Submitted:
23 November 2023
Posted:
24 November 2023
Read the latest preprint version here
Abstract
This study genetically identified the coat color in a small wild horses’ population and compared the correspondence with the visual coat color registrations. Individual DNA from 90 Sarcidano horses was used for the genetic assignment of the coat color, to explore the correspondence with the individual form containing phenotypical traits. By genotyping and sequencing analyses of the MC1R exon 1 and ASIP exon 3, e real image of the coat color distribution in this population has been obtained. Surprisingly, once we compared the genetic results with the individual forms reporting the phenotypic data for each subject, a certain degree of non-correspondence between the phenotypic and genetic data, in relation to coat color emerged. Genetic results showed a clear prevalence of the Chestnuts (n=58), a quite large number of Blacks (n=28) and a very small presence of Bays (n=4), whereas phenotypic distribution resulted in 38 Chestnuts, 40 Bays, only 2 Blacks, and 10 Greys (without the possibility of recognizing the true color they carried). Chestnut resulted a very representative coat color, while many horses visually identified as Bays, were genetically Blacks. This discrepancy suggests the importance of a sure coat color identification to ensure an adequate features registration, reliable also to predict the offspring’s coat color.
Keywords:
Sarcidano Horse pigmentation
; visual/genetic recording
; coat color distribution
; offspring’s coat color
1. Introduction
The Sarcidano Horse is an autochthonous semi-feral breed of small horses native to Sarcidano, a region in the hearth of Sardinia (Italy). It belongs to the 26 minor Italian breeds of limited diffusion collected in the Stud Book managed, since 2018, by ANAREAI, the national association for the local horse and donkey breeds [1]. These horses are locally known for their sturdy build and were traditionally used for rural work and riding. They live in conditions of geographical isolation, due to insularity and to the wild living environment, and are recognized for their endurance and adaptability to the local terrain. According to the breed standard, Sarcidano horses typically come only in basic coat colors, Chestnut, Bay, Black and sometimes Gray, without spotting or color dilution. Only two loci are involved in that hair coloration: Extension locus, coding melanocortin-1-receptor (MC1R) gene, and Agouti locus, coding the agouti-signaling-protein (ASIP) gene, that is MC1R gene antagonist. The correct functioning of the MC1R gene produces the MC1R protein, driving the eumelanin (black pigment) synthesis by melanocytes, producing Black coats [2,3]. Contrarily, alteration in the MC1R gene Exon 1, produced by a C to a T transition, leads to phaeomelanin (reddish pigment) synthesis, producing Chestnut coats [4]. The ASIP gene produces the ASIP protein, which has the power to antagonize MC1R gene functioning in the melanocytes located in the body but not in the extremities, thus producing Bay coats, resulting in reddish-brown body color with a black point coloration on the mane, tail, ear edges and lower legs [5]. Antagonism of MC1R by ASIP gene is efficient only in dominant/non-mutant variant of the MC1R gene, whereas when the receptor is already defective, because of the C>T mutation explained above, the antagonism cannot occur, still producing a Chestnut coat [2].
In a previous study on this breed, we focused on the genetic distribution of the coat color [6]. Just thanks to that investigation, carried out on 70 horses and aimed to contributing to the general knowledge of this breed, we became aware of a certain inconsistency in the color assigned thorough visual evaluation compared to that detected by the genetic investigation. Since the genetic color is the only one that can be transmitted to offspring, the correct assignment of the parental coat color is essential to ensure both correct individual identification and the prediction of foal color, as stated in other studies conducted on other local breeds [7,8,9]. In a wild population coat color provides camouflage and protection in natural environments, helping horses to blend into their surroundings and evade predators. Domestication and human selection introduced a variety of coat colors and patterns, that currently are typical of specific breeds [10]. Diluted or spotted coats, for example, allowed easier recognition of domestic from wild animals, and were therefore probably sought in the early stages of domestication [11]. Thus, the study of the coat color distribution could be also suggestive of possible crossing with domestic breeds, so contributing to the observation of the genetic isolation of local breeds. The correct coat color registration in individual documents avoids recognition errors, allows appropriate archiving of data, therefore granting to correctly assign relationships, to recognize family units and to predict the offspring’s coat color. Therefore, the aim of the present research was to compare visual and genetic coat color registration in a larger number of Sarcidano horses to develop an effective and low-cost system for correctly recording individual phenotypes.
2. Materials and Methods
2.1. Animals and Ethics statement
This study used blood samples provided by the veterinarians who captured Sarcidano horses at different times for periodic checks or extraordinary events, such as microchip implantation or the need to move animals to safer areas based on environmental conditions. Blood samples were collected from the horses under appropriate animal use approval by the Organism in charge for the Animal Welfare and Experimentation (OPBSA) of the University of Sassari (Protocol number: 2018UNISSMEV 0000177). More information on animals, capture methods and on blood and phenotype data collection is available on Cosso et al. manuscript [6]. No further ethics statement was needed for the present study. This study involved 90 of the approximately 120 Sarcidano Horses recorded in 2018, corresponding to the 75% of the entire registered population. The current number of registered Sarcidano horses is 109 heads, but since many microchipped animals are still not registered, it is difficult to determine the exact number of living horses.
2.2. Experimental design
Individual blood samples from 90 Sarcidano horses were subjected to DNA extraction using the NucleoSpin® Blood kit (Macherey-Nagel, Germany), following the provided protocol. Polymerase chain reaction (PCR) technique was carried out to amplify parts of MC1R and ASIP genes following methods reported by Cosso et al., 2022 [6].
2.2.1. Mutation detection
All the research procedures are those reported by Cosso et al. (2022) to which please refer for an accurate explanation of the used protocols and methods. Here we reported only some details useful for a better understanding of the present paper. Mutations in MC1R gene Exon 1 and ASIP gene Exon 3 are known to control base coat color set in horses. A pair of primers were used to amplify by polymerase chain reaction (PCR) a 320-base pair (bp) fragment, corresponding to part of the unique Exon 1 of the MC1R gene (Forward: 5’– CCT CGG GCT GAC CAC CAA CCA GAC GGG GCC – 3’ and Reverse: 5’– CCA TGG AGC CGC AGA TGA GCA CAT – 3’); another primers’ pair (Forward: 5’ – CTT TTG TCT CTC TTT GAA GCA TTG – 3’ and Reverse: 5’– GAG AAG TCC AAG GCC TAC CTT G – 3’) was designed to delimit a 102/91-bp polymorphic fragment, corresponding to the entire Exon 3 of the ASIP gene. Both pairs of primers and all the obtained results refer to the latest horse genome version EquCab3.0 (GCF_002863925.1).
MC1R gene polymorphisms were detected by restriction fragment length polymorphism (RFLP) technique, using TaqI endonuclease for the digestion of the obtained PCR products, thus allowed to recognize a C with a T transition within the amplicons’ nucleotide sequence.
Instead, the polymorphism within the ASIP gene Exon 3 was produced by an 11-bp deletion thanks to which it was possible to attribute the individual genotype directly through electrophoretic reading of the amplicons, producing 102- or 91-bp fragments, that identify the wild dominant or the mutant recessive allele, respectively. Results section shows how the combinations among MC1R and ASIP genotypes determined the real genetic coat colors. To confirm the assigned genotypes, so establishing the reliability of the used methods, 40 samples randomly chosen were sequenced in forward and reverse direction by a commercial service.
2.2.2. Statistical analysis
Allelic frequencies were calculated by direct counting of the detected genotypes in the MC1R and ASIP genes.
The observed genotypes and the horse coat color were compared using the χ2 test by means of the R statistical software (Version 4.3.2 R Core Team 2023 R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria; https://www.R-project.org/ accessed on 30 October 2023.
3. Results and Discussion
Genotyping results showed the genetic coat color distribution in the examined population. The sequencing of the randomly chosen samples confirmed the assigned genotypes, assuring that the used PCR/RFLP methods were reliable and could be therefore used in genetic coat color assignment in horses for faster and cheaper analysis compared to sequencing.
MC1R gene genotyping started by amplification of the 320-bp fragment, corresponding to position 36,979,377 – 36,979,697 of the latest horse genome version EquCab3.0 (GCF_002863925.1). Digestion by TaqI endonuclease detected the C>T transition (rs68458866), that allow to distinguish the wildtype dominant C from the mutant recessive T allele. The resulting genotype set was homozygous wild dominant C/C, heterozygous C/T and homozygous mutant recessive T/T. The main part of the analyzed Sarcidano horses carried the double recessive T/T genotype (58 heads, corresponding to 64% of the total studied population), 27 horses (30%) carried heterozygous genotype C/T, and only 5 horses carried the homozygous wild dominant genotype C/C, equivalent to 6% of the studied population. The resulting allele frequency was 79% for the T and 21% for the C allele (Table 1).
The causative mutation falls at position chr:3: 36,979,560 producing TCC or TTC as possible codons at position 83 of the protein chain (NCBI Reference Sequence: NP_001108006.1), resulting in a Serine being replaced with a Phenylalanine [6,12]. This single mutation in the proteins’ primary structure is expected to alter the alpha-helix structure, thus producing a defective functioning of the MC1 receptor, which becomes unable to be activated by the melanocyte stimulating hormone (MSH). This failure to function, leads to pheomelanin in place of eumelanin production within melanocytes. Thereby, a Black horse exhibits at least a wildtype C allele, while a Chestnut one carries only T/T genotype. Consequently, Agouti locus determines whether a horse carrying at least a C allele will be Bay (if it exhibits at least one recessive type 91 allele) or Black (if it carries homozygous 91/91 genotype); there is no effect of ASIP gene genotype on chestnut-based horses.
ASIP gene genotyping was conducted directly through amplification by PCR of the Exon 3, producing a 102/91-bp fragment. Indeed, a polymorphic 11-bp deletion (rs396813234) allowed to identify the three available genotype set based on the resulting fragments size. The entire (102-bp) nucleotide sequence corresponds to the dominant wildtype allele, here named as “102”, while the 11-bp deletion identifies the mutant recessive allele, here named as “91”. Consequently, the corresponding genotype set for the ASIP gene was 102/102 for the wildtype homozygous dominant, 102/91 for the heterozygous and 91/91 for the homozygous mutant recessive genotypes.
The ASIP gene genotype distribution exhibited only two horses carrying the wildtype dominant 102/102 genotype (corresponding to 2% of the studied population), while the main part of the population exhibited the recessive 91/91 genotype (62 horses, corresponding to 69% of the analyzed Sarcidano horses) and 26 subjects (29%) resulted heterozygous 102/91. Allele frequency resulted 83% for the recessive 91 allele and 17% for the wildtype 102 allele. Both the loci showed a very low frequency of the wild dominant allele and a very high rate of the mutant recessive type (Table 1).
From a genetic point of view more than half the analyzed population resulted Chestnut, well over a quarter were Blacks and a very small portion were Bays. Inconsistencies in visual coat color registration emerged when these results were matched with the phenotypically recorded data, producing an altered distribution of coat colors in the studied population. Indeed, from a phenotypic point of view, the 90 Sarcidano horses were registered as 38 Chestnuts, 40 Bays, only 2 Blacks, and 10 Greys. Grey horses were recorded simply as “grey”, without indication of the possible true color they carried (Figure 1).
The error rate (calculated as number of incorrectly classified color phenotype on the total number of genetically assigned coat colors) was 54%, including also the Grey horses. This large discrepancy between phenotypic and genetic data highlighted how visual observation can greatly alter the coat color assignment, leading to incorrect registration of the individual data.
Different genotype combinations were found for each coat color, as reported in Table 2.
These combinations produced some differences in the color shades varying from light to dark within the basic color set, that could be responsible for the inconsistencies in visual color recognition. Chestnuts resulted in the following combinations: T/T+91/91 (59% of the total chestnuts), T/T+102/91 (38%) and T/T+102/102 (3%). The most represent-ed genotype combination in the studied Sarcidano Horses resulted T/T+91/91, producing a liver Chestnut, 7 of which were visually identified and recorded as Bay. Of the 22 horses carrying T/T+102/91 combination, producing a phenotypic lighter-sorrel Chestnuts, 5 were visually registered as Bays. In Black horses too, different genotype combinations were found: 23 were C/T+91/91 (82% of the Blacks), 21 of which were visually registered as Dark Bays, while 5 were C/C+91/91, of which 4 were visually recognized as Bays. This important discrepancy in recognizing the black coat color has different possible explanations. The non-black areas of a Bay horse can range from a light brown to near-black, whereas Black horses can range from a sun-faded brown to jet black. This range of possible shades can make it difficult to pinpoint the exact coat color and create confusion when recording [13]. Much of the confusion arising because solid black is quite a rare coat color in most horse breeds, therefore many breed standards do not recognize this coat color. Consequently, a very dark horse is always registered as Dark Bay, and many people who fill out individual forms apply this rule to all breeds. However, the genetics of a Bay coat is different from that of a Black one, thus preventing the coat color from being correctly assigned.
In this study only 4 horses were found to be genetically Bays, while visually as many as 40 heads were registered with this coat. All the 4 Bays carried the heterozygous genotype at both the loci, C/T+102/91. The other possible allele combinations for a Bay coat color are C/C+102/102, C/C+102/91 and C/T+102/102, but none of them were found in the studied Sarcidano horses. All these results are summarized in Table 3.
The found inconsistencies in some visual vs genetic coat color recognition means that a subjective perception of shadings can alter the correct identification, so making visual evaluation not completely reliable, as the same horse could be identified differently by different evaluators. Instead, it is crucial to identify coat colors accurately and to correctly re-port them in the Stud Book, for legal and medical certification, and for the correct prediction of the inheritable coat colors, that can only be achieved with genetic investigation of the MC1R and ASIP loci genotypes.
Another source of confusion concerns Grey horses, as they are born Black, Chestnut or Bay but gradually lose color in their coat as they age [14]. This means that an adult Grey horse masks its true color and it is not clear what color its off-spring will inherit. However, since Grey locus is epistatic to the above base coat color genes, if a horse is grey, one can be assured that it has at least one Gray parent, but it is impossible to know what color its foals will be, without knowing what genetic color they carry [15]. The same can be said also for Chestnuts and Bays. In-deed, Chestnuts horses, although are surely carriers of the T/T genotype at the MC1R locus, do not externally show which genotype they have at the ASIP locus; therefore, they can generate offspring of any coat color base, depending on the partner’s genotype.
Even Bay horses do not display the genotype they carry at the ASIP locus and are consequently able to transmit different variants of ASIP alleles: a homozygous 102/102 horse will necessarily pass on the 102-allele to its offspring, while a heterozygous horse (102/91) will have a 50% chance of pass on the 102 allele.
Therefore, when the individual coat color is genetically identified, it is possible to know without any doubt, and already from birth, what type of foal each horse will be able to generate, in absence of other color modifiers, as it is in the Sarcidano Horse. From homozygous MC1R and ASIP loci parents (C/C+102/102) all offspring will be Bay. From C/C+102/91 parents, all offspring will be Dark based, but could be both Dark Bay and true Black. From C/T+102/102 parents, offspring will be Bay as they receive homozygous wild type ASIP alleles, diluting black body color, so that they can be visually appear more red or more black depending on coat color tone and different external and individual factors, such as age, environmental conditions, and nutritional status. Finally, from heterozygous C/T+102/91 parents, any color offspring are possible.
4. Conclusions
To conclude, this study contributes to the growing understanding of coat color genetics in the Sarcidano Horse and to lay out a reliable, easy, and low-cost method for the certain identification of the coat color in horses, avoiding incorrect recording of the data collected and uncertain heritable color prediction. Given the wild nature of the Sarcidano Horse, the visual coat color observation is not always an easy activity, since it occurs out-doors, under any weather condition, without guaranteeing optimal observation time, due to the capture requirements. Inconsistencies in the coat color registration are a common problem for many horse breeds, in which assignment only thorough observation can lead to incorrect attribution of lineage. This could have serious consequences for such a small population that could cause its weakening and even extinction, while the main goal should be to preserve all these ancient native populations. Although further initiatives to defend this important genetic resource would be desirable, at present only a few breeders and local authorities and passionate researchers are actively involved in increasing interest in this small population. Our goal is to plan new captures of Sarcidano horses for the near future to verify the exact number of animals currently living on the Sarcidano plateau. These investigations will also be useful to define any changes in the distribution of the colors of the population from 2018 to today and verify which subjects registered years ago are still alive.
Author Contributions
All authors have made substantial contributions to all the following: the conception and design of the study (MCM, VC, SL, GC), acquisition of data (NC, ESP), analysis and interpretation of data (GC, MCM, VC, SL), and drafting the article and revising it critically for important intellectual content (MCM, VC, SL, GC, NC, ESP). All the authors approved the final version to be submitted. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by grants from Sassari University research fund 2020 (FAR/2020).
Institutional Review Board Statement
All the procedures were reviewed and approved by the Organism in charge for the Animal Welfare and Experimentation (OPBSA) of the University of Sassari (Protocol number: 2018UNISSMEV 0000177).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We are grateful to the Laconi municipality for its availability in this research.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- ANAREAI. Available online: https://www.anareai.it/ (accessed on 15 September 2023).
- Neves, A.P.; Schwengber, E.B.; Albrecht, F.F.; Isola, J.V.; van der Linden, L.D.S. Beyond fifty shades: The genetics of horse colors. In Trends and Advances in Veterinary Genetics; IntechOpen: London, UK, 2017. [Google Scholar]
- Sponenberg, D.P.; Bellone, R. Equine color genetics, 4th ed., Hoboken, New Jersey, NJ, USA, 2017; John Wiley & Sons, ISBN: 978-1-119-13058-1.
- Shang, S.; Yu, Y.; Zhao, Y.; Dang, W.; Zhang, J.; Qin, X.; Irwin, D.M.; Qin Wang, Q.; Liu, F.; Wang, Z.; Zhang, S.; Wang, Z. Synergy between MC1R and ASIP for coat color in horses (Equus caballus). J. Anim. Sci. 2019, 97, 1578–1585. [Google Scholar] [CrossRef] [PubMed]
- Thiruvenkadan, A.K.; Kandasamy, N.; Panneerselvam, S. Coat colour inheritance in horses. Livestock Sci., 2008, 117, 109–129. [Google Scholar] [CrossRef]
- Cosso, G.; Carcangiu, V.; Luridiana, S.; Fiori, S.; Columbano, N.; Masala, G.; Careddu, G.M.; Sanna Passino, E.; Mura, M.C. Characterization of the Sarcidano Horse Coat Color Genes. Animals 2022, 12, 2677. [Google Scholar] [CrossRef] [PubMed]
- Mackowski, M; Wodas, L.; Brooks, S.A.; Cieslak, J. TBX3 and ASIP genotypes reveal discrepancies in officially recorded coat colors of Hucul horses. Animal 2019, 1811–1816. [Google Scholar] [CrossRef]
- Silva, I.L.S.; Junqueira, G.S.B.; Oliveira, C.A.A.; Costa, R.B.; DE Camargo, G.M.F. Inconsistencies in horse coat color registration: A case study. J. Equine Sci. 2020, 31, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Cieslak, J. , Brooks, S.A., Wodas, L., Mantaj, W., Borowska, A., Sliwowska, J.H., Ziarniak, K., Mackowski, M. Genetic Background of the Polish Primitive Horse (Konik) Coat Color Variation-New Insight into Dun Dilution Phenotypic Effect. J. Hered. 2021, 112, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Cieslak, M.; Reissmann, M.; Hofreiter, M.; Lydwig, A. Colours of domestication. Biol. Rev. 2011, 86, 885–899. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, A.; Pruvost, M.; Reissmann, M.; Benecke, N.; Brockmann, G.A.; Castaños, P.; Cieslak, M.; Lippold, S.; Llorente, L.; Malaspinas, A.S.; et al. Coat color variation at the beginning of horse domestication. Science, 2009, 324, 485. [Google Scholar] [CrossRef] [PubMed]
- Marklund, L.; Moller, M.J.; Sandberg, K.; Andersson, L. A missense mutation in the gene for melanocyte-stimulating hormone receptor (MC1R) is associated with the chestnut coat color in horses. Mamm. Genome 1996, 7, 895–899. [Google Scholar] [CrossRef] [PubMed]
- Rieder, S.; Taourit, S.; Mariat, D.; Langlois, B.; Guerin, G. Mutations in the agouti (ASIP), the extension (MC1R), and the brown (TYRP1) loci and their association to coat color phenotypes in horses (Equus caballus). Mamm. Genome 2001, 12, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Kavar, T.; Čeh, E.; Dovč, P. A simplified PCR-based method for detection of gray coat color allele in horse. Mol. Cell Probes. 2012, 256–258. [Google Scholar] [CrossRef]
- Rosengren Pielberg, G.; Golovko, A.; Sundström, E.; Curik, I.; Lennartsson, J.; Seltenhammer, M.H.; Druml, T.; Binns, M.; Fitzsimmons, C.; Lindgren, G.; et al. A cis-acting regulatory mutation causes premature hair graying and susceptibility to melanoma in the horse. Nat. Genet. 2008, 40, 1004–1009. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Differences between genetic and visual coat color assignment in the 90 studied Sarcidano horses. Grey horses are present only as phenotypic coat color as they are carrier of a basic genetic color, which is highlighted by genetic analysis.
Figure 1.
Differences between genetic and visual coat color assignment in the 90 studied Sarcidano horses. Grey horses are present only as phenotypic coat color as they are carrier of a basic genetic color, which is highlighted by genetic analysis.
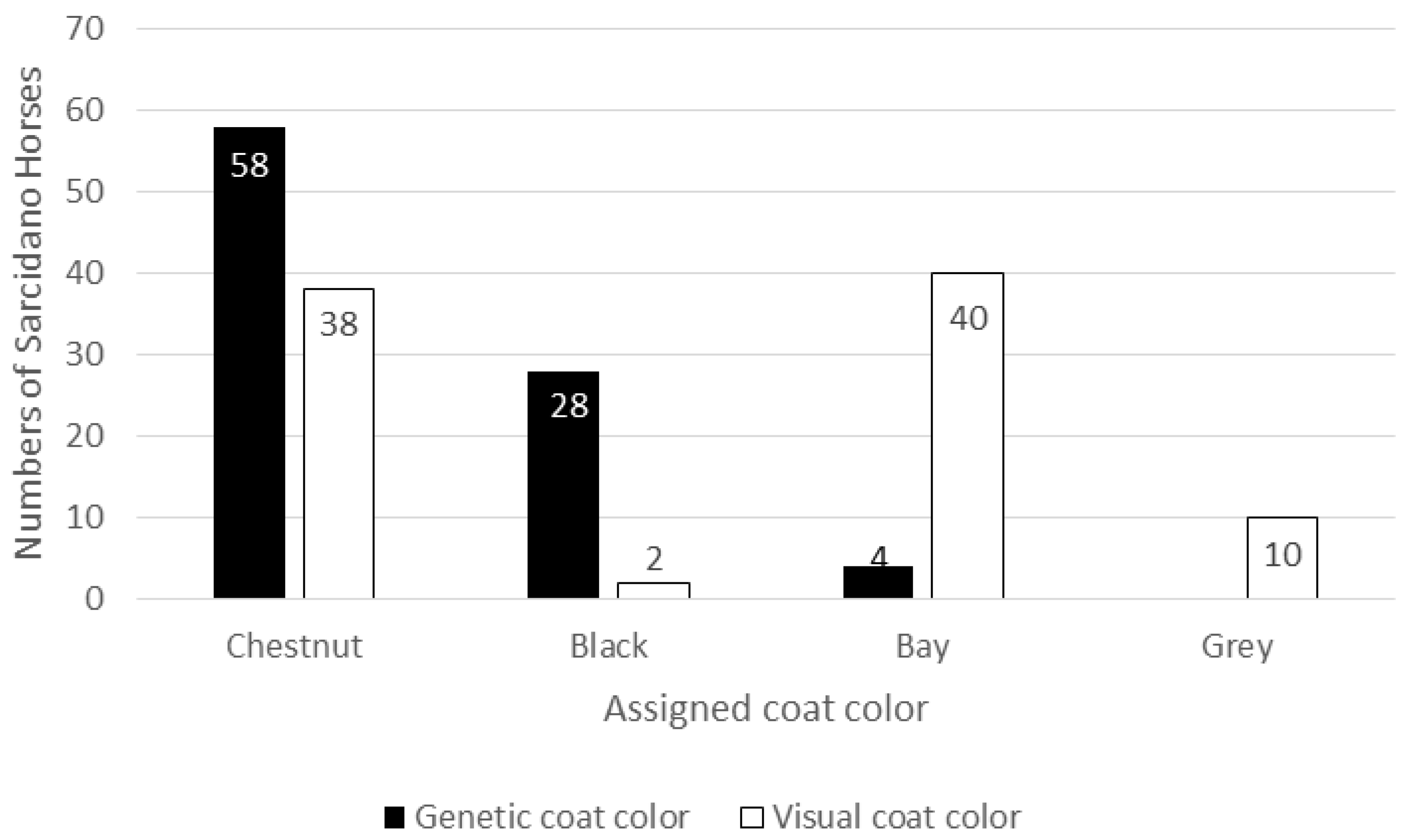

Table 1.
Genotypes and alleles distribution of the MC1R and ASIP genes polymorphisms according to the genetic coat color assignment in the 90 genotyped Sarcidano Horses.
Table 1.
Genotypes and alleles distribution of the MC1R and ASIP genes polymorphisms according to the genetic coat color assignment in the 90 genotyped Sarcidano Horses.
| MC1R | ASIP | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Genotypes | Alleles | Genotypes | Alleles | ||||||||
| C/C | C/T | T/T | C | T | 102/102 | 102/91 | 91/91 | 102 | 91 | ||
| Coat color | Black | 5 | 23 | 0 | 33 | 23 | 0 | 0 | 28 | 0 | 56 |
| Bay | 0 | 4 | 0 | 4 | 4 | 0 | 4 | 0 | 4 | 4 | |
| Chestnut | 0 | 0 | 58 | 0 | 116 | 2 | 22 | 34 | 26 | 90 | |
Table 2.
Combined distribution of MC1R and ASIP loci genotypes and base coat color phenotypes of the 90 genotyped Sarcidano Horse.
Table 2.
Combined distribution of MC1R and ASIP loci genotypes and base coat color phenotypes of the 90 genotyped Sarcidano Horse.
| MC1R genotype | ||||
|---|---|---|---|---|
| C/C | C/T | T/T | ||
| ASIPgenotype | 102/102 | 0 Bay | 0 Bay | 2 Chestnut |
| 102/91 | 0 Bay | 4 Bay | 22 Chestnut * | |
| 91/91 | 5 Black | 23 Black** | 34 Chestnut *** | |
* Three of which were phenotypically Grey; ** One of which was phenotypically Grey; 3*** Six of which were phenotypically Grey.
Table 3.
Cumulative combined distribution of visual and genetic coat color recorded in the 90 studied Sarcidano horses expressed as a percentage.
Table 3.
Cumulative combined distribution of visual and genetic coat color recorded in the 90 studied Sarcidano horses expressed as a percentage.
| Visual Coat Color (VCC) | ||||||
|---|---|---|---|---|---|---|
| Bay | Grey | Black | Chestnut | Total GCC | ||
| Genetic Coat Color (GCC) | Bay | 0.03 | 0.00 | 0.00 | 0.01 | 0.04 |
| Black | 0.28 | 0.00 | 0.02 | 0.01 | 0.31 | |
| Chestnut | 0.13 | 0.11 | 0.00 | 0.40 | 0.64 | |
| Total VCC | 0.44 | 0.11 | 0.02 | 0.42 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions, or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



