
Submitted:
30 August 2023
Posted:
04 September 2023
You are already at the latest version
Abstract
Fipronil is an insecticide widely used in the agricultural and veterinary sectors for its efficacy in pest control. The presence of fipronil in the environment is mainly due to agricultural and domestic practices and is frequently found in different types of environmental matrices in concentrations ranging from µg/L to mg/L and can be hazardous to non-target organisms due to its high toxicity. This study was carried out to obtain and characterize microorganisms from soil which are capable of biodegrading fipronil that could be of great biotechnological interest. For this purpose, a bioprospecting was carried out using fipronil (0.6 g/L) as the main source of carbon and nitrogen for growth. Once obtained, the strain was identified by sequencing the 16S ribosomal RNA (rRNA) gene and the capacity to degrade fipronil was monitored by GC-MS. Our study showed the presence in soil samples of the strain identified as Enterobacter chengduensis, which was able to metabolize fipronil and its metabolites during the mineralization process. Enterobacter chengduensis was able to biodegrade fipronil (96%) and its metabolites fipronil-sulfone (92%) and fipronil-sulfide (79%) in 14 days. Overall, the results of this study provided a bacterium with great potential that could contribute to the degradation of fipronil in the environment.
Keywords:
Pesticide
; Bioprospecting
; Metabolites
; Biodegradation
1. Introduction
Fipronil is an insecticide widely used in the agricultural and veterinary sectors due to its efficacy in pest control [1]. This compound, which belongs to the phenylpyrazole group of insecticides, acts directly on the target nervous system, blocking the gamma-aminobutyric acid (GABA) receptors and altering the permeability of the chloride ion flux, causing paralysis and the death of insects [2].Due to its potential for action, according to data from the Brazilian Institute of Environment and Renewable Natural Resources (IBAMA), it is estimated that only in 2020 more than 2,000 tons of fipronil were commercialized [3]. Although this compound is highly effective against insects tolerant to other classes of insecticides, in some countries in Europe and China its use is banned due to its high toxicological potential [2,3,4].
This prohibition, in part, is related to the problem that only 1% of the pesticides applied in agriculture reach the crops, the rest is dispersed in the environment [5]. Prado and colleagues [6], reported that only 7% of the applied fipronil remains on crops. As it leaches through the soil due to natural factors (e.g., rain, leaching and wind), the presence of fipronil and its metabolites in soil, water bodies and sediments has become alarming [2]. Indirect contact of fipronil with surfaces, such as treated clothing and animals, are also potential sources of contamination and poisoning [7]. Thus, the presence of this compound in the environment is mainly due to agricultural and domestic practices, being frequently found in different matrices varying in concentrations from µg/L to mg/L [8]. Finally, the use of compounds such as fipronil poses risks to the environment due to their long persistence, with a half-life of up to 200 days [1].
Fipronil also possesses non-specific actions, in other words, this compound is potentially toxic to non-target living organisms, mainly aquatic organisms. In this context, in the recent study by Park et al. [8], fipronil was found to be toxic to zebrafish (Danio rerio) embryos at 2.5 mg/L and above. In other studies, with environmentally relevant concentrations, lethality was observed for the microcrustacean Daphnia magna from 0.07-0.3 mg/L, and inhibition of photosynthetic activity of the chlorophycean microalga Chlamydomonas reinhardtii at 2.4 mg/L [9]. Biochemical and genetic response modifications were also observed in organisms exposed to fipronil. For example, Monteiro et al. [10], elucidated the inhibition of antioxidant activity and of motor protein expression and globin biosynthesis in Chironomus riparius exposed to 0.08 µg/L for 48 h. Similarly, El-Murr and colleagues [11] identified an increase in liver enzymes, indicating hepatotoxicity, and showed a decrease in antioxidant activity in the Nile tilapia (Oreochromis niloticus) exposed to 42 µg/L fipronil for 96h.
As mentioned earlier, fipronil and its metabolites originated during the degradation process, are responsible for most of the contamination of agricultural fields [12]. These metabolites come from the mineralization process of the fipronil molecule, produced through its oxidation, reduction, hydrolysis, and photolysis producing fipronil sulfone, fipronil sulfide, fipronil amide and fipronil disulfenyl, respectively [1]. Due to its high harmfulness, especially of fipronil sulfone, a metabolite even more toxic than fipronil, studies focused on the remediation of these molecules using microorganisms have become an approach widely used in order to minimize the impacts caused by environmental exposure to these compounds [2,3,6,7,13,14]. Given these approaches, biological processes are the most recommended [2] and the microorganisms selected from the environment contaminated with these compounds are capable of using fipronil as a source of carbon and nitrogen, making the biodegradation process more promising [6,15].
In this context, it is important to try to minimize the environmental impacts associated with the use of fipronil, as well as to identify microorganisms with the potential to metabolize fipronil and its derivatives. Consequently, this research aims to answer the hypothesis of whether from fipronil-contaminated soils we can isolate microorganisms capable of biodegrading this compound and with the potential to be used in bioremediation. For this purpose, we bioprospected and analyzed the profile of metabolites produced during the metabolization of fipronil, as well as the biodegradation kinetics of this compound, aiming to find, in contaminated soil, the microorganism with the highest capacity to degrade this important agricultural compound.
2. Materials and Methods
2.1. Soil Sampling and Microorganisms isolation
The microorganism used in this study was isolated from samples of soil cultivated with corn having a history of fipronil application. The soil was collected at depths of 0–10 cm from the Experimental Farm of the Federal University of Grande Dourados, Dourados, MS, Brazil (22°48′53′′ S and 54°44′31′′ W). Ten grams of the soil was diluted in 90 mL saline (0.9%) and a serial dilution (10-1 -10-4), was carried out. Samples (100 µL) were plated out by the spread plate method using solid ATZ-R, previously described by Prado et al. [6],containing 0.6 g/L fipronil and incubated at 30 °C for 48 h. Selected bacterial colonies were purified by streak plate and subsequently subjected to a Gram stain to check for the presence of spores and cell wall composition, as described by Prado et al. [6]. The most relevant steps used in this work are described in detail in Figure 1.
2.2. Molecular Identification of Strain and phylogenetic tree
The molecular identification of the strain was made by amplifying the 16S rDNA gene and sequencing by the Sanger method. The analyses were performed by the Facility Centralized Multi-User Laboratory for Large-scale DNA Sequencing and Gene Expression Analysis at Unesp in Jaboticabal, SP, Brazil (LMSEQ), using primers FD1 (CCGAATTCGTCGACAACAGAGTTTGATCCTGGCTCAG), and RD1(CCCGGGATCCAAGCTTAAGGAGGTGATCCAGCC) [16]After DNA sequencing, the assembly process was carried out using CAP3 software. The contig was deposited in the GenBank database (https://www.ncbi.nlm.nih.gov/genbank/) under accession code: OR365541.
The Basic Local Alignment Search Tool (BLAST) was used for sequence comparison against Silva database [17,18] and Genbank database [18]. To evaluate the results between the databases (Silva and GenBank), sequence combinations with the highest similarity scores and the lowest e-values were used.
A phylogenetic tree was built to support the results obtained in the similarity analysis. The evolutionary history was inferred using the Neighbor-Joining method [19]. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates) are shown next to the branches. The evolutionary distances were computed using the p-distance method and are in the units of the number of base differences per site. The rate variation among sites was modeled with a gamma distribution (shape parameter = 1). This analysis involved 7 nucleotide sequences. All positions containing gaps and missing data were eliminated (complete deletion option). There were a total of 1299 positions in the final dataset. Evolutionary analyses were conducted in MEGA11 [20].
2.3. Pre-inoculum preparation
The microorganisms were incubated in Erlenmeyer flasks containing liquid ATZ-R enriched with 0.6 g/L of fipronil and 1% yeast extract for 48 h at 30 °C and orbital agitation at 100 rpm. Then, the cells were washed with ATZ-R to remove the remaining yeast extract. Bacterial cells were centrifuged for 10 min at 5 °C, 3500 rpm, and the supernatant was discarded. The process was carried out three times. Cells were resuspended in ATZ-R to an optical density (OD) of 0.8 to 1.0 (108 cells/mL) at 600 nm and used as inoculum for the degradation experiments.
2.4. Fipronil Degradation
Assays were performed in triplicate with each flask containing 30 mL of liquid ATZ-R, fipronil (0.6 g/L) and 1 mL of inoculum (108 cells/mL), as described by Prado et al. [6]. Bacterial growth was evaluated through dry biomass. Biotic controls (without fipronil) were performed for all sampling time points. Abiotic controls (without the bacteria) were performed at 0 and 14 days to evaluate abiotic degradation of fipronil. Dry biomass analysis was also performed to evaluate the sterility of the control culture. Cultures were incubated at 30°C at 140 rpm for 14 days and samples were collected at 7 time points (0, 3, 5, 7, 10, 12 and 14 days) as described by Uniyal et al. [13]. Samples were centrifuged at 4°C at 3500 rpm for 15 min and the supernatant stored at -20°C for GC-MS analysis and the biomass pellet was used for growth analysis.
The total degradation values of the samples were calculated using equation 1 (Eq. 1), in which total degradation (TD) is obtained by dividing the difference between the initial fipronil concentration (FCi) and the final fipronil concentration (FCf) by abiotic degradation (AD).
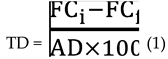
Biological degradation (BD) equals TD minus AD, multiplied by the percentage factor (100), as follows in Eq. 2
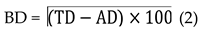
The pre-inoculum was prepared in liquid ATZ-R containing 1% yeast extract and 0.6 g/L fipronil. After 48 h of growth, the cells were washed to completely remove the remaining yeast extract. Bacterial cells were centrifuged for 10 min at 5°C, 3500 rpm. The supernatant was discarded, and the cells were washed three times with ATZ-R and fipronil 0.6 g/L. Cells were resuspended in ATZ-R to an OD of 0.8 to 1.0 (108 cells/mL) at 600 nm and used as inoculum for the degradation experiments.
2.5. GC-MS conditions and analysis of fipronil biodegradation and metabolites quantification
Fipronil (Chem Service, 99.5% purity), fipronil-sulfide (BASF 98.8% purity) and fipronil-sulfone (BASF 99.7% purity) stock standard solutions of 10 ng/µL were prepared in acetone. The working standard solutions of 100, 200, 500, 1000 and 2000 pg/µL were prepared from a stock solution after dilution in acetone and used for calibration curves. The samples were extracted with ethyl acetate in a 1:1 (v/v) ratio and homogenized for 2 min in orbital shakers. The organic phase that formed was collected and concentrated by rotary evaporation and, thereafter, resuspended in 4 mL of acetone. Fipronil and its metabolites (fipronil-sulfide and fipronil-sulfone) were quantified using GC-MS (TRACE 1300 CG-MS Thermo) in a single ion-monitoring mode and equipped with an automatic injector. Separation was carried out using a TG-1 MS capillary column (30 m × 0.25 mm × 0.50 µm). The oven temperature was: 60°C for 1 min, heated to 200°C at a rate of 30°C min-1 and kept for 5 min, heated to 270°C at a rate of 30°C min-1 and kept for 5 min and heated to 300°C at a rate of 30°C/min and kept for 11 min. The total run was 30 min. Helium was the carrier gas at a flow rate of 1 mL/min. The injector temperature was 280°C in a splitless injection mode and transfer line at a temperature of 250 °C.
2.6. Statistical analysis
All experiments were evaluated by analysis of variance (ANOVA) followed by the Fisher's test. The significant difference was calculated using the Fisher Test mean test (p < 0.05) (Minitab 20.0).
3. Results and Discussion
3.1. Isolation and molecular identification
Seven bacteria capable of growing in ATZ-R medium containing fipronil (0.6 g/L) as the main carbon source were isolated (results not shown), from the soil samples. After successive growth evaluations, the isolate G2.8 showed the best growth potential during 96 h, being selected for the fipronil biodegradation assays. In a preliminary analysis of the morphology of the isolate, it was observed that it is a Gram-negative rod, catalase positive and oxidase negative. The selective pressure due to the high toxicity of fipronil probably proved to be a limiting factor in obtaining more isolates from soil samples. In other studies, microorganisms capable of degrading fipronil have been isolated and studied [21,22,23], however, many studies aimed at isolating microorganisms for subsequent biotechnological application have been limited as more than 99% of microorganisms cannot be cultured on media [24], and therefore limited knowledge about the degradation pathway of fipronil is available.
In line with the above, once the isolate with the ability to grow with fipronil was isolated, it was molecularly identified by amplification of the 16S rDNA gene and sequenced. After sequencing and assembly of the 16S rDNA gene of isolate G2.8 OR365541, the contig was aligned with the most similar sequences in the GenBank database. Among the most similar sequences strain G2.8 OR365541 clustered closely with Enterobacter chengduensis NR179167.1, supported by a bootstrap value of 99. Figure 2, shows the clustering of the DNA sequence of the 16S rRNA gene of the bacteria E. chengduensis.
Regarding the genus of the microorganism isolated in this work, Enterobacter belongs to the group of gram-negative bacteria of the large phylum Proteobacteria. These microorganisms are widely distributed in the environment, and one of the predominant phyla has already been described in a metagenomic study with fipronil [25]. However, no studies have reported the potential of bacteria of the genus Enterobacter as fipronil degraders, only representatives of the phylum Proteobacteria. For example, Cappeline et al. [26], identified that the species Burkholderia thailandensis was able to degrade fipronil and its metabolites fipronil-sulfide and fipronil-sulfone. In another study, Bhatti et al. [7] identified that a non-pathogenic strain of Escherichia coli was able to bioaccumulate and biotransform fipronil. In the same vein, Kumar and colleagues [27], observed that soil bacteria of the phylum Paracoccus sp. degraded 80 µg/kg fipronil in sandy soil after 30 days. Finally, Imaniar et al. [28], identified that the species Pseudomonas aeruginosa was able to degrade 65% of fipronil (40 mg/L) after 3 days.
In addition, studies have shown that the genus Enterobacter has a high capacity for biodegradation of toxic compounds, e.g., synthetic polymers (polyethylene and polypropylene), through the bacterial consortium of Enterobacter sp. and Pseudomonas sp. [29]; insecticide (endosulfan), with Enterobacter asburiae JAS5 and Enterobacter cloacae JAS7 [30]; azo dyes (reactive yellow 145 and reactive red 180), by Enterobacter hormaechei species [31].
Regarding the species isolated in this study, the bacterium E. chengduensis is a new species described in 2019 in China [32,33]. This species has been reported due to its potential to metabolize lignocellulosic compounds [34]. However, no study reports its potential as a pesticide biodegradation strain. Most studies report that the species is related to human infections, an opportunistic pathogen normally associated with other species such as E. asburiae, E. cloacae, E. hormaechei, Enterobacter kobei and Enterobacter ludwigii [27,28].
3.1. Fipronil degradation
As mentioned above, fipronil contamination of soils can alter the diversity of microbial communities [9,35,36]. This is because the high toxicity of fipronil and the relatively low water activity found in soils probably act as a selective pressure for the growth of microorganisms, in other words, only those microorganisms possessing the enzymatic machinery that enable them to degrade these compounds are able to survive. An example of this has been observed in the recently published study by Guima et al. [20], where the authors observed the variation of the microbial community in the presence of fipronil, furthermore the authors have shown that the phyla represented by Proteobacteria, Actinobacteria, and Firmicutes have been benefited in this condition. Additionally, some genera of bacteria may also possess the capacity for chemotaxis, which helps them to identify the presence of contaminants through chemical signals, as is the case with actinobacteria and diazotrophic bacteria, helping them to survive in contaminated environments [1]. In this way, bioprospecting microorganisms from fipronil-contaminated environments can accelerate the biodegradation process [2,6,13,15], as the bioprospected microorganisms, which have been able to grow in the presence of these compounds, should have the enzymatic machinery necessary for the metabolization of fipronil.
In line with the previous work, once the bacteria that showed the best growth in the presence of fipronil had been bioprospected and identified here, their ability to degrade this compound was determined. To do this, firstly, the growth kinetics of E. chengduensis was evaluated by assessing the dry biomass. Table 1 shows that the highest growth rate obtained by E. chengduensis occurred on day 7. Furthermore, no microbial growth was observed in the biotic and abiotic controls, showing that E. chengduensis was dependent on and grew exclusively using fipronil as an energy source.
Secondly, the degradation taxa of fipronil and its derivatives were quantified by GS-MS. In this regard, it was observed that after 14 days, E. chengduensis was able to degrade 96% of the fipronil present in the sample (Figure 3A), which highlighted the biodegradation capacity of the bioprospected bacteria. On the other hand, it was observed that the highest degradation rate occurred between the first and third day of the experiment (Figure 3A). Simultaneously, it was identified that the highest biomass increase occurred after 5 days (Table 1). Furthermore, it was also found that 40% up to 45% of the degradation occurring was due to environmental factors (chemical degradation, photodegradation, adsorption, among other possible processes), as observed by the decrease in the abiotic control values (Figure 3A). Finally, it was observed that there was a 50% reduction from the initially inoculated biomass to the final time, demonstrating that Enterobacter chengduensis was exclusively dependent on the presence of fipronil for its growth (Table 1).
Similar results were obtained in the study by Viana et al. [3], where the authors observed that Bacillus amyloliquefaciens strain RFD1C was able to degrade 93% of fipronil (10 mg/L) over a period of 5 days. Similarly, Uniyal et al. [13], identified that a strain of Stenotrophomonas acidaminiphyla, was able to degrade 70% of fipronil (50 mg/L) in 14 days. In another study, Bhatt and colleague [15],demonstrated the ability of 76 % degradation of fipronil (50 mg/L) by Bacillus sp. strain FA3 over 15 days. These results are also in line with those obtained by Gangola et al. [37], who observed a 93% degradation rate of fipronil (450 mg/L) after 15 days of growth by Bacillus sp. strain 3C. In another similar study, Abraham et al. [33],also obtained 100% degradation of fipronil (500 mg/L) after 15 days by Streptomyces rochei strain AJAG7. Finally, some studies have also demonstrated the ability of fungi such as Trametes versicolor [34], and Aspergillus glaucus AJAG1 [35], to degrade fipronil. Taken together, these findings suggest that microorganisms isolated from fipronil-contaminated environments show good performance in the biodegradation process of this compound, which could be a promising tool for the decontamination of fipronil-contaminated agricultural environments.
Regarding the production of metabolites during the fipronil degradation process, over 14 days, a higher production of the metabolite fipronil-sulfone produced by the fipronil oxidation pathway was observed (Figure 2B) in relation to fipronil-sulfide produced by the reduction of this compound (Figure 3C). On the other hand, it was observed, by comparison with the abiotic control, that fipronil-sulfone showed a 25% degradation, while fipronil-sulfide showed a 17% degradation in 14 days. Both metabolites showed an increase at 5 days (Figure 3B,C) and degradation at 12 days. These data suggest that the presence of Enterobacter chengduensis could influence the biodegradation process of fipronil, being responsible for the simultaneous production and degradation of metabolites in an accelerated manner.
The observed increase in fipronil sulphone production by Enterobacter chengduensis could be related to biomass decomposition (Table 1). Another factor that could be related is that the microorganism possesses the active oxidase enzyme (see Isolation and molecular identification), presenting an increased oxidation capacity. Little is known about the mechanisms related to the pathways and metabolization process of fipronil. The observations of Bhatt and colleagues [2], further support the hypothesis that aerobic microorganisms with active oxidase, such as Enterobacter chengduensis biosprospected here, have a higher capacity to oxidize fipronil to generate fipronil-sulfone, while microaerophiles or anaerobes further reduce fipronil, forming fipronil-sulfide.
Finally, although the metabolic pathways of fipronil have not been described, it is known that its metabolism occurs through biochemical processes. Specifically, it has been described that the detoxification of this molecule occurs through mechanisms related to the cytochrome P450 complex, divided into two stages, I and II [41]. In step I, oxidation of the pyrazole ring of fipronil occurs. Step II occurs through detoxification by catalytic enzymes, which first promote hydroxylation of the aromatic ring (target position), with consequent glycosylation of the added hydroxyl[1]. Even without the exact identification of the fipronil degradation pathways, some P450 enzymes (example: CYP18, CYP302, CYP4 and CYP6) have been identified as fipronil detoxification enzyme genes [42]. In this sense, the findings obtained with this study motivate future research and search for microorganisms with potential to degrade fipronil like the one described here, as well as to understand the metabolic mechanisms used by them and thus use this information for the development of new tools, such as bacterial consortia, to try to minimize the impacts of human activity on agricultural soil.
4. Conclusions
In this work, from soils contaminated with fipronil, we have been able to isolate microorganisms capable of biodegrading this compound and with the potential to be used in bioremediation. Therefore, the hypothesis initially proposed was confirmed.
In this sense, from the data obtained, it can be affirmed that the microorganisms present in the soil contaminated by fipronil, can attribute expressive results in the degradation of fipronil. It was identified that the Enterobacter chengduensis strain, isolated from maize crop soil contaminated by fipronil showed high fipronil degradation capacity in relatively short periods of time. Probably, this activity is observed due to its prolonged exposure to the pesticide. Moreover, it was observed that the metabolites produced during fipronil degradation did not present toxicity to the microorganisms, thus allowing their functional metabolism during the mineralization process.
Consequently, Enterobacter chengduensis could be a good candidate to contribute to environmental protection, being used in bioremediation processes in areas contaminated with fipronil minimizing the negative impacts of human intervention in the environment.
Author Contributions
Conceptualization, C.P. (Caio Prado) and M.B. (Maricy Bonfá); methodology, C.P., R.P. (Rodrigo Pereira) and R.J. (Rômulo Júnior); software, C.P., R.P. and R.J.; validation, C.P. and R.J.; formal analysis, C.P., R.P. and R.J; investigation, C.P., R.P., R.J., F.P. (Francine Piubeli) and M.B.; resources, R.P., R.J and M.B..; data curation, C.P., R.P., F.P.,; writing—original draft preparation, C.P. and M.B.; writing—review and editing, C.P., R.P., L.D. (Lucia Durrant), F.P. and M.B.; visualization, L.D., F.P., M.B.; supervision, F.P. and M.B.; project administration, R.P. and M.B.; funding acquisition, M.B. All authors have read and agreed to the published version of the manuscript.
Funding
Fundect—Support Foundation for the Development of Education (n.050/2015 siafem. 025215) , Science and Technology of the State of Mato Grosso do Sul. CNPq—National Council for Scientific and Technological Development.
Acknowledgments
The Centralized Multiuser Laboratory for Large Scale DNA Sequencing—LMSeq (Facility FAPESP proc. No. 2009/53984-2) of UNESP Campus Jaboticabal, SP Brazil. Universidade Federal da Grande Dourados, MS Brazil—UFGD.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bonfá, M.R.L.; do Prado, C.C.A.; Piubeli, F.A.; Durrant, L.R. Fipronil Microbial Degradation: An Overview From Bioremediation to Metabolic Pathways. In Pesticides Bioremediation; Springer International Publishing: Cham, 2022; pp. 81–107. [Google Scholar]
- Bhatt, P.; Gangola, S.; Ramola, S.; Bilal, M.; Bhatt, K.; Huang, Y.; Zhou, Z.; Chen, S. Insights into the Toxicity and Biodegradation of Fipronil in Contaminated Environment. Microbiol Res 2023, 266, 127247. [Google Scholar] [CrossRef] [PubMed]
- Viana, J.G.; Birolli, W.G.; Porto, A.L.M. Biodegradation of the Pesticides Bifenthrin and Fipronil by Bacillus Isolated from Orange Leaves. Appl Biochem Biotechnol 2023, 195, 3295–3310. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.Y.; Lim, J.W.; Lim, M.C.; Song, N.E.; Woo, M.A. Aptamer-Based Fluorescent Assay for Simple and Sensitive Detection of Fipronil in Liquid Eggs. Biotechnology and Bioprocess Engineering 2020, 25, 246–254. [Google Scholar] [CrossRef]
- Gangola, S.; Sharma, A.; Joshi, S.; Bhandari, G.; Prakash, O.; Govarthanan, M.; Kim, W.; Bhatt, P. Novel Mechanism and Degradation Kinetics of Pesticides Mixture Using Bacillus Sp. Strain 3C in Contaminated Sites. Pestic Biochem Physiol 2022, 181, 104996. [Google Scholar] [CrossRef]
- Prado, C.C.A. do; Pereira, R.M.; Durrant, L.R.; Júnior, R.P.S.; Bonfá, M.R.L. Fipronil Biodegradation and Metabolization by Bacillus Megaterium Strain E1. Journal of Chemical Technology & Biotechnology 2021. [Google Scholar] [CrossRef]
- Bhatti, S.; Satyanarayana, G.N.V.; Patel, D.K.; Satish, A. Bioaccumulation, Biotransformation and Toxic Effect of Fipronil in Escherichia Coli. Chemosphere 2019, 231, 207–215. [Google Scholar] [CrossRef]
- Park, H.; Lee, J.-Y.; Park, S.; Song, G.; Lim, W. Developmental Toxicity of Fipronil in Early Development of Zebrafish (Danio Rerio) Larvae: Disrupted Vascular Formation with Angiogenic Failure and Inhibited Neurogenesis. J Hazard Mater 2020, 385, 121531. [Google Scholar] [CrossRef]
- Pino-Otín, M.R.; Ballestero, D.; Navarro, E.; Mainar, A.M.; Val, J. Effects of the Insecticide Fipronil in Freshwater Model Organisms and Microbial and Periphyton Communities. Science of the Total Environment 2020, 1–17. [Google Scholar] [CrossRef]
- Monteiro, H.R.; Pestana, J.L.T.; Novais, S.C.; Leston, S.; Ramos, F.; Soares, A.M.V.M.; Devreese, B.; Lemos, M.F.L. Assessment of Fipronil Toxicity to the Freshwater Midge Chironomus Riparius: Molecular, Biochemical, and Organismal Responses. Aquatic Toxicology 2019, 216, 105292. [Google Scholar] [CrossRef]
- El-Murr, A. elhakeem I.; Abd El Hakim, Y.; Neamat-Allah, A.N.F.; Baeshen, M.; Ali, H.A. Immune-Protective, Antioxidant and Relative Genes Expression Impacts of β-Glucan against Fipronil Toxicity in Nile Tilapia, Oreochromis Niloticus. Fish Shellfish Immunol 2019, 94, 427–433. [Google Scholar] [CrossRef]
- Baldaniya, D.M.; Singh, S.; Saini, L.K.; Gandhi, K.D. Persistence and Dissipation Behaviour of Fipronil and Its Metabolites in Sugarcane Grown Soil of South Gujarat. Int J Chem Stud 2020, 8, 1524–1527. [Google Scholar] [CrossRef]
- Uniyal, S.; Paliwal, R.; Sharma, R.K.; Rai, J.P.N. Degradation of Fipronil by Stenotrophomonas Acidaminiphila Isolated from Rhizospheric Soil of Zea Mays. 3 Biotech 2016, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Wu, H.; Guo, J.; Kimaro, F.M.E. Microbial Degradation of Fipronil in Clay Loam Soil. Water Air Soil Pollut 2004, 153, 35–44. [Google Scholar] [CrossRef]
- Bhatt, P.; Sharma, A.; Rene, E.R.; Jagadeesh, A.; Zhang, W.; Chen, S. Bioremediation of Fipronil Using Bacillus Sp. FA3 : Mechanism, Kinetics and Resource Recovery Potential from Contaminated Environments. Journal of Water Process Engineering 2021, 39, 101712. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S Ribosomal DNA Amplification for Phylogenetic Study. J Bacteriol 1991, 173, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res 2013, 41, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Benson, D.A.; Karsch-mizrachi, I.; Lipman, D.J.; Ostell, J.; Rapp, B.A.; Wheeler, D.L. GenBank. 2000, 28, 15–18. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol Biol Evol 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol Biol Evol 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Mandal, K.; Singh, B.; Jariyal, M.; Gupta, V.K. Bioremediation of Fipronil by a Bacillus Firmus Isolate from Soil. Chemosphere 2014, 101, 55–60. [Google Scholar] [CrossRef]
- Zhou, Z.; Wu, X.; Lin, Z.; Pang, S.; Mishra, S.; Chen, S. Biodegradation of Fipronil: Current State of Mechanisms of Biodegradation and Future Perspectives. Appl Microbiol Biotechnol 2021, 105, 7695–7708. [Google Scholar] [CrossRef] [PubMed]
- Uniyal, S.; Paliwal, R.; Sharma, R.K.; Rai, J.P.N. Degradation of Fipronil by Stenotrophomonas Acidaminiphila Isolated from Rhizospheric Soil of Zea Mays. 3 Biotech 2016, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Elizaquível, P.; Pérez-Cataluña, A.; Yépez, A.; Aristimuño, C.; Jiménez, E.; Cocconcelli, P.S.; Vignolo, G.; Aznar, R. Pyrosequencing vs. Culture-Dependent Approaches to Analyze Lactic Acid Bacteria Associated to Chicha, a Traditional Maize-Based Fermented Beverage from Northwestern Argentina. Int J Food Microbiol 2015, 198, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Guima, S.E.S.; Piubeli, F.; Bonfá, M.R.L.; Pereira, R.M. New Insights into the Effect of Fipronil on the Soil Bacterial Community. Microorganisms 2022, 11, 52. [Google Scholar] [CrossRef]
- Cappelini, L.T.D.; Alberice, J.A.; Eugênio, P.F.M.; Pozzi, E.; Urbaczek, A.C.; Diniz, L.G.R.; Carrilho, E.N.V.M.; Carrilho, E.; Vieira, E.M. Burkholderia Thailandensis: The Main Bacteria Biodegrading Fipronil in Fertilized Soil with Assessment by a QuEChERS/GC-MS Method. J. Braz. Chem. Soc 2018, 29, 1934–1943. [Google Scholar] [CrossRef]
- Kumar, R.; Singh, B.; Gupta, V.K. Biodegradation of Fipronil by Paracoccus Sp. in Different Types of Soil. Bull Environ Contam Toxicol 2012, 88, 781–787. [Google Scholar] [CrossRef]
- Imaniar, K.R.; Tazkiaturrizki, T.; Rinanti, A. Fipronil Removal at Various Temperature and Pollutant Concentration by Using Pseudomonas Aeruginosa and Brevibacterium Sp. in Liquid Media. IOP Conf Ser Mater Sci Eng 2021, 1098, 052071. [Google Scholar] [CrossRef]
- Skariyachan, S.; Taskeen, N.; Kishore, A.P.; Krishna, B.V.; Naidu, G. Novel Consortia of Enterobacter and Pseudomonas Formulated from Cow Dung Exhibited Enhanced Biodegradation of Polyethylene and Polypropylene. J Environ Manage 2021, 284, 112030. [Google Scholar] [CrossRef]
- Abraham, J.; Silambarasan, S. Biomineralization and Formulation of Endosulfan Degrading Bacterial and Fungal Consortiums. Pestic Biochem Physiol 2014, 116, 24–31. [Google Scholar] [CrossRef]
- Thangaraj, S.; Bankole, P.O.; Sadasivam, S.K. Microbial Degradation of Azo Dyes by Textile Effluent Adapted, Enterobacter Hormaechei under Microaerophilic Condition. Microbiol Res 2021, 250, 126805. [Google Scholar] [CrossRef]
- Wu, W.; Feng, Y.; Zong, Z. Characterization of a Strain Representing a New Enterobacter Species, Enterobacter Chengduensis Sp. Nov. Antonie Van Leeuwenhoek 2019, 112, 491–500. [Google Scholar] [CrossRef]
- Wu, W.; Wei, L.; Feng, Y.; Kang, M.; Zong, Z. Enterobacter Huaxiensis Sp. Nov. and Enterobacter Chuandaensis Sp. Nov., Recovered from Human Blood. Int J Syst Evol Microbiol 2019, 69, 708–714. [Google Scholar] [CrossRef]
- Xie, R.; Dong, C.; Wang, S.; Danso, B.; Dar, M.A.; Pandit, R.S.; Pawar, K.D.; Geng, A.; Zhu, D.; Li, X.; et al. Host-Specific Diversity of Culturable Bacteria in the Gut Systems of Fungus-Growing Termites and Their Potential Functions towards Lignocellulose Bioconversion. Insects 2023, 14, 403. [Google Scholar] [CrossRef]
- Sim, J.X.F.; Drigo, B.; Doolette, C.L.; Vasileiadis, S.; Donner, E.; Karpouzas, D.G.; Lombi, E. Repeated Applications of Fipronil, Propyzamide and Flutriafol Affect Soil Microbial Functions and Community Composition: A Laboratory-to-Field Assessment. Chemosphere 2023, 331, 138850. [Google Scholar] [CrossRef] [PubMed]
- Tomazini, R.; Saia, F.T.; van der Zaan, B.; Grosseli, G.M.; Fadini, P.S.; de Oliveira, R.G.M.; Gregoracci, G.B.; Mozetto, A.; van Vugt-Lussenburg, B.M.A.; Brouwer, A.; et al. Biodegradation of Fipronil: Transformation Products, Microbial Characterisation and Toxicity Assessment. Water Air Soil Pollut 2021, 232, 123. [Google Scholar] [CrossRef]
- Gangola, S.; Bhatt, P.; Kumar, A.J.; Bhandari, G.; Joshi, S.; Punetha, A.; Bhatt, K.; Rene, E.R. Biotechnological Tools to Elucidate the Mechanism of Pesticide Degradation in the Environment. Chemosphere 2022, 296, 133916. [Google Scholar] [CrossRef] [PubMed]
- Abraham, J.; Gajendiran, A. Biodegradation of Fi Pronil and Its Metabolite Fi Pronil Sulfone by Streptomyces Rochei Strain AJAG7 and Its Use in Bioremediation of Contaminated Soil. Pestic Biochem Physiol 2019, 155, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Wolfand, J.; Lefevre, G.; Luthy, R. Metabolization and Degradation Kinetics of the Urban-Use Pesticide Fipronil by White Rot Fungus: Trametes Versicolor. Environ Sci Process Impacts 2016, 18, 1256–1265. [Google Scholar] [CrossRef]
- Gajendiran, A.; Abraham, J. Biomineralisation of Fipronil and Its Major Metabolite, Fipronil Sulfone, by Aspergillus Glaucus Strain AJAG1 with Enzymes Studies and Bioformulation. 3 Biotech 2017, 7, 1–15. [Google Scholar] [CrossRef]
- Girvan, H.M.; Munro, A.W. Applications of Microbial Cytochrome P450 Enzymes in Biotechnology and Synthetic Biology. Curr Opin Chem Biol 2016, 31, 136–145. [Google Scholar] [CrossRef]
- Zhou, Z.; Wu, X.; Lin, Z.; Pang, S.; Mishra, S.; Chen, S. Biodegradation of Fipronil: Current State of Mechanisms of Biodegradation and Future Perspectives. Appl Microbiol Biotechnol 2021, 105, 7695–7708. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic representation of the main steps carried out in this work.
Figure 2.
Phylogenetic Tree constructed by Neighbour-joining method with 16S rRNA gene sequences of different Enterobacater species. The circle in the phylogenetic tree indicates the 16S rRNA sequence of the isolated bacterium.
Figure 2.
Phylogenetic Tree constructed by Neighbour-joining method with 16S rRNA gene sequences of different Enterobacater species. The circle in the phylogenetic tree indicates the 16S rRNA sequence of the isolated bacterium.

Figure 3.
Profile of fipronil biodegradation and metabolite production by isolate G2.8. A) fipronil, B) fipronil-sulfone and C) fipronil-sulfide. Statistics analyses by Fisher's test (p<0.05) presented as mean±standard deviation.
Figure 3.
Profile of fipronil biodegradation and metabolite production by isolate G2.8. A) fipronil, B) fipronil-sulfone and C) fipronil-sulfide. Statistics analyses by Fisher's test (p<0.05) presented as mean±standard deviation.
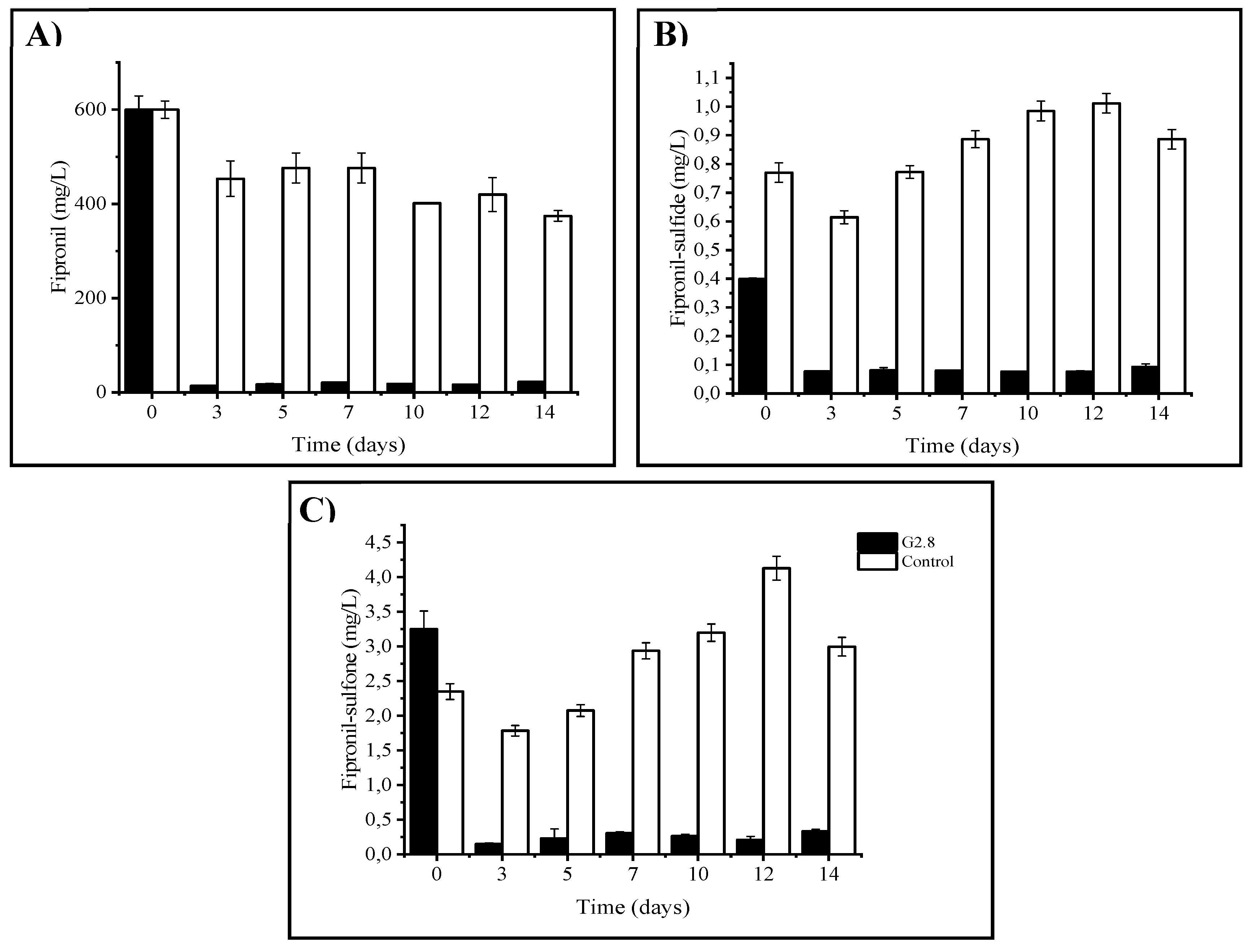

Table 1.
Growth kinetics of Enterobacter chengduensis.
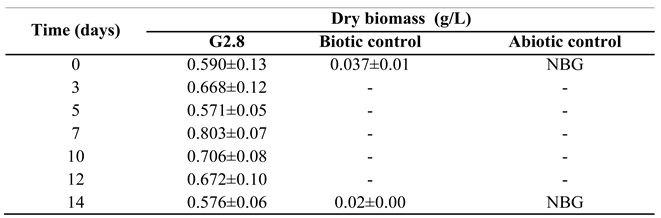
Statistics analyzed by Fisher's test (p<0.05) presented with mean ± standard deviation. Biotic and abiotic control were performed only at the beginning and at the end of the trials; NBG: no bacterial growth.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



