Submitted:
16 October 2023
Posted:
17 October 2023
Read the latest preprint version here
Abstract
Haemosporidians are a group of vector-borne parasites belonging to the order Haemosporida. They infect avian hosts and require blood-sucking insects (Diptera) for their transmission. The occur-rence and diversity of haemosporidian parasites are shaped primarily by the specificity of the parasite and the susceptibility of the host/vector. In this study, the presence and distribution of haemosporidians in blood samples from birds in urbanized and natural habitats were estimated using microscopic and molecular approaches. Birds in urbanized habitats were infected with four different species of Plasmodium: Plasmodium (Haemamoeba) relictum, P. (Novyella) vaughani, P. (H.) matutinum, P. (G.) circumflexum, two different species of Haemoproteus: Haemoproteus (Parahaemo-proteus) majoris, H. (P.) parabelopolskyi, and Leucocytozoon sp. The species H. (P.) balmorali, H. (P.) concavocentralis, H. (P.) minutus, and H. (P.) pallidus were additionally identified in birds in natural habitats. Typically, juvenile birds are essential markers of parasite species transmitted in the study area. The juveniles in the urbanized habitats carried the P. (H.) relictum, P. (N.) vaughani, P. (G.) circumflexum, H. (P.) parabelopolskyi, H. (P.) majoris, and Leucocytozoon species. The most abundant parasite was H. (P.) parabelopolskyi, which was found in both types of habitats. The prevalence of the Haemoproteus and Plasmodium species in birds in natural habitats (in totally 43.80%) was sig-nificantly higher than that in birds in urbanized habitats (in totally 21.94%) (p < 0.05), also in spring. There was no statistically significant difference between types of habitats for total infection rates with Leucocytozoon sp. (p > 0.05), neither overall nor in the spring.
Keywords:
Plasmodium
; Haemoproteus
; Leucocytozoon
; Avian diseases
; Slovakia
; adults
; juveniles
1. Introduction
Avian haemosporidians belonging to order Haemosporida (Alveolata, Apicomplexa, Sporozoa) is classified in four families Haemoproteidae (Haemoproteus), Plasmodiidae (Plasmodium), Leucocytozoidae ((Leucocytozoon) and Garniidae ). It is are a group of vector-borne parasites that infect avian hosts and require blood-sucking insects (Diptera) to use as vectors and complete their life cycle.. Haemoproteus species are transmitted by biting midges (Ceratopogonidae) and hippoboscid flies (Hippoboscidae), Plasmodium species by mosquitoes (Culicidae) and Leucocytozoon species by simuliid flies (Simuliidae). Their life cycle is complicated as they change hosts, and ways of reproduction, and morphologically form different developmental stages [1,2]. Parasites can persist in birds for many years or even a lifetime, and they serve as a source of infection for vectors, which can also lead to the infection of offspring during the breeding season/reproduction period.
A majority of related past studies are based on microscopy of blood smears where a set of morphological characteristics has served as the basis for species determination [1,2]. After molecular methods began to be employed in parasitology, the number of studies increased rapidly. A gold standard used in the protocol for detection of haemosporidians is based on the cytochrome b gene, which was established by Hellgren et al. [3]. The routine application of PCR, followed by sequencing, for detecting and identifying haemosporidian lineages has led to a massive expansion in parasite genetic diversity. Bensch et al. [4] created the MalAvi database [5], which summarize 6729 parasite lineages and knowledge about their host ranges and geographical distributions facilitating understanding of the evolutionary and ecological factors that drive this complex multi-host-multiparasite system easier.
The information about the presence of haemosporidian parasites in Slovakia is currently limited . In the past, most studies focused on the presence of haemosporidians in Slovakia were based on the microscopy analysis of blood smears [6,7,8,9,10]. PCR and sequencing were used for the first time in Slovakia by Berthová et al. [11] and expanded by Šujanová et al. [12].
Here, we investigated the presence of haemosporidians in wild birds, both adults and juveniles, in Slovakia using microscopic and molecular approaches. In order to enrich the knowledge about the occurrence and distribution of haemosporidians in Slovakia, it is necessary to identify the species whose transmission takes place here; we compared the occurrence of haemosporidians in urbanized and natural habitats with local bird populations and migrating bird populations, respectively.
2. Materials and Methods
2.1. Study sites
Birds were mist-netted in urbanized and natural habitat types in Slovakia from April to August 2012 and from April to October 2013 in the morning. The birds were captured, ringed, blood sampled, and released under the permission of the Ministry of Environment of the Slovak Republic No. 9368/2011-2.2. The locations are described in detail in Kazimírová et al. [13] and Berthová et al. [14]. Bratislava and Prievidza represent urbanized habitats in city areas, with anthropogenic activities such as walking, cycling, hiking, and horsing. Bratislava is located in the Small Carpathian Mountains in the southwest of Slovakia at 202 – 334 m above sea level (asl). The Prievidza district (289 m asl), located in central Slovakia, is a forest-steppe rural area. Both study sites are characteristic of local bird populations. Drienovec represents natural habitat far from a city area and is without significant human activities. It is a forest-steppe natural area (181 m asl) in the Slovak Karst National Park in -southeast Slovakia. An ornithological observatory for studying migrating birds is situated there.
2.2. Mist-netting and sampling birds
After ringing the birds, the sex and age of each individual were determined using the field guide by Svensson [15] and Hromádko et al. [16,17,18]. A small amount of blood (approximately 50 µl) was taken by puncture of the vena ulnaris cutanea using a 1 ml X-tra-fine needle (HMD Healthcare, UK), and the blood smears were prepared, while the rest of blood was stored in 96% ethanol. All birds were released immediately after sampling. Juveniles were considered to be all first year individuals captured in the calendar year of hatching. In total, 346 birds representing 42 species from 18 families were caught, and 114 of them were juveniles. Blood was collected from 317 individual birds.
2.3. Microscopic analysis of haemosporidian
A drop of blood was used to prepare the blood smears, and two smears were prepared for each individual of 302 birds. All smears were air-dried, fixed in methanol (96%), and stained with Giemsa stain as described in Valkiūnas [1]. The smears were examined by a Leica DM4500B microscope (Germany), and pictures were taken with a Leica DFC480 camera (Germany). Approximately 100 fields were scanned under ×400 magnification, and then a minimum of 100 fields were examined under high magnification (×1000) (oil immersion). The intensity of infection was determined according to the count of infected blood cells per 10,000 erythrocytes as follows: low (1-10 parasites), medium (11-100 parasites) and high (more than 100 parasites) according to Valkiūnas [1]. Pictures for the identification of parasites were edited using the program Leica Image Manager (Germany). Determination of the haemosporidians was performed based on key data according to Valkiūnas [1,2]. All the smears were deposited at the Institute of Virology, Biomedical Research Center, SAS, Bratislava.
2.4. Molecular analysis of haemosporidian
DNA from the 317 blood samples was extracted using a NucleoSpin® Tissue commercial kit (Macherey–Nagel, Germany). Approximately 50 µl of the blood stored in ethanol was transferred to a sterile 1.5 ml Eppendorf tube, and the ethanol was allowed to evaporate. When the blood was completely dry, 200 µl of lysis buffer B3 and 25 µl of proteinase K were added. The mixture was gently vortexed and incubated at 70 °C for 1 hour. The following procedure was performed according to the manufacturer’s instructions. The concentration and the quality of the DNA were assessed using a NanoPhotometer Pearl (Implen, Germany). The DNA samples were stored at -20 °C until the analyses. The presence of blood parasites was examined by a nested PCR assay targeting the cytochrome b (cyt b) gene mtDNA according to Bensch et al. [19] and Hellgren et al. [3]. Briefly, in the first step, HaemNF1 and HaemNR3 primers were used to amplify the 617 bp fragment. In the second step, the nested PCR with primers HaemF and HaemR2 amplified 480 bp fragments of Haemoproteus and Plasmodium , and the set of HaemFL and HaemR2L primers amplified a 478 bp fragment of Leucocytozoon. Sterile distilled water was used as a negative control, and the samples previously detected as PCR-positive and confirmed by sequencing [11] were used as positive controls. For the separation and visualization of the PCR products, 1 % agarose gel stained with DNA Stain G (Serva, Germany) was used. The PCR products of the expected size were considered positive and were sent for the purification and sequencing of both DNA strands to Macrogen (The Netherlands; http://www.macrogen.com). All the cyt b sequences were edited in MEGA6 software [20] and identified by performing a nucleotide blast in GenBank on NCBI [21]. The obtained sequences were compared with known ones in the MalAvi database [4,5]. Sequences of poor quality were marked as unusable sequences (us), and sequences with double peaks, which denote co-infections, were marked as dp.
Samples which were microscopically positive for haemosporians were examined for the intensity of haemosporidium infection using quantitative real-time PCR (qPCR) that targeted 182 bp fragment of the cyt b gene [12,22]. All reactions were carried out using GoTaq qPCR Master Mix (Promega, USA) on a CFX96 real-time thermocycler (Bio-Rad, Hercules, CA). The following cycling conditions were used: 95 °C for 2 min, followed by 40 cycles of 95 °C for 30 s and 64 °C for 35 s with a plate read, followed by a final melt curve analysis using the instrument’s default settings [12]. A synthetic double-stranded DNA product (Eurofins Genomics, France) designed from a 220 bp fragment of the conserved rDNA region of Plasmodium relictum (accession number NC012426) was used as the positive control [22]. The synthetic DNA was diluted to a starting concentration of 106 copies/µl. The starting solution was serially diluted 10-fold to prepare a series of solutions from the 106 copies of genomic DNA (gDNA) per µl down to 1 copy/µl. All samples were run in duplicates [13]. Parasite intensity, which refers to the number of parasite DNA copies per 100 avian red blood cells, was determined for each sample based on the qPCR data and sample DNA concentration following the method of Friedl & Groscurth [23], with theassumption that the average genome size of a passerine bird was 2.8 pg [24].
2.5. Statistical analysis
3. Results
3.1. Haemosporidian infections based on the microscopical and molecular analyses
Overall, the most frequently captured bird species was the Eurasian blackcap, Sylvia atricapilla (17.3 %). However, the European robin Erithacus rubeculla was the most frequently caught species among the birds in the natural habitats (42.98%) and the great tit Parus major was the most common of the birds caught in the urbanized habitat (16.44%), (Figure S1).
The presence of haemosporidian parasites on the blood smears was detected in 52 individuals (17.22%). The most abundant species was H. (Parahaemoproteus) parabelopolskyi on blood smears of 21 birds (Figure 1), which is a host specific primarily to the black cap Sylvia atricapilla, and it was detected in birds from both habitat types. The second most numerous species was H. (P.) balmorali (Figure 2), which was found on blood smears of 13 bird samples obtained from only one host species, the European robin E. rubeculla, and only in birds from the natural habitats. No haemosporidians were identified in 18 bird samples. The meronts of Plasmodium were detected on blood smears of five birds. Leucocytozoon was found only on blood smears of 2 individuals.
The DNA of parasites by nested PCR was detected in 112/317 individuals (35.3%; CI 30.1-40.6%) of 18 bird species. Parasites of the genera Haemoproteus and Plasmodium were present in 96 individuals (30.28 %; CI 25.2-35.3 %), and parasites of the genus Leucocytozoon were present in 29 individuals (9.15 %; CI 6.0-12.3%), Table 1. Thirteen birds (4.10 %; CI 1.9-6.3%) were co-infected, Table 2.
The infection rate of the genera Haemoproteus and Plasmodium in the birds caught in natural habitats was found to be 43.80% (53/121; CI 34.8-53.1%). The total infection rate of thebirds from urbanized habitats was in totally 21.94% (43/196; CI 16.4-28.4%), in spring 10.67% (8/75; CI 4.7-19.9%). The prevalence of the Haemoproteus and Plasmodium species in the birds in natural habitats was significantly higher that of the birds in urbanized habitats, in total and in spring (p < 0.05). Of the birds in natural habitats, 8.26% (10/121; CI: 4.0-14.7%) were infected with Leucocytozoon in total 9.69% (19/196; CI: 5.9-14.7%), in spring 8.00% (6/75; CI 3.0-16.6%) of the birds in urbanized habitats were infected with Leucocytozoon. There was no statistically significant difference in total infection rates for Leucocytozoon neither overall nor in the spring (p > 0.05). Co-infections were found in total 3.57% (7/196; CI 1.5-7.2%), in spring in 1.33% (1/75; CI 0.0-7.2%) of the birds in urbanized habitats and in 4.96% (6/121; CI 1.8-10.5%) of the birds in natural habitats, and there was no statistically significant difference between the two groups (p > 0.05).
All 125 positive samples were sequenced. The samples with infections (PCR positive for the presence of species of Haemoproteus and Plasmodium, and thereby Leucocytozoon) were sequenced using both sets of inner primers. After comparing the obtained 81 sequences with known sequences in the GenBank and MalAvi databases, we identified ten species: H. (P.) balmorali, H. (P.) concavocentralis, H. (P.) majoris, H. (P.) minutus, H. (P.) pallidus, H. (P.) parabelopolskyi, P. (G.) circumflexum, P. (H.) relictum, P. (N.) vaughani, and P. (H.) matutinum. Sequences from 13 individuals were unusable, and 2 samples showed double peaks, indicating co-infections (Table 1).
Birds from the urbanized habitats were infected with six Haemoproteus spp. and Plasmodium spp. (evaluated by nested PCR based on cyt b), and birds from the natural habitat were infected with nine. The most abundant parasite species in the birds from the urbanized habitats were H. (P.) parabelopolskyi SAYT01, SYAT02, SYAT07, SYAT10 lineages, with a prevalence of 39.29%, P. (G.) circumflexum SYABOR02 lineage (21.74%), P. (H.) relictum SGS1 lineage (10.00%), and P. matutinum LINN1 lineage (8.82%). Haemoproteus (P.) majoris CWT4 lineage and P. (N.) vaughani SYAT05 were not considered in the analyses because of the low number of infected hosts. In the birds from the natural habitats, H. (P.) parabelopolskyi SYAT01, SYAT02 lineages infections were also the most prevalent (40.63%), followed by H. (P.) balmorali ROBIN1/LULU1 lineage (15.91%), P. (H.) relictum SGS1, DP, COLL1 lineages (8.16%), and P. (G.) circumflexum TURDUS1 lineage (6.52%) (excluding H. (P.) concavocentralis HAWF2 lineage, H. (P.) minutus TURDUS2 lineage, P. (N.) vaughaini SYAT05 lineage, P. matutinum LINN1 lineage because of the low number of tested birds). Haemoproteus (P.) parabelopolskyi was detected in birds from both habitat types but only in one host Eurasian blackcap S. atricapilla. Interestingly, we found this parasite in the blood of a seven-year-old female, which was confirmed by ringing data from 2006. Otherwise, H. (P.) balmorali ROBIN1/LULU1 lineage was present in a wide range of hosts (five species), though only in birds captured in the natural habitats (Table 1). No unique lineages were recorded.
The intensity of parasites using qPCR analysis was verified with 52 microscopically positive samples. The intensity varied from 1 to 15,000 DNA copies per 100 avian red blood cells in both types of habitats. Low parasitemia (1-10 parasites per 10,000 erythrocytes) estimated from blood smears by microscopy corresponded on average to 1-1,000 DNA copies per 100 avian red blood cells. Medium parasitemia (11-100 parasites per 10,000 erythrocytes) estimated from blood smears by microscopy corresponded on average to 1,000-10,000 DNA copies per 100 avian red blood cells. And, the high parasitemia (more than 100 parasites per 10,000 erythrocytes) estimated from blood smears by microscopy corresponded on average more than 10,000 DNA copies per 100 avian red blood cells.
Co-infection withmore haemosporidian genera was detected in 4.10% of the individuals (13/317; CI 1.9-6.3%). Co-infections with Haemoproteus and Leucocytozoon parasites were detected in five birds, and co-infections with Plasmodium and Leucocytozoon were detected in eight individuals, by microscopy and molecular biology (Table 2). Co-infections with Haemoproteus and Plasmodium were not analyzed in detail in this study.
3.2. Haemosporidian infections in juvenile birds based on the microscopical and molecular analyses
Important markers of haemosporidians transmitted in Slovakia include juvenile birds caught in urbanized habitats. In our study, out of 114 juveniles, 26 were infected with Haemoproteus and Plasmodium (22.81%; CI 15.1-30.5%), 8 were infected with Leucocytozoon (7.02%; CI 2.3-11.7%) and 6 carried co-infections (5.26%; CI 1.2-9.4%). The juvenile birds the great tit Parus major, Eurasian blue tit Cyanistes caeruleus, and Eurasian blackcap S. atricapilla caught in urbanized habitats carried P. (H.) relictum SGS1 lineage, and the common blackbird T. merula carried P. (N.) vaughani SYAT05 lineage, the song thrush Turdus philomelos was infected with P. (G.) circumflexum SYBOR02 lineage, the Eurasian blackcap S. atricapilla was infected with H. (P.) parabelopolskyi SYAT02 lineage, and the common whitethroat Sylvia communis was infected with H. (P.) majoris CWT4 lineage. The lack of gametocytes made species determination by microscopy impossible for six PCR-positive juveniles the yellowhammer Emberiza citrinella, the common blackbird T. merula, and the common chaffinch Fringilla coelebs that were infected with Haemoproteus spp. The European robin Erithacus rubecula and the common blackbird T. merula were infected with Plasmodium spp., and Leucocytozoon spp. was found in the great tit P. major, the Eurasian blue tit C. caeruleus, the common blackbird T. merula, the Eurasian blackcap S. atricapilla, and the song thrush T. philomelos.
Mature gametocytes were confirmed in the peripheral blood of 11 juvenile birds: H. (P.) parabelopolskyi in 6 Eurasian blackcap S. atricapilla, P. (N.) vaughani in one the common blackbird T. merula, Haemoproteus sp. in one yellowhammer E. citrinella, two S. atricapilla and Plasmodium sp. in one the European robin E. rubecula.
4. Discussion
Many studies have compared the detection limit of microscopy and PCR, and while some have estimated PCR as being more sensitive, others have not [3,27,28,29,30,31,32,33]. Different methodological approaches have led to differing prevalence rates of parasites [34,35,36,37,38,39]. Despite the simplicity and speed of PCR analyses, microscopy remains essential [40]. Detection of a parasite’s DNA in a host’s blood does not confirm the host’s competence in the parasite life cycle. The presence of a parasite’s DNA in the bloodstream of a bird after taking blood meal by an insect does not mean that merozoites can infect erythrocytes and, consequently, develop the gametocytes necessary for assembling the life cycle. Another consideration in PCR is the overlooking of co-infections (what can be distinguished as double peaks in chromatograms) in samples, as general PCR protocols tend to favour the amplification of a parasite with higher parasitemia or the amplification with the best matching sequence to the primers [41]. Whether a bird is susceptible to parasites can be confirmed only by finding gametocyte-infected erythrocytes, and even better by experimental infections. Therefore, microscopical analyses of blood smears remain essential.
In our study, as mentioned above, microscopy failed to identify any species of haemosporidians in 18 samples. It could be caused by e.g. poor preparation of blood smears, poor staining, or deformation of erythrocytes. The meronts of Plasmodium spp. were detected in five birds. They are numerous in the blood of avian hosts in the early stages of infection. The absence of the necessary number of gametocytes enabled the identification of the parasite species. These facts could explain differences between positive findings by the microscopical method and the method of molecular biology.
The widely used nested PCR test mentioned above is sufficient to identify the genus and lineage. However, it is ineffective in distinguishing mixed Haemoproteus and Plasmodium infections because it favours amplification of the most abundant parasite in the sample or the parasite for which the primers are more suitable [34,42]. Mixed infections are very common and have been shown to be particularly virulent [40]. Thus, knowledge of mixed infection is useful and additional molecular biology methods, such as the one-step multiplex PCR developed by Ciloglu et al [42], are essential. We can expect to find more birds with mixed infections using these methods and would therefore like to apply it in the future.
In the present study, we aimed to extent knowledge about the presence of haemosporidians in the blood samples of birds in urbanized and natural habitats and identify which species are transmitted in Slovakia using microscopy and molecular detection. Here, we report ten species of haemosporidian parasites found in the birds in Slovakia. More diverse species were found in the birds in the natural habitats, where most of the birds were migratory species compared to those in the urbanized habitats with local populations (Table 1). In combination with suitable vectors, infected birds could serve as reservoirs of infection for other birds (because of sharing the same habitat for rest and at night). Differences in prevalence may be attributable not only to vectors and hosts but also to abiotic environmental factors such as precipitation and mean annual temperature [43,44,45,46,47]. In our study, it was shown that the birds caught in the natural habitats had significantly higher infection rates than those of the local populations in the urbanized habitats. Haemoproteus dominated in the natural habitats (40.38% vs. 29.89%), which was comparable to a previous study [12], and Plasmodium dominated in urbanized habitats (33.3% vs. 18.0%). The highest haemosporidian infections were recorded in the Eurasian blackcap, the great tit, the common chaffinch, the common blackbird, and the Eurasian blue tit, and these results are related to host abundance and are similar to the results of Šujanová et al. [12]. As recorded previously, Haemoproteus species is the most frequently reported haemosporidian parasite in birds, and this result is not limited to Slovakia [34,48,49,50].
To identify which parasites are transmitted in Slovakia, it was necessary to analyze the juveniles. Unlike adult birds that can become infected in any year or locality, young birds in their first year of life (before autumn migration) can be infected only by parasites actively transmitted by vectors in their breeding area [1,6,7,8]. In the populations in the natural habitats, the juveniles were not mist-netted, , so their infection status remains unknown. Unlike in the local populations in the urbanized habitats, in the caught juveniles, the blood analysis showed the presence of H. (P.) parabelopolskyi, H. (P.) majoris, P. (G.) circumflexum, P. (N.) vaughani, P. (H.) relictum, and Leucocytozoon sp. Nourani et al. [51] showed age does not influence the infection rate of Haemoproteus.
Haemoproteus parasites are the most diverse group of avian haemosporidians [50]. Due to limited knowledge of life cycles and tissue merogony of the majority of Haemoproteus species, these pathogens usually have been considered benign [52]. However, recent molecular studies indicate that they may cause severe and even lethal diseases if infections occur in non-adapted avian hosts [53]. The findings in our study about the species H. (P.) parabelopolskyi and H. (P.) balmorali were interesting. Haemoproteus (P.) parabelopolskyi was the most numerous species found (26 individuals infected). As Pérez-Tris et al. [54] found, this species is a strict host specialist in the Eurasian blackcap S. atricapilla. However, H. (P.) parabelopolskyi can be occasionally found in birds of the Acrocephalidae family [2]. We reported the occurrence of this parasite in both habitat types, which was likely due to the fact that the Eurasian blackcap S. atricapilla is one of the most numerous bird species in Slovakia. Evidence of the presence of a suitable vector was confirmed by the infections in eight juveniles. Incredibly marvellous for us was the finding of a seven-year-old female Eurasian blackcap S. atricapilla infected with this parasite. The infection was confirmed by microscopy, PCR, and sequencing. The microscopy showed a very weak intensity of infection, and together, the ability of active flight predicted the chronic state of infection. We are unable to say when the individual became infected, but this finding illustrates the usage of migration routes in east Slovakia. In contrast, H. (P.) balmorali (the second most prevalent species, with 19 infected individuals from 6 host species) was detected in the natural habitats eventhough these species were common in the other habitats. Hauptmanová et al. [10] noted the presence of this parasite for the first time in Slovakia in the blood of a European robin (E. rubecula) caught in the eastern part of Slovakia. Unfortunately, the authors did not specify the number of infected birds. As vertebrate hosts, Valkiūnas [1] referenced numerous species of the Passeriformes order belonging to Muscicapidae and Turdidae (over 40 species). This could be a marker for the presence of a suitable vector in the east of the country, but to verify this premise, other studies should confirm or disconfirm this proposition. Because of evidence of sporogony in the salivary glands of experimentally infected Culicoides impunctatus, Valkiūnas [1] identified it as a suitable vector. According to the Fauna Europaea database [55,56], C. impunctatus was recorded in Slovakia, as well as in the surrounding states.
5. Conclusions
This study contributes to the knowledge about the prevalence, and morphological and molecular richness of the haemosporidian parasites circulating in free-living birds in urbanized and natural habitats in Slovakia. The prevalence of the Haemoproteus and Plasmodium species in birds in natural habitats was significantly higher than that in birds in urbanized habitats (p < 0.05), though the prevalence of Leucocytozoon was not higher for urban habitats, neither overall nor in the spring. In 42 avian species 10 haemosporidian species H. (P.) balmorali, H. (P.) concavocentralis, H. (P.) majoris, H. (P.) minutus, H. (P.) pallidus, H. (P.) parabelopolskyi, P. (G.) circumflexum, P. (H.) relictum, P. (N.) vaughani, and P. matutinum were identified. However, the species H. (P.) majoris, H. (P.) parabelopolskyi, P. (G.) circumflexum, P. (H.) relictum, and P. (N.) vaughani were present also in the juveniles, which indicates the presence of suitable vectors and the active transmission of parasites in Slovakia.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Frequencies of the avian species captured in urbanized (UH) and natural (NH) habitats in Slovakia.
Author Contributions
Formal analysis, Lenka Minichová and Eva Špitalská; Funding acquisition, Eva Špitalská; Investigation, Lenka Minichová, Vladimír Slobodník, Roman Slobodník, Milan Olekšák and Zuzana Hamšíková; Methodology, Lenka Minichová and Eva Špitalská; Project administration, Eva Špitalská; Supervision, Ľudovít Škultéty and Eva Špitalská; Writing – original draft, Lenka Minichová; Writing – review & editing, Eva Špitalská. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Scientific Grant Agency of Ministry of Education, Science, Research and Sport of the Slovak Republic and Slovak Academy of Science, grant number VEGA 2/0021/21.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the first and corresponding author upon request.
Compliance with ethical standards: The experiments presented in this paper comply with current laws of the Slovak Republic. Birds were captured, ringed, blood sampled and released under the permission of the Ministry of Environment of the Slovak Republic No. 9368/2011-2.2.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Valkiūnas, G. Avian Malaria Parasites and Other Haemosporidia; CRC press: Boca Raton, FL, USA, 2005. [Google Scholar]
- Valkiūnas, G.; Iezhova, T.A. Keys to the avian Haemoproteus parasites (Haemosporida, Haemoproteidae). Malaria Journal 2022, 21, 269. [Google Scholar] [CrossRef]
- Hellgren, O.; Waldenström, J.; Bensch, S. A new PCR assay for simultaneous studies of Leucocytozoon, Plasmodium, and Haemoproteus from avian blood. J. Parasitol. 2004, 90, 797–802. [Google Scholar] [CrossRef]
- Bensch, S.; Hellgren, O.; Pérez-Tris, J. MalAvi: A public database of malaria parasites and related haemosporidians in avian hosts based on mitochondrial cytochrome b lineages. Mol. Ecol. Resour. 2009, 9, 1353–1358. [Google Scholar] [CrossRef]
- MalAvi database. Available online: http://mbio-serv2.mbioekol.lu.se/Malavi/index.html (accessed on 8 March 2023).
- Kučera, J. Blood parasites of birds in Central Europe. 1.Survey of literature. The incidence in domestic birds and general remark to the incidence in wild birds. Folia Parasitologica 1981, 28, 13–22. [Google Scholar]
- Kučera, J. Blood parasites of birds in Central Europe. 2. Leucocytozoon. Folia Parasitologica 1981, 28, 193–203. [Google Scholar]
- Kučera, J. Blood parasites of birds in Central Europe. 3. Plasmodium and Haemoproteus. Folia Parasitologica 1981, 28, 303–312. [Google Scholar] [PubMed]
- Hauptmanová, K.; Benedikt, V.; Literák, I. Blood Parasites in Passerine Birds in Slovakian East Carpathians. Acta Protozool. 2006, 45, 105–109. [Google Scholar]
- Haas, M.; Kisková, J. Absence of blood parasites in the Alpine Accentor Prunella collaris. Oecologia Montana 2010, 19, 30–34. [Google Scholar]
- Berthová, L.; Országhová, Z.; Valkiūnas, G. The first report of nine species of haemosporidian parasites (Haemosporida: Haemoproteus, Plasmodium and Leucocytozoon) in wild birds from Slovakia. Biologia 2012, 67, 931–933. [Google Scholar] [CrossRef]
- Šujanová, A.; Špitalská, E.; Václav, R. Seasonal dynamics and diversity of haemosporidians in a natural woodland bird community in Slovakia. Diversity 2021, 13, 439. [Google Scholar] [CrossRef]
- Kazimírová, M.; Hamšíková, Z.; Kocianová, E.; Marini, G.; Mojšová, M.; Mahríková, L.; Berthová, L.; Slovák, M.; Rosá, R. Relative density of host-seeking ticks in different habitat types of south-western Slovakia. Exp. Appl. Acarol. 2016, 69, 205–224. [Google Scholar] [CrossRef]
- Berthová, L.; Slobodník, V.; Slobodník, R.; Olekšák, M.; Sekeyová, Z.; Svitálková, Z.; Kazimírová, M.; Špitalská, E. The natural infection of birds and ticks feeding on birds with Rickettsia spp. and Coxiella burnetii in Slovakia. Exp. Appl. Acarol. 2016, 68, 299–314. [Google Scholar] [CrossRef]
- Svensson, L. Identification guide to European passerines, 4th ed.; Lars Svensson, Stockholm, 1992. [Google Scholar]
- Hromádko, M.; Horáček, J.; Chytil, J.; Pithart, K.; Škopek. J. Guide to the determination of our passerines, Publisher ZUV, Hradec Králové (in Czech). 1992.
- Hromádko, M.; Horáček, J.; Chytil, J.; Pithart, K.; Škopek. J. Guide to the determination of our passerines, Publisher ZUV, Hradec Králové (in Czech). 1993.
- Hromádko, M.; Horáček, J.; Chytil, J.; Pithart, K.; Škopek., J. Guide to the determination of our passerines; Publisher ZUV, Hradec Králové (in Czech), 1998. [Google Scholar]
- Bensch, S.; Stjernman, M.; Hasselquist, D.; Ostman, O.; Hansson, B.; Westerdahl, H.; Pinheiro, R.T. Host specificity in avian blood parasites: A study of plasmodium and haemoproteus mitochondrial DNA amplified from birds. Proc. Biol. Sci. 2000, 267, 1583–1589. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A. , Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- GenBank database. Available online: http://blast.ncbi.nlm.nih.gov/ (accessed on 22 February 2023).
- Bell, J.A.; Weckstein, J.D.; Fecchio, A.; Tkach, V.V. A new real-time pcr protocol for detection of avian haemosporidians. Parasites Vectors 2015, 8, 383. [Google Scholar] [CrossRef] [PubMed]
- Friedl, T.W.P.; Groscurth, E. A real-time PCR protocol for simple and fast quantification of blood parasite infections in evolutionary and ecological studies and some data on intensities of blood parasite infections in a subtropical weaverbird. J. Ornithol. 2012, 153, 239–247. [Google Scholar] [CrossRef]
- Tiersch, T.R.; Wachtel, S.S. On the evolution of genome size of birds. J. Hered. 1991, 82, 363–368. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeo. Electronica 2001, 4, 4, http://palaeo-electronica.org/2001_1/past/issue1_01.htm. [Google Scholar]
- EasyCalculation. Available online: https://www.easycalculation.com/statistics (accessed on 22 February 2023).
- Valkiũnas, G.; Izheova, T.A.; Križanauskiene, A.; Palinauskas, V.; Sehgal, R.N.M.; Bensch, S. A comparative analysis of microscopy and PCR-based detection methods for blood parasites. J. Parasitol. 2008, 94, 1395–1401. [Google Scholar] [CrossRef] [PubMed]
- Jarvi, S.I.; Schultz, J.J.; Atkinson, C.T. PCR diagnostics underestimate the prevalence of avian malaria (Plasmodium relictum) in experimentally-infected passerines. J. Parasitol. 2002, 88, 153–158. [Google Scholar] [CrossRef]
- Richard, F.A.; Sehgal, R.N.M.; Jones, H.I.; Smith. T.B. A comparative analysis of PCR-based detection methods for avian malaria. J. Parasitol. 2002, 88, 819–822. [Google Scholar] [CrossRef]
- Waldenstrom, J. , Bensch, S.; Hasselquist, D.; Ostman, O. A new nested polymerase chain reaction method very efficient in detecting Plasmodium and Haemoproteus infections from avian blood. J. Parasitol. 2004, 90, 191–194. [Google Scholar] [CrossRef]
- Krone, O.; Valkiũnas, G.; Lessow, O.; Müller, K.; Iezhova, T.A.; Fickel, J.; Bensch, S. Haemosporidian blood parasites in european birds of prey and owls. J. Parasitol. 2087, 94, 709–715. [Google Scholar] [CrossRef]
- Fallon, S.; Ricklefs, R.E. Parasitemia in PCR-detected Plasmodium and Haemoproteus infections in birds. J. Avian Biol. 2008, 39, 514–522. [Google Scholar] [CrossRef]
- Garamszegi, L.Z. The sensitivity of microscopy and PCR-based detection methods affecting estimates of prevalence of blood parasites in birds. J. Parasitol. 2010, 96, 1197–1203. [Google Scholar] [CrossRef]
- Schumm, Y.R.; Bakaloudis, D.; Barboutis, C.; Cecere, J.G.; Eraud, C.; Fischer, D.; Hering, J.; Hillerich, K.; Lormée, H.; Mader, V.; Masello, J.F.; Metzger, B.; Rocha, G.; Spina, F.; Quillfeldt, P. Prevalence and genetic diversity of avian haemosporidian parasites in wild bird species of the order Columbiformes. Parasitol. Res., 2021, 120, 1405–1420. [Google Scholar] [CrossRef] [PubMed]
- Lynton-Jenkis, J.G.; Chaine, A.S.; Russell, A.F.; Bonneaud, C. Parasite detection and quantification in avian blood is dependent on storage medium and duration. Ecol. Evolution, 2023, 13, e9819. [Google Scholar] [CrossRef] [PubMed]
- Noni, V.; Tan, C.S. Prevalence of haemosporidia in Asian Glossy Starling with discovery of misbinding of Haemoproteus-specific primer to Plasmodium genera in Sarawak, Malaysian Borneo. BMC Vet Res 2023, 19, 66. [Google Scholar] [CrossRef] [PubMed]
- Ehsani Amrei, S.F.; Motevalli Haghi, A.; Nateghpour, M.; Mohebali, M.; Youssefi, M.; Raiesi, O.; Mahdavi, S.; Farivar, L. Detection of Malaria Parasites and Other Haemosporidia in Migratory and Native Birds in Mazandaran and Golestan Provinces, Iran. J Arthropod Borne Dis. 2022, 16, 26–33. [Google Scholar] [CrossRef]
- Rahmati, M.; Norouzian, H.; Nayebzadeh, H.; Shokrani, H. Molecular Detection and Phylogenetic Analysis of Avian Haemosporidian Parasites in House Sparrows (Passer domesticus) of Western Iran. J Wildl Dis. 2023. [Google Scholar] [CrossRef]
- Chavarría, X.; Matta, N.E.; Cadena-Ortíz, H.; Alarcón, I.; Bahamonde-Vinueza, D.; González, A.D.; Bonaccorso, E. Haemosporidian parasites in the ash-breasted Sierra finch (Geospizopsis plebejus): insights from an Andean dry forest population. Parasitology, 2022, 150, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Valkiūnas, G.; Bensch, S.; Iezhova, T.A.; Križanauskiene, A.; Hellgren, O.; Bolshakov, C.V. Nested cytochorome B polymerase chain reaction diagnostics underestimate mixed infections of avian blood haemosporidian parasites: Microscopy is still essential. J. Parasitol. 2006, 92, 418–422. [Google Scholar] [CrossRef] [PubMed]
- Zehtindjiev, P.; Križanauskienė, A.; Bensch, S.; Palinauskas, V.; Asghar, M.; Dimirov, D.; Scebba, S.; Valkiūnas, G. A new morphologically distinct avian malaria parasite that fails detection by established polymerase chain reaction-based protocols for amplification of the cytochrome b gene. J. Parasitol. 2012, 98, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Ciloglu A, Ellis VA, Bernotienė R, Valkiūnas G, Bensch S. A new one-step multiplex PCR assay for simultaneous detection and identification of avian haemosporidian parasites. Parasitol Res 2019, 118, 191–201. [CrossRef] [PubMed]
- Basto, N.; Rodríguez, O.A.; Marinkelle, C.J.; Gutiérrez, R.; Matta, N. Haematozoa in birds from La Macarena National Natural Park (Colombia). Caldasia. 2006, 28, 371–7. [Google Scholar]
- Doussang, D. , González-Acuña, D., Torres-Fuentes, L.G. et al. Spatial distribution, prevalence and diversity of haemosporidians in the rufous-collared sparrow, Zonotrichia capensis. Parasites Vectors 2019, 12, 2. [Google Scholar] [CrossRef]
- Ciloglu, A.; Ergen, A.G.; Inci, A.; Dik, B.; Duzlu, O.; Onder, Z.; Yetismis, G.; Bensch, S.; Valkiūnas, G.; Yildrim, A. Prevalence and genetic diversity of avian haemosporidian parasites at an intersection point of bird migration routes: Sultan Marshes National Park, Turkey. Acta Trop. 2020, 210, 105465. [Google Scholar] [CrossRef]
- Quillfeldt, P.; Arriero, E.; Martínez, J.; Masello, J.F.; Merino, S. Prevalence of blood parasites in seabirds- a review. Front Zool. 2011, 8, 26. [Google Scholar] [CrossRef]
- Chakarov, N.; Kampen, H.; Wiegmann, A.; Werner, D.; Bensch, S. Blood parasites in vectors reveal a united blackfly community in the upper canopy. Parasites Vectors 2020, 13, 309. [Google Scholar] [CrossRef]
- Carlson, J.; Martínez-Gómez, J.E.; Valkiūnas, G.; Loiseau, C.; Bell, D.A.; Sehgal, R.N. Diversity and phylogenetic relationships of hemosporidian parasites in birds of Socorro Island, México and their role in the re-introduction of the Socorro Dove (Zenaida graysoni). J Parasitol 2013, 99, 270–276. [Google Scholar] [CrossRef]
- Heym, E.C.; Kampen, H.; Krone, O.; Schäfer, M.; Werner, D. Molecular detection of vector-borne pathogens from mosquitoes collected in two zoological gardens in Germany. Parasitol Res 2019, 118, 2097–2105. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Ilgūnas, M.; Bukauskaitė, D.; Chagas, C.R.F.; Bernotienė, R.; Himmel, T.; Harl, J.; Weissenböck, H.; Iezhova, T.A. Molecular characterization of six widespread avian haemoproteids, with description of three new Haemoproteus species. Acta Tropica 2019, 197, 105051. [Google Scholar] [CrossRef] [PubMed]
- Nourani, L.; Aliabadian, M.; Dinparast Djadid, N.; Mirshamsi, O. Occurrence of Haemoproteus spp. (Haemosporida: Haemoproteidae) in New Host Records of Passerine Birds from the East of Iran. Iran J Parasitol. 2018, 13, 267–274. [Google Scholar] [PubMed]
- Bennet, G.F.; Peirce, M.A.; Ashford, R.W. Avian haematozoa: mortality and pathogenicity. J. Nat. Hist. 1993, 27, 993–1001. [Google Scholar] [CrossRef]
- Ortiz-Catedral, L.; Brunton, D.; Stidworthy, M.F.; Elsheikha, H.M.; Pennycott, T.; Schulze, C.; Braun, M.; Wink, M.; Gerlach, H.; Pendl, H.; Gruber, A.D.; Ewen, J.; Pérez-Tris, J.; Valkiūnas, G.; Olias, P. Haemoproteus minutus is highly virulent for Australasian and South American parrots. Parasit. Vectors 2019, 12, 40. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Tris, J.; Hellgren, O.; Križanauskienė, A.; Waldenström, J.; Secondi, J.; Bonneaud, C.; Fjeldsȧ, J.; Hasselquist, D.; Bensch, S. Within-Host Speciation of Malaria Parasites. PLoS ONE 2007, 2, e235. [Google Scholar] [CrossRef]
- de Jong, Y.; Verbeek, M.; Michelsen, V.; Bjørn, P.; Los, W.; Steeman, F.; Bailly, N.; Basire, C.; Chylarecki, P.; Stloukal, E.; Hagedorn, G.; Wetzel, F.; Glöckler, F.; Kroupa, A.; Korb, G.; Hoffmann, A.; Häuser, C.; Kohlbecker, A.; Müller, A.; Güntsch, A.; Stoev, P.; Penev, L. Fauna Europaea – all European animal species on the web. Biodiversity Data Journal 2014, 2, e4034. [Google Scholar] [CrossRef]
- Fauna Europaea database Available online:. Available online: http://www.faunaeur.org/distribution_table.php (accessed on 22 February 2023).
Figure 1.
Gametocytes of H. (P.) parabelopolskyi in blood of the Eurasian blackcap Sylvia atricapilla.
Figure 1.
Gametocytes of H. (P.) parabelopolskyi in blood of the Eurasian blackcap Sylvia atricapilla.
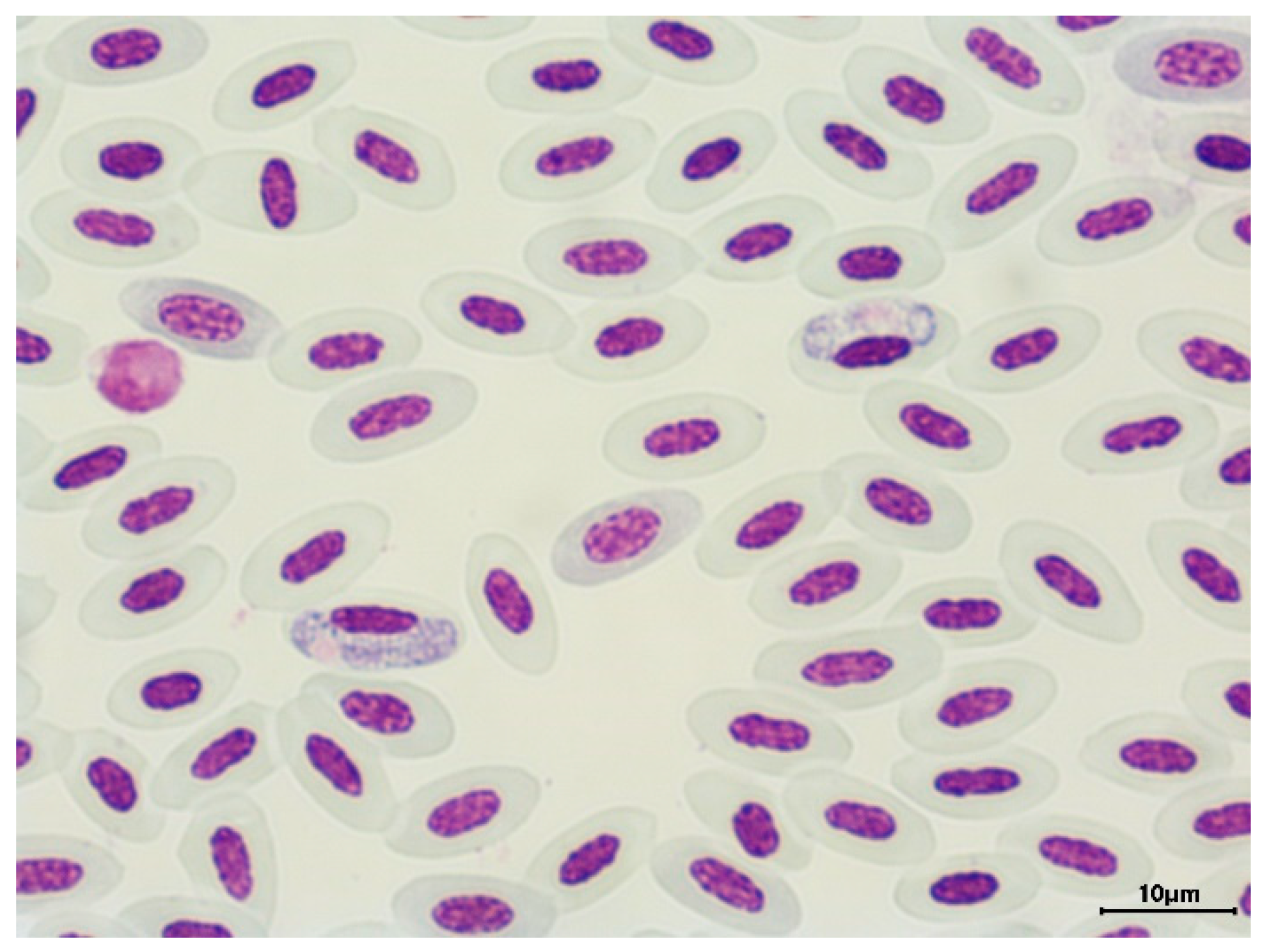
Figure 2.
Gametocytes of H. (P.) balmorali in the blood of the European robin Erithacus rubecula.
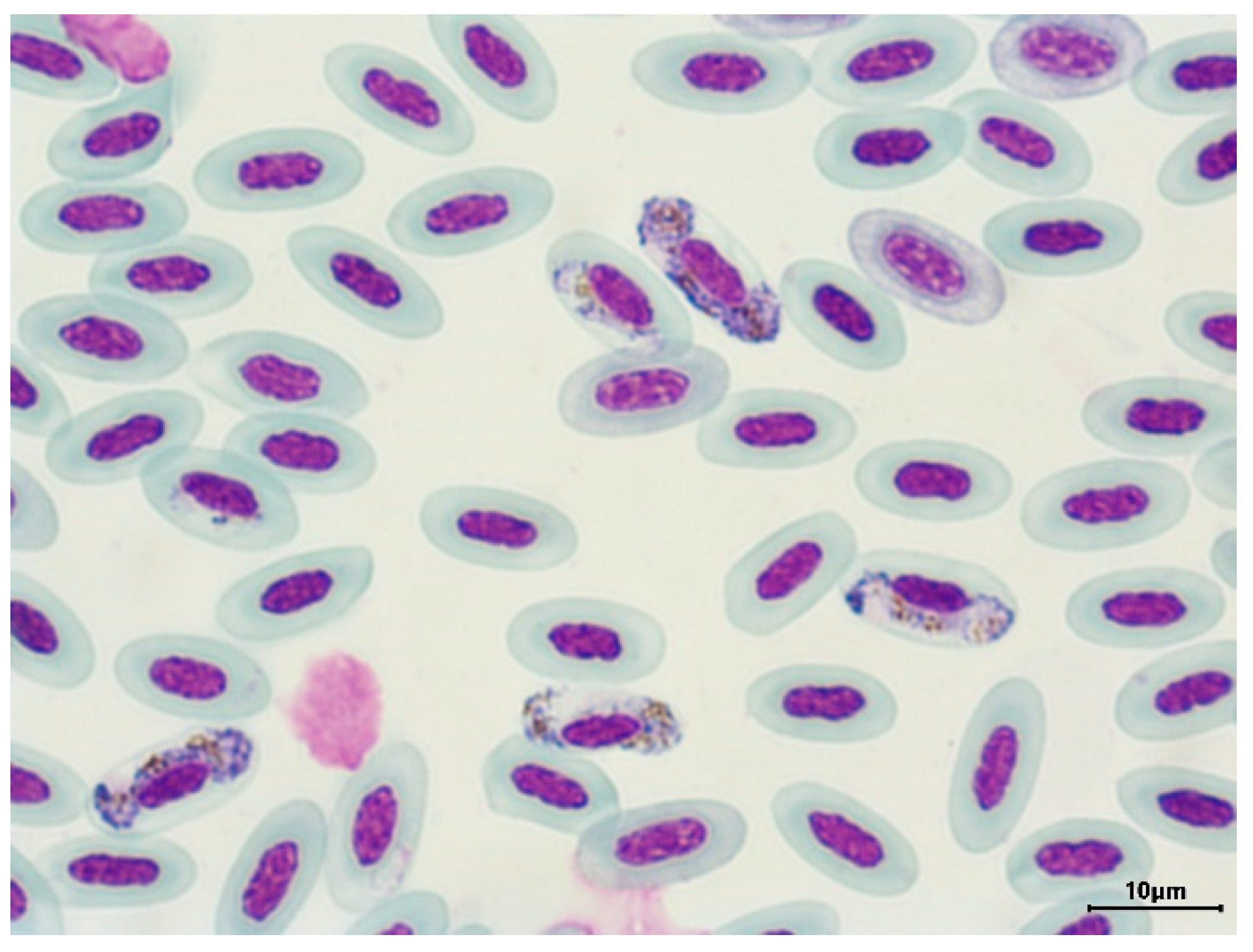

Table 1.
List of the haemosporidian parasites detected by cyt b molecular analysis in the blood samples of the birds from urbanized and natural habitats (us - unusable sequences, dp - double peaks). Numbers in parentheses refer to the number of positive birds of given lineage.
Table 1.
List of the haemosporidian parasites detected by cyt b molecular analysis in the blood samples of the birds from urbanized and natural habitats (us - unusable sequences, dp - double peaks). Numbers in parentheses refer to the number of positive birds of given lineage.
| Bird species | Prevalence | Haemosporidian species | Haemosporidian Lineage in MALAVI | Haemosporidian Lineage in GenBank |
| Urbanized habitats | ||||
| Parus major | 5/37 | P. (Haemamoeba) relictum | SGS1 (8) | MK652232 |
| Parus montanus | 1/6 | |||
| Cyanistes caeruleus | 1/9 | |||
| Sylvia atricapilla | 1/28 | |||
| Sylvia atricapilla | 11/28 | H. (P.) parabelopolskyi | SYAT02 (7) SYAT10 (1) SYAT01 (2) SYAT07 (1) |
JN164719 AY831757 AY831750 AY831754 |
| Sylvia communis | 1/1 | H. (P.) majoris | CWT4 | KJ488675 |
| Fringilla coelebs | 4/6 | Haemoproteus (P.) sp. | CCF2 (3) CCF1 (1) |
AF495551 AF495550 |
| Emberiza citrinella | 1/1 | EMCIR01 | GU085190 | |
| Sylvia atricapilla | 3/28 | us (3) | ||
| Turdus merula | 1/9 | us | ||
| Turdus merula | 6/9 | P. (N.) vaughani | SYAT05 (6) | DQ847271 |
| Turdus merula | 1/9 | P. (H.) matutinum | LINN1 (3) | DQ847270 |
| Erithacus rubecula | 1/10 | |||
| Emberiza schoeniculus | 1/15 | |||
| Emberiza schoeniculus | 2/15 | P. (Giovannolaia) circumflexum | SYBOR02 (2) | DQ368392 |
| Turdus philomelos | 2/2 | BT7 TURDUS1 |
MK062195 KM361492 |
|
| Prunella modularis | 1/6 | TURDUS1 | KM361492 | |
| Parus major | 12/37 | Leucocytozoon sp. | PARUS19 (4) PARUS28 (1) PARUS4 (4) PARUS18 (1) STUR1/TURPEL01 (1) us (1) |
HM234027 JX855047 AY393795 HM234026 DQ847246/ KT376971 |
| Cyanistes caeruleus | 3/9 | PARUS19 PARUS4 PARUS50 |
HM234027 AY393795 JX855065 |
|
| Turdus merula | 1/9 | ON932273 | ||
| Turdus philomelos | 1/2 | dp | ||
| Sylvia atricapilla | 1/28 | PARUS4 | AY393795 |
|
| Prunella modularis | 1/6 | PRUMOD01 | KJ488638 | |
| 62/196 | ||||
| Natural habitats | ||||
| Sylvia atricapilla | 13/32 | H. (P.) parabelopolskyi | SYAT01 (9) SYAT02 (4) |
JN164718 JN164719 |
| Erithacus rubecula | 10/42 | H. (Parahaemoproteus) balmorali | ROBIN1/LULU1 | AY393807 |
| Carduelis carduelis | 1/11 | |||
| Emberiza schoeniculus | 1/2 | |||
| Sylvia atricapilla | 1/32 | |||
| T. philomelos | 1/1 | |||
|
Coccothraustes coccothraustes |
1/3 | H. (P.) concavocentralis | HAWF2 | GQ396708 |
| Turdus merula | 1/4 | H. (P.) minutus | TURDUS2 | KM361485 |
| Erithacus rubecula | 1/42 | H. (P.) pallidus | SYAT03 | JN164720 |
| Carduelis carduelis | 1/11 | |||
| Erithacus rubecula | 5/42 | Haemoproteus (P.) sp | CIRCUM01 (1) us (4) |
KC994896 |
|
Coccothraustes coccothraustes |
1/3 | HAWF1 | DQ368348 | |
| Carduelis carduelis | 1/11 | us | ||
| Emberiza citrinella | 1/4 | CCF6 | DQ368341 | |
| Scolopax rusticola | 1/1 | us | ||
| Sylvia atricapilla | 1/32 | us | ||
| Sylvia curruca | 1/4 | LWT1 | AF495563 | |
| Erithacus rubecula | 2/42 | P. (G.) circumflexum | TURDUS1 | KM361492 |
| Emberiza citrinella | 1/4 | |||
| Emberiza cia | 1/2 | P. (H.) relictum | SGS1 | JX196866 |
| Erithacus rubecula | 1/42 | DP | ||
| Passer montanus | 1/2 | SGS1 | MK652232 | |
| Fringilla coelebs | 1/3 | COLL1 | AY831747 | |
| Turdus merula | 2/4 | P. (N.) vaughani | SYAT05 (2) | OP546094 |
| Turdus merula | 1/4 | P. matutinum | LINN1 | DQ847270 |
| Cyanistes caeruleus | 1/4 | Plasmodium sp. | us | |
| Erithacus rubecula | 5/42 | Leucocytozoon sp. | BT2 BT5 SFC8 SYCON06 PARUS19 |
AY393802 AY393798 DQ847234 KP688305 HM234027 |
|
Coccothraustes coccothraustes |
3/3 | HAWF1 (1) us (2) |
DQ368348 | |
| Scolopax rusticola | 1/1 | SCORUS01 | LC230149 | |
| Cyanistes caeruleus | 1/4 | PARUS4 | AY393795 | |
| 63/121 | ||||
Table 2.
List of birds co-infected with haemosporidian parasites. Identified lineages are listed.
| Avian species | Haemosporidian species |
| Urbanized habitats | |
| Turdus merula | P. (N.) vaughani SYAT05, L. (L.) sp. |
| Cyanistes caeruleus | P. (H.) relictum SGS1, L. (L.) sp. PARUS19 |
| Parus major | P. (H.) relictum SGS1, L. (L.) sp. PARUS4 |
| Parus major | P. (H.) relictum SGS1, L. (L.) sp. PARUS18 |
| Parus major | P. (H.) relictum SGS1, L. (L.) sp. STUR1/TURPEL01 |
| Sylvia atricapilla | P. (H.) sp., L. (L.) sp. PARUS4 |
| Turdus philomelos | P. (G.) circumflexum BT7, L. (L.) sp. |
| Natural habitats | |
| Erithacus rubecula | H. (P.) sp., L. (L.) sp. BT2 |
| Scolopax rusticola | H. (P.) sp., L. (L.) sp. |
| Erithacus rubecula | H. (P.) balmorali ROBIN1/LULU1, L. (L.) sp. SFC8 |
| Coccothraustes coccothraustes | H. (P.) sp. HAWF1, L. (L.) sp. |
| Erithacus rubecula | P. (G.) circumflexum TURDUS1, L. (L.) sp. SYCON06 |
| Coccothraustes coccothraustes | H. (P.) concavocentralis HAWF2, L. (L.) sp. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



