
Submitted:
05 January 2026
Posted:
06 January 2026
You are already at the latest version
Abstract
Prospecting for a future breeding site is an essential component of informed natal and breeding dispersal. It allows individuals to reduce the uncertainty of their environment by gathering personal and social information about the local quality of alternative breeding areas, and make informed emigration and settlement decisions. Although this process has been studied in territorial and social animal species for decades, it is still understudied and not well-understood. Using 124 empirical studies which have explicitly described prospecting in a context of breeding habitat selection in birds, mammals, fish and invertebrates, I review why, how, when and which individuals prospect according to various life history traits and sociality. Multiple classes of individuals were identified prospecting and they do so at different times of the year, depending on whether they immediately use the information gathered for settling in a new breeding area or delay it for a future breeding season. Spatial patterns and occurrence of prospecting were hard to generalize over taxa or even species due to substantial intra and inter-specific variability. From this synthesis, I identify persisting knowledge gaps related with the different spatial and temporal scales involved with prospecting and provide key objectives and research directions. I notably propose to examine prospecting at the interface of movement and behavioural ecology. This will be possible by combining different methodologies including tracking and biologging devices, field monitoring surveys and experimental approaches. Overall, a better understanding of the spatial and behavioural processes underlying prospecting will significantly enhance our ability to predict species responses to environmental changes and inform more effectively management plans for threatened species.
Keywords:
behavioural decisions
; breeding habitat selection
; environmental change
; exploratory movements
; extra-territorial forays
; incursions
; information use
; social cues
; movement ecology
I. Introduction
Finding a place to breed is a vital decision for an individual, since the quality of a breeding site or territory can directly and indirectly affect individual breeding success and survival (Clobert et al. 2012). Some individuals can decide to breed where they were born and remain faithful to their breeding site for their entire life. Yet, environmental conditions vary over time and philopatry may not always be the best strategy to maximize individual lifetime fitness (Doligez et al. 2003, Delgado et al. 2011, Ponchon et al. 2015b). Kin competition, inbreeding avoidance or degrading local environmental conditions may also drive individuals to seek for a new breeding site, leading to dispersal. Dispersal is generally subdivided into three main stages: (1) emigration from the natal or current breeding site, (2) transience, when individuals move through the landscape, and (3) settlement in a new breeding site (Clobert et al. 2012).
In the field, numerous correlative and experimental studies have demonstrated that individuals could gather and use information about their environment to select their breeding habitat and thereby, make informed emigration and settlement decisions (Reed et al. 1999). This complex decision-making process involves two distinct steps: the information gathering phase, called prospecting, and the information use phase which determines settlement decisions. Prospecting basically consists in exploring the environment and could be associated with exploratory movements but in a specific context of breeding habitat selection, prospecting is defined as the visits of individuals to breeding areas or territories other than their current one, from where they can gather information about the local quality of a habitat and its breeding sites. It can occur before emigration, when individuals may consider whether to leave their current breeding site, or during transience, after individuals have decided to disperse towards a new breeding site (Reed et al. 1999). During prospecting, individuals can acquire personal information from their direct interaction with their environment but also from social information, based on the presence and/or performance of their conspecifics or heterospecifics (Dall et al. 2005). Overall, prospecting is expected to help individuals increase their fitness prospects by leading them to leave breeding sites of low quality and settle in better ones (Ponchon et al. 2021).
Prospecting for breeding habitat selection has been identified in many colonial, territorial and social species, and its ecological and evolutionary significance has largely been emphasized, notably its crucial implications on non-random gene flow and population connectivity (Reed et al. 1999, Delgado et al. 2011, Ponchon et al. 2013, Boulinier et al. 2016). Yet, to date, no synthesis covering a wide range of taxa currently exists. One difficulty to assess the universality of prospecting relates to the challenge of monitoring the movements of free-ranging individuals at a fine temporal resolution and over long periods (Ponchon et al. 2013). Moreover, studies often focus on one dispersal stage: either emigration, or settlement (Reed et al. 1999). Hence, prospecting is most of the time missed or ignored.
On the other side, many different terms are used for prospecting: exploratory movements, extra-territorial visits, extra-territorial forays, hidden long-dispersal movements, pre-dispersal excursions, intrusions or incursions, which complicates the literature search (Reed et al. 1999). Moreover, visits of individuals in alternative breeding sites are sometimes reported but prospecting is not explicitly named (e.g. Choden et al. 2019). In other cases, prospecting incorrectly refers to the return of immature seabirds to their natal colony before their recruitment (e.g. Becker and Bradley 2007, Bosman et al. 2013). This is not prospecting, as it does not involve gene flow towards non-natal colonies (Reed et al. 1999). The use of the same word in different contexts and conversely, different words in the same context is therefore confusing and hampers a general comprehension of prospecting for breeding habitat selection. Finally, the types of personal and social information collected and their use for breeding habitat selection have been widely reviewed from a behavioural perspective but the spatial process underlying information gathering during prospecting has seldom been addressed (e.g. Valone and Templeton 2002, Blanchet et al. 2010).
This review aims at providing evidence for the widespread occurrence of prospecting based on 124 empirical studies which have explicitly described prospecting in a context of breeding habitat selection. Those empirical examples, covering 94 different species of birds, mammals, fish and invertebrates, are used to synthesize why, how, when and which individuals prospect according to their life history traits and sociality. Then, I outline persisting knowledge gaps and propose an integrative framework at the interplay of behavioral and movement ecology with key objectives to point out future research direction.
II. Study of Prospecting Movements over the Last Two Decades
A first extensive review documenting prospecting in birds was published 25 years ago (Reed et al. 1999). Most of the studies cited relied on the direct observations of ringed birds. Yet, this method limits the detection of prospecting individuals at spatial scales larger than the monitored areas. Since then, a broader range of approaches for tracking organisms have yielded new perspectives in the study of prospecting (Ponchon et al. 2013).
The following section compiles evidence of prospecting based on a search of papers published from 1999 to 2022 for birds and any time up to 2022 for other animal taxa because of their rarity. Studies were identified through Google Scholar searches and via citations by relevant articles. The following search terms were used: ‘prospecting’ AND breeding habitat selection; ‘prospecting’ AND ‘information use; ‘prospecting’ AND ‘dispersal’; ‘extra-territorial movements’ AND dispersal; ‘exploratory movements’ AND ‘breeding habitat selection’; ‘forays’ AND “prospecting” AND ‘breeding habitat selection’. To be included, studies had to explicitly describe visits or movements of individuals to other breeding territories or habitats or social groups than their own in a context of breeding habitat selection or dispersal. When social information was addressed, studies focusing on heterospecific information were excluded, as they only correlated information use with settlement decisions without explicitly considering prospecting movements (e.g. Kivelä et al. 2014 and references therein).
I found 105 studies which discussed or explicitly mentioned prospecting in a context of breeding habitat selection, natal dispersal or breeding dispersal, 13 studies reporting extra-territorial movements exclusively in a context of extra-pair copulation and 6 studies addressing prospecting in a context of parasitic breeding birds (see a detailed list of all 124 references and their main attributes at https://github.com/auponchon/Prospecting_review/blob/master/data/References_used_for_review.xlsx). Species sociality was either territorial (51%), cooperative or social (21%), colonial (19%), nest parasitic (5%) or non-territorial/non-colonial (5%).
Among taxa, birds were the most represented (71%), followed by mammals (24%; Figure 1a). A majority of studies (65%) used a tracking system to follow prospecting but only 25% deployed devices recording the continuous movement of animals in their environment (e.g. GPS or Platform Terminal Transmitters=PTT; Figure 1b), all focusing either on birds or mammals (Table 1). The number of publications documenting prospecting increased in the 2000s, but later remained stable (Figure 1c) despite the development of tracking devices which facilitated the identification of prospecting over distances up to hundreds of kilometres (Table 1). 84% of studies addressed at least one specific factor driving prospecting while 10% did not include any environmental, social or individual factor (=general exploration) and 6% described prospecting opportunistically (Figure 1d).
II.1. Why? Function of Prospecting for Breeding Habitat Selection
Breeding habitat selection can be viewed as a hierarchical process where individuals first select a relatively homogeneous favourable environment to live in (= a habitat) within which they further select a site for reproduction (Orians and Wittenberger 1991).
At the scale of a habitat, prospecting may allow individuals to assess the general quality of the environment by gathering personal information from their direct interaction with the environment (Dall et al. 2005). Individuals are expected to rely on all the visual, auditory, olfactory or chemical cues that are available to them. Accordingly, 19% of studies showed or suggested correlations between prospecting and habitat quality reflected by the physical structure of the environment (e.g. vegetation structure, Arlt and Pärt 2008; Rémy et al. 2011; forest cover, Cox and Kesler 2012, Trainor et al. 2013; snow depth, Therrien et al. 2015), the potential presence of predators (Thomson et al. 2013) and the current availability of vacant territories (Armstrong et al. 1997, Vangen et al. 2001, Bruinzeel and van de Pol 2004, Soulsbury et al. 2011, Veiga et al. 2012). 6% of studies also suggested that by repeatedly visiting the same habitat or social groups, individuals accumulated personal information and progressively became more familiar with their surroundings (Haughland and Larsen 2004, Selonen and Hanski 2006, Jungwirth et al. 2015, Patchett et al. 2022). Repeated prospecting visits also allowed individuals to update information to fine-tune current investment in reproduction and territory defence (Sicotte and Andrew 2004, Young et al. 2005) or monitor changes in neighbouring social groups (Mayer et al. 2017, Barve et al. 2020, Kelly et al. 2020).
Within a habitat, prospecting may allow colonial or territorial species to locate and start acquiring a new breeding site or territory (Reed et al. 1999). In social or cooperative species, individuals are not directly searching for a territory but rather a social group defending a common territory. In the 16 different social and cooperative species reviewed, prospecting allowed individuals to be accepted more easily by the targeted group and potentially acquire a higher breeding position (fish: Jungwirth et al. 2015; birds: Williams and Rabenold 2005, Kingma et al. 2016; mammals: Teichroeb et al. 2011, Mares et al. 2014). It also represented an opportunity for males to expand their territory (Mayer et al. 2017), change their territory more rapidly (Kelly et al. 2020) or acquire new mates via female transfer or group take-over, creating new breeding opportunities (Doolan and Macdonald 1996, Sicotte and Andrew 2004).
Avian brood parasites such as cuckoos, goldeneyes or cowbirds lay their eggs in nests of other bird species which provide parental care for the parasite eggs and chicks. Among the five reviewed species, females prospected to search for a nest site where they could safely lay eggs and where hosts were assessed as being of good-quality (Honza et al. 2002, Scardamaglia et al. 2016, White et al. 2017). In addition, prospecting allowed females to identify successful active nests cavities with lower predation risk and lay their parasite eggs in safer places (Pöysä et al. 1999, Pöysä 2006).
Finally, 3 mammal and 8 passerine species visited other territories during the breeding/fertility period to increase extra-pair copulation opportunities (e.g. Naguib et al. 2001, Stutchbury et al. 2005, Young et al. 2007, Ward et al. 2014, Carter et al. 2018). This type of prospecting is unlikely to trigger dispersal, as individuals are not searching for a new breeding site but rather a temporary mate (Debeffe et al. 2014 and references therein, but see Williams and Rabenold 2005). It may still be called prospecting because such movements ultimately affect gene flows. But the motivation of “extra-pair copulation” should always be precised, as it may trigger different behavioural and space use patterns compared to prospecting for breeding habitat selection (e.g. in red foxes Vulpes vulpes; Soulsbury et al. 2011).
Even if prospecting is expected to be primarily used to select a future breeding site or social group, it was not always followed by dispersal. In some cases, prospecting individuals eventually remained philopatric, possibly because their current breeding site was of better quality than the prospected ones or because they preferred remaining in a familiar environment (Vangen et al. 2001, Haughland and Larsen 2004, Ducros et al. 2020).
Altogether, the empirical studies reviewed confirm that prospecting is a powerful way of gathering and updating information on the environment to ultimately decide both whether to leave the natal/current breeding territory or social group and subsequently, where to settle.
II.2. Who? Classes of Prospectors
Prospecting is expected to help individuals select a new breeding site so theoretically, it should be primarily displayed by individuals in need of a new breeding site or not currently investing in reproduction, i.e. young or non-breeding individuals (Reed et al. 1999). Life stage and breeding status should thus play a major role in the occurrence of prospecting and 38% of studies addressed it (Figure 1d).
In territorial or social species where individuals quickly become sexually mature and can breed as soon as their first year, juveniles are expected to rapidly leave their natal patch when becoming independent (Greenwood 1980). Therefore, they may engage in prospecting not only to explore and get familiar with their environment but also locate potential future breeding territories. This was the case for 8 mammal species (Harrison et al. 1991, Vangen et al. 2001, Haughland and Larsen 2004, Selonen and Hanski 2006, Mabry and Stamps 2008, Rémy et al. 2011, Samelius et al. 2012, Debeffe et al. 2013).
In colonial or territorial species with a longer immaturity period such as seabirds or raptors, individuals may wait several months/years before actually settling in a territory. Accordingly, immatures/subadults of 14 species were regularly observed prospecting (e.g. Bradley et al. 1999, Balbontín and Ferrer 2009, Fasciolo et al. 2016, Campioni et al. 2017, Ciaglo et al. 2021, Poessel et al. 2022). For social and colonial species becoming highly faithful to their breeding territory once they had settled, young individuals prospected more than adults (Sánchez-Tójar et al. 2017, Campioni et al. 2017, Wolfson et al. 2020, Kelly et al. 2020, Poessel et al. 2022). On the contrary, when breeding adults reassessed their site fidelity regularly, prospecting was frequent, especially in birds willing to renest within the same breeding season after an early breeding failure or living in unstable environments (Ward 2005, Thomson et al. 2013, Martinović et al. 2019).
Breeding performance is also expected to be a major driver of prospecting. In birds, it is mostly expected to occur in adult failed breeders or non-breeders (=floaters), because those individuals generally have to decide whether to emigrate and where (Reed et al. 1999). In line with this, 5 studies detected prospecting only in failed breeders, not in successful ones (Amrhein et al. 2004, Calabuig et al. 2010, Fijn et al. 2014, Ponchon et al. 2015a, 2017b). On the contrary, successful breeders are expected to fully invest in reproduction to maximize offspring survival, potentially leaving no time for prospecting (Reed et al. 1999). Yet, prospecting was still observed in active successful breeders (Blakey et al. 2019, Martinović et al. 2019, Cooper and Marra 2020, Oro et al. 2021). Moreover, a recent study has revealed prospecting occurrence in 14 species of active breeding seabirds, especially those living in unstable environments (Kralj et al. 2023). Those findings challenge the general assumption that prospecting mostly follow individual breeding failure in long-lived colonial species living in predictable environments (Reed et al. 1999, Doligez et al. 2003) and this requires a complete reassessment of prospecting occurrence regarding interactions between breeding status and environmental variability.
According to the way individuals select their breeding site, i.e. whether they directly acquire it or indirectly select it through a mate/social group which already owns a site, prospecting is expected to depend on sex (Reed et al. 1999). Among the 47 studies which directly compared prospecting between males and females, 53% found that males prospected more than females (e.g. Doligez et al. 2004, Selonen and Hanski 2006, Balbontín and Ferrer 2009, Ponchon et al. 2017b), 21% found that females prospected more than males (e.g. Ward 2005, Calabuig et al. 2010, Rémy et al. 2011, Debeffe et al. 2014, Blakey et al. 2019), and 26% found no difference (e.g. Conradt et al. 2000, Thomson et al. 2013, Jungwirth et al. 2015, Campioni et al. 2017, Poessel et al. 2022). No clear pattern emerged according to taxa or sociality, except that females never prospected more than males in seabirds. Hypotheses regarding why such variability arises between and within taxa are still scarce. Understanding how sex drives prospecting patterns and strategies would help better understand sex-biased dispersal, which has multiple crucial ecological and evolutionary consequences at the individual and population level (Li and Kokko 2019).
In 4 cooperative bird, mammal and fish species, subordinates and helpers prospected more than dominants, as they were more likely to seek for new breeding opportunities in neighbouring social groups and evaluate potential competitors (Young et al. 2005, Kesler and Haig 2007, Mayer et al. 2017). Yet, dominants still prospected to find opportunities to expand their current territory, especially when they owned small or low-quality ones (Jungwirth et al. 2015, Mayer et al. 2017, Barve et al. 2020).
In 3 studies addressing dispersal status, individuals that later dispersed (=dispersers) performed more prospecting trips before definitely leaving their territory compared to philopatric individuals (Soulsbury et al. 2011, Debeffe et al. 2014, Oro et al. 2021). In 4 other cases, individuals prospecting the highest number of patches or travelling the furthest were not the ones which later dispersed (Dique et al. 2003, Selonen and Hanski 2010, Jungwirth et al. 2015, Ponchon et al. 2017b). Therefore, prospecting occurrence cannot be used as a reliable predictor of dispersal.
Finally, 5 studies revealed that individuals could prospect in groups: males of social species such as meerkats (Doolan and Macdonald 1996), primates (Sicotte and Andrew 2004), jays (Williams and Rabenold 2005) or badgers (Roper et al. 2003) formed coalitions to prospect, while trogons, territorial passerines, constituted assemblages of both sexes (Riehl 2008). Visiting other breeding territories in groups may allow individuals to acquire information more safely, such as through diluting predation risk and possibly outcompeting conspecifics more easily to acquire new social status or extend an existing territory. This thereby may provide various indirect fitness benefits which may promote prospecting.
Overall, the review reveals that multiple classes of individuals prospect, including successfully breeding or dominant individuals which are theoretically not particularly expected to do so (Table 2). It points out the lack of understanding in the combined effects of individual and social factors on prospecting occurrence and patterns.
II.3. When and What? Timing of Prospecting and Available Cues
Thanks to a variety of personal and social cues (Dall et al. 2005), prospecting individuals may evaluate the quality of alternative breeding habitats at various times of the year, during or outside the breeding season (Table 3). The timing of prospecting thus depends on when individuals use the information gathered to settle in a new breeding site or social group: immediately, for the current breeding season or with a delay, for the next one (Reed et al. 1999). It also depends on the types of personal and social information available to individuals. In the reviewed studies, 27% examined the timing of prospecting, 28% correlated prospecting with the gathering of social cues and 10% addressed these two factors simultaneously.
Prospecting during the pre-breeding or nest building/territory acquisition season implies that individuals are likely to immediately use the gathered information. Yet, individuals cannot directly rely on the breeding performance of their conspecifics, as mating has not taken place yet. Instead, some species relied on physical features of the environment (e.g. snow cover and depth for snowy owl Bubo scandiacus Therrien et al. 2015), breeding status and timing of singing around dawn in nightingales Luscinia megarhynchos (Amrhein et al. 2004, Roth et al. 2009) or the general activity of conspecifics (brown jays Cyanocorax morio Williams and Rabenold 2005). When prospecting occurs during laying and egg incubation in birds, public information, which corresponds to social information based on the breeding performance of conspecifics (Danchin et al. 2004), becomes available. The number of incubating conspecifics and the number of eggs in nests constituted good proxies of the current environmental quality for many prospecting bird species (Ponchon et al. 2015a, Martinović et al. 2019, Oro et al. 2021), including avian brood parasite birds (Honza et al. 2002, Pöysä 2006, White et al. 2017).
The later prospecting occurs during the breeding season, the more likely individuals are to use information for the next breeding season. Yet, this delay between information gathering and use may only be beneficial when the environmental variability is predictable over time (Doligez et al. 2003). Accordingly, a peak of prospecting was generally observed during the chick-rearing period in birds, when public information based on the number and quality of fledglings was the most available and the most reliable (Doligez et al. 2004, Ward 2005, Veiga et al. 2012).
Prospecting may happen during the post-breeding season, when territories have been deserted. During this period, social information may be less available and more uncertain. Yet, individuals still used cues to assess the quality of breeding sites such as the location of occupied breeding sites (Arlt and Pärt 2008, Ciaglo et al. 2021, Patchett et al. 2022) or eggshell fragments and membranes in successful nests (Pöysä 2006).
Outside the breeding season, prospecting mainly occurred in non-migratory mammals (Deuel et al. 2017, Mayer et al. 2017, Mancinelli and Ciucci 2018, Kelly et al. 2020, but see Sánchez-Tójar et al. 2017 for birds). By doing so, individuals monitored neighbouring group composition, conspecific competitive abilities and sexual status, notably through personal information such as olfactory cues.
II.4. How? Patterns of Prospecting
Prospecting can be fundamentally described by two main behavioural tactics: the ‘best-of-n’ strategy and the ‘sequential sampling’ (Reed et al. 1999). The best-of-n strategy consists for prospecting individuals to visit one or several alternative breeding areas while still regularly coming back to their natal/current breeding site. It allows individuals to gather information about different breeding areas and eventually select the one they consider the best among the ones visited. This strategy was observed in species with high mobility abilities but facing limited number of alternative breeding areas (Balbontín and Ferrer 2009, Kesler and Haig 2007, Gaughran et al. 2019, Roper et al. 2003, Ponchon et al. 2015a). The sequential sampling occurs when individuals prospect continuously after definitely leaving their current natal or breeding area. Individuals visit one patch after the other and based on an implicit threshold, they settle in the first breeding site they consider suitable enough (Stamps et al. 2005). This strategy was mainly displayed by species with limited mobility and thus, limited ability to easily find suitable patches in a heterogeneous landscape (Dale et al. 2006) or by species with high constraints in breeding site selection (Armstrong et al. 1997, Therrien et al. 2015).
Regardless of behavioural tactics, taxa or life history traits, this synthesis highlighted an extreme variability in prospecting strategies, within and between species, populations and individuals. For instance, both prospecting strategies were observed in flying squirrels Pteromys volans, where the best-of-n strategy was linked with short-distance dispersal and sequential search with long-distance dispersal (Selonen and Hanski 2010). Likewise, some individuals from the same population exhibited prospecting while others fully remained philopatric to their natal patch and others just dispersed directly to a new territory without prospecting (Harrison et al. 1991, Armstrong et al. 1997, Mabry and Stamps 2008, Ducros et al. 2020, Engler and Krone 2022). Thus, some individuals may be more prone to engage in prospecting than others, possibly due to a cost/benefits trade-off between philopatry and dispersal which in turn may depend on intrinsic, social and/or environmental factors. Personality has been proposed to explain some inter-individual variability observed in movement strategies, with bolder and more aggressive individuals exploring more and farther their environment (Burkhalter et al. 2015, Spiegel et al. 2017). This could also be the case for prospecting but very few empirical studies have tested it so far and their results were inconclusive (e.g. Schuett, Laaksonen & Laaksonen, 2012).
One factor likely influencing prospecting and seldom acknowledged in the reviewed studies was the spatial scale of prospecting (Wiens 1976). Indeed, an individual moving over a few hundred meters in the same breeding habitat may be considered as prospecting as soon as the movement is a response to the local quality of the environment, such as poor territory features, increased presence of predators or strong competition to acquire a territory (e.g. Bradley et al., 1999). At the same time, prospecting and subsequent dispersal movements at large spatial scale might be more risky and time consuming for individuals (Stamps et al. 2005) so there may be an important trade-off between the spatial scale of the environmental factor to escape from and the spatial scale of prospecting movements and dispersal. Adopting the wrong spatial scale of observation to study individual prospecting and dispersal may notably under- or over-estimate population connectivity (Doligez et al. 2008). Despite its obvious importance, the hierarchical spatial aspect of prospecting needs to be better considered when studying prospecting (Gaillard et al. 2010).
II.5. How? Costs of Prospecting
As for dispersal, prospecting is likely to entail multiple costs (Stamps et al. 2005) but only 5% of studies explicitly addressed it (Figure 1d). First, prospecting may require time and energy to access the different breeding areas, and thus, may need to be traded against other essential activities such territory defence, mate bonding, grooming or foraging for food. For example, in black-legged kittiwakes Rissa tridactyla, prospecting failed breeders spent less time foraging compared to non-prospecting successfully breeding ones (Ponchon et al. 2015a).
When individuals visit other breeding areas than their own, they may potentially cross unfamiliar or hostile environments. Accordingly, predation risk and confrontation with aggressive conspecifics entailed injuries (Crawford 2015, Kingma et al. 2016, Mayer et al. 2017) while long-distance and risky movements increased energetic demands and stress levels (Young and Monfort 2009), leading to lower body condition (Kingma et al. 2016). Further, an increase of the frequency of prospecting over years drastically reduced individual survival (Cram et al. 2018).
Prospecting may only be beneficial for individuals if the information gathered actually reflect the environmental quality and are thereby informative and reliable (Dall et al. 2005). Missing the right timing or using erroneous cues may lead individuals to settle in suboptimal territories which may ultimately reduce their fitness, as experimentally observed in social bobolinks Dolichonyx oryzivorus (Nocera et al. 2006).
Locally, prospecting often involves a close inspection of the environment and encounters with conspecifics, which may facilitate disease transmission (Boulinier et al. 2016, Gaughran et al. 2019). As prospecting can occur at large spatial scales (Table 1), it could further accelerate disease circulation to distant populations. Yet, linking prospecting to disease propagation still remains a challenge, as both the infectious status of prospectors and individuals present in the visited breeding areas would have to be monitored regularly, which might not be easy at large spatial scales and in dense populations. Integrating prospecting in eco-epidemiological models might be key to better understand the dynamics of host-pathogens interactions and their effects on population dynamics (Boulinier et al. 2016).
III. Future Research Directions
III.1. Prospecting as Both a Behavioural and Movement Process
The present literature review provides evidence of prospecting for breeding habitat selection in a wide range of species and taxa. However, most of the studies were not replicated and we still currently lack a unified conceptual framework predicting (1) how and where individuals prospect for a breeding habitat according to local and regional environmental conditions and (2) what type of information and cues they actually gather based on personal and social cues.
This is because prospecting involves two inextricable processes that are challenging to study simultaneously with the same methodology: a spatial process linked with individual movement and a behavioural process linked with information gathering. For a long time, observational and experimental behavioural studies have brought actual evidence of information gathering by individuals. They correlated potential available cues with the actual visits of individuals in other breeding areas but also the settlement decisions subsequently observed (e.g. Doligez et al. 1999, Calabuig et al. 2010, Rémy et al. 2011). However, detailed data of individual movements were lacking. More recently, studies using tracking devices have highlighted the spatial process of prospecting (Table 1) but the information gathering process was only implicit: movements were assigned to prospecting because they were subjectively defined as visits in other known or suspected breeding areas. In fact, they rarely provided direct data on how prospecting movements and fine-scale behaviour determined subsequent breeding site selection and settlement. This persisting limit could be closed by integrating prospecting behaviour reflected by the gathering and use of information into the movement ecology framework (Nathan et al. 2008), where individual movement paths are the results of external factors, internal state, navigation and motion capacity (Figure 2).
By doing so, collecting observational data on the spatial and social environment of individuals and carrying out experiments in the field would provide information on the behavioural components of prospecting while tracking individuals with bio-loggers and using cutting-edge modelling tools would provide information on the spatial components of prospecting (Figure 2). It would reveal patterns that may further predict who is likely to prospect and when according to individual, social and environmental factors and resulting movement paths. Moreover, quantifying more precisely how each component entails prospecting costs on individual breeding success and survival will help understand their short- and long-term ecological and evolutionary consequences both at the individual and population level.
III.2. How to Fill the Persisting Gaps?
Objective 1: Acknowledging prospecting as a movement as important as foraging or migration.
In movement ecology, prospecting tends to be considered as a classic exploratory movement with no fitness consequence. Yet, identifying prospecting correctly and using the same word and definition for all species and social contexts may foster further research in the field and help better understand informed dispersal. Then, an effort should be made to identify potential territories and suitable habitats that could be prospected directly in the field and determine the type of information actually available for prospectors depending on the timing of prospecting. This will be more achievable for colonial or social species living in dense groups, as their breeding sites are easier to identify compared to less aggregated territorial species.
Objective 2: Using tracking devices to address prospecting movements.
The constant improvement of miniaturized tracking devices, with longer batteries, higher temporal and spatial resolution, have offered new opportunities to study the spatial process of prospecting (Ponchon et al. 2013). But this will require a careful and thorough inspection of individual tracks, as prospecting could be overlooked because they are not considered important or worth highlighting in the main results and discussion. In addition, the use of ancillary data (e.g. accelerometry, altitude, heart rate, cameras etc) will further validate and thereby strengthen behavioural inferences (e.g. Saldanha et al. 2023), which will facilitate the interpretation of individual behaviours, especially when the function of different movements may be unclear due to similarity of movement patterns between activities (e.g. Poessel et al. 2022).
Objective 3: Applying appropriate analytical tools to infer prospecting behaviour
The rapid development of cutting-edge statistical models and machine learning tools examining space use from individual movement trajectories have opened large avenues for inferring animal behaviour (Nathan et al. 2022) and offers promising tools for the study of prospecting at a hierarchy of spatial scales. For example, the R package recurse (Bracis et al. 2018) may be used to quantify systematically and robustly recurrent visits of individuals in breeding habitats at a fine spatial scale while momentuHMM (McClintock and Michelot 2018) may infer distinct behavioural states including prospecting from hidden Markov models. Additionally, habitat selection functions may be used to examine how individuals select particular habitats according to different covariates and different life stages (Northrup et al. 2022). Environmental variables (e.g. environmental predictability, air temperature, food conditions, human activities) could be included to investigate the potential factors favouring or impeding prospecting, both from a behavioural and movement perspective.
Objective 4: Combining tracking data with field-based monitoring surveys and experiments
For the moment, the acquisition of tracking data is limited by battery capacity and species body size (Nathan et al. 2022). Relying only on tracking devices may prevent researchers from relating information gathering during prospecting with information use during the settlement stage, especially when those two processes are disrupted in time, as in migratory species (Clobert et al. 2012). This is why a combination of spatial and behavioural approaches are needed to relate the different stages of informed dispersal encompassing prospecting (Figure 2). This step will be key to examine prospecting at a hierarchy of spatial and temporal scales which will reflect the scales at which individual collect information on their environment and thereby, the scale at which individuals decide to settle (Doligez et al. 2008). Further, experiments such as modifying social cues by placing decoys or playing playbacks in different suitable habitats, territories or social groups while tracking individuals from different classes will help uncover the drivers and mechanisms of prospecting.
Objective 5: Improving theoretical models to broaden the conceptual framework of prospecting
Despite its crucial ecological and evolutionary consequences on population dynamics, structure and persistence (Delgado et al. 2014, Ponchon et al. 2015b, Ponchon and Travis 2022), prospecting has rarely been explicitly and thoroughly implemented into theoretical models (Ponchon et al. 2021). Consequently, clear hypotheses and predictions are still missing when it comes to understand how sociality, life history traits (e.g. longevity, fecundity), species mobility or individual personality shape prospecting occurrence and patterns in different taxa and what the fitness benefits are.
One way to better integrate prospecting in theoretical studies would be to explicitly implement the different phases of informed dispersal: 1) emigration decision, 2) prospecting (“best-of-n strategy; (Ponchon et al. 2021)) or spatially explicit searching phase (sequential sampling; (Delgado et al. 2014)) and 3) settlement decision. There is also a much-needed work on sex-biased and age-specific prospecting strategies to better understand the underlying mechanism of informed dispersal and their consequences on individual fitness. Newly developed theory would then become testable, driving new experiments in the field with the use of tracking devices. Such results would in turn feed back into the theory to fine-tune the way prospecting is modelled. Overall, with a strong fundamental understanding of prospecting, we would be in a better position to build more realistic predictive models of species’ response to environmental changes, fully embedding informed dispersal (Urban et al. 2016).
III.3. Prospecting in a Changing World
Prospecting has been identified as one key individual behavioural strategy to respond to rapid environmental change, as it may help individuals leave habitats of low quality and settle in habitats of better quality (Ponchon et al. 2015b, 2021). The resulting dispersal at the individual level is expected to have direct and immediate consequences at the population level, with a non-random spatial redistribution of individuals (Ponchon et al. 2015b). Yet, environmental change and the occurrence of extreme climate events may disrupt the availability and reliability of the cues used by individuals and may jeopardize the benefits of informed dispersal. In particular, if environmental or social cues incorrectly reflected the local quality of the environment at the time of prospecting, individuals may gather erroneous information about the environment and may end making poor settlement decisions, thereby jeopardizing their fitness (McNamara et al. 2011). The use of such cues would become maladaptive and would lead the population to ecological traps, as already evidenced in red-necked grebes Podiceps grisegena (Kloskowski 2021).
Conversely, if individuals could appropriately switch the cues they rely on and adjust the timing of their collection, prospecting could constitute a powerful way to efficiently overcome environmental change and track more efficiently breeding habitats of good quality. Nevertheless, the time of adaptation of using new cues would have to be quicker than environmental changes (Ponchon et al. 2015b). In particular, prospecting could lead to the colonization of new breeding areas, including some out of the current species range (Kokko and Lopéz-Sepulcre 2006). Yet, the speed and success of colonization may highly depend on individual preferences between settling in an empty habitat or settling in habitats already occupied by conspecifics. This implies that in addition to the timing prospecting and the types of cues, the balance between the use of personal information versus social information should be crucial in the success of populations to track climate change (Ponchon and Travis 2022).
New evidence of prospecting from species living in ephemeral environments and during active breeding have also challenged long-standing theory (Kralj et al. 2023), suggesting that prospecting could actually constitute an adaptive response to rapid environmental changes. However, this would only be the case if individuals used information immediately after prospecting, so that the value of information would still correctly reflect the local quality of the habitat and individuals would still choose habitats of better quality (McNamara et al. 2011).
In addition to natural adaptation, which might sometimes be too slow for species to keep pace with environmental changes, humans could help species make better settlement decisions by providing artificial cues such as decoys or playbacks to attract them towards habitats of better quality when their current ones suffer from severe degradation. Having a good knowledge of prospecting may favour successful conservation actions by identifying the right types of habitats to be colonized and by providing the appropriate cues to the right individuals at the right time of year to make it the most effective. Several colonization experiments have already been carried out in birds and results have generally been encouraging (Ahlering et al. 2010). Yet, such conservation strategies would have to be monitored scrupulously to ensure actions are not detrimental to the population and do not trigger unforeseen negative consequences.
IV. Conclusions
By bringing together a large and scattered pool of literature, this review confirms that prospecting is widespread, not only in birds, as previously stressed, but also in many other animal taxa, which display a large range of life history traits and socialities. Nevertheless, prospecting occurrence, patterns and strategies are highly variable within and among species but also within populations of the same species, which complicates their fine understanding. Empirical work can be enhanced with the combination of long-term monitoring surveys, the deployment of tracking devices, the use of appropriate mathematical models and experimental approaches to simultaneously address prospecting at the interface of behavioural and movement ecology. Additionally, theoretical work will be boosted by explicitly integrating prospecting within informed dispersal. It will help understand its ecological and evolutionary consequences at the individual and population level and will be key in drawing new hypotheses depending on environmental variability, species life history traits and sociality. Fostering research on prospecting and informed dispersal will shed light on this potential adaptive response that may help species persist on the long-term thanks to dispersal from degrading environments to better ones. It will also help inform more effective management plans by attracting endangered species toward habitats of better quality.
Acknowledgments
I thank Thierry Boulinier who got me into the fascinating world of prospecting and with whom I have shared so many passionate discussions over the years. This work was initiated during my Marie-Sklodowska-Curie project EcoEvoProspectS (grant agreement no. 753420) and I thank attendees of the purple patch pumpkin meeting held in Corbières (France) back in Octobre 2018 who shared their opinion about my initial ideas. I am particularly grateful to Justin Travis, David Grémillet, Valeria Romano and two anonymous reviewers who provided constructive comments on earlier versions of this review.
References
- Ahlering, M. A.; Arlt, D.; Betts, M. G.; Fletcher, R. J.; Nocera, J. J.; Ward, M. Research needs and recommendations for the use of conspecific-attraction methods in the conservation of migratory songbirds. The Condor 2010, 112, 252–264. [Google Scholar] [CrossRef]
- Amrhein, V.; Kunc, H. P.; Naguib, M. Non–territorial nightingales prospect territories during the dawn chorus. Proceedings of the Royal Society of London. Series B: Biological Sciences 2004, 271, S167–S169. [Google Scholar] [CrossRef] [PubMed]
- Arlt, D.; Pärt, T. Post-breeding information gathering and breeding territory shifts in northern wheatears. Journal of Animal Ecology 2008, 77, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, J. D.; Braithwaite, V. A.; Huntingford, F. A. Spatial Strategies of Wild Atlantic Salmon Parr: Exploration and Settlement in Unfamiliar Areas. Journal of Animal Ecology 1997, 66, 203–211. [Google Scholar] [CrossRef]
- Balbontín, J.; Ferrer, M. Movements of juvenile Bonelli’s Eagles Aquila fasciata during dispersal. Bird Study 2009, 56, 86–95. [Google Scholar] [CrossRef]
- Barve, S.; Hagemeyer, N. D. G.; Winter, R. E.; Chamberlain, S. D.; Koenig, W. D.; Winkler, D. W.; Walters, E. L. Wandering woodpeckers: foray behavior in a social bird. Ecology 2020, 101, e02943. [Google Scholar] [CrossRef]
- Becker, P. H.; Bradley, J. S. The role of intrinsic factors for the recruitment process in long-lived birds. Journal of Ornithology 2007, 148, 377–384. [Google Scholar] [CrossRef]
- Blakey, R. V.; Siegel, R. B.; Webb, E. B.; Dillingham, C. P.; Bauer, R. L.; Johnson, M.; Kesler, D. C. Space use, forays, and habitat selection by California Spotted Owls (Strix occidentalis occidentalis) during the breeding season: New insights from high resolution GPS tracking. Forest Ecology and Management 2019, 432, 912–922. [Google Scholar] [CrossRef]
- Blanchet, S.; Clobert, J.; Danchin, E. The role of public information in ecology and conservation: an emphasis on inadvertent social information. Annals of the New-York Academy of Sciences 2010, 1195, 149–168. [Google Scholar] [CrossRef]
- Bosman, D. S.; Vercruijsse, H. J. P.; Stienen, E. W. M.; Vincx, M.; Lens, L. Age of first breeding interacts with pre- and post-recruitment experience in shaping breeding phenology in a long-lived gull. PLOS ONE 2013, 8, e82093. [Google Scholar] [CrossRef]
- Boulinier, T.; Kada, S.; Ponchon, A.; Dupraz, M.; Dietrich, M.; Gamble, A.; Bourret, V.; Duriez, O.; Bazire, R.; Tornos, J.; Tveraa, T.; Chambert, T.; Garnier, R.; McCoy, K. D. Migration, Prospecting, Dispersal? What Host Movement Matters for Infectious Agent Circulation? Integrative and Comparative Biology 2016, 56, 330–342. [Google Scholar] [CrossRef] [PubMed]
- Bracis, C.; Bildstein, K. L.; Mueller, T. Revisitation analysis uncovers spatio-temporal patterns in animal movement data. Ecography 2018, 41, 1801–1811. [Google Scholar] [CrossRef]
- Bradley, J. S.; Gunn, B. M.; Skira, I. J.; Meathrel, C. E.; Wooller, R. D. Age-dependent prospecting and recruitment to a breeding colony of Short-tailed Shearwaters Puffinus tenuirostris. Ibis 1999, 141, 277–285. [Google Scholar] [CrossRef]
- Bruinzeel, L. W.; van de Pol, M. Site attachment of floaters predicts success in territory acquisition. Behavioral Ecology 2004, 15, 290–296. [Google Scholar] [CrossRef]
- Burkhalter, J. C.; Fefferman, N. H.; Lockwood, J. L. The impact of personality on the success of prospecting behavior in changing landscapes. Current Zoology 2015, 61, 557–568. [Google Scholar] [CrossRef]
- Cadahía Lorenzo, L.; López-López, P.; Urios, V.; Soutullo, A.; Balmaseda, J. J. Negro. Natal dispersal and recruitment of two Bonelli’s Eagles Aquila fasciata: a four-year satellite tracking study. Acta Ornithologica 2009, 44, 193–198. [Google Scholar] [CrossRef]
- Calabuig, G.; Ortego, J.; Aparicio, J. M.; Cordero, P. J. Intercolony movements and prospecting behaviour in the colonial lesser kestrel. Animal Behaviour 2010, 79, 811–817. [Google Scholar] [CrossRef]
- Campioni, L.; Granadeiro, J. P.; Catry, P. Albatrosses prospect before choosing a home: intrinsic and extrinsic sources of variability in visit rates. Animal Behaviour 2017, 128, 85–93. [Google Scholar] [CrossRef]
- Carter, G.; Vorisek, S.; Ritchison, G. Extra-territorial movements by female Indigo Buntings (Passerina cyanea). The Wilson Journal of Ornithology 2018, 130, 1032–1035. [Google Scholar] [CrossRef]
- Casazza, M. L.; McDuie, F.; Lorenz, A. A.; Keiter, D.; Yee, J.; Overton, C. T.; Peterson, S. H.; Feldheim, C. L.; Ackerman, J. T. Good prospects: high-resolution telemetry data suggests novel brood site selection behaviour in waterfowl. Animal Behaviour 2020, 164, 163–172. [Google Scholar] [CrossRef]
- Choden, K.; Ravon, S.; Epstein, J. H.; Hoem, T.; Furey, N.; Gely, M.; Jolivot, A.; Hul, V.; Neung, C.; Tran, A.; Cappelle, J. Pteropus lylei primarily forages in residential areas in Kandal, Cambodia. Ecology and Evolution 2019, 9, 4181–4191. [Google Scholar] [CrossRef] [PubMed]
- Ciaglo, M.; Calhoun, R.; Yanco, S. W.; Wunder, M. B.; Stricker, C. A.; Linkhart, B. D. Evidence of postbreeding prospecting in a long-distance migrant. Ecology and Evolution 2021, 11, 599–611. [Google Scholar] [CrossRef] [PubMed]
- Clobert, J., M. Baguette, T. G. Benton, and J. M. Bullock. 2012. Dispersal ecology and evolution. Oxford University Press, Oxford.
- Conradt, L.; Bodsworth, E. J.; Roper, T. J.; Thomas, C. D. Non-random dispersal in the butterfly Maniola jurtina: implications for metapopulation models. Proceedings of the Royal Society of London. Series B: Biological Sciences 2000, 267, 1505–1510. [Google Scholar] [CrossRef] [PubMed]
- Cooper, N. W.; Marra, P. P. Hidden Long-Distance Movements by a Migratory Bird. Current Biology 2020, 30, 4056–4062.e3. [Google Scholar] [CrossRef] [PubMed]
- Cox, A. S.; Kesler, D. C. Prospecting behavior and the influenceof forest cover on natal dispersal in aresident bird. Behavioral Ecology 2012, 23, 1068–1077. [Google Scholar] [CrossRef]
- Cram, D. L.; Monaghan, P.; Gillespie, R.; Dantzer, B.; Duncan, C.; Spence-Jones, H.; Clutton-Brock, T. Rank-Related Contrasts in Longevity Arise from Extra-Group Excursions Not Delayed Senescence in a Cooperative Mammal. Current Biology 2018, 28, 2934–2939. [Google Scholar] [CrossRef]
- Crawford, J. Conspecific Aggression by Beavers (Castor canadensis) in the Sangamon River Basin in Central Illinois: Correlates with Habitat, Age, Sex and Season. The American Midland Naturalist 2015, 173, 145–155. [Google Scholar] [CrossRef]
- Dale, S.; Steifetten, Ø.; Osiejuk, T. S.; Losak, K.; Cygan, J. P. How do birds search for breeding areas at the landscape level? Interpatch movements of male ortolan buntings. Ecography 2006, 29, 886–898. [Google Scholar] [CrossRef]
- Dall, S. R. X.; Giraldeau, L. A.; Olsson, O.; McNamara, J. M.; Stephens, D. W. Information and its use by animals in evolutionary ecology. Trends in Ecology & Evolution 2005, 20, 187–193. [Google Scholar] [CrossRef]
- Danchin, E.; Giraldeau, L. A.; Valone, T. S.; Wagner, R. H. Public information: from nosy neighbors to cultural evolution. Science 2004, 305, 487–491. [Google Scholar] [CrossRef]
- Debeffe, L.; Focardi, S.; Bonenfant, C.; Hewison, A. J. M.; Morellet, N.; Vanpé, C.; Heurich, M.; Kjellander, P.; Linnell, J. D. C.; Mysterud, A.; Pellerin, M.; Sustr, P.; Urbano, F.; Cagnacci, F. A one night stand? Reproductive excursions of female roe deer as a breeding dispersal tactic. Oecologia 2014, 176, 431–443. [Google Scholar] [CrossRef] [PubMed]
- Debeffe, L.; Morellet, N.; Cargnelutti, B.; Lourtet, B.; Coulon, A.; Gaillard, J. M.; Bon, R.; Hewison, A. J. M. Exploration as a key component of natal dispersal: dispersers explore more than philopatric individuals in roe deer. Animal Behaviour 2013, 86, 143–151. [Google Scholar] [CrossRef]
- Delgado, M. M.; Bartoń, K. A.; Bonte, D.; Travis, J. M. J. Prospecting and dispersal: their eco-evolutionary dynamics and implications for population patterns. Proceedings of the Royal Society B 2014, 281. [Google Scholar] [CrossRef] [PubMed]
- Delgado, M.; Ratikainen, I. I.; Kokko, H. Inertia: the discrepancy between individual and common good in dispersal and prospecting behaviour. Biological Reviews 2011, 86, 717–732. [Google Scholar] [CrossRef]
- Deuel, N.; Conner, L. M.; Miller, K. V.; Chamberlain, M.; Cherry, M.; Tannenbaum, L. Gray fox home range, spatial overlap, mated pair interactions and extra-territorial forays in southwestern Georgia, USA. Wildlife Biology 2017. [Google Scholar] [CrossRef]
- Dique, D. S.; Thompson, J.; Preece, H. J.; de Villiers, D. L.; Carrick, F. N. Dispersal patterns in a regional koala population in south-east Queensland. Wildlife Research 2003, 30, 281–290. [Google Scholar] [CrossRef]
- Doligez, B.; Berthouly, A.; Doligez, D.; Tanner, M.; Saladin, V.; Bonfils, D.; Richner, H. Spatial scale of local breeding habitat quality and adjustment of breeding decisions. Ecology 2008, 89, 1436–1444. [Google Scholar] [CrossRef]
- Doligez, B.; Cadet, C.; Danchin, E.; Boulinier, T. When to use public information for breeding habitat selection? The role of environmental predictability and density-dependance. Animal Behaviour 2003, 66, 973–988. [Google Scholar] [CrossRef]
- Doligez, B.; Danchin, E.; Clobert, J.; Gustafsson, L. The use of conspecific reproductive success for breeding habitat selection in a non-colonial, hole-nesting species, the collared flycatcher. Journal of Animal Ecology 1999, 68, 1193–1206. [Google Scholar] [CrossRef]
- Doligez, B.; Pärt, T.; Danchin, E. Prospecting in the collared flycatcher : gathering public information for future breeding habitat selection? Animal Behaviour 2004, 67, 457–466. [Google Scholar] [CrossRef]
- Doolan, S. P.; Macdonald, D. W. Dispersal and extra-territorial prospecting by slender-tailed meerkats (Suricata suricatta) in the south-western Kalahari. Journal of Zoology 1996, 240, 59–73. [Google Scholar] [CrossRef]
- Ducros, D.; Morellet, N.; Patin, R.; Atmeh, K.; Debeffe, L.; Cargnelutti, B.; Chaval, Y.; Lourtet, B.; Coulon, A.; Hewison, A. J. M. Beyond dispersal versus philopatry? Alternative behavioural tactics of juvenile roe deer in a heterogeneous landscape. Oikos 2020, 129, 81–92. [Google Scholar] [CrossRef]
- Eikenaar, C.; Richardson, D. S.; Brouwer, L.; Komdeur, J. Sex biased natal dispersal in a closed, saturated population of Seychelles warblers Acrocephalus sechellensis. Journal of Avian Biology 2008, 39, 73–80. [Google Scholar] [CrossRef]
- Engler, M.; Krone, O. Movement patterns of the White-tailed Sea Eagle (Haliaeetus albicilla): post-fledging behaviour, natal dispersal onset and the role of the natal environment. Ibis 2022, 164, 188–201. [Google Scholar] [CrossRef]
- Fasciolo, A.; Delgado, M. D. M.; Cortés, G.; Soutullo, Á.; Penteriani, V. Limited prospecting behaviour of juvenile Eagle Owls Bubo bubo during natal dispersal: implications for conservation. Bird Study 2016, 63, 128–135. [Google Scholar] [CrossRef]
- Fijn, R. C.; Wolf, P.; Courtens, W.; Verstraete, H.; Stienen, E. W. M.; Iliszko, L.; Poot, M. J. M. Post-breeding prospecting trips of adult Sandwich Terns Thalasseus sandvicensis. Bird Study 2014, 61, 566–571. [Google Scholar] [CrossRef]
- Gaillard, J.-M.; Hebblewhite, M.; Loison, A.; Fuller, M.; Powell, R. A.; Basille, M.; Van Moorter, B. Habitat–performance relationships: finding the right metric at a given spatial scale. Philosophical Transactions of the Royal Society B 2010, 365, 2255–2265. [Google Scholar] [CrossRef]
- Gaughran, A.; MacWhite, T.; Mullen, E.; Maher, P.; Kelly, D. J.; Good, M.; Marples, N. M. Dispersal patterns in a medium-density Irish badger population: Implications for understanding the dynamics of tuberculosis transmission. Ecology and Evolution 2019, 9, 13142–13152. [Google Scholar] [CrossRef]
- Greenwood, P. J. Mating systems, philopatry and dispersal in birds and mammals. Animal Behaviour 1980, 28, 1140–1162. [Google Scholar] [CrossRef]
- Harrison, D. J.; Harrison, J. A.; O’Donoghue, M. Predispersal Movements of Coyote (Canis latrans) Pups in Eastern Maine. Journal of Mammalogy 1991, 72, 756–763. [Google Scholar] [CrossRef]
- Haughland, D. L.; Larsen, K. W. Exploration correlates with settlement: red squirrel dispersal in contrasting habitats. Journal of Animal Ecology 2004, 73, 1024–1034. [Google Scholar] [CrossRef]
- Honza, M.; Taborsky, B.; Taborsky, M.; Teuschl, Y.; Vogl, W.; Moksnes, A.; Røskaft, E. Behaviour of female common cuckoos, Cuculus canorus, in the vicinity of host nests before and during egg laying: a radiotelemetry study. Animal Behaviour 2002, 64, 861–868. [Google Scholar] [CrossRef]
- Jungwirth, A.; Walker, J.; Taborsky, M. Prospecting precedes dispersal and increases survival chances in cooperatively breeding cichlids. Animal Behaviour 2015, 106, 107–114. [Google Scholar] [CrossRef]
- Kelly, D. J.; Gaughran, A.; Mullen, E.; MacWhite, T.; Maher, P.; Good, M.; Marples, N. M. Extra Territorial Excursions by European badgers are not limited by age, sex or season. Scientific Reports 2020, 10, 9665. [Google Scholar] [CrossRef] [PubMed]
- Kesler, D. C.; Haig, S. M. Territoriality, Prospecting, and Dispersal in Cooperatively Breeding Micronesian Kingfishers (Todiramphus Cinnamominus Reichenbachii). The Auk 2007, 124, 381–395. [Google Scholar] [CrossRef]
- Kingma, S. A.; Bebbington, K.; Hammers, M.; Richardson, D. S.; Komdeur, J. Delayed dispersal and the costs and benefits of different routes to independent breeding in a cooperatively breeding bird. Evolution 2016, 70, 2595–2610. [Google Scholar] [CrossRef] [PubMed]
- Kivelä, S. M.; Seppänen, J.-T.; Ovaskainen, O.; Doligez, B.; Gustafsson, L.; Mönkkönen, M.; Forsman, J. T. The past and the present in decision-making: the use of conspecific and heterospecific cues in nest site selection. Ecology 2014, 95, 3428–3439. [Google Scholar] [CrossRef]
- Kloskowski, J. Win-stay/lose-switch, prospecting-based settlement strategy may not be adaptive under rapid environmental change. Scientific Reports 2021, 11, 570. [Google Scholar] [CrossRef]
- Kokko, H.; Lopéz-Sepulcre, A. From individual dispersal to species ranges: perspectives for a changing world. Science 2006, 313, 789–791. [Google Scholar] [CrossRef]
- Kolodzinski, J. J.; Tannenbaum, Lawrence V.; Muller, Lisa I.; Osborn, David A.; Adams, Kent A.; Conner, Mark C.; Ford, W. Mark; Miller, Karl V. Excursive Behaviors by Female White-tailed Deer during Estrus at Two Mid-Atlantic Sites. The American Midland Naturalist 2010, 163, 366–373. [Google Scholar] [CrossRef]
- Kralj, J.; Ponchon, A.; Oro, D.; Amadesi, B.; Arizaga, J.; Baccetti, N.; Boulinier, T.; Cecere, J. G.; Corcoran, R. M.; Corman, A.-M.; Enners, L.; Fleishman, A.; Garthe, S.; Grémillet, D.; Harding, A. M. A.; Igual, J. M.; Jurinović, L.; Kubetzki, U.; Lyons, D. E.; Orben, R.; Paredes, R.; Pirello, S.; Recorbet, B.; Shaffer, S. A.; Schwemmer, P.; Serra, L.; Spelt, A.; Tavecchia, G.; Tengeres, J.; Tome, D.; Williamson, C.; Windsor, S.; Young, H.; Zenatello, M.; Fijn, R. C. Active breeding seabirds prospect alternative breeding colonies. Oecologia 2023. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-Y.; Kokko, H. Sex-biased dispersal: a review of the theory. Biological Reviews 2019, 94, 721–736. [Google Scholar] [CrossRef] [PubMed]
- Mabry, K. E.; Stamps, J. A. Searching for a New Home: Decision Making by Dispersing Brush Mice. The American Naturalist 2008, 172, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Mancinelli, S.; Ciucci, P. Beyond home: Preliminary data on wolf extraterritorial forays and dispersal in Central Italy. Mammalian Biology 2018, 93, 51–55. [Google Scholar] [CrossRef]
- Mares, R.; Bateman, A. W.; English, S.; Clutton-Brock, T. H.; Young, A. J. Timing of predispersal prospecting is influenced by environmental, social and state-dependent factors in meerkats. Animal Behaviour 2014, 88, 185–193. [Google Scholar] [CrossRef]
- Martinović, M.; Galov, A.; Svetličić, I.; Tome, D.; Jurinović, L.; Ječmenica, B.; Basle, T.; Božič, L.; Kralj, J. Prospecting of breeding adult Common terns in an unstable environment. Ethology Ecology & Evolution 2019, 31, 457–468. [Google Scholar] [CrossRef]
- Mayer, M.; Zedrosser, A.; Rosell, F. Extra-territorial movements differ between territory holders and subordinates in a large, monogamous rodent. Scientific Reports 2017, 7, 15261. [Google Scholar] [CrossRef]
- McClintock, B. T.; Michelot, T. momentuHMM: R package for generalized hidden Markov models of animal movement. Methods in Ecology and Evolution 2018, 9, 1518–1530. [Google Scholar] [CrossRef]
- McNamara, J. M.; Barta, Z.; Klaassen, M.; Baue, S. Cues and the optimal timing of activities under environmental changes. Ecology Letters 2011, 14, 1183–1190. [Google Scholar] [CrossRef]
- Naguib, M.; Altenkamp, R.; Griessmann, B. Nightingales in space: song and extra-territorial forays of radio tagged song birds. Journal für Ornithologie 2001, 142, 306–312. [Google Scholar] [CrossRef]
- Nathan, R.; Getz, W. M.; Revilla, E.; Holyoak, M.; Kadmon, R.; Saltz, D.; Smouse, P. E. A movement ecology paradigm for unifying organismal movement research. Proceedings of the National Academy of Sciences 2008, 105, 19052–19059. [Google Scholar] [CrossRef] [PubMed]
- Nathan, R.; Monk, C. T.; Arlinghaus, R.; Adam, T.; Alós, J.; Assaf, M.; Baktoft, H.; Beardsworth, C. E.; Bertram, M. G.; Bijleveld, A. I.; Brodin, T.; Brooks, J. L.; Campos-Candela, A.; Cooke, S. J.; Gjelland, K. Ø.; Gupte, P. R.; Harel, R.; Hellström, G.; Jeltsch, F.; Killen, S. S.; Klefoth, T.; Langrock, R.; Lennox, R. J.; Lourie, E.; Madden, J. R.; Orchan, Y.; Pauwels, I. S.; Říha, M.; Roeleke, M.; Schlägel, U. E.; Shohami, D.; Signer, J.; Toledo, S.; Vilk, O.; Westrelin, S.; Whiteside, M. A.; Jarić, I. Big-data approaches lead to an increased understanding of the ecology of animal movement. Science 2022, 375, eabg1780. [Google Scholar] [CrossRef] [PubMed]
- Nocera, J. J.; Forbes, G. J.; Giraldeau, L. A. Inadvertent social information in breeding site selection of natal dispersing birds. Proceedings of the Royal Society B 2006, 273, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Northrup, J. M.; Vander Wal, E.; Bonar, M.; Fieberg, J.; Laforge, M. P.; Leclerc, M.; Prokopenko, C. M.; Gerber, B. D. Conceptual and methodological advances in habitat-selection modeling: guidelines for ecology and evolution. Ecological Applications 2022, 32, e02470. [Google Scholar] [CrossRef] [PubMed]
- Orians, G. H.; Wittenberger, J. F. Spatial and temporal scales in habitat selection. The American Naturalist 1991, 137, S29–S49. [Google Scholar] [CrossRef]
- Oro, D.; Bécares, J.; Bartumeus, F.; Arcos, J. M. High frequency of prospecting for informed dispersal and colonisation in a social species at large spatial scale. Oecologia 2021, 197, 395–409. [Google Scholar] [CrossRef] [PubMed]
- Pärt, T.; Doligez, B. Gathering public information for habitat selection: prospecting birds cue on parental activity. Proceedings of the Royal Society B 2003, 270, 1809–1813. [Google Scholar] [CrossRef]
- Patchett, R.; Styles, P.; King, J. Robins; Kirschel, A. N. G.; Cresswell, W. The potential function of post-fledging dispersal behavior in first breeding territory selection for males of a migratory bird. Current Zoology 2022, 68, 708–715. [Google Scholar] [CrossRef]
- Péron, C.; Grémillet, D. Tracking through life stages: adult, immature and juvenile autumn migration in a long-lived seabird. PLOS ONE 2013, 8, e72713. [Google Scholar] [CrossRef]
- Poessel, S. A.; Woodbridge, B.; Smith, B. W.; Murphy, R. K.; Bedrosian, B. E.; Bell, D. A.; Bittner, D.; Bloom, P. H.; Crandall, R. H.; Domenech, R.; Fisher, R. N.; Haggerty, P. K.; Slater, S. J.; Tracey, J. A.; Watson, J. W.; Katzner, T. E. Interpreting long-distance movements of non-migratory golden eagles: Prospecting and nomadism? Ecosphere 2022, 13, e4072. [Google Scholar] [CrossRef]
- Ponchon, A.; Aulert, C.; Le Guillou, G.; Gallien, F.; Péron, C.; Grémillet, D. Spatial overlaps of foraging and resting areas of black-legged kittiwakes breeding in the English Channel with existing marine protected areas. Marine Biology 2017a, 164, 119. [Google Scholar] [CrossRef]
- Ponchon, A.; Chambert, T.; Lobato, E.; Tveraa, T.; Grémillet, D.; Boulinier, T. Breeding failure induces large scale prospecting movements in the black-legged kittiwake. Journal of Experimental Marine Biology and Ecology 2015a, 473, 138–145. [Google Scholar] [CrossRef]
- Ponchon, A.; Garnier, R.; Grémillet, D.; Boulinier, T. Predicting population responses to environmental change: the importance of considering informed dispersal strategies in spatially structured population models. Diversity and Distributions 2015b, 21, 88–100. [Google Scholar] [CrossRef]
- Ponchon, A.; Grémillet, D.; Doligez, B.; Chambert, T.; Tveraa, T.; González-Solís, J.; Boulinier, T. Tracking prospecting movements involved in breeding habitat selection: insights, pitfalls and perspectives. Methods in Ecology and Evolution 2013, 4, 143–150. [Google Scholar] [CrossRef]
- Ponchon, A.; Iliszko, L.; Grémillet, D.; Tveraa, T.; Boulinier, T. Intense prospecting movements of failed breeders nesting in an unsuccessful breeding subcolony. Animal Behaviour 2017b, 124, 183–191. [Google Scholar] [CrossRef]
- Ponchon, A.; Scarpa, A.; Bocedi, G.; Palmer, S. C. F.; Travis, J. M. J. Prospecting and informed dispersal: Understanding and predicting their joint eco-evolutionary dynamics. Ecology and Evolution 2021, 11, 15289–15302. [Google Scholar] [CrossRef]
- Ponchon, A.; Travis, J. M. J. Informed dispersal based on prospecting impacts the rate and shape of range expansions. Ecography 2022, 2022, e06190. [Google Scholar] [CrossRef]
- Pöysä, H. Public information and conspecific nest parasitism in goldeneyes: targeting safe nests by parasites. Behavioral Ecology 2006, 17, 459–465. [Google Scholar] [CrossRef]
- Pöysä, H.; Milonoff, M.; Ruusila, V.; Virtanen, J. Nest-Site Selection in Relation to Habitat Edge: Experiments in the Common Goldeneye. Journal of Avian Biology 1999, 30, 79–84. [Google Scholar] [CrossRef]
- Reed, J. M.; Boulinier, T.; Danchin, E.; Oring, L. W. Informed dispersal: prospecting by birds for breeding sites. Current Ornithology 1999, 15, 189–259. [Google Scholar]
- Rémy, A.; Le Galliard, J.-F.; Gundersen, G.; Steen, H.; Andreassen, H. P. Effects of individual condition and habitat quality on natal dispersal behaviour in a small rodent. Journal of Animal Ecology 2011, 80, 929–937. [Google Scholar] [CrossRef] [PubMed]
- Riehl, C. Communal Calling And Prospecting By Black-Headed Trogons Trogon melanocephalus. The Wilson Journal of Ornithology 2008, 120, 248–255. [Google Scholar] [CrossRef]
- Roper, T. J.; Ostler, J. R.; Conradt, L. The process of dispersal in badgers Meles meles. Mammal Review 2003, 33, 314–318. [Google Scholar] [CrossRef]
- Roth, T.; Sprau, P.; Schmidt, R.; Naguib, M.; Amrhein, V. Sex-specific timing of mate searching and territory prospecting in the nightingale: nocturnal life of females. Proceeedings of the Royal Society B 2009, 276, 2045–2050. [Google Scholar] [CrossRef] [PubMed]
- Saldanha, S.; Cox, S. L.; Militão, T.; González-Solís, J. Animal behaviour on the move: the use of auxiliary information and semi-supervision to improve behavioural inferences from Hidden Markov Models applied to GPS tracking datasets. Movement Ecology 2023, 11, 41. [Google Scholar] [CrossRef]
- Samelius, G.; Andrén, H.; Liberg, O.; Linnell, J. D. C.; Odden, J.; Ahlqvist, P.; Segerström, P.; Sköld, K. Spatial and temporal variation in natal dispersal by Eurasian lynx in Scandinavia. Journal of Zoology 2012, 286, 120–130. [Google Scholar] [CrossRef]
- Sánchez-Tójar, A.; Winney, I.; Girndt, A.; Simons, M. J. P.; Nakagawa, S.; Burke, T.; Schroeder, J. Winter territory prospecting is associated with life-history stage but not activity in a passerine. Journal of Avian Biology 2017, 48, 407–416. [Google Scholar] [CrossRef]
- Scardamaglia, R. C.; Fiorini, V. D.; Kacelnik, A.; Reboreda, J. C. Planning host exploitation through prospecting visits by parasitic cowbirds. Behavioral Ecology and Sociobiology 2016, 71, 23. [Google Scholar] [CrossRef]
- Schuett, W.; Laaksonen, J.; Laaksonen, T. Prospecting at conspecific nests and exploration in a novel environment are associated with reproductive success in the jackdaw. Behavioral Ecology and Sociobiology 2012, 66, 1341–1350. [Google Scholar] [CrossRef]
- Selonen, V.; Hanski, I. K. Habitat exploration and use in dispersing juvenile flying squirrels. Journal of Animal Ecology 2006, 75, 1440–1449. [Google Scholar] [CrossRef]
- Selonen, V.; Hanski, I. K. Decision making in dispersing Siberian flying squirrels. Behavioral Ecology 2010, 21, 219–225. [Google Scholar] [CrossRef]
- Sicotte, P.; Andrew, J. M. Inter-group encounters and male incursions in Colobus vellerosus in Central Ghana. Behaviour 2004, 141, 533–553. [Google Scholar] [CrossRef]
- Soulsbury, C. D.; Iossa, G.; Baker, P. J.; White, P. C. L.; Harris, S. Behavioral and spatial analysis of extraterritorial movements in red foxes (Vulpes vulpes). Journal of Mammalogy 2011, 92, 190–199. [Google Scholar] [CrossRef]
- Spiegel, O.; Leu, S. T.; Bull, C. M.; Sih, A. What’s your move? Movement as a link between personality and spatial dynamics in animal populations. Ecology Letters 2017, 20, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Stamps, J. A.; Krishnan, V. V.; Reid, M. L. Search costs and habitat selection by dispersers. Ecology 2005, 86, 510–518. [Google Scholar] [CrossRef]
- Stutchbury, B. J. M.; Pitcher, T. E.; Norris, D. R.; Tuttle, E. M.; Gonser, R. A. Does male extra-territory foray effort affect fertilization success in hooded warblers Wilsonia citrina? Journal of Avian Biology 2005, 36, 471–477. [Google Scholar] [CrossRef]
- Teichroeb, J. A.; Wikberg, E. C.; Sicotte, P. Dispersal in male ursine colobus monkeys (Colobus vellerosus): influence of age, rank and contact with other groups on dispersal decisions. Behaviour 2011, 148, 765–793. [Google Scholar] [CrossRef]
- Therrien, J.-F.; Pinaud, D.; Gauthier, G.; Lecomte, N.; Bildstein, K. L.; Bety, J. Is pre-breeding prospecting behaviour affected by snow cover in the irruptive snowy owl? A test using state-space modelling and environmental data annotated via Movebank. Movement Ecology 2015, 3, 1. [Google Scholar] [CrossRef]
- Thomson, R. L.; Sirkiä, P. M.; Villers, A.; Laaksonen, T. Temporal peaks in social information: prospectors investigate conspecific nests after a simulated predator visit. Behavioral Ecology and Sociobiology 2013, 67, 905–911. [Google Scholar] [CrossRef]
- Trainor, A. M.; Walters, J. R.; Morris, W. F.; Sexton, J.; Moody, A. Empirical estimation of dispersal resistance surfaces: a case study with red-cockaded woodpeckers. Landscape Ecology 2013, 28, 755–767. [Google Scholar] [CrossRef]
- Urban, M. C.; Bocedi, G.; Hendry, A. P.; Mihoub, J.-B.; Pe’er, G.; Singer, A.; Bridle, J. R.; Crozier, L. G.; De Meester, L.; Godsoe, W.; Gonzalez, A.; Hellmann, J. J.; Holt, R. D.; Huth, A.; Johst, K.; Krug, C. B.; Leadley, P. W.; Palmer, S. C. F.; Pantel, J. H.; Schmitz, A.; Zollner, P. A.; Travis, J. M. J. Improving the forecast for biodiversity under climate change. Science 2016, 353. [Google Scholar] [CrossRef] [PubMed]
- Valone, T. J.; Templeton, J. J. Public information for the assessment of quality: a widespread social phenomenon. Philosophical Transactions of the Royal Society B 2002, 357, 1549–1557. [Google Scholar] [CrossRef] [PubMed]
- Vangen, K. M.; Persson, J.; Landa, A.; Andersen, R.; Segerström, P. Characteristics of dispersal in wolverines. Canadian Journal of Zoology 2001, 79, 1641–1649. [Google Scholar] [CrossRef]
- Veiga, J. P.; Polo, V.; Arenas, M.; Sánchez, S. Intruders in Nests of the Spotless Starling: Prospecting for Public Information or for Immediate Nesting Resources? Ethology 2012, 118, 917–924. [Google Scholar] [CrossRef]
- Votier, S.; Grecian, W.; Patrick, S.; Newton, J. Inter-colony movements, at-sea behaviour and foraging in an immature seabird: results from GPS-PPT tracking, radio-tracking and stable isotope analysis. Marine Biology 2011, 158, 355–362. [Google Scholar] [CrossRef]
- Ward, M. Habitat selection by dispersing yellow-headed blackbirds: evidence of prospecting and the use of public information. Oecologia 2005, 145, 650–657. [Google Scholar] [CrossRef]
- Ward, M. P.; Alessi, M.; Benson, T. J.; Chiavacci, S. J. The active nightlife of diurnal birds: extraterritorial forays and nocturnal activity patterns. Animal Behaviour 2014, 88, 175–184. [Google Scholar] [CrossRef]
- White, D. J.; Davies, H. B.; Agyapong, S.; Seegmiller, N. Nest prospecting brown-headed cowbirds “parasitize” social information when the value of personal information is lacking. Proceedings of the Royal Society B: Biological Sciences 2017, 284, 20171083. [Google Scholar] [CrossRef]
- Wiens, J. A. Population responses to patchy environments. Annual Review of Ecology, Evolution, and Systematics 1976, 7, 81–120. [Google Scholar] [CrossRef]
- Williams, D. A.; Rabenold, K. N. Male-biased dispersal, female philopatry, and routes to fitness in a social corvid. Journal of Animal Ecology 2005, 74, 150–159. [Google Scholar] [CrossRef]
- Wolfson, D. W.; Fieberg, J. R.; Andersen, D. E. Juvenile Sandhill Cranes exhibit wider ranging and more exploratory movements than adults during the breeding season. Ibis 2020, 162, 556–562. [Google Scholar] [CrossRef]
- Young, A. J.; Carlson, A. A.; Clutton-Brock, T. Trade-offs between extraterritorial prospecting and helping in a cooperative mammal. Animal Behaviour 2005, 70, 829–837. [Google Scholar] [CrossRef]
- Young, A. J.; Monfort, S. L. Stress and the costs of extra-territorial movement in a social carnivore. Biology Letters 2009, 5, 439–441. [Google Scholar] [CrossRef] [PubMed]
- Young, A. J.; Spong, G.; Clutton-Brock, T. Subordinate male meerkats prospect for extra-group paternity: alternative reproductive tactics in a cooperative mammal. Proceedings of the Royal Society B: Biological Sciences 2007, 274, 1603–1609. [Google Scholar] [CrossRef]
Figure 1.
(a) Percentage of the different taxa studied (b) Percentage of the different methods used (VHF=Very High Frequency; RFID=Radio Frequency IDentification; PTT=Platform Terminal Transmitter; Direct obs = direct observations) (c) number of publications per year and (d) main factors examined in the 124 publications reviewed.
Figure 1.
(a) Percentage of the different taxa studied (b) Percentage of the different methods used (VHF=Very High Frequency; RFID=Radio Frequency IDentification; PTT=Platform Terminal Transmitter; Direct obs = direct observations) (c) number of publications per year and (d) main factors examined in the 124 publications reviewed.
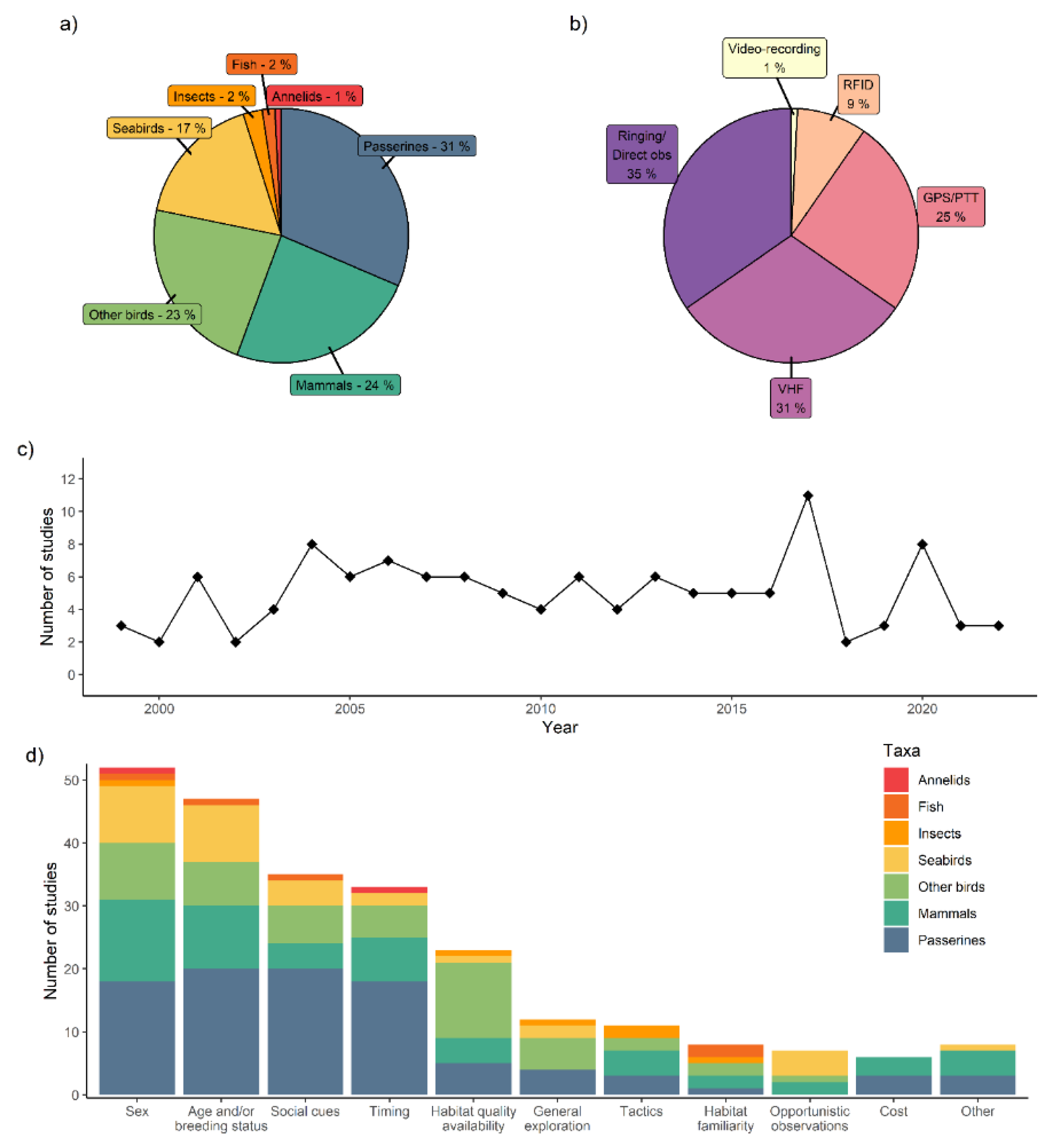
Figure 2.
Conceptual framework integrating the spatial, social and individual components of prospecting behaviour into the movement ecology framework (grey area) through the use of different methodologies: field observations and experiments for behaviour and tracking for movements. Figure created by Aurore Ponchon using icons from Microsoft Office Powerpoint LTSC Professionnel Plus 2021.
Figure 2.
Conceptual framework integrating the spatial, social and individual components of prospecting behaviour into the movement ecology framework (grey area) through the use of different methodologies: field observations and experiments for behaviour and tracking for movements. Figure created by Aurore Ponchon using icons from Microsoft Office Powerpoint LTSC Professionnel Plus 2021.
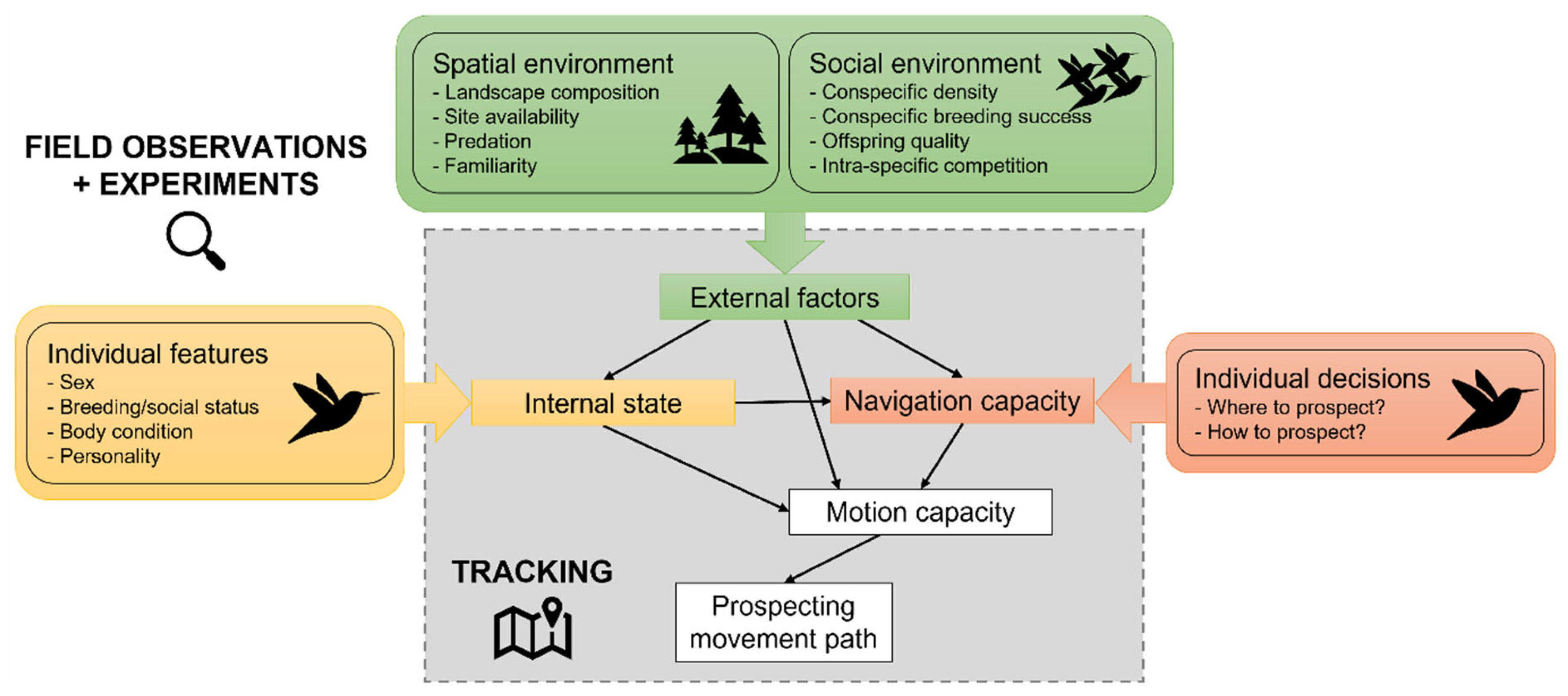

Table 1.
Species tracked with devices allowing the recording of continuous movements of individuals over time and associated maximal prospecting distance (GPS=Global Positioning System; UHF=Ultra High Frequency; VHF= Very High Frequency; GSM=Global System for Mobile communications; PTT=Platform Terminal Transmitter).
Table 1.
Species tracked with devices allowing the recording of continuous movements of individuals over time and associated maximal prospecting distance (GPS=Global Positioning System; UHF=Ultra High Frequency; VHF= Very High Frequency; GSM=Global System for Mobile communications; PTT=Platform Terminal Transmitter).
| Species | Scientific name | Taxon | Class of prospectors | Tracking device | Maximal prospecting distance (km) | Reference |
| Black-legged kittiwake | Rissa tridactyla | seabird | Failed breeders | GPS-UHF | 550 | (Boulinier et al. 2016) |
| Bonelli's eagle | Aquila fasciata | raptor | Juveniles | PTT | 435 | (Cadahía Lorenzo et al. 2009) |
| Audouin's gull | Larus audouinii | seabird | Failed and successful breeders | GPS + PTT | 360 | (Oro et al. 2021) |
| Golden eagle | Aquila chrysaetos | raptor | Juveniles and subadults | PTT | > 300 | (Poessel et al. 2022) |
| Black-legged kittiwake | Rissa tridactyla | seabird | Failed breeders | GPS-UHF | 220 | (Ponchon et al. 2017a) |
| White-tailed sea eagle | Haliaeetus albicilla | raptor | Juveniles | GPS-GSM | 200 | (Engler and Krone 2022) |
| Sandwich tern | Thalasseus sandvicensis | seabird | Failed breeders | GPS-UHF | 170 | (Fijn et al. 2014) |
| Black-browed albatross | Thalassarche melanophris | seabird | Immatures | GPS | 160 | (Campioni et al. 2017) |
| Wolf | Canis lupus | mammal | Adults | GPS | 147 | (Mancinelli and Ciucci 2018) |
| Northern gannet | Morus bassanus | seabird | Immatures | GPS-PTT+VHF | > 100 | (Votier et al. 2011) |
| Cory's shearwater | Calonectris diomedea | seabird | Immatures | PTT | > 100 | (Péron and Grémillet 2013) |
| European badger | Meles meles | mammal | Subadults (2-3 years-old) | GPS | > 100 | (Gaughran et al. 2019) |
| Black-legged kittiwake | Rissa tridatyla | seabird | Failed breeders | GPS | 40 | (Ponchon et al. 2015a) |
| Roe deer | Capreolus capreolus | mammal | Juveniles | GPS | 25 | (Debeffe et al. 2013) |
| Gray fox | Urocyon cinereoargenteus | mammal | Adults | GPS | 23.2 | (Deuel et al. 2017) |
| Common tern | Sterna hirundo | seabird | Successful and failed breeders | GPS-UHF | 19.5 | (Martinović et al. 2019) |
| Eurasian beaver | Castor fiber | mammal | Dominants and subordinates | GPS | 11.3 | (Mayer et al. 2017) |
| Californian spotted owl | Strix occidentalis occidentalis | raptor | Female breeders | GPS-UHF | > 10 | (Blakey et al. 2019) |
| Mallard, gadwall, cinnamon teal |
Anas platyrhynchos - Mareca strepera – Spatula cyanoptera |
waterfowl | Juveniles, immatures and adult females | GPS-GSM | 3 | (Casazza et al. 2020) |
| European badger | Meles meles | mammal | Juveniles and adults | GPS | 2 | (Kelly et al. 2020) |
| White-tailed deer | Odocoileus virginianus | mammal | Breeding females | GPS | < 2 | (Kolodzinski et al. 2010) |
Table 2.
Main factors driving prospecting extracted from the 124 empirical studies reviewed.
| Class of factors | Factors | Class of prospectors | Example of references |
| Individual factors | Sex | Males, females | (Selonen and Hanski 2010, Jungwirth et al. 2015, Blakey et al. 2019) |
| Age | Juveniles, immatures, subadults, adults | (Campioni et al. 2017, Wolfson et al. 2020) | |
| Breeding status | Failed or successful breeders, non-breeders (=floaters) | (Kingma et al. 2016, Ponchon et al. 2017b, Martinović et al. 2019) | |
| Dispersal status | Philopatric or dispersing individuals | (Debeffe et al. 2013, Oro et al. 2021) | |
| Social factors | Social status | Dominants, helpers, subordinates | (Kingma et al. 2016, Cram et al. 2018) |
| Inbreeding | Kin-related individuals | (Eikenaar et al. 2008, Kingma et al. 2016) | |
| Environmental factors | Habitat size/quality | Territory owners | (Haughland and Larsen 2004, Mayer et al. 2017) |
| Habitat familiarity | All | (Armstrong et al. 1997, Bruinzeel and van de Pol 2004) | |
| Territory quality | Territory owners | (Barve et al. 2020) | |
| Predation risk | All | (Thomson et al. 2013) |
Table 3.
Examples of personal and social cues used by prospecting species with different socialites throughout the year or during specific periods based on the 124 reviewed empirical studies.
Table 3.
Examples of personal and social cues used by prospecting species with different socialites throughout the year or during specific periods based on the 124 reviewed empirical studies.
| Type of information | Cues | Species | Availability over time | Examples of references |
| Personal information | Habitat physical structure | All | Permanent | (Therrien et al. 2015) |
| Presence/activity of predators | All | Permanent | (Vangen et al. 2001) | |
| Vacant breeding sites | Territorial, colonial | Breeding season | (Bruinzeel and van de Pol 2004, Veiga et al. 2012) | |
| Empty breeding sites | Territorial, colonial | Non-breeding season | (Pöysä 2006, Ciaglo et al. 2021) | |
| Individual own breeding status/performance | All | Permanent | (Ponchon et al. 2015a, 2017b) | |
| Social information | Conspecific presence/activity | All | Breeding season | (Williams and Rabenold 2005) |
| Conspecific breeding performance | Territorial, colonial | Breeding season | (Calabuig et al. 2010) | |
| Number of eggs in nests | Birds | Incubation period | (Pöysä 2006, Martinović et al. 2019) | |
| Number and quality of offspring | All | Offspring rearing | (Pärt and Doligez 2003, Ward 2005, Veiga et al. 2012) | |
| Host body condition | Parasitic birds | Laying season | (Scardamaglia et al. 2016) | |
| Group composition | Social or cooperative | Permanent | (Mayer et al. 2017, Barve et al. 2020) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



