
Submitted:
05 January 2024
Posted:
05 January 2024
You are already at the latest version
Abstract
DNA-binding transcription factors are broadly characterized as proteins that bind to specific sequences within genomic DNA and modulate the expression of downstream genes. This study focused on KojR—a transcription factor involved in the metabolism of kojic acid, which is an organic acid synthesized in Aspergillus oryzae and is known for its tyrosinase-inhibitory properties. However, the regulatory mechanism underlying KojR-mediated kojic acid synthesis in remains unclear. Hence, we aimed to obtain a comprehensive identification of KojR-associated genes using genomic Systematic Evolution of Ligands by Exponential enrichment (gSELEX-Seq) and RNA-Seq. During the genome-wide exploration of KojR-binding sites via gSELEX-Seq and identification of KojR-dependent differentially expressed genes (DEGs) using RNA-Seq, we confirmed that KojR directly regulates kojT, as was previously reported. We also observed that kojA expression, which may be controlled by KojR, was significantly reduced in a ΔkojR strain. Notably, no binding of KojR to the kojA promoter region was detected. Furthermore, certain KojR-dependent DEGs identified in the present study were associated with enzymes implicated in the carbon metabolic pathway of A. oryzae. This strongly indicates that KojR plays a central role in carbon metabolism in A. oryzae.
Keywords:
Transcriptome
; Transcription factor
; Aspergillus oryzae
; Kojic acid (KA)
; Bioinformatics
1. Introduction
Kojic acid (KA) is highly valuable in several industries. For example, in the cosmetics industry, it is popular for use in skin lightening applications owing to its ability to inhibit tyrosinase and reduce melanin production [1]. Additionally, the antibacterial, antifungal, and anti-inflammatory properties of KA and its derivatives increase its potential for use in the pharmaceutical industry [2].
In Japan, Aspergillus oryzae, has been historically used in the brewing and food industries, produces KA as a secondary metabolite [3] and can be used in fermentative KA production. However, the KA synthesis pathway in A. oryzae remains unclear despite various pathways having been suggested. In fact, a consensus is yet to be reached even a century after the initial observation was made [3]. An interesting review on fungal biosynthesis of KA was published very recently, and the article presented the metabolic pathways of KA [4]; however, the genes pertaining to each metabolic reaction are yet to be identified.
A. oryzae houses kojR (AO090113000137), which encodes a Zn(II)2Cys6 transcriptional activator that is integral to the KA biosynthesis [5,6]. kojR deletion significantly downregulates KA synthesis [6,7]. kojR is located between kojA (AO090113000136) and kojT (AO090113000138) and plays a pivotal role in regulating their expression [5,6,7]. Chang et al. identified an 11-bp palindromic sequence within the kojA and kojT promoter regions in 23 Aspergillus species including A. oryzae and A. flavus [8]. They prepared A. flavus mutants using CRISPR/Cas9 by deleting this consensus sequences in the kojA and kojT promoters and showed that several of these mutants lost their ability to synthesize KA [8]. These results have suggested that KojR directly regulates kojA and kojT expression in A. flavus.
Additionally, KojR is involved in gliotoxin biosynthesis in A. fumigatus and protects against gliotoxin in A. fumigatus, A. nidulans, and A. oryzae [9], implying that the regulatory mechanism of KojR is not confined to the group of genes associated with KA biosynthesis. However, the KojR binding sites in the genome and the genes whose expression is directly regulated by KojR have not been comprehensively investigated.
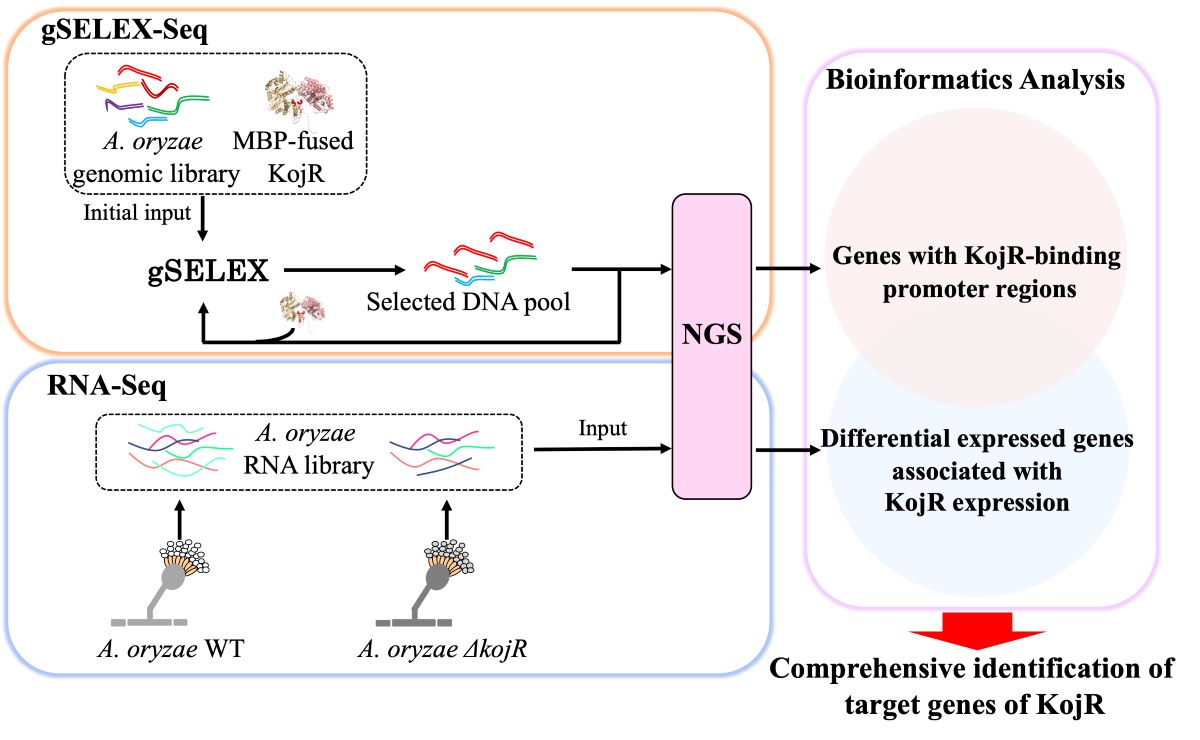
We have previously used the genomic systematic evolution of ligands by exponential enrichment with high-throughput DNA sequencing (gSELEX-Seq) technique and a differential expression analysis method, such as DNA microarray analysis or RNA-Seq, and established a system for the comprehensive analysis of transcriptional regulatory networks. This has enabled us to successfully identify the direct target genes of a given transcription factor (TF) [10,11]. Furthermore, based on the information obtained using the analysis system regarding the binding sites of an A. oryzae TF, AoXlnR, we have shown that the type and number of binding sites correlates with the level of differential expression and that the AoXlnR-dependent differential expression of downstream genes can be predicted based on the parameters of the DNA structure in the vicinity of the binding sites [11,12].
In this study, we aimed to elucidate the mechanism of KojR-mediated KA synthesis in A. oryzae through genome-wide investigation of KojR binding sites using gSELEX-Seq and identification of KojR-dependent differential expression genes (DEGs) by RNA-Seq with a DkojR strain. Our findings indicate that kojT, which encodes a putative transporter in A. oryzae, is directly regulated by KojR. We further examined the C-source metabolic pathway in A. oryzae and its relevance to KojR-dependent DEGs.
2. Materials and Methods
2.1. A. oryzae Strains, Growth Conditions, and Transformation
Wild-type A. oryzae strain RIB40 [13] was used in this study. Transformation of A. oryzae strains was performed as follows: The A. oryzae RIB40 strain was seeded and incubated overnight at 37 °C. The mycelia were collected with a nylon mesh filter (110 mesh), washed with 0.8 M NaCl, and incubated with 20 mL × 2 protoplastogenic solution [0.3% Yatalase (Takara, Shiga, Japan), 0.03% Lysing enzyme from Trichoderma harzianum (Sigma-Aldrich, St. Louis, MO, USA), 0.8 M NaCl, and 10 mM sodium phosphate buffer (pH 5.8)] with shaking at 30 °C for 2 h. The protoplastidized solution was consecutively filtered through 110 and 305 nylon mesh filters and centrifuged (1,500 rpm, 5 min, at RT) using a swinging rotor to remove the supernatant. The obtained pellet was washed with 0.8 M NaCl and centrifuged using a swinging rotor (1,500 rpm, 5 min, at RT) to remove the supernatant. The washing step was repeated twice. Next, 10 mL of Sol I (0.8 M NaCl and 10 mM CaCl2) was gently added, and the mixture was centrifuged using a swing rotor (1,500 rpm, 5 min, at RT). Most of the supernatant was removed but approximately 300–500 μL was retained, and the pellet was resuspended in the remaining supernatant. DNA of interest for transformation (approximately 10 µg) and 12.5 µL of Sol II (0.25 g/mL PEG6000, 50 mM CaCl2, and 10 mM Tris-HCl (pH 7.5)) were added to 50 µL of the protoplast suspension, which was gently suspended and placed on ice for 20 min. An additional 500 µL of Sol II was added, gently suspended, and allowed to stand on ice for 5 min. Then, 900 µL of Sol I was added and gently suspended. The suspensions were inoculated onto plates with regeneration media (70 mM NaNO3, 11 mM KH2PO4, 7 mM KCl, 22% (w/v) sorbitol, 2% (w/v) D-glucose, 0.15% (v/v) trace element solution, 2 mM MgSO4, and 0.01% pyrithiamine (pH 6.5)) and incubated at 37 °C for several days.
2.2. Construction of KojR FL/BD Expression Vector
The gene encoding the KojR constructs was sourced from gBlock Gene Fragments (Integrated DNA Technologies , Coralville, IA, USA). Using this construct as the template, fragments for kojR or kojR-BD were amplified using HO-kojR-F and HO-kojR-R, or using HO-kojR-F, and HO-kojR-BD-R (Supplementary Table S1). These fragments were then cloned through the SLiCE method [14] into a vector fragment amplified with PMAL-IFC-F1 and PMAL-IFC-R1 (Supplementary Table S1) using pMalc2AmyR1-411[15] as the template (pMalc2KojR or pMalc2KojR-BD).
A plasmid construct for cell-free expression of maltose binding protein (MBP)-fused full-length KojR was prepared using the following procedure. The full-length KojR gene fragment was amplified using kojR-F and kojRFull-R (Supplementary Table S1) with pMalc2KojR as the template. The amplicon was cloned using the NEBuilder HiFi DNA Assembly Master Mix (New England BioLabs, Ipswich, MA, USA) into a vector fragment amplified with gibson_Full-F and gibson_kojR-R (Supplementary Table S1) using pRSETb-MBP-DrDsxF (Kojima, T., unpublished) as the template (pRSETb-MBP-KojR-FL).
A plasmid construct for cell-free expression of MBP-fused KojR1-70 (pRSETb-MBP-KojR-BD) was prepared using the procedure described above with the following exceptions: the N-terminal region of the KojR gene fragment was amplified using kojR-F and kojRBD-R (Supplementary Table S1) with pMalc2KojR-BD as the template. The vector fragment was amplified with gibson_BD-F and gibson_KojR-R primers (Supplementary Table S1) using pRSETb-MBP-DrDsxF as the template.
2.3. Expression of MBP-Fused KojR Using Cell-Free Protein Synthesis System
Template DNA for the cell-free protein expression of MBP-fused full-length KojR or MBP-fused KojR1-70 was prepared using PCR with F1-primer and R1-primer (Supplementary Table S1); pRSETb-MBP-KojR-FL or pRSETb-MBP-KojR-BD was used as the template, respectively. Cell-free protein synthesis was performed using the PUREfrex 2.1 kit (GeneFrontier, Chiba, Japan) according to the manufacturer’s instructions. Briefly, cysteine, GSH, Solution I, Solution II, and Solution III from the kit, template DNA for the synthesis of MBP-fused full-length KojR or MBP-fused KojR1-70, and ZnCl2 (final conc. 2 mM) were mixed to obtain a reaction mixture (10 µL). The mixture was incubated at 37 °C for 2 h and centrifuged at 15,000 rpm for 5 min at RT to separate the soluble and insoluble fractions. The soluble fraction was used for the binding reaction in gSELEX (described in the next section).
SDS-PAGE was performed to confirm the expressed proteins using a cell-free protein synthesis system, and 0.1 μL of fluorescently labeled lysine tRNA (FluoroTect GreenLys tRNA, Promega, Tokyo, Japan) was added to the cell-free protein synthesis reaction mixture to visualize the expressed protein. The insoluble fraction recovered after the reaction was suspended in 10 μL of 1×PBS and applied to the analytical gel. A scanner capable of detecting green fluorescence was used to visualize the expressed proteins. The Protein Molecular Weight Marker (Takara) was fluorescently labeled using the Fluorescein Labeling Kit-NH2 (Dojindo Laboratory, Kumamoto, Japan) and used as the molecular weight marker.
2.4. In Vitro Selection of KojR-Bound DNA Fragments from an A. oryzae Genomic Library Using gSELEX-Seq
The A. oryzae genomic library was prepared according to a previously reported procedures [11]. gSELEX was performed using the A. oryzae genomic library. First, 9 µL of cell-free expressed KojR FL or KojR BD solution and 1 µL of 5 ng/µL A. oryzae genomic library solution were mixed and gently by rotation for 30 min at RT. Next, 5 μL of amylose resin (New England BioLabs) was washed with 50 μL of MBP w/o EDTA buffer (200 mM NaCl, 20 mM Tris–HCl, and 10 mM 2-mercaptoethanol (pH 7.5)). The resin was suspended in a 1.5-mL tube in 90 μL of fresh MBP w/o EDTA buffer and mixed with the MBP-KojR FL/BD-binding reaction mixture. The suspension was mixed overnight using a rotator at 4 °C, following which the resin was recovered through centrifuging at 300 × g for 2 min at 4 °C. The resin was washed with 50 μL of MBP w/o EDTA buffer to remove the free DNA fragments. After removing as much of the supernatant as was possible, the resin was suspended in 10 μL of MBP w/o EDTA elution buffer (200 mM NaCl, 20 mM Tris–HCl, 10 mM 2-mercaptoethanol, and 20 mM maltose (pH 7.5)). The suspension was mixed using a rotator for 15 min at 4 °C, and the supernatant was recovered after centrifugation at 300 × g for 2 min at 4 °C. The selected DNA fragments were amplified using a PCR with Nextera-Read1 and Nextera-Read2 (Supplementary Table S1); the following program was used: preheating at 94 °C for 3 min; 17 cycles of 94 °C for 15 s, 68 °C for 10 s, and 72 °C for 20 s; and a final extension at 72 °C for 5 min. After purification, the DNA pool was used for the second round of selection, in which the selected DNA pool was prepared using the same procedure as that used in the first round with the following exception: 5 ng of the DNA pool (selected in the first round) and 1 mU of poly(deoxyinosinic-deoxycytidylic) acid sodium salt (polydIdC) (Sigma-Aldrich) were added to the binding reaction mixture containing the cell-free KojR. In the third round, 5 ng (KojR FL) or 7.5 ng (KojR BD) of the DNA pool selected in the second round was used with 1 mU of polydIdC in the binding reaction. The concentration of each purified DNA pool was assessed using the Quant-iT dsDNA Broad-Range Assay Kit (Thermo Fisher Scientific, Waltham, MA, USA) according to the manufacturer’s instructions.
2.5. DNA Sequencing and Data Analysis in gSELEX-Seq
Sequencing libraries using each DNA pool obtained using gSELEX were prepared with the NEBNext Ultra II DNA Library Prep Kit for Illumina and NEBNext Multiplex Oligos for Illumina (New England BioLabs). The libraries were applied to Illumina NextSeq550 to obtain sequence data in 81 b read length in paired-end mode. It should be noted that the DNA pool from the unselected A. oryzae genomic library was also sequenced for use as a control tag in detecting the peaks of the KojR-binding site. All sequencing data will be made available through the DNA Databank of Japan (DDBJ; accession number: DRA016969).
Genome-wide analysis of KojR binding sites using sequencing data obtained using NextSeq 550 was performed according to the following procedure: 1) The 5' adapter was stripped from the reads using Cutadapt (ver. 4.3) [16] with the following parameters: -g AGATGTGTATAAGAGACAG -g CTGTCTCTTATACACATCT, followed by removal of the unpaired reads using Trimmomatic (ver. 0.39) [17]. 2) The trimmed paired-end reads were mapped using Bowtie2 (ver. 2.2.3) [18] into an A. oryzae genome file (aor0-5.fna) obtained from the Comprehensive Aspergillus oryzae Genome Database (CAoGD) (<https://nribf21.nrib.go.jp/CAoGD/>) using default settings. The generated SAM files were converted into BAM files using SAMtools (ver. 0.1.18) [19]. 3) HOMER (v4.11.1) findPeaks (<http://homer.ucsd.edu/homer/ngs/peaks.html>) was used to identify KojR FL or KojR BD binding stites with the following parameters: -i control tag directory -LP 0.01 -P 0.01, followed by preparation of a bed file based on the position values of detected peaks. Additionally, motif analysis of KojR FL- or KojR BD-bound sequences was performed using Homer findMotifsGenome.pl (<http://homer.ucsd.edu/homer/ngs/peakMotifs.html >) based on the acquired peak regions. 4) The obtained bed files were converted into FASTA files using the aor0-5. fna and BEDTools fastaFromBed (ver. v2.17.0) [20]. 5) The promoters possibly regulated by KojR FL or KojR BD were annotated as follows: the sequences of the peak regions obtained from the third selection round were annotated using A. oryzae upstream 1000 dataset [11] and BLAST+ (ver. 2.13.0+) (<https://blast.ncbi.nlm.nih.gov/doc/blast-help/downloadblastdata.html>) with the following parameters: blastn -evalue 0.1 -outfmt 6.
2.6. Construction of an A. oryzae ΔkojR Strain Using CRISPR/Cas9
A mutation was introduced into the KojR-binding domain-coding region of the A. oryzae RIB40 genome using the CRISPR/Cas9 system reported in a previous study [21]. Linear DNA with the wA target site of ppAsAC9gwA [21] replaced with a 21 bp kojR target site was prepared using PCR with ppAsAC9gwA as the template along with ppAsAC9gwA_F7 and ppAsAC9gwA_R7 (Supplementary Table S1). The linear fragment was re-circled using the SLiCE method [14] and introduced into Escherichia coli DH5a for selection to obtain the plasmid ppAsAC9g_kojR_BD for the preparation of ΔkojR strain. ppAsAC9g_kojR_BD was transformed into A. oryzae RIB40 strain to introduce mutations into the kojR target site in the genome. The mycelia of the transformants on the regeneration medium plate were collected, suspended in 50 µL of S.W., and incubated at 95 °C for 15 min. The suspension was used as the template for the amplification of the target mutation site in kojR using PCR using the kojR_CP_F and kojR_CP_R primers (Supplementary Table S1). The amplified products were subjected to DNA sequencing analysis using kojR_seq primer (Supplementary Table S1) for the investigation of mutation and gene deletion/integration. The loss of KojR function of the ΔkojR strain was confirmed using the non-pigmentation of the culture medium after 3 days of incubation in the KA medium containing FeCl3.
2.7. Identifications of DEGs Associated with KojR Expression Using RNA-Seq
Spores of A. oryzae RIB40 or ΔkojR strain were inoculated (5–6 × 106) into 50 mL of KA production medium (10% (w/v) D-glucose, 0.25% (w/v) yeast extract, 5 mM K2HPO4, and 4 mM MgSO4 (pH 6.0)) [6] with 1 mM FeCl3 and incubated with shaking for 3 days at 37 °C. The cultured mycelia were collected, frozen at -80 °C, divided into approximately 0.1 g samples that were added to each 2-mL screw cap tube, and disrupted using Multi-beads shocker (Yasui Kikai, Osaka, Japan) (2,500 rpm, 10 s; off time 3 s × 3 cycles). Total RNA was extracted from each disrupted mycelium using a PureLink RNA Mini Kit (Thermo Fisher Scientific) according to the manufacturer’s instructions.
cDNA fragments of six samples (3 × RIB40 and 3 × ΔkojR) were subjected to sequencing analysis using the MinION nanopore sequencer (Oxford Nanopore Technologies, Oxford, UK). Each cDNA library was prepared using the cDNA-PCR Sequencing Kit (SQK-PCS109) and PCR Barcoding Kit (SQK-PBK004) (Oxford Nanopore Technologies) according to the manufacturer’s protocol. However, one of the RIB40-derived samples was not used in subsequent analyses because it did not yield a sufficient amount of data. All sequencing data will be made available through the DNA Databank of Japan (DDBJ; accession number: DRA016970).
The sequenced data obtained using MinION was subjected to differential expression analysis as follows: 1) Polythymine and polyadenine sequences at the end of the reads were sequentially removed using Cutadapt (ver. 4.3) [16] with the following parameters: -g TTTTTTTT -n 20 -O 10; -a AAAAAAAAAA -n 10 -O 10. 2) Mapping was performed using minimap2 (ver. 2.17-r954-dirty) [22] with the following parameters: -ax splice -m 20 using the A. oryzae genome file (aor0-5.fna). The generated SAM files were converted into BAM files using SAMtools (ver. 0.1.18) [19]. 3) Counts of reads mapped to the coding sequences (CDS) of genes in the A. oryzae genome were performed using FeatureCounts (ver. 1.5.2) [23] with the following parameters: --readExtension5 200 --readExtension3 200 using an A. oryzae GTF file (aor0-5.gtf) obtained using CAoGD. The --readExtension5 and --readExtension3 arguments were specified to account for the lower counts due to the degradation of RNA applied to sequencing. False discovery rate (FDR) was calculated using the χ2 test with the Benjamini–Hochberg method to detect DEGs (FDR < 0.05).
2.8. KEGG Pathway Analysis of DEGs
3. Results
3.1. Preparation of MBP-Fused KojR for Use in gSELEX
The predicted conformation of the MBP-KojR fusion protein confirmed that the location of the DNA binding domain (Zn(II)2Cys6 region, amino acids from 15th to 58th of KojR) in the MBP fusion full-length KojR or MBP fusion KojR1-70 (70 amino acids N-terminal side of KojR), expressed from the DNA construct used in this study, is spatially distant from the MBP region (Supplementary Figure S1). The target protein expressed by using a cell-free protein synthesis system with DNA constructs as templates were preferentially expressed per the designed size (Supplementary Figure S2).
3.2. Genome-Wide Identification of KojR Binding Sites Using gSELEX-Seq
The DNA pool derived from the third selection round of gSELEX using either MBP-fused full-length KojR or MBP-fused KojR1-70, was sequenced using the Illumina NextSeq 550 system, and genome-wide KojR-binding sites were identified using bioinformatics analysis. The region detected as the peak was subjected to sequence motif analysis, and 5´- CGGCTAATGCGG -3´ was detected as a KojR-binding motif in the result of gSELEX-Seq using full-length MBP-fused KojR (Figure 1A). 5´- (G/C)GG(T/A)ATAG(T/C)C(T/G)T -3´ was detected as the binding motif in gSELEX-Seq using MBP-fused KojR1-70, although this sequence was somewaht less conserved than in the case of full length KojR (Figure 1B).
Furthermore, when the regions detected as peaks were compared with the 1,000 bp regions upstream of all genes in A. oryzae, 32 promoter regions were detected that might potentially bind to the MBP-fused full-length KojR (Table 1). Additionally, it was found that 16 genes possess promoter regions where the MBP-fused KojR1-70 region is likely to bind (Supplementary Table S2). The promoter region of kojT (AO090113000138) was detected in both cases using MBP-fused full-length KojR and MBP-fused KojR1-70.
3.3. Identification of KojR Expression-Dependent DEGs Using RNA-Seq
To comprehensively identify genes dependent on KojR expression, total RNA sequences extracted from A. oryzae ΔkojR and A. oryzae RIB40 were analyzed using nanopore sequencing. After processing the acquired sequence data of the 11,902 A. oryzae genes using various bioinformatics tools, we identified 100 genes whose expression levels varied significantly between A. oryzae ΔkojR and A. oryzae RIB40 strains (Figure 2 and Supplementary Table S3). AO090113000136 (kojA) showed the most significant differential expression among all the analyzed genes with a marked increase in expression observed in the RIB40 strain than in the ΔkojR strain. AO090113000138 (kojT) also showed a significant increase in expression in the RIB40 strain compared with that of the ΔkojR strain. Notably, the number of genes with increased expression in the RIB40 strain was similar to the number of genes with decreased expression, compared to the ΔkojR strain (Figure 2). Certain genes, including AO090010000444 that encoded a predicted phosphofructokinase subunit protein, showed particularly marked increases in expression in ΔkojR strain than in the RIB40 strain (Supplementary Table S3).
3.4. Comprehensive Identification of Target Genes of KojR Using Integrated Analysis-Based Binding Sites on A. oryzae Genome and DEGs Information
The dataset of the genes with KojR binding sites in the promoter region obtained by performing gSELEX-Seq using the MBP-fused full length KojR or MBP-fused KojR1-70 and the dataset of KojR-dependent DEGs obtained by performing RNA-Seq in the ΔkojR strain were compared in an integrated manner (Figure 3). The intersections between the datasets obtained using gSELEX-Seq from MBP-fused KojR and MBP-fused KojR1-70 included AO090113000138 (kojT), AO090038000029, and AO090701000448. Furthermore, comparing the datasets obtaind from gSELEX-Seq to the KojR-dependent DEG dataset showed that only AO090113000138 (kojT) was detected in the intersection (Figure 3).
4. Discussion
KojR is a transcriptional regulator that Marui et al. reported in 2011 to be strongly related to KA synthesis in A. oryzae [6]. Although KojR has been predicted to bind to the CGG triplet, which is a typical target DNA sequence of this type because it possesses a Zn(II)2Cys6 zinc finger domain, the identification of the genome-wide binding site and target genes has not yet been reported. In the present study, we attempted to elucidate the regulatory mechanism underlying KA synthesis through a detailed functional analysis of KojR.
We comprehensively identified the KojR binding sites at the genomic level using gSELEX-Seq, which does not involve an intracellular TF–DNA binding reaction, such as chromatin immunoprecipitation, and rapidly and easily identifies the binding sites of the TF of interest. Previous approaches using gSELEX-Seq have used only the N-terminal region (including the DNA-binding domain) of the target TF that was prepared using an E. coli recombinant protein expression system as the MBP fusion protein [10,11]. Although this strategy, which uses only the N-terminal region of the target TF, allows for efficient preparation of active MBP-fused TFs, the lack of a C-terminal region risks impairing the native intracellular functions such as stability and dimerization. In fact, gSELEX-Seq with AoXlnR1-183, the N-terminal region of AoXlnR, provided a DNA motif, which may be due to binding to the MBP-fused AoXlnR1-183 monomer [11].
To circumvent the problems associated with the restricted expression of full-length TFs in E. coli cells, a cell-free protein synthesis system was used in this study to express the target TF. To evaluate this strategy, a cell-free protein synthesis system was applied using MBP-fused full length KojR or MBP-fused KojR1-70 expression DNA constructs as templates. Subsequently, pronounced MBP-fused KojR expression was detected in the soluble fraction (Supplementary Figure S2). This result shows that gSELEX-Seq can be made more rapid and simpler by using a cell-free protein synthesis system as it circumvents the step where E. coli is cell cultured for expressing the recombinant protein, which is time-consuming and often limits the expression of active proteins from eukaryotes. Notably, MBP-fused full-length KojR was successfully expressed in the soluble fraction (Supplementary Figure S2). This result strongly reinforces the effectiveness of the strategy of using cell-free protein synthesis systems for exogenous protein expression, as this may overcome the problems associated with the lack of a C-terminal region described earlier. However, the expression level of MBP-fused full-length KojR was considerably lower than that of MBP-fused KojR1-70 (Supplementary Figure S2). The difference in the quantity of the synthesized products may be attributed to the preferential degradation of MBP-fused full-length KojR, as evidenced by the presence of several minor bands of smaller molecular weight in the soluble fraction after the expression of MBP-fused full-length KojR (Supplementary Figure S2).
Three rounds of gSELEX were performed with MBP-fused proteins prepared using a cell-free protein synthesis system, and the obtained DNA pool was applied to an Illumina NextSeq 550 to map the obtained read data to the A. oryzae genome. Analysis of the consensus sequence of the peak regions detected through this mapping clearly detected the CGG triplet in both MBP-fused full-length KojR and MBP-fused KojR1-70 (Figure 1). The results showed that the target sequences of the Zn(II)2Cys6-type TF KojR were preferentially detected, demonstrating the effectiveness of using gSELEX-Seq with KojR expressed using a cell-free protein synthesis system. Additionally, two CGG triplets were detected only for MBP-fused full-length KojR (Figure 1), suggesting that MBP-fused full-length KojR formed a dimer during binding DNA selection process of gSELEX.
Next, we defined the promoter region as the segment 1,000 bp upstream of the start codon of the A. oryzae gene and generated lists of genes that contained one or more peaks detected in response to KojR binding. The list contained 32 genes for the MBP-fused full-length KojR (Table 1) and 16 genes for the MBP-fused KojR1-70 (Supplementary Table S2). In both lists, kojT, a gene in the KA synthesis-related gene cluster, was identified as the top gene in terms of peak detection significance. These results strongly suggest the presence of a KojR-binding site in the promoter region of kojT. In fact, four CGGN6CGG sequences were densely distributed within the peak region of the kojT promoter (data not shown). In contrast, no significant peak was detected in the kojA promoter region for KojR binding either with MBP-fused full-length KojR or MBP-fused KojR1-70 (Table 1 and Supplementary Table S2). The possibility that in vitro binding reaction conditions in gSELEX, such as KojR concentration, affect the binding of KojR to the kojA promoter region should not be completely excluded. However, at the very least, the results indicate that KojR binds preferentially to the kojT promoter region rather than to the kojA promoter region.
Previous studies have suggested the possibility that KojR binds to the promoter of kojA and induces the activation of kojA expression [6,26]; however, the gSELEX-Seq results in this study, in which the kojA promoter region was not selected as the KojR-binding region, make this hypothesis difficult to support. Chang et al. proposed the KojR-binding motif as a highly common sequence in the promoter region of kojA and kojT, CGRCTWAGYCG—which is relatively consistent with those of both full-length KojR and KojR1-70 shown in Figure 1. In fact, there is one site in the A. oryzae kojA promoter region that is exactly the same as the CGACTTTGCCG targeted by Chang et al. and two sites in the A. oryzae kojT promoter region that exactly match the proposed consensus sequence CGRCTWAGYCG. In addition, Chang et al. showed that KA synthesis is suppressed by introducing mutations in the sequences around the target binding motif in the kojA promoter region of A. flavus [8]. Although these results suggest that KojR binds to the kojA promoter and induces its expression in A. flavus, the possibility that the binding motif is the target site of the Zn(II)2Cys6 transcription factor involved in KA synthesis other than KojR cannot be ruled out. Definitive conclusions regarding the presence or absence of KojR-binding sites in the kojA promoter may be obtained by preparing kojA promoters with various mutations in the KojR-binding site and quantitatively evaluating the binding affinity for KojR using surface plasmon resonance (SPR) or biolayer interferometry (BLI).
Notably, only three promoter regions (kojT, AO090038000029, and AO090701000448) were included in the intersection set when comparing the gene promoter sets detected using MBP-fused full-length KojR and MBP-fused KojR1-70 (Figure 3, Table 1, and Supplementary Table S2). The differences in the selected promoter regions may be attributed to the concentration of KojR in the in vitro binding reaction and to the effects of KojR stability and dimerization described earlier. Since conditions as close as possible to those of intact cells are more desirable, it would be appropriate to give priority to the promoter list obtained when using full-length KojR fused with MBP.
In parallel with the gSELEX-Seq analysis, we performed a comprehensive identification of KojR expression-dependent DEGs using RNA-Seq with the ΔkojR strain. kojA and kojT in the KA synthesis-related gene cluster were detected as DEGs, and their expression was reduced by the deletion of kojR (Figure 2 and Supplementary Table S3). This result is fully consistent with the previous report that expression of kojA and kojT is suppressed in the ΔkojR strain, as shown by qRT-PCR analysis [6]. The results of this study obtained using RNA-Seq reinforce the association between KojR and the expression of kojA and kojT.
Terabayashi et al. reported genes whose expression was induced in response to the increased synthesis of KA [5]. A comparison of the gene list with the DEGs obtained in this study revealed duplications of AO090120000112, AO090011000414, AO090001000237, and AO090003000018 in addition to kojA and kojT. Notably AO090001000237 is an ortholog of VeA, which is associated with secondary metabolism in A. nidulans (Supplementary Table S3). AO090011000414 was expected to exhibit glyceraldehyde-3-phosphate dehydrogenase activity during glycolysis (Supplementary Table S3). This finding reinforces the involvement of KojR in the metabolism of carbon sources in A. oryzae. Notably, Terabayashi et al. identified AO090011000414 as a gene whose expression is induced by increased KA synthesis, whereas we identified AO090011000414 as a DEG whose expression is higher than that in RIB40 in the ΔkojR strain in which KA synthesis is suppressed. This vital difference suggests that the expression of genes involved in glycolysis is regulated intricately during secondary metabolite synthesis. Interestingly, the DEGs in this study included a GATA-type transcription factor, namely, AO090012000768 (nsdD) Supplementary Table S3), which may be essential for KA synthesis in previous reports [27].
In contrast, AO090012000623 (nrtA) [28], AO090003000489 (laeA) [29], AO090009000298 (AoKap1) [30], AO090120000102 (AoKap2) [31], AO090113000139 (Aokap4) [32], AO090009000515 (Aokap5) [33], AO090113000133 (Aokap6) [34], AO090005000940 (AoZip2) [35], AO090012000498 (AozfA) [36], AO090003001186 (KpeA) [27], AO090020000046 (lreA) [27] and AO090009000320 (creB) [27], which may be related to KA synthesis according to previous reports, were not included in the DEGs in this study. Particularly, Chen et al. reported decreased expression of Aokap6 owing to kojR deletion; however, the degree of change was relatively low [34]. Factors that contribute to the lack of observed expressional variations associated with the deletion of KojR in the genes described above include differences in sensitivity accompanied by analysis conditions, timing of RNA extraction and incubation time, and the possibility that KojR plays a role further downstream in the KA synthesis pathway.
Finally, metabolic pathway mapping using the KEGG pathway [24] was performed to evaluate the role of the 100 DEGs, which were detected by RNA-Seq analysis, in the carbon-source metabolic pathway in A. oryzae cells. In total, 5 DEGs (AO090010000444, AO090011000414, AO090038000395, AO090003000415, and AO090009000557) were mapped onto the carbon metabolism and glycolysis pathways in A. oryzae (Figure 4). AO090010000444, AO090011000414, and AO090038000395 are involved in the glycolytic system and were upregulated in the ΔkojR strain. In contrast, AO090003000415 and AO090009000557 were involved in the citrate cycle and were downregulated in the ΔkojR strain. The mapping results suggest that KojR may be involved in the inhibition of glycolysis but contribute to TCA cycle activation (Figure 4). However, since no KojR binding sites were detected in the promoter regions of the genes analyzed using gSELEX-Seq (Table 1 and Supplementary Table S2), it is possible that these DEGs are indirectly regulated by KojR. Taken together, these results suggested that KojR is indirectly but closely related to glucose-initiated KA synthesis.
5. Conclusions
In this study, we performed a comprehensive search for KojR target genes and showed that kojT is directly regulated by KojR. We also identified a group of genes associated with KojR expression and showed that KojR expression is closely related to the expression of a group of enzymes involved in C-source metabolism. We believe that the findings of this study will contribute greatly to the elucidation of the KA biosynthesis mechanism in A. oryzae using C sources such as glucose.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Supplementary Figure S1: Predicted 3D structure of MBP-fused KojR used in gSELEX; Supplementary Figure S2: SDS-PAGE analysis of MBP fusion proteins containing full-length KojR or KojR DNA binding domain synthesized by cell-free protein synthesis system; Supplementary Table S1: Oligonucleotides used in this study; Supplementary Table S2: Genes containing KojR1-70 binding region in 1,000 bp upstream of the initiation codon; Supplementary Table S3: DEGs detected using RNA-Seq in A. oryzae ΔkojR and RIB40 strains.
Author Contributions
Conceptualization, T.M., H.O., T.K. and H.N.; methodology, T.M., H.O. and T.K.; validation, J.-i.M., M.S., M.K., H.N. and T.K.; formal analysis, T.M. and T.K.; investigation, T.M., H.O., R.G., R.T. and T.K.; data curation, T.K.; writing—original draft preparation, T.M. and T.K.; writing—review and editing, H.O., J.-i.M., M.S., M.K., H.N. and T.K.; visualization, T.M. and T.K.; supervision, T.K.; project administration, T.K. and H.N.; funding acquisition, T.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported in part by the JSPS KAKENHI Grant-in-Aid for Scientific Research (C) (Grant Number JP19K05766). This work was financially supported in part by the Sapporo Bioscience Foundation.
Data Availability Statement
The reported nucleotide sequence data are available in the DDBJ Sequenced Read Archive under the accession numbers DRA016969 and DRA016970.
Acknowledgments
The authors would like to thank Mika Nomoto, Yasuomi Tada, Akiko Akama, and Mikako Yamaguchi (Center for Gene Research, Nagoya University) for their technical support with next-generation sequencing (NextSeq550). We would also like to thank Yoshiaki Inukai (International Center for Research and Education in Agriculture, Nagoya University) for the use of Multi-beads shocker. We would like to thank Editage (www.editage.jp) for English language editing.
Conflicts of Interest
The funders had no role in the study design; collection, analyses, or interpretation of data; writing of the manuscript; or decision to publish the results.
References
- Saeedi:, M.; Eslamifar, M.; Khezri, K. Kojic Acid Applications in Cosmetic and Pharmaceutical Preparations. Biomedicine & Pharmacotherapy 2019, 110, 582–593. [Google Scholar] [CrossRef]
- Brtko, J. Biological Functions of Kojic Acid and Its Derivatives in Medicine, Cosmetics, and Food Industry: Insights into Health Aspects. Archiv der Pharmazie 2022, 355, 2200215. [Google Scholar] [CrossRef]
- Bentley, R. From Miso, Sake and Shoyu to Cosmetics: A Century of Science for Kojic Acid. ChemInform 2007, 38. [Google Scholar] [CrossRef]
- Chib, S.; Jamwal, V.L.; Kumar, V.; Gandhi, S.G.; Saran, S. Fungal Production of Kojic Acid and Its Industrial Applications. Appl Microbiol Biotechnol 2023, 107, 2111–2130. [Google Scholar] [CrossRef]
- Terabayashi, Y.; Sano, M.; Yamane, N.; Marui, J.; Tamano, K.; Sagara, J.; Dohmoto, M.; Oda, K.; Ohshima, E.; Tachibana, K.; et al. Identification and Characterization of Genes Responsible for Biosynthesis of Kojic Acid, an Industrially Important Compound from Aspergillus oryzae. Fungal Genetics and Biology 2010, 47, 953–961. [Google Scholar] [CrossRef]
- Marui, J.; Yamane, N.; Ohashi-Kunihiro, S.; Ando, T.; Terabayashi, Y.; Sano, M.; Ohashi, S.; Ohshima, E.; Tachibana, K.; Higa, Y.; et al. Kojic Acid Biosynthesis in Aspergillus oryzae Is Regulated by a Zn(II)2Cys6 Transcriptional Activator and Induced by Kojic Acid at the Transcriptional Level. Journal of Bioscience and Bioengineering 2011, 112, 40–43. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, H.; Chen, Z.; Fan, J.; Chen, T.; Zeng, B.; Zhang, Z. Construction of Single, Double, or Triple Mutants within Kojic Acid Synthesis Genes kojA, kojR, and kojT by the CRISPR/Cas9 Tool in Aspergillus oryzae. Folia Microbiol 2022, 67, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.-K.; Scharfenstein, L.L.; Mahoney, N.; Kong, Q. Kojic Acid Gene Clusters and the Transcriptional Activation Mechanism of Aspergillus flavus KojR on Expression of Clustered Genes. JoF 2023, 9, 259. [Google Scholar] [CrossRef]
- De Castro, P.A.; Colabardini, A.C.; Moraes, M.; Horta, M.A.C.; Knowles, S.L.; Raja, H.A.; Oberlies, N.H.; Koyama, Y.; Ogawa, M.; Gomi, K.; et al. Regulation of Gliotoxin Biosynthesis and Protection in Aspergillus Species. PLoS Genet 2022, 18, e1009965. [Google Scholar] [CrossRef] [PubMed]
- Kojima, T.; Kunitake, E.; Ihara, K.; Kobayashi, T.; Nakano, H. A Robust Analytical Pipeline for Genome-Wide Identification of the Genes Regulated by a Transcription Factor: Combinatorial Analysis Performed Using gSELEX-Seq and RNA-Seq. PLOS ONE 2016, 11, e0159011. [Google Scholar] [CrossRef] [PubMed]
- Oka, H.; Kojima, T.; Ihara, K.; Kobayashi, T.; Nakano, H. Comprehensive Investigation of the Gene Expression System Regulated by an Aspergillus oryzae Transcription Factor XlnR Using Integrated Mining of gSELEX-Seq and Microarray Data. BMC Genomics 2019, 20, 16. [Google Scholar] [CrossRef]
- Oka, H.; Kojima, T.; Kato, R.; Ihara, K.; Nakano, H. Construction of Transcript Regulation Mechanism Prediction Models Based on Binding Motif Environment of Transcription Factor AoXlnR in Aspergillus oryzae. bioRxiv 2021. [Google Scholar] [CrossRef]
- Machida, M.; Asai, K.; Sano, M.; Tanaka, T.; Kumagai, T.; Terai, G.; Kusumoto, K.-I.; Arima, T.; Akita, O.; Kashiwagi, Y.; et al. Genome Sequencing and Analysis of Aspergillus oryzae. Nature 2005, 438, 1157–1161. [Google Scholar] [CrossRef]
- Okegawa, Y.; Motohashi, K. A Simple and Ultra-Low Cost Homemade Seamless Ligation Cloning Extract (SLiCE) as an Alternative to a Commercially Available Seamless DNA Cloning Kit. Biochemistry and Biophysics Reports 2015, 4, 148–151. [Google Scholar] [CrossRef] [PubMed]
- Tani, S.; Itoh, T.; Kato, M.; Kobayashi, T.; Tsukagoshi, N. In vivo and in vitro analyses of the AmyR binding site of the Aspergillus nidulans agdA promoter; requirement of the CGG direct repeat for induction and high affinity binding of AmyR. Bioscience, Biotechnology, and Biochemistry 2001, 65, 1568–74. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet.journal 17, 10–12. [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nature Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup The Sequence Alignment/Map Format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Quinlan, A.R.; Hall, I.M. BEDTools: A Flexible Suite of Utilities for Comparing Genomic Features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef] [PubMed]
- Katayama, T.; Nakamura, H.; Zhang, Y.; Pascal, A.; Fujii, W.; Maruyama, J. Forced Recycling of an AMA1-Based Genome-Editing Plasmid Allows for Efficient Multiple Gene Deletion/Integration in the Industrial Filamentous Fungus Aspergillus oryzae. Appl Environ Microbiol 2019, 85, e01896–18. [Google Scholar] [CrossRef]
- Li, H. Minimap2: Pairwise Alignment for Nucleotide Sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Sato, Y. KEGG Mapper for Inferring Cellular Functions from Protein Sequences. Protein Science 2020, 29, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Kutmon, M.; Van Iersel, M.P.; Bohler, A.; Kelder, T.; Nunes, N.; Pico, A.R.; Evelo, C.T. PathVisio 3: An Extendable Pathway Analysis Toolbox. PLoS Comput Biol 2015, 11, e1004085. [Google Scholar] [CrossRef]
- Tamano, K.; Kuninaga, M.; Kojima, N.; Umemura, M.; Machida, M.; Koike, H. Use of the kojA Promoter, Involved in Kojic Acid Biosynthesis, for Polyketide Production in Aspergillus oryzae: Implications for Long-Term Production. BMC Biotechnol 2019, 19, 70. [Google Scholar] [CrossRef]
- Arakawa, G.; Kudo, H.; Yanase, A.; Eguchi, Y.; Kodama, H.; Ogawa, M.; Koyama, Y.; Shindo, H.; Hosaka, M.; Tokuoka, M. A Unique Zn(II)2-Cys6-Type Protein, KpeA, Is Involved in Secondary Metabolism and Conidiation in Aspergillus oryzae. Fungal Genetics and Biology 2019, 127, 35–44. [Google Scholar] [CrossRef]
- Sano, M. Aspergillus oryzae nrtA Affects Kojic Acid Production. Bioscience, Biotechnology, and Biochemistry 2016, 80, 1776–1780. [Google Scholar] [CrossRef]
- Oda, K.; Kobayashi, A.; Ohashi, S.; Sano, M. Aspergillus oryzae laeA Regulates Kojic Acid Synthesis Genes. Bioscience, Biotechnology, and Biochemistry 2011, 75, 1832–1834. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, H.; Chen, Z.; Fan, J.; Chen, T.; Zeng, B.; Zhang, Z. Identification and Characterization of a Novel Gene Aokap1 Involved in Growth and Kojic Acid Synthesis in Aspergillus oryzae. Arch Microbiol 2022, 204, 67. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, H.; Chen, Z.; Fan, J.; Chen, T.; Xiao, Y.; Jie, J.; Zeng, B.; Zhang, Z. Overexpression of a Novel Gene Aokap2 Affects the Growth and Kojic Acid Production in Aspergillus oryzae. Mol Biol Rep 2022, 49, 2745–2754. [Google Scholar] [CrossRef]
- Chen, T.; Chen, Z.; Li, Y.; Zeng, B.; Zhang, Z. A Novel Major Facilitator Superfamily Transporter Gene Aokap4 near the Kojic Acid Gene Cluster Is Involved in Growth and Kojic Acid Production in Aspergillus oryzae. JoF 2022, 8, 885. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, Z.; Zhang, F.; Chen, T.; Fan, J.; Deng, X.; Lei, X.; Zeng, B.; Zhang, Z. The C2H2-Type Zinc-Finger Regulator AoKap5 Is Required for the Growth and Kojic Acid Synthesis in Aspergillus oryzae. Fungal Genetics and Biology 2023, 167, 103813. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Chen, T.; Wang, H.; Jiang, C.; Liu, Y.; Wu, X.; Li, Y.; Zeng, B.; Zhang, Z. Disruption of Aokap6 near the Kojic Acid Gene Cluster Affects the Growth and Kojic Acid Production in Aspergillus oryzae. World J Microbiol Biotechnol 2022, 38, 175. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Fan, J.; Long, C.; He, B.; Hu, Z.; Jiang, C.; Li, Y.; Ma, L.; Wen, J.; Zou, X.; et al. Identification and Characterization of the ZRT, IRT-like Protein (ZIP) Family Genes Reveal Their Involvement in Growth and Kojic Acid Production in Aspergillus oryzae. Journal of Industrial Microbiology and Biotechnology 2019, 46, 1769–1780. [Google Scholar] [CrossRef]
- Chen, Z.; Chen, T.; Zhang, H.; Li, Y.; Fan, J.; Yao, L.; Zeng, B.; Zhang, Z. Functional Role of a Novel Zinc Finger Protein, AoZFA, in Growth and Kojic Acid Synthesis in Aspergillus oryzae. Appl Environ Microbiol 2023, e00909–23. [Google Scholar] [CrossRef]
Figure 1.
KojR binding motif detected using gSELEX-Seq. (A) Full-length KojR-binding motif (B) The binding motif of KojR1-70. de novo motif analysis of the KojR-binding site was performed using Homer (v4.11.1). The highest-ranked motif was indicated based on the criteria based on p-values in full-length KojR or KojR1-70.
Figure 1.
KojR binding motif detected using gSELEX-Seq. (A) Full-length KojR-binding motif (B) The binding motif of KojR1-70. de novo motif analysis of the KojR-binding site was performed using Homer (v4.11.1). The highest-ranked motif was indicated based on the criteria based on p-values in full-length KojR or KojR1-70.

Figure 2.
MA-plot based on RNA-Seq read data using A. oryzae ΔkojR strain and A. oryzae RIB40. X-axis, average value of log2(average number of reads for each gene + 1) in A. oryzae RIB40 (WT) and A. oryzae ΔkojR (MT) strains; Y-axis, difference in average value of log2(average number of reads for each gene + 1) in WT and MT. FDR values were calculated using multiple testing using χ2 values and correction using the Benjamini-Hochberg method.
Figure 2.
MA-plot based on RNA-Seq read data using A. oryzae ΔkojR strain and A. oryzae RIB40. X-axis, average value of log2(average number of reads for each gene + 1) in A. oryzae RIB40 (WT) and A. oryzae ΔkojR (MT) strains; Y-axis, difference in average value of log2(average number of reads for each gene + 1) in WT and MT. FDR values were calculated using multiple testing using χ2 values and correction using the Benjamini-Hochberg method.
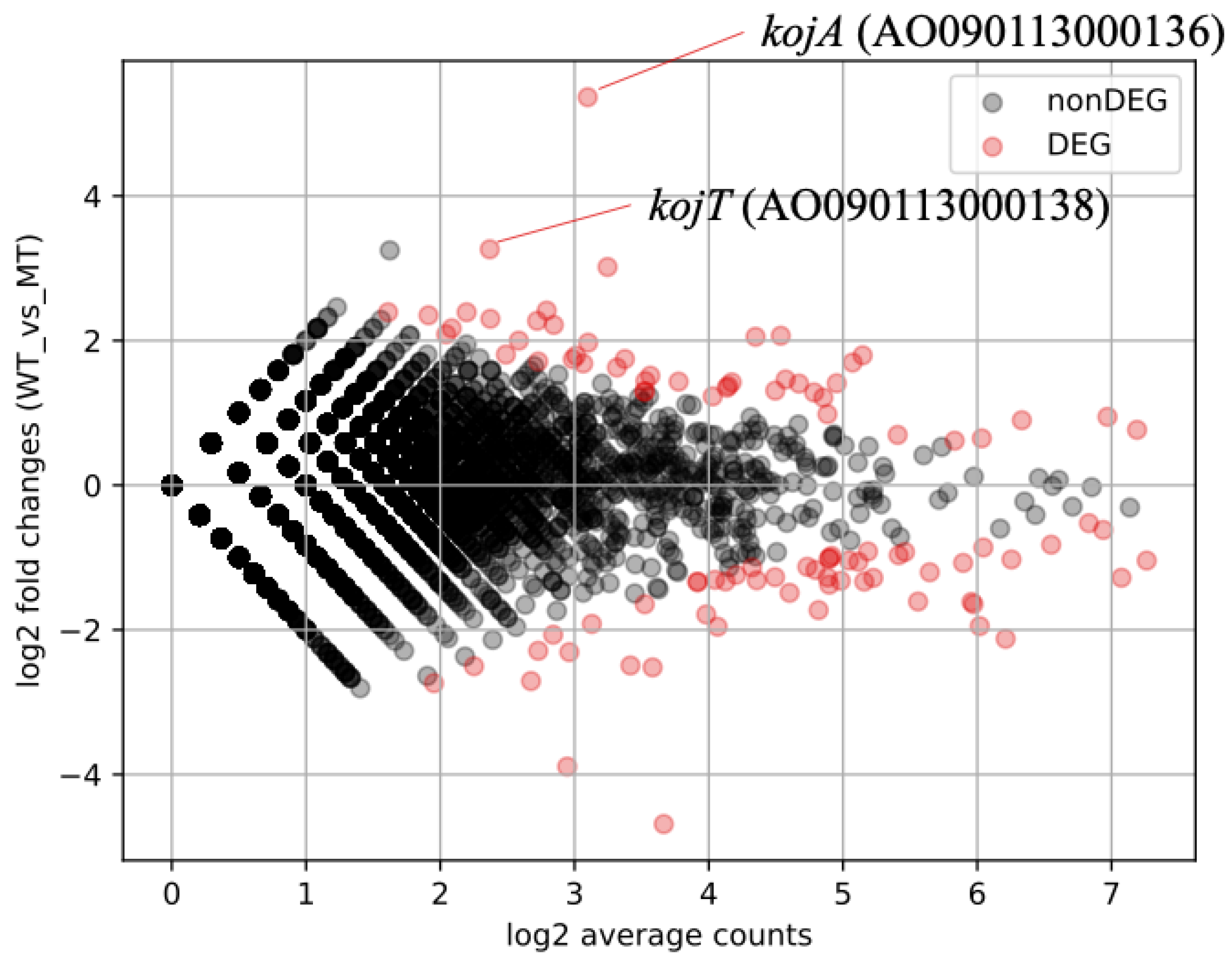
Figure 3.
Venn diagram of the number of KojR-related genes obtained using gSELEX-Seq and RNA-Seq. FL, genes controlled by the candidate KojR-regulated promoters obtained using gSELEX from MBP-fused full-length KojR; BD, genes controlled by the candidate KojR-regulated promoters obtained using gSELEX from MBP-fused KojR1-70; DEGs, DEGs obtained using RNA-seq from A. oryzae ΔkojR strain and A. oryzae RIB40.
Figure 3.
Venn diagram of the number of KojR-related genes obtained using gSELEX-Seq and RNA-Seq. FL, genes controlled by the candidate KojR-regulated promoters obtained using gSELEX from MBP-fused full-length KojR; BD, genes controlled by the candidate KojR-regulated promoters obtained using gSELEX from MBP-fused KojR1-70; DEGs, DEGs obtained using RNA-seq from A. oryzae ΔkojR strain and A. oryzae RIB40.
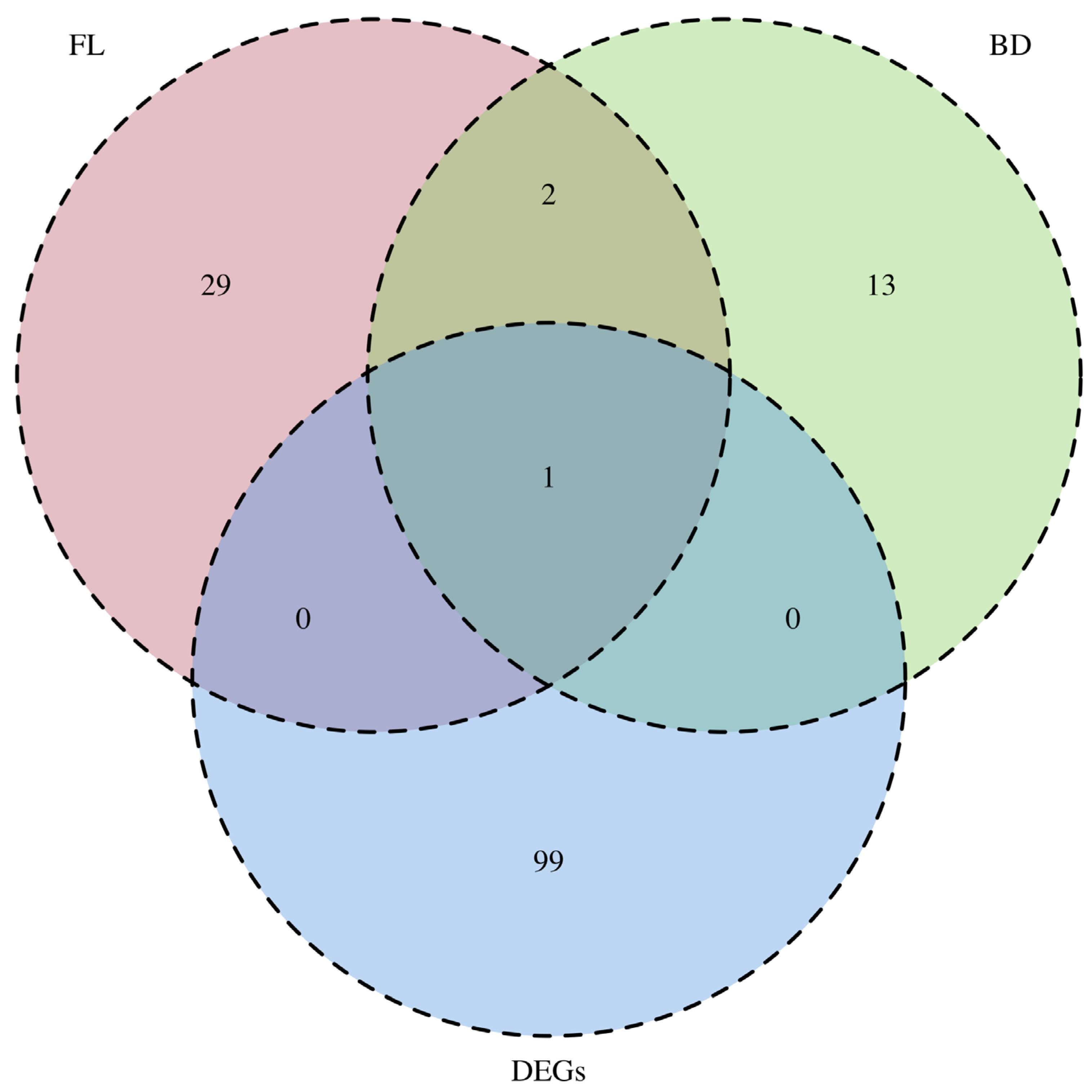
Figure 4.
Mapping of differentially expressed genes (DEGs) associated with kojR expression to A. oryzae carbon metabolic pathway. Red, DEG whose expression levels in A. oryzae RIB40 are higher than that in ΔkojR strain; Blue, DEG with lower expression levels in A. oryzae RIB40 than that in ΔkojR strain. A metabolic map was created using the PathVisio software (Ver. 3.3.0) [25] based on the carbon metabolism (aor01200) and glycolysis/gluconeogenesis (aor00010) of A. oryzae in KEGG [24].
Figure 4.
Mapping of differentially expressed genes (DEGs) associated with kojR expression to A. oryzae carbon metabolic pathway. Red, DEG whose expression levels in A. oryzae RIB40 are higher than that in ΔkojR strain; Blue, DEG with lower expression levels in A. oryzae RIB40 than that in ΔkojR strain. A metabolic map was created using the PathVisio software (Ver. 3.3.0) [25] based on the carbon metabolism (aor01200) and glycolysis/gluconeogenesis (aor00010) of A. oryzae in KEGG [24].
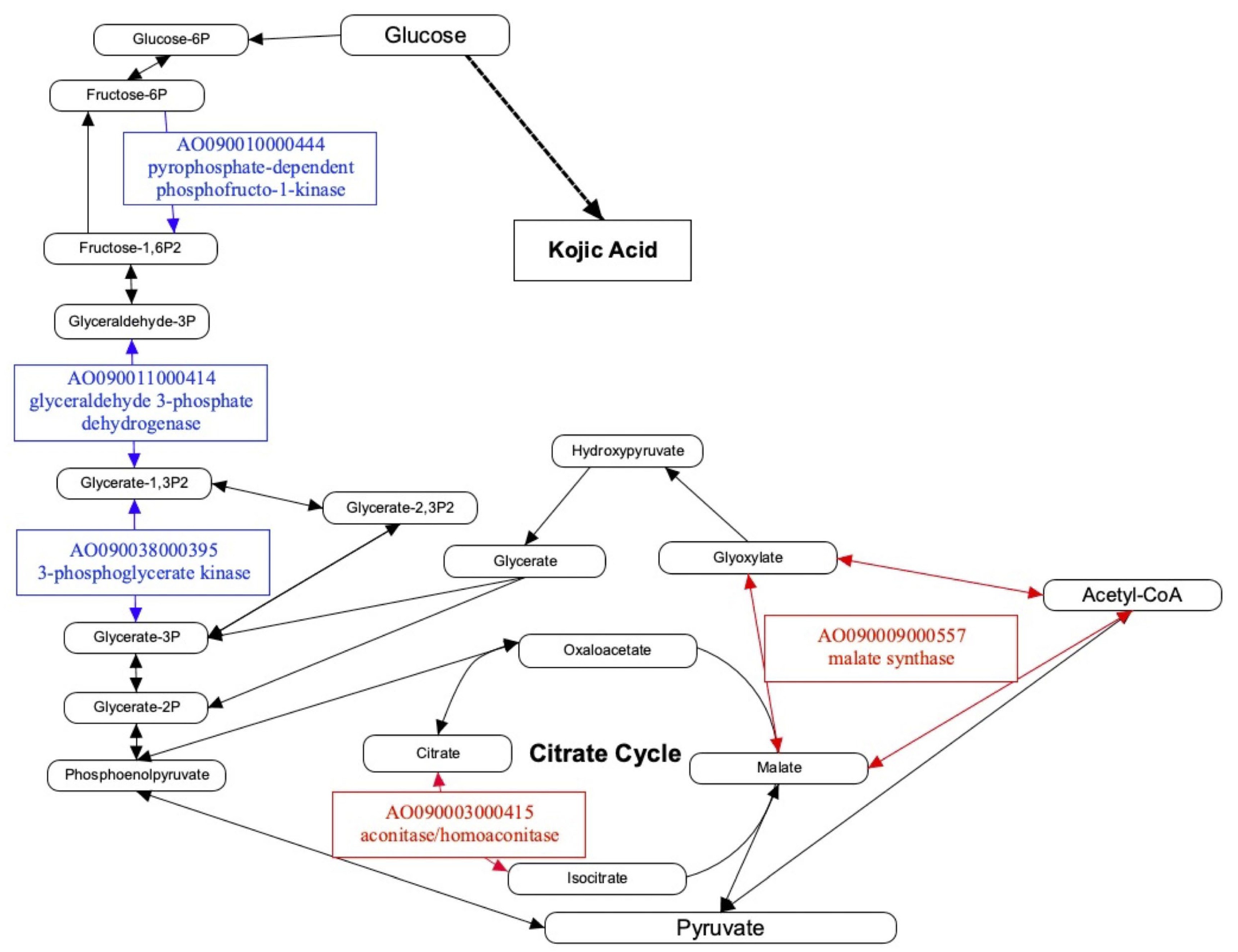

Table 1.
Genes containing full-length KojR binding region 1000 bp upstream of the initiation codon.
| Peak Region | Gene ID | Original Description |
|---|---|---|
| Chr5_A_oryzae_RIB40:4372480-4372692 | AO090113000138 | Putative transporter; present in the kojic acid biosynthetic gene cluster |
| Chr6_A_oryzae_RIB40:3584694-3584906 | AO090038000029 | Has domain(s) with predicted peroxiredoxin activity and role in oxidation-reduction process |
| Chr5_A_oryzae_RIB40:1165262-1165474 | AO090701000448 | Has domain(s) with predicted heme binding activity |
| Chr5_A_oryzae_RIB40:643370-643582 | AO090701000654 | Protein of unknown function |
| Chr5_A_oryzae_RIB40:1158260-1158472 | AO090701000450 | Has domain(s) with predicted catalytic activity, hydrolase activity, and role in metabolic process |
| Chr1_A_oryzae_RIB40:5569621-5569833 | AO090005000336 | Has domain(s) with predicted sequence-specific DNA binding RNA polymerase II transcription factor activity, zinc ion binding activity, and role in regulation of transcription, DNA-templated and nucleus localization |
| Chr7_A_oryzae_RIB40:229700-229912 | AO090011000078 | Ortholog of A. niger CBS 513.88 : An01g06430, A. versicolor : Aspve1_0082366, A. niger ATCC 1015 : 36036-mRNA, and A. zonatus : Aspzo1_0026572 |
| Chr1_A_oryzae_RIB40:4941497-4941709 | AO090005000571 | Has domain(s) with predicted NAD binding, oxidoreductase activity, acting on the CH-OH group of donors, NAD or NADP as acceptor activity, and role in oxidation-reduction process |
| Chr2_A_oryzae_RIB40:4505321-4505533 | AO090003000890 | Protein of unknown function |
| Chr4_A_oryzae_RIB40:4149411-4149623 | AO090102000151 | Ortholog(s) have ubiquitin–protein transferase activity and role in protein import into peroxisome matrix and integral component of peroxisomal membrane localization |
| Chr1_A_oryzae_RIB40:6476509-6476721 | AO090308000007 | Protein of unknown function |
| Chr3_A_oryzae_RIB40:391752-391964 | AO090023000155 | Protein of unknown function |
| Chr6_A_oryzae_RIB40:3081726-3081938 | AO090038000221 | Protein of unknown function |
| Chr7_A_oryzae_RIB40:1923126-1923338 | AO090011000751 | Protein of unknown function |
| Chr2_A_oryzae_RIB40:5473553-5473765 | AO090003001242 | Has domain(s) with predicted ATP binding and inositol pentakisphosphate 2-kinase activity |
| Chr6_A_oryzae_RIB40:1409858-1410070 | AO090020000161 | Ortholog of A. nidulans FGSC A4 : AN6458, A. fumigatus Af293 : Afu3g07420, A. niger CBS 513.88 : An02g10960, An01g01420, An12g05390, and A. oryzae RIB40 : AO090005000921, AO090003001427 |
| Chr6_A_oryzae_RIB40:1409858-1410070 | AO090020000162 | Has domain(s) with predicted UDP-N-acetylmuramate dehydrogenase activity, flavin adenine dinucleotide binding, oxidoreductase activity, oxidoreductase activity, acting on CH-OH group of donors activity |
| Chr6_A_oryzae_RIB40:4172129-4172341 | AO090138000010 | Has domain(s) with predicted 2-dehydropantoate 2-reductase activity, NADP binding, coenzyme binding, oxidoreductase activity, oxidoreductase activity, acting on the CH-OH group of donors, and NAD or NADP as acceptor activity |
| Chr5_A_oryzae_RIB40:1114034-1114246 | AO090701000470 | Ortholog of A. fumigatus Af293 : Afu2g16985, A. wentii : Aspwe1_0171101, A. clavatus NRRL 1 : ACLA_075940, and A. zonatus : Aspzo1_0015705 |
| Chr3_A_oryzae_RIB40:1794701-1794913 | AO090023000683 | Protein of unknown function |
| Chr6_A_oryzae_RIB40:3558859-3559071 | AO090038000040 | Has domain(s) with predicted catalytic activity, catechol 1,2-dioxygenase activity, ferric iron binding, iron ion binding and oxidoreductase activity, more |
| Chr1_A_oryzae_RIB40:3825460-3825672 | AO090005000971 | Has domain(s) with predicted role in biosynthetic process |
| Chr5_A_oryzae_RIB40:4076380-4076592 | AO090113000012 | Ortholog of A. nidulans FGSC A4 : AN6909/BEST2, A. niger CBS 513.88 : An14g05100, A. wentii : Aspwe1_0153378, A. sydowii : Aspsy1_0054771, and A. terreus NIH2624 : ATET_06151 |
| Chr7_A_oryzae_RIB40:2355701-2355913 | AO090011000905 | Ortholog(s) have sequence-specific DNA binding transcription factor activity |
| Chr7_A_oryzae_RIB40:2498610-2498822 | AO090011000954 | Ortholog(s) have cytosol localization |
| Chr8_A_oryzae_RIB40:1654282-1654494 | AO090010000667 | Has domain(s) with predicted iron ion binding and oxidoreductase activity; role in fatty acid biosynthetic process and oxidation-reduction process |
| Chr3_A_oryzae_RIB40:244139-244351 | AO090023000096 | Has domain(s) with predicted catalytic activity, coenzyme binding activity, and role in cellular metabolic process |
| Chr8_A_oryzae_RIB40:1372821-1373033 | AO090010000775 | Ortholog(s) have UDP-N-acetylglucosamine transmembrane transporter activity and role in UDP-N-acetylglucosamine transport, UDP-glucose transport, fungal-type cell wall chitin biosynthetic process, and transmembrane transport |
| Chr1_A_oryzae_RIB40:3919266-3919478 | AO090005000937 | Ortholog of A. nidulans FGSC A4 : AN1323 and A. flavus NRRL 3357 : AFL2T_00913 |
| Chr6_A_oryzae_RIB40:442735-442947 | AO090020000539 | Ortholog(s) have Golgi apparatus, endoplasmic reticulum localization |
| Chr8_A_oryzae_RIB40:2804269-2804481 | AO090010000224 | Ortholog(s) have role in ethanol metabolic process and mitochondrial inner membrane localization |
| Chr8_A_oryzae_RIB40:2804269-2804481 | AO090010000223 | 40S ribosomal protein S2-like protein; predominantly expressed in the hyphal tip region |
* Original description is based on information provided by the Comprehensive Aspergillus oryzae Genome Database (CAoGD) (<https://nribf21.nrib.go.jp/CAoGD/>.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



