
Submitted:
16 November 2023
Posted:
17 November 2023
You are already at the latest version
Abstract
This study delves into the impact of contrasting management practices on epiphytic bryolichenic communities, shedding light on their divergent responses to management regimes on Castanea sativa orchards. Lichens and bryophytes were sampled in managed and abandoned plots, in 95 trees, in north and south sides and at two heights in Galicia (NW Spain). The studied groups exhibit opposing reactions to these management practices, bryophytes suffer adverse effects in managed stands, experiencing reduced cover and species richness compared to abandoned orchards, while lichens displayed heightened cover and species diversity. The size of trees, included as a covariate in our analyses, displayed no significant impact on the overall species richness of lichens or bryophytes, although it did influence the cover of specific functional traits. These differing outcomes are linked to alterations in environmental conditions brought by management interventions. Furthermore, the study uncovers divergent responses within the taxonomic and functional composition of epiphytic communities. Different species and functional groups exhibit varying reactions to changing environmental conditions, making predictions a complex endeavor. In conclusion, this research emphasizes the need for management strategies that account for the diverse ecological requirements of different species and functional groups. Since no single management regime will suit all species or functional groups.
Keywords:
lichens
; bryophytes
; biodiversity
; Castanea sativa
; managed
; unmanaged
; agroforestry system
1. Introduction
The management of trees as a source of food and timber is closely linked with the history of the human being, in fact, there is evidence that human societies all over the world managed forests long before they started to write [1].
Castanea sativa Mill., the sweet chestnut, covers more than 2,5 million hectares in Europe, of which 2,2 million hectares are chestnut forests, and the remaining 0,3 are mixed forests with chestnut [2]. Chestnut forests are considered habitat of Community Interest (9260) in the European Natura 2000 network (Annex I of the “Habitats” Directive 92/43/CEE). Chestnuts trees are distributed from the Caspian Sea to the Atlantic Ocean, and from southern Europe to the north (southern England, Belgium); chestnut forests are concentrated in countries close to the Mediterranean basin, occupying large areas in Italy, France, Spain, Portugal, Greece and Turkey [3,4]. In Spain, the chestnut is distributed over an area of 272,400 ha, of which 154,500 ha are pure [5]. In Galicia (NW Spain) chestnut trees cover more than 49,300 ha [6].
In the Iberian Peninsula, Castanea sativa is a native species, since the presence of pollen and other fossil remains has revealed the pre-Roman existence of Castanea in the northern Iberian Peninsula [7]. Its importance as a cultivated species increased from the Middle Ages onwards, as in other parts of Europe, probably encouraged by monastic orders [2,4,8]. Its distribution in the Old Continent has always been closely linked to human activities, as chestnut fruits have been an essential part of the diet of traditional communities for centuries, and its wood has also been highly valued [9,10].
Traditional sweet chestnut management involves the organisation of grafted trees in two types of monospecific systems with different stand densities: orchards or open stands where the main product is the fruit, and coppiced forests regenerated from adventitious or dormant buds, where the main product is the wood [9]. Both systems are interesting examples of socio-ecological production landscapes [10,11], as they are an important source of timber, firewood, and chestnut fruit, but also of other goods and services: such as honey, agroforestry pasture, litter for fertilisation or mulching [10,12], climate regulation through carbon storage [5], erosion control and water purification, and high biodiversity [13,14,15,16,17].
The most common morphology of chestnut trees, both in Galicia and in other European areas, is that of stands in which the understorey vegetation is removed and the trees are pollarded at a height of between 2 and 4 metres, and grafted at some point below the pollard. Periodic repetition of pruning, together with poor crown grafting leading to water accumulation, causes slow decay from the top centre of the pruning point. Over time, the rot slowly progresses to the base of the trunk, creating hollow trunks that continue to maintain vigorous and productive crowns. Over the years, this produces old trees with rough bark, cavities and cracks, scarred areas, and parts with dead wood [18].
Since the end of the 18th and the beginning of the 19th century, chestnut stands in NW Spain have suffered a significant decline as a result of changes in the agricultural system: mainly the introduction of new agricultural crops, the abandonment of traditional cultural practices and their replacement by pine or eucalyptus plantations. On the other hand, diseases caused by the fungi Phytophthora cambivora (Petri.) Buissman, P. cinnamoni Rands and Cryphonectria parasitica (Murril.) M.E. Barr. together with the plague of chestnut gall wasp Dryocosmus kuriphilus Yasumatsu have caused great damage to these agroforestry systems [19,20]. Another threat is that of forest fires, which may increase in frequency, size and intensity due to the increasing rates of biomass accumulation associated with land abandonment and to climate warming [21]. This was exemplified in the summer of 2022, when a large fire affected 13,000 hectares in our study area (‘O Courel’), including one of the plots studied in this research, in the village of Vilar.
Abandonment brings about important changes in the structure of chestnut stands. Both chestnut orchards and coppices are artificially maintained as monocultures, so that, in the absence of management, they tend to be colonised by other tree species, giving way to successional dynamics towards a more spontaneous forest, with higher tree diversity as well as canopy and understorey cover, as observed in many European regions in recent decades [9]. As forest management can significantly alter the moisture, temperature and light conditions whitin forests and these are important determinants of epiphytic communities [22,23], changes associated with abandonment can have important effects on these communities. For example, canopy thinning in forests favours the replacement of more hygrophilous species with more heliophilous species [24,25], whereas abandonment of this practice may have the opposite effect.
Changes in lichen communities associated with forest management have been shown in several studies across Europe in forests dominated by beech, pine, oak, aspen, spruce or hornbeam [26,27,28,29,30], as regards work on chestnut forests, partial studies have been carried out in Northern Italy [15,31], in the Swiss Alps [32] and in Montenegro [33]. In addition, there is a paucity of studies that collectively show the changes that occur in bryolichenic communities (both bryophytes and lichens) in relation to forest management [34]. Indeed, there are also very few studies on the bryophyte flora growing on chestnut in Europe [17].
In this study, we aim to contribute to closing this gap in the literature by investigating the effects of management vs. abandonment on epiphytic bryolichen communities in chestnut forests, focusing on the effects on lichen and bryophyte richness and abundance, as well as on the taxonomic and functional composition of these communities. It is expected that changes in forest microclimate due to management activities or abandonment will affect lichens and bryophytes differently, given their contrasting requirements, as well as species or functional groups that differ in their adaptations to environmental conditions.
2. Materials and Methods
2.1. Study area context
This research was carried out in the ‘O Courel’ mountain range (hereafter referred to as ‘O Courel’), north-west Spain, within the Eurosiberian biogeographical region. It is characterised by a mountainous landscape with a wide range of altitudes, from 400 m in the valley of the river Lor to 1,643 m on Mount Formigueiros. The climate of the region is Temperate, but this area is very close to the border between the Temperate and the Mediterranean climates. The landscape has a topography dominated by steep slopes (75% of which are steeper than 17º). Shale, quartzite and limestone are particularly dominant, with small outcrops of diabase [35]. It is included in the Ancares-Courel Special Area of Conservation (SAC), which covers 21,020 ha and is part of the Natura 2000 European network of protected areas.
Throughout ‘O Courel’, there are many chestnut orchards associated with villages that have been totally or partially abandoned, which has led to the abandonment of much of these orchards and their gradual transformation into mixed woodland. It is only in the few villages that are still inhabited that we can find at least partially managed orchards, since the number of people and working hours devoted to their upkeep have diminished considerably. Managed and unmanaged areas can easily be found in close proximity, within the same village, although current management appears to be limited to understorey clearance with little pruning.
2.2. Sampling design and procedure
We studied species abundance, richness, community composition in terms of both species and functional groups of epiphytic lichens and bryophytes on managed and unmanaged chestnut orchards in 8 ‘O Courel’ villages: Vilasibil (Vs), Aldea do Mazo (A), Romeor-Mostaz (R), Mercurín (M), Ferramulín (F), Santa Eufemia (St), Seoane (S) and Vilar (Vr). (Figure 1).
Figure 1.
Map of the study area showing chestnut orchards locations.
In each of these localities we selected managed and unmanaged (abandoned) orchard areas according to the intensity of land use, as revealed by the vegetation structure: (1) unmanaged areas were abandoned or semi-abandoned, the understorey vegetation was not cleared annually, and the understorey was dominated by ferns and some shrubs such as blackberry and heather; young trees of chestnut and other species (mainly Acer sp.) were also common, as a result of natural regeneration; (2) managed areas were moderately to intensively used, understorey vegetation was cleared once or twice a year, by burning or cutting, and chestnuts were harvested; the understorey development was limited and consisted of a thin and sparse herbaceous cover or bare soil. Only in one of these managed areas (Ferramulín site) was the planting of new specimens observed, with the typical crown grafting.
In each of the eight sites, we selected 5-7 trees per treatment (managed and unmanaged areas), for a total of 95 trees in 16 plots. Since epiphyte communities change with tree grow and as bark structure changes [36,37,38,39,40], tree size plays an important role in lichen and bryophyte colonisation [41,42]. we paired the trees in managed and unmanaged areas by size and used tree size as a covariate in the analyses, in order to avoid the effect of size as a confounding factor. The size of the selected trees ranged from 80 cm to 330 cm in perimeter, but within each pair the perimeter was similar (± 20cm). Tree size is usually correlated with tree age [28], and in Britain, where the climate is similar to our study area, these tree sizes correspond to an age range of 50 – 170 years [43]. Trees with a perimeter ≤ 100 cm were difficult to find in some of the plots, indicating their poor management. Trees were selected at least 15 m away from the edge of the orchards, avoiding leaning trunks and trees with injuries or with low branches.
In each selected tree we sampled lichens and bryophytes in 20 x 20 cm quadrats, which were divided into of 2 x 2 cm squares, in both the N and S sides of the trunks and at two heights, at the base of the trunk and at breast height. The base of the trunk is expected to be more affected by the different levels of understorey development between managed and unmanaged areas, which may modify the light and humidity conditions and, therefore, affect the epiphyte communities. All taxa present in the sampling quadrats were recorded and their abundance calculated as the area (in cm2) covered by each taxon. A total of 380 quadrats were sampled on 95 trees, 50 in managed areas and 45 in abandoned areas in 8 localities. Sampling took place from September 2015 to June 2018.
Identification of the lichen and bryophyte species was carried out by visual inspection of specimens using a hand lens with 20x magnification. Those taxa that could not be visually identified in the field were collected and taken to the laboratory, where they were identified using the classical methodology based on morphological and anatomical characteristics of lichens and bryophytes and chemical analyses of lichens, using identification keys and/or specific monographs [44,45,46,47,48,49]. The nomenclature of the taxa follows the Index Fungorum (www.indexfungorum.org) and/or [50] for lichens. For bryophytes, the nomenclature presented follows [50] for mosses, liverworts and hornworts.
2.3. Lichen functional groups
Lichen species were classified into functional groups according to seven traits: (1) Photobiont type: cyanolichens (with cyanobacteria), or chlorolichens (with green algae non-Trentepohlia or Trentepohlia), (2) Growth form: crustose, squamulose, foliose (broad or narrow), compound, gelatinous and fruticose (flat or round); (3) Size: macro- or microlichens; (4) Reproductive strategy: sexual, asexual by isidia or asexual by soredia; (5) Type of ascoma: apothecia, lirellae or perithecia; (6) Ascospores septation: simple, septate or muriform and (7) Chemistry: with or without chemical substances. This classification follows [50].
Lichens were also classified according to their ecological requirements, based on [50]: (1) light conditions (1 for lichens of very shaded sites, 2 for those of shaded sites, 3 for those of sites with plenty of diffuse light but scarce direct solar irradiation, 4 for those of sun-exposed sites, and 5 for those of sites with very high direct solar irradiation); (2) aridity (1 for hygrophytic lichens, in sites with a very high frequency of fog, 2 for rather hygrophytic lichens, intermediate between 1 and 2, 3 for mesophytic, 4 for xerophytic, and 5 for very xerophytic lichens); (3) pH of the substratum (1 for lichens of very acid substrata, 2 for those of acid substrata, 3 for those of subacid to subneutral substrata, 4 for those of slightly basic substrata, and 5 for those of basic substrata); (4) eutrophication (1 for lichens of sites with no eutrophication, 2 for those of very weak eutrophication, 3 for those of weak eutrophication, 4 for those of rather high eutrophication, and 5 for those of very high eutrophication; (5) tolerance to human disturbance or poleotolerance (0 lichens on old trees in undisturbed forests, 1 lichens of natural or semi-natural habitats, 2 species in moderately disturbed areas, 3 for those in heavily disturbed areas) (Table S3).
2.4. Bryophyte functional groups
Bryophyte species were classified into functional groups according to morphological and vegetative dispersal traits [52]: (1) life forms - annual, turf, cushions, mat, weft, dendroid; (2) life strategies – fugitives, colonists (LS-col.), annual shuttle, short-lived shuttle, perennial (long-lived) shuttle (LS-long-liv), perennial stayers (LS-peren st.); (3) presence or absence of multicellular buds on short stalks, formed in gemmae cups or leave axes (veg. gem.); (4) presence or absence of stoloniferous or specially modified branches; (5) presence or absence of caducous leaves from new plants (veg. leav.).
Bryophytes were classified according to their ecological requirements [52]: (1) light conditions (1 for bryophytes in deep shade, 3 for those in shade, 5 in semi-shade, 7 in well-lit places, but also occurring in partial shade, 9 in full light and Ind for indifferent bryophytes); (2) temperature (1 for bryophytes of cold conditions, found in high mountains, 3 cool conditions, 5 fairly warm conditions, 7 warm conditions, 9 extremely warm conditions and Ind for indifferent bryophytes); (3) moisture 1 for bryophytes of extremely dry sites, 3 for bryophytes found more often on dry sites than moist sites, 5 for those of moist sites, 7 moist sites, 9 wet sites and Ind for indifferent bryophytes); (4) soil acidity (1 for bryophytes on extremely acid soils, 3 for those on acid soils, 5 on moderately acid soils, 7 on weakly acid to weakly basic soils, 9 on basic soils and Ind for bryophytes indifferent to pH); (5) nutrient content (1 for bryophytes on sites with very low nutrient content, 3 on low nutrient content, 5 on medium nutrient content, 7 on nutrient-rich sites, 9, where no bryophytes occur); (6) human influence (1 absent, 3 weak, 5 moderate, 7 strong, 9 very strong); (7) forest type (1 for bryophytes largely restricted to closed forest, 1.2 for bryophytes that prefer forest edges, 2.1 for bryophytes that occur both in forest and open land, 2.2 for bryophytes that may occur in forest but prefer open land) (Table S4). For ecological requirements (1) to (6), intermediate states between those previously described received intermediate (even) numbers. For two bryophyte species (liverwort: Cephaloziella stellullifera; moss: Ulota crispula, there was no available information from the life-history trait database of forest bryophytes [52], and therefore, were not used in the subsequent statistical analyses regarding functional groups.
2.5. Data analysis
Species accumulation curves were calculated using EstimateS [53]. Species accumulation curves (or sample-based rarefaction curves, [54]) were calculated using Mao Tau randomisations [55] and represent the cumulative number of species as a function of the number of samples. In each randomisation, sampled species were randomly drawn from the dataset without replacement and added to the curve in a random order. We used 500 runs of randomised sampling order.
We analysed the effect of the management, orientation (N vs. S) and height in the tree trunk on the number of lichen and bryophyte taxa (richness, S) and in their abundance (surface cover density, as per sampling quadrat), using ANCOVA (Analysis of covariance) with locality as block and tree size (perimeter) as covariate. Variables were square-root transformed to meet normality and homoscedasticity requirements. However, data transformation was not successful for bryophyte richness. Therefore, in this case, we first performed an analysis of the effect of management, orientation and height using a GLM with a negative binomial function distribution and log-ratio link function and, since height had no significant effect (χ21 df = 0.3, P = 0.560), we pooled the data from the two sampled heights in the bole and performed the ANCOVA analysis as with the other variables, with previous transformation with the square root.
Species and functional community composition were analysed using PERMANOVA (Permutational Multivariate Analysis of Variance), with management as fixed factor, locality as random factor and tree perimeter as a covariate. To graph the differences in composition (in species and in functional traits) we used NMDS (Non-metric Multidimensional Scaling), with PRIMER 6.1.12 software, showing the distance between stands (managed and unmanaged in each locality) in the space defined by the lichen or bryophyte taxa or traits, respectively. Differences in the cover of specific functional trait categories between managed and unmanaged plots were analysed by ANCOVA, with management type as a fixed effect, site as a block and tree perimeter as a covariate. Variables were transformed with the arcsine, square root or logarithm when necessary to meet the requirements of normality and homoscedasticity. Some functional traits did not meet these requirements even after transformation. These were the least frequent traits, with 0 coverage in many of the trees. In these cases, we pooled the abundance of all the trees per site and analysed the effect of management with an ANOVA, with site as a block but no covariate.
3. Results
3.1. Abundance
Lichen cover density (i.e. the average cover per sampling quadrat) was significantly affected by management (Table 1, Figure 2), being higher in managed than in abandoned stands overall (21.1% higher in managed stands, effect adjusted for the covariate tree size), a trend that was fairly consistent across sites and position in the trunk (N vs. S and at the base vs. breast height). Orientation and height in the tree trunk had no significant effect on lichen cover density (Table 1). The effect of tree size was marginally significant (P = 0.062), but with a very weak, almost negligible, correlation strength (r = - 0.11), which was consistent for the two management treatments.
Management type also had a significant effect on bryophyte cover density, but with the opposite effect when compared to lichens: it was higher overall in unmanaged than in managed stands (17.7% increase in abandoned stands; effect adjusted for the covariate tree size). Orientation was also important in determining bryophyte cover density with a stronger effect than management; it was consistently higher on the north side of the trunks than on the south side of the tree trunks (57.0% increase on the north side, effect adjusted for the covariate tree size) (Table 1, Figure 2).
Figure 2.
Lichen and bryophyte cover density (per sampling quadrat) in managed and unmanaged stands in north and south orientations (mean ± SE).
Figure 2.
Lichen and bryophyte cover density (per sampling quadrat) in managed and unmanaged stands in north and south orientations (mean ± SE).
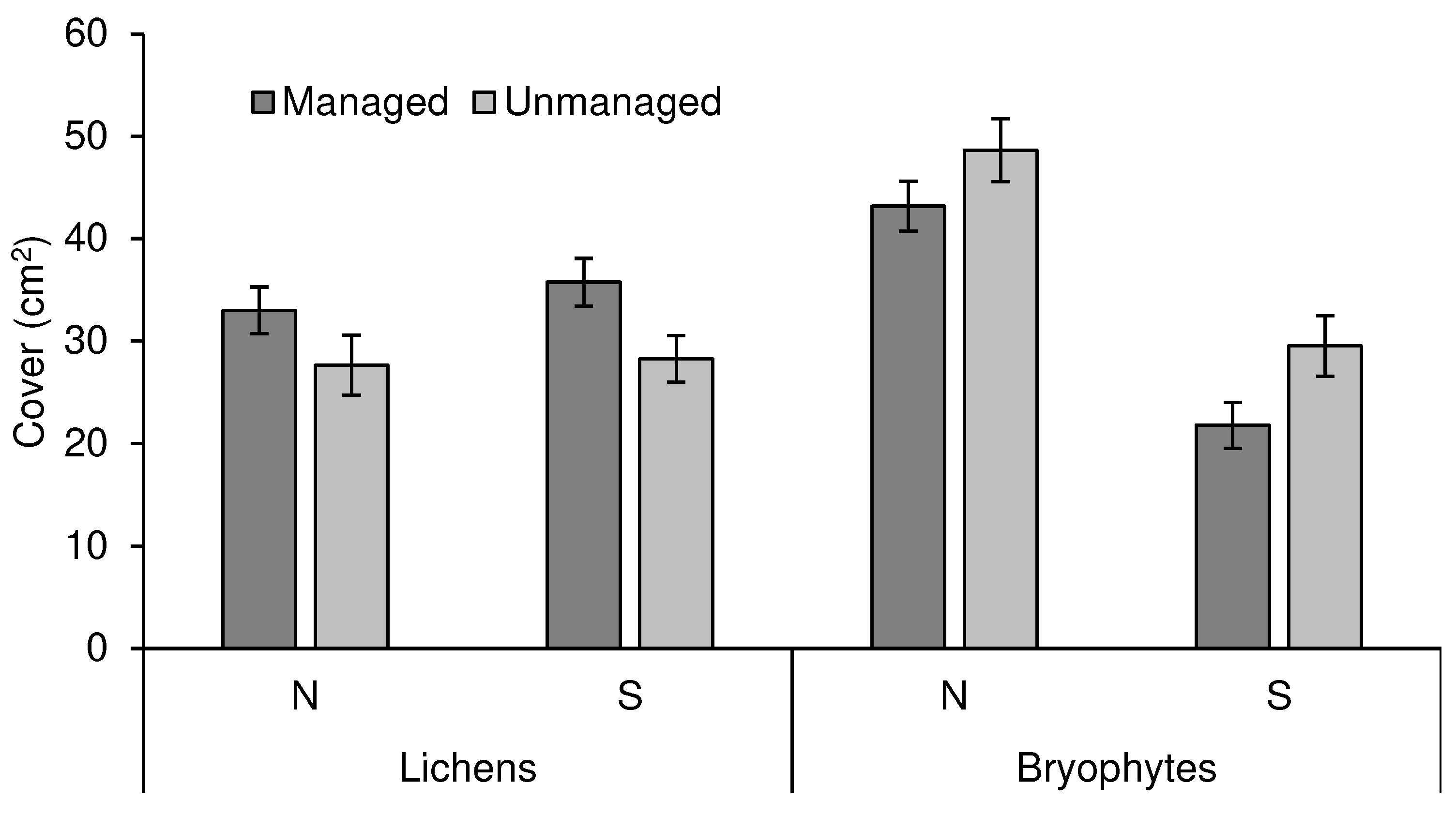
The total cover density, which includes lichens and bryophytes, was significantly influenced by management and orientation in the tree trunk. The average total cover was 66.9% of the surface, being slightly higher in managed (71.0%) than in unmanaged stands (62.5%), and 27.7% higher on the north side of the trunk than in the south side (76.2 vs. 57.7), a difference driven by bryophytes, as lichen cover was almost the same in both orientations (all means and effects were adjusted for the covariate, tree size) (Table 1).
3.2. Species richness
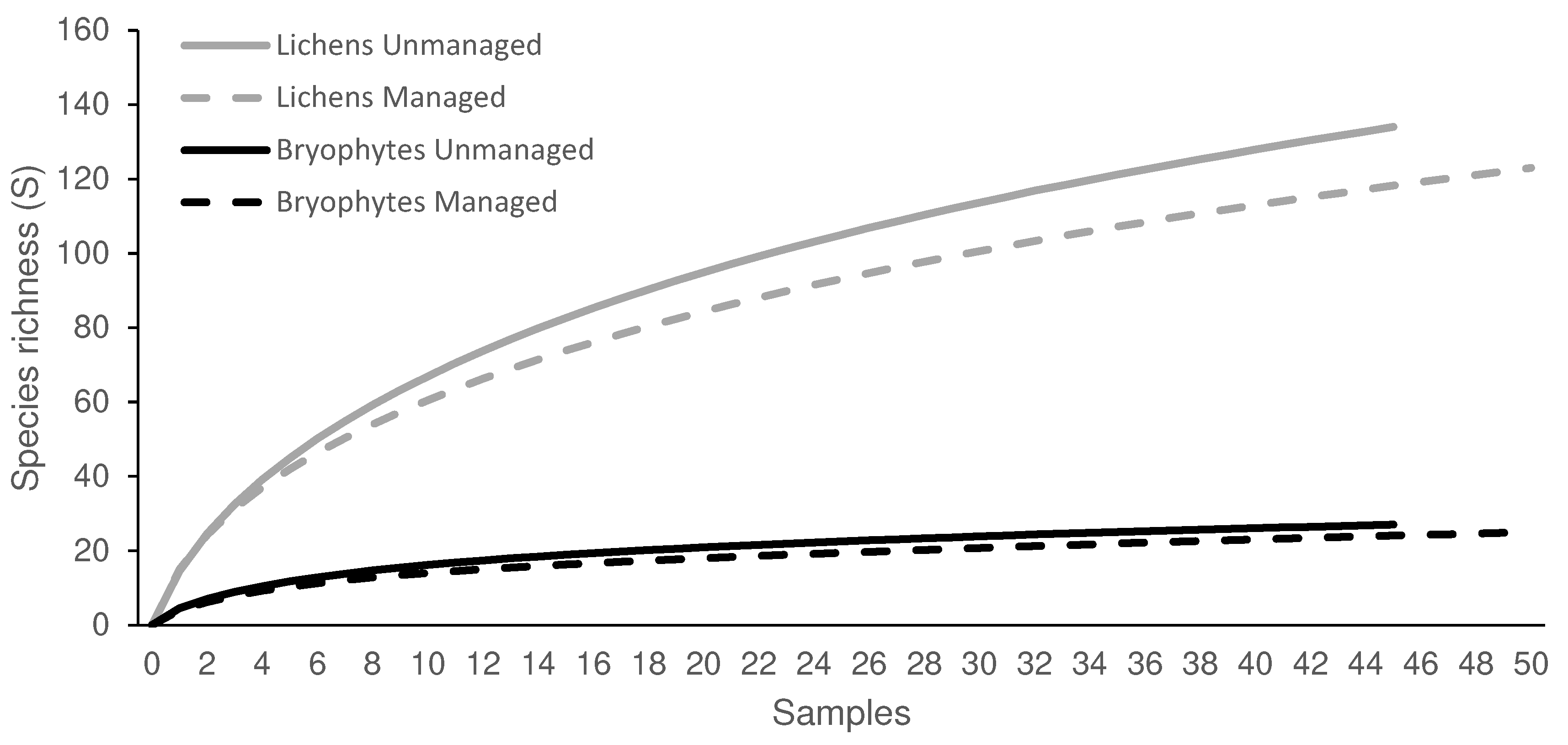
Species accumulation curves’ slopes tended to stabilise towards the end, especially for bryophytes, suggesting that, although the species richness would increase with increasing sample sizes, this increase would be limited (Figure 3). The total number of species recorded was much higher for lichens than for bryophytes, and in both cases higher in unmanaged than in managed stands, although very slightly in the case of bryophytes (2 species more). We recorded a total of 167 lichen taxa (146 identified at the species level and 21 at the genus level) in 70 genera and in all sampled plots (in both managed and unmanaged stands, in all sites, orientations and heights in the tree trunks; (Table S1). The total number of bryophyte taxa was 32, in 21 genera, in all the sampled plots (Table S2).
Figure 3.
Species accumulation curves for lichens and bryophytes in managed and unmanaged (abandoned) stands, representing the average number of species accumulated as the number of samples (trees) grows.
Figure 3.
Species accumulation curves for lichens and bryophytes in managed and unmanaged (abandoned) stands, representing the average number of species accumulated as the number of samples (trees) grows.
The density of taxa per lichens, i.e. the average species richness per sampling quadrat, was significantly influenced by the aspect and height, while the effect of management differed according to aspect (significant interaction between these factors), with management having a positive effect on diversity on the north side of the trunks, while the differences on the south side were not significant (Table 2, Figure 4).
For bryophytes, the density of taxa was significantly affected by the management, with higher richness in unmanaged stands, and by the orientation in the tree trunk (Table 2, Figure 4), with more species on the north side of the trunk than on the south side of trunks, a trend that was consistent across most trees. The effect of height was found to be non-significant factor in a previous test and was not included in this analysis (as explained in Methods).
3.3. Taxonomic composition
Management had a significant effect on the taxonomic composition of both lichens and bryophytes (Table 3), an effect that differed between sites (significant interaction between management and site). Tree size, as measured by the bole perimeter, also had a significant effect on taxonomic composition of both lichens and bryophytes, and this effect was consistent across the two management types (non-significant interaction between management and the covariate; Table 3).
The NMDS ordination of taxonomic composition (Figure 5) shows some aggregation of samples by management for both lichens and bryophytes, but despite its significant effect, there was no clear separation between managed and unmanaged stands, since the taxonomic composition was also affected by site and the effect of management differed between those sites, as shown in Table 3.
Some lichen species were associated with in either managed or unmanaged stands, and some of them were among the most abundant species and occurred predominantly in one type of management. Thus, considering only those species that occurred in at least 7 of the 16 sampled sites, 12 species had at least twice as much cover in unmanaged than managed plots, while 6 taxa had at least twice as much cover in managed than abandoned sites: 1) those predominantly associated with unmanaged plots were the cyanomorph Ricasolia amplissima with 13.4 times more cover in unmanaged than managed sites, followed by Placynthiella icmalea (13.3 times more), Normandina pulchella and Lecanora intumescens (8.1 times more), Fuscopannaria mediterranea (6.1), Lepraria lobificans (2.9), Nephroma parile (2.6), Cladonia pyxidata (2.4), Lecanora sp. (2.3), Pertusaria leioplaca (2.2), Parmeliella parvula (2.0), while 2) the taxa predominantly associated with managed plots were Usnea sp., with 8.7 times higher cover in managed sites, Hypogymnia physodes (3.6), H. tubulosa (3.1), Parmelia saxatilis (2.6), Evernia prunastri (2.3) and P. sulcata (2.1). There were also species exclusive to one type of management (25 and 37, in managed and unmanaged stands, respectively), but this mostly corresponded to a single occurrence on a single tree in a single plot in most cases (19 species in managed plots and 20 species in unmanaged plots), or to species occurring only in 2 or 3 plots.
A group of species were present in 3 plots or less, with the range of distribution varying from a single occurrence on a single tree in a single plot (19 species in managed plots, e.g. Lecanora cf. farinaria, Leucodermia leucomelos or Micarea viridileprosa and 20 species in unmanaged plots, e.g. Bacidia cf. absistens, Bryobilimbia cf sanguineoatra, Gyalidea ophiospora or Gyalideopsis muscicola) to 1 species on 4 trees in 2 orchards (Hypotrachyna minarum in managed plots) and 1 species on 6 trees in 3 orchards (Peltigera collina in unmanaged plots) (Table S1).
For bryophytes, there were also species associated with either managed or abandoned environments. Among the most common species (those occurring in at least half of the sampled sites), the following species were predominantly associated with unmanaged sites: Dicranum scoparium (5.6 times more abundant in unmanaged sites), Frullania dilatata (3.6), Cepahoziella stellullifera (3.0), Frullania tamarisci (2.5) and Orthotrichum lyellii (2.0). There were also a few-exclusive exclusive species (5 vs. 7 in managed vs. unmanaged sites), mostly single occurrences on a single tree in a single plot (Table S2).
3.4. Functional composition
Of the 167 lichen taxa, most were chlorolichens, six of them with Trentepohlia as photobiont and 23 species with cyanobacteria. Lichenized propagules (soredia and isidia) were the main reproductive strategy in more than half of the species (53%), about 45% of the taxa were hygrophytic (preferring more shaded habitats), more than 43% were lichens occurring in natural or semi-natural habitats and 10% were indicator species of old-growth forests [50].
Out of 28 bryophyte taxa with information about functional traits, 10 species are restricted to closed forest and 16 occurs in forests as well as in open land. Perennial stayers and perennial (long-lived) shuttle were the main reproductive life-strategies among bryophyte species (75%), about 78% taxa prefer rather shaded habitats and 75% taxa occur in weak human impact sites.
The functional composition followed a similar pattern to the taxonomic composition, both for lichens and bryophytes, with a significant effect of management, but also of the site and the tree size (covariate), with the effect of management being consistent across sites for bryophytes but varying between sites for lichens (significant interaction between management and site) (Table 4). The NMDS ordination shows these effects graphically. As for the taxonomic composition, the plots show a non-random distribution, but instead some aggregation (similarity) according to management, indicating to its effect. However, there was no clear separation between managed and unmanaged sites, both for lichens and bryophytes, due to the additional effects of the location (Figure 6).
The effect of management by functional trait, both in the sign of the effect (positive or negative effects of management/abandonment) and in its intensity (higher or lower changes in abundance depending on the type of management (Table 5). Among lichens, this effect was particularly strong and especially intense (effect ratio > 0.5 in absolute value, i.e. more than 50% change relative to the grand mean) for microlichens and lichens with septate ascospores, among those favoured by abandonment, and for foliose lichens among those favoured by management. Macrolichens, chlorolichens, lichens with apothecia and with substances were also favoured by management, but to a lesser extent (effect ratios ranged from - 0.23 to - 0.33, Table 9). Regarding ecological requirements, only for tolerance to human disturbance and eutrophication we had statistically significant effects. This effect was more pronounced for the former, where almost the whole gradient (except for one level) showed significant effects: abandonment had increasingly negative effects the more the lichen species were associated to human disturbances (from poleotolerance 1 to 3, i.e. from lichens associated with natural or semi-natural habitats to those from highly disturbed areas). With regard to eutrophication, we had significant effects at only two out of five levels, which makes the interpretation of the results more difficult: lichens associated with both no eutrophication (Eutro1) and low eutrophication (Eutro3) showed a significant positive effect of management, this effect being more intense in the latter. The effects on other ecological requirements were not significant.
Bryophytes with long-lived shuttle life strategies (LS long-liv) and with vegetative regeneration (veg. gem.) were positively affected by abandonment. Among ecological requirements, there were significant effects in some cases, but only in one or two levels of the whole gradient, making it difficult to understand these effects (Table 5).
There was a significant effect of the covariate for many functional traits for both lichens and bryophytes (e.g. for crustose and foliose lichens, macrolichens, chlorolichens, bryophytes with short and long-lived shuttles, and those with vegetative regeneration; Table 5), with different trends depending on the trait. The most pronounced trends were for lichens with chemical substances and chlorolichens in managed plots, which showed a decreasing cover density up to a perimeter of 200 cm, and then an increasing trend in larger trees (r2 = 0.219 and 0.191, respectively; polynomial regression grade 2), and for bryophytes with long-lived shuttles in abandoned plots, where cover density decreased with increasing tree size (r2 = 0.180; linear regression). The trend observed for lichens with chemical substances and chlorolichens in managed stands was also observed for foliose lichens, macrolichens, lichens with sexual reproduction and with apothecia, although to a lesser extent (r2 ranged between 0.11 and 0.14).
4. Discussion
Contrasting management regimes had effects on the epiphytic bryolichenic communities, with opposite effects on lichens and bryophytes: while management had negative effects for bryophytes (resulting in lower cover and richness overall), it had positive effects on lichens (higher cover and more species richness in the north sides of the trees). Management also affected the taxonomic and functional composition of the communities, with some species and functional groups appearing to be associated with either managed or unmanaged (abandoned) stands. Changes in environmental conditions, such as light, temperature and humidity, resulting from management action [22,56] may explain these effects, and the contrasting responses of lichens and bryophytes. By removing trees and understorey vegetation, management has important effects on vegetation structure, which ultimately affects forest microclimate, with changes in light penetration, temperature regimes and buffering capacity, transpiration and rain interception, etc., with closer, abandoned stands being cooler and wetter than managed stands [57,58,59]. Similarly, in the northern hemisphere, northern slopes receive less solar radiation and are cooler and more humid. Higher humidity favours bryophytes, by reducing the risk of desiccation, to which lichens are more tolerant [60], affecting them both directly, via their contrasting physiology, and indirectly, by altering their competitive interactions. This resulted in improved performance for bryophytes in the cooler and wetter abandoned orchards and for lichens in the sunnier and drier managed stands.
Studies conducted across Europe on the effect of management on lichen and bryophyte richness show contrasting results. For lichen richness, there are studies that show no effect of management ([61] comparing intensively managed and non-intensively managed environments), but most studies show a clear effect, either negative, with the more intensive the management leading to lower lichen diversity [62,63,64,65,66], or positive, in line with our study, e.g. [67] in beech forests in NE Germany. As for lichens, most studies on bryophytes have shown their sensitivity to management practices (e.g. [27] and references therein), with either positive or negative effects, which may not apply to all species [33,67].
Differences in the effect of management between forest systems are not surprising, as differences in climate, tree species and vegetation structure, as well as in management regimes (e.g. more or less intensive and using different methods) would determine the contrast between managed and unmanaged sites in environmental conditions. For example, a management regime that involves opening the canopy is not expected to have the same effect on a very dense canopy forest (e.g. beech forest) as on more open canopy systems (e.g. birch forest), nor on dry vs. humid climates. Moreover, some variables may act as confounders with effects on both the treatment and the response. [65], in their review of published research on the effects of management in different tree formations and on different organisms, conclude that it is important to take into account possible confounding factors, and highlight the role of stand and tree age when analysing the effects of management (see also [28]). For this reason, we used tree size as a covariate in our analyses, but it had no significant effect on the cover density or overall species richness of either lichens or bryophytes as a whole (it was marginally significant for lichen density, but with almost negligible correlation). However, it was a significant factor for many functional traits, which showed different trends with no general pattern across all types of lichens or bryophytes.
In addition to the general effects on lichen and bryophyte cover and species richness, there were also effects on the taxonomic and functional composition of epiphytic communities, with contrasting responses between species and functional groups. These contrasting responses are not surprising, as different species and functional groups are expected to respond differently to changes in environmental conditions (see for example, the different ecological requirements of the species recorded in this study, Tables S3 and S4).
Among the most representative lichen taxa of the managed environments were: Evernia prunastri, Hypogymnia physodes, H. tubulosa, Parmelia saxatilis, P. sulcata and Usnea sp. These species show traits that were clearly influenced by management: foliose growth form, macrolichens, chlorolichens, apothecia and chemistry, with a positive effect in all cases (Table 5).
This result is consistent with [56], who observed that fruticose lichens and broad-lobed lichens usually require higher light conditions than other growth forms and that excessive water availability reduces the presence of foliose lichens, explaining their negative response to abandonment. [30] studied the drivers of species composition of epiphytic bryophytes and chlorolichens in managed temperate forests and found that the main conditioning factor for bryophytes was moisture, whereas the main conditioning factor for chlorolichens was light. Again, this seems consistent with our results, as foliose green algal lichens were more abundant in the managed plots, as were macrolichens, most of which are chlorolichens. Green algal photobionts can photosynthesise at high light intensities [68], which may explain the improved performance of chlorolichens in the managed plots, with more open canopies. In contrast, closer, shadier and wetter conditions, typical of abandoned stands, have been associated with the presence of cyanolichens, many of which are also restricted to old-growth forests [23,69], where they are often associated with bryophytes [38]. In our study, although we found more bryophytes in abandoned stands, with higher cover and richness, the effect of abandonment on cyanolichens was not significant.
Asexual reproduction in lichens was similar to sexual in richness, but with some differences in cover, as species with sexual reproduction by apothecia had significantly higher cover in managed plots. [66] studied the patterns of lichen functional groups along 6 main biogeographical regions of Europe and show that the smaller size of spores is adapted to long-distance dispersal, which could explain the higher performance of these species in managed environments (more open) where spore dispersal is favoured. In addition, among sexually reproducing species, those with septate spores are favoured in unmanaged environments. This type of spore increases the chances of germination [70] by producing germinating hyphae from, at least, the cells at the tips of the spore [71]. This strategy appears to be successful in lichenisation, as the unmanaged plots are dominated by bryophytes
Among bryophytes, abandonment was particularly positive for those with perennial (long-lived) shuttle as life-strategies. These species thrive in more stable environments [52], such as abandoned plots compared to managed plots, due to reduced human disturbance. Consistently, abandoned plots had a negative effect on lichens adapted to human disturbance (poleotolerance), with increasing negative effects from lichens adapted to natural or semi-natural habitats to those from highly disturbed areas (Poleo 1 to 3 in Table 9). This was the clearest trend in terms of ecological requirements for lichens, whereas the effects found for eutrophication levels are more difficult to interpret, as they were only significant in only in two out of five levels, making it difficult to identify a trend in this regard. This was also the case for bryophytes, which are characterised by different ecological requirements, with no clear trends in any case.
We expected to see more clear differences in the effects of management on lichens and bryophytes with contrasting ecological requirements, at least in terms of light, temperature or human disturbance, which are clearly affected by management. However, in addition to the direct effects on a particular species or functional group, changing environmental conditions also affect its herbivores, competitors or mutualists, so the overall effect of the changing conditions on a particular species or group can be difficult to predict because direct and indirect effects often go in different directions [72]. Low statistical power was also an issue as well, especially for those traits or ecological requirements with more levels (as defined in [50,52]), as this reduces the number of species per group and the sample sizes. Future studies should also combine in-situ microclimate observations to better assess the impacts of forest management on microclimate heterogeneity, since recent studies concluded that microclimate-heterogeneity in European beech forests is largely determined by the type of forest management and forest structure [73].
In addition to these functional groups, there was also a small group of five species that was clearly associated with dead wood in our sampling, which are typically found in older trees as a result of pruning and grafting, regardless of whether the plots were managed or abandoned. Three of them (Calicium abietinum, C. parvum and Chaenotheca brunneola) are calicioid species known to be indicators of forest age and continuity [74], and the other two (Parmeliopsis ambigua and P. hyperopta) are narrow-lobed foliose species.
The contrasting effects of management on lichens and bryophytes, and on species and groups within them, highlight the importance of considering different taxonomic groups in biodiversity assessment in order to obtain complementary information [75]. Another study on the effects of management in chestnut orchards, carried out in our area of study area [13], focused on the biodiversity of vascular plants, birds, beetles and ants, and also found contrasting effects in different taxonomic groups: management had a significant effect on the richness of vascular plants, with a progressive decrease as management increased, but no clear effect on the richness of animals. Other multi-taxon studies carried out in different systems and geographical areas are along the same lines, showing different effects on different groups, such as the study by [76], comparing the effects of management on birds and lichens in boreal forests, or [77], in Central Europe, focusing on beetles, fungi and lichens. However, [62] showed similar negative effects of management on the richness of both animals (birds, mammals and lizards) and lichens in Mediterranean environments. Nevertheless, the paucity of studies encompassing the entire epiphytic community is striking and calls for more studies that include both lichens and bryophytes, as their contrasting ecological needs favour contrasting responses. Moreover, the varied responses to management in different forests, climates or management regimes highlight the need for further investigation in different systems and environments, including the matrix in which the forest systems are immersed, which may also vary in terms of management, and will ultimately determine the availability of propagules arriving on a given plot. The lack of general cross-taxa congruence in response to management means that management recommendations should balance the needs of different taxonomic groups in order to maintain forest biodiversity. Our results, and those of other studies, show that there is no single management type that suits all species or functional types. Therefore, diversified landscapes, with a variety of forest patches with different management regimes and degrees of abandonment, as in the study area, are probably the best option to provide the habitat heterogeneity to sustain high levels of biodiversity at the landscape scale [78].
5. Conclusions
The type of management of chestnut orchards had effects in the cover and richness of lichens and bryophytes, with opposite effects: abandonment favoured bryophyte cover and species richness density (per unit area) whereas management benefited lichen cover and richness density, albeit only in the north sides of the trunks for the latter.
It also affected the composition of species and of functional types, both for lichens and bryophytes. The most representative lichen species in managed plots belonged to functional groups significantly favoured by management, such as foliose growth form, macrolichens and chlorolichens. Abandonment, in contrast, was especially positive for bryophytes with perennial shuttle as life-strategies, typical of more stable environments, while having a negative effect on lichens adapted to human disturbance.
Diversified landscapes, with a variety of management regimes and degrees of abandonment, would probably represent the best option to favour epiphytic diversity in this case, given the contrasting responses to management/abandoned found between lichens and bryophytes, as well as among species and functional groups within them.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Lichen taxa collected; Table S2: Bryophyte taxa collected; Table S3: Lichen Traits and Ecological Requirements; Table S4: Bryophyte Traits and Ecological Requirements.
Author Contributions
Graciela Paz-Bermúdez: Conceptualization, Methodology, Investigation, Writing – original draft, Writing – review & editing, Supervision. Ana Belén Fernández-Salegui: Methodology, Investigation, Writing – review & editing. Helena Hespanhol: Methodology, Investigation, Writing – review & editing. M. Eugenia López de Silanes: Methodology, Investigation, Writing – review & editing. Cristina Vieira: Methodology, Investigation, Writing – review & editing. María Calviño-Cancela: Methodology, Investigation, Formal analysis, Writing – review & editing, Supervision. All authors have read and agreed to the published version of the manuscript.
Acknowledgments
We are very grateful to Pedro Álvarez, for producing the map included in this paper and for sharing so generously his wisdom and love for chestnut trees.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rietbergen, S. The History and Impact of Forest Management. In: The Forest Handbook. Applying Forest Science for Sustainable Management, Evans, J. Ed.; Blackwell Science, London, 2001; Volume 2, pp. 1-24. [CrossRef]
- Conedera, M.; Manetti, M.C.; Giuduci, F.; Amorini, E. Distribution and economic potential of the sweet chestnut (Castanea sativa Mill.) in Europe. Ecol. Mediterr. 2004; 30, 179–193. [Google Scholar] [CrossRef]
- Fernández-Cruz, J.; Fernández-López, J. Morphological, molecular and statistical tools to identify Castanea species and their hybrids. Conserv. Genet. 2012, 13, 1589–1600. [Google Scholar] [CrossRef]
- Krebs, P.; Pezzatti, G.B.; Beffa, G.; Tinner, W.; Conedera, M. Revising the Sweet Chestnut (Castanea sativa Mill.) Refugia History of the Last Glacial Period with Extended Pollen and Macrofossil Evidence. Quat. Sci. Rev. [CrossRef]
- Menéndez-Miguélez, M.; Álvarez-Álvarez, P.; Pardos, M.; Madrigal, G.; Ruiz-Peinado, R.; López-Senespleda, E.; Del Río, M.; Calama, R. Development of tools to estimate the contribution of young sweet chestnut plantations to climate-change mitigation. For. Ecol. Manage. 2023, 530, 120761. [Google Scholar] [CrossRef]
- DXPOF. Consellería do Medio Rural. Programa estratéxico do castiñeiro e da produción da castaña, Xunta de Galicia, Santiago de Compostela. 2022; 1–73.
- Carrión, J.S.; Yll, E.I.; Walker, M.J.; Legaz, A.J.; Chaín, C.; López, A. Glacial refugia of temperate, Mediterranean and Ibero North African flora in south-eastern Spain: new evidence from cave pollen at two Neanderthal man sites. Global Ecol. Biogeogr. 2003, 12, 119–129. [Google Scholar] [CrossRef]
- Ríos Rodríguez, M.L. Transformación agraria. Los terrenos de monte y la economía campesina (ss. XII-XIV) Semata. Ciencias Sociais e Humanidades, 1997; 9, 145–172. [Google Scholar]
- Conedera, M.; Stanga, P.; Oester, B.; Bachmann, P. Different post-culture dynamics in abandoned chestnut orchards and coppices. For. Snow Landsc. Res. 2001, 76, 487–492. [Google Scholar]
- Roces-Díaz, J.V.; Díaz-Varela, R.A.; Barrio-Anta, M.; Álvarez-Álvarez, P. Sweet chestnut agroforestry systems in North-western Spain: Classification, spatial distribution and an ecosystem services assessment. For. Syst. 2018, 27, e03S. [Google Scholar] [CrossRef]
- Roces-Díaz, J.V.; Díaz-Varela, R.A.; Álvarez-Álvarez, P.; Recondo, C.; Díaz-Varela, E.R. A multiscale analysis of ecosystem services supply in the NW Iberian Peninsula from a functional perspective. Ecol. Indic. 2015, 50, 24–34. [Google Scholar] [CrossRef]
- Aumeeruddy-Thomas, Y.; Therville, C.; Lemarchand, C.; Lauriac, A.; Richard, F. Resilience of Sweet Chestnut and Truffle Holm-Oak Rural Forests in Languedoc-Roussillon, France: Roles of Social-Ecological Legacies, Domestication, and Innovations. Ecol. Soc. 2012, 17. [Google Scholar] [CrossRef]
- Guitián, J.; Guitián, P.; Munilla, I.; Guitián, J.; Garrido, J.; Penín, L.; Domínguez, P.; Guitián, L. Biodiversity in chestnut woodlots: Management regimen vs. Woodlot size. Open J. For. 2012, 2, 200–206. [Google Scholar] [CrossRef]
- Salaverri, L.J.; Guitián, J.; Munilla, I.; Sobral, M. Bird richness decreases with the abandonment of agriculture in a rural region of SW Europe. Reg. Environ. Change 2019, 19, 245–250. [Google Scholar] [CrossRef]
- Matteucci, E.; Benesperi, R.; Giordani, P.; Piervittori, R.; Isocrono, D. Epiphytic lichen communities in chestnut stands in Central-North Italy. Biologia 2012, 67, 61–70. [Google Scholar] [CrossRef]
- Parisi, F.; Lombardi, F.; Marziliano, P.A.; Russo, D.; de Cristofaro, A.; Marchetti, M.; Tognetti, R. Diversity of saproxylic beetle communities in chestnut agroforestry systems. iForest 2020, 13, 456–465. [Google Scholar] [CrossRef]
- Privitera, M.; Puglisi, M. The ecology of bryophytes in the chestnut forests of mount Etna (Sicily, Italy). Ecol. Mediterr. 2000, 26, 43–52. [Google Scholar] [CrossRef]
- Fernández López, J. Guía de cultivo do castiñeiro para a produción de castaña. Xunta de Galicia, Santiago de Compostela, España, 2014; pp. 1–134. [CrossRef]
- Díaz-Varela, R.A.; Álvarez-Álvarez, P.; Díaz-Varela, E.; Calvo-Iglesias, S. Prediction of stand quality characteristics in sweet chestnut forests in NW Spain by combining terrain attributes, spectral textural features and landscapes metrics. For. Ecol. Manage. 2011, 261, 1961–1972. [Google Scholar] [CrossRef]
- Lombardero, M.J.; Ayres, M.P.; Álvarez-Álvarez, P.; Castedo-Dorado, F. Defensive patterns of chestnut genotypes (Castanea spp.) against the gall wasp, Dryocosmus kuriphilus. Front. For. Global Change 2022, 5, 1–17. [Google Scholar] [CrossRef]
- Aparício, B.A.; Santos, J.A.; Freitas, T.R.; Sá, A.C.L.; Pereira, J.M.C.; Fernandes, P.M. Unravelling the efect of climate change on fire danger and fire behaviour in the Transboundary Biosphere Reserve of Meseta Ibérica (Portugal Spain). Clim. Change 2022, 173. [Google Scholar] [CrossRef]
- Moen, J.; Jonsson, B.G. Edge effects on Liverworts and lichens in forests patches in a mosaic of Boreal Forest and Wetland. Conserv. Biol. 2003, 17, 380–388. [Google Scholar] [CrossRef]
- Belinchón, R.; Martínez, I.; Otálora, M.A.G.; Aragón, G.; Dimas, J.; Escudero, A. Fragment quality and matrix affect epiphytic performance in a Mediterranean forest landscape. Am. J. Bot. 2009, 96, 1974–1982. [Google Scholar] [CrossRef] [PubMed]
- Gradstein, S.R. Epiphytes of tropical montane forests – impact of deforestation and climate change. In The Tropical Mountain Forest. Patterns and Processes in a Biodiversity Hotspot; Gradstein, S.R., Homeier, J., Gansert, D., Eds.; University of Göttingen Press, Göttingen, Deutschland, 2008, pp. 51–65.
- Benítez, A.R.; Prieto, M.; Aragón, G. Large trees and dense canopies: Key factors for maintaining high epiphytic diversity on trunk bases (bryophytes and lichens) in tropical montane forests. Forestry 2015, 88, 521–527. [Google Scholar] [CrossRef]
- Cacciatori, C.; Czerepko, J.; Lech, P. Long-term changes in bryophyte diversity of central European managed forests depending on site environmental features. Biodiversity Conserv. 2022, 31, 2657–2681. [Google Scholar] [CrossRef]
- Müller, J.; Boch, S.; Prati, D.; Socher, S.A.; Pommer, U.; Hessenmöller, D.; Schall, P.; Schulze, E.D.; Fischer, M. Effects of forest management on bryophyte species richness in Central European forests. For. Ecol. Manage. 2019, 432, 850–859. [Google Scholar] [CrossRef]
- Nascimbene, J.; Thor, G.; Nimis, P.L. Effects of forest management on epiphytic lichens in temperate deciduous forests of Europe – A review. For. Ecol. Manage. 2013, 298, 27–38. [Google Scholar] [CrossRef]
- Noualhaguet, M.; Timothy, T.; Work, T.T.; Soubeyrand, M.; Nicole, J.; Fenton, N.F. Bryophyte community responses 20 years after forest management in boreal mixedwood forest. For. Ecol. Manage. 2023, 531, 120804. [Google Scholar] [CrossRef]
- Ódor, P.; Király, I.; Tinya, F.; Bortignon, F.; Nascimbene, J. Patterns and drivers of species composition of epiphytic bryophytes and lichens in managed temperate forests. For. Ecol. Manage. 2013, 306, 256–265. [Google Scholar] [CrossRef]
- Pezzi, G.; Gambini, S.; Buldrini, F.; Ferretti, F.; Muzzi, E.; Maresi, G.; Nascimbene, J. Contrasting patterns of tree features, lichen, and plant diversity in managed and abandoned old growth chestnut orchards of the northern Apennines (Italy). For. Ecol. Manage. 2020, 470, 118207. [Google Scholar] [CrossRef]
- Matteucci, E.; Isocrono, D.; Favero-Longo, S.E.; Moretti, M. Comunità licheniche epifite dei castagneti da fruto del Cantone Ticino, Svizzera. In Le selve castanili della Svizzera italiana Aspetti storici, paesaggistici, ecologici e gestionali, Moretti, M., Moretti, G., Conedera, M., Eds.; Societa ticinese di scienze natural, Lugano, Svizzera, 2021, pp. 109–121.
- Mayrhofer, H.; Drescher, A.; Stešević, D.; Bilovitz, P.O. Lichenized fungi of a chestnut grove in Livari (Rumija, Montenegro). Acta Bot. Croat. 2013, 72, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Czerepko, J.; Gawrýs, R.; Szymczyk, R.; Pisarek, W.; Janek, M.; Haidt, A.; Kowalewska, A.; Piegdoń, A.; Stebel, A.; Kukwa, M.; Cacciatori, C. How sensitive are epiphytic and epixylic cryptogams as indicators of forest naturalness? Testing bryophyte and lichen predictive power in stands under different management regimes in the Białowieża forest. Ecol. Indic. 2021, 125, 107532. [Google Scholar] [CrossRef]
- Pérez-Alberti, A. Geomorphology of O Courel. Grupo de Desenvolvemento Rural Ribeira Sacra-Courel. Lugo, España 2019; pp. 1-71. [CrossRef]
- Albertos, B.; Garilleti, R.; Lara, F.; Mazimpaka, V. Especificidad de los briófitos epífitos frente al forófito en un robledal mixto gallego. Bol. Soc. Esp. Briol. 2001, 18/19, 25–36. [Google Scholar]
- Calviño-Cancela, M.; Neumann, M.; López de Silanes, M.E. Contrasting patterns of lichen abundance and diversity in Eucalyptus globulus and Pinus pinaster plantations with tree age. For. Ecol. Manage. 2020, 462, 117994. [Google Scholar] [CrossRef]
- James, P.W.; Hawksworth, D.L.; Rose, F. Lichen communities in the British Isles: a preliminary conspectus. In Lichen Ecology, Seaward; M.R.D. Ed.; Academic Press, London, England, 1977; pp. 295–413.
- Nascimbene, J.; Marini, L.; Motta, R.; Nimis, P.L. Influence of tree age, tree size and crown structure on lichen communities in mature Alpine spruce forests. Biodiversity Conserv. 2009, 18, 1509–1522. [Google Scholar] [CrossRef]
- Slack, N.G. Host specificity of bryophytic epiphytes in eastern North America (Geography and Ecology of Bryophytes). J. Hat. Bot. Lab. 1976, 41, 107–132. [Google Scholar]
- Ranius, T.; Johansson, P.; Berg, N.; Niklasson, M. The influence of tree age and microhabitat quality on the occurrence of crustose lichens associated with old oaks. J. Veg. Sci. 2008, 19, 653–662. [Google Scholar] [CrossRef]
- Lara, F.; Mazimpaka, V. Succession of epiphytic bryophytes in a Quercus pyrenaica forest from Spain Central Range (Iberian Peninsula). Nova Hedwigia 1998, 67, 125–138. [Google Scholar] [CrossRef]
- Jarman, R.; Moir, A.K.; Webb, J.; Chambers, F.M. Sweet chestnut (Castanea sativa Mill.) in Britain: its dendrochronological potential. Arboric. J. 2017, 39, 100–124. [Google Scholar] [CrossRef]
- Burgaz, A.R.; Ahti, T. Cladoniaceae. Flora Liquenológica Ibérica, Sociedad Española de Liquenología, SEL, Madrid, España, 2009; 4, pp. 1-11.
- Carballal, R.; Paz-Bermúdez, G.; López de Silanes, M.E.; Valcárcel, C. Pannariaceae. Flora Liquenológica Ibérica, Sociedad Española de Liquenología, SEL, Pontevedra, España, 2010; 6, pp. 1–44.
- Casas, C.; Brugués, M.; Cros, R.M.; Sérgio, C. Handbook of mosses of the Iberian Peninsula and the Balearic Islands. Institut d'Estudis Catalans. Secció de Ciències Biològiques. Barcelona, España, 2009; pp. 1–177.
- Casas, C.; Brugués, M.; Cros, R.M.; Sérgio, C.; Marta, I. Handbook of Liverworts and Hornworts of the Iberian Peninsula and the Balearic Islands, 1st ed.; Institut d'Estudis Catalans; Secció de Ciències Biològiques: Barcelona, España, 2009; pp. 1–177. [Google Scholar]
- Czarnota, P. The lichen genus Micarea (Lecanorales, Ascomycota) in Poland. Pol. Bot. Stud. 2007, 23, 1–199. [Google Scholar]
- Smith, C.W.; Aptroot, A.; Coppins, B.J.; Fletcher, A.; Gilbert, O.L.; James, P.W.; Wolseley, P.A. The Lichens of Great Britain and Ireland, 1st ed.; British Lichen Society: London, UK, 2009; pp. 1–1046. [Google Scholar]
- ITALIC - The Information System on Italian Lichens. Available online: https://dryades.units.it/italic (accessed on day month 2022).
- Hodgetts, N.G.; Söderström, L.; Blockeel, T.L.; Caspari, S.; Ignatov, M.S.; Konstantinova, N.A.; Lockhart, N.; Papp, B.; Schröck, C.; Sim-Sim, M.; Bell, D.; Bell, N.E.; Blom, H.H.; Bruggeman-Nannenga, M.A.; Brugués, M.; Enroth, J.; Flatberg, K.I.; Garilleti, R.; Hedenäs, L.; Holyoak, D.T.; Hugonnot, V.; Kariyawasam, I.; Köckingeru, H.; Kučera, J.; Lara, F.; Porley, R.D. An annotated checklist of bryophytes of Europe, Macaronesia and Cyprus. J. Bryol. 2020, 42, 1–116. [Google Scholar] [CrossRef]
- Bernhardt-Römermann, M.; Poschlod, P.; Hentschel, J. BryForTrait – A life-history trait database of forest bryophytes. J. Veg. Sci. 2018, 29, 798–800. [Google Scholar] [CrossRef]
- Colwell, R.K. Estimates: Statistical Estimation of Species Richness and Shared Species from Samples. Version 7.5. User’s Guide and Application. 2005. Available online http://purl.oclc.org/estimates (accessed on 10 05 2023).
- Gotelli, N.J.; Colwell, R.K. Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecol. Lett. 2001, 4, 379–391. [Google Scholar] [CrossRef]
- Colwell, R.K.; Mao, C.X.; Chang, J. Interpolating, extrapolating, and comparing incidence-based species accumulation curves. Ecology 2004, 85, 2717–2727. [Google Scholar] [CrossRef]
- Giordani, P.; Incerti, G.; Rizzi, G.; Rellini, I.; Nimis, P.L.; Modenesi, P. Functional traits of cryptogams in Mediterranean ecosystems are driven by water, light and substrate interactions. J. Veg. Sci. 2013, 25, 778–792. [Google Scholar] [CrossRef]
- Chen, J.; Saunders, S.C.; Crow, T.R.; Naiman, R.J.; Brosofske, K.D.; Mroz, G.D.; Brookshire, B.L.; Franklin, J.F. Microcliminate in forest ecosystem and landscape ecology. Variations in local climate can be used to monitor and compare the effects of different management regimes. BioScience 1999, 47, 288–297. [Google Scholar] [CrossRef]
- Ogée, J.; Brunet, Y.; Loustau, D.; Berbigier, P.; Delzon, S. MuSICA, a CO2, water and energy multilayer, multileaf pine forest model: evaluation from hourly to yearly time scales and sensitivity analysis. Global Change Biol. 2003, 9, 697–717. [Google Scholar] [CrossRef]
- Luyssaert, S.; Jammet, M.; Stoy, P.C.; Estel, S.; Pongratz, J.; Ceschia, E.; Churkina, G.; Don, A.; Erb, K.; Ferlicoq, M.; Gielen, B.; Grünwald, T.; Houghton, R.A.; Klumpp, K.; Knohl, A.; Kolb, T.; Kuemmerle, T.; Laurila, T.; Lohila, T.; Loustau, D.; McGrath, M.J.; Meyfroidt, R.; Moors, E.J.; Naudts, K.; Novick, K.; Otto, J.; Pilegaard, K.; Pio, C.A.; Rambal, S.; Rebmann, C.; Ryder, J.; Suyker, A.E.; Varlagin, A.; Wattenbach Molman, A.J. Land management and land-cover change have impacts of similar magnitude on surface temperature. Nat. Clim. Change 2014, 4, 389–393. [Google Scholar] [CrossRef]
- Frahm, J.P. Climatic habitat differences of epiphytic lichens and bryophytes. Cryptogamie Bryol. 2003, 24, 3–14. [Google Scholar]
- Sitzia, T.; Campagnaro, T.; Dainese, M.; Cassol, M.; Dal Cortivo, M.; Gatti, E.; Padovan, F.; Sommacal, M.; Nascimbene, J. Contrasting multi-taxa diversity patterns between abandoned and non-intensively managed forests in the southern dolomites. iForest 2017, 10, 845–850. [Google Scholar] [CrossRef]
- Aragón, G.; Martínez, I.; Izquierdo, P.; Belinchón, R.; Escudero, A. Effects of forest management on epiphytic lichen diversity in Mediterranean forests. Appl. Veg. Sci. 2010, 13, 183–194. [Google Scholar] [CrossRef]
- Ardelean, I.V.; Keller, C.; Scheidegger, C. Effects of Management on Lichen Species Richness, Ecological Traits and Community Structure in the Rodnei Mountains National Park (Romania). Plos One 2015, 10, e0145808. [Google Scholar] [CrossRef] [PubMed]
- Bergamini, A.; Scheidegger, C.; Carvalho, P.; Davey, S.; Dietrich, M.; Dubs, F.; Farkas, E.; Groner, U.; Kärkkäinen, K.; Keller, C.; Lökös, L.; Lommi, S.; Máguas, C.; Mitchell, R.; Rico, V.J.; Aragón, G.; Truscott, A.M.; Wolseley, P.A.; Watt, A. Perfomance of macrolichens and lichen genera as indicators of lichen species richness and composition. Conservation Biology 2005, 19, 1051–1062. [Google Scholar] [CrossRef]
- Paillet, Y.; Bergès, L.; Hjältén, J.; Ódor, P.; Avon, C.; Bernhardt-Römermann, M.; Bijlsma, R.J.; De Bruyn, L. , Fuhr, M.; Grandin, U.; Kanka, R., Lundin, L.; Luque, S.; Magura, T.; Matesanz, S.; Mészáros, I.; Sebastià, M.T.; Schmidt, W.; Standovár, T.; Tóthmérész, B.; Uotila, A.; Valladares, F.; Vellak, K.; Virtanen, R. Biodiversity Differences between Managed and Unmanaged Forests: Meta-Analysis of Species Richness in Europe. Conserv. Biol 2009, 24, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Stofer, S.; Bergamini, A.; Aragón, G.; Carvalho, P.; Coppins, B.J.; Davey, S.; Dietrich, M.; Farkas, E.; Kärkkäinen, K.; Keller, C.; Lökös, L.; Lommi, S.; Máguas, C.; Mitchell, R.; Pinho, P. , Rico, V.J.; Truscott, A.M.; Wolseley, P.A.; Watt, A.; Scheidegger, C. Species richness of lichen functional groups in relation to land use intensity. Lichenologist 2006, 38, 331–353. [Google Scholar] [CrossRef]
- Friedel, A.; Oheimb, G.v.; Dengler, J.; Härdtle, W. Species diversity and species composition of epiphytic bryophytes and lichens – a comparison of managed and unmanaged beech forests in NE Germany. Feddes Repert. 2006, 117, 172–185. [Google Scholar] [CrossRef]
- Benítez, A.R.; Aragón, G.; González, Y.; Prieto, M. Functional traits of epiphytic lichens in response to forest disturbance and as predictors of total richness and diversity. Ecol. Indic. 2018, 86, 18–26. [Google Scholar] [CrossRef]
- Rikkinen, J. Cyanolichens. Biodiversity Conserv. 2015, 24, 973–993. [Google Scholar] [CrossRef]
- Pentecost, A. Some observations on the size and shape of lichen ascospores in relation to ecology and taxonomy. New Phytol. 1981, 89, 667–678. [Google Scholar] [CrossRef]
- Sanders, W.B.; Lücking, R. Reproductive strategies, relichelization and thallus development observed in situ in leaf-dwelling lichen communities. New Phytol. 2002, 155, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Greiser, C.; Ehrlén, J.; Luoto, M.; Meineri, E.; Merinero, S.; Willman, B.; Hylander, K. Warm range margin of boreal bryophytes and lichens not directly limited by temperatures. J. Ecol. 2021, 109, 3724–3736. [Google Scholar] [CrossRef]
- Menge, J.H.; Magdon, P.; Wöllauer, S.; Ehbrecht, M. Impacts of forest management on stand and landscape-level microclimate heterogeneity of European beech forests. Landscape Ecol. 2023, 38, 903–917. [Google Scholar] [CrossRef]
- Selva, S.B. Indicator Species — Restricted Taxa Approach in Coniferous and Hardwood Forests of Northeastern America, In Monitoring with Lichens, Monitoring Lichens, 1st ed.; Nimis, P.L., Scheidegger, C., Wolseley, P., Eds.; NATO Science Series; Springer: Dordrecht, Netherlands, 2002; Volume 7, pp. 349–352. [Google Scholar] [CrossRef]
- Vaz, A.S.; Hespanhol, H.; Vieira, C.; Alves, P.; Honrado, J.; Marques, J. Different responses but complementary views: patterns of cross-taxa diversity under contrasting coastal dynamics in secondary sand dunes. Plant Biosyst. 2020, 154, 553–559. [Google Scholar] [CrossRef]
- Klein, J.; Thor, G.; Low, M.; Sjögren, J.; Lindberg, E.; Eggers, S. What is good for birds is not always good for lichens: Interactions between forest structure and species richness in managed boreal forests. For. Ecol. Manage. 2020, 473, 118327. [Google Scholar] [CrossRef]
- Horák, J.; Pavlíček, J.; Kout, J.; Halda, J.P. Winners and losers in the wilderness: response of biodiversity to the abandonment of ancient forest pastures. Biodiversity Conserv. 2018, 27, 3019–3029. [Google Scholar] [CrossRef]
- Duflot, R.; Fahrig, L.; Mönkkönen, M. Management diversity begets biodiversity in production forest landscapes. Biol. Conserv. 2022, 268, 109514. [Google Scholar] [CrossRef]
Figure 4.
Lichen and bryophyte species richness density (per sampling quadrat) in managed and unmanaged stands in north and south orientations (mean ± SE).
Figure 4.
Lichen and bryophyte species richness density (per sampling quadrat) in managed and unmanaged stands in north and south orientations (mean ± SE).
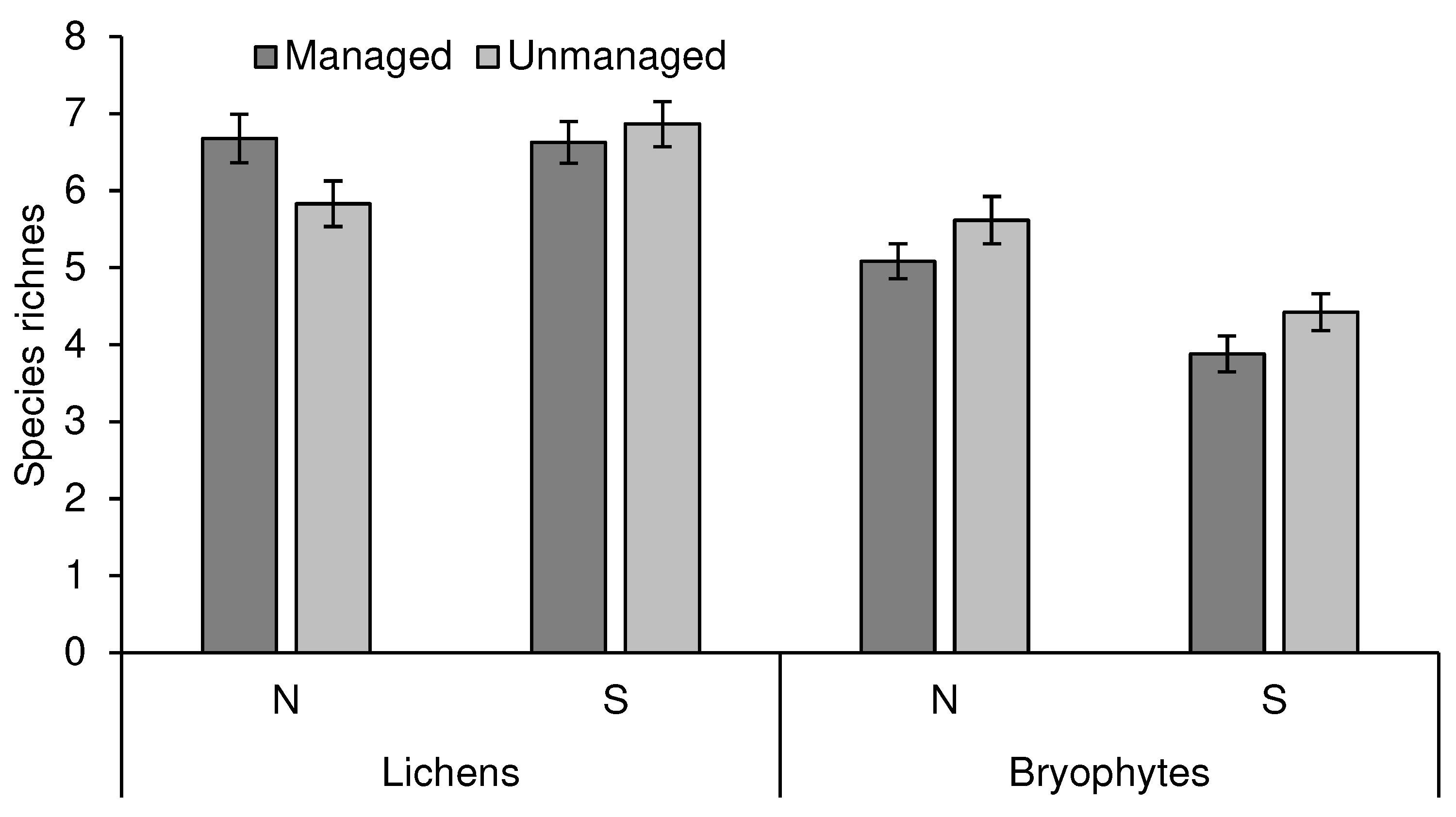
Figure 5.
Taxonomic composition. NMDS ordinations showing Bray-Curtis similarities between managed and abandoned stands of each site in the taxa space, for lichens (top) and bryophytes (bottom). Abbreviations correspond to plots.
Figure 5.
Taxonomic composition. NMDS ordinations showing Bray-Curtis similarities between managed and abandoned stands of each site in the taxa space, for lichens (top) and bryophytes (bottom). Abbreviations correspond to plots.
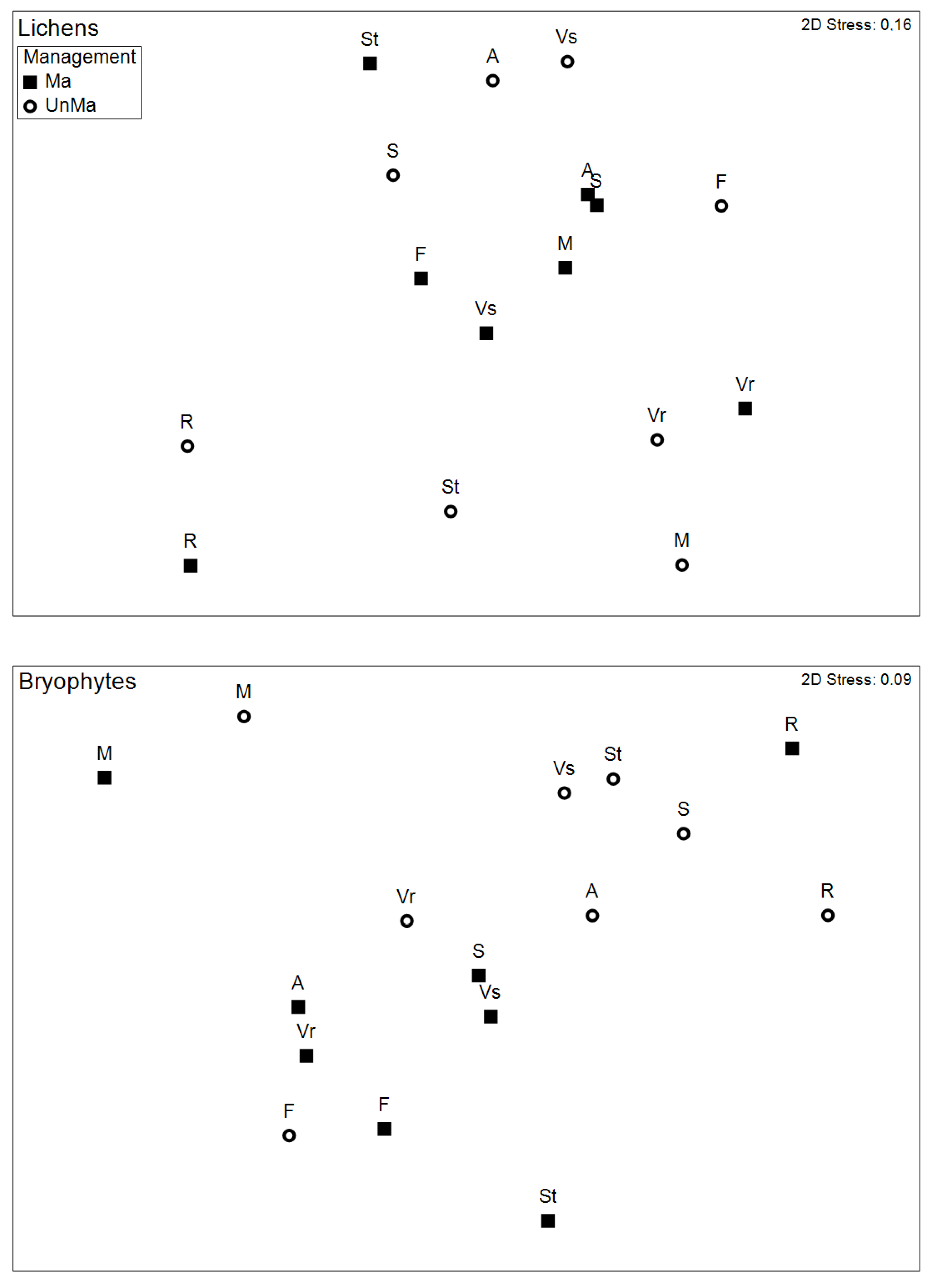
Figure 6.
Functional composition. NMDS ordinations showing Bray-Curtis similarities between managed and unmanaged stands of each site in the functional trait space, for lichens (top) and bryophytes (bottom), respectively. Abbreviations correspond to plots.
Figure 6.
Functional composition. NMDS ordinations showing Bray-Curtis similarities between managed and unmanaged stands of each site in the functional trait space, for lichens (top) and bryophytes (bottom), respectively. Abbreviations correspond to plots.
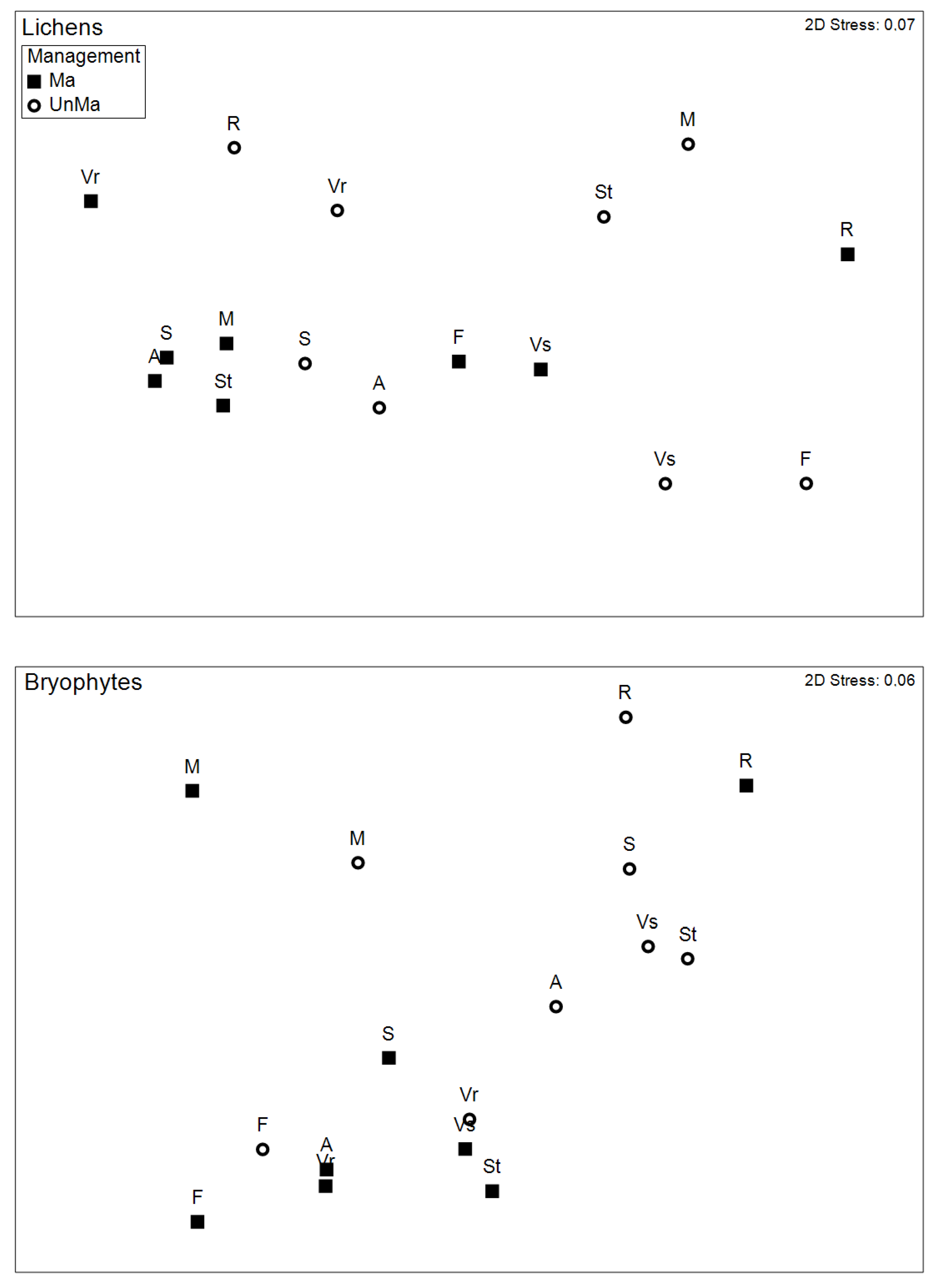

Table 1.
Results of the effect of management, orientation and height and their interactions on lichen, bryophyte and total (lichen plus bryophyte) cover density using ANCOVA with site as block and tree size (perimeter) as covariate. P-values are highlighted in bold. Covariate efficiency was 1 in all cases.
Table 1.
Results of the effect of management, orientation and height and their interactions on lichen, bryophyte and total (lichen plus bryophyte) cover density using ANCOVA with site as block and tree size (perimeter) as covariate. P-values are highlighted in bold. Covariate efficiency was 1 in all cases.
| Lichens | Bryophytes | Total | |||||
| Source of Variation | d.f. | F | P | F | P | F | P |
| Management | 1 | 9.7 | 0.002 | 5.6 | 0.019 | 6.0 | 0.015 |
| Orientation | 1 | 1 | 0.31 | 62 | <0.001 | 28.6 | <0.001 |
| Height | 1 | 0 | 0.84 | 2.1 | 0.145 | 0.1 | 0.801 |
| Manag.:Orient. | 1 | 0 | 0.87 | 0.3 | 0.59 | 0.6 | 0.447 |
| Manag.:Heigh. | 1 | 1.2 | 0.277 | 0 | 0.945 | 2.3 | 0.129 |
| Orient.:Heigh. | 1 | 0.2 | 0.686 | 1.6 | 0.21 | 0.02 | 0.878 |
| Manag.:Orient.:Heigh. | 1 | 0 | 0.926 | 0.1 | 0.717 | 0.3 | 0.559 |
| Covariate (Tree size) | 1 | 3.5 | 0.062 | 0.3 | 0.565 | 0.6 | 0.455 |
| Total | 372 | ||||||
Table 2.
Results of the effects of management, orientation and height, and their interactions, on lichen species richness, and of management and orientation, and their interactions, on bryophyte species richness (height was excluded, after a previous analysis showed no significant effect, in order to meet the requirements of normality and homoscedasticity, as explained in Methods) analysed using ANCOVA, with locality as block and tree size (perimeter) as covariate. P-values are shown in bold. The covariate efficiency was 1 in all cases.
Table 2.
Results of the effects of management, orientation and height, and their interactions, on lichen species richness, and of management and orientation, and their interactions, on bryophyte species richness (height was excluded, after a previous analysis showed no significant effect, in order to meet the requirements of normality and homoscedasticity, as explained in Methods) analysed using ANCOVA, with locality as block and tree size (perimeter) as covariate. P-values are shown in bold. The covariate efficiency was 1 in all cases.
| Lichens | Bryophytes | |||||
| Source of Variation | d.f. | F | P | d.f. | F | P |
| Management | 1 | 1.7 | 0.191 | 1 | 4.2 | 0.041 |
| Orientation | 1 | 4.4 | 0.036 | 1 | 23.0 | <0.001 |
| Height | 1 | 9.2 | 0.003 | . | . | . |
| Manag.: Orient. | 1 | 4.6 | 0.033 | 1 | 0.2 | 0.664 |
| Manag.: Heigh. | 1 | 0 | 0.938 | - | - | - |
| Orient.: Heigh. | 1 | 1.9 | 0.171 | - | - | - |
| Manag.:Orient.: Heigh. | 1 | 0.1 | 0.806 | - | - | - |
| Covariate | 1 | 0.9 | 0.343 | 1 | 0.5 | 0.48 |
| Residual | 364 | 1 | 181 | |||
Table 3.
Results of the effect of management, locality and the covariate tree size (perimeter), and their interactions, on the taxonomic composition of lichens and bryophytes, as analysed with PERMANOVA.
Table 3.
Results of the effect of management, locality and the covariate tree size (perimeter), and their interactions, on the taxonomic composition of lichens and bryophytes, as analysed with PERMANOVA.
| Lichens | Bryophytes | ||||
| Source of Variation | d.f. | Pseudo-F | P | Pseudo-F | P |
| Management | 1 | 1.96 | 0.015 | 4.16 | 0.005 |
| Locality | 7 | 4.02 | 0.001 | 4.86 | 0.001 |
| Manag.: Local. | 7 | 1.97 | 0.001 | 1.52 | 0.042 |
| Covariate (Tree size) | 1 | 7.43 | 0.001 | 7.41 | 0.001 |
| Covar.: Manag. | 1 | 1.05 | 0.405 | 1.63 | 0.138 |
| Covar.: Locality | 7 | 1.22 | 0.053 | 1.42 | 0.073 |
| Residual | 70 | ||||
Table 4.
Results of the effect of management site and their interaction, on the functional composition of lichens and bryophytes as analysed with PERMANOVA.
Table 4.
Results of the effect of management site and their interaction, on the functional composition of lichens and bryophytes as analysed with PERMANOVA.
| Lichens | Bryophytes | ||||
| Source of Variation | d.f. | Pseudo-F | P | Pseudo-F | P |
| Management | 1 | 4.75 | 0.004 | 6.12 | 0.001 |
| Locality | 7 | 3.62 | 0.001 | 4.36 | 0.001 |
| Manag.: Local. | 7 | 2.26 | 0.002 | 1.13 | 0.276 |
| Covariate (Tree size) | 1 | 9.53 | 0.001 | 5.20 | 0.003 |
| Covar.: Manag. | 1 | 0.97 | 0.380 | 2.24 | 0.055 |
| Covar.: Locality | 7 | 1.28 | 0.192 | 1.31 | 0.129 |
| Residual | 70 | ||||
Table 5.
Effect of management on lichen and bryophytes functional traits and ecological requirements using ANCOVA with sites as blocks and tree size (perimeter) as covariate. For traits marked with *, we used ANOVA, without the covariate, because they were very scarce and the large number of trees with 0 cover complicated the analyses; for this reason, we accumulated the cover for all trees in each site, and dropped the covariate (tree size) in these cases. The efficiency of the covariate was 1 in all cases. P-values < 0.05 are shown in bold. When estimating management effects (corrected for the covariate tree size), negative values reflect lower cover in unmanaged plots (negative effect of abandonment), whereas positive values reflect higher cover in unmanaged plots (positive effect of abandonment). The table shows the absolute difference between mean cover in managed and unmanaged plots, the SE of this difference, the grand mean (pooling both management types) and the effect ratio, as the difference of the means divided by grand mean.
Table 5.
Effect of management on lichen and bryophytes functional traits and ecological requirements using ANCOVA with sites as blocks and tree size (perimeter) as covariate. For traits marked with *, we used ANOVA, without the covariate, because they were very scarce and the large number of trees with 0 cover complicated the analyses; for this reason, we accumulated the cover for all trees in each site, and dropped the covariate (tree size) in these cases. The efficiency of the covariate was 1 in all cases. P-values < 0.05 are shown in bold. When estimating management effects (corrected for the covariate tree size), negative values reflect lower cover in unmanaged plots (negative effect of abandonment), whereas positive values reflect higher cover in unmanaged plots (positive effect of abandonment). The table shows the absolute difference between mean cover in managed and unmanaged plots, the SE of this difference, the grand mean (pooling both management types) and the effect ratio, as the difference of the means divided by grand mean.
| Functional traits | Fixed Effect: Management | Covariate: Tree size | Management effect size | ||||||
| (adjusted for tree size) | |||||||||
| F | P-value | F | P-value | Diffs. of means | SE | Grand mean | Effect ratio | ||
| Lichens | |||||||||
| Growth form | Cr | 2.09 | 0.152 | 6.1 | 0.016 | 2.5 | 6.38 | 40.7 | 0.06 |
| Fol | 7.93 | 0.006 | 5.7 | 0.019 | -28.4 | 9.62 | 59.1 | -0.48 | |
| *Squa | 2.21 | 0.181 | 2.42 | 0.974 | 1.5 | 1.61 | |||
| *Frut | 0.16 | 0.699 | -1.45 | 1.307 | 3.62 | -0.40 | |||
| *Compound | 0.05 | 0.831 | 3.1 | 3.17 | 7.7 | 0.40 | |||
| *G | 0.18 | 0.68 | 0.32 | 0.203 | 0.16 | 2.00 | |||
| Size | Macro | 8.36 | 0.005 | 5.4 | 0.023 | -31.9 | 11.13 | 95.4 | -0.33 |
| Micro | 4.38 | 0.039 | 0.6 | 0.447 | 10 | 4.79 | 17.2 | 0.58 | |
| PhotoB | Chlorolich. | 5.6 | 0.02 | 6.8 | 0.011 | -27.7 | 11.73 | 103.3 | -0.27 |
| Cyanolich. | 2.62 | 0.109 | 0.7 | 0.407 | 6 | 3.71 | 9.3 | 0.65 | |
| Trentepoh. | 1.47 | 0.229 | 0.3 | 0.585 | -0.091 | 0.075 | 0.099 | -0.92 | |
| Reproduct. | Asex_isi | 0.14 | 0.705 | 0.2 | 0.63 | -3.8 | 6.63 | 28.4 | -0.13 |
| Asex_sor | 1.6 | 0.209 | 5.9 | 0.018 | 0.055 | 0.044 | 0.488 | 0.11 | |
| Sex | 1.91 | 0.171 | 11 | 0.002 | -0.051 | 0.037 | 0.348 | -0.15 | |
| Ascoma | Apotecia | 5.14 | 0.026 | 10 | 0.002 | -24.9 | 10.96 | 99.3 | -0.25 |
| *Lirellae | 1.9 | 0.21 | -0.102 | 0.071 | 0.073 | -1.40 | |||
| Ascospores septation | SeptaSimp. | 3.65 | 0.06 | 3.3 | 0.074 | -32.5 | 10.91 | 81.7 | -0.40 |
| Septate | 9.44 | 0.003 | 3 | 0.087 | 8.8 | 4.21 | 13.8 | 0.64 | |
| SeptaMurif. | 0.75 | 0.389 | 0.2 | 0.688 | -0.033 | 0.038 | 0.128 | -0.26 | |
| Chemistry | Substances | 5.38 | 0.023 | 4.2 | 0.045 | -24.8 | 10.69 | 106.5 | -0.23 |
| No Subst. | 0.81 | 0.371 | 3.8 | 0.055 | 2.4 | 2.64 | 5.8 | 0.41 | |
| Ecological requirements | |||||||||
| pH | pH1 | 0.2 | 0.659 | 5.5 | 0.022 | -28.2 | 8.96 | 65.4 | -0.43 |
| pH2 | 0.76 | 0.387 | 2.7 | 0.105 | -25.9 | 10.84 | 110.1 | -0.24 | |
| pH3 | 0.1 | 0.75 | 11 | 0.001 | -17.5 | 10.71 | 75.8 | -0.23 | |
| *pH4 | 0.55 | 0.483 | 1.1 | 1.101 | 0.87 | 1.26 | |||
| *pH5 | 1.26 | 0.299 | 1.1 | 1.059 | 0.57 | 1.93 | |||
| Aridity | Arid1 | 2.2 | 0.142 | 1 | 0.314 | 5.2 | 4.59 | 12.1 | 0.43 |
| Arid2 | 0.23 | 0.632 | 6.7 | 0.012 | -17.3 | 10.37 | 92.5 | -0.19 | |
| Arid3 | 3.43 | 0.068 | 0.6 | 0.46 | -33.6 | 10.39 | 93.7 | -0.36 | |
| *Arid4 | 2.85 | 0.135 | 4.5 | 3 | 5 | 0.90 | |||
| *Arid5 | 0.34 | 0.579 | 0.097 | 0.144 | 0.101 | 0.96 | |||
| Light | *Light1 | 1.7 | 0.233 | -0.219 | 0.195 | 0.119 | -1.84 | ||
| Light2 | 0.01 | 0.915 | 13 | <0.001 | -6.2 | 4.14 | 20.9 | -0.30 | |
| Light3 | 1.69 | 0.197 | 7.3 | 0.008 | -24.9 | 10.79 | 108.8 | -0.23 | |
| Light4 | 1.11 | 0.295 | 6 | 0.016 | -26.2 | 11.01 | 85.4 | -0.31 | |
| Light5 | 0.45 | 0.506 | 13 | <0.001 | -14.6 | 7.36 | 28.2 | -0.52 | |
| Eutro | Eutro1 | 8.93 | 0.004 | 13 | <0.001 | -18.6 | 9.64 | 97.6 | -0.19 |
| Eutro2 | 0 | 0.949 | 0.1 | 0.824 | -30 | 11.48 | 96 | -0.31 | |
| Eutro3 | 14.5 | <0.001 | 21 | <0.001 | -35.4 | 10.07 | 67.9 | -0.52 | |
| *Eutro4 | 1.78 | 0.224 | 1.46 | 1.152 | 1.08 | 1.35 | |||
| *Eutro5 | 3.27 | 0.113 | 0.171 | 0.141 | 0.132 | 1.30 | |||
| Human disturbances | *Poleo0 | 1.11 | 0.327 | 1.02 | 1.023 | 2.22 | 0.46 | ||
| Poleo1 | 9.87 | 0.002 | 25 | <0.001 | -23 | 10.78 | 110.3 | -0.21 | |
| Poleo2 | 5.66 | 0.02 | 4.4 | 0.038 | -38.1 | 10.62 | 77.4 | -0.49 | |
| Poleo3 | 7.49 | 0.008 | 23 | <0.001 | -26.5 | 8.9 | 42.6 | -0.62 | |
| Fixed Effect: Management | Covariate: Tree size | Management effect size (adjusted for tree size) | |||||||
| F | P-value | F | P-value | cov. efficiency. | Diffs. of means | SE | Grand mean | Effect ratio | |
| Bryophytes | |||||||||
| Life form | Turf | 2.87 | 0.094 | 0.03 | 0.867 | 20.8 | 128.5 | 12.3 | 0.16 |
| *Cushion | 0.44 | 0.52 | 0.52 | 0.78 | 1.1 | 0.46 | |||
| *Mat | 0.68 | 0.425 | 0.37 | 0.44 | 0.7 | 0.56 | |||
| *Weft | 0.4 | 0.535 | 2.9 | 4.52 | 6.3 | 0.46 | |||
| Life strategies | LS-long-liv | 12.5 | <.001 | 6.13 | 0.015 | 22.6 | 6.18 | 24.3 | 0.93 |
| *LS-col | 0.19 | 0.667 | 0.35 | 0.80 | 1.2 | 0.29 | |||
| LS-peren st. | 1.64 | 0.204 | 3.97 | 0.05 | -15.5 | 12.13 | 95.0 | -0.16 | |
| LS-short-liv | 3.04 | 0.085 | 5.99 | 0.016 | 16.6 | 7.63 | 19.5 | 0.85 | |
| Forest Openness | *ForClosed | 0.42 | 0.525 | -2.1 | 4.76 | 4.9 | -0.43 | ||
| *ForEdges | 0.84 | 0.376 | 2.4 | 3.39 | 3.3 | 0.73 | |||
| *ForOpen | 1.86 | 0.194 | 24 | 17.64 | 134.1 | 0.18 | |||
| Open | 1.06 | 0.305 | 1.32 | 0.254 | 0.0022 | 0.0021 | 0.0011 | 2.00 | |
| Veg.. | Veg._gem. | 5.86 | 0.018 | 11.6 | <.001 | 32.5 | 10.50 | 34.5 | 0.94 |
| *Veg._leav. | 0.25 | 0.623 | 1 | 2.00 | 3.1 | 0.32 | |||
| Ecological requirements | |||||||||
| Light | *L-4 | 0.18 | 0.677 | -1.7 | 4.69 | 4.6 | -0.37 | ||
| *L-5 | 2.67 | 0.124 | 2.45 | 1.50 | 2.7 | 0.91 | |||
| L-7 | 4.22 | 0.043 | 0.28 | 0.596 | 18.3 | 7.68 | 25.5 | 0.72 | |
| *L-8 | 5.26 | 0.038 | 23.4 | 10.17 | 20.4 | 1.15 | |||
| L-Indif | 1.63 | 0.205 | 0.76 | 0.387 | -16 | 12.50 | 88.1 | -0.18 | |
| Temperat. | T-3 | 12.98 | <.001 | 3.33 | 0.072 | 24.4 | 6.99 | 25.3 | 0.96 |
| *T-4 | 0.48 | 0.501 | -2.1 | 4.53 | 5.0 | -0.42 | |||
| T-6 | 1.25 | 0.267 | 1.81 | 0.182 | 14 | 7.50 | 18.5 | 0.76 | |
| T-Indif | 0.25 | 0.641 | 0.43 | 0.542 | -14.6 | 12.44 | 89.8 | -0.16 | |
| Moisture | *F-3 | 0.84 | 0.374 | 0.72 | 0.78 | 1.0 | 0.71 | ||
| F-4 | 16.81 | <.001 | 0.7 | 0.407 | 38.3 | 10.26 | 44.0 | 0.87 | |
| F-5 | 1.83 | 0.18 | 0.17 | 0.679 | 2.41 | 1.58 | 3.4 | 0.72 | |
| F-6 | 1.63 | 0.205 | 0.76 | 0.387 | -16 | 12.50 | 88.1 | -0.18 | |
| *F-Indif | 0.8 | 0.388 | -16.2 | 18.14 | 87.1 | -0.19 | |||
| Soil pH | *R-3 | 0.29 | 0.601 | -0.28 | 0.53 | 0.9 | -0.33 | ||
| *R-4 | 0.01 | 0.935 | -0.4 | 4.49 | 6.4 | -0.06 | |||
| R-5 | 14.81 | <.001 | 4.51 | 0.037 | 37.8 | 10.27 | 39.5 | 0.96 | |
| *R-6 | 1.18 | 0.296 | 3.7 | 4.52 | 6.3 | 0.59 | |||
| R-Indif | 1.63 | 0.205 | 0.76 | 0.387 | -16 | 12.50 | 88.1 | -0.18 | |
| Nutrients | *N-2 | 1.4 | 0.257 | 5.6 | 4.76 | 10.1 | 0.55 | ||
| N-3 | 0.47 | 0.493 | 4.33 | 0.041 | -0.1 | 5.35 | 8.5 | -0.01 | |
| N-4 | 2.6 | 0.11 | 0.69 | 0.409 | 20 | 12.40 | 123.3 | 0.16 | |
| Human disturbanc. | *H-2 | 0.72 | 0.412 | -0.44 | 1.52 | 2.31 | -0.19 | ||
| H-3 | 11.05 | 0.001 | 0.65 | 0.423 | 18.3 | 7.89 | 22.7 | 0.81 | |
| *H-4 | 3.62 | 0.078 | 8.7 | 4.61 | 9.4 | 0.93 | |||
| H-5 | 1.58 | 0.212 | 1.12 | 0.293 | -15.4 | 12.31 | 89.5 | -0.17 | |
| H-6 | 1.25 | 0.267 | 1.81 | 0.182 | 14 | 7.50 | 18.5 | 0.76 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



