Submitted:
18 November 2023
Posted:
21 November 2023
You are already at the latest version
Abstract
Terrestrial algae are the group of photosynthetic organisms, which can survive in extreme conditions. pH is one of the most important factor, influencing on distribution of algae, both in aquatic and terrestrial ecosystems. The influence of different pH levels on cell volume and other morphological characters of authentic and reference strains of Chlorella vulgaris, Bracteacoccus minor, Pseudoccomyxa simplex, Chlorococcum infusionum, and Vischeria magna were studied. Chlorella vulgaris and Pseudoccomyxa simplex were the most resistant species, retaining its morphology in range of pH 4-11,5. The change of pH towards acidic and alkaline levels caused the increase of volume of Pseudoccomixa simplex and Vischeria magna cells according to polynomial regression model. Volume of Chlorella vulgaris cells increased from low to high pH according to linear regression model. Changes of pH levels did not have a significant impact on volume of Bracteacoccus minor and Chlorococcum infusionum cells. Low and high levels of pH caused thickening of cell walls of Chlorococcum infusionum and increase of oil containing substances in Vischeria magna and Bracteacoccus minor cells. Our study revealed a high resistance of studied species to extreme level of pH, which allow recommend these strains for broader use in biotechnology and natures protection.
Keywords:
adaptation
; authentic strains
; cell wall
; discoloration
; protoplast
; tolerance
; Bracteacoccus minor
; Chlorella vulgaris
; Chlorococcum infusionum
; Pseudococcomyxa simples
; Vischeria magna
1. Introduction
The concentration of hydrogen ions has a fundamental role in determining the boundaries of the existence of living matter. Most living organisms exist at pH from 4 to 9. The limiting concentration of hydrogen ions, above and below which currently known organisms stop growing and multiplying, fluctuate within pH from 1 to 11 [1]. If the pH value does not approach its extreme values, communities can regulate changes in this factor by changing the intensity of respiration and the functioning of the body's enzyme systems [2].
There are numerous data in the literature on the effect of pH on various groups of organisms, including algae. Algae have a wide range of tolerance to pH, but the limits of resistance to this factor in representatives of various systematic groups are not the same [3,4,5] The influence of pH on the rate of vegetative division [6], zoospores differentiation [7], the amount of available iron in the medium [8,9], the ratio of nitrogen and carbon, autotrophic and heterotrophic nutrition [10,11,12,13,14,15], proteins, lipids, carbohydrates, fatty acids, chlorophyll content and biomass [16,17], algae assemblages in water and terrestrial ecosystems [18,19,20], algal cell membrane permeability [21], the morphology of algae [22] was established.
It should be noted that the vast majority of algological studies (including environmental ones) are devoted to the investigation of aquatic algae, while terrestrial forms remain extremely insufficiently studied. At the same time, it is well known, that terrestrial algae are highly resistant to extreme levels of ecological factors. This fact allows to use it as a model objects for studying the mechanisms of resistance of living organisms [23], including extreme pH values.
In this connection the study of the autecology of the most widespread species, which are detected in various habitats around the world. Among this species are Bracteacoccus minor (Schmidle ex Chodat) Petrová, Chlorococcum infusionum (Schrank) Meneghini, Chlorella vulgaris Beijerinck, Pseudococcomyxa simplex (Mainx) Fott, and Vischeria magna (J. B. Petersen) Kryvenda, Rybalka, Wolf & Friedl is very important.
Bracteacoccus minor is widely distributed in the terrestrial habits [24,25,26,27] and belongs to the cosmopolitan [28]. In addition, this species is often found in extreme habitats. For example, it was widely distributed in spruce forests that were exposed to acid rain in the Czech Republic [29].
Chlorococcum infusionum is the type species of the genus Chlorococcum [40]. Chlorococcum infusionum has been found both in soil and stagnant water bodies [24]. It is assumed, that this species belongs to cosmopolitans, it was found both in neutral and acid soils, as well as on porous acid rocks [30]. Chlorococcum infusionum was common in all zones in almost all types of soils – from waterlogged to high-altitude desert, and from acid podzolic to carbonate deeply saline [31,32,33,34]. This species detected in polluted urban soils [35]. Chlorococcum infusionum had a high indicator value for the soils of spruce forests in the background areas of the middle and southern taiga [36].
Chlorella vulgaris is the type species of the genus Chlorella [40]. This species is common in reservoirs of various types, soil, terrestrial substrates, and belongs to the cosmopolitan [24,25,28,34,37]. Chlorella vulgaris is widely distributed in soils and in subaerial habitats. This species has a large number of local physiological and ecological races living in different conditions [30]. Chlorella vulgaris was identified the in salt marshes, acidic and polluted soils [38]. This species remained the only one taxa in conditions of severe oil pollution [39] and severe acidification during precipitation near metallurgical enterprises [38]. However, it should be noted, that most of the findings of this species need to be verified, since it is quite common practice to assign any small green spherical algae to Chlorella vulgaris [30].
Pseudococcomyxa simplex is the type species of the genus Pseudococcomyxa [40]. Pseudococcomyxa simplex is one of the widespread species of terrestrial algae [24, 25, 28, 32, 33. 41]. It has a wide ecological plasticity, for example, these algae are highly resistant to low pH. The species was found in very acidic soil with a pH <3 in Italy [28]. In addition, Pseudococcomyxa simplex was found in all the studied sites during the investigation of the of spruce forests algae exposed to acid rain in Northern Bohemia (Czech Republic) [29].
Vischeria magna has a wide distribution around the world, it has been found in the North America [42,43], Europa [25,29,33], Africa [26], Asia [36]. This species was also found in the territories with extreme habitat conditions, for example, different types of salt marshes. Vischeria magna was detected in the crustal salt flats in the chernozem, chestnut and brown zones, in the middle salt flats – the chernozem and chestnut zones, in the deep salt flats – the chernozem, chestnut, brown zones, in the rejuvenated salt flats – the chernozem zone, in the meadow salt flats – the chernozem zone [44]. Besides, this alga was found in the acid rain fall zone in spruce forests in the Czech Republic and Germany [29].
Analysis of the literature data on the influence of ecological factors on microscopic algae revealed two main groups of criteria, used to assess the degree of effect:
1. Morphological criteria: cell size and shape [22,45], cell envelope ultrastructure [46], chloroplast morphology and coloration [45], cytoplasm, membrane and mitochondria structures [47], cell wall integrity [48].
2. Physiological criteria: growth reactions [49,50,51,52,53,54], photosynthesis intensity [55], carbohydrate and protein content [16], pigment composition and assimilation [56], chemical composition [17], chlorophyll “a” and “b” fluorescence [57], transmembrane electrochemical gradient [58], fatty acid production [59], cell division, and biological volume (cell volume) [45].
The term "biological volume" or "volume" is widely used to assess the functioning of aquatic forms of algae. It reflects both morphological and physiological features of living organisms. It has been shown that picoplankton algae with a very small sizes (< 2 microns in diameter) have a larger ratio of surface area to volume, which ensures effective absorption of nutrients and photons [60]. This indicator is widely used to characterize the state of aquatic bacteria [61], dinoflagellates [62], and infusoria [63], as well as microscopic algae [22,64,65]. It has been established, that biological volume plays a very important role in the ecology of algae. For example, in marine diatoms cell volume is associated with metabolic rate [67,68], growth rate [69], photosynthetic capacity [67], respiration rate [70,71], as well as asexual reproduction [72]. In addition, cell volume influence on nutrient uptake [73], light absorption [74], and primary production in marine ecosystems [71,75]. A correlation between the evolution of the genome size and the volume of diatoms was established [76]. It was stressed, that that the cell volume of diatoms is an important component of the global carbon cycle.
The studies of nitrogen uptake depending on the size of micro- and macroalgae revealed that microalgae absorbed nitrogen per unit of biomass much faster at both high and low concentrations of the element. In addition, microalgae had a greater degree of affinity for nitrogen than macroalgae [77]. These differences in absorption rates among small and large algae were usually attributed to size-specific processes depending on the relative surface area (SA:V). Regression analysis confirmed that size-dependent differences in kinetic parameters can be related to the relative surface area for a wide range of algae of different sizes.
Biological volume is closely related to the ratio of surface area to volume (S/V ratio). Under stress conditions organisms try to reduce the ratio of surface area to volume. The most advantageous in this respect is the spherical shape of the cells [78]. So it is not surprising, that many terrestrial microscopic algae living in extreme environmental conditions, have a spherical shape. In the study of morphological diversity, evolutionary relatedness, and size constraints of freshwater algae and cyanobacteria was proposed, that “the morphospace may serve as a proxy for an ecospace” and in future this morphospace could be used for demonstration of the current ecological processes [79].
The aim of the study was estimation of resistance of algae with wide geographical distribution to pH and influence of pH levels on morphology and biological volume of this taxa.
2. Results
The peculiarities of the effect of pH on the cell morphology and volume of each species had its own characteristics, which are described below.
2.1. Bracteacoccus minor
At pH 2-2.5 complete discoloration and destruction of Bracteacoccus minor cells was observed. At pH 3 part of the cells lost the green color (Figure 1B). In the pH range 4-10 the morphology of algae cells differed little from the control (Figure 1A–F), and the appearance of large pear-shaped cells was observed. It should be noted, that such cells were often observed under normal conditions. In pH 3.5-5 some cells contain orange oil droplets (Figures 1B and 2). At pH 10.5-11 discoloration of most of the cells was observed, and at pH 12-13.5, all Bracteacoccus minor cells were destroyed (Figure 1E,F).
With growth pH an increase in the volume of algae cells according to the linear regression model (r: 0.066995; Wilks’ lambda: 0.9955; F: 9.209; p (regr): 0.002439) was dtected (Figure 3). However, these changes were very insignificant, which is evidence of the relative stability of Bracteacoccus minor morphology to extreme levels of environmental factors.
2.2. Chlorococcum infusionum
In the acidic pH from 2 to 3.5 loss of green color and destruction of Chlorococcum infusionum cells were observed (Figure 4 A,E). At pH 4-9.5 the algae retain the usual morphology, which did not differ from the morphology of algae in the control variant (Figure 4A). At a pH of 10-10.5 the destruction of part of the cells, which manifested in compression of the protoplast, its departure from the cell wall and strong granulation was observed (Figure 4E). In the alkaline rahge of pH 11-13.5 all cells were discolored with a completely destroyed protoplast (Figure 3F).
With an increase of pH a decrease the of Chlorococcum infusionum cells volume was detected. This changes were described by a linear regression model (r: -0.07645; Wilks’ lambda: 0.9942; F: 11.13; p (regr): 0.0008664) (Figure 5), however, as in the experiments with Bracteacoccus minor, this influence was very insignificant.
Extreme pH values caused thickening of the mucilage, surrounding Chlorococcum infusionum cells. These cahnges corresponded to the polynomial regression model (χ2: 281.25; AIC: -2.0218; R2: 0.052168; F: 58.617; p (regr): 1.6547E-25) (Figure 6). Possibly, this phenomenon is a protective reaction of algae from the damaging effects of acidic and alkaline pH values.
2.3. Chlorella vulgaris
At pH 2-3 complete discoloration of Chlorella vulgaris cells was observed (Figure 7A,B). At a pH of 3.5 about 30% of the cells lost the green color, the remaining cells had usual morphology (Figure 7B). In the pH range from 4 to 11.5 the algae retained morphological features - green color, cup-shaped cloroplast with the pyrenoid and cell wall integrity (Figure 7E,F). At pH 12 discoloration of all Chlorella vulgaris cells were observed.
With a change in pH an increase of the volume of Chlorella vulgaris cells was noted according to the linear regression model (r: 0.32475; Wilks’ lambda: 0.8945; F: 305.7; p (regr): 8.51E-65) (Figure 8).
Our revealed a high resistance of Chlorella vulgaris to acid and alkaline pH, which is consistent with information about the wide distribution of this species.
2.4. Pseudococcomyxa simplex
At pH 2 loss of green color and "wrinkling" of Pseudococcomyxa simplex cells was noted (Figure 9, A, B). At pH 2.5, about 50% of the cells were broadly ellipsoid in their shape (Figure 9C). The appearance of larger cells was also recorded at a pH of 8.5-10.5 (Figure 9E). At a pH of 3.5 about 30% of the cells were discolored (Figure 9D). In the pH range of 4-11.5 the morphology of the algae was almost the same with the control variant (Figure 9A–E). At pH 12 complete discoloration of the algae cells was observed (Figure 9F).
With a change in pH a change in the volume of algae cells, described by a polynomial regression model, was detected. The reliability of this change was confirmed by the value of p (regr), which was significantly less than 0.05 (χ2: 1,719E05; AIC: 4,572; R^2: 0.10593; F: 105.62; p (regr): 4,4585 E-44) (Figure 10).
It should be noted that at low and high pH Pseudococcomyxa simplex cells became similar to cells of «Avernensia» form [80,81], differing from Pseudococcomyxa by more rounded cells. Perhaps, «Avernensia» is the morphological form of Pseudococcomyxa in insufficient nature conditions. This fact should be taken into account during the algae identification. The discovered phenomenon once again demonstrates the morphological variability, often observed in eukaryotic algae.
2.5. Vischeria magna
At pH 2 complete discoloration of the Vischeria magna culture was observed. At pH 2.5 the algae cells were either completely discolored or bright orange (Figure 11A,B). Bright orange cells were also observed at pH 3 (Figure 11B). In the pH range 3.5-11 the morphology of most algae cells was the same with the control variant (Figure 11A), and zoospores were observed at in this pH range with the maximum at pH 3.5 (Figure 11D).
At high pH values from 11.5 to 12, 80% of Vischeria magna cells were discolored and destroyed (Figure 11E,F). At a pH of 12.5 -13.5 the culture was completely discolored.
The experiments revealed an increase of cell volume in the acid and alkaline pH range according to the polynomial regression model (χ2:1,3735E07; AIC: 8,6522; R^2: 0.28156; F: 471.47; p (regr): 1,7153E-173) (Figure 12).
The study of pH influence on morphology and cell volume of Bracteacoccus minor, Chlorococcum infusionum, Chlorella vulgaris, Pseudococcomyxa simplex, and Vischeria magna revealed, that algae retain their morphological features at pH 3.5-4 and 9.5-11.5 (Table 1). Low and high pH levels caused mostly discoloration and destruction of cells (Table 1), but specific reaction of the species were also observed (Table 1). Extreme pH levels increase mucilage production around Chlorococcum infusionum cells (Figure 4F, Table 1). Low pH caused also the appearance of broadly ellipsoid and compressed cells of Pseudococcomyxa simplex cells (Table 1, Figure 9B), and orange granules in Vischeria magna cells (Table 1, Figure 11B).
3. Discussion
Our investigations revealed high tolerance of investigated algae species to low and high pH levels, which retain their basic morphological features in the range of pH from 3.5-5 till 9.5-11.5 (Table 1). For some genus and species, these results correspond to previously published data. For example, Chlorella pyrenoidosa and Chlorella ellipsoidea can grow in media with pH 3.5 and 2 correcspondingly, although the upper limit for it growth is about pH 10 [82].
The morphological disorders of algae cells at low and high pH are associated with the damaging effect of H+ and OH− ions. Under the conditions of very low pH cells experience stress because it have to maintain a neutral pH level in the cytoplasm, since H+ ions continuously penetrate plasma membranes [83]. Besides, low pH values can damage cell walls as a result of the weakening of hydrogen bonds in molecules, which can lead to an uncontrolled increase in cell size [84]. It was established, that acidification can influence on metal bioavailability by proton inhibition of facilitated metal uptake and by impacting on membrane permeability [21]. It is known, that at high pH values many elements necessary for organisms, such as Fe2+, Ca2+, Mg2+, Zn2+, become insoluble and precipitate as carbonates, hydroxides or phosphates. The concentration of hydrogen ions in the environment can affect the equilibrium of electric charges on the cell surface, increasing the total positive charge at low pH values or the total negative charge at high [1]. Most likely, alkaliphilic algae, like other microorganisms, exclude hydroxyl groups, or, conversely, retain hydrogen ions [1].
Algae strains, resistant to extreme pH, have a great potential for using in the biotechnology. pH is a critical parameter influencing on the mass transfer rate of acidic gases like H2S and CO2 in conventional gas-liquid contactors [85,86]. At pH>9 contaminants in the bigas die, while resistant to pH algae strains survive [87,88,89]. The use of carbonated alkaline culture media in algal-bacterial photobioreactors support a long term effective biogas supporting [89].
The resistance of algae to wade range of pH was discussed in previous investigations [90,91,92,93]. Euglena mutabilis, typical species in acidic environments, achieve the maximal growth at pH 3.0-4.0 in a growth range at 2.0–9.0 pH [90]. For representatives of Mougeotia, in laboratory conditions optimal pH was 8.0, in nature – pH 5.2 [91]. Chlamydomonas acidophila can grow at pH 1.5–7.0 (Gerloff-Elias et al., 2005). The optimum pH of three axenic strains of Chloromonas tughillensis was at 3.0–7.0 pH, and of three non-axenic strains of Chloromonas chenangoensis – at pH 3.0–8.0 [93].
Some algae have tolerance to acidic envirinmets [45,84,94,95,96]. These algae iclude taxa, investigated in our study. For example, in investigations of the algoflora of the Krušne hory Mts forests in Northern Bohemia (Czech Republic), polluted by acid rain, Bracteacoccus minor and Pseudococcomyxa simplex in all investigated sites were found. Vischeria magna and Chlorella vulgaris were also very frequent in the studied area [29]. Pseudococcomyxa simplex and represenatives of genus Chlorella – Chlorella saccharophila and Chlorella protococcoides – were attributed to acidiphilic and acidotolerant algae [84].
However, it should be noted, that the floristic lists, containing this data, are not confirmed by molecular-genetic methods. Therefore, information about the distribution of these species, which are difficult to determine, should be interpreted with the caution.
The data about alcaliphilic and alcalitolerant algae are also known. It was found, that eutrophic algae and cyanobacteria (Gonatozygon monotaenium, Gloeocapsa sp.) can grow at pH values above 9 [97]. Representative of the genus Klebsormidium – Klebsormidium acidophilum and Klebsormidium dissectum – were detected at pH 3.0 and pH 4.8–6.2 accordingly [98]. High pH (8-8.5) together with the high salinity promote the growth oh the green filamentous algae Chaetomorpha valida [99].
Our studies also revealed a high resistance of Chlorella vulgaris to acid and alkaline pH, which is consistent with information about the ubiquism of this species. As already noted above, Chlorella vulgaris was found in acidic soils [38]. But according to earlier investigations, the maximum growth of Chlorella vulgaris was observed at a pH of 6.31-6.84 [100]. The optimal pH for Chlorella sorokiniana DOE1412 is also approximately 6.0 for cell growth and lipid production [101].
The data about resistance of Chlorococcum infusionum to different pH values are inconsistent. Some studies revealed, that Chlorococcum infusionum preferred neutral or slightly alkaline pH values [102]. At the same time, there are information about the resistance of Chlorococcum species to extreme pH. Thus, in the study of adaptation of the strain MS-1 Chlorococcum sp. to extreme alkalinity and salinity, the ability of algae to adapt and grow at a pH of more than 12 was shown [103]. Simultaneously Chlorococcum infusionum was found in very acidic habitats, contaminated with heavy metals [104].
Our exeprimemts also demonstrated that Chlorococcum infusionum is highly resistante to extremely acidic and alkaline pH. It is possible, that one of the adaptations against this negative effect is the ability to produce mucilage around the cells, which reduces the damaging effect of H+ and OH- ions. Possibly, this mechanism is also inherent in other groups of algae. Obvious mucilage produaction was observed for Hormidium ruvulare Kützing (the old synonim of Klebsormidium rivulare (Kützing) M.O.Morison & Sheath) at pH 2.75-3.00 [45].
We found, that changes of pH, especially at pH 3.5, caused mass zoospores germination on Vischeria magna. In the previous publication it was siggested, that spore germination observed at the pH levels, in wich algae can grow [105]. It was noted, that probably akinetes could germinate in acidic pH due to destruction of akinete walls at low pH [48]. Possibly, the similar mechanism caused the germination the zoospores of Vischeria magna at low and high pH.
At low pH (2.5-3) appearance of orange oil granules in Vischeria magna was observed. This effect possibly use in biotechnlogy for inreasing of oil production during cultivation of representatives of Eustigmatohyceae. In previous publication the impact of extreme levels of ecological factors on members of this family was detected. For eaxmple, the influence of salt stress on a decrease in total a very-long-chain fatty acid with high nutraceutical value productivity in an authentic Vischeria punctata strain IPPAS H-242 was observed [106]. It was found, that using nitrate–N, peptone-N sources and NaCl allows enlarging the oil production in Vischeria sp. WL1 strain [107].
We detected the appearance of oil granules in cells of Bracteacoccus minor at pH 3.5-5. According to previous data, in stress conditions the representatives of genus Bracteacoccus increase the oil production. For example, Bracteacoccus bullatus was suitable for synthesis of oil with a high content of unsaturated fatty acids at temperatures 12–18 °C [108].
In previous investigations a number of the physiological mechanisms for surviving the algae at low pH values were described [22,84,92,96,109,110]. These mechanisms include maintaining a positive membrane potential [109], reducing the permeability of the plasma membrane to protons or maintaining an active proton pump to preserve neutral cytosol [84,96,110]. The resistant plasma membrane also provides general tolerance of acidophilic and acidotolerant algae to heavy metals and toxic anions and prevents their entry to the algae cell [84,94,96]. In addition, it has been suggested that morphological adaptations of microalgae to extremely low pH values exist [111].
It is generally believed that cells should have a low surface area to volume ratio (S/V) to reduce stress caused by hydrogen cations penetrating through cell walls and membranes. A decrease in the S/V ratio can be achieved by increasing the volume of cells [22].
Our study revealed several models of changes algae cells biovolume in extreme pH levels:
1. Minor changes according to the linear regression model – this type of reaction was characteristic for Bracteacoccus minor and Chlorococcum infusionum. It is necessary to note, that desmid Euastrum binale also keep the cell size when the pH changes [22]. Possibly for protection in extreme pH levels this species used increase of oil content in cells (Bracteacoccus minor) or mucilage production (Chlorococcum infusionum).
2. Increase in acid and alkaline pH range according to polynomial regression model – such changes were observed in Pseudoccomixa simplex and Vischeria magna cells. Perhaps these algae could use the regulation of biovolume in stress conditions of low and high pH. Vischeria magna in addition increase the synthesis oil containing metabolites.
3. Increase in alkaline pH range according to the linear regression model – was found in experiments with Chlorella vulgaris.
Possibly, the different reactions of algae biovolume at low and high pH could be explained by their taxonomic variability and related genetic, morphological, biochemical, physiological and ecological peculiarities.
Algae strains, resistant to extreme pH, have a great potential for using in the biotechnology. pH is a critical parameter influencing on the mass transfer rate of acidic gases like H2S and CO2 in conventional gas-liquid contactors [85,86]. At pH>9 contaminants in the biogas die, while resistant to pH algae strains survive [87,88,89]. The use of carbonated alkaline culture media in algal-bacterial photobioreactors support a long term effective biogas supporting [89].
Besides, the resistant to high and low pH strains can be used for industrial wastewater treatment, that pH of which ranges from 2 to 11 [112,113].
It should be emphasized the studied strains are authentic (excluding Vischeria magna) and represent reference samples of species. This allows to consider their reaction to a change in pH as an example, and the reaction of other strains can later be compared with the reaction of investigated taxa. In addition, the studied strains are available in several major collections of algae and can be used in further physiological, biochemical and biotechnological research.
Thus, the investigated authentic and reference strains of Chlorella vulgaris, Bracteacoccus minor, Pseudoccomyxa simplex, Chlorococcum infusionum, and Vischeria magna were resistant to extreme pH levels and use several physiological mechanisms for protection in conditions of low and high pH values. These mechanisms include the changes of volume, increasing mucilage and oil containing substances production as well as enhancing of zoospore production. The results obtained allow to recommend the studied strains for broader use in biotechnology and nature protection.
4. Materials and Methods
4.1. Strains cultivation and methodic of experiments
In the study authentic strains or strains with correct identification were used. This strains were received from Collection of algae culture of Taras Shevchenko Kiev National University (ACKU), which, in turn, received these strains from the Culture Collection of Algae at the University of Göttingen (SAG). The following strains in the experiments were used: Chlorella vulgaris – ACKU 531-06 (SAG (211-11b), Bracteacoccus minor – ACKU 506-06 (SAG 221-1), Pseudoccomixa simplex – ACKU 559-06 (SAG (216-9a), Chlorococcum infusionum ACKU 539-06 (SAG 10.86) and UTEX (UTEX 2351 Eustigmatos magna =Vischeria magna).
Before the experiments, the pH meter electrodes were calibrated using standard buffer solutions (pH=6.87 and pH=9.18). In experiments pH levels from 2 to 13.5 with an interval of 0.5 were studied. The liquid Bold media [114] with a pH of 6.5 was a control. The different pH values were achieved by adding 0.1 molar solutions of NaOH or HCl, which were added into the Bold media. The pH of the media was measured using a pH meter "Multitest IPL-311" with an ESC electrode No. 1060317. Small amount of algae biomass was put into the tubes with the different pH concentrations. The tubes were incubated 7 days at 20–23° C with illumination of 40 μmol/m2 per second using 18W cool fluorescent tubes (Philips TLD18W/33, Philips Lighting Poland S.A., Pila, Poland).
In each pH concentrations 5 tubes were examined, from each tube at least 20 cells were analyzed. For this purpose, in each concentration the micrographs, containing at least 100 algae cells, were taken. After that in each concentration measurements of the length, width and other dimensions at least 100 algae cells were made. For taking the algae pictures and analyzing the algae cells parameters the Axio Vision 4.9 software was used.
4.2. Statistical analysis
Statistical processing of the results was carried out using regression analysis [115]. For calculation the biological volume of algae (V), formulas of geometric shapes that most accurately corresponded to the shape of their body were used [116]. The volume of globular cells of Chlorella vulgaris, Bracteacoccus minor, Chlorococcum infusionum, Vischeria magna was calculated by the formula of the sphere volume:
where a is diameter of the sphere (Sun, Liu, 2003).
V=π/6 a3 (1),
It should be noted that the young cells of Chlorella vulgaris and Chlorococcum infu-sionum have an elongated shape. Therefore, for calculating the volume of cells of these species and Pseudococcomyxa simplex cells the formula of an ellipsoid was used:
where a is the length of the major axis, b is the length of the minor axis (Sun, Liu, 2003).
V=π/6 b2 a
During the analysis of the pH influence on the volume of Chlorella vulgaris and Chlorococcum infusionum cells, the volumes of both spherical and ellipsoid cells were taken into account.
Statistical processing of the results was carried out using the Past 2.14 software [117]. The morphological variations related to the changing pH values were analyzed. For this purpose, the most suitable regression model was chosen.
Funding
This research was funded by the Ministry of Education of the Russian Federation, grant number BWUZ-2023-0006.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors are thankful to Yu. Z. Gabidullin for their help with the preparation of the figures.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kushner, D.J. Microbial life in extreme conditions; Mir: Moscow, Russia, 1981. [Google Scholar]
- Odum, Yu. Fundamentals of Ecology; Mir: Moscow, Russia, 1975. [Google Scholar]
- Coleman, J.R.; Colman, B. Effect of external pH on carbon accumulation and CO2 fixation in blue-green alga. Proceedings of 5th Int. Congr. Photosynth. Malkidik, 89–90. 1980. 123. [Google Scholar]
- Reddy, H.; Balasubramanian, A.; Shantaram, M.V. Influence of pH on growth (14 C)-carbon metabolism and nitrogen fixation by two blue-green algae. Phycos 1980, 1980. 19, 45–51. [Google Scholar]
- Rachlin, J.; Grosso, A. The effects of pH on the growth of Chlorella vulgaris and its interactions with cadmium toxicity. Arch. Environ. Contam. And Toxicol 1991, 1991. 20, 505–508. [Google Scholar] [CrossRef]
- Taylor, R.W. The effect of the pH on the division rate of the coccolithophonid Cricoshaera elongate. Journal of Phycology 1966, 2, 33–42. [Google Scholar] [CrossRef]
- McLean, R. J.; Trainor, F. R. Fasciculochloris, a new chlorosphaeracean alga from Connecticut soil. Phycologia 1965, 4, 145–148. [Google Scholar] [CrossRef]
- Hopkins, E.F.; Wann, F.B. Relation of hydrogen-ion concentration to growth of Chlorella and to the availability of iron. Bot. Gaz. 1926, 18–24. [Google Scholar] [CrossRef]
- Maciasr, F. M. Effect of pH of the medium on the availability of chelated iron for Chlamydomonas mundane. The Journal of Protozoology 1965, 12, 302–311. [Google Scholar] [CrossRef] [PubMed]
- Hood, D.W.; Park, K. Bicarbonate utilization by marine phytoplankton in photosynthesis. Physiologia Plantarium 1962, 15, 43–45. [Google Scholar] [CrossRef]
- Rao, K.V.N. Determination of optimum pH range for the growth of Oocystis marssonii Lemm. in the three media differing in nitrogen source. Indian Journal of Plant Physiology 1963, 6, 2–10. [Google Scholar]
- Spencer, C.P. Theoretical aspects of the control of pH in natural sea water and synthetic culture media for marine algae. Botanica Marina 1966, 9, 3–4. [Google Scholar] [CrossRef]
- Ikemori, M. , Noshida, K. Inorganic carbon sourse and the inhibitory effect of diamox on the photosynthesis of marine algae, Ulva pertusa. Ann. Rept. Noto Mar. Lab. 1967, 7, 63–81. [Google Scholar]
- Thomas, E.A.; Tregunna, E.B. Bicarbonate ion assimilation in photosynthesis by Sargassum muticum. Canadian Journal of Botany 1968, 46, 256–261. [Google Scholar] [CrossRef]
- Raven, J. The mechanism of photosynthetic use of bicarbonate by Hydrodictyon africanum. Exp. Bot. 1968, 46, 82–85. [Google Scholar] [CrossRef]
- Khaili, Z. , Asker M.S., El-Sayed S., Kobbia I.A. Responces of Dunaliella bardawil and Chlorella ellipsoidea to pH stress. Acta Bot. Hung. 2010, 52, 123–135. [Google Scholar] [CrossRef]
- Jabir, T.F.; Noor Abbood, H.A.; Salman, F.S.; Hafit, A.Y. Influence of pH, pesticide and radiation interactions on the chemical composition of Chlorella vulgaris algae. IOP Conf. Ser. Earth Environ. Sci. 2021, 722, 012046. [Google Scholar] [CrossRef]
- Gollerbach, M.M.; Shtina, E.A. Soil Algae; Nauka: Leningrad, Russia, 1969; p. 228. (In Russian) [Google Scholar]
- Schneider, S.C.; Kahlert, M.; Kelly, M.G. Interactions between pH and nutrients on benthic algae in streams and consequences for ecological status assessment and species richness patterns. Science of The Total Environment 2013, 444, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Thomas, B.T.; Rajan, R.; Saranyamol, S. T.; Bhagya, M. V. influence of pH on the diversity of soil algae. Journal of Global Biosciences 2017, 6, 5088–5094. [Google Scholar]
- Lavoie, M.; Le Faucheur, S.; Boullemant, A.; Fortin, C.; Campbell, P.G.C. The influence of pH on algal cell membrane permeability and its implications for the uptake of lipophilic metal complexes. Journal of Phycology, 48. [CrossRef]
- Černá, K.; Neustupa, J. The pH-related morphological variations of two acidophilic species of Desmidiales (Viridiplantae) isolated from a lowland peat bog, Czech Republic. Aquatic Ecology 2010, 44, 409–419. [Google Scholar] [CrossRef]
- Shtina, E.A.; Gollerbach, M.M. Ecology of the soil algae; M.: Nauka, 1976. (In Russian) [Google Scholar]
- Andreeva, V.M. Soil and aerophilic green algae (Chlorophyta: Tetrasporales, Chlorococcales, Chlorosarcinales); Nauka, Saint Petersburg, Russia, 1998; 351 p.
- Kostikov, I.; Romanenko, P.; Demchenko, P.; Darienko, T.M.; Mikhayljuk, T.I.; Rybchinskiy, O.V.; Solonenko, A.M. Soil Algae of Ukraine; Phytosotsiologichniy Center: Kiev, Ukraine, 2001; p. 300. [Google Scholar]
- Büdel, B.; Darienko, T; Deutschewitz, K. ; Dojani, S.; Friedl, T.; Mohr, K.I.; Salisch, M.; Reisser, W.; Weber, B. Southern African biological soil crusts are ubiquitous and highly diverse in drylands, being restricted by rainfall frequency. Microbial Ecology 2009, 57, 229–247. [Google Scholar] [CrossRef] [PubMed]
- Škaloud, P. Species composition and diversity of aero-terrestrial algae and cyanobacteria of the Boreč Hill ventaroles. Fottea 2009, 9, 65–80. [Google Scholar] [CrossRef]
- Ettl, H. , Gärtner G. Syllabus der Boden-, Luft- und Flechtenalgen, S: Fischer Verlag, 1995. [Google Scholar]
- Lukešová, A.; Hoffmann, L. Soil algae flora from acid rain impacted forest areas of the Krušne hory Mts. 1. Algal communities. Vegatatio 1996, 125, 123–136. [Google Scholar] [CrossRef]
- John, D.M.; Witton, B.A.; Brook, A.J. The freshwater algal flora of the British Isles: an identification guide to freshwater and terrestrial algae; Cambridge University Press, Cambridge, UK, 2005; 702 p.
- Johansen, J.R.; Rushfort, S.R.; Brotherson, J.D. The algal flora of Navajo National Monumant, Arizona, U.S.A. Nova Hedwigia 1983, 38, 501–553. [Google Scholar]
- Broady, P.A. Diversity, distribution and dispersal of Antarctic terrestrial algae. Biodiversity and Conservation 1996, 5, 1307–1335. [Google Scholar] [CrossRef]
- Lukešova, A. Soil algae in brown coal and lignite post-mining areas in Central Europe (Czech Republic and Germany). Restoration Ecology. 2001, 9, 341–350. [Google Scholar] [CrossRef]
- Flechtner, V.R.; Johansen, J.R. , Belnap, J. The biological soil crusts of the San Nicolas Island: enigmatic algae from a geographically isolated ecosystems. Western North American Naturalist 2008, 68, 405–436. [Google Scholar] [CrossRef]
- Moskvich, N.P. The experience of using algae in the study of the sanitary state of the soil. Botanical Journal 1973, 58, 412–416. [Google Scholar]
- Novakovskaya, I.V.; Patova, E.N. Soil algae of spruce forests and their changes in conditions of aerotechnogenic pollution. Komi Scientific Center: Syktyvkar, Russia, 2012. 128 p.
- Novichkova-Ivanova, L.N. Changes of synusions of soil algae of Franz Josef Land. Botanical Journal 1963, 47, 42. [Google Scholar]
- Shtina, E.A. Soil algae as pioneers of the overgrowth of technogenic substrates and indicators of the state of disturbed lands. Journal of General Biology 1985, 46, 435–443. [Google Scholar]
- Neganova, L.B.; Shilova, I.I.; Shtina, E.A. Algoflora of technogenic sands of oil and gas producing areas of the Middle Ob region and the impact of oil pollution on it. Ecology 1978, 3, 29–35. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. World-wide electronic publication, National University of Ireland, Galway. . AlgaeBase. https://www.algaebase. 22 November 2023. [Google Scholar]
- Kaštovska, K.; Elster, J.; Stibal, M.; Šantrůčkova, H. Microbial assemblages in soil microbial succession after glacial retreat in Svalbard (High Arctic). Microbial Ecology 2005, 50, 396–407. [Google Scholar] [CrossRef] [PubMed]
- Gomez, Sh.R.; Johansen, J.R.; Lowe, R.L. Epilithic aerial algae of Great Smoky Mountains National Park. Biologia, Bratislava.
- Johansen, J.R.; Lowe, R.; Gomez, Sh.R.; Kociolek, J.P.; Makosky, S.A. New algal records for the Great Smoky Mountains National Park, U.S.A., with an annotated checklist of all reported algal species for the park. Algological Studies. [CrossRef]
- Shtina, E.L.; Bolyshev, N.I. Algae communities in soils of dry and desert steppes. Botanical Journal 1963, 48, 670–680. [Google Scholar]
- Hargreaves, J.; Whitton, B. Effect of pH on growth of acid stream algae. Eur. J. Phycol. 1976, 11, 215–223. [Google Scholar] [CrossRef]
- Zuppini, A.; Gerotto, C.; Baldan, B. Programmed cell death and adaptation: two different types of abiotic stress response in a unicellular chlorophyte. Plant and Cell Physiology 2010, 51, 884–895. [Google Scholar] [CrossRef] [PubMed]
- Bastien, C.; Côté, R. Effects du cuirve sur l’ultrastructure de Scenedesmus quadricauda et Chlorella vulgaris. Inf. Rev. Gesamt. Hydrobiol. 1989, 74, 57–71. [Google Scholar]
- Agrawal, S.C. Effects of different factors on the akinete germination in Pithophora oedogonia (Mont.) Wittrock. J.Basic Microbiol. 1986, 26, 195–199. [Google Scholar] [CrossRef]
- Rothe, S.; Schumann, R.; Karsten, U. Effect of temperature and photon influence rate on growth rates of two epiphytic diatom species from Kongsfjorden. Ber.Polar- und Meeresforsch. 2004, 492, 136–146. [Google Scholar]
- Liu, C.G.; Jin, X.C.; Sun, L.; Zhong, Y.; Dai, S.G.; Zhuang, Y.Y. Influence of рН on growth and species composition of algae in freshwater ecosystems. J. Agro-Environ. Sci. 2005, 24, 294–298. [Google Scholar]
- Zhan, Y.-J.; Wang, X.-L.; Yang, R.-J.; Zhang, Y.-Y. Influence of Cu (II) on growth of 8 species of marine microalgae. Environ.Sci. 2006, 27, 720–726. [Google Scholar]
- Middelboe, A.L.; Hansen, P.J. Direct effects of pH and inorganic carbon on macrolalgal photosynthesis and growth. Mar. Biol. Res. 2007, 3, 134–144. [Google Scholar] [CrossRef]
- Mitrovic, S.M.; Hitchcock, J.N.; Davie, A.W.; Ryan, D.A. Growth responses of Cyclotella meneghiniana (Bacillariophyceae) to various temperatures. J. Plankton Res. 2010, 32, 1217–1221. [Google Scholar] [CrossRef]
- Gigova, L.; Ivanova, N.; Gacheva, G.; Andreeva, R.; Furnadzhieva, S. Response of Trachydiscus minutes (Xanthophyceae) to temperature and light. Journal of Phycology.
- Karandashova, I.V.; Elanskaya, I.V. Genetic control and mechanisms of cyanobacteria resistance to salt and hyperosmotic stress. Genetics 2005, 41, 1589–1600. [Google Scholar] [CrossRef]
- Backor, M.; Fahselt, D.; Davidson, R.D.; Wu, C.T. Effects of copper on wild and tolerant strains of the lichen photobiont Trebouxia erici (Chlorophyta) and possible tolerance mechanism. Arch. Environ. Contam. And Toxicol. 2003, 45, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Piovár, J.; Stavrou, E.; Kimákova, T.; Kadukova, J. , Bačkor, M. Influence of long-term exposure to copper on the lichen photobiont Trebouxia erici and free-living algae Scenedesmus quadricauda. Plant Growth Regul. 2011, 63, 81–88. [CrossRef]
- Messerli, M.A.; Amaral-Zettler, L.A.; Zettler, E.; Jung, S.-K.; Smith, P.J.S.; Sogin, M.L. Life in acidic pH imposes as increased energetic cost for a eukaryotic acidophile. J. Exp. Biol. 2005, 208, 2569–2579. [Google Scholar] [CrossRef] [PubMed]
- Almutairi, A. W.; & Toulibah, H. E.; & Toulibah, H. E. Effect of salinity and pH on fatty acid profile of the Green Algae Tetraselmis suecica. Journal of Petroleum & Environmental Biotechnology 2017, 8, 1–6. [Google Scholar] [CrossRef]
- Li, K.W.; McLaughlin, F.; Lovejoy, C.; Carmack, E.C. Smallest algae thrive as the Arctic Ocean freshens. Science 2009, 326, 539. [Google Scholar] [CrossRef] [PubMed]
- Bratbak, G. Bacterial biovolume and biomass estimation. Applied and Environmental Microbiology 1985, 49, 1488–1493. [Google Scholar] [CrossRef] [PubMed]
- Sunda, W.G. , Huntsman, S.A. Interrelated influence of iron, light and cell size on marine phytoplankton growth. Nature. [CrossRef]
- Weisse, T.; Scheffel, U.; Stadler, P.; Foissner, W. Local adaptation among geographically distant clones of the cosmopolitan freshwater ciliate Meseres corlissi. II. Response to pH. Aquatic Microbial Ecology 2007, 47, 289–297. [Google Scholar] [CrossRef]
- Hillebrand, H.; Dürselen, C.-D.; Kirschtel, D.; Pollingher, U.; Zohary, T. Biovolume calculation for pelagic and benthic microalgae. Journal of Phycology 1999, 35, 403–424. [Google Scholar] [CrossRef]
- Sun, J.; Liu, D. Geometric models for calculating cell biovolume and surface area for phytoplankton. Journal of Plankton Research 2003, 25, 1331–1346. [Google Scholar] [CrossRef]
- Borics, G.; Lerf, V.; Enikő, T.; Stanković, I.; Pickó, L.; Béres, V.; Várbíró, G. Biovolume and surface area calculations for microalgae, using realistic 3D models. The Science of the Total Environment 2021, 773, 145538. [Google Scholar] [CrossRef] [PubMed]
- Finkel, Z. V. Light absorption and size scaling of the lightlimited metabolism in marine diatoms. Limnol. Oceanogr. 2001, 46, 86–94. [Google Scholar] [CrossRef]
- Raven, J.A.; Kubler, J. E. New light on the scaling of metabolic rate with the size of algae. Journal of Phycology 2002, 38, 11–16. [Google Scholar] [CrossRef]
- Tang, E.P.Y. The allelometry of algal growth rates. Journal of Plankton Research 1995, 17, 1325–1335. [Google Scholar] [CrossRef]
- Tang, E.P.Y. , Peters, R.H. The allelometry of algal respiration. Journal of Plankton Research 1995, 17, 303–315. [Google Scholar] [CrossRef]
- Sarthou, G.; Timmermans, K.R.; Blain, S.; Treguer, P. Growth physiology and fate of diatoms in the ocean: a review. Journal of Sea Research 2005, 53, 25–42. [Google Scholar] [CrossRef]
- Round, F.E.; Crawford, R.M.; Mann, D.G. The Diatom: Biology and Morphology of the Genera. Cambridge University Press, Cambridge, UK; 1990. 747 p.
- Irwin, A.J.; Finkel, Z.V.; Schofield, O.M.E.; Falkowski, P.G. Scaling-up from nutrient physiology to the size-structure of phytoplankton communities. Journal of Plankton Research 2006, 28, 459–471. [Google Scholar] [CrossRef]
- Agusti, S. Allometric scaling of light absorption and scattering by phytoplankton cells. Can. J. Fish. Aquat. Sci. 1991, 48, 763–767. [Google Scholar] [CrossRef]
- Joint, I.; Pomroy, A. Allometric estimation of the productivity of phytoplankton assemblages. Mar. Ecol. Prog. Ser. 1988, 47, 161–168. [Google Scholar] [CrossRef]
- Connolly, J.A.; Oliver, M.J.; Beaulieu, J.M.; Knight, C.A.; Tomanek, L.; Moline, M.A. Correlated evolution of genome size and cella volume in diatoms (Bacillariophyceae). Journal of Phycology 2008, 44, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Hein, M. , Pedersen, M.F., Sand-Jensen, K. Size-dependent nitrogen uptake in micro- and macroalgae. Marine Ecology Progress Series 1995, 118, 247–253. [CrossRef]
- Vogel, S. Life's Devices: The Physical World of Animals and Plants. Princeton University Press, Princeton, NJ, USA, 1988; 369 p.
- Borics, G.; Várbíró, G.; Falucskai, J.; Végvári, Z.; T-Krasznai, E.; Görgényi, J.; B-Béres, V.; Lerf, V. A two-dimensional morphospace for cyanobacteria and microalgae: Morphological diversity, evolutionary relatedness, and size constraints. Freshw. Biol. 2023, 68, 115–126. [Google Scholar] [CrossRef]
- Polishuk,V. ; Kostikov, I.Yu.; Taran, N.Yu.; Voitsitsky, V.M.; Budzanivska, I.; Khyzhnyak, S.; Trokhymets, V.M. The complex studying of Antarctic biota. Ukr. Antarct. J. 2009, 8, 293–301. [Google Scholar] [CrossRef]
- Neustupa, J.; Skaloud, P. Diversity of subaerial algae and cyanobacteria on tree bark in tropical mountain habitats. Biologia 2008, 63, 806–812. [Google Scholar] [CrossRef]
- Kessler, E.; Kramer, H. Physiologische untersuchungen an einer ungewöhnlich saureresistenten Chlorella. Arch. Mikrobiol. 1960, 37, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Gimmler, H.; Weis, U. Dunaliella acidophila – life at pH 1.0. In Dunaliella, physiology, biochemistry and biotechnology. M. Avron, A. Ben Amotz. CRC Press: Boca Raton, USA, 1992; 99–133.
- Gross, W. Ecophysiology of algae living in highly acidic environments. Hydrobiologia 2000, 433, 31–37. [Google Scholar] [CrossRef]
- Bahr, M.; Díaz, I.; Dominguez, A.; González Sánchez, A.; Muñoz, R. Microalgal biotechnology as a platform for an integral biogas upgrading and nutrient removal from anaerobic effluents. Environ. Sci. Technol. 2014, 2014. 48, 573–81. [Google Scholar] [CrossRef]
- González-Sánchez, A.; Revah, S.; Deshusses, M.A. Alkaline biofiltration of H2S odors. Environ. Sci. Technol. 2008, 42, 7398–7404. [Google Scholar] [CrossRef] [PubMed]
- González-Sánchez, A.; Revah, S. The effect of chemical oxidation on the biological 396 sulfide oxidation by an alkaliphilic sulfoxidizing bacterial consortium. Enzyme Microb. Technol. 2007, 40, 292–298. [Google Scholar] [CrossRef]
- Markou, G.; Vandamme, D.; Muylaert, K. Microalgal and cyanobacterial cultivation: The supply of nutrients. Water Res. 2014, 65, 186–202. [Google Scholar] [CrossRef] [PubMed]
- Franco-Morgado, M.; Alcántara, C.; Noyola, A.; Muñoz, R.; González-Sánchez, A. A study of photosynthetic biogas upgrading based on a high rate algal pond under alkaline conditions: Influence of the illumination regime. Sci. Total Environ. 2017, 592, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Olaveson, M. M.; Stokes, P. M. 1989: Responses of the acidophilic Euglena mutabilis (Euglenophyceae) to carbon enrichment at pH 3. Journal of Phycology.
- Graham, J. M.; Arancibia-Avila, P.; Graham, L. E. Effects of pH and selected metals on growth of the filamentous green alga Mougeotia under acidic conditions. Journal of Limnology and Oceanography 1996, 41, 263–270. [Google Scholar] [CrossRef]
- Gerloff-Elias, A.; Spijkerman, E.; Pröschold, T. Effect of external pH on the growth, photosynthesis and photosynthetic electron transport of Chlamydomonas acidophila Negoro, isolated from an extremely acidic lake (pH 2.6). Plant, Cell and Environment, 1218. [Google Scholar]
- Hoham, R.W.; Filbin, R.W.; Frey, F.M.; Pusack, T.J.; Ryba, J.B.; Mcdermott, P.D.; Fields, R.A. The optimum pH of the green snow algae, Chloromonas tughillensis and Chloromonas chenangoensis, from upstate New York. Arctic, Antarctic and Alpine Research. [CrossRef]
- Spijkerman, E.; Barua, D.; Gerloff-Elias, A.; Kern, J.; Gaedke, U.; Heckathorn, S.A. Stress responses and metal tolerance of Chlamydomonas acidophila in metalenriched lake water and artificial medium. Extremophiles 2007, 11, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Ňancucheo, I.; Johnson, B.D. Acidophilic algae isolated from mine-impacted environments and their roles in sustaining heterotrophic acidophiles. Front. Microbiol. 2012, 3, 325. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, M.R.; Senhorinho, G.N.A.; Scott, J.A. Microalgae under Environmental Stress as a Source of Antioxidants. Algal Res. 2020, 52, 102104. [Google Scholar] [CrossRef]
- Moss, B. The influence of environmental factors on the distribution of freshwater algae: An experimental study: II. The role of pH and the carbon dioxide-bicarbonate system. J. Ecol. [CrossRef]
- Novis, P. Taxonomy of Klebsormidium (Klebsormidales, Chlorophyceae) in New Zealand streams, and the significance of low pH habitats. Phycologia.
- Xing, R.; Ma, W.; Shao, Y.; Cao, X.; Chen, L.; Jiang, A. Factors that affect the growth and photosynthesis of the filamentous green algae, Chaetomorpha valida, in static sea cucumber aquaculture ponds with high salinity and high pH. PeerJ 2019, 7, e6468. [Google Scholar] [CrossRef] [PubMed]
- Mayo, A.W. Effects of temperature and pH on the kinetic growth of unialga Chlorella vulgaris cultures containing bacteria. Water Environment Research 1997, 69, 64–72. [Google Scholar] [CrossRef]
- Qiu, R.; Gao, S.; Lopez, P.A.; Ogden, K.L. Effects of pH on cell growth, lipid production and CO2 addition of microalgae Chlorella sorokiniana. Algal Res. 2017, 28, 192–199. [Google Scholar] [CrossRef]
- Karemore, A.; Pal, R.; Sen, R. Strategic enhancement of algal biomass and lipid in Chlorococcum infusionum as bioenergy feedstock. Algal Res. 2013, 2, 113–121. [Google Scholar] [CrossRef]
- Xiang, W.-Z.; Wu, H.-L.; Xie, K.; He, H.; Xiao, W. Extreme features of Chlorococcum sp. and rapid induction of ataxantin. J. Trop. Oceanogr. 2007, 26, 50–54. [Google Scholar]
- Shubert, E.; Rusu, L.E. ; Bartok, A-M.; Moncrieff, C.B. Distribution and abundance of edaphic algae adopted to higly acidic, metal rich soils. In Algae and Extreme Environments. J. Elster, O. Lhotsky. Nova Hedwigia 123. J. Cramer, Stuttgart, Germany, 2001; pp. 411–425.
- Agrawal, S.C. Factors affecting spore germination in algae — review. Folia Microbiologica 2009, 2009. 54, 273–302. [Google Scholar] [CrossRef]
- Sinetova, M.A.; Sidorov, R.A.; Medvedeva, A.A.; Starikov, A.Y.; Markelova, A.G.; Allakhverdiev, S.I.; Los, D.A. Effect of salt stress on physiological parameters of microalgae Vischeria punctata strain IPPAS H-242, a superproducer of eicosapentaenoic acid. J. Biotechnol. 2021, 331, 63–73. [Google Scholar] [CrossRef] [PubMed]
- She, Y.; Gao, X.; Jing, X.; Wang, J.; Dong, Y.; Cui, J.; Xue, H.; Li, Z.; Zhu, D. Effects of nitrogen source and NaCl stress on oil production in Vischeria sp. WL1 (Eustigmatophyceae) isolated from dryland biological soil crusts in China. J. Appl. Phycol, 1291. [Google Scholar] [CrossRef]
- Lukavský, J.; Kopecký, J.; Kubácˇ, D.; Kvíderová, J.; Procházková, L.; Řezanka, T. The alga Bracteacoccus bullatus (Chlorophyceae) isolated from snow, as a source of oil comprising essential unsaturated fatty acids and carotenoids. J. Appl. Phycol. [CrossRef]
- Remis, D.; Treffny, B.; Gimmler, H. Light-induced H+ transport across the plasma membrane of the acid-resistant green alga Dunaliella acidophila. Plant Physiol Biochem. 1994, 1994. 32, 1–10. [Google Scholar]
- Gimmler, H. Acidophilic and acidotolerant algae. In Algal adaptations to environmental stresses. L.C. Rai, J.P. Gaur. Springer: Berlin, Germany, 2001; 259–290.
- Nixdorf, B.; Fyson, A.; Krumbeck, H. Review: plant life in extremely acidic water. Environ. Exp. Bot. 2001, 46, 203–211. [Google Scholar] [CrossRef]
- Grzmil, B.; Wronkowski, J. Removal of phosphates and fluorides from industrial wastewater. Desalination 2006, 189, 261–268. [Google Scholar] [CrossRef]
- Mohsenpour, S.F.; Hennige, S.; Willoughby, N.; Adeloye, A.; Gutierrez, T. Integrating micro-algae into wastewater treatment: A review. Science of The Total Environment 2021, 752. [Google Scholar] [CrossRef] [PubMed]
- Bishoff, H.; Bold, H. Phycological Studies. IV. Some algae from enchanted rock and related algae species. Univ. Texas. Publ.: Texas, USA, 1963; 6318: 91–95.
- Jongman, R.H.G.; Ter Braak, C.J.F.; Van Tongeren, O.F.R. Data Analysis in Community and Landscape Ecology. Cambridge University Press: Cambridge, UK, 1996; 29–50.
- Sun, J.; Liu, D. Geometric models for calculating cell biovolume and surface area for phytoplankton. Journal of Plankton Research 2003, 25, 1331–1346. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: paleontological statistics software package for education and data analysis. 2001. Palaeontol. Electron, 1–9. 10.1016/j.bcp.2008.05. [CrossRef]
Figure 1.
Influence of pH on Bracteacoccus minor morphology. (A) control variant (рН 6,5); (B) рН 3; (C) pH 8,5; (D) рН 9; (E) рН 10,5; (F) рН 11. Scale bar 10 μm.
Figure 1.
Influence of pH on Bracteacoccus minor morphology. (A) control variant (рН 6,5); (B) рН 3; (C) pH 8,5; (D) рН 9; (E) рН 10,5; (F) рН 11. Scale bar 10 μm.
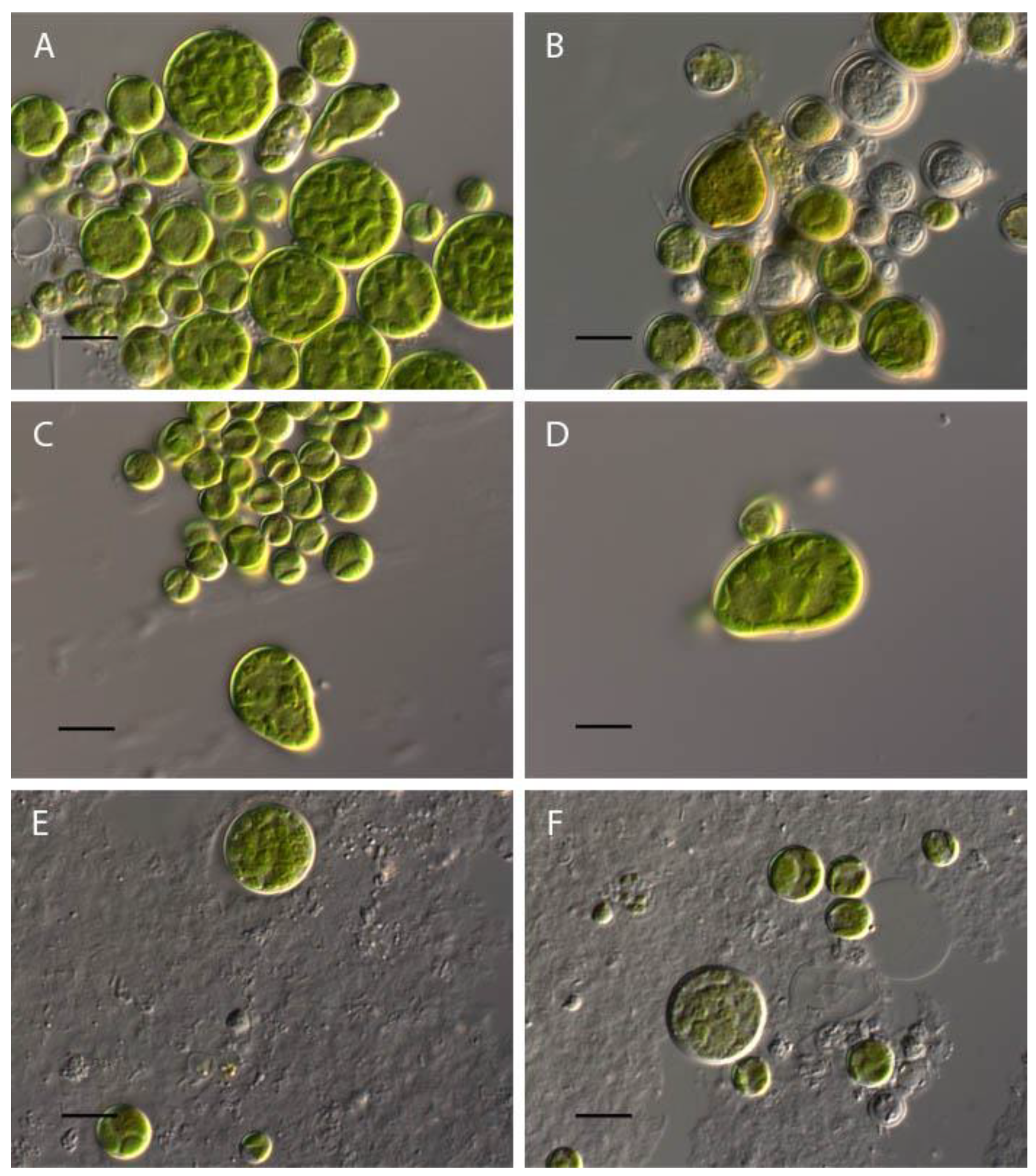
Figure 2.
Oil containing vacuoles in Bracteacccus minor cells (shown by white arrow). Scale bar 10 μm.
Figure 2.
Oil containing vacuoles in Bracteacccus minor cells (shown by white arrow). Scale bar 10 μm.
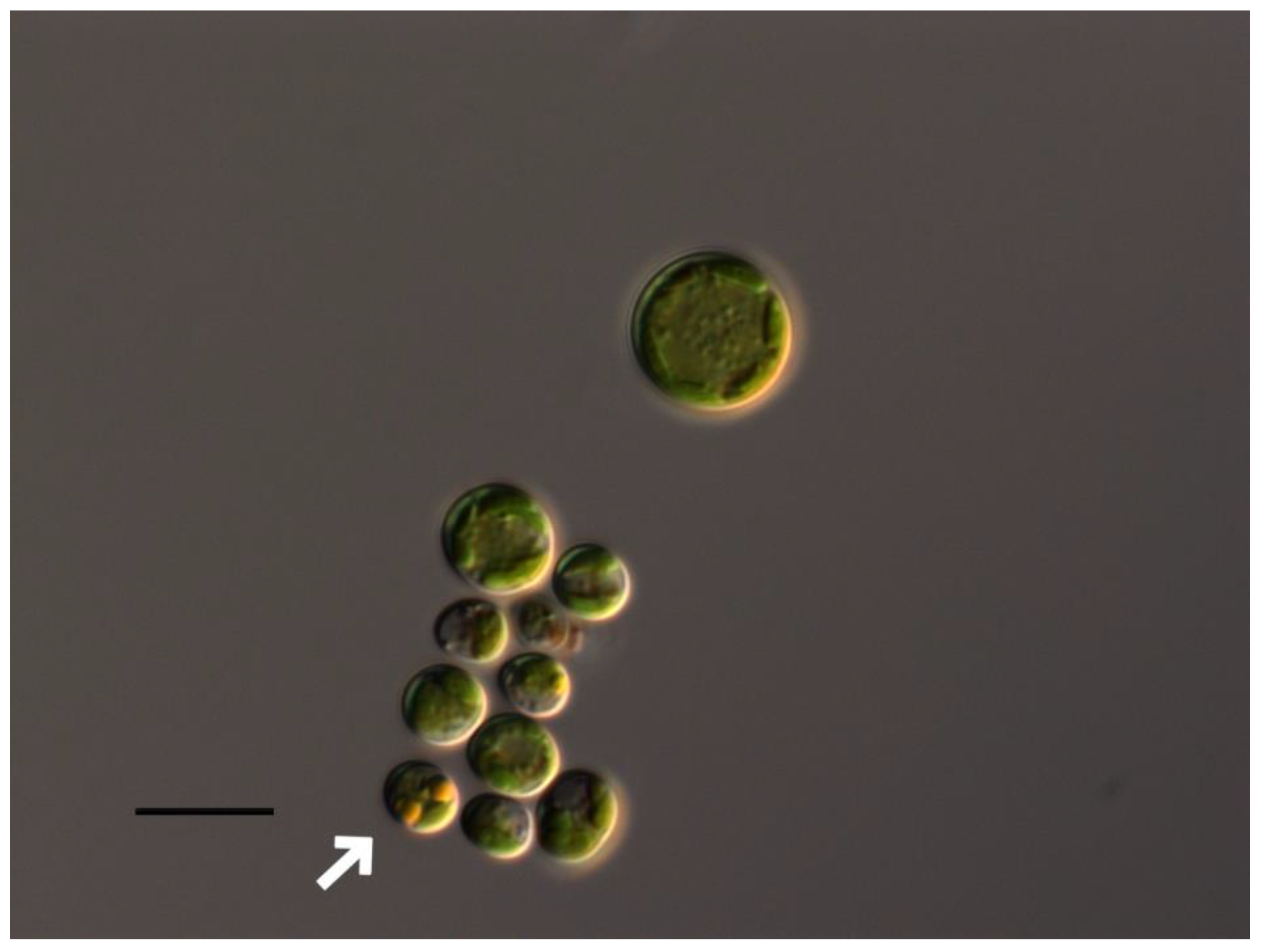
Figure 3.
– Influence of pH on cell volume of Bracteacoccus minor.
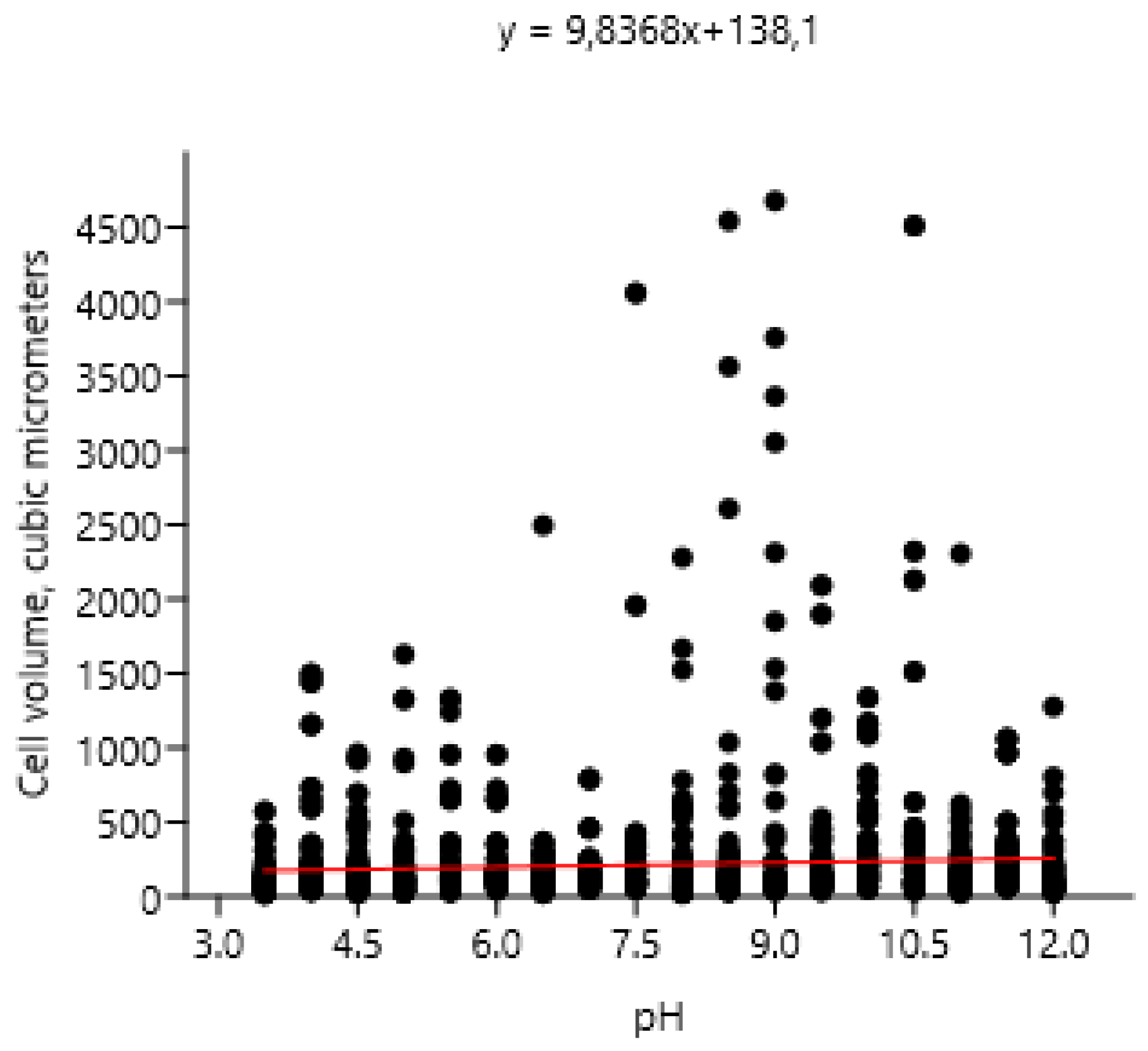
Figure 4.
Influence of pH on Chlorococcum infusionum morphology. (A) control variant (рН 6,5); (B) рН 2; (C) pH 3; (D) рН 3.5; (E) рН 10; (F) рН 12. Scale bar 10 μm.
Figure 4.
Influence of pH on Chlorococcum infusionum morphology. (A) control variant (рН 6,5); (B) рН 2; (C) pH 3; (D) рН 3.5; (E) рН 10; (F) рН 12. Scale bar 10 μm.
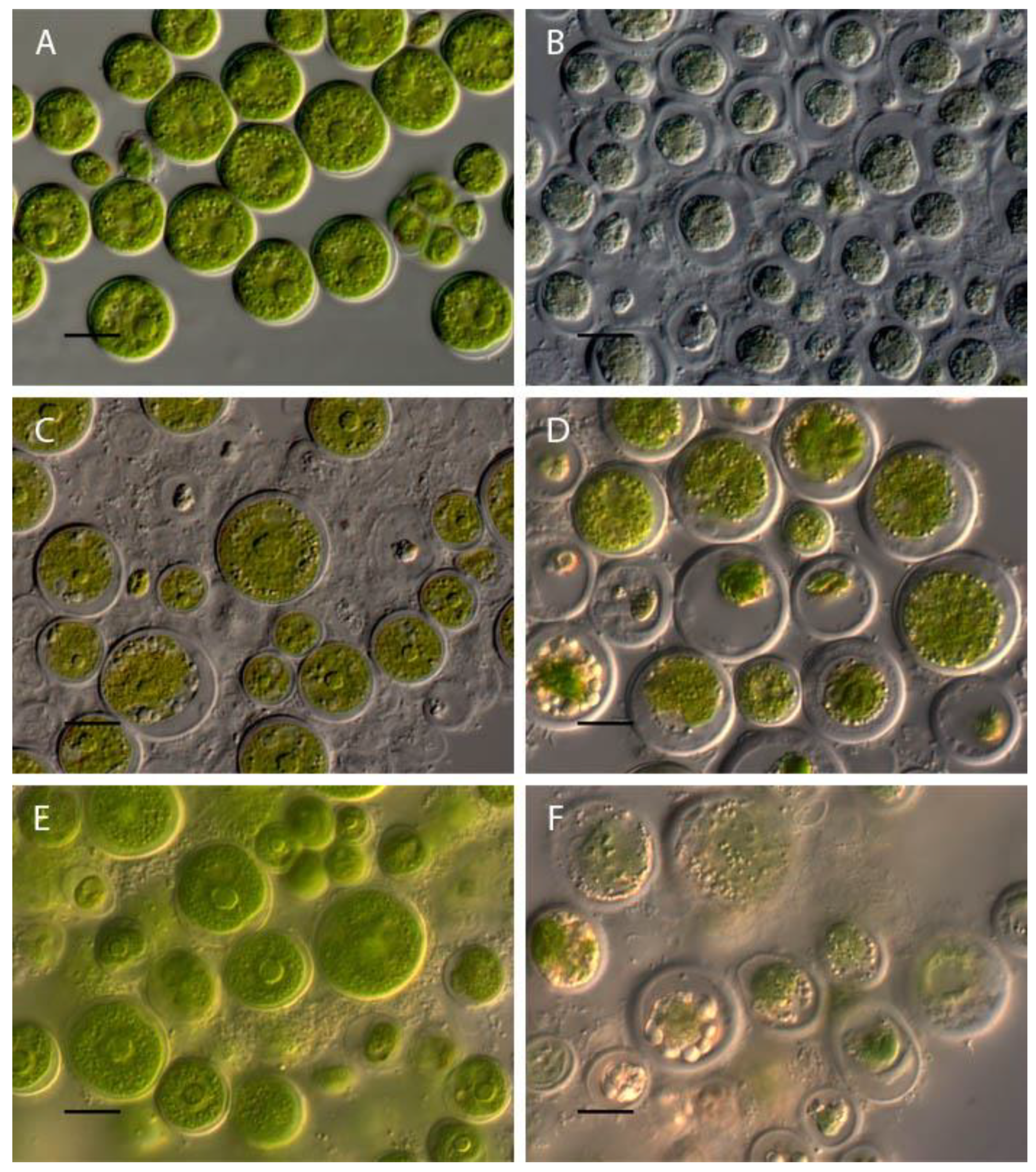
Figure 5.
Influence of pH on cell volume of Chlorococcum infusionum.
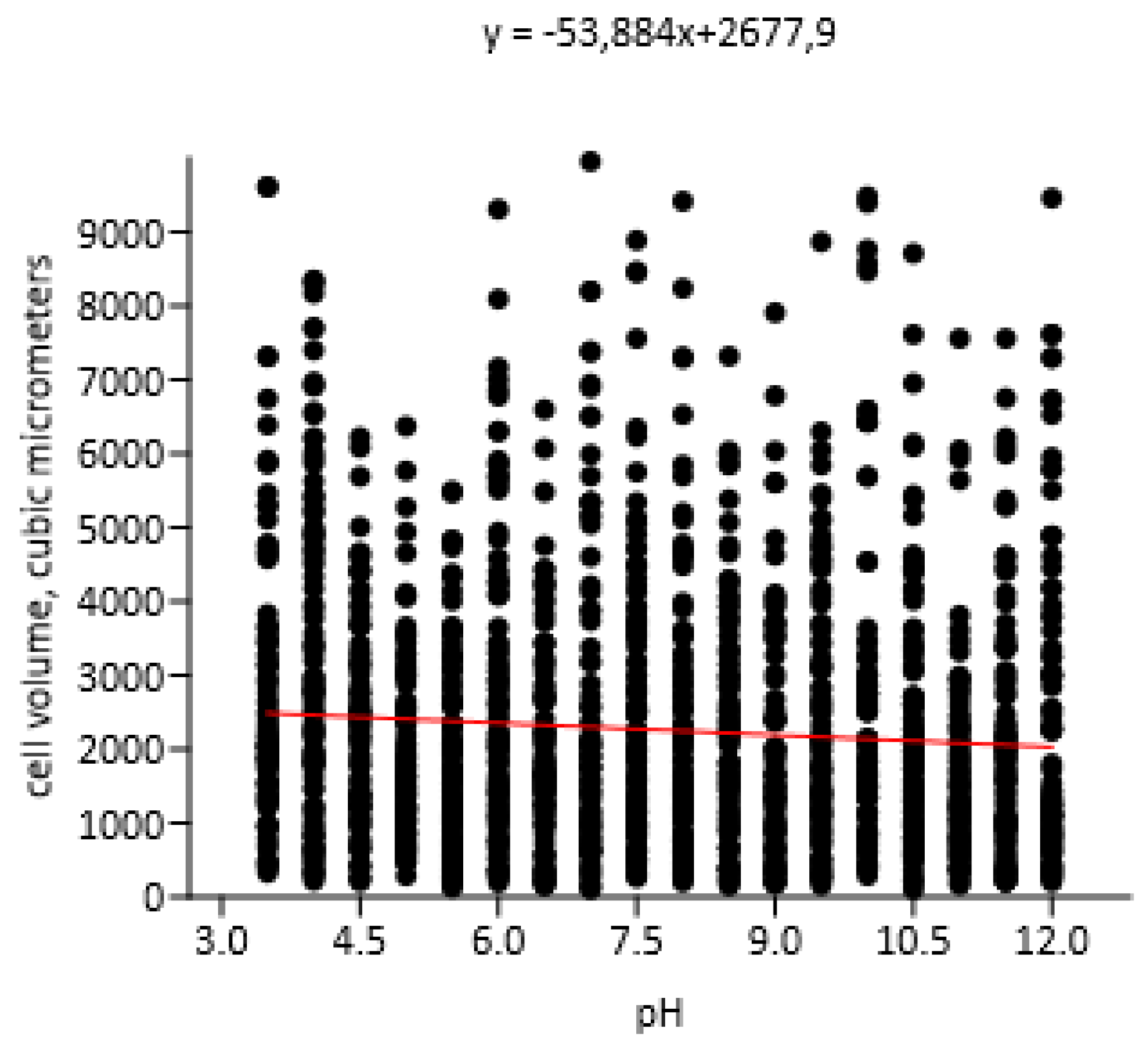
Figure 6.
- Influence of pH on mucilage thickness of Chlorococcum infusionum.
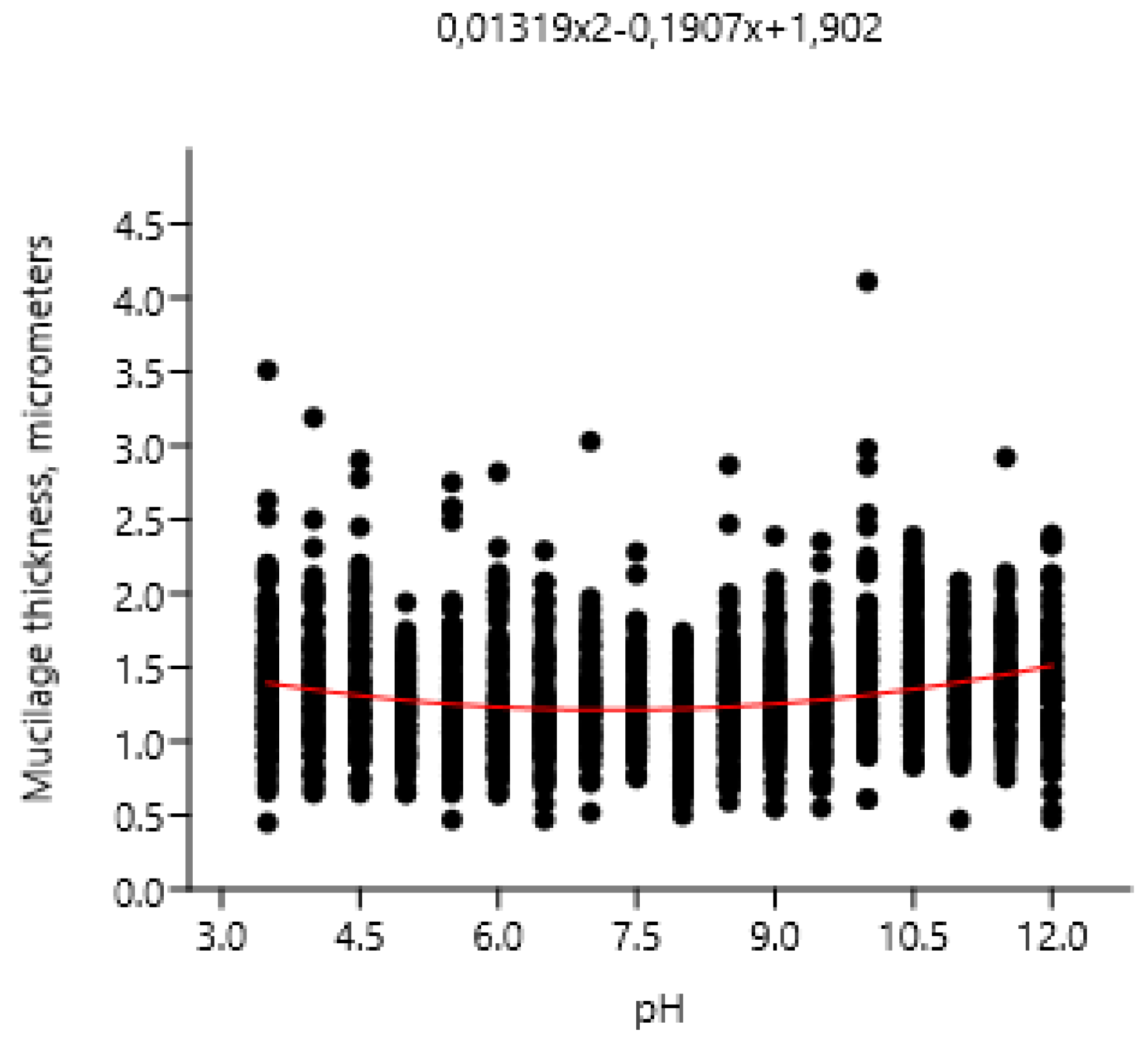
Figure 7.
Influence of pH on Chlorella vulgaris morphology. (A) control variant (рН 6.5); (B) рН 2; (C) pH 3.5; (D) рН 4; (E) рН 11.5; (F) рН 12. Scale bar 10 μm.
Figure 7.
Influence of pH on Chlorella vulgaris morphology. (A) control variant (рН 6.5); (B) рН 2; (C) pH 3.5; (D) рН 4; (E) рН 11.5; (F) рН 12. Scale bar 10 μm.
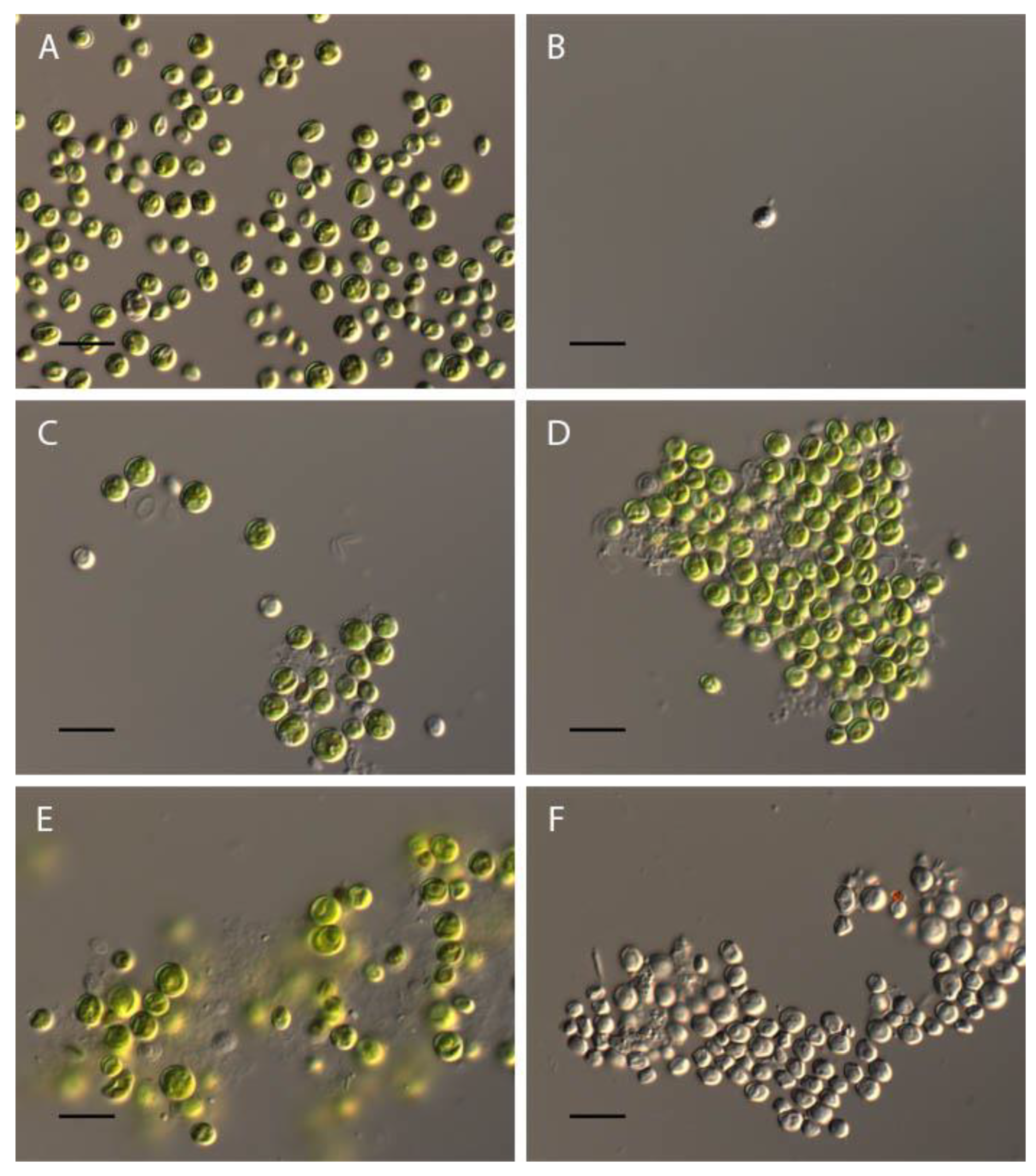
Figure 8.
Influence of pH on cell volume of Chlorella vulgaris.
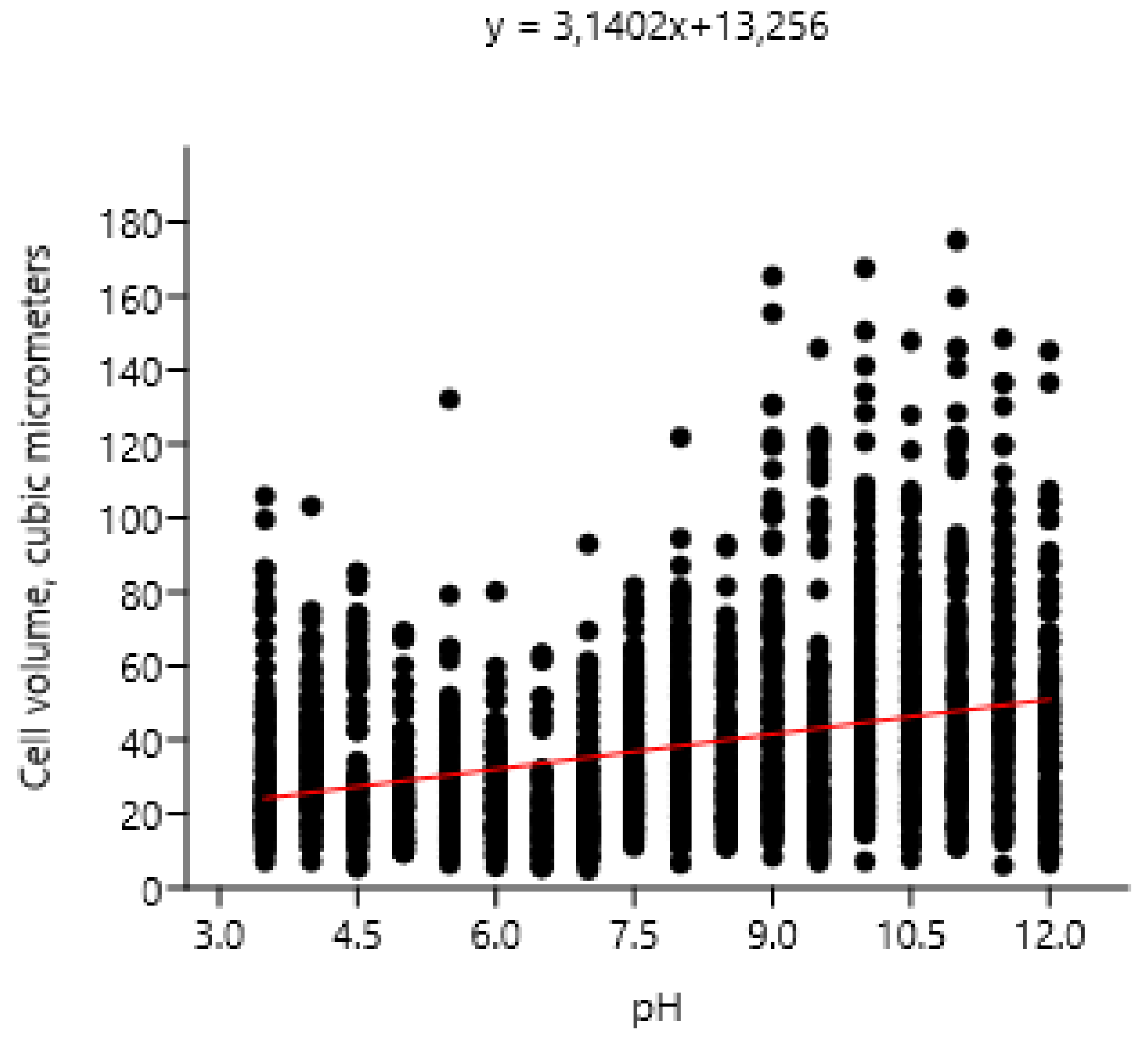
Figure 9.
Influence of pH on Pseudococcomyxa simplex morphology. (A) control variant (рН 6.5); (B) рН 2; (C) pH 2.5; (D) рН 3.5; (E) рН 8.5; (F) рН 12. Scale bar 10 μm.
Figure 9.
Influence of pH on Pseudococcomyxa simplex morphology. (A) control variant (рН 6.5); (B) рН 2; (C) pH 2.5; (D) рН 3.5; (E) рН 8.5; (F) рН 12. Scale bar 10 μm.
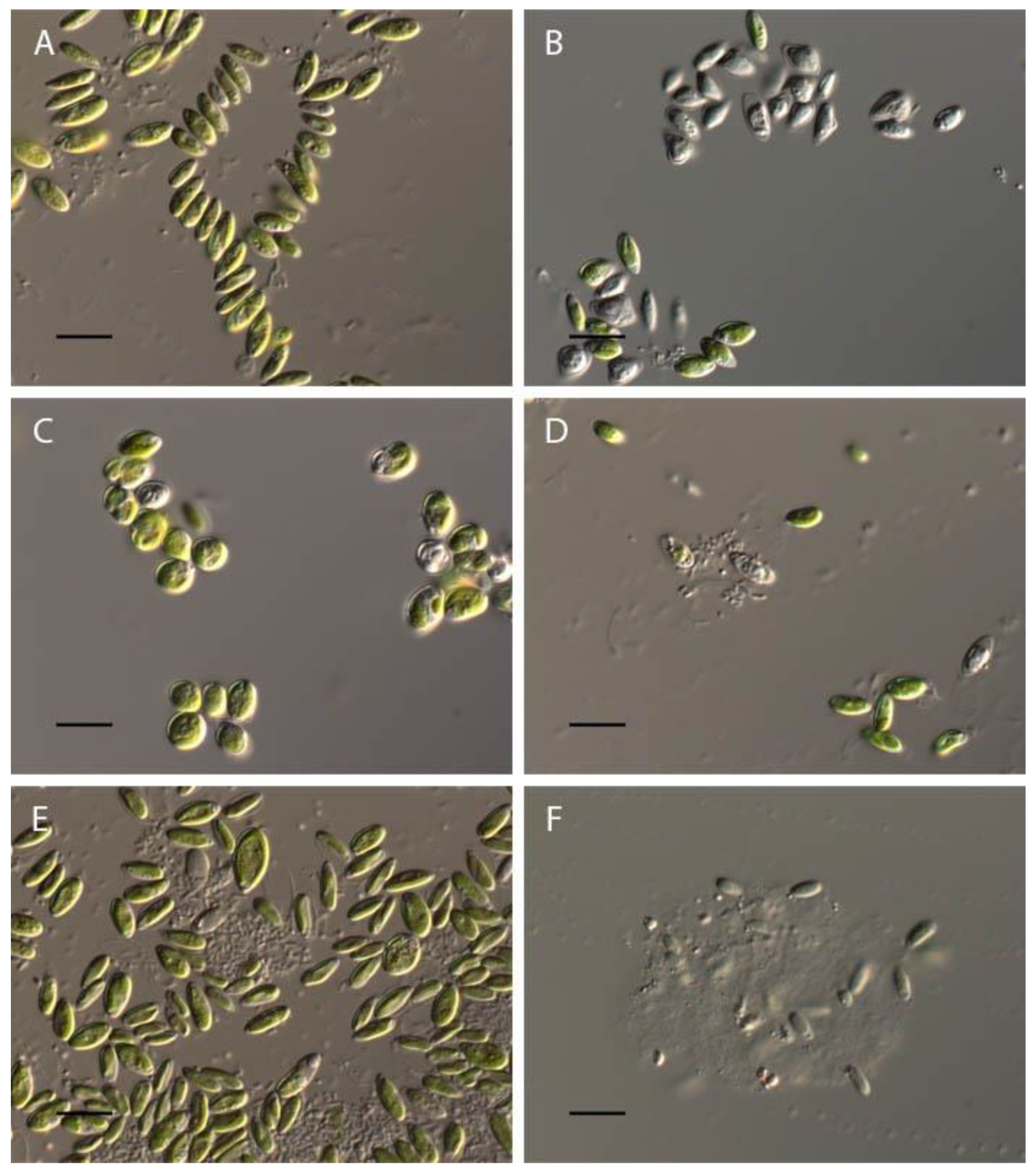
Figure 10.
Influence of pH on cell volume of Pseudococcomyxa simplex.
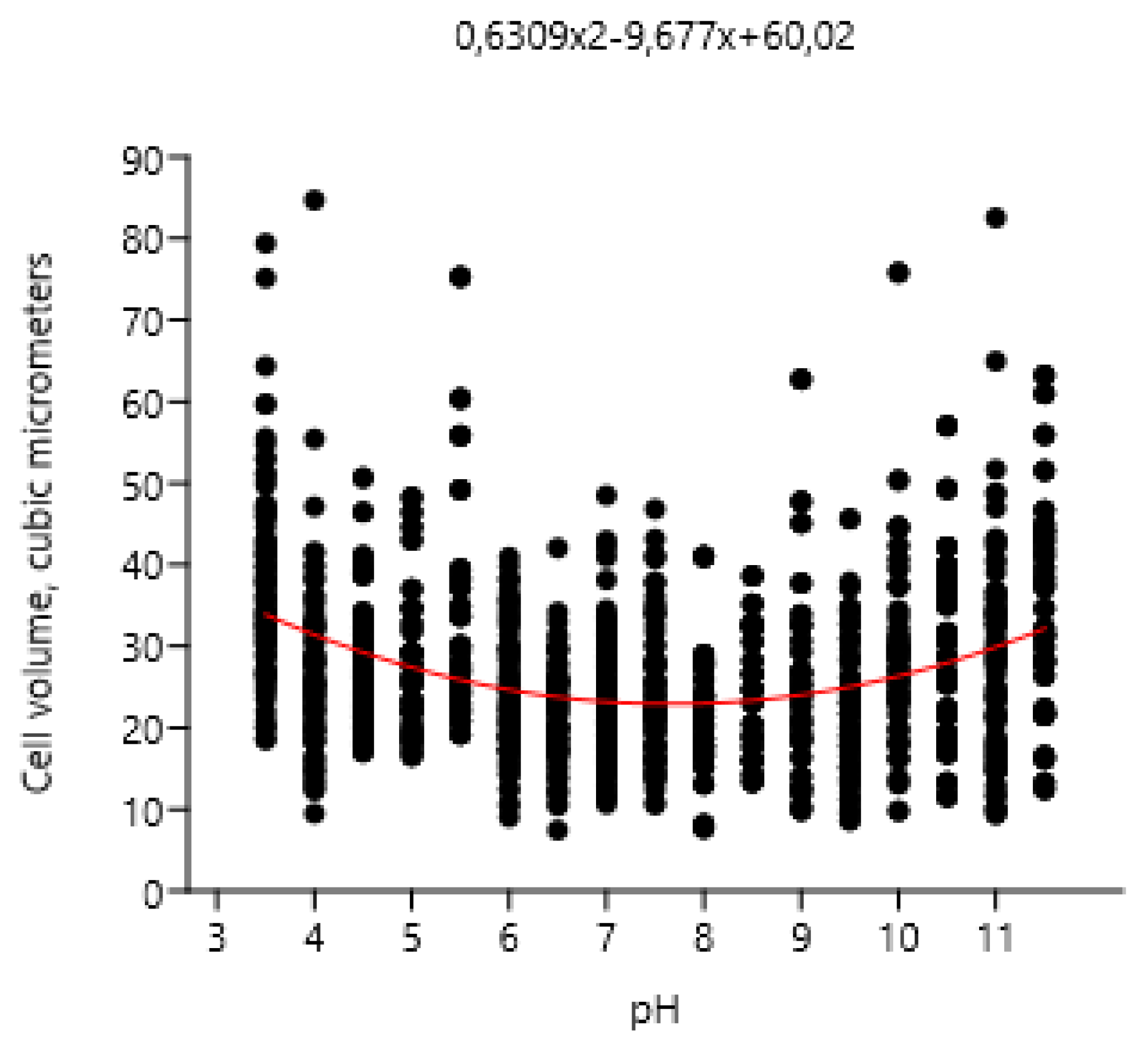
Figure 11.
Influence of pH on Vischeria magna morphology. (A) control variant (рН 6.5); (B) рН 2.5; (C) pH 3; (D) рН 3.5; (E) рН 11.5; (F) рН 12. Scale bar 10 μm.
Figure 11.
Influence of pH on Vischeria magna morphology. (A) control variant (рН 6.5); (B) рН 2.5; (C) pH 3; (D) рН 3.5; (E) рН 11.5; (F) рН 12. Scale bar 10 μm.
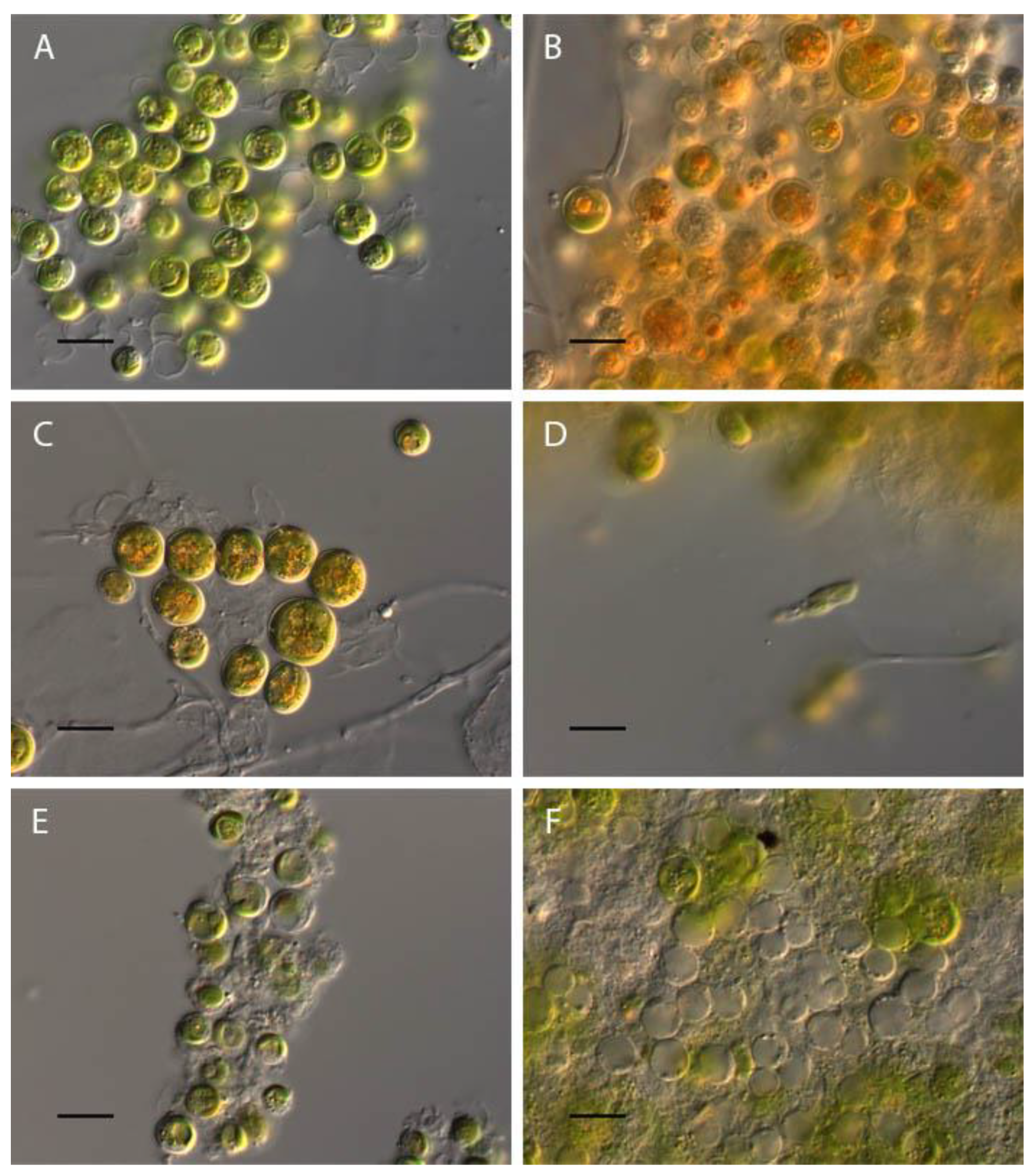
Figure 12.
Influence of pH on cell volume of Vischeria magna.
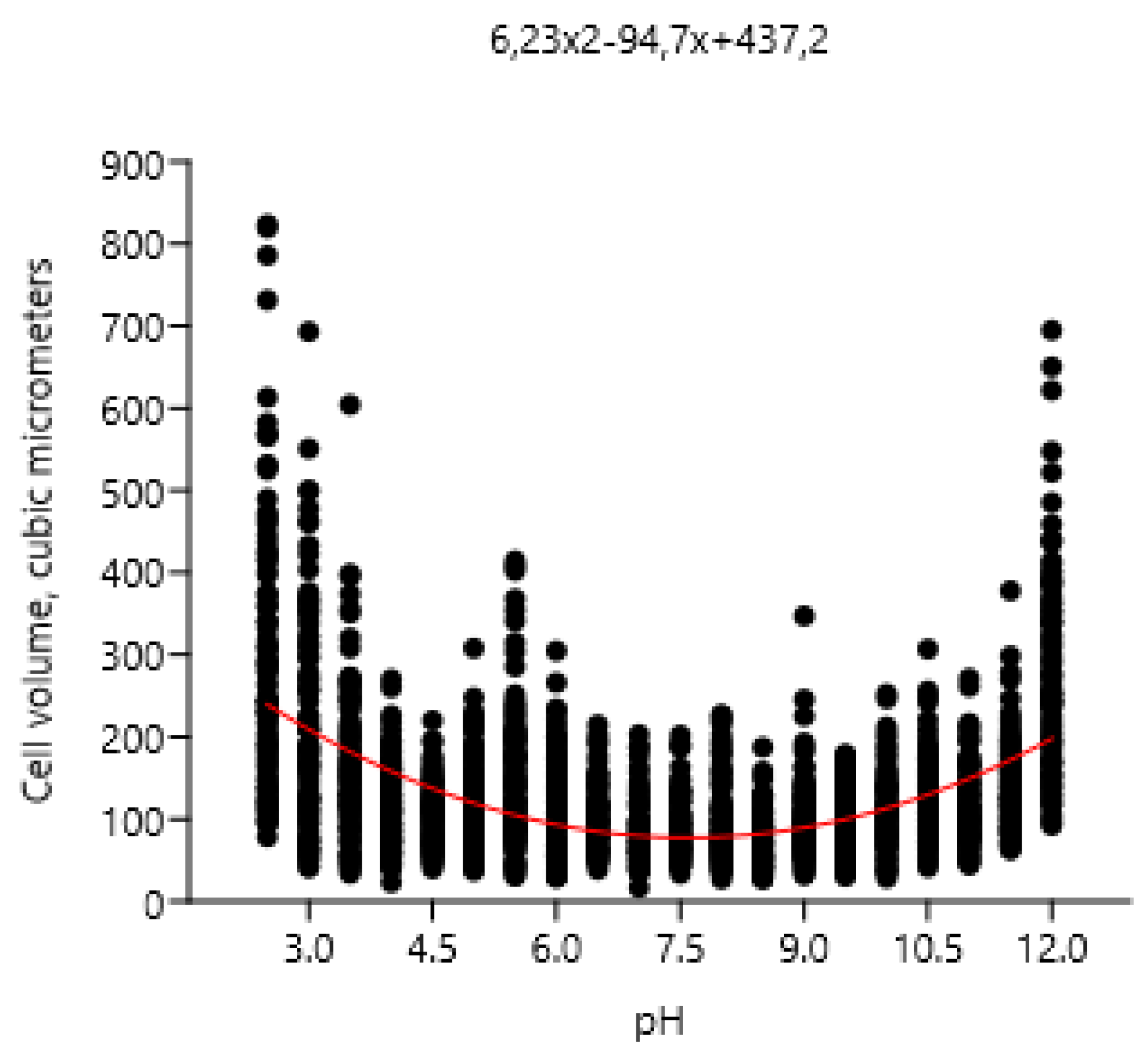

Table 1.
The limits of pH and morphological disturbances at low and high pH.
| Taxa | pH limits | Morphological and physiological features changes at pH< 4 | Morphological and physiological features at pH>10 |
|---|---|---|---|
| Bracteacoccus minor | 4-10 | Complete discoloration and destruction of cells, appearance of orange granules | Discoloration and destruction of cells |
| Chlorococcum infusionum | 4-9,5 | Discoloration and destruction of cells, increase of mucilage production, appearance of large granules in cytoplasm | Discoloration and destruction of cells, increase of mucilage production, appearance of large granules in cytoplasm |
| Chlorella vulgaris | 4-11,5 | Discoloration of cells | Discoloration of cells |
| Pseudococcomyxa simplex | 4-11,5 | Discoloration and "wrinkling" of cells, appearance of the almost round cells | Discoloration of cells |
| Vischeria magna | 3,5-11 | Discoloration and appearance of the orange granules in cells, maximal zoospores production at pH 3.5, increasing of cell volume | Discoloration of cells |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



