
Submitted:
10 October 2023
Posted:
11 October 2023
You are already at the latest version
Abstract
Cocoa bean quality depends mainly on genetic material, edaphoclimatic factors and post-harvest processes such as fermentation. The impact of the fermentation process on the chemical and func-tional composition of different cocoa clones grown in southern Colombia was analyzed. A facto-rial design with repeated measures over time was used to analyze the effect of clone and fermen-tation time on chemical characteristics (bromatology, phenolic compounds and antioxidant ac-tivity). The bromatological characteristics showed significant differences between clones and fermentation time. Clones EET-8 and CCN-51 showed higher contents of acidity (0.51%), fat (63.61%), protein (12.85%) and carbohydrates (1.63 mg g-1). Moisture, acidity and sucrose in-creased their values between day 4 and day 6 of fermentation (P<0.05). Phenolic compounds and antioxidant activity had significant differences between clones, where clones CCN-51 and ICS-95 had high contents of phenols (64.56 mg g-1), flavonoids (3.30 mg g-1) and DPPH reducing capacity (325.55 µmol g-1). In this sense, we consider the FSV-41 clone as the major grain quality index based on the results of chemical composition at bromatological level, antioxidant activity and phenolic compounds.
Keywords:
: theobroma cacao
; bromatological
; phenolic compounds
; antioxidant activity
1. Introduction
Cocoa (Theobroma cacao L.) is a cash crop with tropical distribution and its world production is around 4.955 million tons for the period 2021/22 (International Cocoa Organization [1], production that comes mainly from countries on the African and Latin American continent, which contribute around 70 and 13% of world volumes, respectively [2]. For producing countries, cocoa is an important economic, social, and cultural activity that is mostly developed in family farming [3]. In Latin America and the Caribbean, this activity is carried out by more than 350 thousand families and at least 1.7 million people depend on or directly benefit from its production [4].
In Colombia, there are 52,000 cocoa-producing families that depend directly on this crop [5]. This is produced in 30 of the 32 departments, where the department of Huila is the fourth largest producer of cocoa nationally, with a production of 4,197 t per year [6], production that is recognized for its quality, as it has the distinction "fine aroma", a situation that is recognized to only 7% of world production [7]. The cataloging of fine aroma cocoa for the origin of the department of Huila generates commercial opportunities for a sector that has been growing in area and production in recent years and has also gained recognition abroad, having greater export opportunities [7].
Cocoa bean quality depends on factors such as clone, crop management, climatic and edaphic factors, harvest, and post-harvest process (preconditioning of the pulp, fermentation, drying and roasting) and value-added processes (transformation of the raw material) [8]. In cocoa, fermentation is a prerequisite for the generation of flavor precursors [9], resulting in desirable changes in the chemical and sensory attributes of the product [10]. During this process, different chemical reactions occur that generate a large number of substances, among them, phenolic compounds and antioxidants that affect the organoleptic characteristics of the grain [11]. In addition, during fermentation, variations in temperature, pH and oxygen availability are generated, which affect protein, fat, and carbohydrate content [8].
Another factor affecting bean quality is the clone [12]. In this regard, Ramírez et al. [13] indicated that crosses between high quality clones ensure the production of fine cocoa beans. Likewise, Nazario et al. [14] stated that the highest antioxidant activity against 1-1-diphenyl-2-picrylhydrazyl radical (DPPH) is presented by clones ICS-95 and TSH-1188, whereas clones IMC-67 and CCN-51 did not present good activity. In addition, bean weight and size are a genetically inherited trait, being the clone and location determinants of some physical and chemical characteristics such as bean index, cob index, husk content, fat, fiber and protein [15]. Therefore, the objective of this work was to evaluate the effect of the fermentation process on the chemical and functional composition (content of phenolic compounds and antioxidant activity) in different cocoa clones grown in southern Colombia. To determine this variation, subindices (bromatological, antioxidant activity, phenolic compounds) and a grain quality index were developed in order to obtain a clone with a better chemical composition. The information generated will allow rural producers and cocoa associations to have the necessary information on the chemical composition of different clones during the fermentation process.
2. Materials and Methods
2.1. Study area and selection of cocoa clones
The study was carried out on farms in different municipalities located in the central region of the department of Huila (Rivera, Campoalegre, Hobo, Gigante, Algeciras and Tarqui). For the selection of the different farms (n=11), criteria such as: i. plots in production, ii. plots with an area greater than 1 ha of which 80% of the trees correspond to the same clone, and iii. plots with a shade canopy greater than 30%. Of the total number of selected farms, 32 plots were identified that had the different cocoa clones under study (Table 1). For each clone, four samples of cocoa beans were obtained from pods, each sample obtained from different plots. In each plot, they were harvested 250 mature pods of each clone were systematically harvested at a stage of development between 20 to 22 weeks [16], without the presence of pests (Monalonion dissimultum) and diseases (Moniliophthora roreri and Phytophthora sp.) [17], activity that was carried out between the months of March to June 2018, when the highest production pod is found. The cocoa clones found in the lots of the farms sampled are described in Table 1 which correspond to universal clones which have been introduced commercially and regional clones product of the identification of production traits by FEDECACAO and AGROSAVIA.
2.2. Fermentation, drying and roasting of cocoa beans
After harvesting, fresh cocoa bean samples of 4 kg of each clone were obtained and fermented in wooden crates of 1×1 m, located 1 m above the ground. This process was carried out at the cocoa bean collection center located in the municipality of Campoalegre (department of Huila), belonging to the Association of Small Cocoa Producers of Campoalegre, Huila (ASOPECA). The mass of each cocoa sample was placed in a tulle bag duly labeled with the information of the clone, lot and farm, samples that were subjected to the same fermentation process in order to avoid variations due to this process [21]. From the second day on, the cocoa mass was turned, a process that was carried out every 24 hours until the end of fermentation (day 8). In order to determine the effect of the fermentation process, samples were collected from four bags (replicates) for each clone at 0, 2, 4, 6 and 8 days of fermentation. After this process, the grains of each sample were subjected to the drying process by exposure to the sun until reaching between 7 and 8% moisture [22]. Subsequently, the grains were roasted in a ROASTY model oven, of which the drying was programmed with a development time of 15 minutes with initial and final temperature of 80 and 120 °C, respectively [23]. Finally, the husk of the mass was removed manually, and the beans were ground manually to obtain the cocoa nibs.
2.3. Determination of chemical characteristics of beans from different cocoa clones during the fermentation process
Different bromatological characteristics (8 variables), phenolic compounds (6 variables) and antioxidant activity (2 variables) were determined in each cocoa sample following the methods mentioned in Table 2.
2.4. Statistical analysis
Data were analyzed using linear mixed models (LMM), where clone (CCN-51, EET-8, TSH-565, ICS-1, ICS-60, ICS-95, SCC-61, FSV-41) and fermentation time (0, 2, 4, 6 and 8 days) were the fixed factors and farms as lots were included as random effects. Assumptions of normality and homogeneity of variance were assessed by exploratory analysis of residuals. Differences between factors were analyzed using the LSD Fisher test. Subsequently, a principal component analysis (PCA) was performed to determine the multivariate relationships between the variables evaluated and the cocoa clones during the fermentation process [33]. In order to be able to assess the chemical quality of cocoa, an indicator was generated [23], which was constructed by transforming the values of each of the variables of the three components (Table 2) into scores with values from 0 to 1, using a standardized scoring function, under the criteria: I. More is better, suitable for standardizing the scores of cocoa properties (indicators) in cocoa quality, associated with values close to one (1), II. Less is worse, those properties whose values were close to zero (0). The variables moisture, total acidity, DPPH and FRAP were multiplied by (-1) to invert the value. Subsequently, the variables were summed by components (Table 2) and transformed back to the interval [0, 1]. In this way, an index was generated for each component, these indices were summed and transformed [0, 1] to obtain the indicator of chemical quality of cocoa beans. Subsequently, a Pearson correlation analysis was performed to determine the relationships between the variables characterized[33]; for the analysis, the types of correlations used by Di Rienzo et al. [34] were taken into account: weak correlation (-0.01 to -0.1 or 0.01 to 0.1), medium correlation (-0.11 to -0.50 or 0.11 to 0.50), considerable correlation (-0.51 to -0.75 or 0.51 to 0.75), very strong correlation (-0.76 to -0.90 or 0.76 to 0.90) and perfect correlation (-0.91 to -1 or 0.91 to 1). Finally, a Multiple Factor Analysis (MFA) was performed in which the variables with the highest contribution for each component, their synergies and trade-offs were identified. The LMMs were performed using the lme function of the nlme package, the PCA and the graphical outputs were performed in the packages ade4, ggplot2, factoextra and corrplot in the R language software, version 4.2.0[35], using the RStudio interface [35].
3. Results
3.1. Bromatological characteristics of beans from different cocoa clones during the fermentation process
Bromatological characteristics showed differences (P<0.05) between clones and days of fermentation, but no significant interactions were found (Table 3). Only for fat and total carbohydrates between clones there were no differences, and for sucrase and total carbohydrates at the level of fermentation time. As fermentation time increased, variables such as moisture and acidity increased their values with respect to the initial value; however, ash, fat, and crude protein content, as well as pH values, decreased as a function of fermentation time (Table 3).
3.2. Phenolic content and antioxidant activity of beans from different cocoa clones during the fermentation process
For the variables associated with the content of phenolic compounds and the antioxidant activity of the cocoa beans, there was no interaction between clones and fermentation time, nor was there an effect of fermentation time; only the clone factor was significant for some variables (Table 4). The content of phenolic compounds (total phenols and flavonoids, catechin, epicatechin, theobromine and caffeine) showed statistical differences between clones (P<0.05) (Table 4). The highest concentrations of total phenols were recorded in clones TSH-565 and CCN-51 with values 60.97 and 64.56 mg g-1 respectively, while clones SCC-61 and ICS-60 had the lowest concentrations decreasing by 45.2 and 38.1% with respect to clone CCN-51. Clones CCN-51 and ICS-60 had higher total flavonoid contents, with values of 3.30 and 2.84 and mg g-1 respectively; on the contrary, clones TSH-565 and ICS-1 presented the lowest contents, decreasing their concentration significantly by 61 and 48% with respect to CCN-51 (Table 4).
Catechin and epicatechin had higher contents in clones CCN-51 and ICS-60, with concentrations of 1.90 and 1.95 mg g-1 and 0.48 and 0.49 mg g-1, respectively, while clones TSH-565 and ICS-1 obtained the lowest values, with a reduction of 12.30 and 8.71% for catechin and 10.20 and 6.1% for epicatechin. Clone TSH-565 presented lower theobromine and caffeine contents, decreasing by 6.7 and 7.9% with respect to clone ICS-60 (Table 4). Ferric reducing antioxidant power (FRAP) and 1-1-diphenyl-2-picrylhydrazyl radical (DPPH) decolorization did not show significant differences between clones (Table 4). Their behavior at the clone level ranged from 169.51 to 356.18 µmol g-1 and 350.01 to 168.71 µmol g-1, respectively (Table 4).
Figure 1 shows the association of the variables evaluated both at the clone level (Figure 1a) and fermentation time (Figure 1b) in the biplot plane obtained from the principal components analysis. The principal component analysis (PCA) at the clone level in its first two components explained 70.8% of the variance, component one related attributes such as crude protein and fat content with clones EET-8, TSH-565, ICS-1 and SCC-61. In component two, clones ICS-1 and TSH-565 were related to total phenol content, as were clones ICS-95 and CCN-51 to attributes such as caffeine, theobromine, catechin, epicatechin, total flavonoids, FRAP and DPPH (Figure 1a). As for sugar content (total carbohydrates and sucrose), these were related to clones FSV-41 and ICS-60, while moisture and total acidity did not show any relationship. At the level of fermentation time, the PCA explained 84.6% of the variance, where component one separated the fermentation days in which high contents of fat, crude protein, ash, and pH were higher both at time zero and day two (Figure 1b). In component two the samples were separated by high phenol contents.
When relating the different variables measured in sub-indicators proposed to measure quality in terms of chemical composition, at the clone level we found that FSV-41 presented the highest values for the grain quality index (P<0.05, Figure 2A). Likewise, at the clone level, the bromatological and phenolic compound sub-indicators showed significant differences (P<0.05), with FSV-41 and TSH-565 presenting the highest values, respectively (Figure 2a). At the level of fermentation time, both at the initial moment and on the second day, the highest values were found for the grain quality index and the bromatological sub-indicator (Figure 2b, P<0.05). Finally, the antioxidant activity sub-indicator on days 4 and 6 showed the lowest values for the phenolic compounds index (Figure 2b, P<0.05).
3.3. Correlations between the different bromatological characteristics, phenolic compounds and antioxidant activity of cocoa beans
From the Multiple Factor Analysis performed with the 16 variables, 54.4% of the variance was explained, where those that contributed the highest proportion in principal component one (Figure 3a, P<0.05) were DPPH and FRAP of the phenolic compounds variables (Figure 3b), followed by epicatechin, caffeine, theobromine, flavonoids, catechin and phenols that make up the antioxidant activity variables (Figure 3b). In the principal component two, the variables that contributed the highest proportion (P<0.05) were the majority that make up the bromatological component excluding sucrose and carbohydrates (Figure 3c).
According to the spatial distribution of the variables of each of the components (Antioxidant Activity, Bromatological, Phenolic Compound) in the plane of each of the factors analyzed, clone and fermentation time (Figure 3a and 3b), it is possible to determine the positive relationships as well as the degree of contribution of the variables in each factor. For example, total acidity correlated negatively with pH, ash and fat, and a similar situation occurred between phenols and different variables FRAP, DDPH, among others. When analyzing the contribution of the variables, we found different contributions per factor, as shown by the variables above the red line (Figure 3b and 3c). For example, FRAP and DDPH were the variables with the highest contribution in the clone factor, the opposite behavior in the fermentation time factor. In the case of the variables with the highest contribution in the fermentation time factor, pH and total acidity presented the highest contributions.
4. Discussion
In general, we found a variation in the chemical composition at the clone level and during the fermentation process, in this sense, we highlight how the bromatological variables and those related to antioxidant capacity vary significantly among clones. Likewise, when analyzing the incidence of the fermentation process, differences were found in the bromatological variables and phenolic components. The incidence of these factors on the variables evaluated is discussed below.
4.1. Bromatological characteristics of beans from different cocoa clones during the fermentation process
During the fermentation process, there were no significant variations in some variables; for example, the average moisture content was 2.74%, an adequate value to ensure the storage, transport, and processing of the product[36] as well as the preservation of the product[37]. Regarding the percentage of ash found for clones such as FSV, SCC, CCN and ICS are similar to those reported by Perea et al.[38], with values ranging between 2.7 and 3.5%, while for EET clones it coincides with that stated by Vera et al.[39], who found an average value of 3.2%. In addition, the percentage of ash for all clones is within the stipulations of the AOAC[40] standard, as Perea et al.[38] mention that ash values for cocoa should be less than 4%.
It was verified that as the acidity increased the pH decreased, behavior generated by fermentative reactions under microbiological action on the carbohydrates present in the mucilage [36]. During this process, high levels of acetic acid are generated, which is the predominant acid in the cocoa bean and the cause of their acidification, which is why the less fermented grains have lower acidity and higher pH, because in the absence of an adequate fermentation process, the degradation of carbohydrates is not carried out and these are not released during the drying and roasting process[41].
The fat content for the evaluated clones is adequate, since it presents a value higher than 48% as indicated by NTC 793, and similar to that reported by Perea et al.[38]. Some authors have studied [38,42] the variability of fat content and have concluded that the increase in temperature has a significant effect on the stearoyl-acyl transport protein protein, since it decreases the transfer of glycerides such as oleic, palmitic and stearic acid, reducing the fat percentage since it is mostly constituted by these acids [43]. In addition, fat was reduced during the fermentation process due to the increase in temperature in the first 48 hours that causes a release of fat in the almonds[44], at which time the fatty acids that compose it begin to be lost by leaching [43].
The crude protein percentages of the FSV, SCC, CCN and ICS clones are similar to those reported by Perea et al.[38], with values ranging from 12.9 to 14.2%. Adequate protein range, which is demanded by companies that produce feed mixtures, because raw materials with contents higher than 10% protein have a greater impact on the nutritional quality of the product[45]. As for the content during the fermentation process, there is a variation due to the microbiological activity that develops during this process and the requirement of substrates (proteins and minerals) to carry it out, as well as the dragging effect of these by the draining that occurs during fermentation[36].
The sucrose content (°Brix) in the evaluated clones shows a significant increase, being different from that reported by Loureiro et al.[22] and Barrientos et al.[46], since in their research they obtained values of 1.3 and 0.5%, respectively, for fermented cocoa beans. This increase may be related to the shade of the cocoa plantations evaluated, showing that more shade causes a delay in ear maturation by a few days [47], increasing the mucilage content in the bean and thus increasing the concentration of sugars (carbohydrates)[22]. Although these sugars did not present statistical differences in both the clone factor and the fermentation time, there are reactions that are generated specifically during the roasting process that can affect the content of this variable [48,49], specifically during the roasting process. At this moment, a series of transformations are generated that originate aromatic compounds due to the reduction of carbohydrates and proteins that are concentrated in the cocoa bean[11].
4.2. Phenolic content and antioxidant activity of beans from different cocoa clones during the fermentation process
The total phenol content coincides with that reported by Zapata et al.[50], with values ranging from 21.69 to 38.64 mg g-1 in unfermented grains and from 22.58 to 50.23 mg g-1 in fermented grains, for clones such as CCN-51, ICS-1, ICS-60, ICS-95 and TSH-565. In addition, the variability of the results obtained may be due to the characteristics of each clone, since according to Ramón et al.[23] and Zapata et al.[11] there are internal and external factors that affect the quality and quantity of phenolic compounds in plants, such as genetic diversity, maturity stage, environmental variables, extraction method, processing and storage. On the other hand, the content of total phenols although not was significant presented higher values during the first days, this is mainly due to the formation of polymeric proanthocyanidins during this process, since these are equivalent to 58% of the total phenols in the cocoa bean[50].
The content of total flavonoids is similar to that reported by Zzaman et al.[51] in roasted cocoa beans. In addition, these authors also mention that the concentration of this compound is given by the contents of flavanoiles, flavonols, flavones and anthocyanins present in cocoa, where the most abundant flavonoids are catechin and epicatechin, which is why the total flavonoid content is more abundant in clones CCN-51, EET-8 and ICS-60, since they present the highest concentrations in these variables. The reduction of total flavonoid content in the roasting process can take place thanks to the high oxidation during the method, due to the presence of oxygen, or also to the effect of oxidative enzymes, which can also generate flavonoid losses during the conventional roasting process[51].
The catechin and epicatechin contents found in the cocoa beans in the study were adequate, since they coincide with those reported by Zapata et al.[11] in clones such as CCN-51, ICS-1, ICS-60, ICS-95 and TSH-565, which clarify that these compounds are influenced by clone and roasting. These same authors report that clone CCN-51 has the highest catechin content in roasted grains, which is similar to that found in this work. Epicatechin decreased considerably, this reduction may be due to the increase in temperature in the roasting process, which causes this compound to modify its structure and rearrange or transform into another from hydroxyl groups (H+ and OH-)[52]. Likewise, the catechin content, although it does not present significant differences during the fermentation process, there was a tendency to decrease this content. This is mainly due to the fact that this type of polyphenols are oxidized to quinones and these compounds can complex with amino acids, peptides and proteins and polymerize with other flavonoids to form tannins[11], in addition, the reduction can also be caused by the diffusion of phenolic compounds in the exudates released during this process[50].
Theobromine and caffeine values are similar to those reported by Zapata et al.[11], with ranges from 0.41 to 3.11 mg g-1 and from 0.16 to 1.21 mg g-1 for theobromine and caffeine, respectively, determining the type of cocoa clone as a main source of variation in the content of these alkaloids[53]. However, during the fermentation process, although not was statistic significant the reduction of theobromine and caffeine content is mainly due to the diffusion of these alkaloids with cell liquids[53]; moreover, the content of these compounds is higher during the first days of fermentation, and gradually decreases as fermentation time increases. The above is due to the accumulation through the grain coat during the first days of the fermentation process which increases the content[50,53]. However, theobromine and caffeine content decreases considerably after the roasting process, because during this process there is an increase of substances related to dicetopiperazines, which interact with these alkaloids and reduce their concentration [11].
The antioxidant activity of the free radical DPPH presented values similar to those reported by Zapata et al. [50] in five of the eight clones evaluated (CCN-51, ICS-1, ICS-60, ICS-95 and TSH-565), obtaining values between 224.52 and 350.01 µmol g-1. These results are related by the total flavonoid content, since there is a linear relationship between DPPH values and total flavonoids, because this radical reacts with hydrogen (H) donors and phenolic compounds [50,54,55]. On the other hand, the reducing capacity of the FRAP free radical presented values similar to those found by Perea et al. [56] (2009) in fermented cocoa beans where they report an average value of 361.15 µmol g-1.
4.3. Correlation of bromatological characteristics, phenolic compounds, and antioxidant activity of beans from different cocoa clones during the fermentation process
The percentage of ash correlates positively with the percentage of fat, crude protein and pH, this is due to the fact that these variables present higher values in the first days of fermentation and gradually decrease with time. The decline in protein takes place thanks to the demand for these molecules and minerals, plus the draining that occurs during fermentation [57]. The decline in fat percentage is due to the increase in temperature that occurs during the first 48 hours [44], where losses by leaching of fatty acids begin since this process only takes place from the third day [43]. Likewise, pH decreases due to the production of acetic acid [36], an acid that increases with the passing of days in the fermentation process, which in turn is responsible for the increase in grain acidity, negatively correlated with ash content.
On the other hand, the percentage of ash is reduced due to the exudation produced by the dough during fermentation, so it tends to lose soluble minerals that lead to an imbalance in the concentration of this substance in the kernel [58,59]. When the fermentation process is not carried out properly, there is no demand for proteins, minerals, increased temperature, formation of acetic acid and leaching, so that the protein, fat, pH and ash contents increase and acidity decreases.
Cocoa bean pH and acidity showed a significant negative correlation. Similar data were reported by Jinap and Dimick [60] in cocoa beans from different cocoa bean producing countries, finding a correlation coefficient of -0.94. This relationship may be given by the high concentration of acetic and lactic acid presented by the almonds during the fermentation process [57], since the correlation between acetic and lactic acid with pH was 0.86, while for total acidity it was 0.91, indicating that these acids could be the main responsible for the high acidity and low pH in cocoa almonds in the first days of fermentation[60].
Fat percentage presents a negative correlation with total carbohydrate content; this is probably due to the glyoxylate cycle that takes place during the fermentation process. In this cycle, glucose is generated from fatty acids because most of the metabolic energy required for its development is in the form of triacylglycerols, decreasing fat and increasing sugar content [61]. In addition, the percentage of fat also presents a negative correlation with the content of total phenols, agreeing with Locatelli et al. [62], who report a decrease in lipids in cocoa beans with the presence of phenolic compounds. This is because the concentration of phenols increases with the fermentation process, reducing the fat content, due to the fact that phenols promote the lipoperoxidation process [63], which reduces the oxidative degradation of lipids, causing them to be lost by the diffusion of exudates released during fermentation.
Total phenols show a negative correlation with flavonoids, flavanols (catechin and epicatechin), methylxanthine-type alkaloids (theobromine and caffeine) and fat. This is due to the increase of phenols and the reduction of flavonoids, flavonols, methylxanthines and fat during the fermentation and roasting processes. In fermentation, the formation of polymeric proanthocyanidins responsible for the increase of phenols takes place[50]. In addition, in this process also takes place the oxidation of epicatechin and catechin turning into quinones, which help to reduce the astringency and bitter taste of cocoa[11]. The decrease of these flavanols is also caused by the diffusion of phenolic compounds in the exudates released during this process[50].
The content of total alkaloids is given by the values of methylxanthines (theobromine and caffeine) which causes a decrease in the fermentation process, in addition, the reduction of these compounds is also influenced by the roasting process, where oxidation takes place due to the presence of oxygen or the effect of oxidative enzymes [51]. Likewise, the content of methylxanthines in cocoa beans decreases after the roasting process[11], during this process there is an increase of dicetopiperazine amide (DKP), a compound that interacts with alkaloids, decreasing the content of theobromine and caffeine. On the other hand, the decrease in fat is due to the reduction of fatty acids (oleic, palmitic and stearic acid) due to the increase in temperature in the fermentation process after the third day [64]. However, flavonoids present a positive correlation with antioxidant activity, estimated by FRAP and DPPH technique; this is because the flavonoid content largely determines the scavenging capacity of DPPH radicals, due to the fact that this radical reacts with H donors and phenolic compounds [55]. Likewise, the flavonoids of cocoa beans present high iron (Fe3+) reducing power doing this work by donating hydrogen or singlet oxygen carrying out the redox power[65]. Authors such as Szeto et al. [66] also reported that flavonoids have antioxidant properties through electron donation, which is related to the number of phenolic compounds and the location of the hydroxyl groups and the presence of the galloyl group (gelotannins).
Likewise, total flavonoids present a positive correlation with flavanols (catechin and epicatechin), this correlation is carried out because flavonoids are composed of chalcones, xanthones, flavonols, flavones, flavanones, isofiavonones, anthocyanidins, flavanols and condensed tannins [59] where flavanols comprise 37% of flavonoids. Similarly, the main catechin is epicatechin, which accounts for about 30% of the flavonoid content of the bean [67], thus, increasing flavonoid content increases flavanols. In addition, total flavonoids also show a positive correlation with methylxanthine-type alkaloids (theobromine and caffeine). To date this relationship is not clear [68], and it is known that there is a possible synergistic interaction between flavonoids and methylxanthines, but further studies are needed to corroborate this claim[50]. Also, total flavonoids, catechin, epicatechin, caffeine and catechin present a positive correlation with DPPH and FRAP. This is because the molecules responsible for the antioxidant activity of cocoa are procyanidins, anthocyanins, flavonones and glycosidic flavonols[58], which are mostly present in flavonoids.
The positive correlation between catechin and epicatechin is due to the fact that catechins are comprised of epicatechin, epigallocatechin, gallocatechin, and catechin [69]. On the other hand, the positive correlation between methylxanthines (caffeine and theobromine) and flavanols (catechin or epicatechin) occurs because they are the secondary metabolites most synthesized by the plant [24]. The positive correlation between caffeine and theobromine is associated with the fact that these two alkaloids are the most abundant within the methylxanthine family [70], which comprise between 0.8 and 2% of the total dry weight of the cocoa bean in the case of theobromine, while caffeine in cocoa ranges around 0.2%, varying by clone type and degree of fermentation [71]. In addition, the positive correlation between the antioxidant activity estimated by FRAP and DPPH techniques is due to the fact that both are used to estimate the oxidation capacity of cocoa [65].
5. Conclusions
The fermentation process of the cocoa beans only significantly affected different variables of the bromatological composition (moisture, ash, pH, acidity, fat and crude protein) without affecting the antioxidant activity and phenolic compounds. At the clone level and based on the results obtained with the quality indices in terms of chemical composition (Bromatological, Antioxidant Activity, Phenolic Compound), the FVS-41 clone was the best qualified.
Author Contributions
Conceptualization, P.A.P-H., H.E.H., and J.C.S.; data curation, P.A.P-H., H.E.H., J.C.S. and F.C.; formal analysis, H.E.H., J.C.S. and F.C.; investigation, P.A.P-H., H.E.H., and J.C.S.; methodology, P.A.P-H., H.E.H., V.Y.R-T., N.C., F.C., and J.C.S.; funding acquisition, H.E.H., and J.C.S.; project administration, H.E.H., and J.C.S.; resources, H.E.H., and J.C.S.; supervision, J.C.S.; writing—original draft, P.A.P-H., H.E.H., and J.C.S.; writing—review and editing, P.A.P-H., H.E.H., V.Y.R-T., N.C., F.C., and J.C.S. All authors have read and agreed to the published version of the manuscript.
Data Availability Statement
Data are available from the authors upon request.
Acknowledgments
The authors would like to thank the following collaborators: Ministry of National Education (MEN) and to rural households in the department of Meta that supported the investigation.
Conflicts of Interest
The authors have declared that no competing interests exist.
References
- ICCO. ICCO quarterly bulletin of cocoa statistics. The International Cocoa Organization (ICCO) Cocoa Producing and Cocoa Consuming Countries. Supply & Demand QBCS XLVIII No. 1. 2022.
- ICCO. Quarterly Bulletin of Cocoa Statistcs. Vol. XLIII, Quarterly Bulletin of cocoa statistics. Côte d’Ivoire; 2017. 20–29 p.
- Hernández-Núñez, H.E.; Gutiérrez-Montes, I.; Bernal-Núñez, A.P.; Gutiérrez-García, G.A.; Suárez, J.C.; Casanoves, F.; Flora, C.B. Cacao cultivation as a livelihood strategy: contributions to the well-being of Colombian rural households. Agric. Hum. Values 2021, 39, 201–216. [Google Scholar] [CrossRef]
- Barrera V, Alwang J, Casanova T, Domínguez J, Escudero L, Loor G, et al. La cadena de valor del cacao en y el bienestar de los productores de la provincia de Manabí-Ecuador [Internet]. Iniap. 2019 [cited 2023 Feb 19]. 1–204 p. Available from: http://repositorio.iniap.gob. 4100.
- FEDECACAO. Colombia Cacaotera. Federación Nacional de Cacaoteros [Internet]. 2022 [cited 2023 Feb 19];24. Available from: https://drive.google.
- González, X. La producción de cacao alcanzó cifra récord en 2020 y llegó a las 63.416 toneladas [Internet]. La República. 2021 [cited 2023 Feb 19]. p. Agronegocios. Available from: https://www.google.com/search?q=La+producción+de+cacao+alcanzó+cifra+récord+en+2020+y+llegó+a+las+63.416+toneladas. 1676. [Google Scholar]
- Johanna Gómez González K, Carolina Londoño López V. Analysis of variables for the export of Colombian cocoa to European countries. 2017.
- Kongor, J.E.; Hinneh, M.; Van de Walle, D.; Afoakwa, E.O.; Boeckx, P.; Dewettinck, K. Factors influencing quality variation in cocoa (Theobroma cacao) bean flavour profile — A review. Food Res. Int. 2016, 82, 44–52. [Google Scholar] [CrossRef]
- Hue, C.; Gunata, Z.; Breysse, A.; Davrieux, F.; Boulanger, R.; Sauvage, F. Impact of fermentation on nitrogenous compounds of cocoa beans (Theobroma cacao L.) from various origins. Food Chem. 2016, 192, 958–964. [Google Scholar] [CrossRef]
- Rodriguez-Campos, J.; Escalona-Buendía, H.; Orozco-Avila, I.; Lugo-Cervantes, E.; Jaramillo-Flores, M. Dynamics of volatile and non-volatile compounds in cocoa (Theobroma cacao L.) during fermentation and drying processes using principal components analysis. Food Res. Int. 2011, 44, 250–258. [Google Scholar] [CrossRef]
- Zapata Bustamante S, Tamayo Tenorio A, Alberto Rojano B. Efecto del Tostado Sobre los Metabolitos Secundarios y la Actividad Antioxidante de Clones de Cacao Colombiano. Rev Fac Nac Agron Medellín [Internet]. 2015 Jan 1 [cited 2023 Feb 19];68(1):7497–507. Available from: https://revistas.unal.edu.co/index. 4783.
- Martinez, N. Evaluación de componentes físicos, químicos, organolépticos y de rendimiento de clones universales y regionales de cacao (Theobroma cacao L.) en las zonas productoras de Santander, Arauca y Huila. Magister. Bogotá: Universidad Nacional de Colombia, Cienci. 2016.
- Ramírez González MB, Cely Niño VH, Ramírez SI. Actividad antioxidante de clones de cacao (Theobroma cacao L.) finos y aromáticos cultivados en el estado de Chiapas, México. Perspect EN Nutr HUMANA [Internet]. 2013 [cited 2023 Feb 19];15(1):27–47. Available from: http://www.scielo.org.co/scielo.php? 0124.
- Nazario O, Elizabeth Ordoñez ;, Mandujano Y, Arévalo J. Polifenoles totales, antocianinas, capacidad antioxidante de granos secos y análisis sensorial del licor de cacao (Theobroma cacao L.) criollo y siete clones. Investig y Amaz. 2013;3(1):51–9.
- Sukha, D.A.; Butler, D.R.; Umaharan, P.; Boult, E. The use of an optimised organoleptic assessment protocol to describe and quantify different flavour attributes of cocoa liquors made from Ghana and Trinitario beans. Eur. Food Res. Technol. 2007, 226, 405–413. [Google Scholar] [CrossRef]
- Quezada, Luis. Quevedo, José. García R. Determinación del efecto del grado de madurez de las mazorcas en la producción y la calidad sensorial de (Theobroma cacao L.). Rev Científica Agroecosistemas [Internet]. 2017 Dec 11 [cited 2023 Feb 19];5(1):36–46. Available from: https://aes.ucf.edu.cu/index.
- Roos, W. ÍNDICE MIP DE ALGUNOS CULTIVOS TROPICALES [Internet]. EDICIONES. 2015 [cited 2023 Feb 19]. 119 p. Available from: https://www.cabi.org/wp-content/uploads/Rogg-2000b-IPM-in-tropical-crops.
- Boza, E.J.; Motamayor, J.C.; Amores, F.M.; Cedeño-Amador, S.; Tondo, C.L.; Livingstone, D.S.; Schnell, R.J.; Gutiérrez, O.A. Genetic Characterization of the Cacao Cultivar CCN 51: Its Impact and Significance on Global Cacao Improvement and Production. J. Am. Soc. Hortic. Sci. 2014, 139, 219–229. [Google Scholar] [CrossRef]
- Bekele, F.L.; Bekele, I.; Butler, D.R.; Bidaisee, G.G. Patterns of Morphological Variation in a Sample of Cacao (Theobroma Cacao L.) Germplasm from the International Cocoa Genebank, Trinidad. Genet. Resour. Crop. Evol. 2006, 53, 933–948. [Google Scholar] [CrossRef]
- Johnson, E.S.; Bekele, F.L.; Brown, S.J.; Song, Q.; Zhang, D.; Meinhardt, L.W.; Schnell, R.J. Population Structure and Genetic Diversity of the Trinitario Cacao (Theobroma cacao L.) from Trinidad and Tobago. Crop. Sci. 2009, 49, 564–572. [Google Scholar] [CrossRef]
- Hernández-Núñez HE, Gutiérrez-Montes I, Sánchez-Acosta JR, Rodríguez-Suárez L, Gutiérrez-García GA, Suárez-Salazar JC, et al. Agronomic conditions of cacao cultivation: its relationship with the capitals endowment of Colombian rural households. Agrofor Syst. 2020.
- Loureiro, G.A.H.A.; Araujo, Q.R.; Sodré, G.A.; Valle, R.R.; Souza, J.O.; Ramos, E.M.L.S.; Comerford, N.B.; Grierson, P.F. Cacao quality: Highlighting selected attributes. Food Rev. Int. 2016, 33, 382–405. [Google Scholar] [CrossRef]
- Ramón, V.; Hernández, H.E.; Polania, P.; Suárez, J.C. Spatial Distribution of Cocoa Quality: Relationship between Physicochemical, Functional and Sensory Attributes of Clones from Southern Colombia. Agronomy 2022, 13, 15. [Google Scholar] [CrossRef]
- Samaniego, I.; Espín, S.; Quiroz, J.; Ortiz, B.; Carrillo, W.; García-Viguera, C.; Mena, P. Effect of the growing area on the methylxanthines and flavan-3-ols content in cocoa beans from Ecuador. J. Food Compos. Anal. 2020, 88, 103448. [Google Scholar] [CrossRef]
- AOAC. Official methods of analysis of AOAC International, 19th ed. (970.22). Association of Official Analytical Chemists International. 2012.
- García E, Fernández I. Determinación de proteínas de un alimento por el método Kjeldahl. Valoración con un ácido fuerte. [Internet]. ETSIAMN. Universidad Politécnica de Valencia. Universitat Politècnica de València; 2020 Jun [cited 2023 Feb 19]. Available from: https://riunet.upv. 1025.
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Galeano PG, Paladines MB. Actividad antioxidante de extractos de granos de copoazú (Theobroma grandiflorum). Vitae [Internet]. 2012 [cited 2023 Feb 19];19(1):436–8. Available from: https://www.redalyc.org/pdf/1698/169823914137.
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Hatzidimitriou, E.; Nenadis, N.; Tsimidou, M.Z. Changes in the catechin and epicatechin content of grape seeds on storage under different water activity (aw) conditions. Food Chem. 2007, 105, 1504–1511. [Google Scholar] [CrossRef]
- Brunetto M del R, Gutiérrez L, Delgado Y, Gallignani M, Zambrano A, Gómez Á, et al. Determination of theobromine, theophylline and caffeine in cocoa samples by a high-performance liquid chromatographic method with on-line sample cleanup in a switching-column system. Food Chem. 2007 Jan 1;100(2):459–67.
- Velásquez H, Galeano P. Evaluación fitoquímica y de actividad antioxidante de los rizomas de tres especies del género Cyperus. Momentos Cienc. 2012;9(1):15–21.
- Balzarini M, Di Rienzo J, Tablada M, Gonzalez L, Bruno C, Córdoba M, et al. Estadística y biometrías. Ilustraciones del uso de InfoStat en problemas de agronomía. Editorial Brujas, Córdoba, Argentina. Segunda Edición. 2012. 380 p.
- Rienzo D, Alejandro J, Alicia L, Margot E, Pilar M. Estadistica para las ciencias agropecuarias. Potencia. 2005. 1–329 p.
- R Development Core Team. R: The R Project for Statistical Computing. In: Foundation for Statistical Computing, V., Austria. ISBN 3-900051-07-0 (Ed.). 2023; Available from: https://www.r-project.org/.
- Del M, Amaíz CL, Gutiérrez R, Pérez E, Álvarez C. Efecto del tostado sobre las propiedades físicas, fisicoquímicas, composición proximal y perfil de ácidos grasos de la manteca de granos de cacao del estado Miranda, Venezuela Effect of roasting process on physical and physicochemical properties, proximat [Internet]. Vol. 12, Revista Científica UDO Agrícola. Universidad de Oriente; 2012 [cited 2023 Feb 19]. Available from: https://dialnet.unirioja.es/servlet/articulo? 4688.
- lvarez C, Pérez E, Lares MC. Caracterización física y química de almendras de cacao fermentadas, secas y tostadas cultivadas en la región de Cuyagua, estado Aragua. Agron Trop [Internet]. 2007 [cited 2022 Oct 18];57(4):249–56. Available from: http://ve.scielo.org/scielo.php? 0002.
- Perea JA, Ramirez OL, Villamizar AR. Caracterización fisicoquimica de materiales regionales de cacao colombiano. Biotecnol en el Sect Agropecu y Agroindustrial [Internet]. 2011 [cited 2022 Oct 18];9(1):35–42. Available from: http://www.scielo.org.co/scielo.php? 1692.
- Vera Chang JF, Vallejo Torres C, Párraga Morán DE, Macías Véliz J, Ramos Remache R, Morales Rodríguez W. Atributos físicos-químicos y sensoriales de las almendras de quince clones de cacao nacional (Theobroma cacao L.) en el Ecuador. Cienc y Tecnol [Internet]. 2015 Mar 19 [cited 2022 Oct 18];7(2):21–34. Available from: https://revistas.uteq.edu.ec/index.
- AOAC. Ash (acid-insoluble) of cacao products. AOAC 975.12. [Internet]. 2000 [cited 2023 Feb 20]. Available from: http://www.aoacofficialmethod.org/index.php?
- lvarez R, Portillo E, Portillo A, Villasmil R. Evaluación de las propiedades sensoriales del licor de cacao (theobroma cacao l.) obtenido en forma artesanal e industrial. Rev Agrollanía [Internet]. 2018 [cited 2022 Oct 18];15:1690–8066. 8: Available from: http://localhost, 8080.
- Mustiga, G.M.; Morrissey, J.; Stack, J.C.; DuVal, A.; Royaert, S.; Jansen, J.; Bizzotto, C.; Villela-Dias, C.; Mei, L.; Cahoon, E.B.; et al. Identification of Climate and Genetic Factors That Control Fat Content and Fatty Acid Composition of Theobroma cacao L. Beans. Front. Plant Sci. 2019, 10, 1159. [Google Scholar] [CrossRef]
- Del Rosario Castro M, Hernández JA, Marcilla S, Córdova JS, Solari FA, Chire GC. Efecto del contenido de grasa en la concentración de polifenoles y capacidad antioxidante de Theobroma cacao L. “Cacao.” Cienc Invest [Internet]. 2017 [cited 2023 Feb 19];19(1):19–23. Available from: https://www.researchgate. 3081.
- Servent, A.; Boulanger, R.; Davrieux, F.; Pinot, M.-N.; Tardan, E.; Forestier-Chiron, N.; Hue, C. Assessment of cocoa (Theobroma cacao L.) butter content and composition throughout fermentations. Food Res. Int. 2018, 107, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Andrade JA, Rivera-García J, Chire-Fajardo GC, Ureña-Peralta MO. Propiedades físicas y químicas de cultivares de cacao (Theobroma cacao L.) de Ecuador y Perú. Enfoque UTE [Internet]. 2019 [cited 2023 Feb 19];10(4):1–12. Available from: http://scielo.senescyt.gob.ec/scielo.php? 1390.
- Barrientos, L.D.P.; Oquendo, J.D.T.; Gil Garzón, M.A.; Álvarez, O.L.M. Effect of the solar drying process on the sensory and chemical quality of cocoa (Theobroma cacao L.) cultivated in Antioquia, Colombia. Food Res. Int. 2018, 115, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Imru NO, Wogderess MD, Gidada T V. A study of the effects of shade on growth, production and quality of coffee (COFFEA ARABICA) in Ethiopia. Int J Agric Sci [Internet]. 2015 [cited 2023 Feb 19];5(5):748–52. Available from: http://www.academicjournals.
- Afoakwa EO, Ofosu-Ansah E, Takrama JF, Budu AS, Mensah-Brown H. Changes in chemical quality of cocoa butter during roasting of pulp pre-conditioned and fermented cocoa (Theobroma cacao) beans. Int Food Res J [Internet]. 2014 [cited 2023 Feb 19];21(6):2221–7. Available from: https://www.researchgate. 2763.
- Rohan, T.A.; Stewart, T. The Precursors of Chocolate Aroma: Changes in the Sugars During the Roasting of Cocoa Beans. J. Food Sci. 1966, 31, 206–209. [Google Scholar] [CrossRef]
- Zapata Bustamante S, Tamayo Tenorio A, Alberto Rojano B. Efecto de la fermentación sobre la actividad antioxidante de diferentes clones de cacao Colombiano. Rev Cuba Plantas Med [Internet]. 2013 [cited 2023 Feb 20];18(3):391–404. Available from: http://scielo.sld.cu/scielo.php? 1028.
- Zzaman, W.; Bhat, R.; Abedin, Z.; Yang, T.A. Comparison between Superheated Steam and Convectional Roasting on Changes in the Phenolic Compound and Antioxidant Activity of Cocoa Beans. Food Sci. Technol. Res. 2013, 19, 949–956. [Google Scholar] [CrossRef]
- Lujano E, Manganiello L, Contento A, Rios A. Identification and quantification of (+) - Catechins and Procyanidins in Cocoa from Ocumare de la Costa, Venezuela. Ing Uc. 2019;26(2):192–201.
- Alvarez, L.C.; Alvarez, N.C.; Garcia, P.G.; Salazar, J.C.S. Effect of fermentation time on phenolic content and antioxidant potential in Cupuassu (Theobroma grandiflorum (Willd. ex Spreng.) K.Schum.) beans. 2017, 66, 473–479. [Google Scholar] [CrossRef]
- Kuskoski EM, Asuero AG, Troncoso AM, Mancini-Filho J, Fett R. Aplicación de diversos métodos químicos para determinar actividad antioxidante en pulpa de frutos. Ciência e Tecnol Aliment [Internet]. 2005 Dec [cited 2023 Feb 19];25(4):726–32. Available from: http://www.scielo.br/j/cta/a/B58T9S5zLLxjBL5PVzZXHCF/abstract/?
- Roginsky, V.; Lissi, E.A. Review of methods to determine chain-breaking antioxidant activity in food. Food Chem. 2005, 92, 235–254. [Google Scholar] [CrossRef]
- Perea J, Cadena T, Herrera J. El cacao y sus productos como fuente de antioxidantes: Efecto del procesamiento. Salud UIS [Internet]. 2009 [cited 2023 Feb 19];41:128–34. Available from: http://www.scielo.org.co/scielo.php? 0121.
- Apriyanto, M. Changes in Chemical Properties of Dreid Cocoa (Theobroma cacao) Beans during Fermentation. Int. J. Fermented Foods 2016, 5, 11. [Google Scholar] [CrossRef]
- Acevedo L, Mejía D, Acosta E, Valencia W, Penagos L. Efecto de la temperatura del conchado sobre los polifenoles en un chocolate semi-amargo. Aliment Hoy [Internet]. 2017 [cited 2023 Feb 19];25(41):31–50. Available from: https://alimentoshoy.acta.org.co/index.
- Quiñones M, Miguel M, Aleixandre A. Los polifenoles, compuestos de origen natural con efectos saludables sobre el sistema cardiovascular. Nutr Hosp organo Of la Soc Espa??ola Nutr Parenter y Enter. 2012;27(1):76–89.
- Jinap, S.; Dimick, P.S. Acidic Characteristics of Fermented and Dried Cocoa Beans from Different Countries of Origin. J. Food Sci. 1990, 55, 547–550. [Google Scholar] [CrossRef]
- Afoakwa, E.O.; Quao, J.; Takrama, J.; Budu, A.S.; Saalia, F.K. Chemical composition and physical quality characteristics of Ghanaian cocoa beans as affected by pulp pre-conditioning and fermentation. J. Food Sci. Technol. 2011, 50, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- Locatelli, M.; Travaglia, F.; Giovannelli, L.; Coïsson, J.D.; Bordiga, M.; Pattarino, F.; Arlorio, M. Clovamide and phenolics from cocoa beans (Theobroma cacao L.) inhibit lipid peroxidation in liposomal systems. Food Res. Int. 2013, 50, 129–134. [Google Scholar] [CrossRef]
- Ordoñez ES, Leon-Arevalo A, Rivera-Rojas H, Vargas E. Cuantificación de polifenoles totales y capacidad antioxidante en cáscara y semilla de cacao (Theobroma cacao L.), tuna (Opuntia ficus indica Mill), uva (Vitis Vinífera) y uvilla (Pourouma cecropiifolia). Sci Agropecu [Internet]. 2019 [cited 2023 Feb 19];10(2):175–83. Available from: http://www.scielo.org.pe/scielo.php? 2077.
- Ramos-Escudero, F.; Casimiro-Gonzales, S.; Fernández-Prior. ; Chávez, K.C.; Gómez-Mendoza, J.; de la Fuente-Carmelino, L.; Muñoz, A.M. Colour, fatty acids, bioactive compounds, and total antioxidant capacity in commercial cocoa beans (Theobroma cacao L.). LWT 2021, 147, 111629. [Google Scholar] [CrossRef]
- Othman, A.; Ismail, A.; Ghani, N.A.; Adenan, I. Antioxidant capacity and phenolic content of cocoa beans. Food Chem. 2007, 100, 1523–1530. [Google Scholar] [CrossRef]
- Szeto YT, Tomlinson B, Benzie IFF. Total antioxidant and ascorbic acid content of fresh fruits and vegetables: implications for dietary planning and food preservation. Br J Nutr [Internet]. 2002 Jan [cited 2023 Feb 19];87(1):55–9. Available from: https://www.cambridge. 4579.
- Sotelo, C. L, Alvis B. A, Arrázola P. G. Evaluación de epicatequina, teobromina y cafeína en cáscaras de cacao (Theobroma cacao L.), determinación de su capacidad antioxidante. Rev Colomb Ciencias Hortícolas [Internet]. 2015 Aug 12 [cited 2023 Feb 19];9(1):124. Available from: http://www.scielo.org.co/scielo.php? 2011. [Google Scholar]
- Belščak A, Komes D, Horžić D, Ganić KK, Karlović D. Comparative study of commercially available cocoa products in terms of their bioactive composition. Food Res Int. 2009 Jun 1;42(5–6):707–16.
- Quizhpe PAC. Efecto inhibitorio de la pulpa que recubre las semillas del cacao (Theobroma Cacao) a diferentes concentraciones sobre la cepa de Streptococcus mutans: Estudio in … [Internet]. Quito: UCE; 2018 [cited 2023 Feb 19]. Available from: http://www.dspace.uce.edu. 2500.
- Peláez PP, Bardón I, Camasca P. Methylxanthine and catechin content of fresh and fermented cocoa beans, dried cocoa beans, and cocoa liquor. Sci Agropecu [Internet]. 2016 Dec 31 [cited 2023 Feb 19];7(4):355–65. Available from: http://www.scielo.org.pe/scielo.php? 2077.
- Carrillo, L.C.; Londoño-Londoño, J.; Gil, A. Comparison of polyphenol, methylxanthines and antioxidant activity in Theobroma cacao beans from different cocoa-growing areas in Colombia. Food Res. Int. 2014, 60, 273–280. [Google Scholar] [CrossRef]
Figure 1.
Biplot result of principal component analysis between cocoa bean chemical characteristics at the clone level (Figure 1a) and fermentation time (Figure 1b).
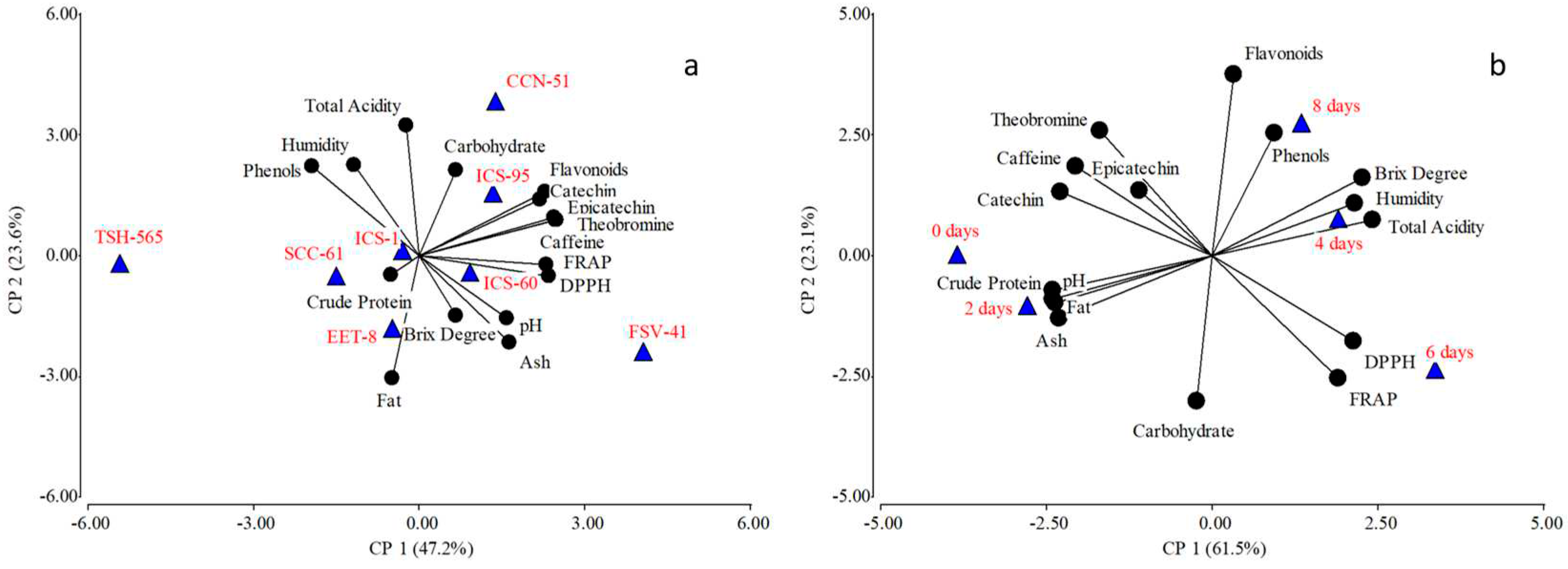
Figure 2.
Indicator and sub-indicators representing cocoa bean quality at both A. clone level and B. fermentation time. Values represent the mean and standard error (n=4). a, b, c: mean statistical differences between each of the factors for clone and days of fermentation.
Figure 2.
Indicator and sub-indicators representing cocoa bean quality at both A. clone level and B. fermentation time. Values represent the mean and standard error (n=4). a, b, c: mean statistical differences between each of the factors for clone and days of fermentation.
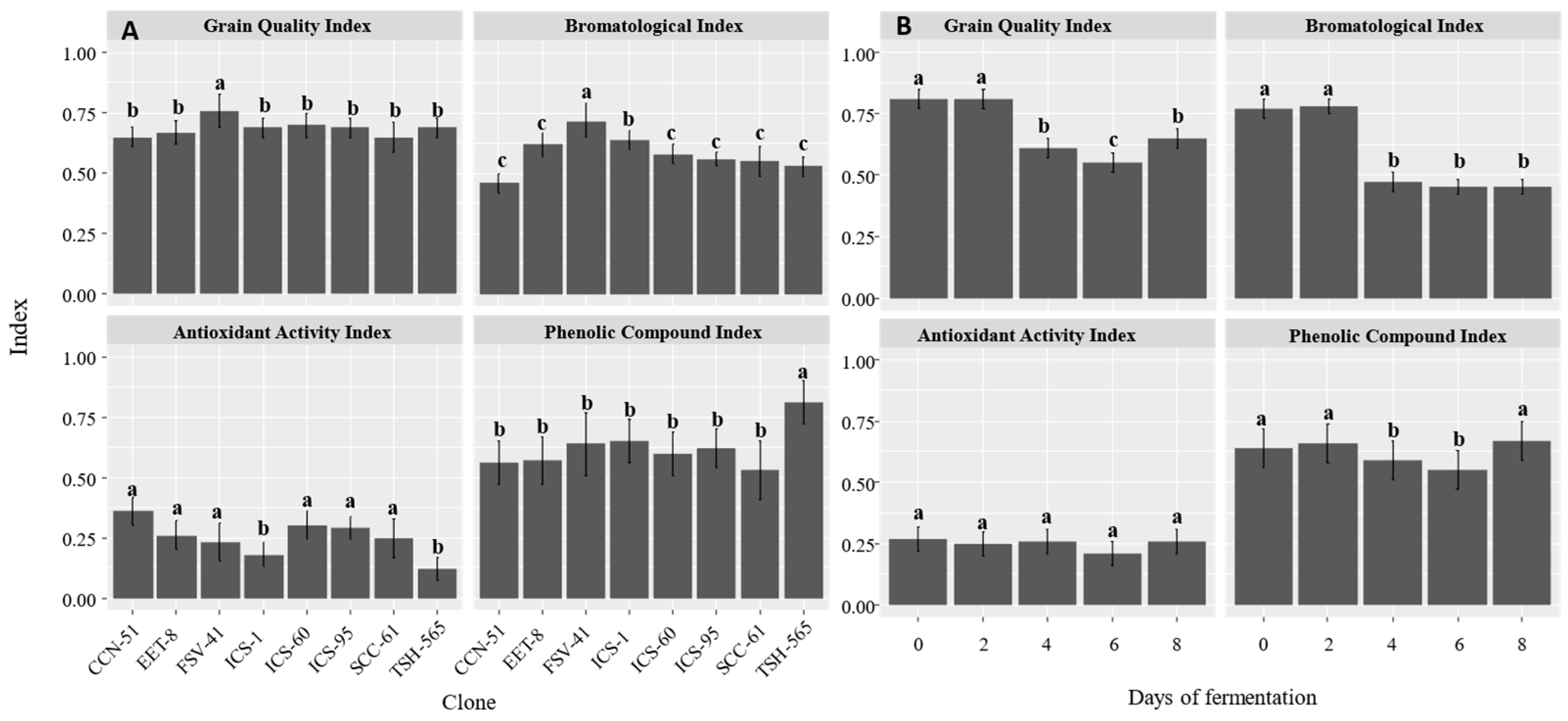
Figure 3.
Most important variables in each sub-indicator that impact the cocoa quality index. a: Contribution of the variables for each sub-indicator in the principal component (PC) 1 and 2. From red to blue means from highest to lowest contribution. b, c: distribution of the variables in a biplot plane obtained through a principal component analysis, obtained through a multiple factor analysis. The variables above the dotted line presented the highest contribution. AA: Antioxidant Activity; Brom: Bromatological; PC: Phenolic Compound.
Figure 3.
Most important variables in each sub-indicator that impact the cocoa quality index. a: Contribution of the variables for each sub-indicator in the principal component (PC) 1 and 2. From red to blue means from highest to lowest contribution. b, c: distribution of the variables in a biplot plane obtained through a principal component analysis, obtained through a multiple factor analysis. The variables above the dotted line presented the highest contribution. AA: Antioxidant Activity; Brom: Bromatological; PC: Phenolic Compound.
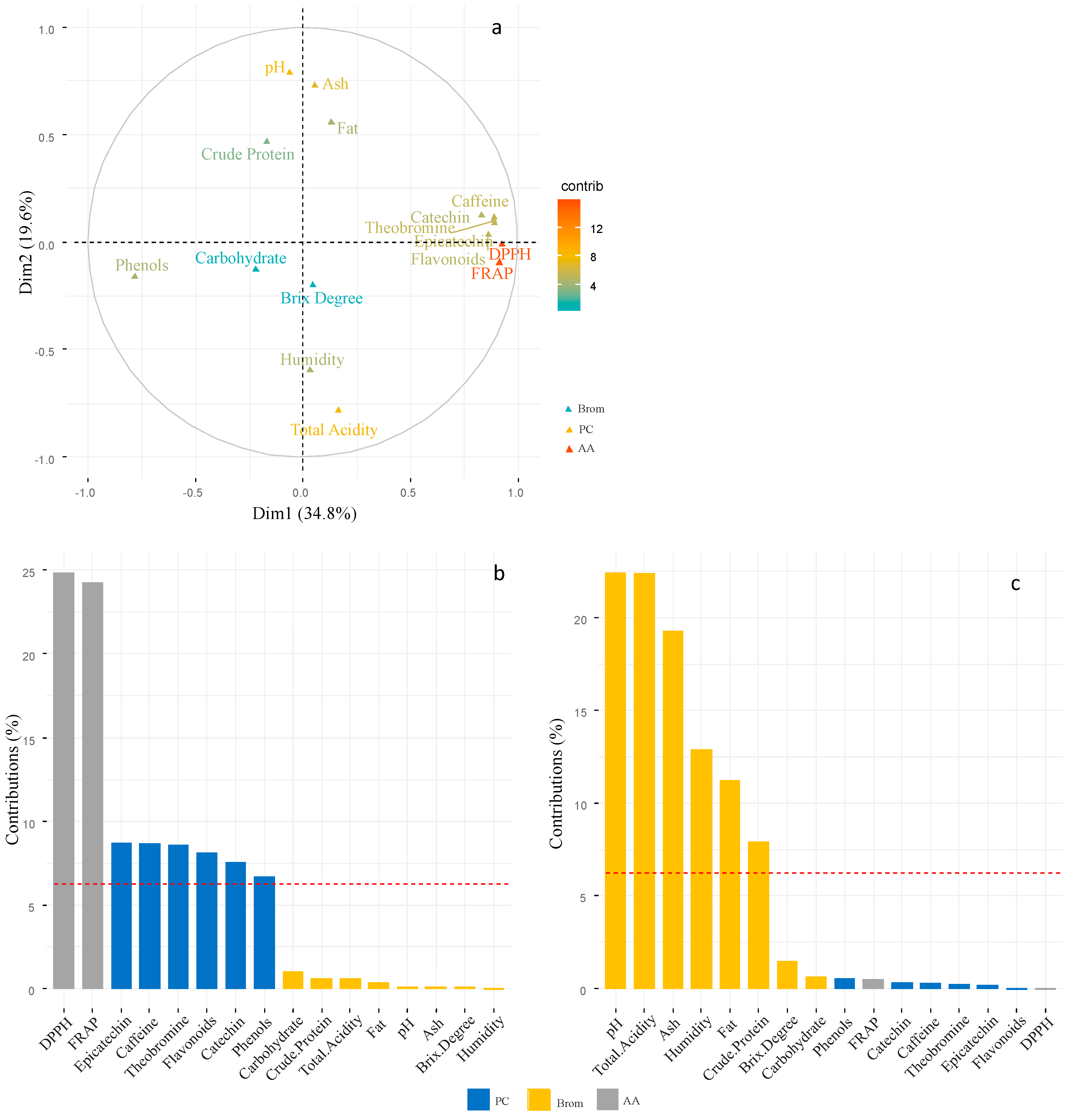

Table 1.
List of cocoa clones, their identification and origin.
| Introduced commercial | ||
|---|---|---|
| Nomenclature | Identification and origin | Characteristics of interest |
| CCN-51 | Castro Naranjal (Ecuador) | Commercially grown. High yield. Resistance to Monilia[18]. |
| EET-8 | United Fruit Company (Costa Rica) | Commercially grown. Good grain index[19]. |
| TSH-565 | Trinidad Selection Hybrid (Trinidad) | Resistance to Monilliphthora perniciosa, high productivity[20]. |
| ICS-1 | Imperial College Selection (Trinidad, Nicaragua y Venezuela) | Present in commercial crops in several countries. Good grain and cob index[19]. |
| ICS-60 | Imperial College Selection (Trinidad, Nicaragua y Venezuela) | Present in commercial crops in several countries. Good grain and cob index [19]. |
| ICS-95 | Imperial College Selection (Trinidad, Nicaragua y Venezuela) | Present in commercial crops in several countries. Good grain and cob index [19]. |
| Regional | ||
| SCC-61 | Selección Colombia Corpoica (Santander), Hibrido trinitario | High grain index[12]. |
| FSV-41 | Fedecacao San Vicente (Santander), Híbrido trinitario | High grain rate, yield and quality[12]. |
Table 2.
Bromatological variables, phenolic compounds and antioxidant activity characterized.
| Component | Variable | Unit | Method |
|---|---|---|---|
| Bromatological | pH | Potentiometric [24] | |
| Moisture | % | Gravimetric [24] | |
| Ash | % | Incineration [25] | |
| Acidity | % | Titling [24] | |
| Fat | % | Soxhlet [24] | |
| Crude protein | % | Kjeldahl [26] | |
| Sucrose | °Brix | Refractometry [22] | |
| Total carbohydrates | mg g-1 | Phenol-Sulfuric [27] | |
| Phenolic compounds | Total phenols | mg g-1 | Folin-Ciocalteu colorimetry [28] |
| Total flavonoids | mg g-1 | Aluminum chloride [29] | |
| Catechin | mg g-1 | HPLC [30] | |
| Epicatechin | mg g-1 | HPLC [30] | |
| Theobromine | mg g-1 | HPLC [31] | |
| Caffeine | mg g-1 | HPLC [31] | |
| Antioxidant activity | DPPH | (µmol g-1) | Colorimetric [32] |
| FRAP | (µmol g-1) | Colorimetric [32] |
Table 3.
Bromatological characteristics evaluated in cocoa clones at different fermentation times (Mean±Standard error).
Table 3.
Bromatological characteristics evaluated in cocoa clones at different fermentation times (Mean±Standard error).
| Factor | Level | Moisture (%) | Ash (%) | pH | Acidity (%) | Fat (%) | Crude Protein (%) | Sucrose °Brix |
Total Carbohydrates (mg g-1) |
|---|---|---|---|---|---|---|---|---|---|
| Clone | CCN 51 | 2.98±0.16a | 3.03±0.09c | 5.14±0.09 | 0.56±0.03ab | 53.25±2.56 | 12.07±0.18c | 2.78±0.54b | 1.63±0.12 |
| EET 8 | 2.54±0.18abc | 3.17±0.10bc | 5.30±0.10 | 0.46±0.03c | 63.61±3.10 | 12.40±0.21bc | 3.42±0.61ab | 1.35±0.13 | |
| FSV 41 | 1.98±0.26c | 3.53±0.14a | 5.32±0.13 | 0.49±0.05bc | 60.66±4.99 | 12.60±0.28abc | 4.43±0.81ab | 1.55±0.18 | |
| ICS 1 | 2.38±0.15bc | 3.35±0.09ab | 5.16±0.09 | 0.54±0.03abc | 58.26±2.56 | 12.85±0.18a | 3.51±0.52ab | 1.62±0.11 | |
| ICS 60 | 2.65±0.18ab | 3.21±0.10bc | 5.17±0.10 | 0.49±0.03bc | 64.45±2.98 | 12.19±0.20c | 3.23±0.59b | 1.55±0.13 | |
| ICS 95 | 2.90±0.13a | 3.37±0.07ab | 5.10±0.08 | 0.59±0.02a | 55.66±2.07 | 12.50±0.16bc | 4.57±0.46a | 1.55±0.10 | |
| SCC 61 | 3.01±0.23a | 3.31±0.13abc | 5.35±0.12 | 0.52±0.04abc | 59.83±3.72 | 12.82±0.26ab | 3.97±0.76ab | 1.22±0.17 | |
| TSH 565 | 2.61±0.15ab | 3.15±0.08c | 5.11±0.09 | 0.55±0.03ab | 60.80±2.50 | 12.12±0.18c | 3.91±0.52ab | 1.53±0.11 | |
| P value | 0.0031 | 0.0017 | 0.1937 | 0.0420 | 0.0624 | 0.0009 | 0.0324 | 0.2789 | |
| Fermentation time(days) | 0 | 2.52±0.15ab | 3.47±0.08a | 5.49±0.08a | 0.42±0.03b | 65.44±2.38a | 12.90±0.17a | 3.29±0.51 | 1.49±0.11 |
| 2 | 2.21±0.14b | 3.58±0.08a | 5.51±0.08a | 0.43±0.03b | 62.37±2.10ab | 12.87±0.16a | 3.28±0.48 | 1.52±0.11 | |
| 4 | 2.75±0.14a | 3.15±0.08b | 4.99±0.08b | 0.61±0.03a | 56.69±2.14bc | 12.21±0.16b | 3.97±0.49 | 1.47±0.11 | |
| 6 | 2.86±0.14a | 3.07±0.08b | 4.98±0.08b | 0.60±0.03a | 55.70±2.10c | 12.14±0.16b | 3.98±0.48 | 1.56±0.11 | |
| 8 | 2.82±0.14a | 3.05±0.08b | 5.05±0.08b | 0.58±0.03a | 54.96±2.10c | 12.12±0.16b | 4.12±0.48 | 1.46±0.11 | |
| P value | 0.0004 | <0.0001 | <0.0001 | <0.0001 | 0.0028 | <0.0001 | 0.2568 | 0.8975 |
a, b, c: Different letters indicate significant differences between cocoa clones and fermentation time (LSD Fisher test, p<0.05). Values represent the mean and standard error (n=4).
Table 4.
Phenolic content and antioxidant activity evaluated in cocoa clones at different fermentation times.
Table 4.
Phenolic content and antioxidant activity evaluated in cocoa clones at different fermentation times.
| Factor | Level | Total Phenols (mg g-1) | Total Flavonoids (mg g-1) |
Catechin (mg g-1) |
Epicatechin (mg g-1) | Theobromine (mg g-1) | Caffeine (mg g-1) | DPPH (µmol g-1) |
FRAP (µmol g-1) |
|---|---|---|---|---|---|---|---|---|---|
| Clone | CCN-51 | 64.56±11.74a | 3.30±0.57a | 1.95±0.06a | 0.49±0.01a | 0.30±0.01a | 0.37±0.01a | 325.55±33.30 | 331.00±34.29 |
| EET-8 | 42.71±7.63abc | 2.73±0.54a | 1.90±0.07a | 0.48±0.01ab | 0.29±0.01a | 0.37±0.01a | 251.47±40.26 | 244.72±41.45 | |
| FSV-41 | 46.46±19.37abc | 2.25±0.78abc | 1.82±0.09abc | 0.47±0.02abc | 0.29±0.01ab | 0.37±0.01ab | 168.71±64.91 | 169.51±66,84 | |
| ICS-1 | 47.55±7.30abc | 1.72±0.44bc | 1.78±0.05bc | 0.46±0.01bc | 0.29±0.01ab | 0.36±0.01ab | 245.23±33.30 | 248.29±34.29 | |
| ICS-60 | 39.93±7.54bc | 2.84±0.53a | 1.93±0.07a | 0.48±0.01a | 0.30±0.01a | 0.38±0.01a | 224.42±38.79 | 226.87±39.94 | |
| ICS-95 | 52.23±7.73ab | 2.55±0.43ab | 1.88±0.05ab | 0.48±0.01ab | 0.30±0.01a | 0.37±0.01a | 256.22±26.95 | 259.61±27.75 | |
| SCC-61 | 35.36±7.66c | 2.41±0.69abc | 1.87±0.08abc | 0.48±0.02ab | 0.29±0.01ab | 0.37±0.01ab | 275.00±48.38 | 278.95±49.82 | |
| TSH-565 | 60.97±8.27a | 1.29±0.42c | 1.71±0.05c | 0.44±0.01c | 0.28±0.01b | 0.35±0.01b | 350.01±32.56 | 356.18±33.42 | |
| P value | 0.0238 | 0.0063 | 0.0022 | 0.0213 | 0.0195 | 0.0117 | 0.0678 | 0.0678 | |
| Fermentation time (days) | 0 | 39.32±8.77 | 2.64±0.46 | 1.90±0.06 | 0.48±0.01 | 0.30±0.01 | 0.37±0.01 | 256.11±32.88 | 259.50±30.43 |
| 2 | 48.91±8.62 | 2.44±0.44 | 1.88±0.05 | 0.47±0.01 | 0.29±0.01 | 0.37±0.01 | 272.95±29.00 | 276.84±29.86 | |
| 4 | 52.91±8.64 | 2.36±0.46 | 1.85±0.05 | 0.47±0.01 | 0.29±0.01 | 0.37±0.01 | 291.95±29.55 | 296.40±30.43 | |
| 6 | 50.50±8.62 | 2.18±0.41 | 1.82±0.05 | 0.47±0.01 | 0.29±0.01 | 0.36±0.01 | 260.08±29.00 | 263.59±29.86 | |
| 8 | 51.97±8.62 | 2.31±0.43 | 1.83±0.05 | 0.47±0.01 | 0.29±0.01 | 0.37±0.01 | 283.11±29.00 | 287.30±29.86 | |
| P value | 0.5806 | 0.8314 | 0.3330 | 0.8945 | 0.2962 | 0.2282 | 0.9101 | 0.9101 |
a, b, c: Different letters indicate significant differences between cocoa clones and fermentation time (LSD Fisher test, p<0.05). Values represent the mean and standard error (n=4).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



