choi, H.; Min, K.; Lee, Y.-W.; Kim, K.-H. A comparison of transcriptional patterns and mycological phenotypes following infection of Fusarium graminearum by four mycoviruses. PLOS One 2014, 9, e100989. [Google Scholar] [CrossRef]
- Nerva, L.; Chitarra, W.; Siciliano, I.; Gaiotti, F.; Ciuffo, M.; Forgia, M.; Varese, G.C.; Turina, M. Mycoviruses mediate mycotoxin regulation in Aspergillus ochraceus. Environ. Microbiol. 2019, 21, 1957–1968. [Google Scholar] [CrossRef] [PubMed]
- Ninomiya, A.; Urayama, S.; Suo, R.; Itoi, S.; Fuji, S.; Moriyama, H.; Hagiwara, D. Mycovirus-induced tenuazonic acid production in a rice blast fungus Magnaporthe oryzae. Front. Microbiol. 2020, 11, 1641. [Google Scholar] [CrossRef] [PubMed]
- Schiwek, S.; Beule, L.; Vinas, M.; Pfordt, A.; von Tiedemann, A.; Karlovsky, P. High-resolution melting (HRM) curve assay for the identification of eight Fusarium species causing ear rot in maize. Pathogens 2020, 9, 270. [Google Scholar] [CrossRef]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. U. S. A. 1998, 95, 2044–2049. [Google Scholar] [CrossRef]
- Forgia, M.; Navarro, B.; Daghino, S.; Cervera, A.; Gisel, A.; Perotto, S.; Aghayeva, D.N.; Akinyuwa, M.F.; Gobbi, E.; Zheludev, I.N.; et al. Hybrids of RNA viruses and viroid-like elements replicate in fungi. Nat. Commun. 2023, 14, 2591. [Google Scholar] [CrossRef]
- Sutela, S., Forgia, M., Vainio, E.J., Chiapello, M., Daghino, S., Vallino, M., Martino, E., Girlanda, M., Perotto, S., Turina, M., The virome from a collection of endomycorrhizal fungi reveals new viral taxa with unprecedented genome organization. Virus Evolution 2020, 6, 76.
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Gonnet, G.H.; Cohen, M.A.; Benner, S.A. Exhaustive matching of the entire protein sequence database. Science 1992, 256, 1443–1445. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Comp Appl Biosci 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef]
- Pringle, C.R.; Easton, A.J. Monopartite negative strand RNA genomes. Seminars in Virology 1997, 8, 49–57. [Google Scholar] [CrossRef]
- Liu, L.; Xie, J.; Cheng, J.; Fu, Y.; Li, G.; Yi, X.; Jiang, D. Fungal negative-stranded RNA virus that is related to Bornaviruses and Nyaviruses. Proc. Nat. Acad. Sci. U.S.A. 2014, 111, 12205–12210. [Google Scholar] [CrossRef]
- Wang, L.; He, H.; Wang, S.; Chen, X.; Qiu, D.; Kondo, H.; Guo, L. Evidence for a novel negative-stranded RNA mycovirus isolated from the plant pathogenic fungus Fusarium graminearum. Virology 2018, 518, 232–240. [Google Scholar] [CrossRef]
- Xu, Z.; Wu, S.; Liu, L.; Cheng, J.; Fu, Y.; Jiang, D.; Xie, J. A mitovirus related to plant mitochondrial gene confers hypovirulence on the phytopathogenic fungus Sclerotinia sclerotiorum. Virus Res. 2015, 197, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Gruber, A.R.; Lorenz, R.; Bernhart, S.H.; Neuböck, R.; Hofacker, I.L. The Vienna RNA websuite. Nucleic Acids Research 2008, 36, W70–W74. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, M.E.; Pearson, M.N. Molecular characterization of three mitoviruses co-infecting a hypovirulent isolate of Sclerotinia sclerotiorum fungus. Virology 2013, 441, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.A.; Baig, D.I.; Bhatti, M.F. An overview of mycoviral curing strategies used in evaluating fungal host fitness. Mol. Biotechnol. 2023, 65, 1547–1564. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.M.; Lim, W.S.; Yea, S.J.; Cho, J.D.; Lee, Y.W.; Kim, K.H. Complexity of dsRNA mycovirus isolated from Fusarium graminearum. Virus Genes 2004, 28, 135–143. [Google Scholar] [CrossRef]
- Bormann, J.; Heinze, C.; Blum, C.; Mentges, M.; Brockmann, A.; Alder, A.; Landt, S.K.; Josephson, B.; Indenbirken, D.; Spohn, M.; et al. Expression of a structural protein of the mycovirus FgV-ch9 negatively affects the transcript level of a novel symptom alleviation factor and causes virus infection-like symptoms in Fusarium graminearum. J. Virol. 2018, 92, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Lee, K.M.; Son, M.; Kim, K.H. Effects of the deletion and over-expression of Fusarium graminearum gene FgHal2 on host response to mycovirus Fusarium graminearum virus 1. Mol. Plant Pathol. 2015, 16, 641–652. [Google Scholar] [CrossRef]
- Kim, J.-E.; Han, K.-H.; Jin, J.; Kim, H.; Kim, J.-C.; Yun, S.-H.; Lee, Y.-W. Putative polyketide synthase and laccase genes for biosynthesis of aurofusarin in Gibberella zeae. Appl. Env. Microbiol. 2005, 71, 1701–1708. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.-M.; Lee, J.; Lee, Y.-W. Characterization of carotenoid biosynthetic genes in the ascomycete Gibberella zeae. FEMS Microbiol. Lett. 2010, 302, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Moonjely, S., Ebert, M., Paton-Glassbrook, D., Noel, Z. A., Roze, L., Shay, R., Watkins, T., Trail, F. Update on the state of research to manage Fusarium head blight. Fungal Genet. Biol. 2023, 169, 103829. [CrossRef]
- Son, M.; Yu, J.; Kim, K.-H. Five Questions about mycoviruses. PLOS Pathogens 2015, 11, e1005172. [Google Scholar] [CrossRef] [PubMed]
- Szabó, B.; Kocsis, R.; Mézes, M. Reproduction inhibiting effects of deoxynivalenol or T-2 toxin contaminated maize on Folsomia candida (Collembola). Acta Zoologica Academiae Scientiarum Hungaricae 2019, 65, 323–334.
- Szabó, B., Bálint, B., Mézes, M., Balogh, K. Agricultural trichothecene mycotoxin contamination affects the life-history and reduced glutathione content of Folsomia candida Willem (Collembola). Acta Zoologica Academiae Scientiarum Hungaricae 2020, 66, 379–392.
- Karlovsky, P. Biological detoxification of fungal toxins and its use in plant breeding, feed and food production. Natural Toxins 1999, 7, 1–23. [Google Scholar] [CrossRef]
- Staaden, S.; Milcu, A.; Rohlfs, M.; Scheu, S. Olfactory cues associated with fungal grazing intensity and secondary metabolite pathway modulate collembola foraging behaviour. Soil Biol. Biochem. 2011, 43, 1411–1416. [Google Scholar] [CrossRef]
- Hedlund, K.; Bengtsson, G.; Rundgren, S. Fungal odour discrimination in two sympatric species of fungivorous Collembolans. Funct. Ecol. 1995, 9, 869–875. [Google Scholar] [CrossRef]
- Bengtsson, G.; Hedlund, K.; Rundgren, S. selective odor perception in the soil Collembola Onychiurus armatus. J. Chem. Ecol. 1991, 17, 2113–2125. [Google Scholar] [CrossRef]
- Nebapure, S.M.; Shankarganesh, K.; Rajna, S.; Naga, K.C.; Pandey, D.; Gambhir, S.; Praveen, K.V.; Subramanian, S. Dynamic changes in virus-induced volatiles in cotton modulate the orientation and oviposition behavior of the whitefly Bemisia tabaci. Front. Physiol. 2022, 13, 1–16. [Google Scholar] [CrossRef]
- Werner, B.J.; Mowry, T.M.; Bosque-Pérez, N.A.; Ding, H.; Eigenbrode, S.D. Changes in green peach aphid responses to potato leafroll virus induced volatiles emitted during disease progression. Environ. Entomol. 2009, 38, 1429–1438. [Google Scholar] [CrossRef]
- Sherif, M.; Kirsch, N.; Splivallo, R.; Pfohl, K.; Karlovsky, P. The role of mycotoxins in interactions between Fusarium graminearum and F. verticillioides growing in saprophytic cultures and co-infecting maize plants. Toxins 2023, 15, 575. [CrossRef]
- Pfordt, A.; Ramos Romero, L.; Schiwek, S.; Karlovsky, P.; von Tiedemann, A. Impact of environmental conditions and agronomic practices on the prevalence of Fusarium species associated with ear- and stalk rot in maize. Pathogens 2020, 9, 236. [Google Scholar] [CrossRef]
- Lysøe, E.; Klemsdal, S.S.; Bone, K.R.; Frandsen, R.J.N.; Johansen, T.; Thrane, U.; Giese, H. The PKS4 Gene of Fusarium graminearum is essential for zearalenone production. Appl Environ. Microbiol. 2006, 72, 3924–3932. [Google Scholar] [CrossRef]
- Maniatis, T., Fritsch, E. F. & Sambrook, J. Molecular Cloning – a Laboratory Manual. Cold Spring Harbor Laboratory: New York, 1989, pp. 94-103.
- Pielhop, T.P.; Popp, C.; Knierim, D.; Margaria, P.; Maiß, E. Three new mycoviruses identified in the apple replant disease (ARD)-associated fungus Rugonectria rugulosa. Virus Genes, 2022,. [CrossRef]
- Koonin, E. V; Senkevich, T.G.; Dolja, V. V The ancient virus world and evolution of cells. Biol. Direct 2006, 1, 27. [Google Scholar]
- Nasir, A.; Caetano-Anollés, G. A phylogenomic data-driven exploration of viral origins and evolution. Sci. Adv. 2015, 1, e1500527. [Google Scholar] [CrossRef] [PubMed]
- Brandfass, C.; Karlovsky, P. Upscaled CTAB-Based DNA extraction and real-time PCR Assays for Fusarium culmorum and F. graminearum DNA in plant material with reduced sampling error. Int. J. Mol. Sci. 2008, 9, 2306–2321. [Google Scholar] [CrossRef] [PubMed]
- Nirenberg, H. Untersuchungen über die morphologische und biologische Differenzierung in der Fusarium-Sektion Liseola. Mitt. Biol. Bundesanst. Land und Forstwirtsch. Berlin-Dahlem 1976, 169, 1–117.
- Schiwek, S.; Alhussein, M.; Rodemann, C.; Budragchaa, T.; Beule, L.; von Tiedemann, A.; Karlovsky, P. Fusarium culmorum produces NX-2 toxin simultaneously with deoxynivalenol and 3-acetyl-deoxynivalenol or nivalenol. Toxins 2022, 14, 456. [Google Scholar] [CrossRef] [PubMed]
- Osaki, H.; Sasaki, A.; Nomiyama, K.; Tomioka, K. Multiple virus infection in a single strain of Fusarium poae shown by deep sequencing. Virus Genes 2016, 52, 835–847. [Google Scholar] [CrossRef]
- Nicholson, P.; Simpson, D.R.; Weston, G.; Rezanoor, H.N.; Lees, A.K.; Parry, D.W.; Joyce, D. Detection and quantification of Fusarium culmorum and Fusarium graminearum in cereals using PCR assays. Physiol. Mol. Plant Path. 1998, 53, 17–37. [Google Scholar] [CrossRef]
Figure 1.
DNase digestion of nucleic acids extracted from F. graminearum 266MV.
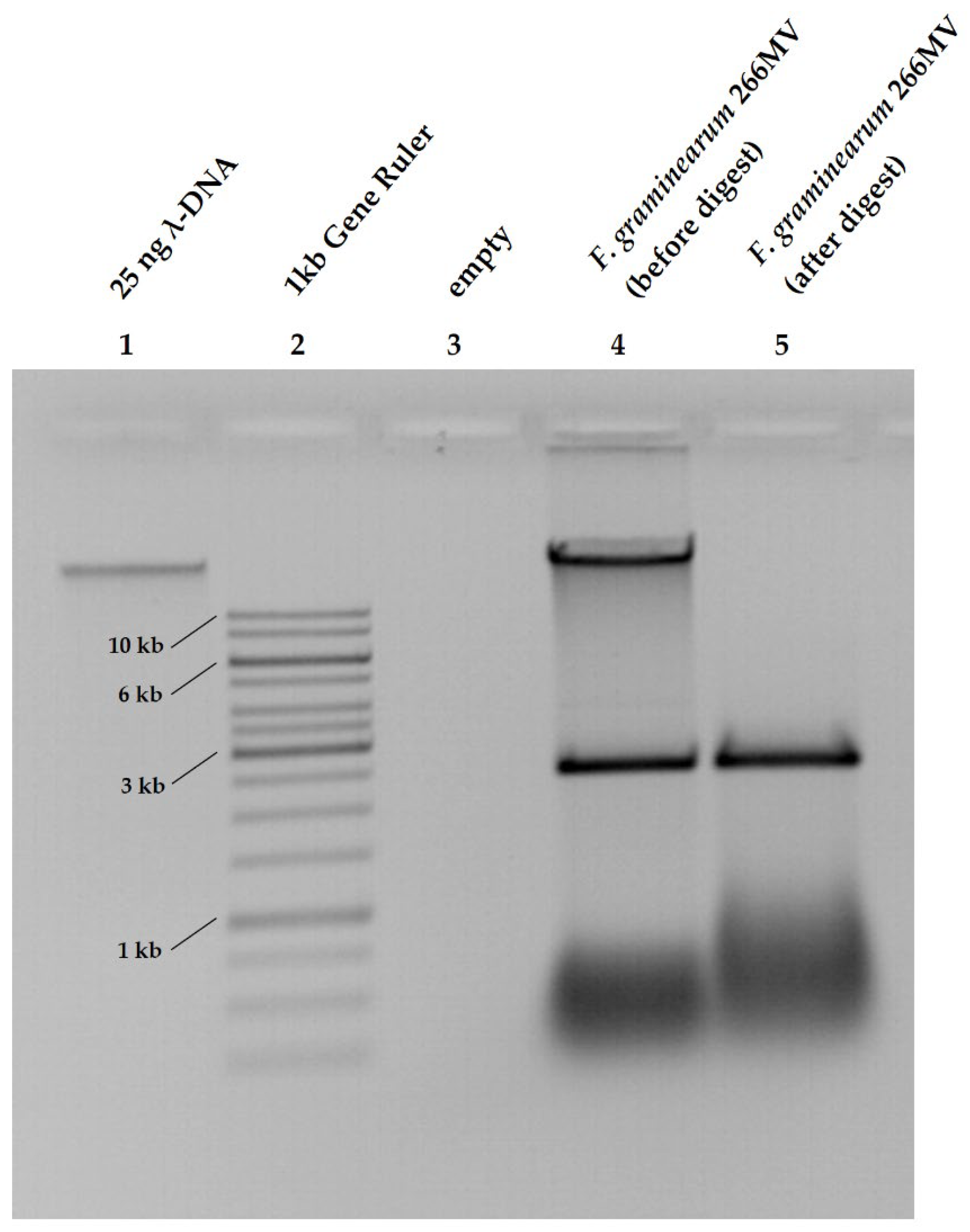
Figure 2.
Comparison of F. graminearum 266MV and 266 differing in the presence of an extrachromosomal RNA element. Left: growth pattern of 266MV (A) and 266 (B) on PDA 14 d after inoculation. Right (C): Agarose gel electrophoresis of total RNA extracted from strains 266 and 266MV.
Figure 2.
Comparison of F. graminearum 266MV and 266 differing in the presence of an extrachromosomal RNA element. Left: growth pattern of 266MV (A) and 266 (B) on PDA 14 d after inoculation. Right (C): Agarose gel electrophoresis of total RNA extracted from strains 266 and 266MV.
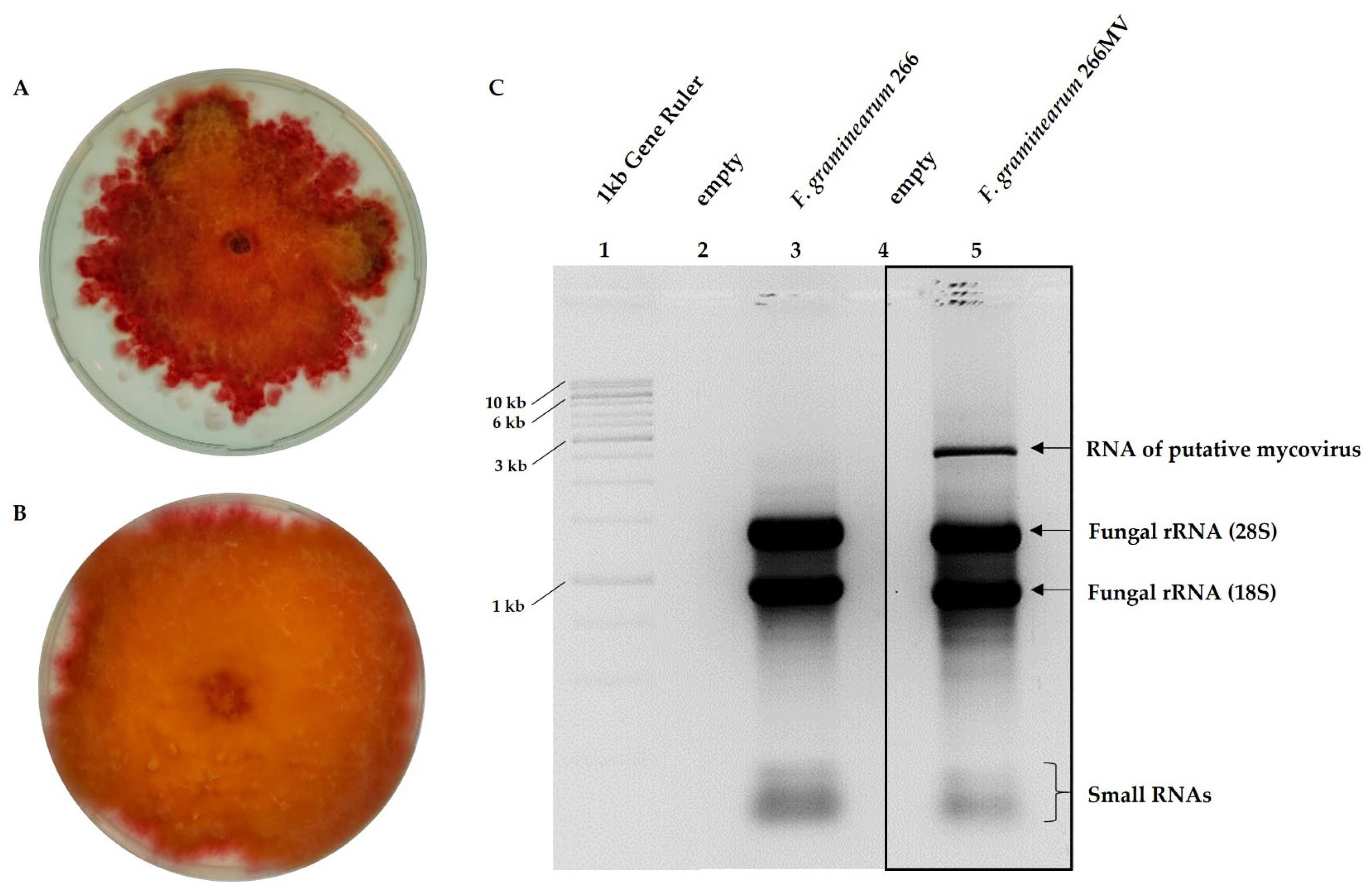
Figure 3.
Transmission electron micrograph of virus-like particles purified from the mycelium of Fusarium graminearum carrying mycoviruses. (A) Wild type strain 266MV; (B) strain 668MV into which mycoviruses have been transfected (Section 3.4). Isometric particles of approximately 38 nm resemble Betapartitiviruses, such as Fusarium poae virus 1 (FpV1). The bar represents 100 nm.
Figure 3.
Transmission electron micrograph of virus-like particles purified from the mycelium of Fusarium graminearum carrying mycoviruses. (A) Wild type strain 266MV; (B) strain 668MV into which mycoviruses have been transfected (Section 3.4). Isometric particles of approximately 38 nm resemble Betapartitiviruses, such as Fusarium poae virus 1 (FpV1). The bar represents 100 nm.
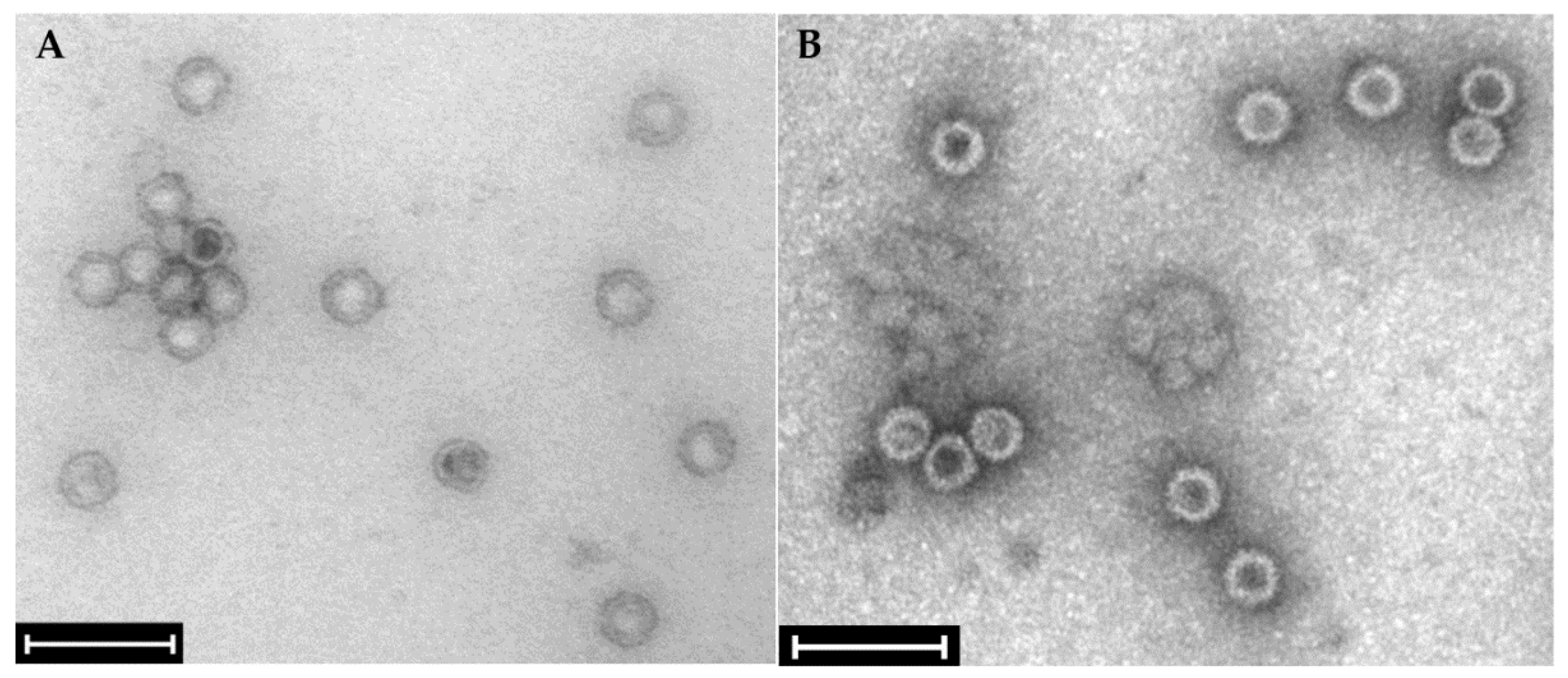
Figure 5.
Genome organization of Fusarium graminearum negative-stranded RNA virus 2 (FgNSRV2). (A) Schematic representation of the genome of FgNSRV2, including five open reading frames (ORFs). Conserved functional domains and accession numbers, identified in BLAST/CDD analysis, are indicated above the ORFs [37]. (B) Alignment of the sequence of ORF1 from FgNSRV2 with highly similar sequences from SlNSRV1 (ALM62220) and FgNSRV1 (ATP75709) from NCBI. Full species names and accession Nos. are shown in Appendix Table A2. (C) Dendrogram constructed from the sequences of ORF4, putatively encoding RdRp, and similar sequences obtained from NCBI. For the construction of the alignment and dendrogram, symbols for amino acid similarity, bootstrap values, and the scale refer to Figure 4.
Figure 5.
Genome organization of Fusarium graminearum negative-stranded RNA virus 2 (FgNSRV2). (A) Schematic representation of the genome of FgNSRV2, including five open reading frames (ORFs). Conserved functional domains and accession numbers, identified in BLAST/CDD analysis, are indicated above the ORFs [37]. (B) Alignment of the sequence of ORF1 from FgNSRV2 with highly similar sequences from SlNSRV1 (ALM62220) and FgNSRV1 (ATP75709) from NCBI. Full species names and accession Nos. are shown in Appendix Table A2. (C) Dendrogram constructed from the sequences of ORF4, putatively encoding RdRp, and similar sequences obtained from NCBI. For the construction of the alignment and dendrogram, symbols for amino acid similarity, bootstrap values, and the scale refer to Figure 4.
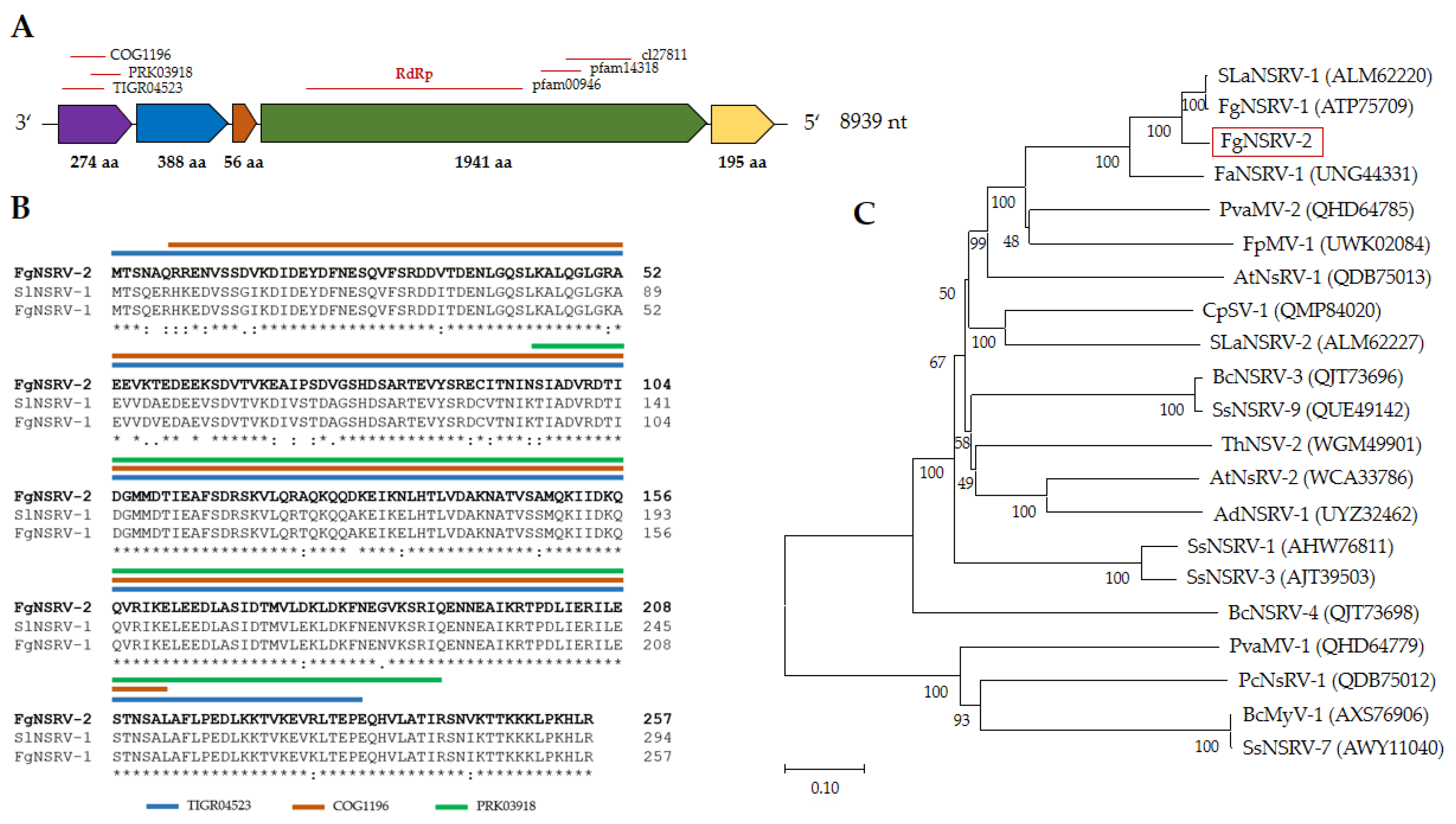
Figure 6.
Alignment of the gene-junction sequences of FgNSRV2 with SsNSRV-1 (KJ186782, [39]) and FgNSRV-1 [40]. Nucleotide sequences of ORFs are labelled according to Figure 5. Numbers indicate the position of the first and last nucleotide of the motif in the genome. The conserved gene-junction sequences (A/U/G)(G/U/A/C)UAUU(U/A)AA(U/G)AAAACUUAGG(A/U)(G/U) is highlighted bold.
Figure 6.
Alignment of the gene-junction sequences of FgNSRV2 with SsNSRV-1 (KJ186782, [39]) and FgNSRV-1 [40]. Nucleotide sequences of ORFs are labelled according to Figure 5. Numbers indicate the position of the first and last nucleotide of the motif in the genome. The conserved gene-junction sequences (A/U/G)(G/U/A/C)UAUU(U/A)AA(U/G)AAAACUUAGG(A/U)(G/U) is highlighted bold.

Figure 7.
Genome of Fusarium graminearum mitovirus 2 (FgMV-2). (A) Single ORF encoding RdRp. (B) Alignment of conserved motifs and (C) dendrogram of RdRp sequences. For the explanation of symbols and description of the software, see Figure 4.
Figure 7.
Genome of Fusarium graminearum mitovirus 2 (FgMV-2). (A) Single ORF encoding RdRp. (B) Alignment of conserved motifs and (C) dendrogram of RdRp sequences. For the explanation of symbols and description of the software, see Figure 4.
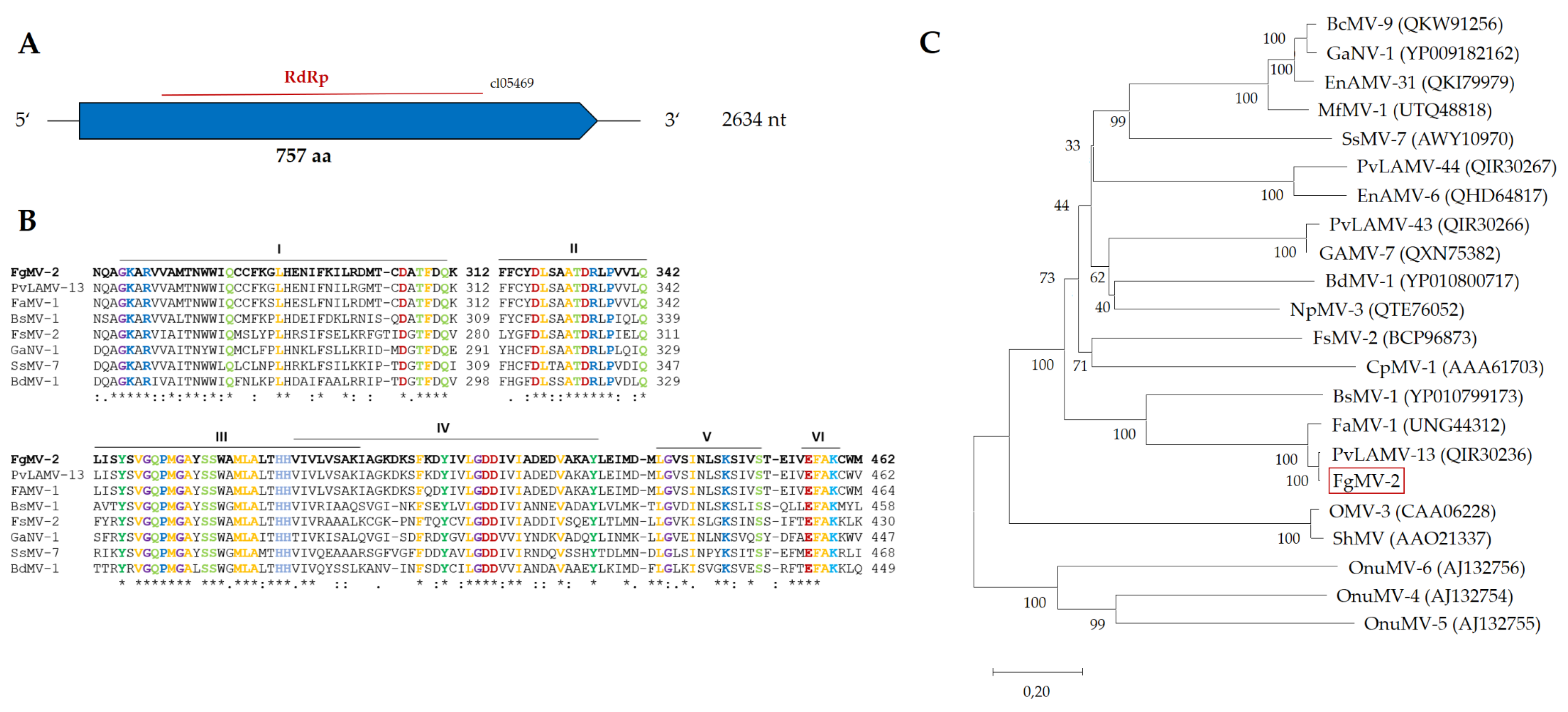
Figure 8.
Effect of mycoviral infection on radial growth and sporulation capacity of F. graminarum. In each panel, results obtained with two isogenic strains differing merely in the presence of mycoviruses are shown. Radial growth rates were estimated on PDA after 48 h of incubation at 25 °C in the dark. Sporulation capacity was estimated by growing cultures in liquid media for 7 d at 25 °C (Section 5.5.). n.s. stands for p > 0.1.
Figure 8.
Effect of mycoviral infection on radial growth and sporulation capacity of F. graminarum. In each panel, results obtained with two isogenic strains differing merely in the presence of mycoviruses are shown. Radial growth rates were estimated on PDA after 48 h of incubation at 25 °C in the dark. Sporulation capacity was estimated by growing cultures in liquid media for 7 d at 25 °C (Section 5.5.). n.s. stands for p > 0.1.
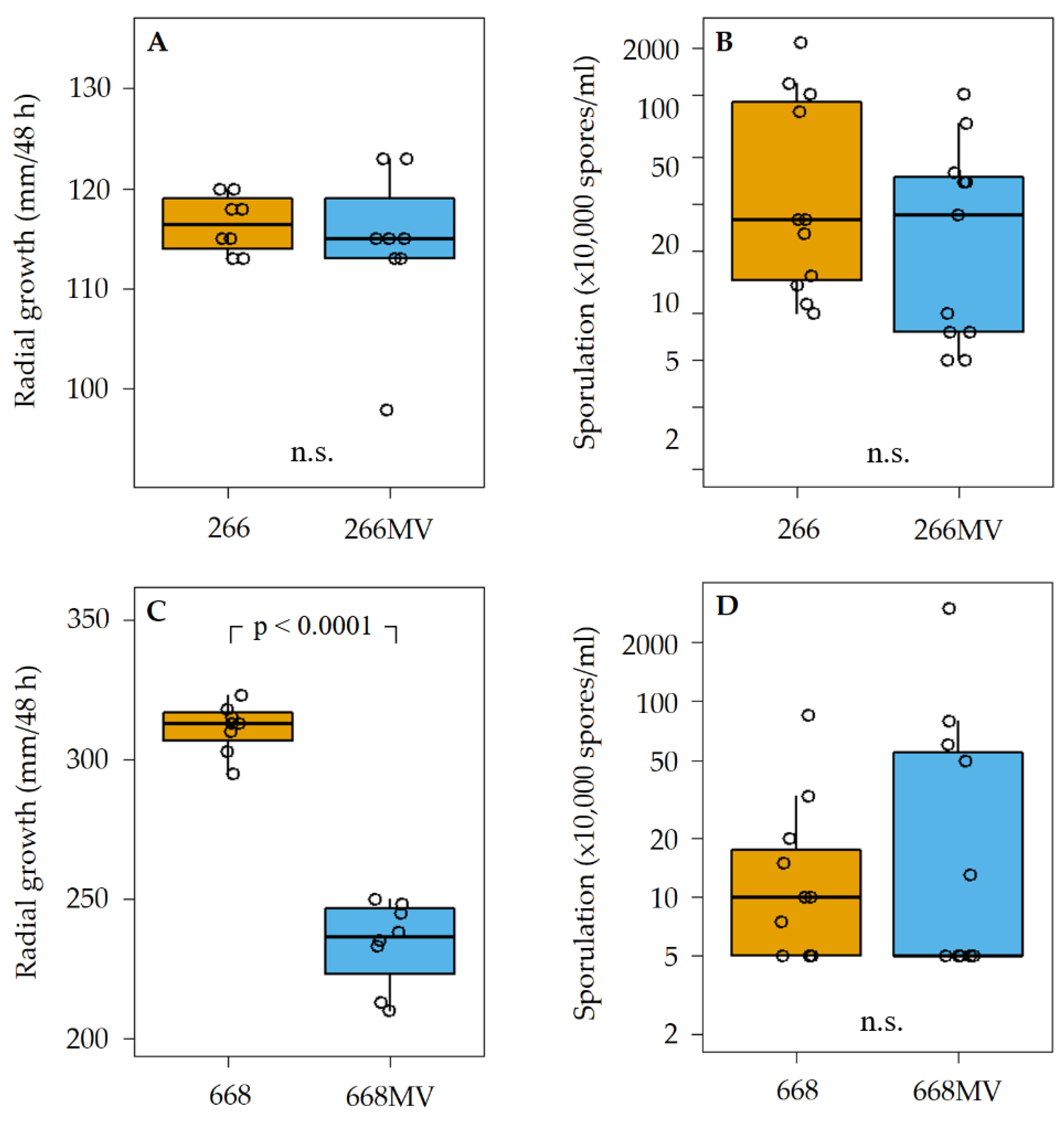
Figure 9.
Effect of infection with mycoviruses on the production of trichothecenes and aurofusarin by F. graminearum. Concentrations are expressed in mg/kg dry weight of rice cultures normalized to fungal biomass. For details regarding culture conditions, metabolite extraction, and HPLC-MS/MS analysis, see Section 5.6.
Figure 9.
Effect of infection with mycoviruses on the production of trichothecenes and aurofusarin by F. graminearum. Concentrations are expressed in mg/kg dry weight of rice cultures normalized to fungal biomass. For details regarding culture conditions, metabolite extraction, and HPLC-MS/MS analysis, see Section 5.6.
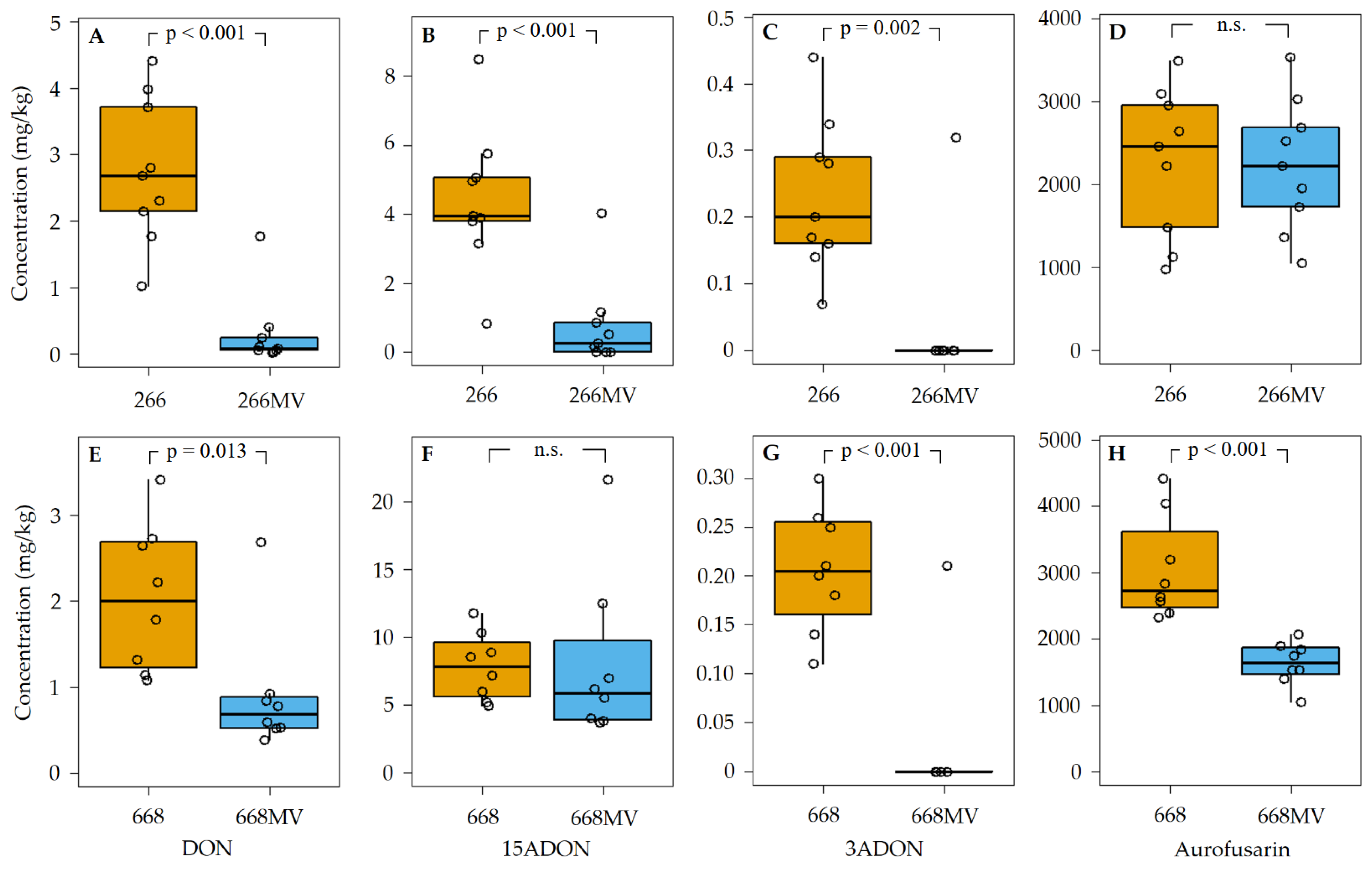
Figure 10.
Effect of mycoviral infection on the emission of volatile metabolites by Fusarium graminearum. (A) Comparison of strains 266 and 266MV. (B) Comparison of strains 668 and 668MV. Sample collection and analysis is described in Section 5.7. Statistic comparison between treatments were performed with Students t-test or Wilcoxon ranked test, according to data structure. Boxplot whiskers extend to the most extend data point within 1.5 interquartile range. The upper box limit represents 3rd quartile, the lower box limit represents 1st quartile, and the box line represents the 2nd quartile.
Figure 10.
Effect of mycoviral infection on the emission of volatile metabolites by Fusarium graminearum. (A) Comparison of strains 266 and 266MV. (B) Comparison of strains 668 and 668MV. Sample collection and analysis is described in Section 5.7. Statistic comparison between treatments were performed with Students t-test or Wilcoxon ranked test, according to data structure. Boxplot whiskers extend to the most extend data point within 1.5 interquartile range. The upper box limit represents 3rd quartile, the lower box limit represents 1st quartile, and the box line represents the 2nd quartile.
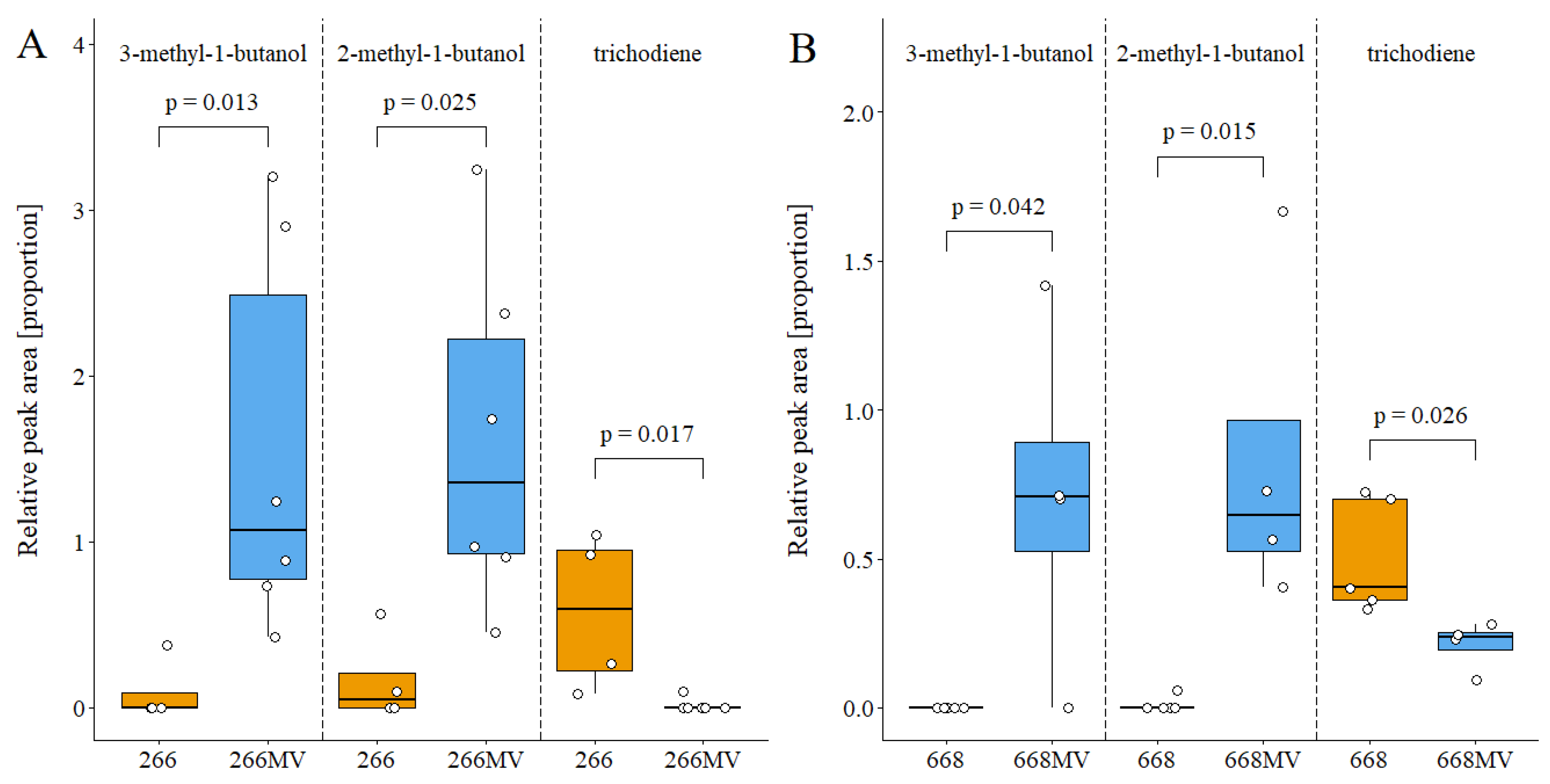
Figure 11.
Effect of mycoviral infection on food preference of Folsomia candida. (A) Strains 266 and 266MV; (B) strains 668 and 668MV. Food choice was determined as the proportion of responding individuals that fed on the mycelium of a particular strain. Response, represented by black dots, was defined as the proportion of individuals feeding on mycelial plugs.
Figure 11.
Effect of mycoviral infection on food preference of Folsomia candida. (A) Strains 266 and 266MV; (B) strains 668 and 668MV. Food choice was determined as the proportion of responding individuals that fed on the mycelium of a particular strain. Response, represented by black dots, was defined as the proportion of individuals feeding on mycelial plugs.
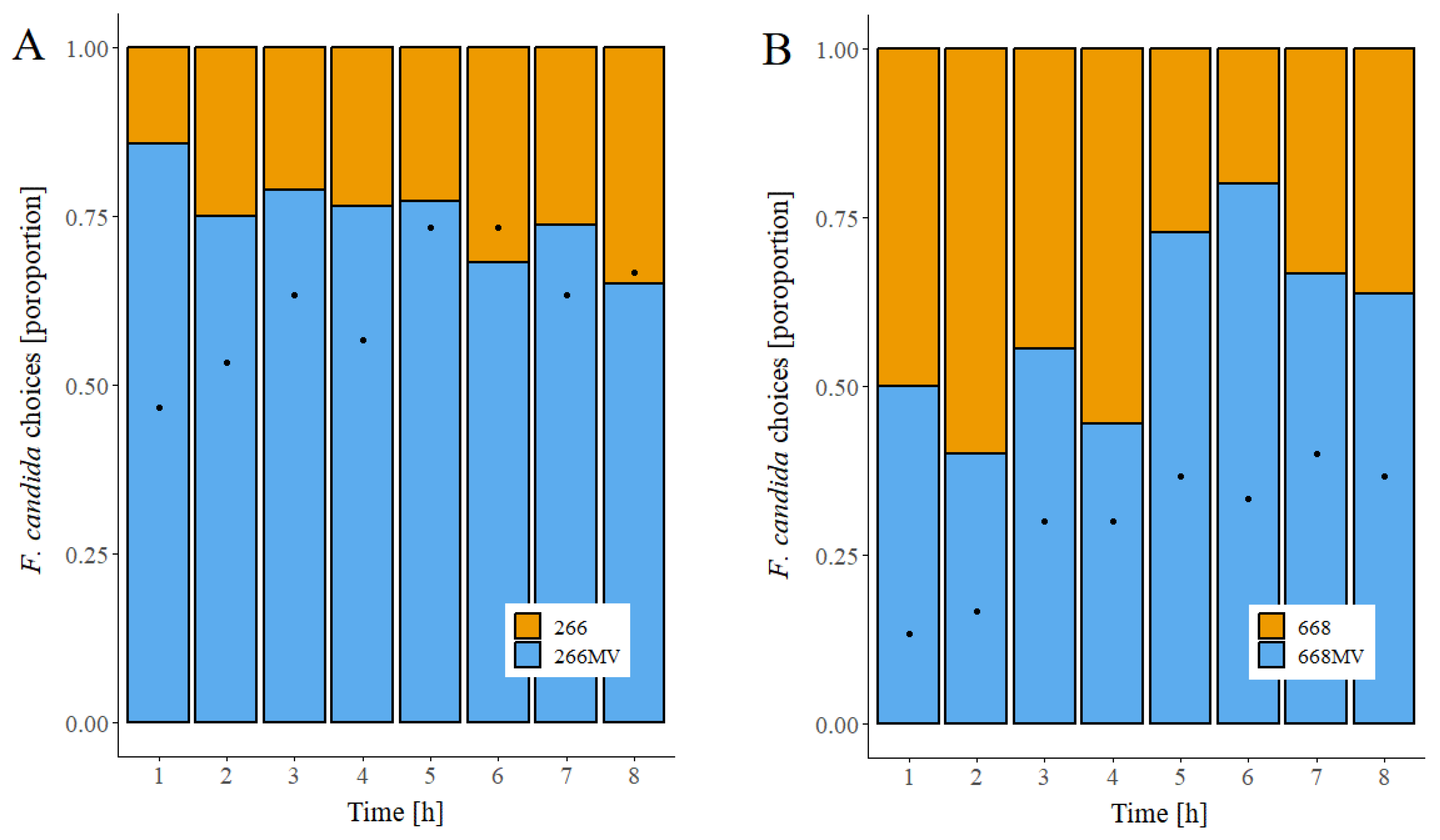
Table 1.
Mycovirus sequences assembled from F. graminearum 266MV.
| Name | Size [nt] | Accession no. | Most similar genomes by NCBI BLASTn | |||
| Closest reference sequenceB | Accession no. | Genome [nt] | Similarity [%] | |||
| FpV1 | 2298 | ON969095 | Fusarium poae virus 1 RNA 1 | LC150606 | 2431 | 98.6 |
| 2205 | ON969096 | Fusarium poae virus 1 RNA 2 | OK524181 | 2212 | 98.8 | |
| FgNSRV2 | 8939 | ON969097 | Soybean leaf-associated negative-stranded RNA virus | NC075296 | 9041 | 98.0 |
| FgMV2 | 2634 | ON969098 | Plasmopara viticola lesion associated mitovirus 13 | MN539774 | 2607 | 96.3 |
| FgAV1 | 4579 | ON969099 | Cryphonectria parasitica ambivirus 1 | MT354566 | 4579 | 73.0 |
Table 2.
Presence of mycoviruses in F. graminearum strains.
| Strain | FpV1 | FgNSRV1 | FgMV2 | FgAV1 |
| 266 | yes | no | yes | no |
| 266MV | yes | yes | yes | yes |
| 668 | no | no | no | no |
| 668MV | yes | no | no | yes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



