Submitted:
24 September 2023
Posted:
25 September 2023
You are already at the latest version
Abstract
Powdery mildew disease, caused by Sphaerotheca fusca, is a major disease affecting cucumbers cultivated in greenhouses. This study was conducted to find defense genes induced by DL-3-amino-butyric acid (BABA) and powdery mildew in cucumber. The 2000 and 5000 mg/L BABA treated cucumber exhibited disease severity of 25% and 5%, respectively, whereas the water treated cucumber leaves were severely infected, with a disease severity of 90%. BABA did not effect spore germination of the powdery mildew pathogen,. showing BABA is not antifungal agent against the pathogen. In quantitative real-time PCR analysis, BABA treated cucumber upregulated transcriptional level of the defense genes CsPAL, CsPR3,CsPR1, CsLOX1, CsLOX23, Cs LecRK6.1, CsWRKY20, and Cupi4 in cucumber to maximum levels at 48 h, whereas CsLecRK6.1 reached maximum expression after 24 h and futher salicylic acid (SA) levels were significantly increased in BABA-treated cucumber plants. In addition, the infected cucumber with powdery mildew enhanced the expression levels from1.6 to 47.3 fold of the defense genes PAL, PR3, PR1, Lox1, Lox 23, LecRK6.1, WRKY20, and Cupi4 compared to heathy cucumber. The results suggests the BABA induced defense response is associated with SA signal pathway-dependent systemic acquired resistance (SAR) in cucumber, which is involved in plant resistance mechanisms.
Keywords:
cucumber
; powdery mildew
; infection
; DL-3-amino-butyric acid
; defense gene
1. Introduction
Cucumber powdery mildew (PM), caused by Sphaerotheca fusca, is a serious airborne disease affecting cucumbers. The disease is very difficult to manage because the conidia can attach to host plants, after which germination begins. One generation is completed in 5–6 days, and PM spreads very quickly among plants [1]. Particularly, the pathogen causes severe damage to domestically cultivated vegetables because the pathogen density increases during continuous cultivation in growing facilities. Although disease control has mainly relied on chemical fungicides, this approach has resulted in the emergence of drug-resistant pathogens. Alternatively, resistant plants can be developed as an effective, environmentally friendly, and economical method for controlling PM.
Plants resist pathogenic bacteria and fungi to protect themselves from diseases. Induced resistance is a physiological “state of enhanced defensive capacity” elicited by specific environmental stimuli, enabling a plant to defend against subsequent biotic challenges [2]. Defense-related genes have been well-characterized in different plant hosts during infection with pathogenic and non-pathogenic pathogens and are useful for evaluating the molecular mechanisms of disease resistance [3,4]. A previous study showed that five defense genes (CsWRKY20, CsLecRK6.1, PR3, PR1-1a and LOX1) were expressed in cucumber leaves after inoculation with Phytophthora melonis [5]. Quantitative real- time polymerase chain reaction (qRT-PCR) showed that non-pathogenic Fusarium oxysporium CS-20 induced the expression of the defense genes PR3, LOX1, and PAL1 in pre-inoculated cucumber roots, whereas the pathogenic fungus-mediated defense response was regulated by PR1 and PR3 [4]. The defense gene LecRK-6.1 is also associated with resistance to PM in wheat, and the salicylic acid (SA) pathway contributes to enhanced resistance to virulent PM fungi [6].
The chemical elicitor DL-3-amino-butyric acid (BABA) induces a systemic defense response against various plant pathogens [7]. BABA enhances resistance to Botrytis cinerea and Pseudomonas syringae in Arabidopsis and induces transcriptional expression of the PR-1 gene, which is correlated with the SA-dependent systemic acquired resistance (SAR) system. Additionally, BABA treatment suppresses Phytophthora blight and induces defense gene expression in pepper [8,9]. The cucumber PM pathogen (S. fuliginea) and defense signal chemicals SA, abscisic acid, and methyl jasmonate were treated on cucumber leaves and it was revealed 23 C-lipoxygenase-related genes (CsLOXs) in the cucumber genome were up- or down regulated in response to the signal chemicals [10]. SA and jasmonic acid (JA) transduce signals in the defense response to SAR and induce systemic resistance (ISR). The expression of defense genes in the SA and JA pathways is induced in BABA-treated plants [2,10,11]. It has been known the PR1 and PR3 genes encoding pathogenicity related protein (PR) are related to the SA signalling pathway[2,4]. .In addition, It was ISR is usually triggered by nonpathogenic micro-organisms and the realated defense genes NPR1, and phenylalanine ammonia lyase (PAL), encoded by the PAL1 are activated by JA/ethylene signaling pathway[4]. The ethylene and lipoxygenase (LOX), encoded by the LOX1 gene, is the first enzyme in the biosynthesis pathway of JA[4,7]. In addition, biochemical and enzymatic defense analyses suggested that BABA treatment significantly regulated catalase, guaiacol peroxidase, phenylalanine ammonia-lyase (PAL) activity, H2O2, and lignin content [12]. Nevertheless, expressional regulations of the defense-related genes and and chemical signal response by PM pathogen infection and BABA are still remained to be elucidated in cucumber plants.
This study was conducted to find the expression fasions of defense-related genes in cucumber by BABA treatment and PM infection and futher to elucidate whether BABA-treated cucumber plants produce chemical signals related to resistance responses to SAR or ISR.
2. Materials and Methods
2.1. Host Plant and Infection
The cucumber variety , Baeckdadagi (Dongwon Nong San seed Co.,Ltd.), Korea) was used as the host plant for PM. Seeds were sown in 48-pore trays pore filled with peat, and the seedlings were transplanted into plastic pots (12 cm) and grown in a greenhouse. The inoculum was obtained from fresh sporulation of naturally infected cucumber leaves. The conidia were isolated from the PM lesions using distilled water containing 0.0003% Tween 20, and a hemocytometer was used to prepare a suspension of 4 × 106 conidia/mL. The conidia suspension was sprayed onto cucumber leaves, and the cucumber plants were maintained in a greenhouse to monitor for disease occurrence. The 2,000 and 5,000 mg/L BABA were used as treatments, and didtilled water (DW) was used as control. The infection degree was evaluated based on a 0–5 scale where 0 = no disease symptoms; 1 = less than 5% PM symptoms, 2 = 5.1–20% PM symptoms, 3 = 20.1–40% of plant disease, 4 = more than 40–60% diseased plant, and 5 = more than 61% of PM symptoms. Disease severity was calculated by reported method [12] as follows: R = [ Σ (a × b)/N × K] × 100% where R –disease severity, a –number of infected leaves rated, b –numerical value of each grade, N –total number of examined plants, K –the highest degree of infection in scale. Ten plants were used for each treatment, with three replicates per treatment.
2.2. Spore Germination
The fleshed conidia were isolated from cucumber PM lesions, spread onto 2% agar-coated glass slides, and covered with a cover glass. The agar blocks were incubated under fluorescent light 48 h at 25℃. The germination rate was measured for randomly selected conidia after two days.
2.3. qRT-PCR
Primers targeting defense genes in cucumber were used in qRT-PCR (Table 1). Total RNA was extracted from infected and BABA-treated cucumber eaves and control treatments using Favorgen kit (Favorgen, Seoul, Austria) according to the manufacturer’s instructions. After treating the RNA smples with RNase-free DNase (Invitrogen mbion DNase I, Seoul, USA orea), 1 µg DNA-free RNA was used for first-strand cDNA synthesis with Reverse transcriptase reagent HelixCript™ Easy (NANOHELIX, Taejeon, korea).The reaction mixture (20 μL) consisted of 10μL of Dyne qPCR 2X PreMIX (DyneBio, Seongnam, Korea), 1000ng/ μL of cDNA, and 10 pmol of each primer. Activation, followed by 40 cycles of 95°C for 15 s, 58°C for 30 s, and 72°C for 30 s. Relative gene expression was normalized to the level of mRNA using the in a LightCycler® 96 System. (Roche, Basel, Switzerland).
2.4. Extraction and Measurement of SA from Cucumber Leaves
The 2,000 and 5,000 mg/L BABA solution was foliarly sprayed on cucumber leaves. After 72 hours, the leaves were collected and ground with liquid nitrogen. 90 g of silicon dioxide and 30 ml of 90% methanol were added to 10 g of the ground leaves, and the supernatant was collected using a mortar bowl. The methanolic extracts were concentrated using a vacuum evaporator. The residues were resuspended in a mixture of 1mL of 5% trichloroacetic acid (Sigma-Aldrich, St. Louis, MO, USA) and 10 mL of 99.8% methanol (w/v). The volume of each extract was adjusted with distilled water to 50 mL and centrifuged at 8,000 ×g for 10 min. The supernatants of the extracts were used to measure the SA content with a liquid chromatograph mass spectrometer (LCMS8050; Shimadzu, Kyoto, Japan) on a Kinetex c18 column (2.6 μm, 100 × 2.1 mm; Phenomenex, Torrance, CA, USA). The calibration curves were linear (R2 > 0.99) over a concentration range of 10–500 ng/mL with acceptable accuracy and precision.
2.5. Statistical Analysis
Data were analyzed using SIGMA PLOT 11.0 (http://www.sigmaplot.com). The significance of the data was evaluated using analysis of variance, followed by Tukey’s multiple range test (P < 0.05).
3. Results
3.1. BABA Inhibits Cucumber PM Disease
To investigate whether BABA protects against cucumber PM, BABA was applied at 2,000, and 5,000 mg/L, after which the cucumber leaves were infected with PM spores. The protective effects were assessed using negative and positive controls. As shown in (Figure 1A) 2000 and 5000 mg/L BABA treated cucumber exhibited disease severity of 25% and 5%, respectively, whereas the water treated cucumber leaves were severely infected, with a disease severity of 90%. In BABA-treated cucumbers, the PM was suppressed, and symptoms were largely absent, whereas in water-treated cucumber leaves, typical PM infection was observed on the surface of the cucumber leaves (Figure 1B). In addition, the inhibitory effect of BABA on PM spore germination was examined to determine whether BABA directly inhibits PM. PM spores treated with 2000 and 5000 mg/L BABA and water showed germination rates of 45%, 42%, and 40%, respectively (Figure 2A). This result suggests that BABA is not fungitoxic towards PM spores and may induce disease resistance, as reported previously [2,11].
3.2. Defense Genes in Cucumber Were Upregulated by BABA and PM Infection
After treating cucumber plants with 2000 ppm BABA, the expression levels of defense genes were quantified using qRT-PCR at 0, 24, 48, and 72 h. (Figure 3) The expression level of defense genes in cucumber plants began increasing at 24 h after BABA treatment; except for the LecRK6.1 gene, which showed the highest expression level at 24 h, the other defense genes evaluated showed the highest expression levels at 48 h, although these levels were decreased at 72 h. PAL, PR3, PR1, Lox1, Lox 23, LecRK6.1, WRKY20, and Cupi4 were transcriptionally expressed at 2.7, 53.1, 25, 2.7, 2.8, 4.4, 0.6, and 21.6-fold higher levels compared to those in non-treated BABA cucumber plants at 48 h after BABA treatment. In addition, we quantified the relative expression of defense genes in cucumbers infected with PM and treated with BABA. The transcriptional expression level of defense genes in PM-infected and uninfected cucumbers was analyzed using qRT-PCR with a primer set targeting the cucumber disease resistance genes shown in Table 1. The transcriptional expression levels of the defense genes PAL, PR3, PR1, Lox1, Lox23, LecRK6.1, WRKY20, and Cupi4 in infected cucumber were increased by 3.4 , 24.2, 7.8, 2.5, 3.9, 29.5, 1.6, and 47.3-fold compared to those in uninfected cucumber (Figure 4).
3.3. BABA Iduces SA Accumulation in Cucumber
The defense genes PAL, PR1, and PR3 were upregulated at the transcriptional level. Defense genes are triggered through an SA-dependent signaling pathway, reflecting that BABA treatment induces SA accumulation in cucumbers [2,13]. To estimate whether BABA-treated cucumbers accumulated SA, the total SA content in BABA-treated cucumbers was determined using liquid chromatography-mass spectrometry. SA levels were increased by 3.8–14-fold in 2,000. and 5,000 mg/L BABA-treated leaves compared with those in water-treated leaves (Figure 5). Another type of resistance, induced systemic resistance (SIR), is triggered by the JA/ET signaling pathway. We confirmed that JA accumulated in BABA-treated cucumbers; however, JA was not detected using liquid chromatography-mass spectrometry (Data not shown). Thus, the result suggests BABA is associated with the SA signaling pathway-dependent SAR response in cucumber.
4. Discussion
Sphaerotheca fuliginea, the causal agent of PM disease, is an obligate parasite that cannot be cultured in artificial media and has a broad host range, particularly in cucurbit crops; the disease is problematic worldwide [14]. In the form of cleistothecia containing a few ascuses, S. fuliginea spends the winter on the remains of diseased plants and is the primary source of infection [14]. It spreads rapidly in cucumber cultivation facilities and is extremely difficult to control, leading to large amounts of damage. Although chemical controls have been effectively applied, their overuse can lead to the emergence of drug-resistant strains and environmental contamination problems. Therefore, disease-resistant plants should be developed. Selecting useful genes identified through searches for disease-resistance genes in cucumber plants can provide basic information for producing disease-resistant varieties. Plants resist pathogenic bacteria and fungi to protect themselves from diseases. Induced resistance is a physiological “state of enhanced defensive capacity” elicited by specific environmental stimuli, whereby a plant’s innate defenses are potentiated against subsequent biotic challenges [15].
In the present study, we assessed the expression patterns of defense genes in cucumber plants treated with BABA. BABA is a non-protein amino acid that rarely occurs in nature and is involved in a wide range of activities as an inducer of resistance to a broad spectrum of plant pathogens such as viruses, bacteria, fungi, and nematodes [11]. Previously, different concentrations of BABA (0.5, 2, and 4 mM) were used to control spinach PM, and 4 mM BABA was the most effective against PM [12]. Additionally, 10–100 mM BABA was used to control lettuce downy mildew, with 10 mM BABA shown to be effective for controlling the disease [16]. We examined the protective effect of different BABA concentrations against PM. It was revealed that more than 2000 mg/L BABA inhibited cucumber PM, indicating that a higher concentration of BABA than that reported previously is required to control cucumber PM.
. The expression of the defense genes PR3, PR1-1a, CsWRKY20, CsLecRK6.1 and Cupi4 was increased when cucumbers were artificially inoculated with P. melonis, which causes damping-off disease [2]. In addition, Fusarium oxysporum-infected cucumber shows strongly enhanced expression of PR3, LOX1, and NPR1 [4], and it was recently reported that phenolic compounds and flavonoid-related metabolites accumulate in the cucumber PM-resistant cultivar BK following infection with S. fuliginea [17]. However, the expression patterns of different defense genes in cucumbers caused by PM infection have not been defined. In this study, the expression levels of the defense genes PAL, PR3, PR1, Lox1, Lox 23, LecRK6.1, WRKY20 and Cupi4 were significantly increased by 1.6 - 47.3-fold under biotic stress by PM infection. This is the first study to provide information on the transcriptional expression levels of different defense genes in both BABA-treated and PM-infected cucumber plants. The defense genes induced in cucumber plants showed expression values similar to those previously reported for other plant pathogens. BABA did not directly affect the germination of PM; therefore, it did not exert a direct antifungal action against the PM pathogen, suggesting that BABA participates in defense mechanisms by inducing disease resistance in plants.
The receptors in the plant cell membrane recognize pathogen-associated molecular patterns (PAMP)and activate the plant immune system, including induction of disease resistance-related gene expression [2]. In this study, the upregulated expression of various defense genes in PM-infected cucumbers may be attributed to the triggering of disease resistance mechanisms related to the innate immune system in the cucumber genome. PR1, PR3, and lipoxygenases (LOX) are strongly induced when plants respond to pathogen infection [2,18]. Pathogenesis-related proteins (PRs) are among the most commonly induced proteins in plant defense mechanisms and play important roles in plant immunity [17]. PR1 strongly inhibits Phytophthora infestans in potatoes [19], and PR1 expression in plant cells is a useful molecular marker for SA-dependent SAR signaling pathway and may be related to its putative direct antimicrobial action [20]. Furthermore, PR3 (endochitinase), which hydrolyzes chitin components by biotic and abiotic stress, is often used as a marker for SA-dependent SAR signaling [2,8,21]. Chitinase degrades the fungal cell wall and degradable products, chitin oligomers, which can serve as elicitors of plant disease resistance [21]. Fusarium wilt pathogen (Fusarium oysporium f.sp. cucumerium)-responsive chitinase genes were profiled using comparative transcriptome analysis, revealing six genes that were significantly upregulated after pathogen infection. Additionally, LOX pathways are crucial for lipid peroxidation during defense responses to infection and inhibit pathogen growth [22]. LOX genes are upregulated in cucumber leaves following infection with the PM pathogens S. fuliginea, SA, JA, and ABA [10]. PAL generates cinnamic acid by catalyzing the non-oxidative deamination of phenylalanine to trans-cinnamate. Given that phenylpropanoids derived from cinnamic acid serve as precursors for a range of phenolic compounds, they play a vital role in the biosynthesis of SA, which is an essential signal in the SAR response of plants [23].
In this study, LecRK6.1, showed the highest expression level among the defense genes in PM-infected and BABA-treated cucumber leaves. Interestingly, the highest expression level of LecRK6.1 was observed at 24 h after BABA treatment rather than at 48 h after BABA treatment. The LecRK family of genes plays important roles under biotic and abiotic stresses in plants by inducing plant innate immunity [1,24]. In cucumbers, CsLecRK6.1, is induced by P. melonis and Phytophthora capsici in resistant cultivars [2,24]. In cucumber plants, the expression of CsLecRK6.1 was increased by P. melonis and P. capsici. LecRK-V confers broad resistance to wheat PM through an SA pathway-dependent disease resistance mechanism [24]. Resistance is activated in plant cells by infection with plant pathogens, which is determined by an intricate network of signaling pathways involving innate immunity and resistance machinery [2]. WRKY is a plant-specific transcription factor with an important role in plant defense. WRKY enhances resistance to plant pathogens by increasing the expression levels of PR genes and inducing the accumulation of phytoalexins [25,26]. In addition, WRKY transcription factors positively regulate plant immunity to pathogens associated with H2O2 production, and a hypersensitive response mimics cell death and activation of phytohormone-mediated signaling pathways[27]. However, we found that the transcriptional expression of WRKY20 was 1.6-fold higher in PM cucumber leaves, whereas its expression was downregulated in BABA-treated cucumber leaves (data not shown). These results suggest that cucumber WRKY20 gene expression was not sufficiently induced by BABA or PM infection. Cupi4, which is homologous to a pathogen-inducible protein, has been isolated from cucumbers infected with bacterial pathogens and characterized as a chemical inducer of SAR, such as SA, pathogenic bacteria, viruses and fungi treated cucumbers led to upregulation of Cupi4 transcript [28]. It was predicted that the gene product was associated with SAR. In this study, expression of the Cupi4 transcript was increased by 21.6–47.3-fold at 48 h in BABA-treated and PM-infected cucumbers, respectively. PAL, PR3, PR1, Lox1, Lox 23, LecRK6.1, WRKY20, and Cupi4 are associated with SA signaling pathway-dependent SAR. They showed relatively high expression levels in BABA-treated cucumber plants. Accordingly, we examined whether SA accumulation occurred in BABA-treated cucumbers. As expected, 14.1 ng/mL and 3.84 ng/mL SA was detected in the BABA 5,000 and 2000 mg/L-treated cucumbers, whereas 0.92 ㎍/mL SA was detected in untreated cucumbers. This result indicates that BABA treatment led to the accumulation of SA, suggesting that SA is involved in the SAR mechanism. The results showed that BABA may be used as an inducer of SAR against cucumber PM and that BABA and PM infection-induced defense genes can be used as target genes for developing PM-resistant plants. Although some studies have been conducted to induce the expression of resistance genes in cucumbers following infection with various plant pathogens, few studies have focused on the expression of resistance genes targeting cucumber PM. We focused on cucumber PM and the expression of defense genes induced by BABA treatment, targeting previously reported genes in cucumbers. In future, additional disease-resistance genes related to PM can be identified by profiling genes expressed during BABA and PM infection at the genome level in cucumber plants.
Conclusions
We found that the transcriptional expression of defense genes in cucumbers was induced by PM infection and BABA treatment. BABA had a protective effect of more than 80% against PM disease and did not affect PM spore germination, indicating that it did not directly inhibit pathogens. In BABA-treated and PM-infected cucumbers, the transcripts of the defense genes PAL, PR3, PR1, LOX1, LOX23, LecRK6.1, WRKY20 and Cupi4 were significantly increased, demonstrating their association with SA-dependent SAR resistance. This study provides a foundation for functional studies of cucumber plant defense responses and signaling pathways against PM.
Acknowledgments
This research was supported by the Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry, and Fisheries (IPET; Grant No. 321103-3)
References
- Reuveni, M.; Agapov, V.; Reuveni, R.; et al. A foliar spray of micronutrient solutions induces local and systemic protection against powdery mildew (Sphaerotheca fuliginia) in cucumber plants. Eur. J. Plant Pathol. 1997, 103, 581-588. [CrossRef]
- Andersen, E.J.; Ali, S.; Byamukama, E.; Yen, Y.; Nepal, M.P.; et al. Disease resistance mechanisms in plants. Genes 2018, 9, 339. [CrossRef]
- Knoth, C.; Ringler, J.; Dangl, J.L.; Eulgem, T.; et al. Arabidopsis WRKY70 is required for full RPP4-mediated disease resistance and basal defense against Hyaloperonospora parasitica. Molecular plant-microbe interactions, 2007, 20, 120-128. [CrossRef]
- Pu, X.; Xie, B.; Li, P.; Mao, Z.; Ling, J.; Shen, H.; Lin, B.; et al. Analysis of the defence-related mechanism in cucumber seedlings in relation to root colonization by nonpathogenic Fusarium oxysporum CS-20. FEMS Microbiol. Lett., 2014, 355, 142-151. [CrossRef]
- Hashemi, L.; Golparvar, A. R.; Nasr-Esfahani, M.; Golabadi, M.; et al. Expression analysis of defense-related genes in cucumber (Cucumis sativus L.) against Phytophthora melonis. Mol. Biol. Rep., 2020, 47, 4933-4944. [CrossRef]
- Wang, Z.; Cheng, J.; Fan, A.; Zhao, J.; Yu, Z.; Li, Y.; Wang, X.; et al. LecRK-V, an L-type lectin receptor kinase in Haynaldia villosa, plays positive role in resistance to wheat powdery mildew. Plant Biotechnol. J., 2018, 16, 50-62. [CrossRef]
- Oh, S. K.; Jang, H. A.; Kim, J.; Choi, D.; Park, Y. I.; Kwon, S. Y.; et al. Expression of cucumber LOX genes in response to powdery mildew and defense-related signal molecules. Can. J. Plant Sci., 2014, 94, 845-850. [CrossRef]
- Kang, D. S.; Min, K. J.; Kwak, A. M.; Lee, S. Y.;Kang, H. W.; et al. Defense response and suppression of Phytophthora blight disease of pepper by water extract from spent mushroom substrate of Lentinula edodes. Plant Pathol. J., 2017, 33, 264. [CrossRef]
- Stamler, R. A.; Holguin, O.; Dungan, B.; Schaub, T.; Sanogo, S.; Goldberg, N.; Randall, J. J. ; et al. BABA and Phytophthora nicotianae induce resistance to Phytophthora capsici in chile pepper (Capsicum annuum).Plos one,2015, 10, e0128327. [CrossRef]
- Oh, S. K.; Jang, H. A.; Kim, J.; Choi, D.; Park, Y. I.; Kwon, S. Y.; et al. Expression of cucumber LOX genes in response to powdery mildew and defense-related signal molecules. Canadian Journal of Plant Science, 2014, 94, 845-850. [CrossRef]
- Jakab, G.; Cottier, V.; Toquin, V.; Rigoli, G.; Zimmerli, L.; Métraux, J. P.; Mauch-Mani, B.; et al. β-Aminobutyric acid-induced resistance in plants. European Journal of plant pathology, 2001, 107, 29-37. [CrossRef]
- Zeighaminejad, R.; Sharifi-Sirchi, G. R.; Mohammadi, H.; & Aminai, M. M.; et al. Induction of resistance against powdery mildew by Beta aminobutyric acid in squash.Journal of Applied Botany and Food Quality, 2016, 89. [CrossRef]
- Cameron, R. K.; PAIVA, N. L.; LAMB, C. J.; DIXON, R. A.; et al. Accumulation of salicylic acid and PR-1 gene transcripts in relation to the systemic acquired resistance (SAR) response induced by Pseudomonas syringae pv. tomato in Arabidopsis. Physiological and Molecular Plant Pathology, 1999, 55, 121-130. [CrossRef]
- George N. Agrios, Plant Pathology, 5th ed, ELSEVIER Academic Press, Amsterdam, 2005, pp. 448.
- van Loon, L. C.; Rep, M.; Pieterse, C. M.; et al. ignificance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol., 2006, 44, 135-162.
- Pajot, E., Le Corre, D. & Silué, D. Phytogard® and DL-β-amino Butyric Acid (BABA) Induce Resistance to Downy Mildew (Bremia Lactucae) in Lettuce (Lactuca sativa L). European Journal of Plant Pathology 107, 861–869 (2001), 861, 866. [CrossRef]
- Zhang, P.; Zhu, Y., & Zhou, S. et al. (2021). Comparative analysis of powdery mildew resistant and susceptible cultivated cucumber (Cucumis sativus L.) varieties to reveal the metabolic responses to Sphaerotheca fuliginea infection. BMC plant biology, 21(1), 1-13. [CrossRef]
- Kavroulakis, N.; Papadopoulou, K. K.; Ntougias, S.; Zervakis, G. I.; Ehaliotis, C.; et al. Cytological and other aspects of pathogenesis-related gene expression in tomato plants grown on a suppressive compost. Annals of botany, 2006, 98, 555-564. [CrossRef]
- Niderman, T.; Genetet, I.; Bruyere, T.; Gees, R.; Stintzi, A.; Legrand, M.; Mosinger, E.; et al. Pathogenesis-related PR-1 proteins are antifungal (isolation and characterization of three 14-kilodalton proteins of tomato and of a basic PR-1 of tobacco with inhibitory activity against Phytophthora infestans).Plant physiology, 1995, 108, 17-27. [CrossRef]
- Wu, C. T. Expression, regulation and functional roles of β-1, 3-glucanase and chitinase in germinating tomato seeds. University of California, Davis. 2003.
- Rawat, S.; Ali, S.; Mittra, B.; & Grover, A.; et al. Expression analysis of chitinase upon challenge inoculation to Alternaria wounding and defense inducers in Brassica juncea. Biotechnology reports, 2017, 13, 72-79. [CrossRef]
- Yang, X. Y.; Jiang, W. J.; & Yu, H. J.; et al. The expression profiling of the lipoxygenase (LOX) family genes during fruit development, abiotic stress and hormonal treatments in cucumber (Cucumis sativus L.). International journal of molecular sciences, 2012, 13, 2481-2500. [CrossRef]
- Kawatra, Anubhuti & Dhankhar, Rakhi & Mohanty, Aparajita & Gulati, Pooja. Biomedical applications of microbial phenylalanine ammonia lyase: Current status and future prospects. Biochimie. 2020. 177. [CrossRef]
- Wang, Z.; Cheng, J.; Fan, A.; Zhao, J.; Yu, Z.; Li, Y.’ Wang, X; et al. LecRK-V, an L-type lectin receptor kinase in Haynaldia villosa, plays positive role in resistance to wheat powdery mildew. Plant biotechnology journal, 2018, 16, 50-62.
- Wang, J.; Tao, F.; Tian, W.; Guo, Z.; Chen, X.; Xu, X.; Hu, X. The wheat WRKY transcription factors TaWRKY49 and TaWRKY62 confer differential high-temperature seedling-plant resistance to Puccinia striiformis f. sp. tritici. PloS one, 2017,12, e0181963. [CrossRef]
- Wen, F.; Wu, X.; Zhang, L.; Xiao, J.; Li, T.; Jia, M.; et al. Molecular Cloning and Characterization of WRKY12, A Pathogen Induced WRKY Transcription Factor from Akebia trifoliata.Genes,14, 1015.
- Wang, X.; Li, J.; Guo, J.; Qiao, Q.; Guo, X.; & Ma, Y.; et al. The WRKY transcription factor PlWRKY65 enhances the resistance of Paeonia lactiflora (herbaceous peony) to Alternaria tenuissima. orticulture Research, 2020, 7. [CrossRef]
- Phuntumart, Vipa & Marro, Pascal & Metraux, Jean-Pierre & Sticher, Liliane. A novel cucumber gene associated with systemic acquired resistance. Plant Science. 2006. 171. 555-564. [CrossRef]
Figure 1.
Protective effect of DL-3-aminobutyric acid (BABA) against cucumber powdery mildew. A: Powdery mildew disease index and B: diseases index observed at 14 days after inoculation. The value represents the mean disease index ± standard deviation. The experiment was repeated three times. DW, distilled water.
Figure 1.
Protective effect of DL-3-aminobutyric acid (BABA) against cucumber powdery mildew. A: Powdery mildew disease index and B: diseases index observed at 14 days after inoculation. The value represents the mean disease index ± standard deviation. The experiment was repeated three times. DW, distilled water.

Figure 2.
Inhibition rate of spore germination on different concentrations of DL-β-aminobutyric acid against cucumber powdery mildew. A: Spore germination rate following treatment with different BABA concentrations and B: conidial germination of powdery mildew pathogen derived from cucumber.
Figure 2.
Inhibition rate of spore germination on different concentrations of DL-β-aminobutyric acid against cucumber powdery mildew. A: Spore germination rate following treatment with different BABA concentrations and B: conidial germination of powdery mildew pathogen derived from cucumber.
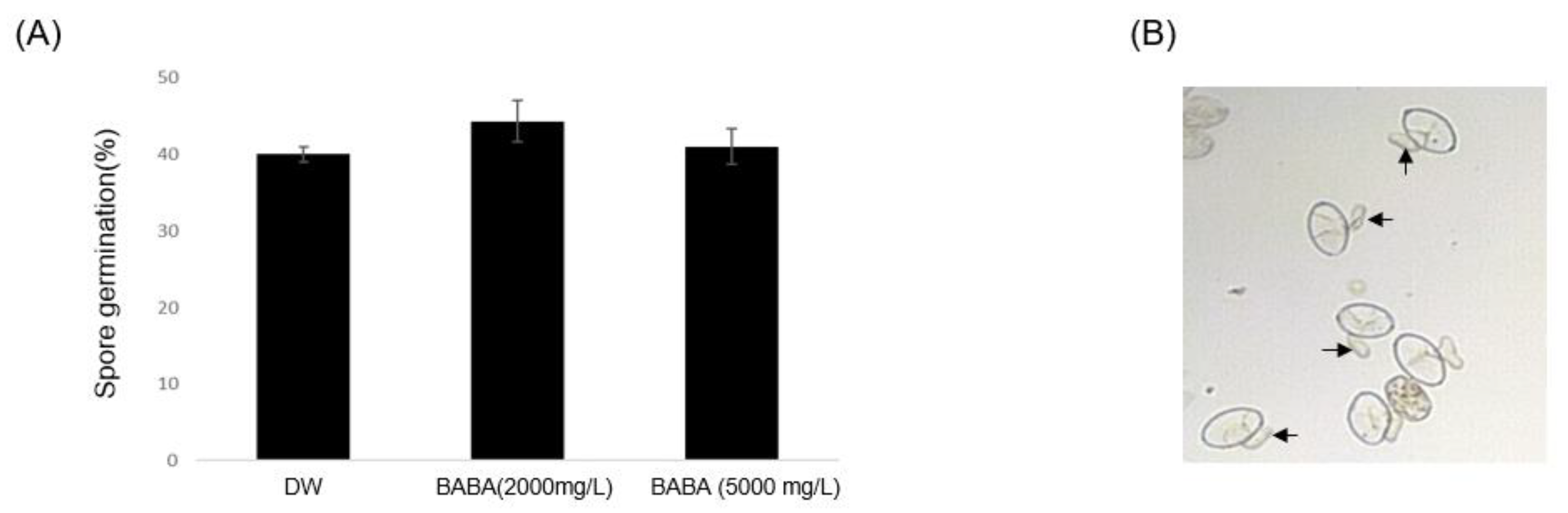
Figure 3.
Expression level of defense genes in cucumber on different hours after treatments of DL-β-aminobutyric acid (BABA). BABA(2000mg/L) was treated on cucumber leaves and different hours (0, 24,48, and 72h) were given after the treatment. Total RNA was extracted from each treated cucumber leaf samples and subjected to qRT-PCR using primers targeting defense genes in Table 1. Each gene expression was normalized to reference gene, CsActin. Expression value is the average of three replications and the bar indicates the standard deviation. Different letters indicate significant differences between treatments (P < 0.05 according to Duncan’s multiple test.
Figure 3.
Expression level of defense genes in cucumber on different hours after treatments of DL-β-aminobutyric acid (BABA). BABA(2000mg/L) was treated on cucumber leaves and different hours (0, 24,48, and 72h) were given after the treatment. Total RNA was extracted from each treated cucumber leaf samples and subjected to qRT-PCR using primers targeting defense genes in Table 1. Each gene expression was normalized to reference gene, CsActin. Expression value is the average of three replications and the bar indicates the standard deviation. Different letters indicate significant differences between treatments (P < 0.05 according to Duncan’s multiple test.
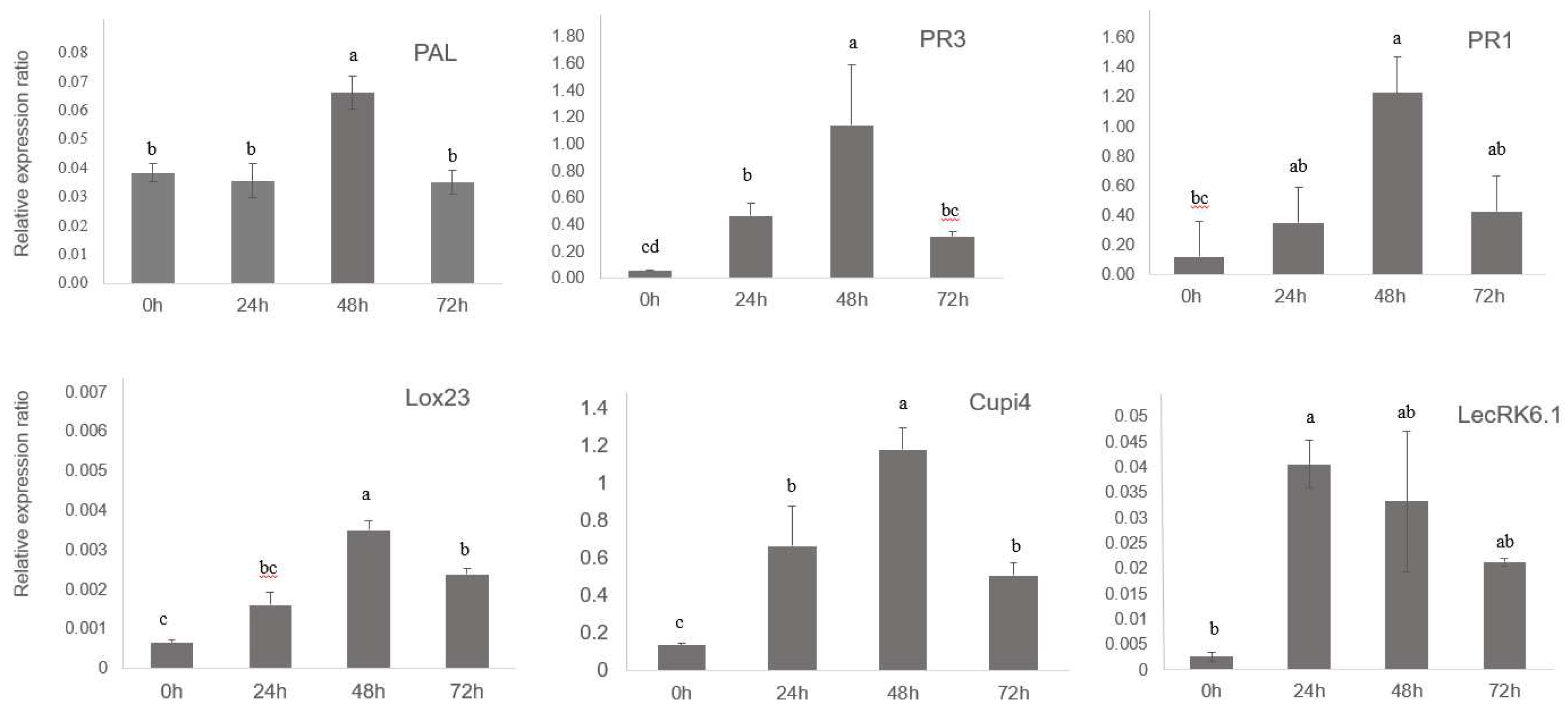
Figure 4.
Quantitative real - time PCR (qRT-PCR) analysis of defense genes in powdery mildew-infected and uninfected cucumber leaves. The expression of each gene was normalized to that of the reference gene, Actin. The expression value is shown as the average of three replicates, and the bar indicates the standard deviation. Different letters indicate significant differences between treatments (P < 0.05 according to Duncan’s multiple test).
Figure 4.
Quantitative real - time PCR (qRT-PCR) analysis of defense genes in powdery mildew-infected and uninfected cucumber leaves. The expression of each gene was normalized to that of the reference gene, Actin. The expression value is shown as the average of three replicates, and the bar indicates the standard deviation. Different letters indicate significant differences between treatments (P < 0.05 according to Duncan’s multiple test).
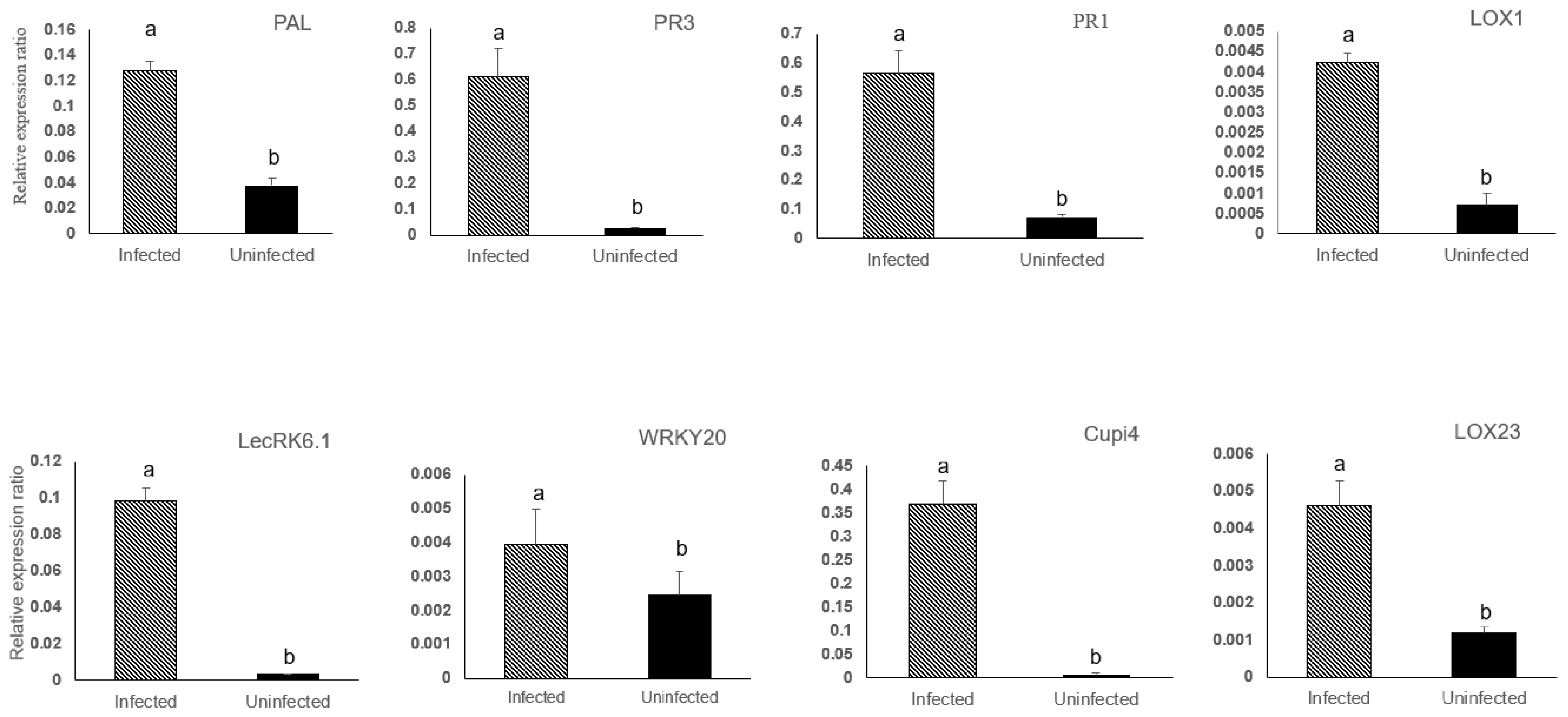
Figure 5.
Accumulation of salicylic acid in BABA-treated cucumber. Values are presented as the mean ± standard deviation of three replicates. Different letters indicate significant differences between treatments (P < 0.05 according to Duncan’s multiple test). BABA, DL-3-aminobutyric acid; DW, distilled water.
Figure 5.
Accumulation of salicylic acid in BABA-treated cucumber. Values are presented as the mean ± standard deviation of three replicates. Different letters indicate significant differences between treatments (P < 0.05 according to Duncan’s multiple test). BABA, DL-3-aminobutyric acid; DW, distilled water.
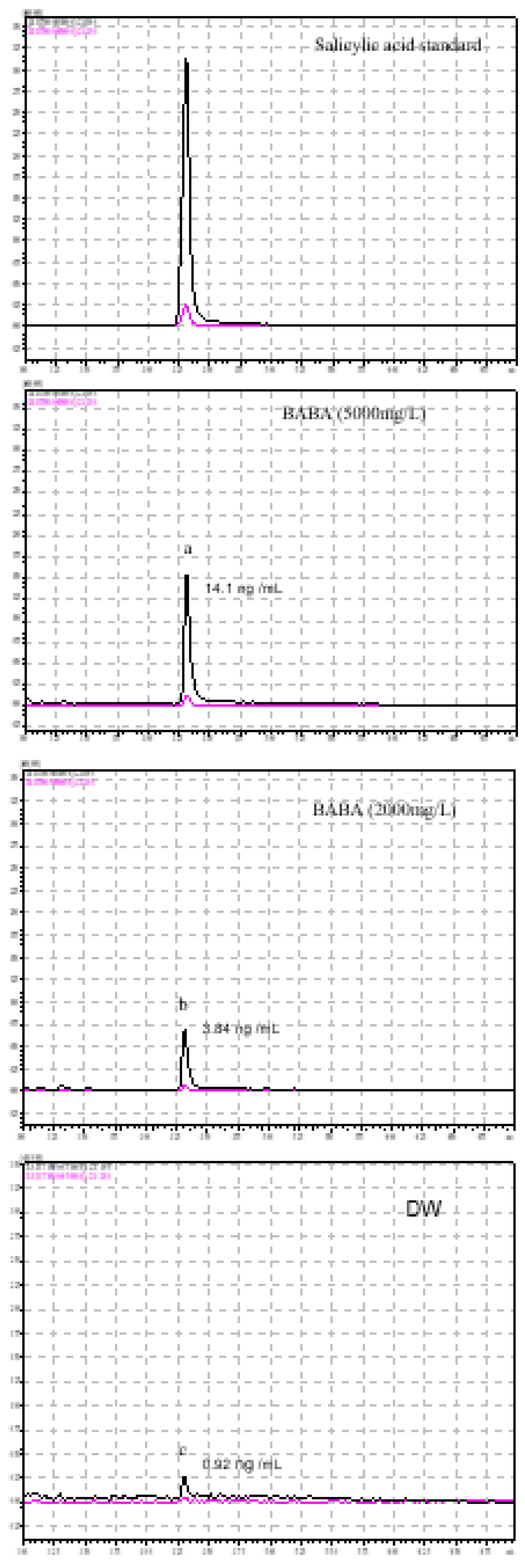

Table 1.
Primer sequences targeting defense genes in cucumber.
| Genes | Primer Sequence |
|---|---|
| CsActin | F : 5′-TCG TGC TGG ATT CTG GTG-3′ |
| R : 5′-GGC AGT GGT GGT GAA CAT-3′ | |
| CsPAL | F : 5′-AAA CAC GTC GGA TAA ATA TGG CTT -3′ |
| R : 5′-CAT CCA TTC AGG CGT TCC AG -3′ | |
| CsPR3 | F : 5′-CAC TGC AAC CCT GAC AAC AAC G -3′ |
| R :5′-AAG TGG CCT GGA ATC CGA CTG -3′ | |
| CsPR1 | F : 5′-CTC AAG ACT TCG TCG GTG TCC A -3′ |
| R : CGC CAG AGT TCA CTA GCC TAC | |
| CsLOX1 | F : 5′-TCT TTG CTT CAG GGT ATC AC -3′ |
| R : 5′-GCA AAT TCT TCA TCA CTA CTC C -3′ | |
| LOX23 | F : 5′-TGC CTC CAA CAC CTT CTT CAA -3′ |
| R : 5′-CTT CCA TAT CAA ATC GCC ACA -3′ | |
| CsLecRK6.1 | F : 5′-CGA CCA CAA CGA AAT GTC ACA C -3′ |
| R : 5′- TTT CTT CCA CAC GCC ACT TCC -3′ | |
| CsWRKY20 | F : 5′-GAA ATA ACG TAC AGA GGG AAG C -3′ |
| R : 5′-CAG GTG CTG TTT GTT GGT TAT G -3′ | |
| Cupi4 | F : 5′-TCA CTG TGG TGT GTG CTC TC -3′ |
| R : - ACT CAA GCC ATT GCC TTC CA-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



