
Submitted:
10 November 2023
Posted:
22 November 2023
You are already at the latest version
Abstract
Epigenetic modifications play a significant role in determining the fate of AFSC in terms of differentiation into multiple lineages. There is a tight correlation among DNA methylation, histone modifications, and small noncoding RNAs during the epigenetic control of stem cells differentiation. The present study investigates the role of miRNAs in differentiation of AFSC and addresses how their unique signatures contribute to lineage-specific differentiation. miRNAs profile was assessed in AFSC after 4 weeks of endothelial and muscular differentiation. Our results showed a decreased expression of for 5 miRNAs (miR-125b-5p, hsa-miR-18a-5p, hsa-miR-137, hsa-miR-21-5p and hsa-let-7a-5p) and an increased expression for 12 miRNAs (miR-134-5p, miR-103a-3p, let-7i-5p, miR-214-3p, let-7c-5p, miR-129-5p, miR-210-3p, let-7d-5p, miR-375, miR-181-5p, miR-125a-5p, hsa-let-7e-5p) in EPC compared with undifferentiated AFSC. Smooth muscle differentiation of AFSC revealed significant changes in 9 miRNAs from total of 84 tested; 3 were downregulated (miR-18a-5p, miR-137, sa-miR-21-5p) and 6 were upregulated (miR-155-5p, miR-20a-5p, let-7i-5p, hsa-miR-134-5p, hsa-miR-214-3p, hsa-miR-375). miRNAs are molecular regulators of cell fate and differentiation, recent advances in molecular biology techniques revealed miRNA regulatory networks that can provide us information of endothelial and muscle cell biology in terms of homeostasis, growth, differentiation, and vascular function.
Keywords:
microRNA
; stem cells
; differentiation
; endothelial progenitor cells
; smooth muscle progenitor cells
1. Introduction
The amniotic fluid contains a variety of cells with broad therapeutic potential, featuring immunomodulatory and wound healing effects without tumorigenic risk, lack of ethical and legal limitations specific to other stem cell preparations, and high differentiation capacity to various lineages. All these properties make them ideal candidates for regenerative medicine applications [1]. Amniotic fluid cells represent a heterogeneous group of mesenchymal cells with foetal origin, with plenty of implications in regenerative medicine. These populations has low immunogenicity, presenting on the surface only HLA class I antigens, so it is less likely to trigger an immune response [2]. Phenotypes observed after culturing the amniotic fluid include epithelial-like cells and fibroblast-like cells. About 0.1–0.5% of stem cells showed c-kit (CD117+) expression. The renewing potential of amniotic fluid derived mesenchymal stem cells (AFSC) emerges from their differentiation potential and reduced immune and inflammatory responses at the implantation site [3]. AFSC can specialize into various cell lines such as bone, nervous, vascular, and cardiac [4,5,6]. Specific differentiation protocols and conditioning media have been devised for cultured AFSC, but seemingly non-mesenchymal derivatives could be obtained only from c-kit positive stem cell clones isolated from amniotic fluid cell pools [4]. When grown in hydrogels, AFSC have the potential to form blood vessels-like networks. On the other hand, when AFSC are indirectly cultivated with cardiac cells, they modulate cardiac regeneration, due to paracrine signalling [5,6,7]. Epigenetic alterations have a substantial role in deciding the fate of AFSC in terms of their differentiation into several tissue lineages. Throughout the epigenetic control of stem cells differentiation there are several processes involved such as DNA methylation, histone alterations, and small noncoding RNAs; however, the precise mechanisms are still unclear so far [8]. microRNAs (miRNAs) are a set of small noncoding RNAs, with lengths of 15–25 base pairs in the mature single-stranded form, modulating various biological processes by regulating the post-transcriptional alterations of specific genes [9]. miRNA can control hundreds of genes involved in both adaptive and innate immune response, as well in growth, differentiation, apoptosis and senescence of the cells by targeting hundreds of messengers RNAs [10]. AFSC have notable potential to differentiate into different cell types, but the epigenetic mechanisms that are associated with their multilineage differentiation ability is still not clearly understood. Studies involving miRNA in AFSC differentiation was initiated starting with year 2014 when it has been demonstrated that AFSC showed a significant capacity to differentiate towards chondrogenic and osteogenic cellular types but not in adipocyte type, due to expression of miR-21, primarily involved in osteogenesis differentiation [11,12]. These results were also demonstrated by Gholizadeh-Ghaleh Aziz et al., who showed that miR-Let7g and miR-302a might be involved in interaction with Nanog and POU5F1 in processes like differentiation, adipogenesis and osteogenesis [13]. Glemžaitė and Navakauskienė highlighted that osteogenic differentiation of AFSC is mediated by specific microRNA expression (miR-17, and miR-148b), chromatin-modifying enzymes, histone modifications (H3K9ac, H4 hyperAc, H3K27me3) [14]. Current literature data shows that miRNAs can control MAPK, WNT, and TGF-β signalling pathways playing a crucial role in the fate of AFSC [15]. Analysis AFSC between passages 1 and 15 by Miranda-Sayago et al., proved major variation regarding the expression of tested miRNAs, generally related to a decrease expression of TP53 and an increased expression of miR-125a, which is an indicator of proliferation and stemness [16]. In this context, AFSCs can be a remarkable study model due to the potential benefits of their use in regenerative medicine. Nevertheless, a better understanding of the basis of multiple-lineage potential might help to successfully utilize these cells for therapeutic purposes [17,18]. The present study investigates the role of miRNAs in differentiation of AFSC and reports how their unique signatures can promote the lineage-specific differentiation.
2. Results
2.1. AFSC differentiated in endothelial and smooth muscle progenitor cells
After 4 weeks of stimulation with endothelial-differentiation growth factors and endothelial specific medium, cultures of adherent AFSC started to form colonies with epithelial-like cobblestone morphology (Figure 1A). The differentiation of AFSC was validated by the representative functional characteristics of endothelial progenitor cells. Cultured on Matrigel for 24 hours, EPC form vessel-like structures (Figure 1B), and after incubation with acetylated-LDL and Ulex europaeus lectin, these cells incorporated and binding these tracers (Figure 1D). The gene expression assay showed increased mRNA expression for: CD31 (151-fold), eNOS (106-fold), CD144 (26-fold) and wVF (6.6-fold); these factors control EPC proliferation, adherence, differentiation, and survival (Figure 1C).
Stimulation of AFSC with FGF, EGF, heparin, IGF, and BSA promoted differentiation toward smooth muscle cells, inducing some phenotypic changes such as spindle shape morphology and elongated actin-rich protrusions could be observed in Figure 1C. The smooth muscle differentiation of AFSC was confirmed by the upregulation of mRNA expression levels for calponin-1 (4.7-fold), smoothelin (50-fold), myosin-heavy chain (1.47-fold), troponin (13-fold), smooth muscle actin (2.7-fold), caldesmon (2.08-fold), cav3.1 (2-fold) (Figure 2B). Whole-cell patch-clamp experiments in the classical (ruptured) or β-escin-perforated configuration were performed on a total number of 24 SPCs, with application of different pharmacological compounds (Tables S1 and S2 of Supplemental data). All cells featured outward-rectifying K+ currents. BK currents (with standard deviation of current fluctuations measured over the last 250 ms of the depolarizing step at +60 mV exceeding 5 pA) were present in 91.7% of the cells, inactivatable outward K+ currents were present in 41.7% of cells), while INa was recorded in only one cell, a major difference compared to previous results on cultured AFSC [19,20]. The inactivatable outward K+ current component was partly inhibited by phrixotoxin-1 (Figure S1 of Supplemental data), proving it was produced by Kv4.2/4.3 channels, while other inhibitors (DIDS, glibenclamide, NaCN) did not exert significant effects. The patch-clamp experiments demonstrated the presence in SPC of calcium-activated potassium channels with big conductance (BK channels – KCa1.1), producing an outward current component at positive potentials when applying a double-ramp protocol (Figure 2C), which could be‘ blocked by iberiotoxin 100 nM. Furthermore, in 1 of 24 cells we recorded two inward current components with different activation thresholds and I-V plots suggestive of ICaT and ICaL, and in another cell only the ICaT component. The fast component was resistant to application of nifedipine 1 μM, but it was inhibited by subsequent application of mibefradil 10 μM (Figure S2 of Supplemental data). These results suggest the differentiation of AFSC into SPC (Figure 2D). The presence of T-type calcium channels was confirmed by the mRNA expression levels of cav3.1.
2.2. miRNA regulates the differentiation of AFSC
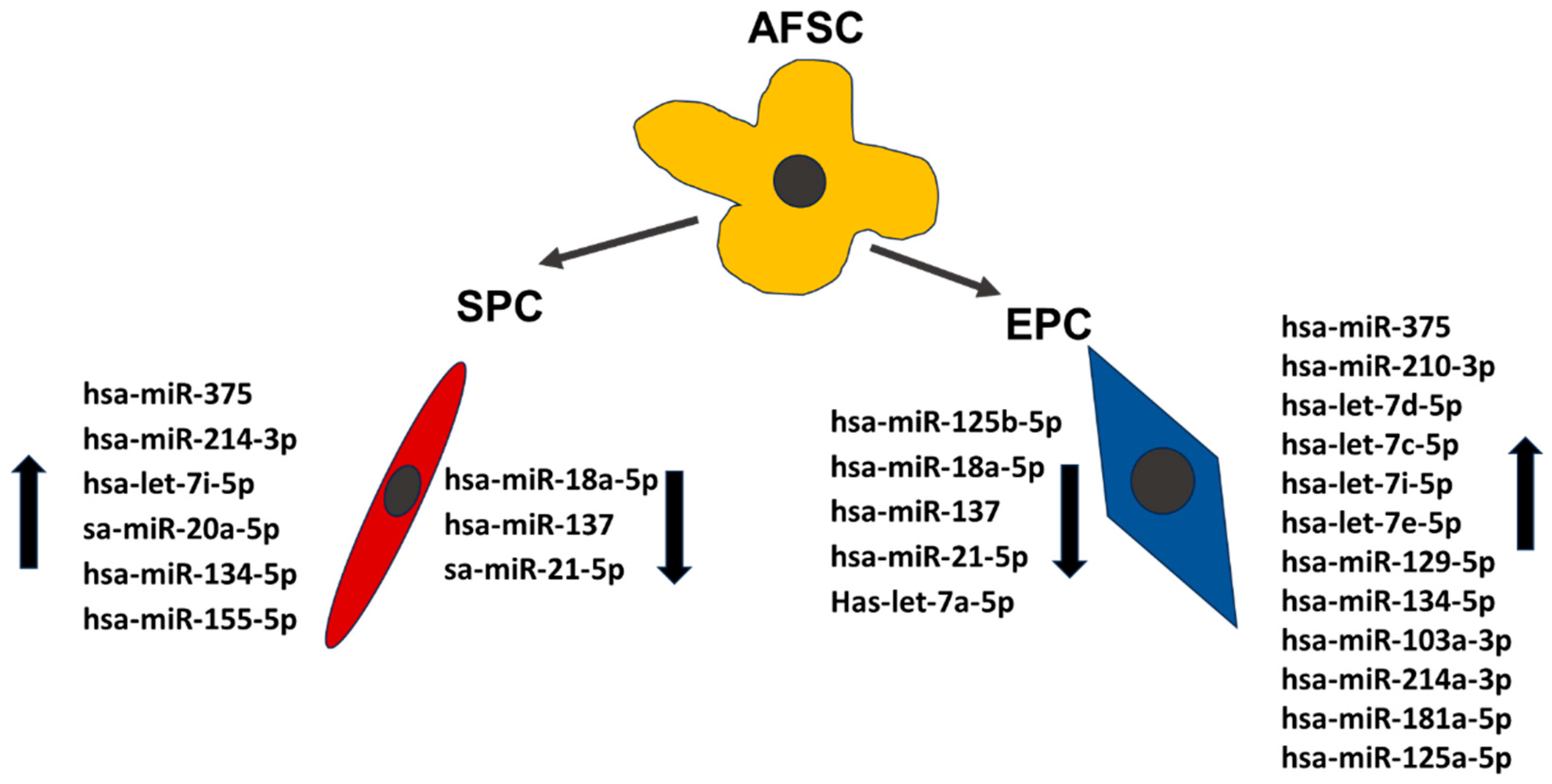
The investigation of the miRNAs profile using “Human Cell Differentiation & Development miRNA array” in AFSC after 4 weeks of endothelial and muscular differentiation stimulation revealed changes in miRNA expression. Among the 84 arrayed miRNAs, EPC derived from AFSC showed increased expression of miR-125a-5p (2.75-fold), miR-181a-5p (2.75-fold), miR-134-5p (7.62-fold), miR-103a-3p (4.89-fold), let-7i-5p (17.39-fold), miR-214-3p (4.20-fold), let-7c-5p (7.36-fold), miR-129-5p (7.46-fold), miR-210-3p (7.11-fold), let-7d-5p (4.86-fold), miR-375 (11-fold). Decreased expression was reveled for let-7e-5p (-3.48-fold), miR-125b-5p (-8.57-fold), miR-18a-5p (-2.69-fold), miR-137 (-2.07-fold), miR-21-5p (-2.03-fold), let-7a-5p (-2.83-fold) (Figure 3).
SPC derived from AFSC showed increased expression of miR-155-5p (4.38-fold), miR-134-5p (2.33-fold), miR-20a-5p (3.16-fold), let-7i-5p (3.78-fold), miR-214-3p (2.48-fold), miR-375 (2.17-fold) and a downregulation of: miR-137 (-9.85-fold), miR-18a-5p (-12.82-fold), miR-21-5p (-2.28-fold) (Figure 4).
3. Discussion
Recent miRNAs studies established a signature that is typical for embryonic stem cells [17,18]. The miRNA clusters act as cell cycle mediators increasing embryonic stem cells proliferation promoting transition from the G phase to S1 phase, other such as miR-291-3p, miR-295 and miR-294, families control pluripotency factors (Oct4, Sox2 and Nanog). By acting on these transcription factors they promote somatic reprogramming, activating mesenchymal-to-epithelial changes necessary for colony formation, differentiation, and inhibition of cellular senescence [19]. Our results showed a decreased expression of hsa-miR-18a-5p, miR-125b-5p, hsa-miR-137, hsa-miR-21-5p and hsa-let-7a-5p in EPC compared with young AFSC (Figure 5). These results agree with other research, who showed that low expression level of miR-125 in endothelial cells may represents the “youth code” that maintains the normal function in human endothelial cells. The increased expression of miR-125 in endothelial cells of aged mice can regulate angiogenesis by targeting RTEF-1 and regulate the expression of VEGF and eNOS [20]. Furthermore, let-7e controls tube formation and migration of endothelial cells via targeting FASLG. Let-7e overexpression activates Th1 and Th17 cells and regulates the inflammatory responses of endothelial cells, directly or indirectly by targeting different inflammatory genes [21]. Among the 84 arrayed microRNAs, EPC derived from AFSC showed increased expression in 12 miRNA (miR-134-5p, miR-103a-3p, let-7i-5p, miR-214-3p, let-7c-5p, miR-129-5p, miR-210-3p, let-7d-5p, miR-375, miR-181-5p, miR-125a-5p, hsa-let-7e-5p) (Figure 5). Zang et al., showed that vildagliptin (dipeptidyl peptidase-4 inhibitor), attenuated endothelial dysfunction in diabetic rats through miRNAs molecule. They demonstrated that vildagliptin inhibit the expression of miR-134-5p that stimulates BDNF and regulates endothelial function [22]. miR-103a-3p promotes EPC migration and angiogenesis, being downregulated in EPCs isolated from patients with deep vein thrombosis. Downregulation of miR-103a-3p is involve in endothelial progenitor cell dysfunction by activation of PTEN protein [23]. Yamakuchi profiling of microRNA in human aortic endothelial cells showed that miR-21, miR-29, miR-126, family, and let-7 family are abundant in endothelial cells [24], in correlation with our results that showed an increased expression of let-7 family. The miR-210 plays an important role in angiogenesis. Several studies suggested that upregulated miR-210 could inhibit proliferation and induce apoptosis in endothelial progenitor cells, damaging the angiogenic properties under oxygen and glucose deprivation conditions in endothelial progenitor cells [25]. Wang et al. reported that miR-210 significantly reduce HIF-1 expression, which affects the mobilization of bone marrow endothelial progenitor cells [26]. Moreover, in vivo study showed that miR-210 decrease the mobilization effect of SDF-1α / CXCL12 on bone marrow-derived angiogenic cells. All these findings suggested that miR-210 is closely related to the number and function of endothelial progenitor cells [27]. Shi et al., found that miR-375–3p was significantly up-regulated following high glucose stress. Functional assays showed that miR-375 regulates the proliferation, migration, tube formation and apoptosis of EPCs in vitro [28]; in our study the level of miR-375 increased 11-fold compared with AFSC. The miR-17–92 cluster is a polycistronic miRNAs localized in intron 3 of the 13q31.3. chromosome. MiR-17–92 contains 6 individual miRNAs: miR-17, miR-18a, miR-19a, miR-20a, miR-19b, and miR-92a implicated in cell proliferation, suppressing apoptosis in cancer cells, and induces tumor angiogenesis. miR-17–92 is stimulated upon VEGF treatment in endothelial cells. The activation of the miR-17–92 cluster by Elk-1 is necessary for endothelial cell proliferation and vascular sprouting. Laminar flow increased miR-21 and nitric oxide synthesis acting by phosphatidylinositol-4,5-bisphosphate 3-kinase/Protein Kinase B Alpha pathway regulating endothelial cell functions. Oscillatory shear flow induces miR-21 expression stimulating endothelial inflammation mediated by peroxisome proliferators-activated receptor mechanisms [29]. Endothelial cells synthesize endothelin-1 (ET-1), one of the most effective vasoconstrictors, its expression and secretion being stimulated by ischemia, hypoxia, and shear stress. MiR-125a/b is engaged in regulation of ET-1 factor secreted by the endothelial cells. Moreover, its expression has been shown to be regulated by oxidized LDLs [30]. Also, in our experiments observed a decreased expression of miR-125b, let-7a but an increased expression of miR-125a in endothelial progenitor cells and avidity for uptake oxidized LDL. Oxidized LDL particles accumulated in the atherosclerotic lesions, enhance the expression of proinflammatory molecules, leading to dysfunction of vascular endothelial cells. Ox-LDL cause downregulation of let-7g in endothelial progenitor cells by interaction of octamer binding transcription factor-1 to the let-7g promoter [31]. Interestingly, also let-7a and let-7b, target the lectin-like LDL-receptor1, which is the receptor for ox-LDL in endothelial cells [32].
Smooth muscle differentiation of AFSC using M231 Medium supplemented with bFGF, EGF, heparin, IGF, and BSA revealed significant changes in nine microRNAs from total of 84 tested; three were downregulated (miR-18a-5p, miR-137, sa-miR-21-5p) and six were upregulated (miR-20a-5p, miR-155-5p, let-7i-5p, hsa-miR-134-5p, hsa-miR-214-3p, hsa-miR-375) compared with AFSC. Pan et al. showed that miR-137 inhibits VSMCs migration and proliferation. They used bioinformatics algorithms and luciferase reporter assay and found that IGFBP-5 (insulin-like growth factor binding protein-5) was a direct target of miR-137 in VSMCs. Also, they demonstrated that MiR-137 overexpression suppressed the activity of mTOR/STAT3 signaling pathway [33]. Furthermore, the miR-18a-5p expression was increased in differentiated VSMCs, whereas it decreased in de-differentiated VSMCs. Syndecan 4 is a target of miR-18-5p in VSMCs; overexpression of syndecan 4 reduced Smad2 expression, whereas knockdown of syndecan4 increased Smad2 level in vascular smooth muscle cells [34]. The miR-155-5p function as an inhibitor in VSMC migration and vascular remodeling in hypertension state. In addition, miR-155-5p direct inhibit the levels of BACH1, and suppress the formation of ROS in SHR VSMCs. Moreover, deletion of miR-155-5p increased the expression of BACH1, and support the production of superoxide anions, in simultaneousness with higher protein expression of NOX2, NOX4, two NAD(P)H oxidases as the main source of ROS in cardiovascular cells. Finally, overexpression of miR-155-5p and BACH1 downregulation repressed the migration of VSMCs. The suppression of BACH1 by miR-155-5p was sufficient to decrease the migration and oxidative stress in VSMCs [35]. A recent study published by Li et al using a knock-out model of miR-214 in VSMC investigated the role of miR-214 in vascular smooth muscle cells by quantification the expression of contractile markers including smooth muscle actin, smooth muscle myosin heavy chain, and calponin. Their results showed that the expressions of these markers were significantly decreased in miR-214 KO VSMC [36]. Our results consent these, by the presence of these contractile markers in differentiated smooth muscle progenitors that corelates with an increased expression of miR-124.
4. Materials and Methods
4.1. Amniotic fluid stem cells differentiation and characterization
The amniotic fluid cell cultures were kindly given by S.C. Personal Genetics S.R.L. diagnostics laboratory upon informed consent of the patients in accordance with the Declartion of Helsinki and national and EU rules. The primary AFSC cultures were obtained by centrifugation of amniotic fluid for 10 min. at 300g The cells were then cultured for 10 days without passages in AmnioMax medium (CN 11269016, ThermoFischer Scientific, Waltham, Massachusetts, USA), with medium changes at every 2 days (37°C, 5%CO2, 21%O2 in a humidified atmosphere). Subsequently 10 days the primary cultures were passaged and cultured in differentiation specific medium supplemented with growth factors. Differentiation of AFSC in endothelial progenitor cells was performed by culturing cells in M200 Medium with 10% FBS (fetal bovine serum), 20 ng/mL insulin growth factor (IGF-1), 40 ng/mL vascular endothelial growth factor (VEGF), 10 ng/mL epidermal growth factor (EGF), 10 ng/ml basic fibroblast growth factor (bFGF), 100 μg/mL streptomycin, 100 μg /mL penicillin, and 50 μg/mL neomycin (Thermo Fischer Scientific, Massachusetts, USA). Smooth muscle cell differentiation of AFSC was obtained by culturing in M231 Medium supplemented with bFGF (2 ng/mL), heparin (5 ng/mL), EGF (0.5 ng/mL), IGF-1 (2 µg/mL) and bovine serum albumin (BSA) (0.2 µg/mL) (Thermo Fischer Scientific, Massachusetts, USA). The cells were maintained in these media for 4 weeks, and passaged when reaching subconfluence (37°C, 5%CO2, 21%O2 in a humidified atmosphere) [37,38].
4.2. Gene expression and functional characterization of EPC
Expression levels of mRNA were assessed by qRT-PCR. Total RNA was isolated from AFSC using RNeasy Mini Kit (Qiagen, Hilden, Germany) and reverse-transcription reaction was carried out using M-MLV polymerase (High-Capacity cDNA Reverse Transcription kit, Thermo Fischer Scientific, USA). mRNA levels of endothelial related genes (ICAM-1, PECAM-1, eNOS, VE-Cadherin, and vWF) were quantified using TaqMan hydrolysis probes (ThermoFischer Scientific, Massachusetts, USA) (Table S3). Quantitative RT-PCR reactions were performed in a real-time thermocycler (TaqMan™ Universal PCR Master Mix, ViiA7, Applied Biosystems, USA), respecting manufacturer’s guidelines. The results were expressed using relative quantitation (2-ΔCt), where ΔCT represents CT difference between values for AFSC and EPC. Dil-Ac-LDL uptake assay. AFSC-derived EPC were incubated with 6 µg/mL Dil-AcLDL-PE (acetylated low density lipoprotein pycoeritrine conjugated, ThermoFischer Scientific, USA) for 2 h at 37 °C with 5% CO2, washed with PBS and fixed with 1% paraformaldehyde (PFA) for 10 minutes at room temperature. Ulex europaeus agglutinin (UEA) binding capacity to EPC. EPC, were incubated with 0.01 mg/mL FITC – Ulex europaeus lectin (Sigma-Aldrich, St. Louis, USA) 2 hours, followed by PBS wash. The nuclei of EPC were counterstained with 1 mg/mL DAPI (ThermoFischer Scientific, Waltham, Massachusetts, USA). The photomicrographs were captured with a digital camera (Digital Net Camera DN100) using an TE300 Eclipse microscope (Nikon, Tokyo, Japan). Matrigel tube-formation assay. Evaluation of blood vessel networks in Matrigel, was made by seedding the EPC into 96-well plates at a density of 3,000 cells per well. Briefly, 50 μL of Matrigel (Sigma-Aldrich, St. Louis, USA) was added in each well of 96-well plate, and left for gelation 30 minutes at 37ºC. After polymerization of Matrigel, EPC suspension was added and incubated for 24 h. Tubes formation was assesed using the same microscope-digital camera system.
4.3. Gene expression assays on smooth muscle progenitor cells (SPC)
mRNA levels of muscular-associated genes (smoothelin, calponin 1, α-SMA (α-actin), Myh11 (myosin heavy chain), α-tropomyosin, caldesmon-1, Cav3.1) were quantified using TaqMan hydrolysis probes (ThermoFischer Scientific, Waltham, Massachusetts, USA) (Table S3).
4.4. Patch-clamp assays on smooth muscle progenitor cells (SPC)
In patch-clamp experiments we used 1.5/0.86 mm diameter borosilicate glass capillaries with inner filament (Gc150f-10, Harvard Apparatus, Holliston, USA) pulled in four steps with a PUL-100 microprocessor-controlled equipment (WPI, Sarasota, FL, USA) and fire-polished with a microforge to yield a resistance in solution between 2-3 MΩ. The patch-clamp system included an inverted microscope (IMT-2, Olympus, Tokyo, Japan) placed on an antivibratory stand in a Faraday cage, a temperature controller (TC202A, Harvard Apparatus, MA, USA), a resistive feedback amplifer (WPC-100, Göttingen, Germany) connected to a Digidata 1322AAD/DA interface controlled by the pClamp software (Axon Instruments, Sunnyvalle, CA, USA). The extracellular solution was composed of: 135mM NaCl, 1.8 mM CaCl2, 5.4 mM KCl, 0.9 mM MgCl2, 10 mM HEPES, 0.33 mM NaH2PO4, 10 mM D-glucose pH 7.40 at 25°C titrated with NaOH. The pipette solution contained: 5 mM EGTA, 140 mM KCl, 10 mM HEPES, pH 7.21 at 25°C titrated with KOH. Some of the experiments were performed in the perforated whole-cell configuration by adding 15 to 30 μM β-escin (Sigma E1378) to the pipette solution [39]. SPCs cultured in 24-well plates (some of them pre-incubated with nifedipine 1 μM in the culture medium for 24-48 h) were detached with accutase (Sigma A6964) (7 min at 37˚C), centrifuged, disposed in 35 mm Petri dishes in external solution and retained for 30 min at 37˚C for reattachment. Several voltage protocols were applied: a double voltage ramp protocol (from -120 mV to +80 mV in 2s and back to -120 mV in the same time, with a holding potential of -70 mV); a standard multi-step voltage protocol to assess several voltage-dependent currents, including T-type and L-type Ca2+ currents (ICaT and ICaL) and transient outward K+ currents (Ito) (holding potential -80 mV, 300 ms voltage steps from -60 mV to +60 mV in 10 mV increments); a protocol for voltage-dependent Na+ currents (INa) (holding potential -100 mV, 6 ms voltage steps from -60 mV to +40 mV in 10 mV increments); a voltage protocol to separate ICaL from INa based on different voltage dependence of steady-state inactivation [40]. The following pharmacological compounds were applied in individual experiments: the specific BK channel blocker iberiotoxin (STI-400, Alomone Labs, Jerusalem, IL) at 100nM, the specific Ito fast (Kv4.2 and 4.3) peptide inhibitor phrixotoxin-1 (Abcam ab141844) at 280 nM, the moderately T-type selective Ca2+ channel inhibitor mibefradil (Sigma M5441) at 5 or 10 μM (estimated IC50 for ICaT 2.7 μM and for ICaL 18.6 μM), the L-type Ca2+ channel inhibitor nifedipine (Sigma N7634) at 1 μM, the non-selective anion exchange inhibitor 4, 4′-diisothiocyano-2, 2′-stilbenedisulfonic acid (DIDS) (Calbiochem 309795) at 100 μM, the sulfonylurea receptor inhibitor glibenclamide (Sigma G0639) at 100 μM, the cytochrome inhibitor sodium cyanide (Sigma-Aldrich 380970) at 5 mM, and β-adrenergic receptor agonist isoprenaline (Sigma I5627) at 1 μM.
4.5. miRNA expression
The expression of miRNA was assessed using The Human Cell Differentiation & Development miScript miRNA PCR Array (Qiagen, Hilden, Germany) that profiles the expression of 84 miRNAs differentially expressed during cellular differentiation and organism development. The data was analyzed using GeneGlobe Data Analysis Center platform (Qiagen, Hilden, Germany).
4.6. Data analysis
Data are described as mean ± SD and mean ± SEM, “n” showing the samples number. Statistical significance was evaluated using either Student’s t two-tailed test for independent samples or its non-parametric variant (Mann-Whitney test), according to results of normality tests, for quantitative data, and Fisher’s exact probability test for categorical frequency data. For the statistical tests was set a critical level p = 0.05. The datasets produced during this study are available from the corresponding author on reasonable request.
5. Conclusions
Stem cells own specific miRNAs profiles which regulate stem cell fate. Stem cells differentiation controlled by miRNA could contribute in obtaining better populations of smooth muscle and endothelial cells for the benefit of regenerative medicine. miRNAs are molecular regulators of cell fate and differentiation during development, recent advances in molecular biology techniques established miRNA regulatory networks that can provide us information of endothelial and muscle cell biology in terms of homeostasis, growth, differentiation, and vascular function. The understanding of miRNAs mechanisms of regulation of cell fate specification and differentiation could be used to differentiate cells to treat numerous diseases, including muscle diseases, neurodegenerative diseases, blood diseases, and cardiac infarction.
Supplementary Materials
The following supporting information can be downloaded at: Preprints.org. Table S1: General electrical characteristics of SPC from the n = 24 whole-cell patch-clamp experiments and applied pharmacological compounds; Table S2: Analysis of principal ionic current components recorded in SPC from the n = 24 whole-cell patch-clamp experiments; Figure S1: Inhibitory effect of phrixotoxin 1 (280 nM) on the Ito fast component, highlighted in the general IK voltage-clamp protocol at the depolarization step at +50 mV; Figure S2: Lack of effect of 1 μM nifedipine and inhibitory effect of 10 μM mibefradil on the T-type Ca2+ current component (ICaT) measured in the depolarizing step at -40 mV with the general IK protocol; Table S3: TaqMan assays used for characterization of progenitor cells.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, F.I. and A.I.S; methodology, A.(P.)I; software, L.B.; validation, F.I., A.(P.)I; A.M.P., C.C., and L.S.; formal analysis, B.A.; investigation, F.I and A.I.S.; resources, F.I.; data curation, B.A., O.I.G and L.B; writing—original draft preparation, F.I. A.(P.)I; writing—review and editing, A.I.S.; visualization, F.I.; supervision, A.I..S; project administration, F.I.; funding acquisition, F.I. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Romanian UEFISCDI Agency, Project PN-III-P1-1.1-TE-2021-0864, No.19/2022, SENSTEM.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data supporting this study’s findings are available from the cor- responding author, AIS upon reasonable request.
Acknowledgments
This work was supported from the Romanian UEFISCDI Agency, Project PN-III-P1-1.1-TE-2021-0864, No.19/2022, SENSTEM.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Loukogeorgakis, S.P.; De Coppi, P. Concise Review: Amniotic Fluid Stem Cells: The Known, the Unknown, and Potential Regenerative Medicine Applications. Stem Cells. 2017, 35(7), 1663-1673. [CrossRef]
- Cananzi, M.; De Coppi, P. CD117(+) amniotic fluid stem cells: state of the art and future perspectives. Organogenesis. 2012, 8(3), 77-88. [CrossRef]
- Resca, E.; Zavatti, M.; Maraldi, T.; Bertoni, L.; Beretti, F.; Guida, M.; La Sala, G.B.; Guillot, P.V.; David, A.L.; Sebire, N.J.; De Pol, A.; De Coppi, P. Enrichment in c-Kit improved differentiation potential of amniotic membrane progenitor/stem cells. Placenta. 2015, 36(1), 18-26. [CrossRef]
- De Coppi, P.; Bartsch, G.Jr.; Siddiqui, M.M.; Xu, T.; Santos, C.C.; Perin, L.; Mostoslavsky, G.; Serre, A.C.; Snyder, E.Y.; Yoo, J.J.; Furth, M.E.; Soker, S.; Atala, A. Isolation of amniotic stem cell lines with potential for therapy. Nat Biotechnol. 2007, 25(1), 100-6. [CrossRef]
- Shamsnajafabadi, H.; Soheili, Z.S. Amniotic fluid characteristics and its application in stem cell therapy: A review. Int J Reprod Biomed. 2022, 20(8), 627-643. [CrossRef]
- Cananzi, M.; Atala, A.; De Coppi, P. Stem cells derived from amniotic fluid: new potentials in regenerative medicine. Reprod Biomed Online. 2009, 18, 17-27. [CrossRef]
- Di Baldassarre, A.; D’Amico, M.A.; Izzicupo, P.; Gaggi, G.; Guarnieri, S.; Mariggiò, M.A.; Antonucci, I.; Corneo, B.; Sirabella, D.; Stuppia, L.; Ghinassi, B. Cardiomyocytes derived from human cardiopoietic amniotic fluids. Sci Rep. 2018, 8(1), 12028. [CrossRef]
- Di Tizio, D.; Di Serafino, A.; Upadhyaya, P.; Sorino, L.; Stuppia, L.; Antonucci, I. The impact of epigenetic signatures on amniotic fluid stem cell fate. Stem Cells Int. 2018, 4274518. [CrossRef]
- Condorelli, G.; Latronico, M.V.; Cavarretta, E. microRNAs in cardiovascular diseases: current knowledge and the road ahead. J Am Coll Cardiol. 2014, 63(21), 2177-87. [CrossRef]
- Milcu, A.I.; Anghel, A.; Muşat, O.; Munteanu, M.; Salavat, M.C.; Iordache, A.; Ungureanu, E.; Bonţe, D.C.; Borugă, O. Implications at the ocular level of miRNAs modifications induced by SARS-CoV-2 infection. Rom J Morphol Embryol. 2022, 63(1), 55-59. [CrossRef]
- Trohatou, O.; Zagoura, D.; Bitsika V.; Pappa, K.I.; Antsaklis, A.; Anagnou, N.P.; Roubelakis, M.G. Sox2 suppression by miR-21 governs human mesenchymal stem cell properties. Stem Cells Translational Medicine, 2014, 3, 1, 54–68. [CrossRef]
- Lazzarini, R.; Olivieri, F.; Ferretti, C.; Mattioli-Belmonte, M.; di Primio, R.; Orciani, M. mRNAs and miRNAs profiling of mesenchymal stem cells derived from amniotic fluid and skin: the double face of the coin. Cell and Tissue Research, 2014, 355, 1, 121–130. [CrossRef]
- Gholizadeh-Ghaleh Aziz, S.; Pashaei-Asl, F.; Fardyazar, Z.; Pashaiasl, M. Isolation, characterization, cryopreservation of human amniotic stem cells and differentiation to osteogenic and adipogenic cells. PLoS One, 2016, vol. 11, no. 7, 2016. [CrossRef]
- Glemžaitė, M.; Navakauskienė, R. Osteogenic differentiation of human amniotic fluid mesenchymal stem cells is determined by epigenetic changes. Stem Cells International, 2016, 2016, 1-10. [CrossRef]
- Lazzarini, R.; Sorgentoni, G.; Caffarini, M.; Sayeed, M.A.; Olivieri, F.; Di Primio, R.; Orciani, M. New miRNAs network in human mesenchymal stem cells derived from skin and amniotic fluid. Int J Immunopathol Pharmacol. 2016, 29(3), 523-528. [CrossRef]
- Miranda-Sayago, J.M.; Fernandez-Arcas, N.; Reyes-Engel, A.; Benito, C..; Narbona, I.; Alonso, A. Changes in CDKN 2 D, TP 53, and mi R 125a expression: potential role in the evaluation of human amniotic fluid-derived mesenchymal stromal cell fitness. Genes to Cells, 2012, 17(8), 673-687. [CrossRef]
- Miyamoto, T.; Furusawa, C.; Kaneko, K. Pluripotency, differentiation, and reprogramming: A gene expression dynamics model with epigenetic feedback regulation. PLoS Comput Biol. 2015, 11(8), e1004476. [CrossRef]
- Bem, J.; Grabowska, I.; Daniszewski, M.; Zawada, D.; Czerwinska, A.M.; Bugajski, L.; Piwocka, K.; Fogtman, A.; Ciemerych, M.A. Transient microRNA expression enhances myogenic potential of mouse embryonic stem cells. Stem Cells. 2018, 36(5), 655-670. [CrossRef]
- Du, P.; Pirouz, M.; Choi, J.; Huebner, A.J.; Clement, K.; Meissner, A.; Hochedlinger, K.; Gregory, R.I. An intermediate pluripotent state controlled by micrornas is required for the naive-to-primed stem cell transition. Cell Stem Cell. 2018, 22(6), 851-864.e5. [CrossRef]
- Che, P.; Liu, J.; Shan, Z.; Wu, R.; Yao, C.; Cui, J.; Zhu, X.; Wang, J.; Burnett, M.S.; Wang, S.; Wang, J. miR-125a-5p impairs endothelial cell angiogenesis in aging mice via RTEF-1 downregulation. Aging Cell. 2014, 13(5), 926-934. [CrossRef]
- Lin, Z.; Ge, J.; Wang, Z.; Ren, J.; Wang, X.; Xiong, H.; Gao, J.; Zhang, Y.; Zhang, Q. Let-7e modulates the inflammatory response in vascular endothelial cells through ceRNA crosstalk. Sci Rep. 2017, 7, 42498. [CrossRef]
- Zhang, Q.; Xiao, X.; Zheng, J.; Li, M.; Yu, M.; Ping, F.; Wang, T.; Wang, X. Vildagliptin, a dipeptidyl peptidase-4 inhibitor, attenuated endothelial dysfunction through miRNAs in diabetic rats. Arch Med Sci. 2019, 17(5), 1378-1387. [CrossRef]
- Zhang, P.; Zhao, Q.; Gong, K.; Long, Y.; Zhang, J.; Li, Y.; Guo, X. Downregulation of miR-103a-3p contributes to endothelial progenitor cell dysfunction in deep vein thrombosis through PTEN targeting. Ann Vasc Surg. 2020, 64, 339-346. [CrossRef]
- Yamakuchi, M. Endothelial senescence and microRNA. Biomol Concepts. 2012, 3(3), 213-23. [CrossRef]
- Zeng, W.; Lei, Q.; Ma, J.; Ju, R. Effects of hypoxic-ischemic pre-treatment on microvesicles derived from endothelial progenitor cells. Exp Ther Med. 2020, 19(3), 2171-2178. [CrossRef]
- Wang, H.; Flach, H.; Onizawa, M.; Wei, L.; McManusm M.T.; Weiss, A. Negative regulation of Hif1a expression and TH17 differentiation by the hypoxia-regulated microRNA miR-210. Nat Immunol. 2014, 15(4), 393-401. [CrossRef]
- Li, X.; Jia, Z.; Zhao, X.; Xu, M.; Chen, M. Expression of miR-210 in the peripheral blood of patients with newly diagnosed type 2 diabetes mellitus and its effect on the number and function of endothelial progenitor cells. Microvasc Res. 2020, 131, 104032. [CrossRef]
- Shi, Z.H.; Han, X.Y.; Yao, M.D.; Liu, C.; Jiang, Q.; Yan, B. Differential MicroRNA expression pattern in endothelial Progenitor cells during diabetic retinopathy. Front Cell Dev Biol. 2021, 9, 773050. [CrossRef]
- Zhou, J.; Wang, K.C.; Wu, W., Subramaniam, S.; Shy, J.Y.; Chiu, J.J.; Li, J.Y.; Chien, S. MicroRNA-21 targets peroxisome proliferators-activated receptor-alpha in an autoregulatory loop to modulate flow-induced endothelial inflammation. Proc Natl Acad Sci U S A. 2011, 108:10355–10360. [CrossRef]
- Li, D.; Yang, P.; Xiong, Q.; Song, X.; Yang, X.; Liu, L.; Yuan, W.; Rui, Y.C. MicroRNA-125a/b-5p inhibits endothelin-1 expression in vascular endothelial cells. J Hypertens. 2010, 28(8), 1646-1654. [CrossRef]
- Chen, K.C.; Hsieh, I.C.; Hsi, E.; Wang, Y.S.; Dai, C.Y.; Chou, W.W; Juo, S.H. Negative feedback regulation between microRNA let-7g and the oxLDL receptor LOX-1. J Cell Sci. 2011, 124, 4115-4124. [CrossRef]
- Bao, M.H.; Zhang. Y.W.; Lou, X.Y.; Cheng, Y.; Zhou, H.H. Protective effects of let-7a and let-7b on oxidized low-density lipoprotein induced endothelial cell injuries. PLoS One. 2014, 9(9):e106540. [CrossRef]
- Pan, J.; Li, K.; Huang, W.; Zhang, X. MiR-137 inhibited cell proliferation and migration of vascular smooth muscle cells via targeting IGFBP-5 and modulating the mTOR/STAT3 signaling. PLoS One. 2017, 12(10), e0186245. [CrossRef]
- Kee, H.J.; Kim, G.R.; Cho, S.N.; Kwon, J.S.; Ahn, Y.; Kook, H.; Jeong, M.H. mir-18a-5p microrna increases vascular smooth muscle cell differentiation by downregulating syndecan4. Korean Circ J. 2014, 44(4), 255-563. [CrossRef]
- Tong, Y.; Zhou, M.H.; Li, S.P.; Zhao, H.M.; Zhang, Y.R.; Chen, D.; Wu, Y.X.; Pang, Q.F. MiR-155-5p attenuates vascular smooth muscle cell oxidative stress and migration via inhibiting BACH1 expression. Biomedicines 2023, 11(6), 1679. [CrossRef]
- Li, Y.; Li, H.; Xing, W.; Li, J.; Du, R.; Cao, D.; Wang, Y.; Yang, X.; Zhong, G.; Zhao, Y.; Sun, W.; Liu, C.; Gao, X.; Li, Y.; Liu, Z.; Jin, X.; Zhao, D.; Tan, Y.; Wang, Y.; Liu, S.; Yuan, M.; Song, J.; Chang, Y.Z.; Gao, F.; Ling, S.; Li, Y. Vascular smooth muscle cell-specific miRNA-214 knockout inhibits angiotensin II-induced hypertension through upregulation of Smad7. FASEB J. 2021, 35(11):e21947. [CrossRef]
- Iordache, F.; Constantinescu, A.; Andrei, E.; Amuzescu, B.; Halitzchi, F.; Savu, L.; Maniu, H. Electrophysiology, immunophenotype, and gene expression characterization of senescent and cryopreserved human amniotic fluid stem cells. J Physiol Sci. 2016, 66(6), 463-476. [CrossRef]
- Airini, R.; Iordache, F.; Alexandru, D.; Savu, L.; Epureanu, F.B.; Mihailescu, D.; Amuzescu, B.; Maniu, H. Senescence-induced immunophenotype, gene expression and electrophysiology changes in human amniocytes. J Cell Mol Med. 2019, 23(11), 7233-7245. [CrossRef]
- Mann, S.A; Heide, J.; Knott, T.; Airini, R,; Epureanu, F.B.; Deftu, A.F.; Deftu, A.T.; Radu, B.M.; Amuzescu, B. Recording of multiple ion current components and action potentials in human induced pluripotent stem cell-derived cardiomyocytes via automated patch-clamp. J Pharmacol Toxicol Methods. 2019, 100, 106599. [CrossRef]
- Scheel, O.; Frech, S.; Amuzescu, B.; Eisfeld, J.; Lin, K.H.; Knott, T. Action Potential Characterization of Human Induced Pluripotent Stem Cell–Derived Cardiomyocytes Using Automated Patch-Clamp Technology. ASSAY and Drug Development Technologies. 2014, 457-469. [CrossRef]
Figure 1.
Markers suggestive of endothelial progenitor commitment of AFSC: (A) Morphology of EPC after 4 weeks of culture with endothelial growth factors; (B) Tube-like structures of EPC on Matrigel extracellular matrix; (C) mRNA expression level for human CD31, eNOS, CD144, vwF (TaqMan qRT-PCR, n = 4, mean ± SEM, p < 0.05, one-way ANOVA); (D) Dil-Acetylatd-LDL uptake (red), UEA lectin binding (green) capacity of endothelial progenitor cells.
Figure 1.
Markers suggestive of endothelial progenitor commitment of AFSC: (A) Morphology of EPC after 4 weeks of culture with endothelial growth factors; (B) Tube-like structures of EPC on Matrigel extracellular matrix; (C) mRNA expression level for human CD31, eNOS, CD144, vwF (TaqMan qRT-PCR, n = 4, mean ± SEM, p < 0.05, one-way ANOVA); (D) Dil-Acetylatd-LDL uptake (red), UEA lectin binding (green) capacity of endothelial progenitor cells.
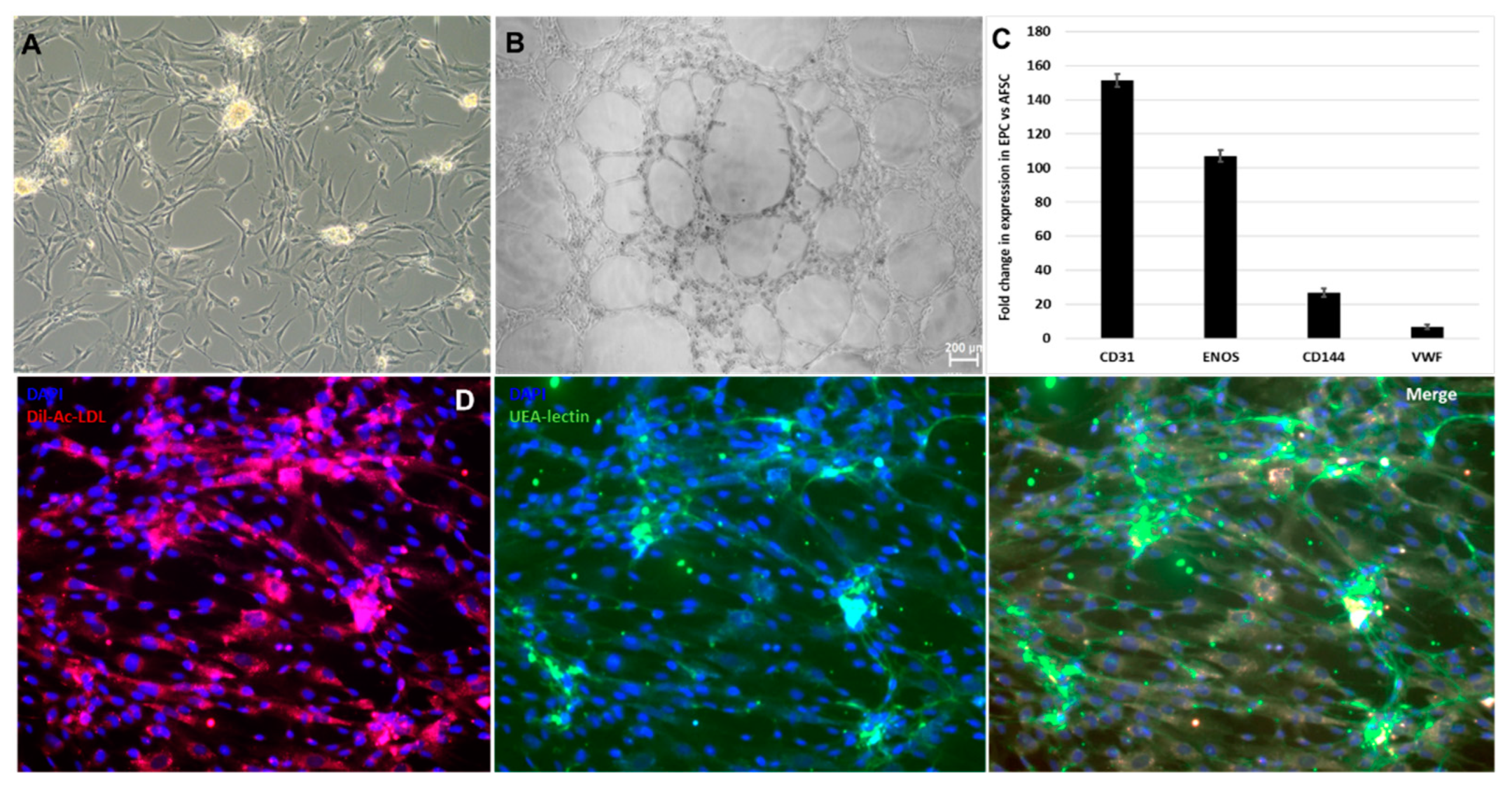
Figure 2.
Markers suggestive of smooth muscle commitment of AFSC. (A) mRNA expression level for human calponin-1, smoothelin, Mhy-11, TPM-1, SMA, caldesmon-1, Cav3.1 (TaqMan qRT-PCR, n = 4, mean ± SEM, p < 0.05, one way ANOVA); (B) Morphology of SPC after 4 weeks of culture with specific growth factors; (C) Double-ramp voltage-clamp protocol with traces recorded before and during application of iberiotoxin 100 nmol/L that blocked big conductance Ca2+-dependent K+ (BK) current fluctuations at positive potentials; (D) Inward current components suggestive of ICaT and ICaL, and their I-V curves (n = 24).
Figure 2.
Markers suggestive of smooth muscle commitment of AFSC. (A) mRNA expression level for human calponin-1, smoothelin, Mhy-11, TPM-1, SMA, caldesmon-1, Cav3.1 (TaqMan qRT-PCR, n = 4, mean ± SEM, p < 0.05, one way ANOVA); (B) Morphology of SPC after 4 weeks of culture with specific growth factors; (C) Double-ramp voltage-clamp protocol with traces recorded before and during application of iberiotoxin 100 nmol/L that blocked big conductance Ca2+-dependent K+ (BK) current fluctuations at positive potentials; (D) Inward current components suggestive of ICaT and ICaL, and their I-V curves (n = 24).
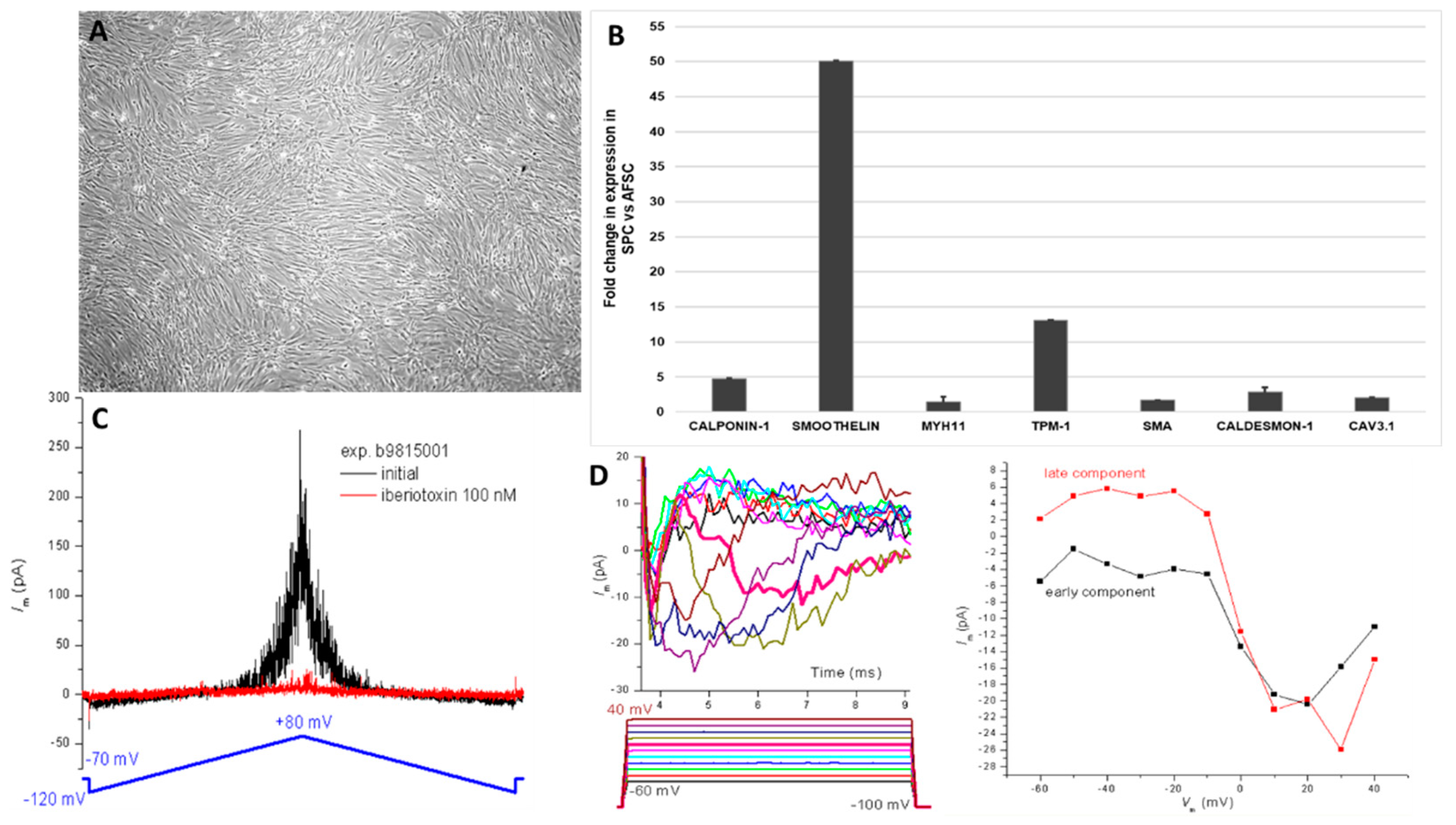
Figure 3.
miRNA expression levels in endothelial progenitor cells after 4 weeks of AFSC differentiation treatment. Fold-change is the normalized miRNA expression in each test sample divided the normalized miRNA expression in the control sample. Fold-change values higher than 2 are indicated the diagram, red-upregulated miRNA, green-downregulated miRNA, n=3, p< 0,005.
Figure 3.
miRNA expression levels in endothelial progenitor cells after 4 weeks of AFSC differentiation treatment. Fold-change is the normalized miRNA expression in each test sample divided the normalized miRNA expression in the control sample. Fold-change values higher than 2 are indicated the diagram, red-upregulated miRNA, green-downregulated miRNA, n=3, p< 0,005.
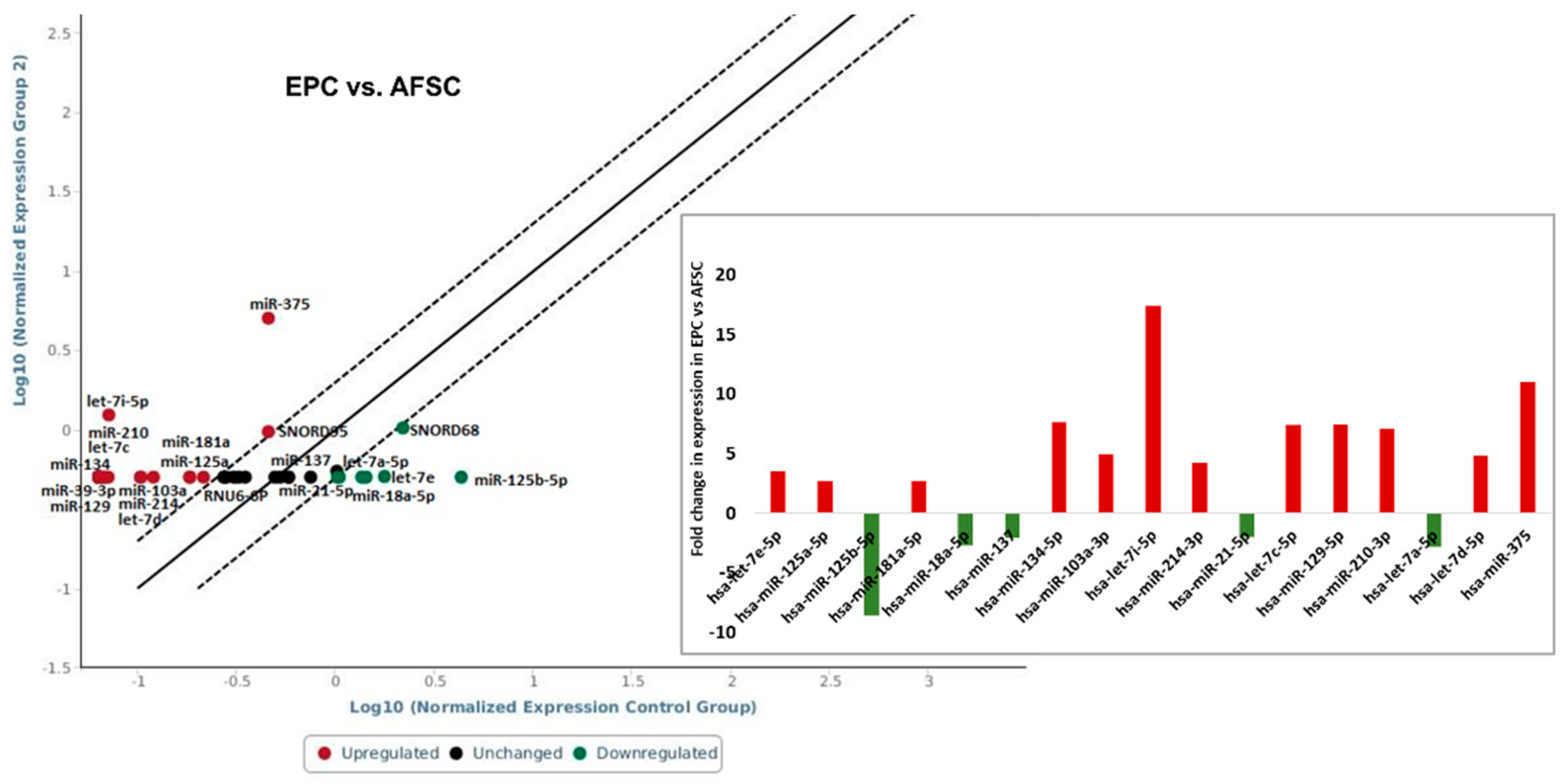
Figure 4.
miRNA expression levels in smooth muscle progenitor cells after 4 weeks of AFSC differentiation treatment. Fold-change is normalized miRNA expression in each test sample divided the normalized miRNA expression in the control sample. Fold-change values higher than 2 are indicated the diagram, red-upregulated miRNA, green-downregulated miRNA, n=3, p< 0,005.
Figure 4.
miRNA expression levels in smooth muscle progenitor cells after 4 weeks of AFSC differentiation treatment. Fold-change is normalized miRNA expression in each test sample divided the normalized miRNA expression in the control sample. Fold-change values higher than 2 are indicated the diagram, red-upregulated miRNA, green-downregulated miRNA, n=3, p< 0,005.
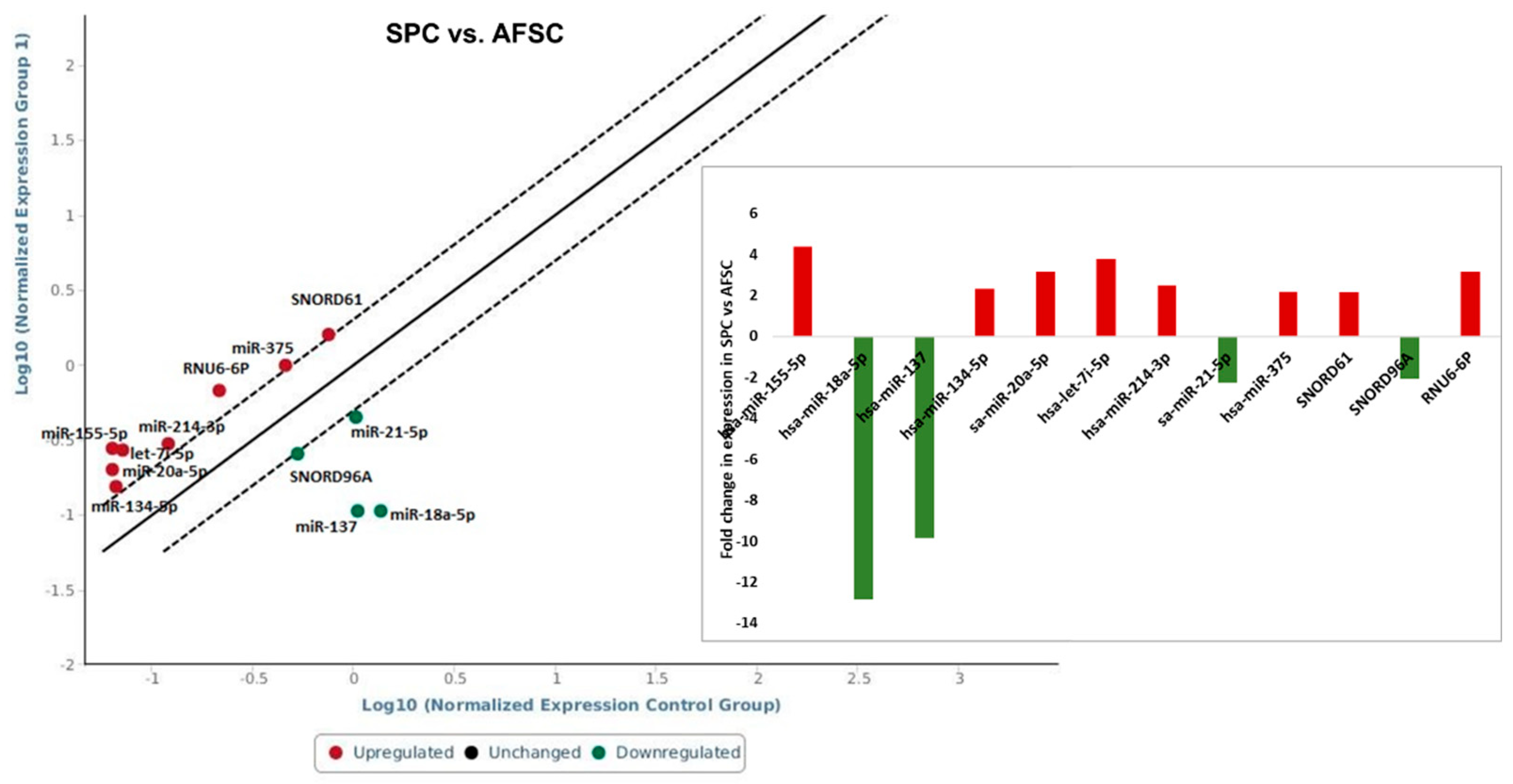
Figure 5.
The expression levels of miRNAs in endothelial and smooth muscle progenitor cells.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



