
Submitted:
25 September 2023
Posted:
27 September 2023
You are already at the latest version
Abstract
Candidatus Accumulibacter belongs to phosphate-accumulating organisms (PAO) which exhibit cyclic metabolism and are capable of intracellular polyphosphate accumulation and their hydrolysis under feast-famine anaerobic-aerobic cycling. In consortia of activated sludge microorganisms, these bacteria are responsible for enhanced biological phosphorus removal (EBPR). The spectrum of the substrates used by Ca. Accumulibacter remains insufficiently studied. It was investigated by measuring the oxygen uptake rates (OUR) of Ca. Accumulibacter-enriched culture supplemented with 17 different organic substrates. The highest oxygen uptake rates values were observed in the presence of tryptone, volatile fatty acids (acetate, propionate, and butyrate), succinate, pyruvate, and amino acids (aspartate and glutamate). Phosphate dynamics in the medium under shifts from anaerobic to aerobic cultivation in batch experiments was studied for these compounds (except for tryptone). All tested substrates were shown to cause phosphate cycling (release in the anaerobic phase and uptake in the aerobic one), with OURs for the substrates correlating with the amount of phosphates consumed during the aerobic phase. It was concluded that OUR may be used as an indicator of the monosubstrates used by Ca. Accumulibacter in the anaerobic/aerobic cycle. The possible pathways for substrate transport and metabolism by Ca. Accumulibacter are discussed using the stoichiometric data and the results of metagenomic analysis.
Keywords:
phosphate-accumulating microorganisms
; EBPR
; Ca. Accumulibacter
; oxygen uptake
1. Introduction
Phosphate-accumulating organisms (PAO) are aerobic or facultatively anaerobic microorganisms removing phosphorus from wastewater due to their ability to accumulate phosphorus inside their cells in amounts significantly exceeding those required for the metabolism of regular bacteria [1,2,3]. According to the current concepts [4,5,6], under anaerobic conditions PAO rapidly consume readily biodegradable organic compounds and store them as intracellular polymers, using the energy derived from hydrolysis of intracellular polyphosphates. Under aerobic conditions or in the presence of an alternative electron acceptor (nitrate or nitrite), they uptake orthophosphate and synthesize intracellular polyphosphates, using the intracellular carbon and energy sources accumulated under anaerobic conditions. Cell biomass growth occurs during the aerobic phase, while during the anaerobic phase the biosynthetic activity of these bacteria is insignificant (apart from accumulation of intracellular storage compounds) [7]. The metabolic properties of PAO are used in the modern technologies for enhanced biological phosphorus removal (EBPR), which use the cyclic shifts in conditions favoring the growth of these microorganisms. The main PAO used in EBPR belong to the genera Accumulibacter, Tetrasphaera, and Dechloromonas [6]. Glycogen-accumulating organisms (GAO), the major competitors of PAO, have a similar metabolism; they are unable to cycle phosphates, but under anaerobic conditions they rapidly consume readily biodegradable organic compounds and accumulate organic storage compounds. Most PAO have not been isolated in pure cultures, we have previously suggested that they possess an obligate cyclic metabolism [8]. However, highly enriched PAO cultures may be obtained under laboratory conditions in the feast-famine anaerobic–aerobic cycling mode.
The amount and composition of readily biodegradable organic matter supplied to the bioreactors or produced in them plays the major part in formation and functioning of the microbial community in EBPR bioreactors. Due to the differences in the spectra of utilized substrates and in their physiological properties, PAO and GAO may occupy different ecological niches and coexist in the EBPR activated sludge [9,10]. Most researchers consider the result of competition between GAO and PAO to be responsible for the relative abundance of these groups in the activated sludge consortia and therefore for the efficiency of biological phosphorus removal [4].
Low-molecular weight volatile fatty acids (VFA), particularly acetate and propionate, are universally considered the preferred carbon sources for Ca. Accumulibacter and most other PAO. The role of butyrate in Ca. Accumulibacter metabolism and its potential for BPR remain uncleared, and the research results are contradictory. Kong et al. [11] did not reveal butyrate consumption by the Rhodocyclus PAO in the activated sludge of EBPR facilities under aerobic or anaerobic conditions. On the contrary, Levantasi et al. [12] used the acetate/propionate/butyrate mixture to obtain a Ca. Accumulibacter-enriched culture, which actively removed phosphorus. Ability to cycle phosphorus upon addition of butyrate was shown for the Ca. Accumulibacter-enriched culture grown on propionate [13]. Begum and Batista [14] investigate the competition between PAO and GAO grown on butyrate as the sole carbon source. While the authors obtained the culture enriched with PAO (Ca. Accumulibacter) and GAO (Defluviicoccus), they noted unstable functioning of the community, as well as the low rates of butyrate consumption and phosphate release in the anaerobic phase. Wang et al. [15] achieved EBPR in a sequencing batch reactor (SBR), in which acetate was gradually replaced by butyrate at 30°C. Although replacement of acetate with butyrate resulted in partial replacement of Ca. Accumulibacter by Rhodocyclaceae, phosphorus removal remained stably high.
Ability of Ca. Accumulibacter and other PAO to use amino acids also remains an open issue. Tian et al. [16] observed a negative effect of glycine on EBPR and the inability of PAO- and GAO-enriched consortia to use glycine as the carbon source. At the same time, glycine stimulated phosphate release under anaerobic conditions and decreased the efficiency of phosphate uptake under aerobic conditions. Over 10 amino acids were shown to be probably involved to phosphorus cycling by the Tetrasphaera, with glycine showing the highest efficiency [17]. Marques et al. [9] also confirmed the ability of Tetrasphaera to utilize glycine and other amino acids, such as aspartate and glutamate, using a combination of microautoradiography and FISH, while Ca. Accumulibacter utilized the products of amino acid fermentation, rather than amino acids per se. On the contrary, according to Oyserman et al. [18], identification of the genes involved an anaerobic glycine utilization by Ca. Accumulibacter and enhanced phosphate release under anaerobic conditions indicated the ability of Ca. Accumulibacter to use glycine as a substrate. In-depth research of amino acid metabolism in Ca. Accumulibacter clade IIF strain SCELSE-1 was carried out by Qiu et al. [19]. They showed that the culture enriched with Ca. Accumulibacter clade IIF was able to metabolize 11 out of 20 α-amino acids, with aspartate, glutamate, asparagine, and glutamine being the most efficient substrates for phosphorus removal. The author suggested that aspartate was desaminated and was used to form polyhydroxyalkanoates (PHA) via core carbon metabolic pathways. Glutamate was stores as polyglutamate. Recent studies by Ziliani et al. [20] showed that, apart from VFA, the culture highly enriched with Ca. Accumulibacter could also use glucose.
Wastewater injected into EBPR bioreactors contains diverse organic compounds. Investigation of the spectrum of the substrates utilized by PAO and of the pathways of their metabolism is required for further elucidation of the physiological properties of these microorganisms, determination of their position in the microbial consortium, and optimization of existing EBPR technologies and development of new ones. Rapid determination of the ability of PAO to use a given substrates is needed for this purpose.
Oxygen uptake rate is among the most important, rapidly determined indicators of the catabolism rates in aerobic microorganisms. Oxygen uptake rate increase upon addition of a substrate to a starving culture usually indicates the ability to use this substrate as an energy source. However, investigation of the metabolic properties of aerobic PAO concentrates usually on the anaerobic phase, since it is considered that the unique metabolic characteristics of PAO, enabling them to compete for the substrate with other aerobic heterotrophic microorganisms, manifest themselves in the anaerobic phase [21]. PAO aerobic metabolism has been investigated mainly as a part of the anaerobic/aerobic cycle [22,23], and catabolism of extracellular substrates in the presence of electron acceptors got insufficient attention.
The goal of the present work was to determine the spectrum of organic substrates used as energy sources by a Ca. Accumulibacter-enriched culture under aerobic conditions and to reveal their relationship to the phosphorus cycling.
2. Materials and Methods
2.1. The bioreactor for obtaining a Ca. Accumulibacter-enriched culture
In acute experiments, the culture grown for a long term (2 years) in SBR with acetate as the main carbon source was used. The 2-L bioreactor was equipped with a system for aeration and nitrogen supply (Eltochpribor, Russia). Supply and removal of the medium were achieved using peristaltic pumps. Automatic control of gas flows and peristaltic pumps was carried out using a LOGО universal logical module (Siemens, Chengdu, China). An IKA C-MAG MS7 magnetic stirrer (Germany) was used for agitation. Anaerobic conditions were achieved by pumping nitrogen (0.3 L/min) into the bioreactor. The temperature was maintained at 18°C; pH was measured with an Expert-01 meter (Econix, Moscow, Russia). It was not adjusted and varied during the cycle from 7.5 to 8.2.
One cycle of SBR operation (6 h) consisted of five stages: medium supply under anaerobic conditions (30 min); anaerobic phase (2 h 25 min); aerobic phase (2 h 30 min); settling stage (30 min); and supernatant removal (5 min). In each SBR cycle, 08 L of the medium was replaced, with the hydraulic retention time of 15 h. The average solid retention time was maintained at 17.5 days.
The medium contained the following (g/L tap water): CH3COONa·3H2O, 0.67; (NH4)2SO4, 0.139; КH2PO4, 0.109; yeast extract, 0.009; MgSO4·7H2O, 0.150; and trace elements solution, 1 mL. Trace elements solution contained the following (g/L distilled water): Na-EDTA, 10; FeCl3·6H2O, 1.5; H3BO3, 0.15; CuSO4·5H2O, 0.03; MnCl2·4H2O, 0.12; Na2MoO4·2H2O, 0.06; ZnSO4·7H2O, 0.12; KI, 0.18; and CoCl2·H2O, 0.15. To suppress nitrification, thiourea was added to the final concentration of 2.5 mg/L. For pH adjustment, 0.5 M HCl was added to the medium (7.5 mL/L).
2.2. Determination of the oxygen uptake rates
At the end of the aerobic phase, a 800-mL sample of the culture was collected and incubated in a refrigerator for 20 h in order to obtain a culture in the standard physiological state with predominance of endogenous metabolism. The culture was then aerated for 2 h at 20°C, with the concentration of dissolved oxygen maintained at 6‒8 mg O2/L. A 100-mL aliquot was diluted 5-fold with the salt buffer (the medium without acetate and yeast extract) and supplemented with 1 mL of phosphate buffer to the final pH of 7.1. The samples thus obtained were dispensed into glass vials and hermetically sealed without the gas phase. Dynamics of endogenous O2 consumption was then determined, followed by O2 consumption after addition of a carbon and energy source. For all substrates, except for Bacto-tryptone, the initial concentration was 200 mg COD/L. Bacto-tryptone was added to the final concentration of 2 g/L. The tested substrates were VFA salts (acetate, propionate, butyrate, and formate), amino acids (glutamic acid, alanine, serine, threonine, aspartic acid, lysine, and glycine), glucose, ethanol, succinate, pyruvate, glycerol, and tryptone. All reagents were produced by Diaem (Moscow, Russia).
The experiments were carried out at 20±0.2°C. Oxygen concentration was measured electrochemically using an inoLab® Oxi 7310 oxygen meter with a StirrOx G sensor (WTW, Germany). The rate of oxygen uptake was calculated at the linear part of the curve of dissolved O2 concentration decrease with subsequent calculation of the specific oxygen uptake rate. If not stated otherwise, the specific oxygen uptake rate (OUR) was used:
OUR = OURsum – OURend, where
OURsum is the total specific oxygen uptake rate and
OURend is the endogenous specific oxygen uptake rate.
2.3. Determination of dynamics of substrate and phosphate in batch experiments
A 200-mL culture sample from the SBR was transferred into a 500-mL vial. The vial was purged with nitrogen (0.3 L/min) for 10 min in order to establish anaerobic conditions, and the substrate was added to the final concentration of 200 mg COD/L (succinate concentration used was 100 mg COD/L). COD was calculated according to the chemical reaction of complete oxidation of an organic substrate with oxygen. After 2 h, air was supplied to the culture (2 L/min), which resulted in oxygen concentrations of at least 2.0 mg O2/L. The culture to which no substrate was added was used as the control. Dynamics of the concentrations of phosphate and the substrate in the medium was monitored during the experiment. The temperature was maintained at 20±0.2°.
2.4. Molecular techniques
2.5. Analytical techniques
Phosphate ion concentration, TSS, and VSS were determined as described previously [24]. The concentrations of VFA, succinate, and pyruvate we determined by HPLC using Staier (Akvilon, Russia).
2.6. Statistical data processing
The concentrations of phosphate ions and organic compounds were carried out in triplicate. Average values and standard deviation were determined for each experimental point. The deviation was within the limits of experimental data scattering and did not exceed 5%. It is shown on the graphs as data scattering.
3. Results
3.1. Bioreactor operation and microbial community composition
After 2 years of SBR cultivation, a stable microbial community developed, which was characterized by dynamics of the substrate (acetate) and phosphate in the medium typical of the PAO phenotype. The molar ratio of phosphorus released in the anaerobic phase to that of consumed acetate was 0.6‒0.76 P-mol/S-mol, indicating high enrichment of the culture with PAO [2,26,27,28,29]. The biomass concentration during the experiment was 4.4‒4.6 g TSS/L (2.8‒2.9 g VSS/L). Phosphorus content in the biomass at the end of the aerobic phase was 14‒16% VSS, being significantly higher than that in the biomass of non-PAO microorganisms. Detailed description of the community development and dynamics of the SBR operational characteristics was published earlier [25].
During the experiment, the community grew as granule-like aggregates; their taxonomic composition determined by the 16S rRNA gene sequencing is presented on Figure 1. The microbial community was mainly represented by members of the phyla Pseudomonadota and Bacteroidota (56.8 and 19.6%, respectively). The PAO Ca. Accumulibacter was the dominant organism of the community (36.9% of the total number of the 16S rRNA gene sequences). Other members of the phylum Pseudomonadota, potentially belonging to PAO, were also present in the community: Dechloromonas (5.02%) and Thauera (7.09%). The GAO Ca. Competibacter comprised an insignificant part of the community (2,74%) The phylum Bacteroidota was mostly represented by heterotrophic organisms capable of degrading complex organic polymers: members of the orders Flavobacteriales, Chitinophagales, Cytophagales, and Ignavibacteriales.
3.2. Oxygen consumption
Prior to the experiments, endogenous respiration (OURend) was on average 1.65±0.48 mg O2/(g TSS h). After addition of the substrates, О2 consumption rate increased for 2-3 min, reached the maximum, and stabilized. The highest OUR for acetate was 32,2±2,8 mg O2/(g TSS h) (50,3±4,4 mg O2/(g VSS h)). While for tryptone the initial OUR was close to that for acetate, it then rapidly decreased, reaching 35‒40% of the initial values after 20 min. Among other substrates, OUR decreased in the series: pyruvate – propionate – butyrate - succinate – aspartate – glutamate – alanine. For the other substrates, OUR was below 10% of the values observed in the presence of acetate (Figure 2).
3.3. Cycle of phosphate release/uptake under anaerobic/aerobic conditions
In spite of predominance of Ca. Accumulibacter in the community, other aerobic microorganisms not belonging to PAO certainly contributed to the total OUR. For investigation of the effect of external carbon and energy sources on the cycle of phosphorus release/uptake, the substrates providing for the highest OUR were chosen (tryptone was not used due to its multicomponent composition). Ethanol and glucose were also used as compounds widespread in natural and artificial ecosystems.
According to their effect on the phosphorus release/uptake cycle, the tested substrates were subdivided into three groups (Figure 3).
Addition of group 1 substrates (acetate, propionate, pyruvate, and succinate) resulted in phosphate release during the anaerobic phase with its subsequent 60‒90% consumption in the aerobic phase. Acetate, propionate, and pyruvate had the most pronounced effect, while succinate had the least effect. During the anaerobic phase, acetate and propionate were completely consumed during the first 30 min (Figure 4), which was accompanied by phosphate release to the concentrations of 145 and 95 mg РО43--P/L, respectively. During the aerobic stage, phosphates were consumed, and their concentration decreased to 19 and 14 mg РО43--P/L, respectively. The rates of pyruvate, succinate, and butyrate were lower than those for acetate and propionate, and phosphate cycling in the anaerobic/aerobic stages was less pronounced. While butyrate caused rapid phosphate release to 69 mg РО43--P/L, the rate of phosphate uptake in the aerobic phase was low. By the end of the aerobic phase, phosphate concentrations in the presence of pyruvate, succinate, and butyrate decreased to 5, 4, and 23 mg РО43--P/L, respectively (Figure 3a).
The substrates of group 2 (amino acids glutamate, aspartate, and alanine) caused phosphate release, but phosphate uptake during the aerobic phase was lower compared to group 1 (Figure 3b). Glutamate and aspartate showed the highest effect.
No noticeable phosphate cycling was found for group 3 substrates, glucose and ethanol (Figure 3c). Upon addition of ethanol, phosphate concentration increased gradually both in the anaerobic and aerobic phases. Glucose had no effect on phosphate dynamics compared to the control.
The ratio between the amounts of phosphate released during the anaerobic phase (amount of hydrolyzed polyphosphate) and the amount of consumed substrate (P-release/substrate-uptake) is one of the major stoichiometric indicators of AO metabolism. In PAO these processes are coupled. Polyphosphate hydrolysis provides for substrate transport by establishing the transmembrane proton gradient and for formation of intracellular storage compounds. The greater the energy expense for substrate transport and accumulation, the higher the P-release/substrate-uptake ratio. While this parameter for VFA depends considerably on the ambient pH, in the near-neutral range with acetate as the substrate it is close to 1 P-mol/S-mol of the substrate (0.5 P-mol/C-mol) [3,30,31].
The P-release/substrate-uptake values calculated using our experimental results and integral characteristics of the parameters of the anaerobic/aerobic cycle and OUR are presented in Table 1. The P-release/substrate-uptake values for amino acids were calculated assuming complete consumption of the substrate during the anaerobic phase. In the case of VFA, the highest value (2.1 P-mol/S-mol) was obtained for butyrate. The values for aspartate and glutamate were 2.0 and 2.1 P-mol/S-mol, respectively. The P-release/substrate-uptake ratio for alanine was significantly lower, 0.3 P-mol/S-mol.
3.4. Abilities of Ca. Accumulibacter transport systems
We have previously sequenced the metagenome of the microbial community [25]. Metagenomic analysis resulted in assembly of the Ca. Accumulibacter genomes represented in the studied microbial community. Analysis of these genomes revealed the major transport systems of Ca. Accumulibacter. The genes encoding the transport systems for all substrates causing noticeable phosphate cycling in the anaerobic/aerobic cycle were revealed . The genes responsible for transport of VFA (actP), pyruvate (btsT), succinate (dctM,P,Q,R,S), and amino acids (gltI, K, J, L, and livK, H, M, G, F) were revealed in Ca. Accumulibacter.
4. Discussion
Analysis of the taxonomic composition (Figure 1) showed predominance of Ca. Accumulibacter (36.9%), the main universally accepted PAO responsible for BPR under anaerobic/aerobic conditions. Members of the genera Thauera (7.09%) and Dechloromonas (5.02%), some representatives of which belong to denitrifying PAO [6,32], were also present in the community. Since the relative abundance of Thauera and Dechloromonas was significantly lower than that of Ca. Accumulibacter, and since high phosphate-accumulating activity of these organisms under anaerobic/aerobic conditions is not highly probable, in our opinion the results obtained in the present work depended mainly on the metabolic properties of Ca. Accumulibacter.
4.1. Response of Ca. Accumulibacter to the substrates
4.1.1. Volatile fatty acids
Dynamics of phosphate cycling and consumption of acetate and propionate were in agreement with the known PAO phenotype of Ca. Accumulibacter. These are the substrates traditionally used as carbon sources to obtain PAO-enriched cultures. The P-release/substrate-uptake values for acetate and propionate were 1.4 and 1.6 P-mol/S-mol (0.7 and 0.53 P-mol/C-mol), respectively. This is close to the similar values reported by other researchers for PAO within the pH range of 7.0‒8.5 [33,34,35] and indicates substrate consumption mainly by PAO, rather than GAO; for the latter, the P-release/substrate-uptake is close to zero. Utilization of butyrate resulted in phosphorus cycling, although the rates of phosphate release and substrate consumption in the anaerobic phase were significantly lower than those for acetate and propionate. The P-release/substrate-uptake ration (2.1 P-mol/S-mol) was higher than in the case of other VFAs, indicating lower efficiency of utilization of this substrate.
The earlier [25] metagenomic analysis of the Ca. Accumulibacter culture used in the present work revealed the genes encoding the main metabolic pathways for acetate, PHA, glycogen, and phosphates, including the genes required for consumption and VFA conversion to PHA. Acetate permease encoded by the actP gene is involved in acetate transport [36]. The product of this gene is used for acetate transport and, to a lesser degree, in that of propionate and (still less actively) of butyrate [37,38]. The VFAs acquired via transport are converted to acetyl-CoA by acetyl-CoA synthetase (acs) or acetate kinase (ackA) and phosphate acetyltransferase (pta) [39]. The genes of acetate transport and conversion to acetyl-CoA have been identified, apart for Ca. Accumulibacter MAG, in the genomes of Competibacter, Siculibacillus, Azonexus, and Myxococcacea. However, the genes encoding acetyl-CoA conversion to PHA have been found only in Ca. Accumulibacter. The efficiency of VFA by Ca. Accumulibacter via acetate permease is probably the factor determining the high activity of phosphate release when acetate is used as the substrate. No specific propionate transporter was (prpP) was found in any of the studied MAGs. Metagenomic analysis did not reveal the genes encoding specific butyrate transport, which explained lower rates of phosphate release during the anaerobic phase of the cycle in the presence of this substrate. Another factor responsible for lower efficiency of propionate and butyrate is the necessity for additional conversions to form acetyl-CoA, requiring propionyl-CoA synthetase (prpE) [40].
Taking into account the results of our experiments and of metagenomic analysis, butyrate may be considered a substrate used by Accumulibacter and potentially acceptable for BPR. In general, our results are in agreement with those of researchers reporting the possible application of butyrate as a substrate for Ca. Accumulibacter and for EBPR [13,14,15]. Since the rates of metabolic processes (phosphate release and substrate consumption in the anaerobic phase and phosphate consumption in the aerobic one) are lower than those for acetate and propionate, optimization of PAO cultivation and phosphorus removal requires adjustment of duration of the phases of the anaerobic/aerobic cycle, which should be longer than with acetate or propionate.
4.1.2. Pyruvate
When pyruvate was used, the rate of phosphate release and it maximum concentration during the anaerobic phase was somewhat lower, although generally close to those observed for acetate and propionate. This was to be expected, since pyruvate is one of the central intermediates in metabolism of organisms, including PAO. Although Arun et al. [41] and Satoh et al. [42] observed phosphate cycling after addition of pyruvate to the activated sludge of EBPR facilities, the ability of Ca. Accumulibacter to carry out this process has not been reported previously. Almost complete consumption of pyruvate during the anaerobic phase and the low P-release/substrate-uptake ratio (1.7 P-mol/S-mol) indicated an efficient system for pyruvate transport into the cells and formation of the intracellular storage material. The relatively high efficiency of phosphorus cycling with pyruvate may be related to the presence of a pyruvate/proton symporter (btsT) in Ca. Accumulibacter MAG, as well as of the lactate transporter (jen1), which is capable of pyruvate and propionate transport, although less efficiently than lactate [43].
4.1.3. Succinate
Succinate is one of the major intermediates in the central metabolic pathways. Under anaerobic conditions, it is the terminal product of organic matter metabolism by many microorganisms, e.g., in anaerobic fermentation of organic waste [44,45,46]. Early research on the BPR mechanism [41,42] showed that emergence of the PAO phenotype in the activated sludge of BPR facilities and in the culture grown in SBR with succinate. However, we found no works on its possible role in the metabolism and phosphate cycling in Ca. Accumulibacter. Among the transport systems of Ca. Accumulibacter we identified the genes of the C4-dicarboxylate transporter system (dctM,P,Q,R,S), which indicated the ability of Ca. Accumulibacter to transport succinate. In our experiments, addition of succinate resulted in its 80% consumption during the anaerobic phase and to a considerable phosphate release (up to 28.5 mg РО43--P/L. The rate of phosphate release was significantly lower than for other studied VFA. During the aerobic phase, over 80% of released phosphate was consumed. The P-release/substrate-uptake ratio for succinate had the lowest value of 1.3 P-mol/S-mol, indicating high efficiency of its transport and intracellular storage. We do not rule out the possibility of succinate consumption by other microorganisms with formation of, e.g., propionate, which may be used by Ca. Accumulibacter. However, the results of the present work indicate that succinate may be considered a promising substrate for Ca. Accumulibacter enrichment and should be taken into account in technological calculations.
4.1.4. Amino acids
The genes of the general L-amino acid transport system permease (aapJ, Q, M, P) may be considered the determinants of aspartate, alanine, and glutamate consumption [47]. Ca. Accumulibacter does not bear the aapP gene. The genomes of Ca. Accumulibacter were also found to contain the genes for transport of branched amino acids (livKHMGF) [47,48].
Among the tested amino acids, the highest OURs were observed upon addition of glutamate, aspartate, and alanine. Our data obtained under batch conditions revealed a considerable stimulating effect of glutamate and aspartate on phosphate release under anaerobic conditions. Metagenomic analysis revealed the presence in Ca. Accumulibacter of the genes of the glutamate/aspartate transport system ATP-binding protein (gltIKJL), which indicated ability to transport these amino acids and accumulate intracellular storage compounds. Dynamics of amino acid concentrations was not determined in the present work, but the P-release/substrate-uptake ratio for glutamate and aspartate was somewhat higher than the values cited in the literature for Ca. Accumulibacter (1.2 and 1.4 P-mol/S-mol) [19], reaching 2.0 and 2.1 P-mol/S-mol, respectively, which was an indirect indication of complete consumption of these substrates during the anaerobic phase. At the same time, the rates of phosphate uptake in batch experiments under aerobic conditions in the presence of glutamate and aspartate were low, and no significant phosphate consumption occurred. Our results are similar to those of Qiu et al. [19], who showed that in the culture enriched with Ca. Accumulibacter clade IIF 11 out of 20 tested amino acids caused measurable phosphorus release, with the highest phosphorus removal efficiency and phosphate cycling observed with glutamate and aspartate. In batch experiments, the authors observed only partial consumption of phosphate released under anaerobic conditions. At the same time, in SBR Qiu et al. observed complete phosphate uptake during the aerobic phase of the SBR cycle.
The effect of alanine on phosphate cycling was significantly less pronounced. While Ca. Accumulibacter was found to possess the gene encoding the branched-chain amino acid transport system, which indicated ability of Ca. Accumulibacter to transport alanine, considering the low rates of phosphate uptake and the low P-release/substrate-uptake value (0.3 P-mol/S-mol), alanine consumption during the anaerobic phase was probably insignificant, and this substrate is not easily available to Ca. Accumulibacter.
4.2. Correlation between OUR and phosphate uptake during the anaerobic stage
Analysis of our results revealed a non-evident correlation between OUR after addition of the substrate and the amount of phosphates consumed during the aerobic phase of the aerobic/anaerobic cycle (Figure 5). The higher was the specific OUR upon substrate addition to the starving culture, the greatest amount of phosphates was consumed during the aerobic phase when the substrate was added in the anaerobic phase. Specific OUR values for acetate, pyruvate, and propionate were considerably lower than for other tested substrates, which were characterized both by lower OUR and lower ability of the organisms to uptake phosphates during the aerobic phase. At the same time, no relation was found between OUR and the rate of phosphate release when the substrates were added during the anaerobic phase of the cycle. Thus, glutamate was characterized by a high rate of phosphate release, while OUR was low. Deviation of the results obtained with pyruvate (point 8 on Figure 5) may be due to its oxidation by the microorganisms other than PAO.
In our opinion, variation in the amount of phosphate consumed during the aerobic phase results from the different rates of catabolism of organic compounds accumulated during the anaerobic phase. During the aerobic phase of the cycle, the lower the rates of the energy-yielding processes, the lower the intensity of polyphosphate accumulation. Intensity of catabolism of organic storage compound should certainly depend on their amount and composition, which change depending on the substrate consumed. In the culture enriched with Ca. Accumulibacter, synthesis of different PHA (PHB, PHV, or poly-beta-hydroxy-2-methylvalerate) occurs depending on the substrate type during the anaerobic phase [19], utilization of which during the aerobic phase is associated with efficiency of phosphate uptake [49]. Apart from PHA, PAO cells may accumulate other storage compounds. Pijuan et al. [13] investigated a PAO-enriched culture and revealed among the intracellular compounds some unidentified monomers forming only in the presence of butyrate and constituting most of the monomers produced by the culture. Based on metagenomic analysis, Qiu et al. [19] concluded that in Ca. Accumulibacter clade IIF aspartate is desaminated and directed to the main carbon metabolic pathways for polylhyroxyalkanoate (PHA) formation, while glutamate is stored as polyglutamate. Under anaerobic conditions, members of the genus Tetrasphaera were shown to accumulate glycogen, glycine, and fermentation products as storage compounds [9]. Decreased efficiency of catabolism of accumulated compounds results in decreased rates of polyphosphate formation. In our opinion, the correlation between OUR and phosphate uptake observed in our experiments for different substrates may indicate that consumption of more energy-efficient substrates results in accumulation of more energy-efficient storage compounds. Moreover, PAO biomass increases during the aerobic phase of the cycle, and regulatory systems may exist, which direct the energy flow primarily to growth, maintenance, and glycogen regeneration. In this case, synthesis of a stored energy source in the form of polyphosphates is possible only under "excessive" catabolism, which becomes more pronounced when easily oxidized substrates are used. However, we are not aware of systematic studies on catabolism regulation of intracellular stored sources of carbon and energy in PAO during the aerobic phase.
Kinetic and stoichiometric parameters of catabolism of the substrates and of phosphate release/uptake evidently depend on a number of factors, including additivity and synergism in the case of simultaneous consumption of several substrates. Additivity of substrate utilization may explain the high rate of tryptone, a multicomponent substrate, which was observed in our experiments. Although no easily oxidized substrates of the VFA group were found in the tryptone used, the initial OUR was much higher than for the studied amino acids and was comparable for the OUR with acetate. At the same time, OUR decreased by 2.5‒3 times after 20–30 min, which indicated rapid consumption and exhaustion of the easily oxidized tryptone components. It should be noticed that simultaneous utilization of several substrates, one of them not being a VFA, has been reported for Ca. Accumulibacter. Thus, in the work by Qiu et al. [19], simultaneous addition of acetate and glutamate or aspartate resulted in almost the same consumption rates for each carbon source.
The present work was not aimed at determination of the ability of the tested substrates to support PAO growth for a long time. However, acute experiments revealed that the community enriched with a typical aerobic PAO Ca. Accumulibacter was able to metabolize a broad spectrum of organic compounds with different rates of their consumption and catabolism and different effects on the anaerobic/aerobic phosphate cycling. Genomic analysis of the Ca. Accumulibacter revealed the ability of this organism to uptake various substrates.
OUR may be used as an express method for search of the new potential PAO substrates. OUR values in PAO-enriched cultures correlate with the amount of phosphate consumed in the aerobic phase of the anaerobic/aerobic cycle. Further improvement of our understanding of the spectrum of the substrates utilized by PAO is certainly of interest both for optimization of BPR technologies and development of new ones and for determination of the role of PAO in natural ecosystems.
5. Conclusions
In the present work, the oxygen uptake rates of Ca. Accumulibacter-enriched culture were measured for 17 organic substrates in order to determine the spectrum of the substrates used by Ca. Accumulibacter. The highest OUR values were observed for tryptone, acetate, propionate, butyrate, succinate, pyruvate, aspartate, and glutamate. Dynamics of phosphate release/uptake upon respective shift from anaerobic to aerobic conditions was studied for these compounds in batch experiments. Based on stoichiometric data and the results of metagenomic analysis, it was concluded that all studied compounds could be used by Ca. Accumulibacter as substrates. A correlation between OUR and the amount of phosphates consumed during the aerobic phase was revealed in batch experiments. OUR was proposed as a rapid indicator for search of the substrates used by Ca. Accumulibacter.
Author Contributions
Conceptualization, A.D.; methodology, A.D. and A.P.; validation, Y.N., N.P. and A.M.; investigation, A.P., A.D. and E.G.; resources, A.M. and Y.B.; data curation, Y.B and E.G.; writing—original draft preparation, A.D. and A.P.; writing—review and editing, N.P., A.M. and Y.N.; supervision, N.P.; project administration, A.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Russian Science Foundation, grant number 21-64-00019, and by the RF Ministry of Science and Higher Education.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wentzel, M.C.; Loewenthal, R.E., Ekama, G.A.; Marais, G.V.R. Enhanced polyphosphate organism cultures in activated sludge systems – Part 1: Enhanced culture development. Water S.A. 1988, 14, 81–92.
- Schuler, A.J.; Jenkins, D. Enhanced biological phosphorus removal from wastewater by biomass with different phosphorus contents, part1: Experimental Results and Comparison with Metabolic Models. Water Environ. Res. 2003, 75, 485–498. [CrossRef]
- Welles, L.; Abbas, B.; Sorokin, D.Y.; Lopez-Vazquez C.M.; Hooijmans C.M, van Loosdrecht M.C.M.; Brdjanovic D. Metabolic response of 'Candidatus Accumulibacter phosphatis' clade II to changes in the influent P/C ratio. Front. Microbiol. 2017, 7, 2121. https://doi.org/10.3389/fmicb.2016.02121. [CrossRef]
- Oehmen, A.; Lemos, P.C.; Carvalho, G.; Yuan, Z.; Keller, J.; Blackall, L.L.; Reis, M.A.M. Advances in enhanced biological phosphorus removal: from micro to macro scale. Water Res. 2007, 41, 2271–2300. [CrossRef]
- Dorofeev, A.G.; Nikolaev, Y.A.; Mardanov, A.V.; Pimenov, N.V. Role of phosphate-accumulating bacteria in biological phosphorus removal from wastewater. Appl. Biochem. Microbiol. 2020, 56, 3-18. https://doi.org/10.1134/S0003683820010056. [CrossRef]
- Zhao, W.; Bi, X.; Peng, Y.; Bai, M. Research advances of the phosphorus-accumulating organisms of Candidatus Accumulibacter, Dechloromonas and Tetrasphaera: Metabolic mechanisms, applications and influencing factors. Chemosphere 2022, 307 Part 1, 135675. https://doi.org/10.1016/j.chemosphere.2022.135675. [CrossRef]
- Paez-Watson, T.; van Loosdrecht, M.C.M.; Wahl, S.A. Predicting the impact of temperature on metabolic fluxes using resource allocation modelling: Application to polyphosphate accumulating organisms. Water Res. 2023, 228, 119365. https://doi.org/10.1016/j.watres.2022.119365. [CrossRef]
- Dorofeev, A.G.; Nikolaev, Y.A.; Mardanov, A.V.; Pimenov, N.V. Cyclic metabolism as a mode of microbial existence. Microbiology 2019, 88, 402-415. https://doi.org/10.1134/S0026261719040052. [CrossRef]
- Marques, R.; Santos, J.; Nguyen, H.; Carvalho, G.; Noronha, J.P.; Nielsen, P.H.; Reis, M.A. M.; Oehmen, A. Metabolism and ecological niche of Tetrasphaera and Ca. Accumulibacter in enhanced biological phosphorus removal. Water Res. 2017, 122, 159–171. https://doi.org/10.1016/j.watres.2017.04.072. [CrossRef]
- Brand, V.R.; Crosby, L.D.; Criddle, C.S. Niche differentiation among three closely related Competibacteraceae clades at a fullscale activated sludge wastewater treatment plant and putative linkages to process performance. Appl. Environ. Microbiol. 2019, 85, e02301-18. https://doi.org/10.1128/AEM.02301-18. [CrossRef]
- Kong, Y.; Nielsen, J.L.; Nielsen, P.H. Microautoradiographic study of Rhodocyclus-related polyphosphate-accumulating bacteria in full-scale enhanced biological phosphorus removal plants. Appl. Environ. Microbiol. 2004, 70, 5383-5390. [CrossRef]
- Levantesi, C.; Serafim, L.S.; Crocetti, G.R.; Lemos, P.C.; Rossetti, S.; Blackall, L.L.; Reis, M.A.M.; Tandoi, V. Analysis of the microbial community structure and function of a laboratory scale enhanced biological phosphorus removal reactor. Environ. Microbiol. 2002, 4, 559–569. [CrossRef]
- Pijuan, M.; Baeza, J.A.; Casas, C.; Lafuente J. Response of an EBPR population developed in an SBR with propionate to different carbon sources. Water Sci. Technol. 2004, 50, 131–138. [CrossRef]
- Begum, S.A.; Batista, J.R. Impact of butyrate on microbial selection in enhanced biological phosphorus removal systems. Environ. Technol. 2014, 35, 2961–2972. [CrossRef]
- Wang, L.; Liu, J.; Oehmen, A.; Le, C.; Geng, Y.; Zhou Y. Butyrate can support PAOs but not GAOs in tropical climates. Water Res. 2021, 193, 116884. https://doi.org/10.1016/j.watres.2021.116884. [CrossRef]
- Tian. Y.; Chen, H.; Chen, L.; Deng, X.; Hu, Z.; Wang, C.; Wei, C.; Qiu, G.; Wuertz, S. Glycine adversely affects enhanced biological phosphorus removal. Water Res. 2022, 209, 117894. https://doi.org/10.1016/j.watres.2021.117894. [CrossRef]
- Nguyen, H.T.; Kristiansen, R.; Vestergaard, M.; Wimmer, R.; Nielsen, P.H. Intracellular accumulation of glycine in polyphosphate-accumulating organisms in activated sludge, a novel storage mechanism under dynamic anaerobic-aerobic conditions. Appl. Environ. Microbiol. 2015, 81, 4809–4818. [CrossRef]
- Oyserman, B.O.; Noguera, D.R.; del Rio, T.G.; Tringe. S.G.; McMahon, K.D. Metatranscriptomic insights on gene expression and regulatory controls in Candidatus Accumulibacter phosphatis. ISME J. 2016, 10, 810–822. [CrossRef]
- Qiu, G.; Liu, X.; Saw, N.; Law, Y.; Zuniga-Montanez, R.; Thi, S.S.; Nguyen, T.Q.N.; Nielsen, P.H.; Williams, R.B.H.; Wuertz, S. Metabolic traits of Candidatus Accumulibacter clade IIF strain SCELSE-1 using amino acids as carbon sources for enhanced biological phosphorus removal. Environ. Sci. Technol. 2020, 54, 2448–2458. https://doi.org/10.1021/acs.est.9b02901. [CrossRef]
- Ziliani, A.; Bovio-Winkler, P.; Cabezas, F.; Etchebehere, C.; Garcia, H.A.; López-Vázquez, C.M.; Brdjanovic, D.; van Loosdrecht, M.C.M.; Rubio-Rincón, F.J. Putative metabolism of Ca. Accumulibacter via the utilization of glucose. Water Res. 2023, 229, 119446. https://doi.org/10.1016/j.watres.2022.119446. [CrossRef]
- McMahon, K.D.; He, S.; Oehmen, A. The microbiology of phosphorus removal. In Microbial Ecology of Activated Sludge; Seviour, R., Nielsen, P.H., Eds.; IWA, London, 2010; pp. 281–319.
- Smolders, G.J.; van der Meij, J.; van Loosdrecht, M.C.M.; Heijnen, J.J. Stoichiometric model of the aerobic metabolism of the biological phosphorus removal process. Biotechnol. Bioeng. 1994, 44, 837–848. [CrossRef]
- Welles, L.; Lopez-Vazquez, C.M.; Hooijmans, C.M.; van Loosdrecht, M. C. M.; Brdjanovic, D. Impact of salinity on the aerobic metabolism of phosphate-accumulating organisms. Appl. Microbiol. Biotechnol. 2015, 99, 3659–3672. [CrossRef]
- Pelevina, A.V.; Berestovskaya, Y.Y.; Grachev, V.A.; Dorofeeva, I.K.; Sorokin, V.V.; Dorofeev, A.G.; Kallistova, A.Yu.; Nikolaev, Yu.A.; Kotlyarov, R.Yu.; Beletskii, A.V.; Ravin, N.V.; Pimenov, N.V.; Mardanov, A.V. A Microbial Consortium Removing Phosphates under Conditions of Cyclic Aerobic-Anaerobic Cultivation. Microbiology 2021, 90, 66–77. https://doi.org/10.1134/S0026261721010082. [CrossRef]
- Pelevina, A.V.; Grouzdev, E.V.; Berestovskaya, Yu.Yu.; Dorofeev, A.G.; Nikolaev, Yu.A.; Kallistova, A.Yu.; Beletsky, A.V.; Ravin, N.V.; Pimenov, N.V.; Mardanov, A.V. New insight into the granule formation in the reactor for enhanced biological phosphorus removal. Front. Microbiol. 2023, in press.
- Brdjanovic, D.; van Loosdrecht, M.C.M.; Hooijmans, C.M.; Alaerts, G.J.; Heijnen, J.J. Temperature effects on physiology of biological phosphorus removal. J. Environ. Eng. 1997, 123, 144–153. [CrossRef]
- Schuler, A.J. Jenkins, D. Enhanced biological phosphorus removal from wastewater by biomass with different phosphorus contents, part II: anaerobic adenosine triphosphate utilization and acetate uptake rates. Water Environ. Res. 2003, 75, 512–522. [CrossRef]
- Bassin, J.P.; Winkler, M.-K.H.; Kleerebezem, R.; Dezotti, M.; van Loosdrecht, M.C.M. Improved phosphate removal by selective sludge discharge in aerobic granular sludge reactors. Biotechnol. Bioeng. 2012, 109, 1919–1928. [CrossRef]
- Acevedo. B.; Murgui, M.; Borr´as, L.; Barat, R. New insights in the metabolic behaviour of PAO under negligible poly-P reserves. Chem. Eng. J. 2017, 311, 82–90. [CrossRef]
- Smolders, G.J.F.; van der Meij, J.; van Loosdrecht, M.C.M.; Heijnen, J.J. Model of the Anaerobic Metabolism of the Biological Phosphorus Removal Process: Stoichiometry and pH Influence. Biotechnol. Bioeng. 1994, 43, 461-470. [CrossRef]
- Liu, W.-T.; Mino, T.; Matsuo, T.; Nakamura, K. Biological phosphorus removal processes - Effect of pH on anaerobic substrate metabolism. Water Sci. Technol. 1996, 34, 25-32. [CrossRef]
- Ren, T.; Chi, Y.; Wang, Y.; Shi, X.; Jin, X.; Jin, P. Diversified metabolism makes novel Thauera strain highly competitive in low carbon wastewater treatment. Water Res. 2021, 206, 117742. https://doi.org/10.1016/j.watres.2021.117742. [CrossRef]
- Oehmen, A.; Zeng, R.J.; Yuan, Z.; Keller, J. Anaerobic metabolism of propionate by polyphosphate-accumulating organisms in enhanced biological phosphorus removal systems. Biotechnol. Bioeng. 2005, 1, 43-53. [CrossRef]
- L´opez-V´azquez, C.M.; Hooijmans, C.M.; Brdjanovic, D.; Gijzen, H.J.; van Loosdrecht, M. C.M. A practical method for quantification of phosphorus- and glycogen accumulating organism populations in activated sludge systems. Water Environ. Res. 2007, 79, 2487–2498. [CrossRef]
- 35. Diaz, R.; Mackey, B.; Chadalavada, S.; Kainthola, J.; Heck. P.; Goel, R. Enhanced Bio-P removal: Past, present, and future – A comprehensive review. Chemosphere 2022, 309, 136518. https://doi.org/10.1016/j.chemosphere.2022.136518. [CrossRef]
- Ribas, D.; Soares-Silva, I.; Vieira, D.; Sousa-Silva, M.; Sá-Pessoa, J.; Azevedo-Silva, J.; Viegas, S.C.; Arraiano, C.M.; Diallinas, G.; Paiva, S.; Soares, P.; Casal, M. The acetate uptake transporter family motif “NPAPLGL (M/S)” is essential for substrate uptake. Fungal Genet. Biol. 2019, 122, 1-10. [CrossRef]
- Gimenez, R.; Nuñez, M.F.; Badia, J.; Aguilar, J.; Baldoma, L. The gene yjcG, cotranscribed with the gene acs, encodes an acetate permease in Escherichia coli. J. Bacteriol. 2003, 185, 6448-6455. [CrossRef]
- Jolkver, E.; Emer, D.; Ballan, S.; Krämer, R.; Eikmanns, B.J.; Marin, K. Identification and characterization of a bacterial transport system for the uptake of pyruvate, propionate, and acetate in Corynebacterium glutamicum. J. Bacteriol. 2009, 191, 940-948.
- Barr J.J.; Dutilh, B.E.; Skennerton, C.T.; Fukushima, T.; Hastie M.L.; Gorman, J.J.; Tyson, G.W.; Bond, P.L. Metagenomic and metaproteomic analyses of Accumulibacter phosphatis-enriched floccular and granular biofilm. Environ. Microbiol. 2016, 18, 273-287. [CrossRef]
- Liu, X.-W.; Wang, H.-H.; Chen, J.-Y.; Li, X.-T.; Chen, G.-Q. Biosynthesis of poly (3-hydroxybutyrate-co-3-hydroxyvalerate) by recombinant Escherichia coli harboring propionyl-CoA synthase gene (prpE) or propionate permease gene (prpP). Biochem. Eng. J. 2009, 43, 72-77. [CrossRef]
- Arun, V.; Mino, T.; Matsuo, T. Metabolism of carboxylic acids located in and around the glycolytic pathway and the TCA cycle in the biological phosphorus removal process. In Proceedings of the Fourteenth Biennial Conference of the International Association on Water Pollution Research and Control, Brighton, U.K., 18–21 July, 1988. https://doi.org/10.1016/B978-1-4832-8439-2.50038-9. [CrossRef]
- Satoh, H.; Ramey, W.D.; Koch F.A.; Oldham W.K.; Mino, T.; Matsuo T. Anaerobic substrate uptake by the enhanced biological phosphorus removal activated sludge treating real sewage. Water Sci. Technol. 1996, 34, 9-16. [CrossRef]
- Skory, C.D.; Hector, R.E.; Gorsich, S.W.; Rich J.O. Analysis of a functional lactate permease in the fungus Rhizopus. Enzyme Microb. Technol. 2010, 46, 43-50. [CrossRef]
- Song, H.; Lee, S.Y. Production of succinic acid by bacterial fermentation. Enzyme Microbial Technol. 2006, 39, 352–361. [CrossRef]
- Wainaina, S.; Lukitawesa; Awasthi, K.M.; Taherzadeh, M.J. Bioengineering of anaerobic digestion for volatile fatty acids, hydrogen or methane production: A critical review. Bioengineered 2019, 10, 437-458. https://doi.org/10.1080/21655979.2019.1673937. [CrossRef]
- Putri, D.N.; Sahlan, M.; Montastruc, L.; Meyer, M.; Negny, S.; Hermansyah, H. Progress of fermentation methods for bio-succinic acid production using agro-industrial waste by Actinobacillus succinogenes. Energy Rep. 2020, 6, 234–239. https://doi.org/10.1016/j.egyr.2019.08.050. [CrossRef]
- 47. Hosie, A. H. F.; Poole, P. S. Bacterial ABC transporters of amino acids. Res. microbiol. 2001, 152, 259-270. [CrossRef]
- Wang, S.; Yan, Z.; Wang, P.; Zheng, X.; Fan, J. Comparative metagenomics reveals the microbial diversity and metabolic potentials in the sediments and surrounding seawaters of Qinhuangdao mariculture area. PloS one 2020, 15, e0234128. https://doi.org/10.1371/journal.pone.0234128. [CrossRef]
- Oehmen, A.; Yuan, Z.; Blackall, L.L.; Keller, J. Comparison of acetate and propionate uptake by polyphosphate accumulating organisms and glycogen accumulating organisms. Biotechnol. Bioeng. 2005, 91, 162–168. [CrossRef]
Figure 1.
Taxonomic composition of the SBR microbial community.
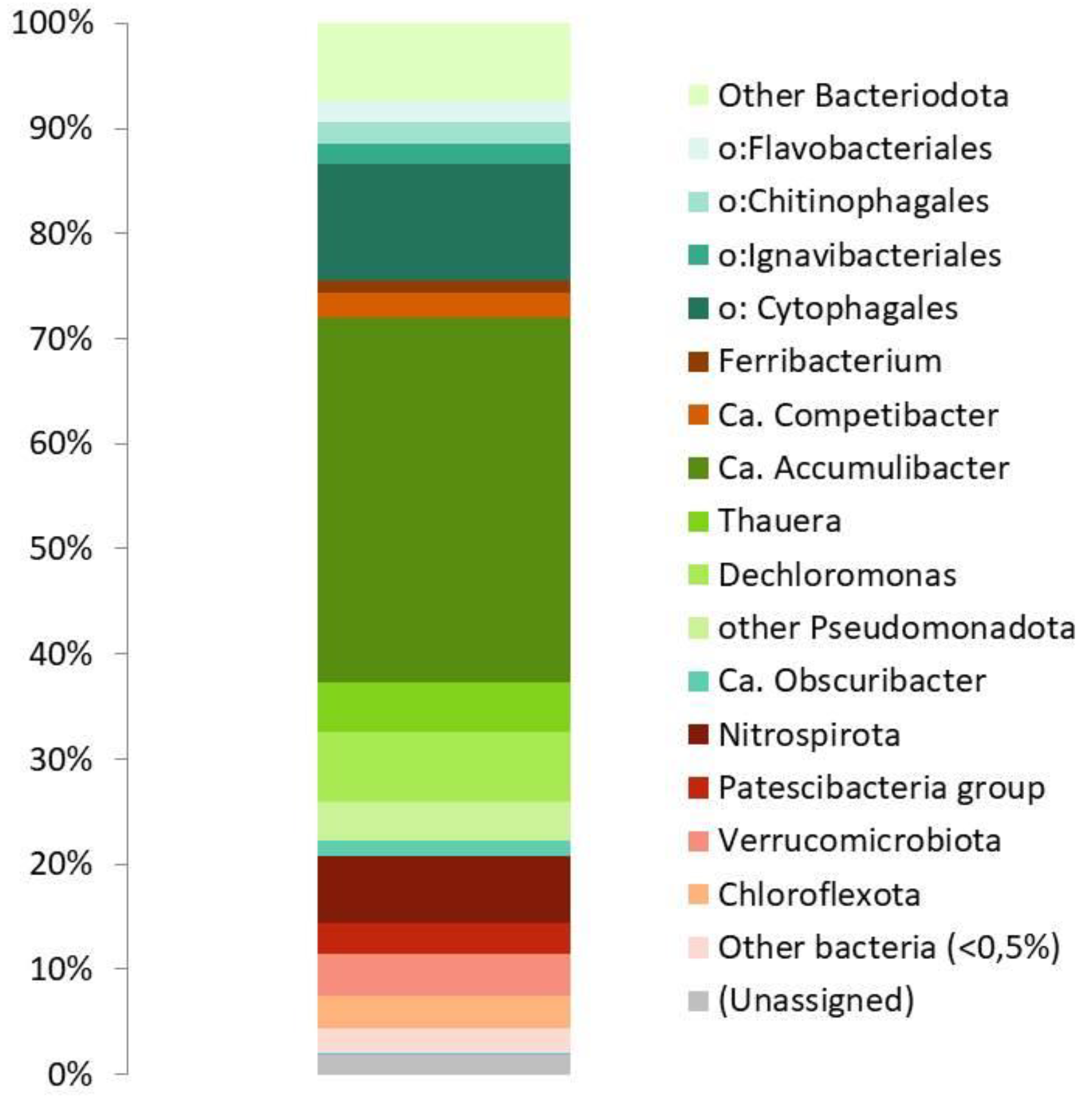
Figure 2.
OUR of the PAO-enriched culture after addition of different substrates. OUR after addition of acetate was taken as 100%.
Figure 2.
OUR of the PAO-enriched culture after addition of different substrates. OUR after addition of acetate was taken as 100%.
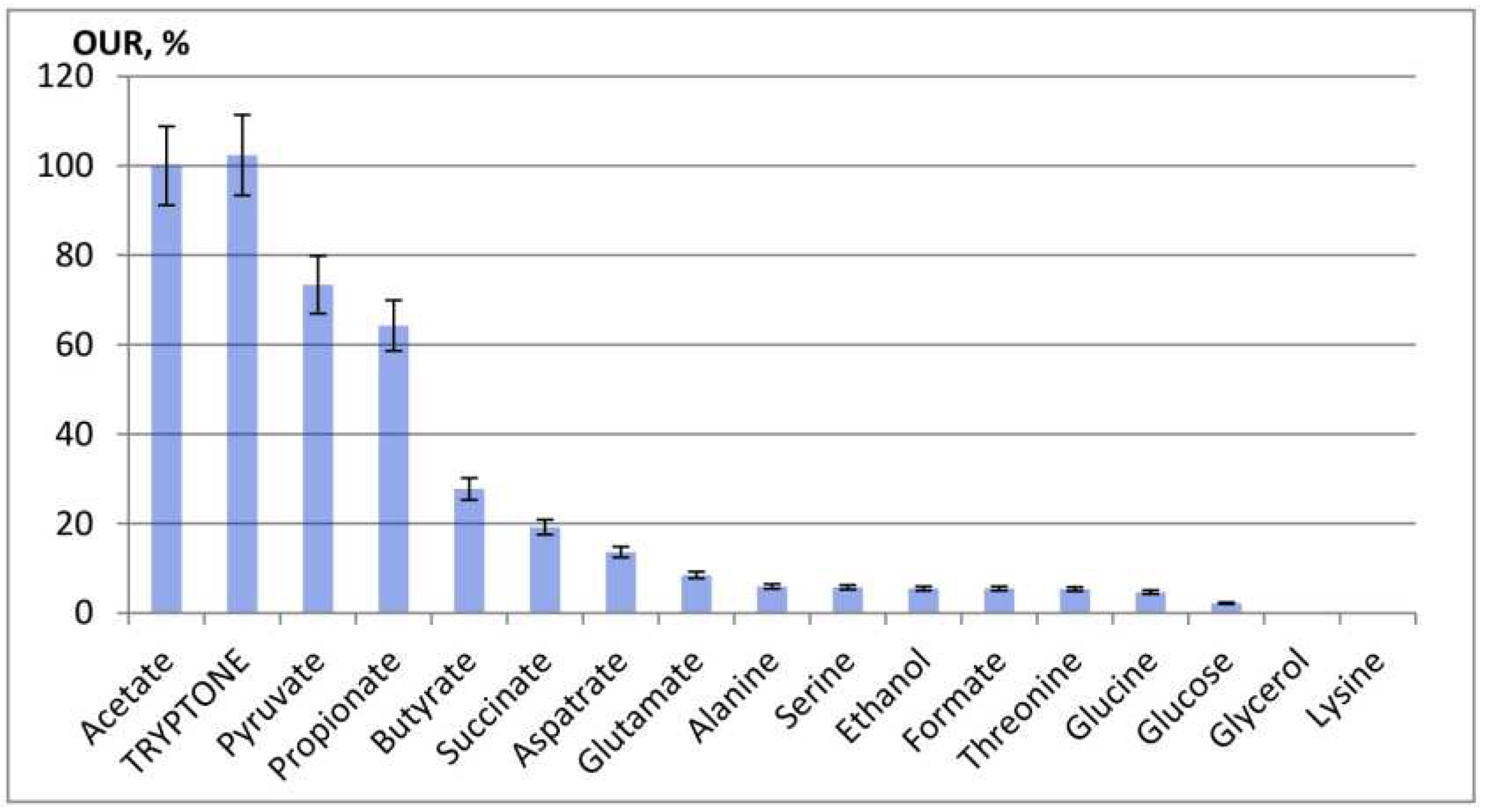
Figure 3.
Phosphate dynamics during the anaerobic/aerobic cycle after substrate addition: substrates of groups 1 (a), 2 (b), and 3 (c). Broken lines indicate shifts from the anaerobic to the aerobic phase.
Figure 3.
Phosphate dynamics during the anaerobic/aerobic cycle after substrate addition: substrates of groups 1 (a), 2 (b), and 3 (c). Broken lines indicate shifts from the anaerobic to the aerobic phase.
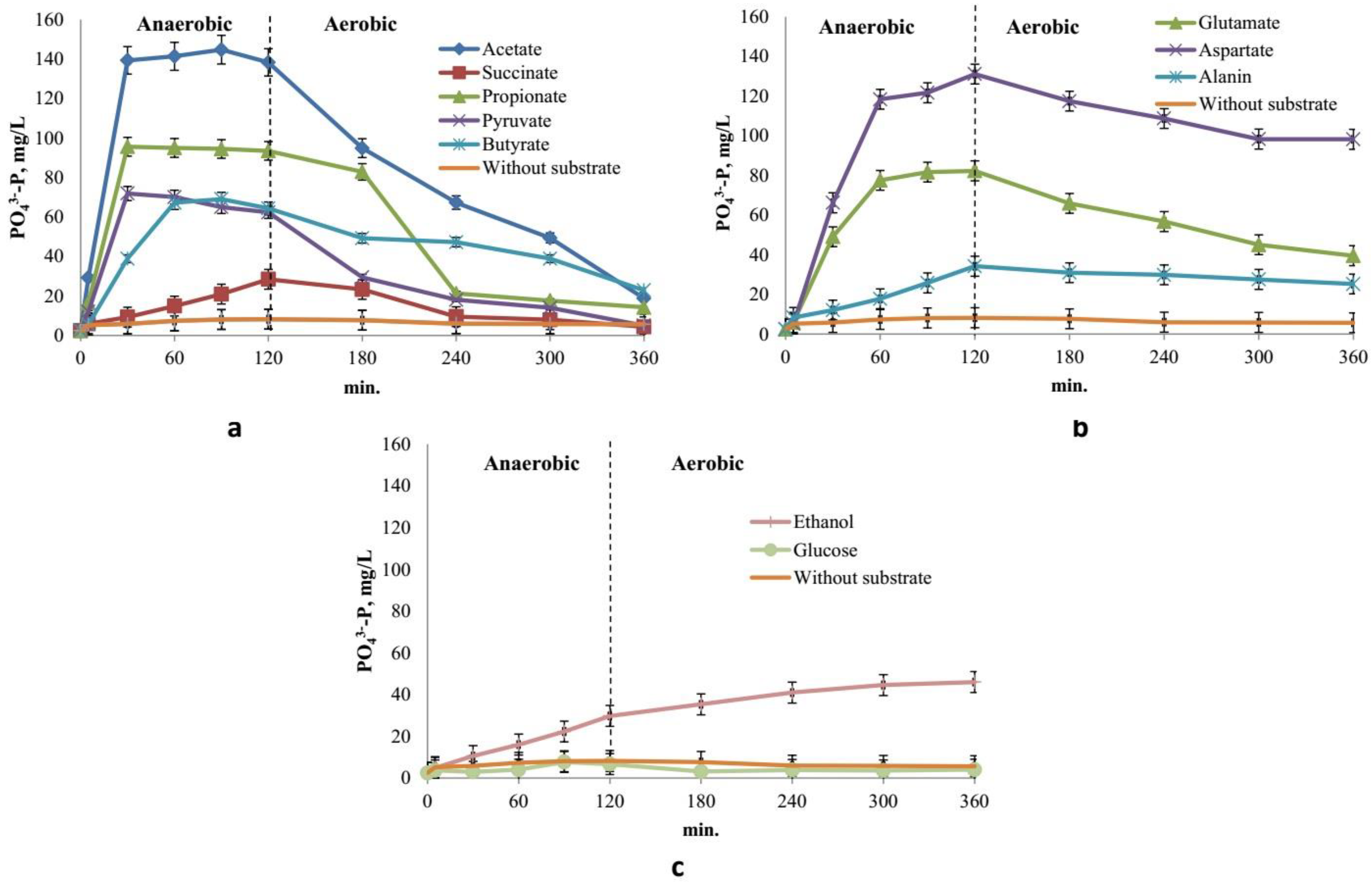
Figure 4.
Dynamics of substrate consumption during the anaerobic/aerobic cycle. The broken line indicates the shift from the anaerobic phase to the aerobic one.
Figure 4.
Dynamics of substrate consumption during the anaerobic/aerobic cycle. The broken line indicates the shift from the anaerobic phase to the aerobic one.
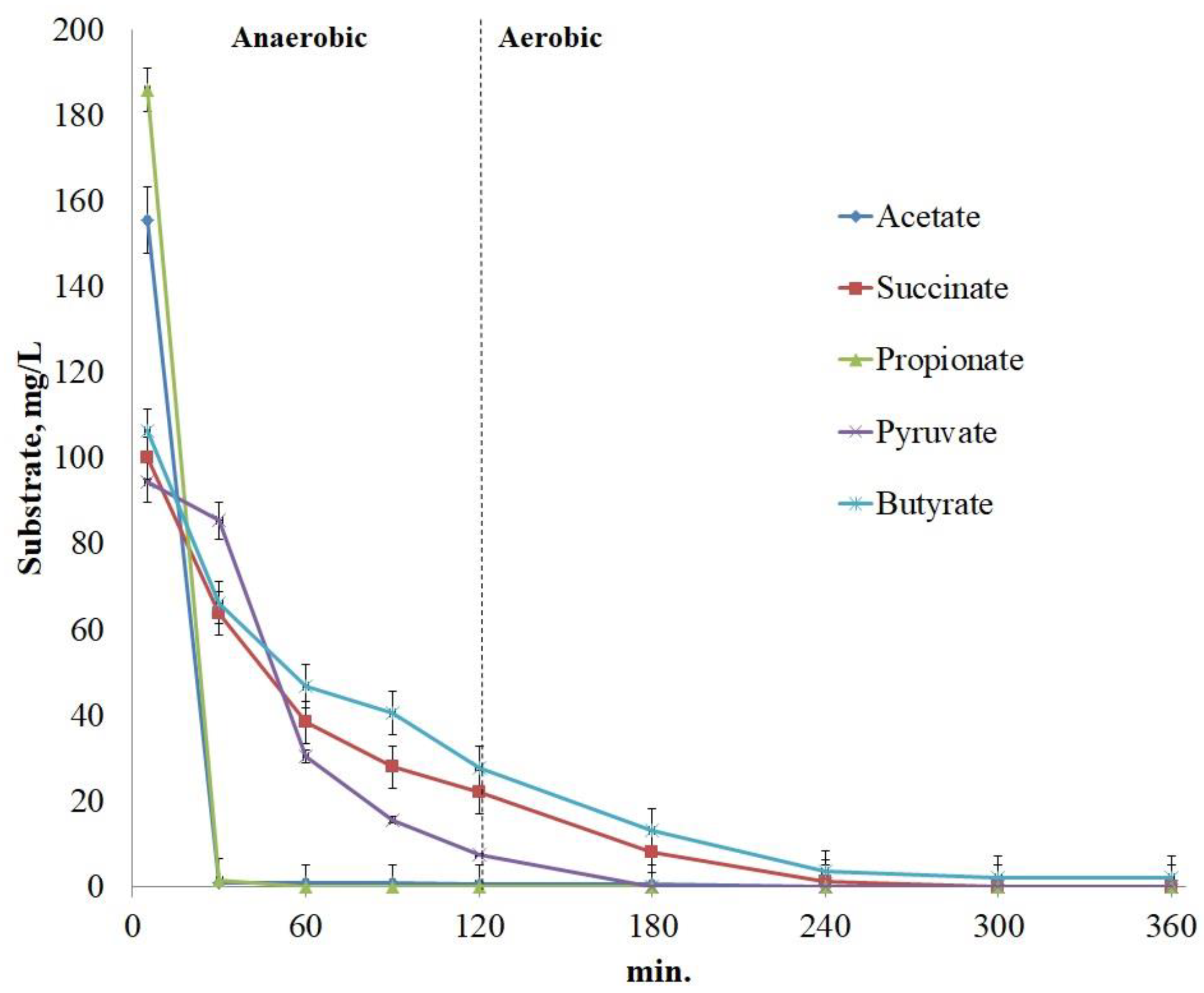
Figure 5.
Relationship between OUR and phosphates uptake during the aerobic phase of the anaerobic/aerobic cycle for different substrates: glucose (1), alanine (2), glutamate (3), aspartate (4), succinate (5), butyrate (6), propionate (7), pyruvate (8), and acetate (9).
Figure 5.
Relationship between OUR and phosphates uptake during the aerobic phase of the anaerobic/aerobic cycle for different substrates: glucose (1), alanine (2), glutamate (3), aspartate (4), succinate (5), butyrate (6), propionate (7), pyruvate (8), and acetate (9).
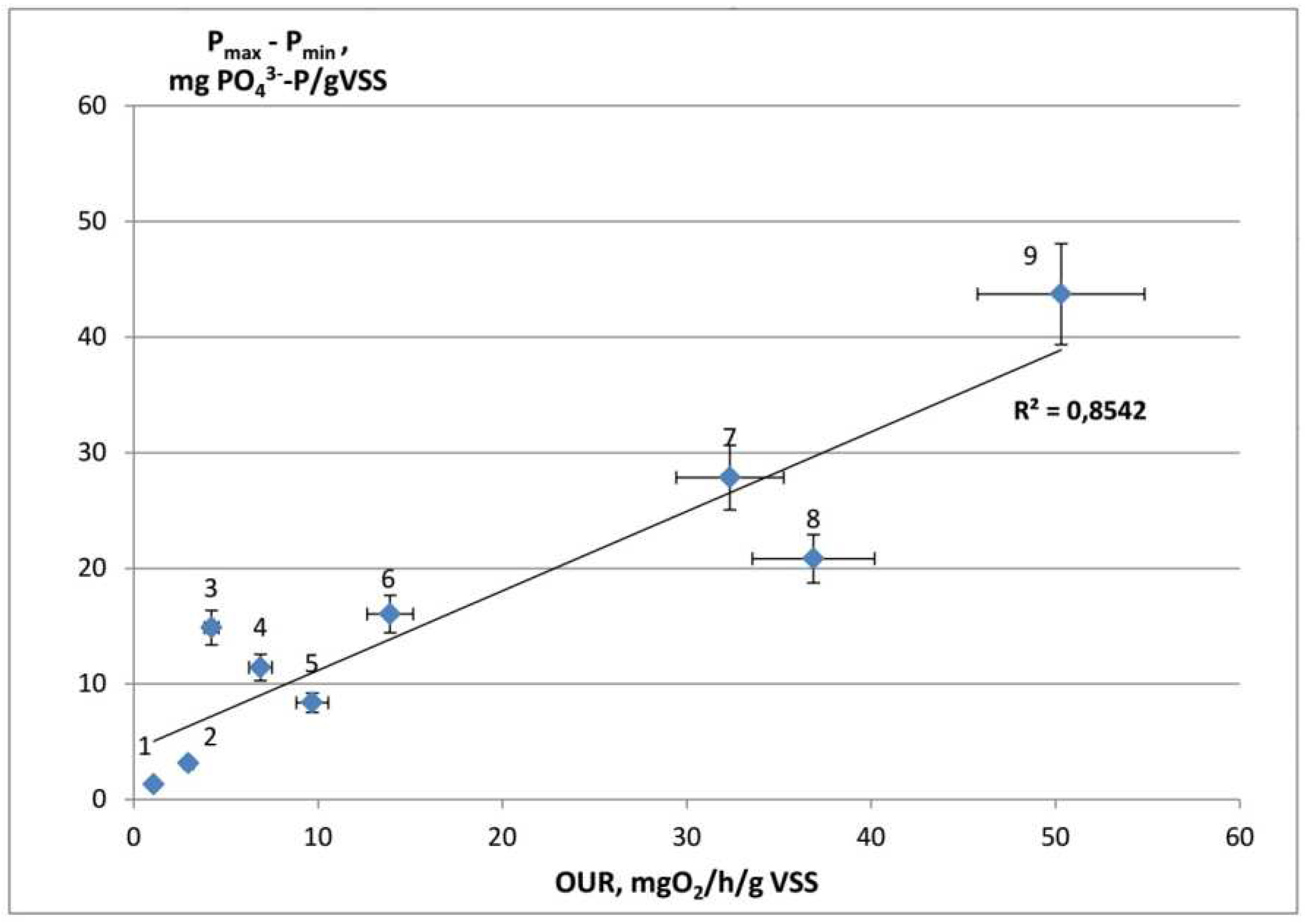

Table 1.
OUR and parameters of the phosphate release/uptake in the aerobic and anaerobic phases of the cycle
Table 1.
OUR and parameters of the phosphate release/uptake in the aerobic and anaerobic phases of the cycle
| Substrate | OUR, mg O2/(g TSS h) |
Anaerobic phase | Aerobic phase | ||
|---|---|---|---|---|---|
| Рmax, mg РО43--P/L |
P-release/ substrate- uptake, P-mol/S-mol |
Average P release rate in the first 30 min, mg РО43--P/(L h) |
Рmax - Рmin, mg РО43--P/L |
||
| Acetate | 32,2 | 144,7 | 1,4 | 273,6 | 125,9 |
| Pyruvate | 23,6 | 65,0 | 1,7 | 138,8 | 60 |
| Propionate | 20,7 | 94,5 | 1,6 | 186,2 | 80,2 |
| Butyrate | 8,9 | 69,1 | 2,1 | 72,8 | 46,2 |
| Succinate 1 | 6,2 | 28,5 | 1,3 | 13,7 | 24,1 |
| D,L-Aspartate | 4,4 | 131,1 | 2,0 2 | 127,3 | 32,9 |
| L- Glutamate | 2,7 | 82,3 | 2,1 2 | 93,1 | 42,8 |
| D,L-Alanine | 1,9 | 34,2 | 0,3 2 | 19,2 | 9 |
| D-Glucose | 1,1 | 7,7 | - | - | 3,7 |
1 added 100 mg COD/L. 2 calculated assuming complete substrate consumption in the anaerobic phase.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



