
Submitted:
20 November 2023
Posted:
24 November 2023
You are already at the latest version
Abstract
Many bacteria have the ability to survive in challenging environments; however, they cannot grow on standard culture media, a phenomenon known as the Viable but Non-Culturable (VBNC) state. Bacteria commonly go into the VBNC state under nutrient-poor environments or under stressful conditions. This review explores the concept of the VBNC state, providing insights into the beneficial bacteria known to employ this strategy. The investigation covers different chemical and physical factors that can induce the latency state, cell features, and gene expression observed in cells in the VBNC state. The revision also covered the significance and applications of beneficial bacteria, methods for evaluating bacterial viability, the ability of bacteria to persist in environments associated with higher organisms, and the factors that facilitate the return to the culturable state. Knowledge about beneficial bacteria capable of entering the VBNC state remains limited, however, beneficial bacteria in this state could face adverse environmental conditions, and return to culturable state when conditions become suitable and continue to exert their beneficial effects. Likewise, this unique feature positions them as potential candidates for healthcare applications, such as the use of probiotic bacteria to enhance human health, applications in industrial microbiology for the production of prebiotics, functional foods, and in the beer and wine industry. Moreover, their use in formulations to increase crop yield and for bacterial bioremediation offers an alternative pathway to harness their beneficial attributes.
Keywords:
VBNC state
; stress
; beneficial bacteria
; rhizosphere
; latency
; survival
1. Introduction
Bacteria in VBNC state exhibit a remarkable phenomenon: they are unable to grow and form colonies on conventional culture media, yet they remain alive and able to restart their metabolic activity [1]. Cells in this status typically display reduced levels of metabolic activity, and undergo significant metabolic alterations, such as: reduction of nutrient transport, respiration rate, and macromolecular synthesis [2]. However, a feature that distinguishes VNBC state is the continuous gene expression within these cells [3].
A typical response of cells entering into the VBNC state is shown in Figure 1, under stressful conditions caused by desiccation in presence and absence of a cytoprotective agent such as trehalose. In presence of trehalose, bacterial cells maintain high avoiding entry into the VBNC state. Conversely, in the absence of the protector a decrease in the number of CFUs/mL is observed under the same environmental stress. After 9 DABD (days after the beginning of desiccation), the cells entered into the VBNC state and remained so until 18 DABD; which was confirmed using several methodologies [4]. Interestingly, after prolonged rehydration or rapid rehydration in presence of plant exudates these bacteria reached high numbers indicating that they had returned to the culturable state [4].
1.1. Conditions that induce VBNC state
Bacterial cells commonly respond to stressful conditions by losing their ability to form colonies in standard culture media, although cells can remain viable for long periods of time. Cells may enter into the VBNC state in response to natural stresses, such as starvation, extreme temperature, high osmotic or oxygen concentrations, or exposure to white light [1]. Additional stressors that may lead to this status are listed in Table 1. In general, extreme environmental conditions could be lethal unless they adopt a latency status, for example, it is known that one of the most critical factors that plays a crucial role on bacterial survival is the availability of nutrients in the surrounding environment. When bacterial cells are subject to nutrient starvation conditions, they may reduce their size and become more resistant to adverse environmental conditions, alternatively, they could induce a latency estate, forming viable but non-culturable cells or resistance structures such as spores [3]. However, spore-forming bacteria are typically not classified within the VBNC state literature [3]. It is interesting that not only environmental stress can induce the VBNC state, various processes and bactericidal substances are able to induce this state as well. For example, milk pasteurization [5], wastewater chlorination [2], and use of food preservatives such as potassium sorbate and sodium benzoate [6] have been documented as inducers of the VBNC state.
2. Bacterial species entering the VBNC state
A substantial portion of bacterial species known to enter the VBNC includes human pathogens such as: Campylobacter spp. Escherichia coli (EHEC strains), Francisella tularensis, Helicobacter pylori, Legionella pneumophila, Listeria monocytogenes, Mycobacterium tuberculosis, Pseudomonas aeruginosa, several species of Salmonella spp., Shigella spp., and numerous pathogens from the genus Vibrio sp., being Vibrio vulnificus, one of the most studied in terms of the VBNC [2]. The medical significance of this phenomenon is multifaceted [17]. For example, it is suggested that the latent phase of Mycobacterium tuberculosis infections is caused by bacteria in the VBNC state [18,19] and the recurrence of disease several years later may be linked to their return to the culturable state within the host [20].
The list of pathogenic bacteria that can adopt the VBNC state as a survival strategy includes not only pathogen affecting humans but also animals such as Photobacterium damselae infecting fish [21], V. vulnificus eel pathogen [22] and V. shiloi that cause bleaching of corals [23,24]. Several plant pathogens have been described, for example, Ralstonia solanacearum in tomato plants [25], Xanthomonas axonopodis colonizing grapefruit plants [26], Erwinia amylovora infecting ripe apples [27], Pseudomonas syringae in tomato, cereals, almond, cherry, plum plants, Acidovorax citrulli infecting a wide variety of Cucurbitaceae causing bacterial spot disease (Bacterial Fruit Blotch) [28] and Agrobacterium tumefaciens which cause tumors in different dicotyledons.
Since the publication of Xu´s et al. study [7] more than 30 years ago, a substantial body of research around the world has focused on documenting the occurrence of the VBNC state in different bacterial species [2,4,12,17,29,30,31,32,33,34,35,36,37].
To date, approximately 101 bacterial species spanning 50 different genera have been reported to exhibit the VBNC phenomenon [2,29,36,38]. In the present review we analyze the beneficial bacterial species where the VBNC status has been reported and the importance of this within their potential in areas such as biotechnology, agriculture, food industry and health.
3. The VBNC state in beneficial bacteria
Beneficial bacteria play a crucial role in maintaining life on our planet. Some realize nitrogen fixation [39], mineral solubilization [40] greenhouse gas consumption, and are thus viewed as gatekeepers preventing excessive methane emissions from escaping the atmosphere [41]. Due to the several processes in which beneficial bacteria participate, they have been used to increase the crop production [42]. Some beneficial properties of these bacteria include plant growth promotion [43,44], control or inhibition of the activity of plant pathogens [45,46], improvements soil structure, bioaccumulation or microbial leaching of inorganics [47] bioremediation of xenobiotic compounds [48,49], or the production of compounds of industrial interest [50,51].
Certain bacteria that interact with their host establish mutually beneficial relationships. Probiotics for instance, interact with humans to improve overall health. They can eliminate or remove pathogens [52], reinforce the epithelial barrier, induce migration of fibroblasts and epithelial cells [53]. In the immune system probiotics are related to modulation and activation of intraepithelial lymphocytes, natural killer cells, and macrophages through induced production of cytokines [54].
On the other hand, there are beneficial bacteria with industrial applications that favor the production of certain foods, prebiotics and beverages. In the brewing and wine industry, bacteria play a crucial role in the fermentation process. Lactic acid bacteria (LAB) and yeast are instrumental in this context. LAB catalyze the conversion of dicarboxylic malic acid into monocarboxylic lactic acid and carbon dioxide (malolactic fermentation MLF) and yeast convert sugars into alcohol (alcoholic fermentation) [55]. During malolactic fermentation by LAB, no free intermediary products are formed achieving a more palatable wine by reducing the tart taste of malic acid. Additionally, malolactic fermentation reduces the amount of residual nutrients available to support microbial growth, enhances wine aroma, improves microbial stability and reduces the acidity of wine making the wine more stable before being bottled [56].
Despite the importance and benefits of bacteria at different levels, unfortunately, the knowledge about bacteria in the VBNC remains limited. The table 2 show the beneficial bacteria so far reported to enter viable non-culturable state under different conditions. This table displays a variety of conditions that can induce the VBNC state. Factors such as temperature, solute concentration, nutrient limitation, pH, desiccation, and industrial processes are determinants for the formation of VBNC cells. The groups of bacteria with more beneficial species capable to enter VBNC state are Alpha-proteobacteria and Gamma-proteobacteria with nine species each one. The groups with only three species that enter VBNC sate are Actinobacteria and Firmicutes. In the case of Betaproteobacteria only one species has been reported to enter the VBNC state.

Table 2.
Main species of beneficial bacteria that enter the Viable non-cultivable state.
| Group of bacteria | Species | Conditions that induce VBNC state | References | |
|---|---|---|---|---|
| Proteobecteria | Alfaproteobacteria | Acetobacter aceti | Treatment with SO2 at a concentration of 30 and 50 mg/L | [57] |
| Acetobacter pasteurianus | High acid stress during fermentation | [58] | ||
| Methylosinus sporium | Freeze drying and Cryopreservation (liquid nitrogen) |
[36,41] | ||
| Methylosinus trichosporium | ||||
| Methylocystis hirsuta | ||||
| Methylocystis parvus | ||||
| Methylocella tundrae | ||||
| Rhizobium leguminosarum | Cupric sulfate to a concentration of 60 ppm | [59] | ||
| Sinorhizobium meliloti | Incubating at 25°C in tap water from The University of North Carolina at Charlote and tap water from The University of Maryland Biotechnology Institute, Center of Marine Biotechnology. Incubating under anoxic conditions in liquid microcosms Incubation in nitrocellulose filters at a relative humidity of 22% for three days at 20°C in the dark. |
[12,60,61] | ||
| Betaproteobacteria | Cupriavidus metallidurans | Incubation into the artificial soil at 30 °C for 12 days, without any C source or H2O | [60] | |
| Gamaproteobacteria | Methylomonas methanica | Lyophilization and Cryopreservation (liquid nitrogen) |
[36,41] | |
| Methylosarcina fibrata | ||||
| Methylocaldum gracile | ||||
| Methylomicrobium alcaliphilum | ||||
| Methylococcus capsulatus | ||||
| Microbulbifer aggregans | Incubation in modified artificial seawater (ASW) by 4 h at 30°C | [61] | ||
| Pseudomonas fluorescens | Incubation in saline solution (NaCl 0.9% w/v) at 37°C | [62] | ||
| Pseudomonas putida KT2440 | Desiccation at 30°C and 50% relative humidity | [4] | ||
| Vibrio fischeri | Incubation at 22°C in nutrient-limited artificial seawater (ASW) |
[63] | ||
| Terrabacteria | Actinobacteria | Bifidobacterium animalis subsp. lactis | Storage in fermented foods Refrigerated storage of butter for 4 weeks Microcapsules with full-fat goat milk and inulin-type fructans |
[64,65,66] |
| Bifidobacterium longum | Storage in fermented foods | [64] | ||
|
Arthrobacter albidus. reclassified as Sinomonas albida |
absence of proteins Rpf (resuscitation promoting factor) in the culture medium |
[65,66] | ||
| Firmicutes | Bacillus coagulans | Incubation at pH 2 for 24 hours and subsequent incubation at 140°C for 5 min | [67] | |
| Lactobacillus plantarum | Treatment for 30 min at 100 °C or with 1 mol/L HCl Incubation in beer at 0°C temperature |
[31] | ||
| Oenococcus oeni | Sulfur dioxide and histidine decarboxylase activity in wines | [57] | ||
This review analyzed the phylogenetic relationship between different beneficial bacterial species reported to enter the VBNC state (Figure 2). The sequences were analyzed by comparing them with the National Center for Biotechnology Information (NCBI) database. To evaluate viable but unculturable strains, phylogenetic trees were constructed by the Neighbour-joining method [68] using ClustalX, Bioedit and Mega 4 software. A bootstrap confidence analysis was applied on 1000 replicates to determine the reliability of the topology obtained [69]. The phylogenetic tree show that these bacteria are very diverse. The furthest phylogenetic group corresponds to Methylosinus, Methylocystis and Methylocella. The genus Rhizobium, Sinorhizobium and Acetobacter are groups that are phylogenetically closer to each other. Methylococcus, Methylocaldum, Methylomicrobium, Methylotuvimicrobium, Methylosarcina and Methylomonas are phylogenetically close with Vibrio, Microbulbifer, and Pseudomonas. Another group that can be identified is formed by Arthrobacter, Bifidobacterium, Bacillus, Lactobacillus, and Oenococcus, with some strains being phylogenetically closely related and others very distantly species. When analyzing the phylogenetic relationships, it can be observed that viable non-cultivable state is not exclusive to any taxonomic group or group of species, and it is a widely distributed strategy in phylogenetically close and distant species.
The beneficial species in which the viable non-culturable state has been reported are described below, highlighting their main applications in different areas such as biotechnology, agro-biotechnology, health and industrial applications.
Within the group of alpha-proteobacteria that enter the VBNC state are organisms belonging to genus: Acetobacter, Methylosinus, Methylocistis, Methylocella, Rhizobium and Sinorhizobium (Table 2).
Acetobacter aceti and Acetobacter pasteurianus have a great relevance in vinegar production, since can transform ethanol into acetic acid through oxidative fermentation [70]. The VBNC state in A. aceti has been documented in wine production [57] and A. pasteurianus enters to VBNC state under high acid stress generated during the fermentation process [58]. Studies are still lacking that could support the idea that A. aceti and A. pasteurianus under a non-culturable state can follow the fermentation process, which would help increase production. The ability of these species to persist under adverse conditions represents a challenge to develop novel ways to improve industrial yields, enhancing wine quality and driving vinegar production on a larger scale.
The methylotrophic bacteria Methylosinus sporium, Methylosinus trichosporium, Methylocystis hirsuta, Methylocystis parvus and Methylocella tundrae are methanotrophic microorganisms also called methane-oxidizing bacteria (MOB) capable of generating energy through the oxidation of methane gas [71]. MOB have different biotechnological applications mainly, biological mitigation of methane greenhouse gas, production of high-value products from methane, and bioremediation of pollutants [72,73]. Because of ecological importance of these microorganisms, it is important understand how different conditions can affect methane consumption. One of the conditions that induces the VBNC state in MOB is freeze-drying and cryopreservation (Table 2). This represents a challenge to researchers, because culturable cells are needed whenever beneficial application. Most studies focus on how to avoid loss of cultivability, but is likely that MOB can return to culturable state by being in an environment with methane gas. This have observed with different pathogen species of bacteria that, when find in favorable conditions, leave from the non-culturable state [38]. Research with this group of bacteria in the VBNC state is scarce, which represents a challenge to understand this survival strategy. In the future, this could stimulate these organisms to consume methane, aiding in the degradation of pollutants and production of high-value biomass.
Other species of alphaproteobacteria are Sinorhizobium meliloti and Rhizobium leguminosarum found mainly in the soil, that have acquired, by horizontal gene transfer, the ability to associate in symbiosis with leguminous plant roots. The association of rhizobia and legumes occurs through a complex signaling process that generates nodules, organs specialized in the fixation of atmospheric nitrogen [74]. Biological nitrogen fixation is a vital process in agriculture, allowing the production of nitrogen through legume-rhizobium symbiosis that contributes to increased nitrogen levels in the soil, resulting in increased plant growth [75]. In addition to the ability to fix nitrogen, R. leguminosarum isolated from fava bean root nodules can generate induced systemic response against infection by Bean yellow mosaic virus [76]. The VBNC state in this bacterium is related with presence of cupric sulfate (Table 2). For S. meliloti the factors induce VBNC state are temperatures of 20 °C to 25 °C, incubation under anoxic conditions and incubation in nitrocellulose filters at low relative humidity (Table 2). It is interesting that under these conditions the bacteria can persist. This could explain their survival capacity under this types stress in environment. The importance of rhizobia for agriculture represents a major challenge to learn more about their beneficial functions in VBNC state, because it is unknown whether when bacteria are under non-culturable state they can continue performing nitrogen fixation, generate induced systemic response or even develop nodules.
In the group of Betaproteobacteria, the only beneficial bacteria where the VBNC state has been documented is Cupriavidus metallidurans. This bacterium is a metallophilic found in environments containing high concentrations of heavy metal, industrial wastes rich in toxic heavy metals, often mixed with recalcitrant organic compounds and hydrocarbons [77]. It is an ideal bacterium for bioaugmentation purposes in environmental applications due to strong resistance to environmental stress factors and its adaptation capacity [78]. Another peculiar application is its ability to synthesize gold of 24-caratgold in one week from gold chloride [79]. These observations suggest that bacteria actively contribute to forming gold grains in surface environments [80]. With this bacterium, the conditions that can induce its loss of cultivability have been studied, but also the conditions that can return to cultivable state. The addition of water and gluconate being sufficient for C. metallidurans to be culturable in 24 hours [60]. Due to its interesting applications and ability to persist in toxic environments, it is an excellent model for studying the mechanisms for coping with heavy metal stress. This unique ability to metabolize toxic substances and enter the VBNC state without water and carbon sources could help to understand how the origin of life occurred.
One of the groups with the most beneficial species in which the VBNC status has been demonstrated is Gammaproteobacteria. Methylomonas methanica, Methylosarcina fibrate, Methylocaldum gracile, Methylomicrobium alcaliphilum and Methylococcus capsulatus are microorganism methylotrophs. This type of bacteria uses methanol and methane as only carbon source [81]. Methylotrophs can habit in soil, water and plants, powering the carbon cycle [82]. When M. fibrate is co-inoculated with species of Methylomonas and Cupriavidus taiwanensis LMG 19424, its growth is highly stimulated [83]. Also, methane oxidation is elevated when methanotrophs interact with algae and moss [84]. Cryopreservation has been one method that induces the VBNC state in methanotrophs (Table 2). The study of VBNC state in methanotrophs is essential due to its role in the carbon cycle and therefore in the planet's life. It is relevant to consider that interaction with others microorganisms or whit plants could be an alternative for these microorganisms to return to the cultivable state, being able to continue exerting their beneficial effects.
Microbulbifer aggregans, other Gammaproteobacteria is a halophilic, Gram-negative bacterium isolated from sediment in the Matang mangrove forest, Malaysia [85]; its importance is centered in its capability to reduce sulfur by the presence of genes involved in this process [61]. Sulfur is an essential element for life, is present in amino acid, proteins, enzymes, vitamins and other biological molecules [86]. M. aggregans is a little explored bacterium, recently, it was observed that non-culturable cells exhibit a change in cell shape from rod to coccus, and the genes responsible for sulfate reduction are upregulated in the VBNC state [61]. These findings demonstrate its importance in the environment where it lives, because effective reduction of sulfur in non-cultivable state may occur in response to changes in the sulfur concentration in the environment where it is found, playing a relevant role in the sulfur cycle in marine environments.
Within the Gammaproteobacteria there is also the genus Pseudomonas. The species of this genus have a great capacity to use different nutrients as carbon source, which explains their ubiquity. Their enzymatic activity makes them an important group of microorganisms responsible for the aerobic degradation of many compounds in different ecosystems [87]. Some species Pseudomonas plant associated promote plant growth by suppressing pathogenic microorganisms, synthesizing growth stimulating plant hormones and promoting increased plant disease resistance [88]. Pseudomonas fluorescens and Pseudomonas putida KT2440 are bacteria able to colonize plants' root and promote their growth. Some strains of P. fluorescens have been shown to degrade a variety of organic compounds, thus important in bioremediation [89]. This bacterium is used for biocontrol to protect plants against soilborne fungal pathogens. One mechanism for the biocontrol by P. fluorescens is the ability to produce antibiotics [90]. Also stimulates the systemic resistance induced (ISR) [91] and production of volatile compounds [92].
On other hand P. putida KT2440 is a bacterium capable of using different aromatic compounds as a carbon source [93], metabolizing xenobiotic compounds, it can colonize the roots of plants such as corn, wheat, strawberry, sugarcane, sugar and spinach [94] and is capable of promoting the growth and health of the plants [95,96]. P. putida KT2440 has been used as part of consortia bacteria formulations to enhance plant growth [96,97].
The potential of P. putida KT2440 and P. fluorescens to exert its beneficial effects can be affected by their exposure to different stress that induce the VBNC state, such as saline environments for P. fluorescens and desiccation for P. putida KT2440 (Table 2). However, it has been shown that plant-bacteria interaction is a mechanism that allows these bacterium return to the cultivable state. Therefore, its use in formulation of stable bacterial inoculants that can stimulate plant growth after rehydration [4], or the development of formulations to perform biological control [34] could be an alternative to reduce the overuse of nitrogen fertilizers and pesticides, decreasing the damage caused by those chemical products [98].
Vibrio fischeri, also belonging to the Gammaproteobacteria, is a luminous marine bacterium that lives free or in symbiosis with different species of fish and squid [99]. The most studied interaction in this microorganism is the symbiosis with the Hawaiian squid, Euprymna scolopes, inducing bioluminescence that the squid uses to avoid predation during nocturnal activity [100]. In the squid-vibrio symbiosis, bacteria found in a ventral tissue called lumen organ. The relationship between V. fischeri and E. scolopes is characterized by daily rhythmic cycles that control the population dynamics of bacteria. At night, when squid take feed, the light organ fills with bioluminescent V. fischeri. At dawn, the squid bury themselves in the sand and ventilate approximately 90% of bacterial population to environment, the remaining bacteria repopulating the crypts and are ready to produce light at dusk [101]. This example of symbiosis shows beneficial microorganisms' important role in their hosts' health and activities. The role of the VBNC state in V. fischeri free in marine environment or symbiosis is still unknown. It has only been discovered that non-culturable cells lose their luminescence in response to fluctuations in salinity [63]. The loss of luminescence could have an application for the diagnosis of marine environments allowing know alterations in salinity, temperature variety and concentration of nutrients relating the number of luminescent bacteria of V. fischeri, interpreting that the less luminescent bacteria, the disturbance present in marine environment is greater.
The actinobacteria group includes Arthrobacter albidus, reclassified as Sinomonas albida is an that was isolated from a seep substrate made of volcanic rock from Niigata, Japan [102]. Among the main applications of this microorganism, it has been demonstrated that it leaves the VBNC state in the presence of RpF proteins, has an efficient flocculant activity [65]. This flocculent activity represents a possible application in wastewater treatment and residual sludge dewatering. Until now, little is known about this species, which represents an alternative and challenge to take advantage of the biotechnological properties that S. albida can provide.
The presence of bacteria in the human digestive tract has various properties related to health, such as regulation of intestinal microbial homeostasis, inhibition of pathogenic bacteria, modulation the immune response, anticancer effects, production of bacteriocins or bioconversion of diet components in bioactive compounds [103]. Recently, production and consumption of products with beneficial strains for human health has increased considerably [104]. Bifidobacterium longum and Bifidobacterium animalis subsp. lactis are multifunctional probiotic Actinobacteria clinically effective as immunomodulatory, anti-inflammatory, antimutagenic, anticancer and alleviation of gastrointestinal diseases [104]. In both species it has been observed that their storage at low temperatures in fermented foods can cause loss of viability (Table 2). In this respect, determination of the viability and activity of probiotic bacteria is of great economic, regulatory and technological importance to ensure that fermented products or formulations with probiotics carried bacteria capable of exerting their beneficial effect.
Another probiotic, Bacillus coagulans is a Firmicute considered safe. This bacterium has the ability to endure high temperatures and genetic stability through several years of commercial production [105]. Its main benefits to human health include modulation of gastrointestinal disorders, immune system stimulation, and lowering cholesterol [106]. This bacterium can form endospores and survive during decades in unfavorable environmental conditions. It was previously thought that their high persistence was due only to ability to form spores, however, it has been observed that B. coagulans can enter the VBNC state as a strategy to face adverse conditions [67]. Little is known about the return to viability when humans consume these beneficial microorganisms. Some studies, mainly with pathogens, have shown that bacteria secrete certain specific proteins to come out of the latency state [107], which could be a possibility for the case of B. longum and B. animalis, this represents a challenge for future research. If we consider that some beneficial bacteria are capable of returning to the culturable state when they interact with their host [4] is very likely think that, although culturable bacteria are not detected in fermented foods, when they are consumed by humans, conditions present in these host may favor its return to cultivable state, being able to exercise its beneficial properties.
Lactobacillus plantarum is a firmicute widely distributed in various environments such as the gastrointestinal, vaginal, and urogenital tracts, in dairy products, vegetables, meat, hay, and wine. This ability to adapt to different conditions demonstrates its metabolic diversity [108] having potential for various applications. The main applications of L. plantarum include fermentation of foods such as cheese, kefir, sauerkraut, fermented meat products, fermented vegetables and beverages [109]. It has been reported that L. plantarum produces antimicrobial substances, such as plantaricin [110] and can remove microcystins, the main toxins produced by cyanobacteria [111], which makes it an alternative as a food preservative or fighting infections. Studies about VBNC state in L. plantarum, reported that can remain latent during beer storage, which contributes to the deterioration of this beverage [31]. Another study showed that L. plantarum in VBNC state can inhibit microcystins, which contributes to the preservation of fermented foods [111]. It is interesting to observe that the VBNC state in this specie can be detrimental for beer production, but beneficial for food preservation, reflecting the versatility of non-cultivable L. plantarum to continue performing its functions. This leads us to think that when used as a probiotic to improve human health, it can provide its benefits without being culturable. In short, it is a microorganism that should be studied in greater depth. Mainly what happens during the VBNC state to take advantage of its diversity of applications.
In the production of alcoholic beverages, the involvement of microorganisms is crucial for a successful fermentation. A significant challenge in the brewing and winemaking industry is the spoilage of wine and beer by lactic acid bacteria, which have the ability to enter the VBNC state [31]. Oenococcus oeni is a firmicute bacterium, belonging to lactic acid bacteria group, adapted to stressful environment of wine. It is widely used as starter microorganism to carry out malolactic fermentation (MLF), where L-malate is converting to L-lactate [112]. The use of starter cultures of O. oeni remains difficult in some wine regions, due to the hostile environment created by low pH and the presence of SO2 and ethanol [113]. The ability of O. oeni to respond these stress conditions has great relevance for increasing wine production at lower cost. So far, knowledge about the VBNC status of O. oeni is limited. It is known that presence of sulfur dioxide induces the loss of viability, and the addition of arginine to medium allows the return to the cultivable state [112]. Due to findings with O. oeni, it is interesting to think that, although bacteria are in VBNC state, they may still be capable of carrying out malolactic fermentation, although perhaps not as efficiently as their cultivable counterparts. When cells are present in a rich environment and added arginine, it returns to the cultivable state, increasing its fermentation efficiency. The generation of new knowledge is challenging, but it also involves changing the paradigm and legislation that require all microorganisms used in food production to be culturable. It is feasible to think that using non-culturable cells would decrease production costs because they would not require an expensive infrastructure to preserve bacteria, since they have all the necessary equipment to preserve themselves.
The VBNC state could be an important reservoir of beneficial bacteria species, as this state constitutes a survival strategy in response to harsh environmental conditions. The capability of bacteria to enter the VBNC state in response to stress could have important biotechnological applications. It is possible consider that VBNC state may reduce negative selection and regulate microbial dominance in soil, as the rhizosphere, where plants could induce a selective bacterial revival by releasing selected organic compounds, directly influencing the diversity present in these environments. In the same way, the hosts themselves could be exerting a selection by releasing compounds that determine the viability of certain bacteria necessary in a given physiological state.
Figure 3.
Methodologies to evaluate bacterial viability in the VBNC state.
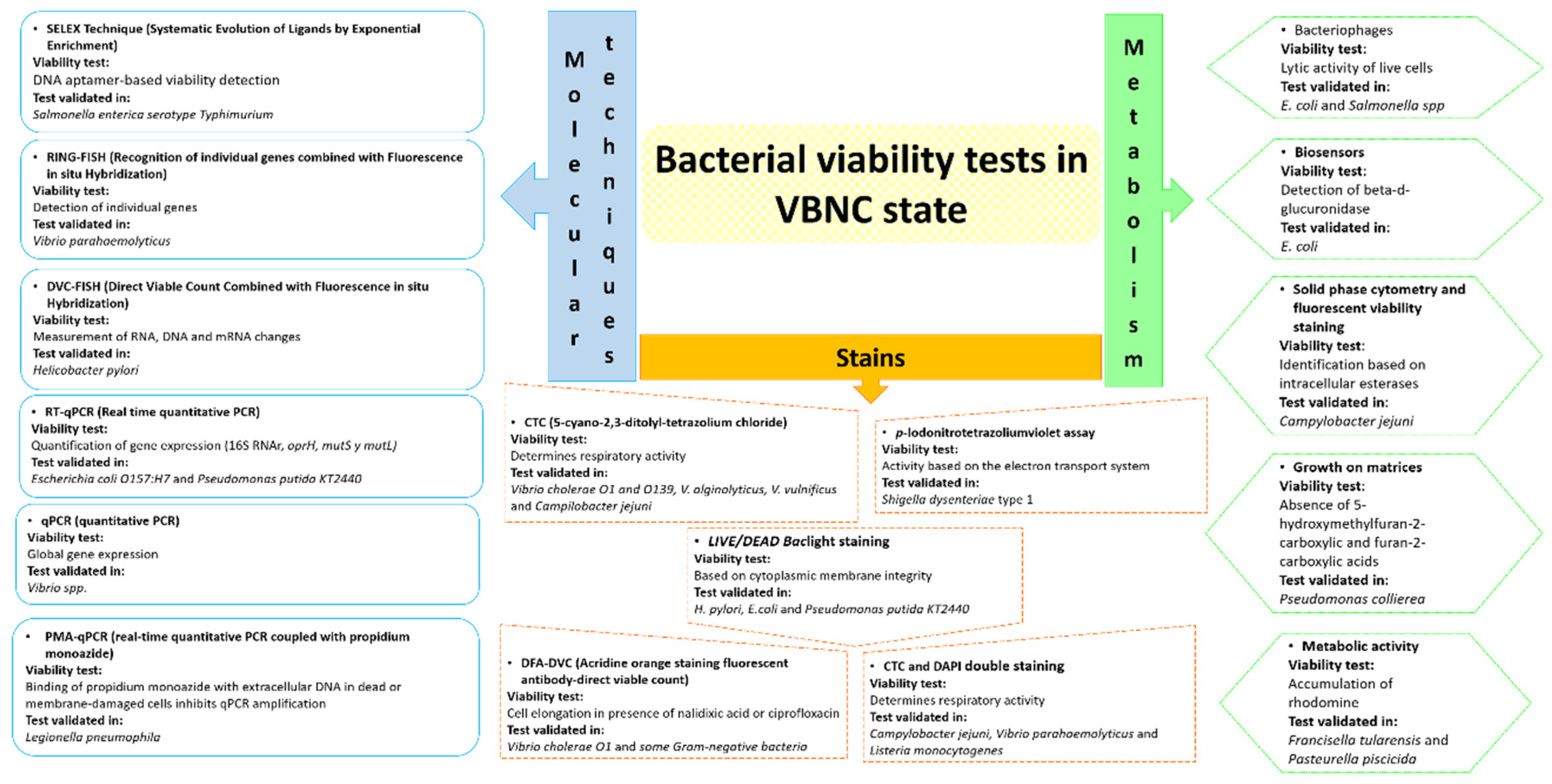
4. Techniques to evaluate bacterial viability in the VBNC state
One challenge that researchers who study bacteria in the VBNC state face is verifying the viability of these cells. Currently, several methodologies have been reported that can be used to determine the viability of a bacterium (Figure 3). The methodologies are divided into three groups: molecular techniques, techniques focused in metabolism and staining techniques. The molecular techniques are based on detecting individual genes or individual and global gene expression in non-culturable cells. According to the literature, the use of transcripts is an excellent alternative, given that mRNA half-lives are typically in the range of seconds to minutes [114,115] and the identification of the mRNA is evidence that cells remain metabolically active. In addition, its detection providing an essential insight into factors that may be regulated the VBNC sate. The techniques focused in metabolism search metabolites or enzymatic activity, generally used methodologies with colorimetric results, biosensors or matrices. The staining techniques mainly are based in detect activity in electron transport system and integrity of cytoplasmatic membrane. Cellular respiration allows quickly identifying metabolically activity cells, only using a compound fluorescent or a compound that reacts and forms a fluorescent compound. The BacLight® Live/Dead kit, is a method frequently used in which, by differential staining and fluorescence microscopy, bacteria in the VBNC state stain green indicating that they have largely intact membranes and thus can be considered to be alive [12]. However, in the case of bacteria during desiccation stress, membranes suffer an apparent sublethal damage during the VBNC state and they stain red [4]. Is important to mention that there is not a decisive test for bacterial viability in the VBNC state, and it is usually proposed that, when trying to prove this fact two or more methodologies should be performed.
5. What happens during the VBNC state?
There are different characteristics shown by cells in the VBNC state. Several bacterial species decrease their size, producing several metabolic changes, including depletion of energy reserves, altered gene expression and DNA replication [3]. Biosynthesis is a process that does not stop during this state; with cells forming new proteins related to starvation and cold shock [116,117]. ATP levels rapidly decrease in dead cells, however, in VBNC cells these levels are high [118]. Other studies have shown that E. coli O157: H7 cells in the VBNC state show continuous gene expression of mobA, rfbE, stx1 and those responsible for synthesizing 16S rRNA [119]. Similarly, in V. vulnificus VBNC cells, a continuous production of messenger RNAs for different genes was observed for 4.5 months [120]. Cells of Shigella dysenteriae in the VBNC state could actively capture methionine to incorporate it into several proteins during 4-8 weeks [121]. Pazos-Rojas et al., (2019) detected the expression of oprH, mutS, mutL genes and 16S RNA in P. putida KT2440 VBNC cells under desiccation stress [4]. These studies suggest that some genes may be effective monitors for viability, but whether they are involved in the entrance or exit from VBNC state is still unknown.
At the structural level, it has been described that VBNC cells may suffer damage to the plasmatic membrane and modifications in the composition of fatty acids, strongly suggesting that changes may be essential for the entrance into this state [4,122]. Cells may undergo biochemical changes in cell walls; in example, the peptidoglycan cell wall of E. coli in the VBNC state suffer some changes. An increment of 3-fold of DAP-DAP crosslinking was observed in peptides carrying covalently bound lipoproteins, as well as a shortening of the average length of glycan chains [123]. Hence, genes involved in peptidoglycan biosynthesis could be a required characteristic of VBNC cells to create a more rigid wall than in dividing cells [124].
Bacteria can detect stressors through histidine kinases bound to the membrane, mediating cellular response through differential expression of the target genes [125]. Therefore, early signaling could be a determining factor for VBNC state induction in bacteria. As mentioned above, cellular energy is one of the key characteristics of the VBNC state, triggering the induction of genes codifying subunits of proton pumps such as NADH:ubiquinone oxidoreductase, a protein essential for processes requiring energy in VBNC cells as well as under normal conditions [126]. This suggests that when bacterial cells enter the VBNC state in response to an environmental change, the activity of complex I of the respiratory chain and NADPH-generating systems are critical for the maintenance of cell viability [126]. In addition, selective permeability to nutrients and metabolites provided by ABC transporters may be a prerequisite for the VBNC state [127]. It is possible to consider that this behavior is widely distributed among bacteria. What has been described above in pathogenic microorganisms may also occur in beneficial bacteria. Nevertheless, further studies are needed to better understand it
6. Proteomics and transcriptomics of bacterial cells in VBNC state
When bacteria enter the VBNC state, various cellular changes may occur. Studying changes at the post-transcriptional and post-translational levels could elucidate which genes could be important for bacteria to perform this phenomenon; however, knowledge is still limited under the VBNC state. H. pylori produce more alkaline phosphatase and α-ketoglutarate oxidoreductase (KOR), while CoA transferase levels decrease in the Krebs cycle. There is also a decrease in urease, leucine arilamidase and naftol-AS- β -1-phosphohidrolase [128]. Based on these observations, it has been proposed that it would be possible to eliminate multi-drug resistant strains of H. pylori by activating the KOR enzyme, preventing cells from entering the VBNC state [129]. On the other hand, latent E. coli (EHEC) O157 isolated from food, had low levels of AhpCF and AceF proteins, but showed an increase in OmpW external membrane proteins [130]. Transcriptomic studies in V. vulnificus, showed a high expression of glutathione S-transferase, suggesting that when the VBNC state is induced by oxidative stress, this enzyme may prevent damage to different cellular components [131,132]. Under saline stress at low temperature several proteins related to transcription, translation, ATP synthesis, gluconeogenesis and antioxidants were over-expressed in V. parahaemolyticus [133]. On the other hand, V. parahaemolyticus in VBNC state induced by low temperature, an over-expression of gene dacB which encodes D-alanyl-D-alanyl carboxypeptidase was observed, resulting in the interruption of cell wall synthesis, and consequently the production of amorphic cells [134]. It is interesting that under saline stress at low temperatures, the protein AhpC (Alkyl hydroperoxide reductase subunit C) of V. parahaemolyticus can delay the entrance into the VBNC state [135]. The protein AhpC is responsible for the detoxification of reactive oxygen species that form in bacterial cells and facilitates the survival of pathogenic bacteria under environmental stresses, suggesting the importance of antioxidative activity during the induction of the VBNC state [135,136]. In VBNC cells of V. cholerae by cold stress an over-expression of different genes was detected, such as polB corresponding to DNA polymerase II, fliG codifying for proteins of the flagellar motor, flaC for protein C in the flagellar subunit and ABC iron transporter genes [137]. It has been observed that the tcp gene which encodes the pilus toxin and is required for bowel colonization in lactating mice, is expressed in coccoid cells of V. cholerae in the VBNC state. The transition from bacillary to coccoid morphology is thought to be a mechanism of survival that V. cholerae uses in response to environmental stress and the expression of the tcp gene in coccoid cells in the VBNC state suggests that this could be very important for the adaptation and survival of V. cholerae during the transition between the host and aquatic environment [138]. In C. jejuni, the expression of the external membrane protein CadF was detected after entering the VBNC state. This protein functions as a mediator of fibronectin binding of Caco2 cells. The expression of this type of proteins suggests that even when bacteria enter the latency state, they can maintain their virulence [139].
The expression of the RpoS sigma factor in the VBNC state has been demonstrated in several bacterial species, with guanosine 3', 5'-bispirophosphate (ppGpp) acting as a positive regulator during the synthesis and function of this factor [140], interestingly it has been observed that RpoS mutant strains quickly lose culturability and cannot return to the culturable state with existing resuscitation methods [132,140]. Considering that the absence of this protein can mean imminent cell death, it could be one of the key genes involved in the entrance and exit from the VBNC state in bacteria.
The activity in the ribosomes is one of the most important processes in all living cells. A low expression level for RaiA proteins (Ribosome-associated inhibitor A) has been observed in the VBNC state, for several bacterial species [132]. This could mean that bacteria decrease their translational activity during the latency status, thereby saving energy required to face stress conditions. As described in this review, different bacterial groups modulate genes responsible for cellular processes such as cell wall reordering, detoxification, transporters and virulence genes.
Transcriptomic studies of P. syringae in VBNC state, with presence of acetosyringone synthesized in some plants in response to bacteria, showed that a complex network of biochemical reactions related to oxidative phosphorylation, ABC transporters, peptidoglycan biosynthesis, Krebs cycle, chemotaxis, two-component system and bacterial secretion systems, are important for keeping cells in the VBNC state. Likely the entrance into this state begins with metabolic pathways that control stimulus-response mechanisms (two-component system) and bacterial movement (chemotaxis), determined by the presence of chemicals in the environment where the cells are located. In addition, bacterial survival could depend on multi-drug resistance efflux pumps, which could give them tolerance to the presence of oxidative residues caused by stress [127]. The same research also found a positive regulation of genes involved in peptidoglycan biosynthesis, genes of the transcriptional factors LysR, Lrp/AsnC, and MarR. It is proposed that keeping cells in VBNC state may be under the control of LysR, which is the most abundant transcription factor in bacteria that regulates several genes (virulence, metabolism, quorum sensing and motility) [141]. Furthermore, the VBNC state could also be regulated by the proteins Lrp/AsnC (leucine-responsive regulatory protein/asparagine synthase C products) and MarR [127]. The MarR protein controls genes involved in the degradation of toxic compounds (including phenols), virulence, export of harmful chemicals and resistance to oxidative stress [142]. Another interesting finding was the repression of genes related to pathogenesis pathways, for example, Type III Secretion System (T3SS) and type VI secretion system proteins, which could be an additional requirement or consequence of the VBNC state in P. syringae.
Within our scant knowledge on beneficial bacteria, there are studies on P. putida KT2440 and C. metallidurans. Transcriptomic studies of P. putida KT2440 in VBNC state caused by desiccation stress [143], shown that 6 genes related to transmembrane transport and oxidation-reduction processes were upregulated. Ethylene glycol porin (PP_ 2662) and substrate-binding protein (PP_2676) genes could transport and degrade polyhydric alcohols, which are accumulated during desiccation stress as compatible solutes. The upregulation of TonB dependent receptor (PP_1446) gene could be a strategy used by P. putida KT2440 to provide ferrous-iron required for vital functions during the VBNC state. P. putida KT2440 cells return to cultivable sate upon 24 h of rehydration, after this return, 148 genes related to transport, oxidation-reduction, regulation of transcription, and biosynthetic process were upregulated while 42 genes related with translation, oxidation-reduction, and the regulation of transcription were downregulated. During prolonged rehydration of P. putida KT2440 cells the catabolism of phenylalanine/tyrosine is activated, maybe to provide energy and carbon source for ubiquinone biosynthesis while maintaining reduced protein synthesis [143].
Studies in C. metallidurans showed that during transition from the culturable state to VBNC state, there is a strong decrease in the expression of proteins involved in different mechanisms such as: basal bacterial metabolism, cellular processes, signaling, information storage, protein synthesis pathways, energetic processes and cell shape regulation [60]. The proteins that showed an increase in their regulation are involved in producing energetic processes and redox reactions. Proteomic analysis of return to culturable state showed that gluconate increased the expression of several proteins related to fundamental bacterial metabolism when used. While, when only water was added to promote the return to the culturable state, the expression of only six proteins was increased. These results suggest that reduced soil carbon or water availability could initiate bacterial VBNC state in soil-like environments. This limitation could induce gene expression and protein synthesis leading to the VBNC state.
7. Resuscitation of bacteria under VBNC state
As mentioned in previous sections, bacteria in the viable but non-culturable state do not grow in routine bacteriological media. However, they are still alive, which may constitute a survival strategy under stressful conditions. The cells must be able to increase their metabolic activity to return to the culturable state [2]. It has been a topic of discussion and a great challenge to demonstrate that cells can return to the culturable state and that observed cells which grow again on media do not simply correspond to the growth of other surviving bacteria. V. vulnificus is one of the best studied microorganisms able to return to the culturable state. This bacterium enters the VBNC state in response to low temperatures (10 °C) [117]. Numerous studies have reported that a simple reversal of stress is sufficient to return VBNC cells to the culturable state, for example, by changing the temperature to that for optimal growth [144,145,146]. Similarly, it was observed for V. parahaemolyticus temperature changes control the VBNC state [147].
In P. putida KT2440 cells in VBNC state induced by desiccation, the colonization of rhizosphere of maize plants, short rehydration in presence of root exudates and prolonged rehydration with only distilled sterile water were shown to allow the return to a culturable state [4].
Interestingly, it has been observed that the interaction with higher organisms can function as a biological mediator to return to the culturable state. For example, in L. pneumophila which enters the VBNC state under starvation and hypochlorite treatment, it can return to the culturable state in presence of protozoa Acanthamoeba polyphaga and Acanthamoeba castellanii [148,149]. In a different system, the association of VBNC cells of the coral pathogen, Vibrio shiloi, with the seafire worm Hermodice carunculate is a strategy for remain and infect coral [150]. The resuscitation of the pathogen L. monocytogenes was observed after inoculation of pathogen free embryonic chicken eggs [151]. Regarding beneficial bacteria, a recent study has shown that VBNC cells of P. putida KT2440, can colonize the rhizosphere of maize plants and this interaction allows the return to the culturable state [4]. Furthermore, root exudates of maize can return these bacteria to the culturable state [98], thus, it is of interest to study bacterial associations with their host since, this influences the survival of the bacteria even in the VBNC state.
Other factors that could be decisive for the return to the culturable state, are based on the study of extracellular proteins Rpfs (resuscitation-promoting factors). Rpf was discovered in Micrococcus luteus as a cytokine, that promotes resuscitation and growth of dormant cells [152]. It was reported that a picomolar concentration of Rpf could increase the viable cell number of dormant M. luteus [153]. Other study reported that Rpf and RipA proteins work synergistically to remodel the cell wall promoting cell division and resuscitation. The cell wall fragments digested by Rpf and RipA serve as signaling molecules binding with a receptor to trigger resuscitation [154]. RpF-like proteins have been reported with the ability to induce return to cultivable state. The YeaZ protein with protease activity present in V. parahaemolyticus, V. harveyi, S. typhimurium, and E. coli has been shown to promote return to culturable state [155]. The promoting effect of YeaZ may be correlated with its protease activities, but mechanism that help recovery cultivability lacks further investigation.
Return to the culturable state in M. tuberculosis and M. smegmatis, is promoted by activity of a peptidoglycan hydrolase, involved in the digestion of the cell wall and cell division. In this way, reorganization of peptidoglycan plays an important role in the VBNC state [124,153,156]. Another type of proteins involved in the return to the culturable state are the so called "autoinducer of growth", which are stable to heat and are secreted by Gram-positive and Gram-negative bacteria growing in culture media with the hormone norepinephrine [157]. Humans release this type of hormone after the occurrence of an injury, thus in humans it is considered a stress-related hormone [157,158]. These findings are of great relevance since it was observed that in the presence of autoinducer proteins in culture media other bacteria such as S. enterica serotype Typhimurium, and two E. coli O157:H7 strains were able to return to the culturable state [159]. These investigations provide a breakthrough in understanding the return to the culturable state of enteropathogens, found in the intestinal tract when the host (human) suffers tissue damage and is under significant physiological stress.
Molecules such as sodium pyruvate have been studied in their role in return to culturable state. This molecule is an intermediate key metabolite in glycolysis and H2O2 degrading compound [160]. It was observed that VBNC cells can return to culturable state in a media supplemented with sodium pyruvate [161]. It was suggested that sodium pyruvate, catalase and superoxide dismutase, due to their H2O2 or reactive oxygen-degrading effect can induce return to cultivable state [144].
The Quorum sensing (QS) is a mechanism widespread communication system in bacteria that induces global gene expression changes [162]. It has been observed that QS can induce an adaptation to stressful conditions and play a role in return of VBNC state. Studies in P. aeruginosa indicated that QS system has a role in activating superoxide dismutase or catalase to regulate the antioxidation activities [163]. In S. typhimurium, QS triggered the expression of catalase to restore the culturable state [164]. It is proposed that QS may help the cell to express genes related to oxidative stress, which as mentioned above play an important role in allowing return to the culturable state of different bacteria. Being certain that the VBNC state is reversible opens up a range of possibilities for using microorganisms in different areas that may offer some benefit.
8. Conclusions
The study of the VBNC state in bacteria remains controversial, because its verification does not completely satisfy all researchers worldwide, even though, many scientists have concluded that this is a survival strategy under stressful conditions. Regardless of the role of the VBNC state in the life cycle of bacteria, it has been fully proven that many bacterial species, especially human pathogens, can carry out this survival strategy. During the VBNC state bacteria maintain their cellular structure and biological functions, such as cellular respiration, and continuous gene expression, having the ability to leave this state and return to a culturable state when conditions become more favorable. Despite the advances made over the last thirty years, it remains a challenge to investigate this state's physiology, biochemistry and genetics, because it is unknown what gene or genes could directly be involved with the loss and recovery of culturability. The study of beneficial bacteria in the VBNC state is still little explored. Expanding knowledge in this field could represent an opportunity for future biotechnological applications, for example, for the biological control of plant pathogens, since even when bacteria enter the VBNC state under stressful conditions, when returning to the culturable state they can continue exercising their biocontrol over species harmful to their hosts. Microbial biocontrol technology avoids the use of pesticides that are highly toxic to the environment. Another application is the formulation of bacterial inoculants with beneficial species capable of entering the VBNC state, to increase its shelf life, so that when applied to the plant rhizosphere, bacteria will be able to return to the culturable state and carry out their beneficial activity on the growth of plants of agricultural interest. The impact of beneficial bacteria on human health represents a challenge to understand the role of the VBNC state and how it can affect or benefit hosts. Changing the topic on the elaboration of consumable biotechnological products using only culturable microorganisms is a real challenge for the industry, however we must understand that bacteria have their preservation system such as the VBNC state that can represent a significant decrease in production costs. This highlights the importance of more precisely understanding the transition between the culturable and non-culturable state.
Author Contributions
Conceptualization, L.A. P.-R., Y. E. M.-G., P. v. D., and M. R. B.-C.; software, L.A. P.-R., A. C.-S., A. L. R.-C., D. A. L.-V.; validation, J. M.-R., Y. E. M.-G., and M. R. B.-C.; data curation, J. M.-R., Y. E. M.-G., and M. R. B.-C.; writing—original draft preparation, L. A. P.-R, J. M.-R., P. v. D., A. R.-U.; writing—review and editing, J. M.-R., Y. E. M.-G., and M. R. B.-C.; visualization, D. A. L.-V., A. R.-U.; supervision, Y. E. M.-G., and M. R. B.-C.; project administration, J. M.-R.; funding acquisition, J. M. R. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Internationalization of Research program at VIEP-BUAP (VIEP-Muñoz-Rojas 100425788).
Acknowledgments
L. A. P.-R., A. C.-S., D. A. L.-V., A. R.-U., J. M-R., Y. E. M.-G., M. R. B.-C. are members of the National Researchers System (Sistema Nacional de Investigadores) of CONAHCyT, and they thank this institution for the support provided.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Oliver, J.D. The Viable but Nonculturable State and cellular resuscitation. Microbial biosystems: New frontiers 2000, 723–730.
- Oliver, J.D. The Viable but Nonculturable State in Bacteria. Journal of Microbiology 2005, 43, 93–100.
- Trevors, J.T. Viable but Non-Culturable (VBNC) Bacteria: Gene expression in planktonic and biofilm cells. Journal of Microbiological Methods 2011, 86, 266–273. [CrossRef]
- Pazos-Rojas, L.A.; Muñoz-Arenas, L.C.; Rodríguez-Andrade, O.; López-Cruz, L.E.; López-Ortega, O.; Lopes-Olivares, F.; Luna-Suarez, S.; Baez, A.; Morales-García, Y.E.; Quintero-Hernández, V.; et al. Desiccation-Induced Viable but Nonculturable state in Pseudomonas putida KT2440, a survival strategy. PLoS ONE 2019, 14, e0219554. [CrossRef]
- Gunasekera, T.S.; Sørensen, A.; Attfield, P.V.; Sørensen, S.J.; Veal, D.A. Inducible gene expression by nonculturable bacteria in milk after pasteurization. Appl Environ Microbiol 2002, 68, 1988–1993. [CrossRef]
- Cunningham, E.; O’Byrne, C.; Oliver, J.D. Effect of weak acids on Listeria monocytogenes survival: Evidence for a Viable but Nonculturable state in response to low pH. Food Control 2009, 20, 1141–1144. [CrossRef]
- Xu, H.-S.; Roberts, N.; Singleton, F.L.; Attwell, R.W.; Grimes, D.J.; Colwell, R.R. Survival and viability of nonculturable Escherichia coli and Vibrio cholerae in the estuarine and marine environment. Microb Ecol 1982, 8, 313–323. [CrossRef]
- Barcina, I.; González, J.M.; Iriberri, J.; Egea, L. Effect of visible light on progressive dormancy of Escherichia coli cells during the survival process in natural fresh water. Appl Environ Microbiol 1989, 55, 246–251. [CrossRef]
- Davies, C.M.; Evison, L.M. Sunlight and the survival of enteric bacteria in natural waters. Journal of Applied Bacteriology 1991, 70, 265–274. [CrossRef]
- Trainor, V.C.; Udy, R.K.; Bremer, P.J.; Cook, G.M. Survival of Streptococcus pyogenes under stress and starvation. FEMS Microbiology Letters 1999, 176, 421–428. [CrossRef]
- Rollins, D.M.; Colwell, R.R. Viable but Nonculturable Stage of Campylobacter jejuni and its role in survival in the natural aquatic environment. Appl Environ Microbiol 1986, 52, 531–538. [CrossRef]
- Vriezen, J.A.; De Bruijn, F.J.; Nüsslein, K.R. Desiccation induces Viable but non-culturable cells in Sinorhizobium meliloti 1021. AMB Expr 2012, 2, 6. [CrossRef]
- Bej, A.K.; Mahbubani, M.H.; Atlas, R.M. Detection of viable Legionella pneumophila in water by Polymerase Chain Reaction and gene probe methods. Appl Environ Microbiol 1991, 57, 597–600. [CrossRef]
- Singh, A.; Yeager, R.; McFeters, G.A. Assessment of in vivo revival, growth, and pathogenicity of Escherichia coli strains after copper-and chlorine-induced injury. Appl Environ Microbiol 1986, 52, 832–837. [CrossRef]
- Chowdhury, M.A.; Huq, A.; Xu, B.; Madeira, F.J.; Colwell, R.R. Effect of alum on free-living and copepod-associated Vibrio cholerae O1 and O139. Appl Environ Microbiol 1997, 63, 3323–3326. [CrossRef]
- Mason, D.J.; Power, E.G.; Talsania, H.; Phillips, I.; Gant, V.A. Antibacterial action of ciprofloxacin. Antimicrob Agents Chemother 1995, 39, 2752–2758. [CrossRef]
- Fakruddin, Md.; Mannan, K.S.B.; Andrews, S. Viable but Nonculturable Bacteria: Food Safety and Public Health Perspective. ISRN Microbiology 2013, 2013, 1–6. [CrossRef]
- Shleeva, M.; Mukamolova, G.V.; Young, M.; Williams, H.D.; Kaprelyants, A.S. Formation of ‘Non-Culturable’ cells of Mycobacterium smegmatis in stationary phase in response to growth under suboptimal conditions and their Rpf-mediated resuscitation. Microbiology 2004, 150, 1687–1697. [CrossRef]
- Young, D.B.; Gideon, H.P.; Wilkinson, R.J. Eliminating latent tuberculosis. Trends in Microbiology 2009, 17, 183–188. [CrossRef]
- Pai, S.R.; Actor, J.K.; Sepulveda, E.; Hunter, R.L.; Jagannath, C. Identification of Viable and Non-Viable Mycobacterium tuberculosis in mouse organs by directed RT-PCR for antigen 85B mRNA. Microbial Pathogenesis 2000, 28, 335–342. [CrossRef]
- Magariños, B.; Romalde, J.L.; Barja, J.L.; Toranzo, A.E. Evidence of a dormant but infective state of the fish pathogen Pasteurella piscicida in seawater and sediment. Appl Environ Microbiol 1994, 60, 180–186. [CrossRef]
- Biosca, E.G.; Amaro, C.; Marco-Noales, E.; Oliver, J.D. Effect of low temperature on starvation-survival of the Eel pathogen Vibrio vulnificus biotype 2. Appl Environ Microbiol 1996, 62, 450–455. [CrossRef]
- Banin, E.; Israely, T.; Kushmaro, A.; Loya, Y.; Orr, E.; Rosenberg, E. Penetration of the coral-bleaching bacterium Vibrio shiloi into Oculina patagonica. Appl Environ Microbiol 2000, 66, 3031–3036. [CrossRef]
- Israely, T.; Banin, E.; Rosenberg, E. Growth, differentiation and death of Vibrio shiloi in coral tissue as a function of seawater temperature. Aquat. Microb. Ecol. 2001, 24, 1–8. [CrossRef]
- Grey, B.E.; Steck, T.R. The Viable But Nonculturable State of Ralstonia solanacearum may be involved in long-term survival and plant infection. Appl Environ Microbiol 2001, 67, 3866–3872. [CrossRef]
- Del Campo, R.; Russi, P.; Mara, P.; Mara, H.; Peyrou, M.; De León, I.P.; Gaggero, C. Xanthomonas axonopodis Pv. citri enters the VBNC state after copper treatment and retains its virulence. FEMS Microbiology Letters 2009, 298, 143–148. [CrossRef]
- Ordax, M.; Biosca, E.G.; Wimalajeewa, S.C.; López, M.M.; Marco-Noales, E. Survival of Erwinia amylovora in mature apple fruit calyces through the Viable but Nonculturable (VBNC) state. Journal of Applied Microbiology 2009, 107, 106–116. [CrossRef]
- Kan, Y.; Jiang, N.; Xu, X.; Lyu, Q.; Gopalakrishnan, V.; Walcott, R.; Burdman, S.; Li, J.; Luo, L. Induction and Resuscitation of the Viable but Non-Culturable (VBNC) state in Acidovorax citrulli, the causal agent of bacterial fruit blotch of cucurbitaceous Crops. Front. Microbiol. 2019, 10, 1081. [CrossRef]
- Pinto, D.; Santos, M.A.; Chambel, L. Thirty years of Viable but Nonculturable state Research: Unsolved Molecular Mechanisms. Critical Reviews in Microbiology 2015, 41, 61–76. [CrossRef]
- Liu, J.; Deng, Y.; Peters, B.M.; Li, L.; Li, B.; Chen, L.; Xu, Z.; Shirtliff, M.E. Transcriptomic analysis on the formation of the viable putative non-culturable state of beer-spoilage Lactobacillus acetotolerans. Sci Rep 2016, 6, 36753. [CrossRef]
- Liu, J.; Li, L.; Li, B.; Peters, B.M.; Deng, Y.; Xu, Z.; Shirtliff, M.E. Study on spoilage capability and VBNC state formation and recovery of Lactobacillus plantarum. Microbial Pathogenesis 2017, 110, 257–261. [CrossRef]
- Liu, J.; Deng, Y.; Li, L.; Li, B.; Li, Y.; Zhou, S.; Shirtliff, M.E.; Xu, Z.; Peters, B.M. Discovery and control of culturable and Viable but Non-Culturable cells of a distinctive Lactobacillus harbinensis strain from spoiled beer. Sci Rep 2018, 8, 11446. [CrossRef]
- Li, Y.; Yang, L.; Fu, J.; Yan, M.; Chen, D.; Zhang, L. The novel loop-mediated isothermal amplification based confirmation methodology on thebacteria in Viable but Non-Culturable (VBNC) state. Microbial Pathogenesis 2017, 111, 280–284. [CrossRef]
- Daranas, N.; Bonaterra, A.; Francés, J.; Cabrefiga, J.; Montesinos, E.; Badosa, E. Monitoring viable cells of the biological control agent Lactobacillus plantarum PM411 in aerial plant surfaces by means of a strain-specific viability quantitative PCRmethod. Appl Environ Microbiol 2018, 84, e00107-18. [CrossRef]
- Santander, R.D.; Figàs-Segura, À.; Biosca, E.G. Erwinia amylovora catalases KatA and KatG are virulence factors and delay the starvation-induced Viable but Non-culturable (VBNC) response. Molecular Plant Pathology 2018, 19, 922–934. [CrossRef]
- Dong, K.; Pan, H.; Yang, D.; Rao, L.; Zhao, L.; Wang, Y.; Liao, X. Induction, detection, formation, and esuscitation of Viable but Non-culturable state microorganisms. Comp Rev Food Sci Food Safe 2020, 19, 149–183. [CrossRef]
- Zhang, X.-H.; Ahmad, W.; Zhu, X.-Y.; Chen, J.; Austin, B. Viable but Nonculturable bacteria and their resuscitation: Implications for cultivating uncultured marine microorganisms. Mar Life Sci Technol 2021, 3, 189–203. [CrossRef]
- Oliver, J.D. Recent findings on the Viable but Nonculturable state in pathogenic bacteria. FEMS Microbiol Rev 2010, 34, 415–425. [CrossRef]
- Dobereiner, J.; Urquiaga, S. Alternatives for Nitrogen nutrition of crops in Tropical agriculture. Nitrogen Economy in Tropical Soils: Proceedings of the International Symposium on Nitrogen Economy in Tropical Soils, held in Trinidad, W.I., January 9–14, 1994.
- Crowley, D.E.; Wang, Y.C.; Reid, C.P.P.; Szaniszlo, P.J. Mechanisms of iron acquisition from siderophores by microorganisms and plants. In Developments in Plant and Soil Sciences; Srpinger, 1991; pp. 179–198 ISBN 978-94-010-5455-3.
- Hoefman, S.; Van Hoorde, K.; Boon, N.; Vandamme, P.; De Vos, P.; Heylen, K. Survival or revival:long-term preservation induces a reversible Viable but Non-Culturable state in methane-oxidizing bacteria. PLoS ONE 2012, 7, e34196. [CrossRef]
- Babalola, O.O. Beneficial bacteria of agricultural importance. Biotechnol Lett 2010, 32, 1559–1570. [CrossRef]
- Lugtenberg, B.; Kamilova, F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [CrossRef]
- Hayat, R.; Ali, S.; Amara, U.; Khalid, R.; Ahmed, I. Soil Beneficial bacteria and their role in plant growth promotion: a review. Ann Microbiol 2010, 60, 579–598. [CrossRef]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced Systemic Resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [CrossRef]
- Fira, D.; Dimkić, I.; Berić, T.; Lozo, J.; Stanković, S. Biologicalcontrol of plant pathogens by Bacillus species. Journal of Biotechnology 2018, 285, 44–55. [CrossRef]
- Brierley, C.L. Microbiological mining. Sci Am 1982, 247, 44–53. [CrossRef]
- Zaidi, S.; Usmani, S.; Singh, B.R.; Musarrat, J. Significance of Bacillus subtilis strain SJ-101 as a bioinoculant for concurrent plant growth promotion and Nickel accumulation in Brassica juncea. Chemosphere 2006, 64, 991–997. [CrossRef]
- Zhuang, X.; Chen, J.; Shim, H.; Bai, Z. New advances in Plant Growth-Promoting Rhizobacteria for bioremediation. Environment International 2007, 33, 406–413. [CrossRef]
- Leroy, F.; De Vuyst, L. Lactic acid bacteria as functional starter cultures for the food fermentation industry. Trends in Food Science & Technology 2004, 15, 67–78. [CrossRef]
- Chemier, J.A.; Fowler, Z.L.; Koffas, M.A.G. Trends in microbial synthesis of natural products and biofuels. In Advances in Enzymology - and Related Areas of Molecular Biology; Toone, E.J., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2008; Vol. 76, pp. 151–217 ISBN 978-0-470-39288-1.
- Tejero-Sariñena, S.; Barlow, J.; Costabile, A.; Gibson, G.R.; Rowland, I. In vitro evaluation of the antimicrobial activity of a range of probiotics against pathogens: evidence for the effects of organic acids. Anaerobe 2012, 18, 530–538. [CrossRef]
- Liévin-Le Moal, V.; Servin, A.L. The front line of enteric host defense against unwelcome intrusion of harmful microorganisms: mucins, antimicrobial peptides, and microbiota. Clin Microbiol Rev 2006, 19, 315–337. [CrossRef]
- Pagnini, C.; Saeed, R.; Bamias, G.; Arseneau, K.O.; Pizarro, T.T.; Cominelli, F. Probiotics promote gut health through stimulation of epithelial innate immunity. Proc. Natl. Acad. Sci. U.S.A. 2010, 107, 454–459. [CrossRef]
- Lonvaud-Funel, A. Lactic Acid Bacteria in the quality improvement and depreciation of wine. In Lactic Acid Bacteria: Genetics, Metabolism and Applications; Konings, W.N., Kuipers, O.P., In ’T Veld, J.H.J.H., Eds.; Springer Netherlands: Dordrecht, 1999; pp. 317–331 ISBN 978-90-481-5312-1.
- Virdis, C.; Sumby, K.; Bartowsky, E.; Jiranek, V. Lactic Acid Bacteria in wine: technological advances and evaluation of their functional role. Front. Microbiol. 2021, 11, 612118. [CrossRef]
- Millet, V.; Lonvaud-Funel, A. The Viable but Non-Culturable state of wine microorganisms during storage. Lett Appl Microbiol 2000, 30, 136–141. [CrossRef]
- Xia, K.; Han, C.; Xu, J.; Liang, X. Toxin-antitoxin HicAB regulates the formation of persister cells responsible for the acid stress resistance in Acetobacter pasteurianus. Appl Microbiol Biotechnol 2021, 105, 725–739. [CrossRef]
- Alexander, E.; Pham, D.; Steck, T.R. The Viable-but-Nonculturable condition is induced by copper in Agrobacterium tumefaciens and Rhizobium leguminosarum. Appl Environ Microbiol 1999, 65, 3754–3756. [CrossRef]
- Giagnoni, L.; Arenella, M.; Galardi, E.; Nannipieri, P.; Renella, G. Bacterial culturability and the Viable but Non-Culturable (VBNC) state studied by a proteomic approach using an artificial soil. Soil Biology and Biochemistry 2018, 118, 51–58. [CrossRef]
- Furusawa, G.; Diyana, T.; Lau, N.-S. Metabolic strategies of dormancy of a marine bacterium Microbulbifer aggregans CCB-MM1: its alternative electron transfer chain and sulfate-reducing Pathway. Genomics 2022, 114, 443–455. [CrossRef]
- Arana, I.; Muela, A.; Orruño, M.; Seco, C.; Garaizabal, I.; Barcina, I. Effect of temperature and starvation upon survival strategies of Pseudomonas fluorescens CHA0: comparison with Escherichia coli. FEMS Microbiology Ecology 2010, 74, 500–509. [CrossRef]
- Ramaiah, N.; Ravel, J.; Straube, W.L.; Hill, R.T.; Colwell, R.R. Entry of Vibrio harveyi and Vibrio fischeri into the Viable but Nonculturable state. Journal of Applied Microbiology 2002, 93, 108–116. [CrossRef]
- Lahtinen, S.J.; Ahokoski, H.; Reinikainen, J.P.; Gueimonde, M.; Nurmi, J.; Ouwehand, A.C.; Salminen, S.J. Degradation of 16S rRNA and attributes of viability of Viable but Nonculturable probiotic bacteria. Lett Appl Microbiol 2008, 46, 693–698. [CrossRef]
- Su, X.; Shen, X.; Ding, L.; Yokota, A. Study on the flocculability of the Arthrobacter sp., an actinomycete resuscitated from the VBNC state. World J Microbiol Biotechnol 2012, 28, 91–97. [CrossRef]
- Zhou, Y.; Chen, X.; Zhang, Y.; Wang, W.; Xu, J. Description of Sinomonas soli sp. nov., reclassification of Arthrobacter echigonensis and Arthrobacter albidus (Ding et al. 2009) as Sinomonas echigonensis comb. nov. and Sinomonas albida comb. nov., respectively, and emended description of the Genus Sinomonas. International Journal of Systematic and Evolutionary Microbiology 2012, 62, 764–769. [CrossRef]
- Majeed, M.; Majeed, S.; Nagabhushanam, K.; Punnapuzha, A.; Philip, S.; Mundkur, L. Rapid assessment of Viable but Non-Culturable Bacillus coagulans MTCC 5856 in commercial formulations using flow cytometry. PLoS ONE 2018, 13, e0192836. [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Molecular Biology and Evolution 1987. [CrossRef]
- Felsenstein, J. Phylogenies and the comparative method. The American Society of Naturalists 1985, 125, 1–15. [CrossRef]
- Andrés-Barrao, C.; Saad, M.M.; Chappuis, M.-L.; Boffa, M.; Perret, X.; Ortega Pérez, R.; Barja, F. Proteome analysis of Acetobacter pasteurianus during acetic acid fermentation. Journal of Proteomics 2012, 75, 1701–1717. [CrossRef]
- Semrau, J.D.; DiSpirito, A.A.; Yoon, S. Methanotrophs and copper. FEMS Microbiol Rev 2010, 34, 496–531. [CrossRef]
- Jiang, H.; Chen, Y.; Jiang, P.; Zhang, C.; Smith, T.J.; Murrell, J.C.; Xing, X.-H. Methanotrophs: multifunctional bacteria with promising applications in environmental bioengineering. Biochemical Engineering Journal 2010, 49, 277–288. [CrossRef]
- Wendlandt, K.-D.; Stottmeister, U.; Helm, J.; Soltmann, B.; Jechorek, M.; Beck, M. The potential of methane-oxidizing bacteria for applications in environmental Biotechnology. Eng. Life Sci. 2010, NA-NA. [CrossRef]
- Coba De La Peña, T.; Fedorova, E.; Pueyo, J.J.; Lucas, M.M. The symbiosome: legume and rhizobia co-evolution toward a Nitrogen-Fixing organelle? Front. Plant Sci. 2018, 8, 2229. [CrossRef]
- Moawad, H.; Abd el-Rahim, W.M.; Abd el-Aleem, D.; Abo Sedera, S.A. Persistence of two Rhizobium etli inoculant strains in clay and silty loam soils. J. Basic Microbiol. 2005, 45, 438–446. [CrossRef]
- Abdelkhalek, A.; Yassin, Y.; Abdel-Megeed, A.; Abd-Elsalam, K.; Moawad, H.; Behiry, S. Rhizobium leguminosarum bv. viciae-mediated silver nanoparticles for controlling bean yellow mosaic virus (BYMV) infection in faba bean plants. Plants 2022, 12, 45. [CrossRef]
- Mergeay, M.; Monchy, S.; Vallaeys, T.; Auquier, V.; Benotmane, A.; Bertin, P.; Taghavi, S.; Dunn, J.; Van Der Lelie, D.; Wattiez, R. Ralstonia metallidurans, a bacterium specifically adapted to toxic metals: towards a catalogue of metal-responsive genes. FEMS Microbiol Rev 2003, 27, 385–410. [CrossRef]
- Diels, L.; Van Roy, S.; Taghavi, S.; Van Houdt, R. From industrial sites to environmental applications with Cupriavidus metallidurans. Antonie van Leeuwenhoek 2009, 96, 247–258. [CrossRef]
- Lal, D.; Nayyar, N.; Kohli, P.; Lal, R. Cupriavidus metallidurans: a modern alchemist. Indian J Microbiol 2013, 53, 114–115. [CrossRef]
- Reith, F.; Etschmann, B.; Grosse, C.; Moors, H.; Benotmane, M.A.; Monsieurs, P.; Grass, G.; Doonan, C.; Vogt, S.; Lai, B.; et al. Mechanisms of gold biomineralization in the bacterium Cupriavidus metallidurans. Proc. Natl. Acad. Sci. U.S.A. 2009, 106, 17757–17762. [CrossRef]
- Iguchi, H.; Yurimoto, H.; Sakai, Y. Interactions of methylotrophs with plants and other heterotrophic bacteria. Microorganisms 2015, 3, 137–151. [CrossRef]
- Semrau, J.D.; DiSpirito, A.A.; Vuilleumier, S. Facultative methanotrophy: false leads, true results, and suggestions for future research: facultative methanotrophy. FEMS Microbiol Lett 2011, 323, 1–12. [CrossRef]
- Stock, M.; Hoefman, S.; Kerckhof, F.-M.; Boon, N.; De Vos, P.; De Baets, B.; Heylen, K.; Waegeman, W. Exploration and prediction of interactions between methanotrophs and heterotrophs. Research in Microbiology 2013, 164, 1045–1054. [CrossRef]
- Liebner, S.; Zeyer, J.; Wagner, D.; Schubert, C.; Pfeiffer, E.-M.; Knoblauch, C. Methane oxidation associated with submerged brown mosses reduces methane emissions from siberian polygonal tundra: Moss-associated methane oxidation. Journal of Ecology 2011, 99, 914–922. [CrossRef]
- Moh, T.H.; Furusawa, G.; Amirul, A.A.-A. Microbulbifer aggregans sp. nov., isolated from estuarine sediment from a mangrove forest. International Journal of Systematic and Evolutionary Microbiology 2017, 67, 4089–4094. [CrossRef]
- Komarnisky, L.A.; Christopherson, R.J.; Basu, T.K. Sulfur: its clinical and toxicologic aspects. Nutrition 2003, 19, 54–61. [CrossRef]
- Spiers, A.J.; Buckling, A.; Rainey, P.B. The Causes of Pseudomonas diversity. Microbiology 2000, 146, 2345–2350. [CrossRef]
- Preston, G.M. Plant perceptions of plant growth-promoting Pseudomonas. Phil. Trans. R. Soc. Lond. B 2004, 359, 907–918. [CrossRef]
- Bunker, S.T.; Bates, T.C.; Oliver, J.D. Effects of temperature on detection of plasmid or chromosomally encoded Gfp -and Lux-labeled Pseudomonas fluorescens in Soil. Environ. Biosafety Res. 2004, 3, 83–90. [CrossRef]
- Wallace, R.L.; Hirkala, D.L.; Nelson, L.M. Efficacy of Pseudomonas fluorescens for control of mucor rot of apple during commercial storage and potential modes of action. Can. J. Microbiol. 2018, 64, 420–431. [CrossRef]
- Van Der Ent, S.; Van Wees, S.C.M.; Pieterse, C.M.J. Jasmonate signaling in plant interactions with resistance-inducing beneficial microbes. Phytochemistry 2009, 70, 1581–1588. [CrossRef]
- Wang, Z.; Zhong, T.; Chen, K.; Du, M.; Chen, G.; Chen, X.; Wang, K.; Zalán, Z.; Takács, K.; Kan, J. Antifungal activity of volatile organic compounds produced by Pseudomonas fluorescens ZX and potential biocontrol of blue mold decay on postharvest citrus. Food Control 2021, 120, 107499. [CrossRef]
- Ramos-González, M.I.; Ramos-Díaz, M.A.; Ramos, J.L. Chromosomal gene capture mediated by the Pseudomonas putida TOL catabolic plasmid. J Bacteriol 1994, 176, 4635–4641. [CrossRef]
- Espinosa-Urgel, M..; Kolter, R..; Ramos, J.-L.. Root colonization by Pseudomonas putida: love at first sight. Microbiology 2002, 148, 341–343. [CrossRef]
- Matilla, M.A.; Ramos, J.L.; Bakker, P.A.H.M.; Doornbos, R.; Badri, D.V.; Vivanco, J.M.; Ramos-González, M.I. Pseudomonas putida KT2440 causes induced systemic resistance and changes in Arabidopsis root exudation. Environ Microbiol Rep 2010, 2, 381–388. [CrossRef]
- Molina-Romero, D.; Morales-García, Y.-E.; Hernández-Tenorio, A.-L.; Netzahuatl-Muñoz, A.-R. Pseudomonas putida estimula el crecimiento de maíz en función de la temperatura. Revista Iberoamericana de Ciencias 2017, 4,80-88.
- Morales-García; Baez A; Quintero-Hernández V; Molina-Romero D; Rivera-Urbalejo A.P.; Pazos-Rojas L.A., L.A.; Muñoz-Rojas J. Bacterial mixtures, the future generation of inoculants for sustainable crop production. In Field Crops: Sustainable Management by PGPR, Sustainable Development and Biodiversity; Springer Nature Switzerland, 2019; Vol. 23, pp. 11–44 ISBN 978-3-030-30925-1.
- Pazos-Rojas, L.A.; Marín-Cevada, V.; García, Y.E.M.; Baez, A.; Villalobos-López, M.A.; Pérez-Santos, M. Uso de microorganismos benéficos para reducir los daños causados por la revolución verde. Revista Iberoamericana de Ciencias 2016, 3, 72–85.
- Meighen, E.A. Molecular biology of bacterial bioluminescence. MICROBIOL. REV. 1991, 55.
- Jones, B.W.; Nishiguchi, M.K. Counterillumination in the Hawaiian bobtail squid, Euprymna scolopes berry (Mollusca: Cephalopoda). Marine Biology 2004, 144, 1151–1155. [CrossRef]
- Lee, K.-H.; Ruby, E.G. Effect of the Squid Host on the Abundance and Distribution of symbiotic Vibrio fischeri in nature. Appl Environ Microbiol 1994, 60, 1565–1571. [CrossRef]
- Ding, L.; Hirose, T.; Yokota, A. Four novel Arthrobacter species isolated from filtration substrate. INTERNATIONAL JOURNAL OF SYSTEMATIC AND EVOLUTIONARY MICROBIOLOGY 2009, 59, 856–862. [CrossRef]
- Soomro, A.H.; Masud, T.; Anwaar, K. Role of Lactic Acid Bacteria (LAB) in food preservation and human health–a review. Pakistan J. of Nutrition 2001, 1, 20–24. [CrossRef]
- Mousavi Khaneghah, A.; Abhari, K.; Eş, I.; Soares, M.B.; Oliveira, R.B.A.; Hosseini, H.; Rezaei, M.; Balthazar, C.F.; Silva, R.; Cruz, A.G.; et al. Interactions between probiotics and pathogenic microorganisms in hosts and foods: a review. Trends in Food Science & Technology 2020, 95, 205–218. [CrossRef]
- Majeed, M.; Nagabhushanam, K.; Natarajan, S.; Sivakumar, A.; Eshuis-de Ruiter, T.; Booij-Veurink, J.; De Vries, Y.P.; Ali, F. Evaluation of genetic and phenotypic consistency of Bacillus coagulans MTCC 5856: a commercial probiotic strain. World J Microbiol Biotechnol 2016, 32, 60. [CrossRef]
- Saw, C.-Y.; Chang, T.-J.; Chen, P.-Y.; Dai, F.-J.; Lau, Y.-Q.; Chen, T.-Y.; Chau, C.-F. Presence of Bacillus coagulans spores and vegetative cells in rat intestine and feces and their physiological effects. Bioscience, Biotechnology, and Biochemistry 2019, 83, 2327–2333. [CrossRef]
- Gupta, R.Kr.; Srivastava, R. Resuscitation promoting factors: a family of microbial proteins in survival and resuscitation of dormant Mycobacteria. Indian J Microbiol 2012, 52, 114–121. [CrossRef]
- Seddik, H.A.; Bendali, F.; Gancel, F.; Fliss, I.; Spano, G.; Drider, D. Lactobacillus plantarum and its probiotic and food potentialities. Probiotics & Antimicro. Prot. 2017, 9, 111–122. [CrossRef]
- Todorov, S.D.; Franco, B.D.G.D.M. Lactobacillus plantarum: characterization of the species and application in food production. Food Reviews International 2010, 26, 205–229. [CrossRef]
- Barbosa, M.S.; Todorov, S.D.; Ivanova, I.V.; Belguesmia, Y.; Choiset, Y.; Rabesona, H.; Chobert, J.-M.; Haertlé, T.; Franco, B.D.G.M. Characterization of a two-peptide plantaricin produced by Lactobacillus plantarum MBSa4 isolated from brazilian salami. Food Control 2016, 60, 103–112. [CrossRef]
- Surono, I.S.; Collado, M.C.; Salminen, S.; Meriluoto, J. Effect of glucose and incubation temperature on metabolically active Lactobacillus plantarum from dadih in removing microcystin-LR. Food and Chemical Toxicology 2008, 46, 502–507. [CrossRef]
- Tonon, T.; Lonvaud-Funel, A. Metabolism of arginine and its positive effect on growth and revival of Oenococcus oeni: degradation of arginine by Oenococcus oeni. Journal of Applied Microbiology 2000, 89, 526–531. [CrossRef]
- Bech-Terkilsen, S.; Westman, J.O.; Swiegers, J.H.; Siegumfeldt, H. Oenococcus oeni , a species born and moulded in wine: a critical review of the stress impacts of wine and the physiological responses. Australian Journal of Grape and Wine Research 2020, 26, 188–206. [CrossRef]
- Conway, T.; K., G. Microarray expression profiling: capturing a genome-wide portrait of the transcriptome: genome-wide portrait. Molecular Microbiology 2003, 47, 879–889. [CrossRef]
- Laalami, S.; Zig, L.; Putzer, H. Initiation of mRNA decay in bacteria. Cell. Mol. Life Sci. 2014, 71, 1799–1828. [CrossRef]
- Morton, D.S.; Oliver, J.D. Induction of carbon starvation-induced proteins in Vibrio vulnificus. Appl Environ Microbiol 1994, 60, 3653–3659. [CrossRef]
- McGovern, V.P.; Oliver, J.D. Induction of cold-responsive proteins in Vibrio vulnificus. J Bacteriol 1995, 177, 4131–4133. [CrossRef]
- Federighi, M.; Tholozan, J.L.; Cappelier, J.M.; Tissier, J.P.; Jouve, J.L. Evidence of non-coccoid Viable but Non-culturable Campylobacter jejuni cells in microcosm water by direct viable count, CTC-DAPI double staining, and scanning electron microscopy. Food Microbiology 1998, 15, 539–550. [CrossRef]
- Yaron, S.; Matthews, K.R. A Reverse Transcriptase-Polymerase Chain Reaction assay for detection of Viable Escherichia coli O157:H7: investigation of specific target genes. J Appl Microbiol 2002, 92, 633–640. [CrossRef]
- Fischer-Le Saux, M.; Hervio-Heath, D.; Loaec, S.; Colwell, R.R.; Pommepuy, M. Detection of cytotoxin-hemolysin mRNA in Nonculturable populations of environmental and clinical Vibrio vulnificus strains in artificial seawater. Appl Environ Microbiol 2002, 68, 5641–5646. [CrossRef]
- Rahman, I.; Shahamat, M.; Kirchman, P.A.; Russek-Cohen, E.; Colwell, R.R. Methionine uptake and cytopathogenicity of Viable but Nonculturable Shigella dysenteriae type 1. Appl Environ Microbiol 1994, 60, 3573–3578. [CrossRef]
- Day, A.P.; Oliver, J.D. Changes in membane fatty acid composition during entry of Vibrio vulnificus into the Viable But Non Culturable state. 2004, 42, 69–73.
- Signoretto, C.; Lleò, M.D.M.; Canepari, P. Modification of the Peptidoglycan of Escherichia coli in the Viable But Nonculturable State. Current Microbiology 2002, 44, 125–131. [CrossRef]
- Signoretto, C.; Del Mar Lleò, M.; Tafi, M.C.; Canepari, P. Cell wall chemical composition of Enterococcus faecalis in the Viable but Nonculturable state. Appl Environ Microbiol 2000, 66, 1953–1959. [CrossRef]
- Mascher, T.; Helmann, J.D.; Unden, G. Stimulus perception in bacterial signal-transducing histidine kinases. Microbiol Mol Biol Rev 2006, 70, 910–938. [CrossRef]
- Friedrich, T.; Scheide, D. The respiratory complex I of bacteria, archaea and eukarya and its module common with membrane-bound multisubunit hydrogenases. FEBS Letters 2000, 479, 1–5. [CrossRef]
- Postnikova, O.A.; Shao, J.; Mock, N.M.; Baker, C.J.; Nemchinov, L.G. Gene expression profiling in Viable but Nonculturable (VBNC) cells of Pseudomonas syringae pv. Syringae. Front. Microbiol. 2015, 6. [CrossRef]
- Hua J.; Ho B. Is the coccoid form of Helicobacter pylori Viable? Microbios 1996, 87, 103–112.
- Tsugawa, H.; Suzuki, H.; Nakagawa, I.; Nishizawa, T.; Saito, Y.; Suematsu, M.; Hibi, T. Alpha-kKetoglutarate oxidoreductase, an essential salvage enzyme of energy metabolism, in coccoid form of Helicobacter pylori. Biochemical and Biophysical Research Communications 2008, 376, 46–51. [CrossRef]
- Asakura, H.; Panutdaporn, N.; Kawamoto, K.; Igimi, S.; Yamamoto, S.; Makino, S. Proteomic characterization of enterohemorrhagic Escherichia coli O157:H7 in the oxidation-induced Viable but Non-Culturable state. Microbiology and Immunology 2007, 51, 875–881. [CrossRef]
- Abe, A.; Ohashi, E.; Ren, H.; Hayashi, T.; Endo, H. Isolation and characterization of a cold-induced nonculturable suppression mutant of Vibrio vulnificus. Microbiological Research 2007, 162, 130–138. [CrossRef]
- Ramamurthy, T.; Ghosh, A.; Pazhani, G.P.; Shinoda, S. Current perspectives on Viable but Non-Culturable (VBNC) pathogenic bacteria. Front. Public Health 2014, 2. [CrossRef]
- Lai, C.-J.; Chen, S.-Y.; Lin, I.-H.; Chang, C.-H.; Wong, H. Change of protein profiles in the induction of the Viable but Nonculturable State of Vibrio parahaemolyticus. International Journal of Food Microbiology 2009, 135, 118–124. [CrossRef]
- Hung, W.; Jane, W.-N.; Wong, H. Association of a d-alanyl- d-alanine carboxypeptidase gene with the formation of aberrantly shaped cells during the induction of Viable but Nonculturable Vibrio parahaemolyticus. Appl Environ Microbiol 2013, 79, 7305–7312. [CrossRef]
- Wang, H.-W.; Chung, C.-H.; Ma, T.-Y.; Wong, H. Roles of alkyl hydroperoxide reductase subunit C (AhpC) in Viable but Nonculturable Vibrio parahaemolyticus. Appl Environ Microbiol 2013, 79, 3734–3743. [CrossRef]
- Mongkolsuk, S.; Whangsuk, W.; Vattanaviboon, P.; Loprasert, S.; Fuangthong, M. A Xanthomonas alkyl hydroperoxide reductase subunit C (ahpC) mutant showed an altered peroxide stress response and complex regulation of the compensatory response of peroxide detoxification enzymes. J Bacteriol 2000, 182, 6845–6849. [CrossRef]
- Asakura, H.; Ishiwa, A.; Arakawa, E.; Makino, S.; Okada, Y.; Yamamoto, S.; Igimi, S. Gene expression profile of Vibrio cholerae in the cold stress-induced Viable but Non-culturable state. Environmental Microbiology 2007, 9, 869–879. [CrossRef]
- Krebs, S.J.; Taylor, R.K. Nutrient-dependent, rapid transition of Vibrio cholerae to coccoid morphology and expression of the toxin co-regulated pilus in this form. Microbiology 2011, 157, 2942–2953. [CrossRef]
- Patrone, V.; Campana, R.; Vallorani, L.; Dominici, S.; Federici, S.; Casadei, L.; Gioacchini, A.M.; Stocchi, V.; Baffone, W. CadF expression in Campylobacter jejuni strains incubated under low-temperature water microcosm conditions which induce the Viable but Non-Culturable (VBNC) state. Antonie van Leeuwenhoek 2013, 103, 979–988. [CrossRef]
- Boaretti, M.; Del Mar Lleo, M.; Bonato, B.; Signoretto, C.; Canepari, P. Involvement of rpoS in the survival of Escherichia coli in the Viable but Non-Culturable state. Environ Microbiol 2003, 5, 986–996. [CrossRef]
- Maddocks, S.E.; Oyston, P.C.F. Structure and function of the LysR-type transcriptional regulator (LTTR) family proteins. Microbiology 2008, 154, 3609–3623. [CrossRef]
- Grove, A. MarR family transcription factors. Current Biology 2013, 23, R142–R143. [CrossRef]
- López-Lara, L.I.; Pazos-Rojas, L.A.; López-Cruz, L.E.; Morales-García, Y.E.; Quintero-Hernández, V.; De La Torre, J.; Van Dillewijn, P.; Muñoz-Rojas, J.; Baez, A. Influence of rehydration on transcriptome during resuscitation of desiccated Pseudomonas putida KT2440. Ann Microbiol 2020, 70, 54. [CrossRef]
- Gupte, A.R.; De Rezende, C.L.E.; Joseph, S.W. Induction and resuscitation of Viable but Nonculturable Salmonella enterica serovar typhimurium DT104. Appl Environ Microbiol 2003, 69, 6669–6675. [CrossRef]
- Du, M.; Chen, J.; Zhang, X.; Li, A.; Li, Y. Characterization and resuscitation of Viable but Nonculturable Vibrio alginolyticus VIB283. Arch Microbiol 2007, 188, 283–288. [CrossRef]
- Wong, H.-C.; Liu, S.-H. Characterization of the low-salinity stress in Vibrio vulnificus. Journal of Food Protection 2008, 71, 416–419. [CrossRef]
- Bates, T.C.; Oliver, J.D. The Viable but Non Culturable state of Kanagawa positive and negative strains of Vibrio parahaemolyticus. J. Microbiol 2004, 42, 74–79.
- Steinert, M.; Emödy, L.; Amann, R.; Hacker, J. Resuscitation of Viable but Nonculturable Legionella pneumophila Philadelphia JR32 by Acanthamoeba castellanii. Appl Environ Microbiol 1997, 63, 2047–2053. [CrossRef]
- García, M.T.; Jones, S.; Pelaz, C.; Millar, R.D.; Abu Kwaik, Y. Acanthamoeba polyphaga Resuscitates Viable Non-culturable Legionella pneumophila after disinfection. Environmental Microbiology 2007, 9, 1267–1277. [CrossRef]
- Sussman, M.; Loya, Y.; Fine, M.; Rosenberg, E. The marine fireworm Hermodice carunculata as a winter reservoir and spring-summer vector for the coral-bleaching pathogen Vibrio shiloi. Environ Microbiol 2003, 5, 250–255. [CrossRef]
- Cappelier, J.M.; Besnard, V.; Roche, S.M.; Velge, P.; Federighi, M. Avirulent Viable But Non Culturable cells of Listeria monocytogenes need the presence of an embryo to be recovered in egg yolk and regain virulence after recovery. Vet. Res. 2007, 38, 573–583. [CrossRef]
- Mukamolova, G.V.; Kaprelyants, A.S.; Kell, D.B. Secretion of an antibacterial factor during resuscitation of dormant cells in Micrococcus luteus cultures held in an extended stationary phase. Antonie van Leeuwenhoek 1995, 67, 289–295. [CrossRef]
- Mukamolova, G.V.; Kaprelyants, A.S.; Young, D.I.; Young, M.; Kell, D.B. A bacterial cytokine. Proc. Natl. Acad. Sci. U.S.A. 1998, 95, 8916–8921. [CrossRef]
- Nikitushkin, V.D.; Demina, G.R.; Shleeva, M.O.; Guryanova, S.V.; Ruggiero, A.; Berisio, R.; Kaprelyants, A.S. A product of RpfB and RipA joint enzymatic action promotes the resuscitation of dormant mycobacteria. The FEBS Journal 2015, 282, 2500–2511. [CrossRef]
- Zhao, R.; Chen, J.; Wang, Y.; Li, Y.; Kong, X.; Han, Y. Proteolytic activity of Vibrio harveyi YeaZ is related with resuscitation on the Viable but Non-Culturable State. Letters in Applied Microbiology 2020, 71, 126–133. [CrossRef]
- Mukamolova, G.V.; Turapov, O.A.; Young, D.I.; Kaprelyants, A.S.; Kell, D.B.; Young, M. A family of autocrine growth factors in Mycobacterium tuberculosis: autocrine growth factors in Mycobacterium tuberculosis. Molecular Microbiology 2002, 46, 623–635. [CrossRef]
- Freestone, P.P.E.; Haigh, R.D.; Williams, P.H.; Lyte, M. Stimulation of bacterial growth by heat-stable, norepinephrine-induced autoinducers. FEMS Microbiology Letters 1999, 172, 53–60. [CrossRef]
- Woolf, P.D. The catecholamine response to multisystem trauma. Arch Surg 1992, 127, 899. [CrossRef]
- Reissbrodt, R.; Rienaecker, I.; Romanova, J.M.; Freestone, P.P.E.; Haigh, R.D.; Lyte, M.; Tschäpe, H.; Williams, P.H. Resuscitation of Salmonella enterica serovar typhimurium and enterohemorrhagic Escherichia coli from the Viable but Nonculturable state by heat-stable enterobacterial autoinducer. Appl Environ Microbiol 2002, 68, 4788–4794. [CrossRef]
- Imazaki, I.; Nakaho, K. Temperature-upshift-mediated revival from the sodium-pyruvate-recoverable Viable but Nonculturable state induced by low temperature in Ralstonia solanacearum: linear regression analysis. J Gen Plant Pathol 2009, 75, 213–226. [CrossRef]
- Zhao, F.; Bi, X.; Hao, Y.; Liao, X. Induction of Viable but Nonculturable Escherichia coli O157:H7 by high pressure CO2 and its characteristics. PLoS ONE 2013, 8, e62388. [CrossRef]
- Abisado, R.G.; Benomar, S.; Klaus, J.R.; Dandekar, A.A.; Chandler, J.R. Bacterial quorum sensing and microbial community interactions. mBio 2018, 9, e02331-17. [CrossRef]
- Hassett, D.J.; Ma, J.; Elkins, J.G.; McDermott, T.R.; Ochsner, U.A.; West, S.E.H.; Huang, C.; Fredericks, J.; Burnett, S.; Stewart, P.S.; et al. Quorum sensing in Pseudomonas aeruginosa controls expression of catalase and superoxide dismutase genes and mediates biofilm susceptibility to hydrogen peroxide. Molecular Microbiology 1999, 34, 1082–1093. [CrossRef]
- Liao, H.; Zhong, X.; Xu, L.; Ma, Q.; Wang, Y.; Cai, Y.; Guo, X. Research in Microbiology 2019, 170, 65–73. [CrossRef]
Figure 1.
CFU/mL (log) of Pseudomonas putida KT2440 under desiccation stress during 18 days after the beginning of desiccation (DABD), in presence (green bars) and absence (blue bars) of trehalose. Adapted from Pazos-Rojas et al., 2019 [4].
Figure 1.
CFU/mL (log) of Pseudomonas putida KT2440 under desiccation stress during 18 days after the beginning of desiccation (DABD), in presence (green bars) and absence (blue bars) of trehalose. Adapted from Pazos-Rojas et al., 2019 [4].
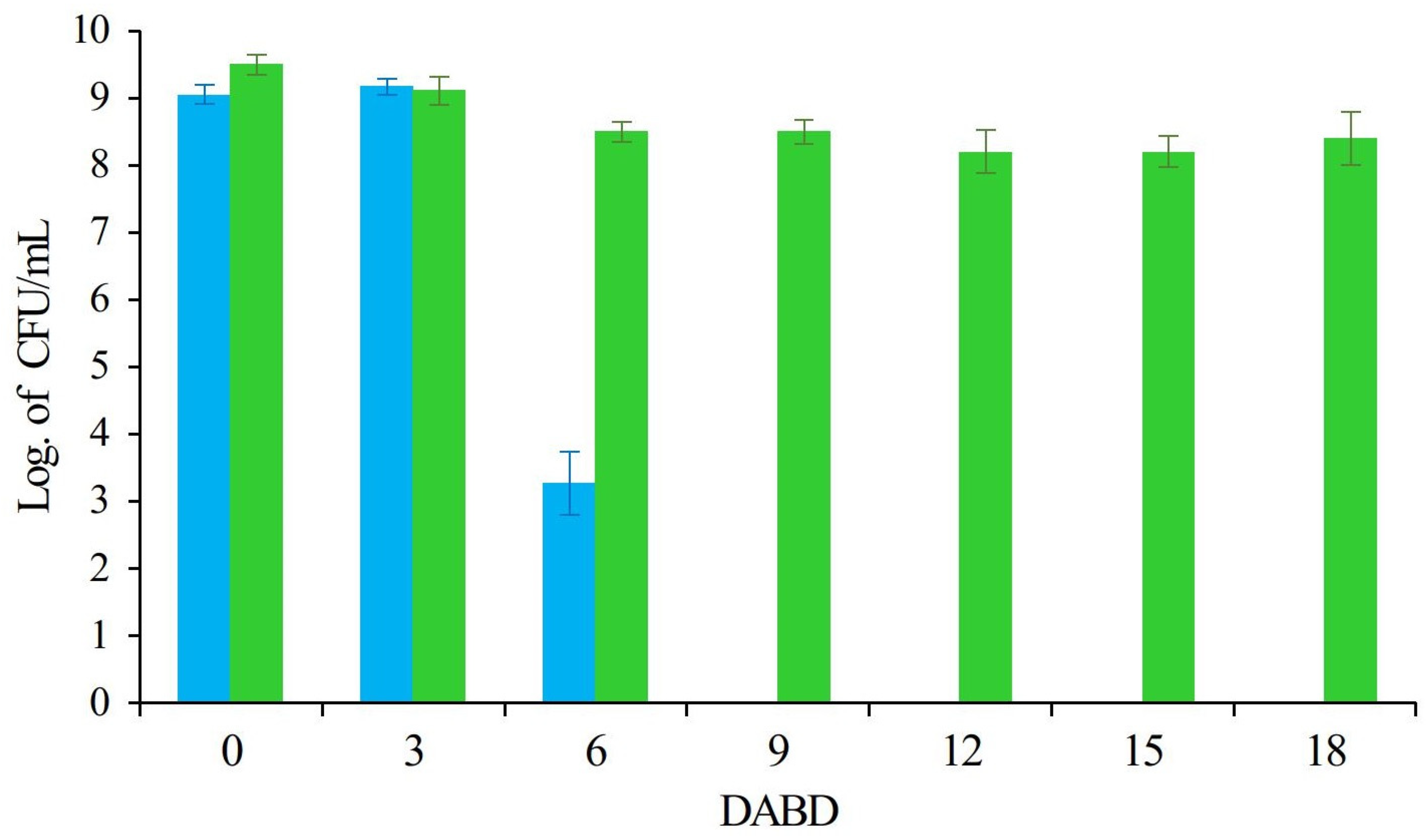
Figure 2.
Phylogenetic tree of beneficial bacterial species entering the VBNC state. Phylogenetic trees were constructed by the Neighbour-joining method [68] using ClustalX, Bioedit and Mega 4 software. A bootstrap confidence analysis was applied on 1000 replicates to determine the reliability of the topology obtained.
Figure 2.
Phylogenetic tree of beneficial bacterial species entering the VBNC state. Phylogenetic trees were constructed by the Neighbour-joining method [68] using ClustalX, Bioedit and Mega 4 software. A bootstrap confidence analysis was applied on 1000 replicates to determine the reliability of the topology obtained.
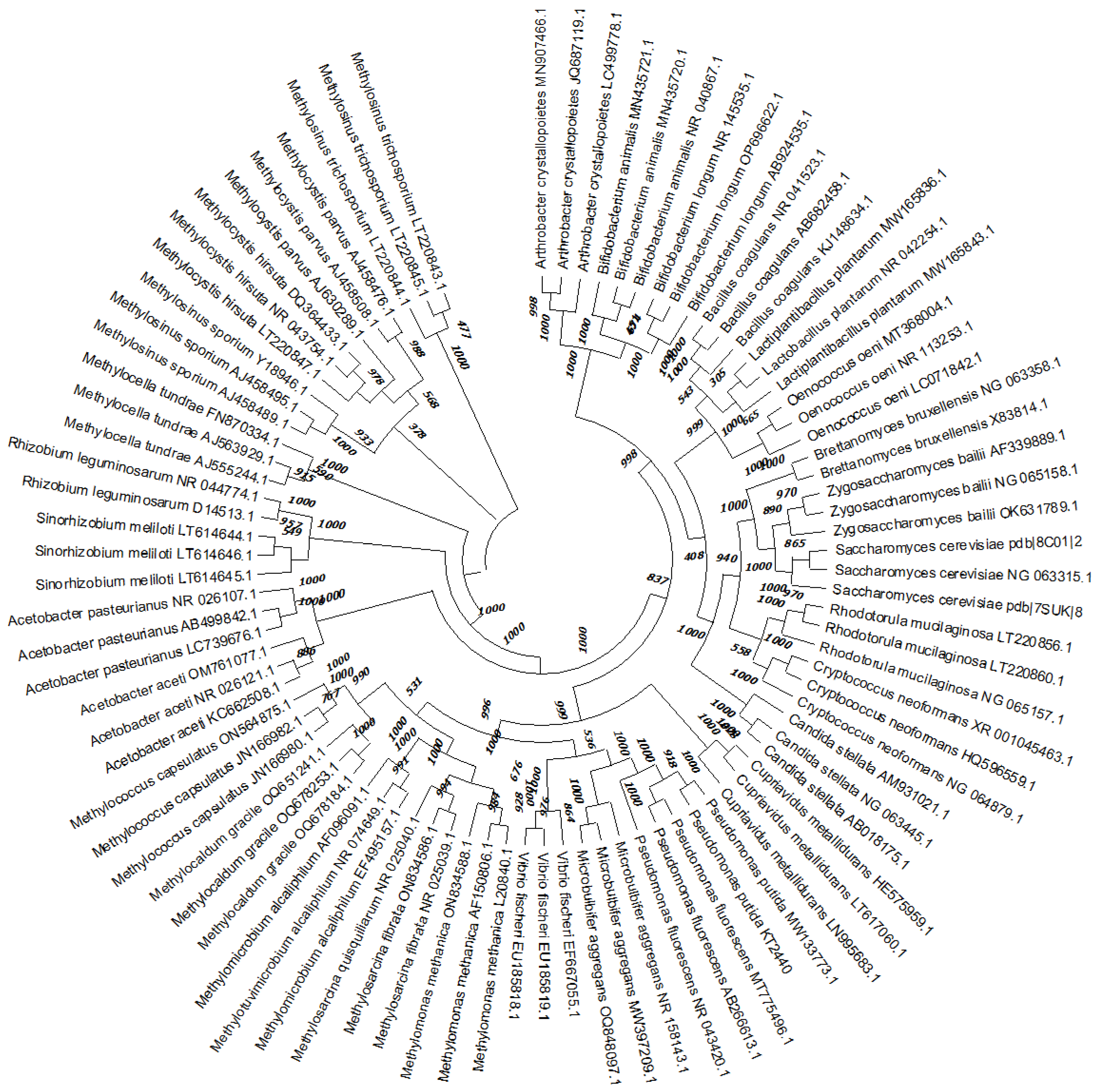
Table 1.
Types of stress that can induce VBNC state in bacteria.
| Type of stress | Example of microorganism | Reference | |
|---|---|---|---|
| Physical parameters | Temperature | Vibrio cholerae incubated in a range of 4 to 6°C | [7] |
| Visible solar and UV radiation | Escherichia coli in microcosm of river water under visible light irradiation | [8] | |
| Osmolarity | E. coli and Salmonella typhimurium in freshwater and seawater | [9] | |
| Chemical parameters | Organic matter | ||
| Nutritional effects. | Streptococcus pyogenes carbon and phosphorus limitation in culture during stationary phase | [10] | |
| Humic acids | E. coli in presence of humic acids significantly decreases the entrance to VBNC status in fresh water but not in seawater | [9] | |
| Aeration | Campylobacter jejuni enters faster into the VBNC state in microcosmos incubated under agitation (3 days), compared to immobile microcosmos (10 days) | [11] | |
| Drying | Sinorhizobium meliloti part of the population enters into the VBNC state with 22% humidity | [12] | |
| P. putida KT2440 enters into the VBNC state under desiccation conditions at 30°C and 50% relative humidity | [4] | ||
| Biocidal agents | Hypochlorite (100ppm) | Legionella pneumophila was detected in culture medium treated with hypochlorite for 10 minutes | [13] |
| Cupper (0.6-1mg/mL) | E. coli ETEC in buffered water at 4 °C produced VBNC cells that maintained their enterotoxigenic activity when reactivated. | [14] | |
| Alum KAI(S04)2 | V. cholerae O1 in artificial seawater with alum, a significant number of cells lost their culturable capacity while keeping viability | [15] | |
| Quinolone and ciprofloxacin | E. coli, exposed to 10-100 times minimum inhibitory concentration (MIC), lost the culturable capacity | [16] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



