
Submitted:
09 August 2023
Posted:
10 August 2023
You are already at the latest version
Abstract
Investigation of bacterial communities is on the rise both in human and veterinary medicine. Their role in health maintenance and pathogenic mechanisms is in the limelight of infectious, metabolic, and cancer research. Among the most considered, gut bacterial communities takes the cake. Their part in animals was assessed mainly to improve animal production, public health, and pet management. In this regard, canaries deserve attention, being a popular pet and source of economic income for bird-keepers, for whom breeding represents a pivotal point. Thus, the aim of the present work was to follow gut bacterial communities’ evolution along on whole reproductive cycle of 12 healthy female canaries. Feces were collected during parental care, molting, and resting phase, and submitted for 16S rRNA sequencing. Data analysis a substantial presence of Lactobacillus aviarius along all the phases, and a relevant shift of microbiota during molting and rest due to an abrupt decrease of Vermiphilaceae family. Although the meaning of such change is not clear, future research may highlight unforeseen scenarios. Moreover, Lactobacillus aviarius may be deemed for normal bacteria flora restoration in debilitated birds, perhaps improving their health and productivity.
Keywords:
Canaries
; Serinus canaria
; gut bacterial microbiota
; bacterial communities
; reproduction
; reproductive cycle
; 16S rRNA gene sequencing
1. Introduction
The gut microbiota is regarded as a full-fledged endocrine organ because of its numerous effects on distant organs and pathways [1]. Commensal bacteria can produce and secrete hormones, and the interaction between hormones and microbes impacts on the metabolism, immunity, and behavior of the host. Changes in the microbiota, particularly in the gut microbial communities, have specific effects on the reproductive endocrine system [2]. In this respect, metagenomic techniques development offered a priceless opportunity to unveil microbial ecology. Among the available technologies, 16S rRNA gene sequencing represents an effective and economically affordable solution, yielding the identification to the genus level of most bacteria characterizing an environment [3,4]. Human medicine greatly took advantage on the application of metagenomic, enhancing the comprehension of microbes-host interactions and learning how to modulate microbial communities’ composition for health purposes. In women, microbiota imbalance was linked to several disease conditions, from cancer to reproductive issues such as endometriosis, polycystic ovary syndrome (PCOS), pregnancy complications, and adverse pregnancies outcomes [1]. The correlation between the shifts in the gut bacterial communities and reproduction was also investigated in many animal species, finding connections between microorganisms and the endocrine system of their host. Animals have complex and species-specific reproductive interactions that are finely tuned, and gut bacterial microbiota was demonstrated to greatly affect physiology and behavior by impacting on neurotransmitters and neuropeptides [5]. As regards birds, gut bacterial microbiota was investigated in a variety of captive and wild avian species, focusing mostly on the interplay between the gut microbiota composition and specific bacteria (especially pathogens), diet, season, and migration of the avian host [6,7]. The relationship between microbiota and reproduction was explored mainly in in laying hens and endangered birds, for commercial and conservation respectively confirming the gut bacterial community’s footprint on reproductive performances [8,9].
Canaries (Serinus canaria) are Fringillidae songbirds appreciated for their voice, colors, and gentle nature. Kept as a pet and increasingly popular, they are receiving more and more attention, making their breeding profitable [10,11]. Canaries are non-migratory birds, whose reproductive cycle is composed of three phases: winter/nonbreeding, breeding, and molt [12]. Reproductive disorders of canaries include egg- binding, dystocia, ovarian cysts, and bacterial infections. Klebsiella, Escherichia, Pantoea, Bacillus and Staphylococcus are reported as the main responsible for bacterial disease conditions [13,14].
So far, few data are available on the gut microbial communities of canaries (Serinus canaria domesticus), but none in relation to reproduction [15,16]. Thus, this study aimed to outline the gut bacterial microbiota of healthy female canaries throughout one whole reproductive cycle, evaluating possible shifts in microbial communities between each phase.
2. Materials and Methods
2.1. Sampling
A total of 12 female Serinus canaria domesticus were included in the study. They were all color canaries, aged between 18 and 24 months. The breeding group consisted of 120 canaries housed in battery cages (60 x 32 x 40 cm). The environmental temperature was controlled in winter, always above 15°C, with 55-70% relative humidity. The canaries were fed with commercial seed mash. Supplements containing vitamins, mineral salts and cuttlefish bones were given during the mating period, while polyunsaturated fatty acids (PUFA) were added to the diet during the molting period. Antibiotics were administered only when disease occurred, and the bacteriological origin was identified. In such cases, a bacteriological culture and an antibiogram were performed to select the most appropriate therapy. No antibiotics were given during the trial. A clinical evaluation of birds in the cage was performed by an experienced physician on all the involved subjects before each sampling. Quality of the feathers, nares, beak, eyes, vent, and feet were regarded as criteria for health assessment. The canaries were sampled three times between July and November 2022. The first sample was taken during the parental case phase, the second during the molting period and the third during the resting phase before the start of a new reproductive cycle. A total of 35 samples were collected, as one of the canaries died before the last sampling. (Table 1).
Prior to each collection, dry heat sterilized waxed paper was placed on the bottom of the cages. Freshly deposited feces were collected from the waxed paper using a disposable sterile scalpel blade (a new blade was used for each collection) and transferred to cryogenic vials (Thermo-Fisher Scientific, Waltham, MA, US). The vials were immediately placed in a cryo-container filled with liquid nitrogen to prevent sample alteration.
2.2. DNA extraction
Total genomic DNA was extracted under a laminar flow cabinet. A commercial kit for DNA isolation was used according to the manufacturer’s instructions (Exgene™ Stool DNA mini, Seoul, Korea) and stored at -20°C until use. DNA concentration was assessed by Qubit fluorometer (Invitrogen), and samples were normalized at 10 ng/µL concentration.
2.3. 16S rRNA sequencing
V3-V4 region of 16S rRNA gene was amplified using the following primers: F, 5′- CCTACGGGNGGCWGCAG -3′, and R, 5′- GACTACHVGGGTATCTAATCC -3′. Primers were modified with forward and reverse overhangs (Forward overhang: 5’ TCGTCGGCAGCGTCAGATGTGTATAAGAGACAG-[locus specific sequence]; Reverse overhang: 5’ GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAG-[locus specific sequence]) necessary for dual index library preparation. For more details see the Illumina MiSeq protocol (16S Metagenomic Sequencing Library protocol n. 15044223 Rev. B). Sequencing was performed on Illumina MiSeq using a 2X300 flow cell V3 chemistry.
2.4. Data analysis
Bacterial microbiota analysis was performed with QIIME 2 2021.11 [17]. Q2 demux plugin was used to demultiplex raw sequences. Quality filter was applied by means of q2-demux plugin and denoising was carried out with DADA2 via q2-dada2 [18]. The amplicon sequences variants (ASVs) were then aligned via q2-alignment with mafft [19]. Aligned sequences were used to produce an approximately-maximum-likelihood phylogenetic tree with FastTree2 via q2-phylogeny [20]. Alpha-diversity metrics, namely Chao1, Faith’s Phylogenetic Diversity, Evenness, Observed Features and Simpson and Shannon Indexes were used [21,22,23,24,25]. Beta diversity metrics were estimated to assess differences between the groups A, B and C. In particular, weighted UniFrac [26], unweighted UniFrac [27], Jaccard distance and Bray-Curtis dissimilarity [28,29], were obtained using q2-diversity. Silva v138.1 was used as a reference for taxonomic annotation of ASVs [30,31]. Classification of the reads had 0.96 precision to the genus level, Recall of 0.93 and F-Measure of 0.95. Statistical computing and visualization were performed in R v4.1 environment [32]. Permutational multivariate analysis of variance (PERMANOVA) test was used to evaluate differences in gut bacterial communities between groups based on 1000 permutations [33]. Results were considered statistically relevant when p value was below 0.05.
3. Results
3.1. Sequencing results and GBC composition
A total of 34 samples were included in the final analysis, due to insufficient DNA amount in one sample. Thus, the groups A, B, and C consisted of 12, 11 and 11 samples respectively. From a minimum of 12,126 to a maximum of 104,841 features per sample were observed, with a total frequency of 2,133,870. In general, 4179 sequences were identified, with an average length of 392.08, with a minimum length of 273 and a maximum length of 448. Globally, 171 orders were assigned within the total samples. The most abundant orders were Lactobacillales (68,96%), Enterobacterales (11,64%), Bacillales (3,67%), Burkholderiales (3,10%), and Staphylococcales (1,50%), accounting for 88,87% of the total reads (Figure 1).
A total of 333 families were identified. The families with the highest relative abundance were Lactobacillaceae, Erwiniaceae, Yersiniaceae, Bacillaceae, Burkholderiaceae, Staphylococcaceae, Rhodobaceraceae, Pseudomonadaceae, Sphingomonadaceae, and Streptococcaceae (Figure 2).
At the genus level, 787 genera were found, with Ligilactobacillus, Pantoea, Serratia, Bacillus, Staphylococcus, Ralstonia, and Pseudomonas being the most observed. In terms of identified species, Ligilactobacillus aviarius, formerly Lactobacillus aviarius [34], was by far the most represented, its feature being found 1.363.443 times out of a total of 2.133.870 global features (63.89%). Lactobacillales were found in all 34 examined samples, and L. aviarius in 32 out of 34 samples.
3.2. Alpha diversity
Alpha diversity was assessed by means of Chao1, Shannon and Simpson’s indexes (Figure 3).
Pielou’s Evenness, Faith phylogenetic diversity, Observed Features, and Shannon indexes were used to assess phylogenetic dissimilarity within and between the groups (Table 2).
The comparison of the obtained values yielded p values respectively of 0.121, 0.013, 0.006 and 0.055 (p<0.05 was considered statistically significant). More in detail, Pielou’s Evenness index comparison between groups A, B, and C suggests that there is statistically relevant difference of number and abundance of the taxa between the communities, only when comparing A and C (p=0.048), (A vs B 0.622, B vs C 0.122). As concerns Phylogenetic diversity (Faith), the phylogenetic distance between the communities belonging to the groups was significant. In particular, group B clustered separately from A and C, having a lower phylogenetic distance between its community components than the other two groups (A vs B 0.026, A vs C 0.218, B vs C 0.009). When considering Observed features (i.e., richness within each group), B richness is lower than the other groups, especially lower than C (p=0.009), (A vs B 0.022; A vs C 0.056; B vs C 0.009). Shannon diversity, which accounts both for diversity and relative abundance of the taxa composing a community, showed a trend in diversity between groups. Pairwise comparison highlighted meaningful differences when comparing A and C, and B and C (A vs B 0.423; A vs C 0.042; B vs C 0.045). Briefly, the group clustering more separately from the others is C, which showed statistically significant differences, especially when compared to B.
3.2 Beta diversity
Beta diversity significance was estimated through Bray-Curtis dissimilarity and UnWeighted Unifrac by Permanova analysis (Figure 4).
Comparisons according to Bray-Curtis dissimilarity yielded a global p value equal to 0.004 among the three groups, while in single comparisons, the major distance of C with regards to the other groups was more striking (A vs B 0.863; A vs C 0.002; B vs C 0.011), meaning that group C has a more different community composition with respect to A and B. UnWeighted Unifrac p value was equal to 0.0009, stating a meaningful difference between the overall composition of the three groups (pairwise results A vs B 0.001; A vs C 0.005; B vs C 0.003), thus accounting both for phylogenetic distance and presence of taxa (Table 3).
The difference between the three groups is attributable to a highest relative abundance of Legionellales and Babeliales at the order level in group A, followed by mild presence in group B and very few in group C (W 133 and 122 respectively). Vermiphilaceae presence in the three groups followed the same pattern as Babeliales at the order level (W 250). In total, 10 out of 13 samples containing Vermiphilaceae belonged to group A, and the remaining 3 were specimens collected from individuals that tested positive for the same family during the parental care phase sampling. More in detail, one canary showed the presence of Vermiphilaceae during parental care and molting phases (1.359% vs 0.053%), and another during all the three phases (6.183% vs 0.190% vs Vermiphilaceae<0.05%). The same was assessed at the genus level (Vermiphilaceae undetermined genus, W 603). At genus level, also Proteus turned out to be determining in the statistical difference between groups, its feature being consistently more observed in group C (W 535). Globally, Proteus sp. was found in 10 out of 34 samples, 8 of which belonged to group C (resting phase), and 2 to group A (parental care phase). The two samples which proved positive for Proteus sp. presence during the parental care phase, were as well positive during the molting phase, and while one showed a reduction in Proteus sp. relative frequency (0.050% vs 0.021%), the other showed an increase (0.102% vs 2.611%).
4. Discussion
The present study provides robust data on the gut bacterial communities of healthy female canaries throughout one reproductive cycle. It was observed a significant shift between three reproductive phases (i.e., parental care, molting and resting phase).
In all the samples examined, Lactobacillaceae was the most consistent family of the gut bacterial microbiota. Lactobacillaceae are recognized as a relevant component of the fecal and cloacal bacterial microbiota in avian hosts, and they were so far found to have a major role in the gut microbiota of many vertebrates. In humans, Lactobacillaceae represent the 1-2% of the overall distal gut population, and despite not being as numerous as in other organisms, species and genotypes belonging to the genus Lactobacillus were proposed as gut health biomarkers [35,36]. Notably, in this study a consistent part of the bacterial communities observed was composed of L. aviarius. The presence and high percentages of L. aviarius in almost all the examined samples suggests its common presence in canaries’ feces and designate it as the main component of the “core” bacterial microbiota. In general, such bacterial components are regarded as a marker of a healthy community [37]. According to studies carried out in vitro, a relationship was suggested between Lactobacillus genus and an improved intestinal barrier function, both due to the increased mucin secretion and to the promotion of goblet cells proliferation [38,39]. In healthy layer hens, L. aviarius was among the predominant species during the laying peak. Also, Lactobacillus species (including L. aviarius) were used as a probiotic, demonstrating the in vivo improvement of intestinal absorption via an increased number of villus wrinkles [40]. It could be assumed that L. aviarius could represent a benchmark for healthy gut bacterial communities of canaries as well, though, as mentioned previously, more specific studies for this species are needed. Our findings on gut bacterial communities’ composition are consistent with previous studies carried out on many avian species, including pheasants, parrots, and chickens [36,41]. In canaries, fecal bacterial microbiota was analyzed in two papers. The first was carried out on 6 canaries’ flocks pooled feces, and family Lactobacillaceae was observed in all the examined flocks and ranged approximately from 10% to 90% of the overall families. Such variability was attributed to diet variations between flocks [15]. The second was carried out on 44 canaries from the same breed in relation to Macrorhabdus ornithogaster infection. The genus Lactobacillus was found to be more abundant in infected birds than in uninfected ones (32% vs. 6%), maybe due to infection-dependent increase in gastric pH, which possibly favored Lactobacillus proliferation [16]. In the present study, all the sampled canaries were clinically healthy, and same feed was administered to all of them throughout the study.
When analyzing Alpha diversity indexes, higher values were observed for group C (resting phase), especially with respect to B (molt), which showed the lowest Alpha diversity indexes values. The reduction in gut bacterial communities’ phylogenetic diversity (Faith), observed features, and relative abundance (Table 1) during this phase can be ascribed to the physiological alterations that come with the molt. More in detail, changes in thyroid hormones, gonadal steroid hormones, and prolactin are involved in the molt process. Among hormones, prolactin seems to play a major role, decreasing gradually along with the light hours and eventually triggering the start of the post-breeding molt. Increase in basal metabolism with respect to non-molting periods, protein synthesis, bone and lipids metabolism and immune system functionality are affected during molt [42,43,44,45]. Feather replacement and changes in tissue metabolism are the main feature of molt, which make it energy consuming for the avian host. In many species, molt is avoided during periods of high energy demand and for this reason it generally follows the reproductive phase [46]. Molt was linked to alterations in the gut bacterial communities’ composition, and a shift towards potentially pathogenic bacteria is reported both in wild birds and poultry. Such changes depend on reduction in light hours and on the fasting/caloric restriction laying hens and wild birds face during molt [47,48,49]. In pet passerines, which do not undergo feed reduction, molt starts in response to changes in daylight hours [50]. Thus, the changes in gut bacterial microbiota observed in the present studies were probably related to molt per se.
Beta diversity analysis showed also significant differences among groups. The diversity pattern was similar to Alpha diversity, although the gap between groups was even more pronounced, both when considering Bray-Curtis dissimilarity (p≤0.01 in comparisons involving group C) and UnWeighted Unifrac (p≤0.005 in all groups comparisons). Therefore, communities’ composition was more different in group C, and the combination of taxonomic composition and phylogenetic distance were significantly different between all three groups. More diverse microbiota has been associated with a better health status of the host. In fact, more diverse ecosystems have a certain degree of redundancy which allows compensation of function whenever a species is lost or removed [51]. The findings of the present study could be suggestive of the importance of the resting period for restoring an optimal bacterial microbiota of the host before the start of a new reproductive cycle.
Regarding gut communities’ composition, differences among groups were largely due to a marked decrease in the orders Babeliales and Legionellales during molt and resting phase. Legionellales were so far found mostly in invertebrates’ gut microbiota, such as clams and ascidians. In the latter, Legionellales are possibly involved in compensative mechanisms during starvation [52,53]. On the other hand, Legionellales are globally distributed in the environment, being found in soil, freshwater, and seawater. Nonetheless, little is known on species and diversity of the bacteria belonging to this order. Apart from pathogenic members of Legionellales, other species have received little attention, were not sequenced, and go therefore unnoticed in 16S rRNA analysis [54]. Within Babeliales order, family Vermiphilaceae decrease was responsible for the shift in microbiota composition. Vermiphilaceae family has been mentioned so far in studies investigating the gut microbiota of lizards (Sceloporus spp.), giant river prawn Macrobrachium rosenbergii, and ascidian (Halocynthia roretzi), in which was put in relation to age, growth rate and season respectively [53,55,56]. Nevertheless, yet little is known on its role in the gut bacterial microbiota dynamics and its ecology in living host communities. At genus level, an increase in Proteus relative abundance during the resting phase was observed. Globally, Proteus sp. was found mostly in group C (8 out 10 samples positive for Proteus sp.). In two canaries, Proteus sp. was observed both during the parental care phase and during the resting phase, but no single individual showed its presence during molt. The restoration of a genus during the resting phase with respect to parental care stage, along with an increase in Alpha and Beta diversity could suggest the re-establishment of the gut bacterial microbiota after molt. Proteus spp. are regarded as common commensals of the gastrointestinal tract microbiota, and in avians, the presence of Proteus sp. was assessed in the gut microbiota of clinically healthy bird species including passerines and psittacines [57,58,59,60,61].
In general, shifts along the reproductive cycle were observed in many passeriformes. In tree swallows (Tachycineta bicolor), microbiota changes between nest building and incubation, and in rufous-collared sparrows (Zonotrichia capensis) fluctuations of cloacal microbiota composition were associated to the breeding condition of the host [62,63]. Our findings are consistent with the available literature, although much land remains to be conquered on microbiota composition in avian hosts.
As regards supplements administration, the action of dietary intake of PUFA on the gut microbiota is uncertain. PUFA are regarded as prebiotics by some authors, while other studies stated that dietary intake of fatty acids may change the fatty acid composition of the gut wall and therefore alter the attachment site of bacteria, promoting or inhibiting microbial colonization [64,65]. Lastly, other researchers found no correlation at all between PUFA administration and microbiota shifts [66]. In the present study, canaries belonging to the examined flock were routinely given dietary PUFA supplementation as an aid for feather regrowth during molt. Thus, all the canaries involved in the study received PUFA, and no control group was made to examine the effect of dietary augmentation of fatty acids on gut microbial communities. Nevertheless, it was not the aim of the present investigation, although it would be interesting to assess possible impact of PUFA on the gut microbiota of canaries.
Finally, as for the choice of the kind of specimen, feces represent a non-invasive sampling method that can be repeated with no consequences for the host. Furthermore, unnecessary handling of the animals was avoided. Although maybe not fully representative of all the ecological niches of the intestine’s bacterial communities (i.e., duodenum, jejunum, ileum, cecum, colon, and cloaca), feces can be used to approach the microbial components of the gut, especially when instantly frozen at least at -80°C [67,68]. It is noteworthy pointing out that feces in birds go through cloaca, which is a compartment gathering the bacterial components from gastrointestinal, reproductive, and urinary systems, and is therefore considered relevant to the health of all the systems involved [36,69].
In conclusion, our study provides a useful reference for the analysis of microbiota changes in the reproductive tract of avian species. The time factor was considered to assess variability in the bacterial communities’ composition, showing that the gut bacterial microbiota is responsive to breeding phases in canaries. This paper lays the groundwork for a clearer understanding of canaries’ ecology and physiology. The interaction between host and bacterial microbiota may help shedding a light on the causes of female canaries’ infertility, with important consequences for multiple fields including reproductive science, conservation, and commercial breeding of canaries. Future investigations will be possibly focused on the evaluation of male gut bacterial microbiota, aiming to describe male canaries’ gut microbial communities, assess the presence of microbial fluctuations along one male reproductive cycle and to make a comparison with females. It would also be interesting to repeat the sampling along with a hormonal evaluation of the individuals, although it would involve handling of the canaries and therefore be stressful for the subjects involved.
Author Contributions
Conceptualization, G.M. and G.T.; methodology, G.M.; software, P.G.T.; validation, A.S. and P.G.T.; formal analysis, J.H.; investigation, A.S.; resources, G.T.; data curation, J.H.; writing—original draft preparation, J.H.; writing—review and editing, G.M.; visualization, A.S.; supervision, G.T.; project administration, P.G.T.; funding acquisition, G.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research and APC were funded by FOI (Federazione Ornicoltori Italiani Onlus), Via Caorsana 94 Loc. Le Mose, 29122 Piacenza (Italy). http://www.foi.it VECO 00067.
Data Availability Statement
Data are available from the authors upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Qi, X.; Yun, C.; Pang, Y.; Qiao, J. The impact of the gut microbiota on the reproductive and metabolic endocrine system. Gut microbes 2021, 13, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Franasiak, J.M.; Scott, R.T. Introduction: microbiome in human reproduction. Fertil Steril 2015, 104, 1341–1343. [Google Scholar] [CrossRef] [PubMed]
- Kwa, W.T.; Sundarajoo, S.; Toh, K.Y.; Lee, J. Application of emerging technologies for gut microbiome research. Singapore med j 2023, 64, 45–52. [Google Scholar] [CrossRef]
- Singer, E.; Bushnell, B.; Coleman-Derr, D.; Bowman, B.; Bowers, R. M.; Levy, A.; Gies, E.A.; Cheng, J.F.; Copeland, A.; Klenk, H.P.; Hallam, S.J.; Hugenholtz, P.; Tringe, S.G.; Woyke, T. High-resolution phylogenetic microbial community profiling. ISME J 2016, 10, 2020–2032. [Google Scholar] [CrossRef]
- Williams, C.L.; Garcia-Reyero, N.; Martyniuk, C.J.; Tubbs, C.W.; Bisesi, J.H. Jr. Regulation of endocrine systems by the microbiome: Perspectives from comparative animal models. Gen Comp Endocrinol. 2020, 292, 113437. [Google Scholar] [CrossRef]
- Góngora, E.; Elliott, K.H.; Whyte, L. Gut microbiome is affected by inter-sexual and inter-seasonal variation in diet for thick-billed murres (Uria lomvia). Sci Rep 2021, 11, 1200. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Chen, J.; Liu, K.; Tang, M.; Yang, Y. The avian gut microbiota: Diversity, influencing factors, and future directions. Front Microbiol 2022, 13, 934272. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.; Qi, G.H.; Wang, J.; Zhang, H.J.; Qiu, K.; Wu, S.G. Intestinal microbiota of layer hens and its association with egg quality and safety. Poult sci 2022, 101, 102008. [Google Scholar] [CrossRef]
- Ran, J.; Wan, Q.H.; Fang, S.G. Gut microbiota of endangered crested ibis: Establishment, diversity, and association with reproductive output. PLoS One. 2021, 16, e0250075. [Google Scholar] [CrossRef]
- Arnaiz-Villena, A.; Areces, C.; Ruiz-Del-Valle, V. El origen de los canarios. Revta Ornitol Práct 2012, 53, 3–11. [Google Scholar]
- Boseret, G.; Losson, B.; Mainil, J.G.; Thiry, E.; Saegerman, C. Zoonoses in pet birds: review and perspectives. Vet Res 2013, 44, 36. [Google Scholar] [CrossRef] [PubMed]
- Wingfield, J.C. Organization of vertebrate annual cycles: implications for control mechanisms. Philos Trans R Soc Lond B Biol Sci 2008, 363, 425–441. [Google Scholar] [CrossRef] [PubMed]
- Di Francesco, C.E.; Todisco, G.; Montani, A.; Profeta, F.; Di Provvido, A.; Foschi, G.; Persiani, T.; Marsilio, F. Reproductive disorders in domestic canaries (Serinus canarius domesticus): a retrospective study on bacterial isolates and their antimicrobial resistance in Italy from 2009 to 2012. Vet Ital 2018, 54, 169–174. [Google Scholar] [CrossRef]
- Rosen, L.B. Topics in Medicine and Surgery. Avian Reproductive Disorders. J Exot Pet Med 2012, 21, 124–131. [Google Scholar] [CrossRef]
- Garcia-Mazcorro, J.F.; Castillo-Carranza, S.A.; Guard, B.; Gomez-Vazquez, J.P.; Dowd, S.E.; Brigthsmith, D.J. Comprehensive Molecular Characterization of Bacterial Communities in Feces of Pet Birds Using 16S Marker Sequencing. Microb Ecol 2017, 73, 224–235. [Google Scholar] [CrossRef]
- Robino, P.; Ferrocino, I.; Rossi, G.; Dogliero, A.; Alessandria, V.; Grosso, L.; Galosi, L.; Tramuta, C.; Cocolin, L.; Nebbia, P. Changes in gut bacterial communities in canaries infected by Macrorhabdus ornithogaster. Avian Pathol 2019, 48, 111–120. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; Bai, Y.; Bisanz, J.E.; Bittinger, K.; Brejnrod, A.; Brislawn, C.J.; Brown, C.T.; Callahan, B.J.; Caraballo-Rodríguez, A.M.; Chase, J.; Cope, E.K.; Da Silva, R.; Diener, C.; Dorrestein, P.C.; Douglas, G.M.; Durall, D.M.; Duvallet, C.; Edwardson, C.F.; Ernst, M.; Estaki, M.; Fouquier, J.; Gauglitz, J.M.; Gibbons, S.M.; Gibson, D.L.; Gonzalez, A.; Gorlick, K.; Guo, J.; Hillmann, B.; Holmes, S.; Holste, H.; Huttenhower, C.; Huttley, G.A.; Janssen, S.; Jarmusch, A.K.; Jiang, L.; Kaehler, B.D.; Kang, K.B.; Keefe, C.R.; Keim, P.; Kelley, S.T.; Knights, D.; Koester, I.; Kosciolek, T.; Kreps, J.; Langille, M.G.I.; Lee, J.; Ley, R.; Liu, Y.X.; Loftfield, E.; Lozupone, C.; Maher, M.; Marotz, C.; Martin, B.D.; McDonald, D.; McIver, L.J.; Melnik, A.V.; Metcalf, J.L.; Morgan, S.C.; Morton, J.T.; Naimey, A.T.; Navas-Molina, J.A.; Nothias, L.F.; Orchanian, S.B.; Pearson, T.; Peoples, S.L.; Petras, D.; Preuss, M.L.; Pruesse, E.; Rasmussen, L.B.; Rivers, A.; Robeson, M.S.; Rosenthal, P.; Segata, N.; Shaffer, M.; Shiffer, A.; Sinha, R.; Song, S.J.; Spear, J.R.; Swafford, A.D.; Thompson, L.R.; Torres, P.J.; Trinh, P.; Tripathi, A.; Turnbaugh, P.J.; Ul-Hasan, S.; van der Hooft, J.J.J.; Vargas, F.; Vázquez-Baeza, Y.; Vogtmann, E.; von Hippel, M.; Walters, W.; Wan, Y.; Wang, M.; Warren, J.; Weber, K.C.; Williamson, C.H.D.; Willis, A.D.; Xu, Z.Z.; Zaneveld, J.R.; Zhang, Y.; Zhu, Q.; Knight, R.; Caporaso, J.G. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat Biotechnol 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: high-resolution sample inference from Illumina amplicon data. Nat Methods 2016, 13, 581–583. [Google Scholar]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic acids res 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2–approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Species Richness: Estimation and Comparison. In Wiley StatsRef: Statistics Reference Online; Balakrishnan, N., Colton, T., Everitt, B., Piegorsch, W., Ruggeri, F., Teugels, J.L. Eds.; John Wiley & Sons Ltd.: Chichester.
- UK, 2016; pp. 1–26. [CrossRef]
- Faith, D.P. Conservation evaluation and phylogenetic diversity. Biol Cons 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J Theor Biol 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst tech j 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Hamady, M.; Kelley, S.T.; Knight, R. Quantitative and qualitative beta diversity measures lead to different insights into factors that structure microbial communities. Appl Environ Microbiol 2007, 73, 1576–1585. [Google Scholar] [CrossRef]
- Lozupone, C.; Knight, R. UniFrac: a new phylogenetic method for comparing microbial communities. Appl Environ Microbiol 2005, 71, 8228–8235. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An ordination of upland forest communities of southern Wisconsin. Ecol Monogr 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Jaccard, P. The Distribution of the Flora in the Alpine Zone. New Phytol 1912, 11, 37–50. [Google Scholar] [CrossRef]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A Comprehensive Online Resource for Quality Checked and Aligned Ribosomal RNA Sequence Data Compatible with ARB. Nucleic Acids Res 2007, 35, 7188–96. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res 2013, 41, D590–96. [Google Scholar] [CrossRef]
- R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria, 2021. Available online: https://www.R-project.org/ accessed on 7th August 2023.
- Anderson, M.J. Permutational multivariate analysis of variance (PERMANOVA). In: Wiley statsref: statistics reference online; Balakrishnan, N., Colton, T., Everitt, B., Piegorsch, W., Ruggeri, F., Teugels, J.L. Eds.; John Wiley & Sons Ltd.: Chichester,.
- UK, 2017; pp. 1–15.
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O'Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; Watanabe, K.; Wuyts, S.; Felis, G.E.; Gänzle, M.G.; Lebeer, S. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int J Syst Evol Microbiol 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Huynh, U.; Zastrow, M.L. Metallobiology of Lactobacillaceae in the gut microbiome. J Inorg Biochem 2023, 238, 112023. [Google Scholar] [CrossRef]
- Mohsin Bukhari, S.; Ahmed Alghamdi, H.; Ur Rehman, K.; Andleeb, S.; Ahmad, S.; Khalid, N. Metagenomics analysis of the fecal microbiota in Ring-necked pheasants (Phasianus colchicus) and Green pheasants (Phasianus versicolor) using next generation sequencing. Saudi J Biol Sci 2022, 29, 1781–1788. [Google Scholar] [CrossRef] [PubMed]
- De Cesare, A.; Sirri, F.; Manfreda, G.; Moniaci, P.; Giardini, A.; Zampiga, M.; Meluzzi, A. Effect of dietary supplementation with Lactobacillus acidophilus D2/CSL (CECT 4529) on caecum microbioma and productive performance in broiler chickens. PLoS One 2017, 12, e0176309. [Google Scholar] [CrossRef]
- Mattar, A.F.; Teitelbaum, D.H.; Drongowski, R.A.; Yongyi, F.; Harmon, C.M.; Coran, A.G. Probiotics up-regulate MUC-2 mucin gene expression in a Caco-2 cell-culture model. Pediatr surg int 2002, 18, 586–590. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kim, S.H.; Whang, K.Y.; Kim, Y.J.; Oh, S. Inhibition of Escherichia coli O157:H7 attachment by interactions between lactic acid bacteria and intestinal epithelial cells. J microbiol biotechnol 2008, 18, 1278–1285. [Google Scholar]
- Liu, L.; Zhou, Z.; Hong, Y.; Jiang, K.; Yu, L.; Xie, X.; Mi, Y.; Zhu, S.J.; Zhang, C.; Li, J. Transplantion of predominant Lactobacilli from native hens to commercial hens could indirectly regulate their ISC activity by improving intestinal microbiota. Microb biotechnol 2022, 15, 1235–1252. [Google Scholar] [CrossRef]
- Walter, J. Ecological role of lactobacilli in the gastrointestinal tract: implications for fundamental and biomedical research. Appl Environ Microbiol 2008, 74, 4985–4996. [Google Scholar] [CrossRef] [PubMed]
- Dawson, A. Avian Molting. In Sturkie's Avian Physiology, 6th ed.; Scanes, C.G., Ed.; Academic Press: Cambridge, MA (US), 2015; pp. 907–917. [Google Scholar] [CrossRef]
- Vézina, F.; Gustowska, A.; Jalvingh, K.M.; Chastel, O.; Piersma, T. Hormonal correlates and thermoregulatory consequences of molting on metabolic rate in a northerly wintering shorebird. Physiol Biochem Zool 2009, 82, 129–142. [Google Scholar] [CrossRef]
- Kuenzel, W.J. Neurobiology of molt in avian species. Poult Sci 2003, 82, 981–991. [Google Scholar] [CrossRef]
- Smiley, K.O. Prolactin and avian parental care: New insights and unanswered questions. Horm Behav 2019, 111, 114–130. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.E. Energetics and nutrition of molt. In Avian energetics and nutritional ecology; Carey, C., Ed.; Chapman and Hall: London, UK, 1996; pp. 158–198. [Google Scholar]
- Cho, H.; Lee, W.Y. Interspecific comparison of the fecal microbiota structure in three Arctic migratory bird species. Ecol Evol 2020, 10, 5582–5594. [Google Scholar] [CrossRef] [PubMed]
- Dewar, M.L.; Arnould, J.P.; Krause, L.; Trathan, P.; Dann, P.; Smith, S.C. Influence of fasting during moult on the faecal microbiota of penguins. PLoS One. 2014, 9, e99996. [Google Scholar] [CrossRef]
- Yaqoob, M.U.; El-Hack, M.E.A.; Hassan, F.; El-Saadony, M.T.; Khafaga, A.F.; Batiha, G.E.; Yehia, N.; Elnesr, S.S.; Alagawany, M.; El-Tarabily, K.A.; Wang, M. The potential mechanistic insights and future implications for the effect of prebiotics on poultry performance, gut microbiome, and intestinal morphology. Poult Sci 2021, 100, 101143. [Google Scholar] [CrossRef]
- Coutteel, P. Veterinary aspects of breeding management in captive passerines. Semin avian exot pet med 2003, 12, 3–10. [Google Scholar] [CrossRef]
- Larsen, O.F.A.; Claassen, E. The mechanistic link between health and gut microbiota diversity. Sci Rep 2018, 8, 2183. [Google Scholar] [CrossRef]
- Offret, C.; Paulino, S.; Gauthier, O.; Château, K.; Bidault, A.; Corporeau, C.; Miner, P.; Petton, B.; Pernet, F.; Fabioux, C.; Paillard, C.; Blay, G.L. The marine intertidal zone shapes oyster and clam digestive bacterial microbiota. FEMS Microbiol Ecol 2020, 96, fiaa078. [Google Scholar] [CrossRef]
- Wei, J.; Gao, H.; Yang, Y.; Liu, H.; Yu, H.; Chen, Z.; Dong, B. Seasonal dynamics and starvation impact on the gut microbiome of urochordate ascidian Halocynthia roretzi. Anim Microbiome 2020, 2, 30. [Google Scholar] [CrossRef]
- Graells, T.; Ishak, H.; Larsson, M.; Guy, L. The all-intracellular order Legionellales is unexpectedly diverse, globally distributed and lowly abundant. FEMS Microbiol Ecol 2018, 94, fiy185. [Google Scholar] [CrossRef]
- Lan, X.; Peng, X.; Du, T.; Xia, Z.; Gao, Q.; Tang, Q.; Yi, S.; Yang, G. Alterations of the Gut Microbiota and Metabolomics Associated with the Different Growth Performances of Macrobrachium rosenbergii Families. Animals, 2023, 13, 1539. [Google Scholar] [CrossRef] [PubMed]
- Assis, B.A.; Bell, T.H.; Engler, H.I.; King, W.L. Shared and unique responses in the microbiome of allopatric lizards reared in a standardized environment. J Exp Zool A: Ecol Integr Physiol, 2023, 339, 5–12. [Google Scholar] [CrossRef]
- Cunha, M.P.V.; Guimarães, M.B.; Davies, Y.M.; Milanelo, L.; Knöbl, T. Bactérias gram-negativas em cardeais (Paroaria coronata e Paroaria dominicana) apreendidos do tráfico de animais silvestres. Braz J Vet Res Anim Sci 2016, 53, 107–111. [Google Scholar] [CrossRef]
- Davies, Y.M.; Guimarães, M.B.; Milanelo, L.; Oliveira, M.G.X.; de Gomes, V.T. de M.; Azevedo, N.P.; Cunha, M.P.V.; Moreno, L.Z.; Romero, D.C.; Christ, A.P.G.; Sato, M.I.Z.; Moreno, A.M.; Ferreira, A.J.P.; Sá, L.R.M.; Knöbl, T. A survey on gram-negative bacteria in saffron finches (Sicalis flaveola) from illegal wildlife trade in Brazil. Braz J Vet Res Anim Sci 2016, 53, 286–294. [Google Scholar] [CrossRef]
- Davies, Y.M.; Franco, L.S.; Barbosa, F.B.; Vanin, C.L.; Gomes, V.T.M.; Moreno, L.Z.; Barbosa, M.R.F.; Sato, M.I.Z.; Moreno, A.M.; Knöbl, T. Use of MALDI-TOF for identification and surveillance of gram-negative bacteria in captive wild psittacines. Braz J Biol 2021, 82, e233523. [Google Scholar] [CrossRef]
- Penner, J.L. Genus XXIX. Proteus, In Bergey's manual of systematic bacteriology, 2nd ed.; Brenner, D.J., Krieg, N.R., Staley, J.T., Garrity, G.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA (US), 2005; Volume 2, pp. 745–753. [Google Scholar]
- Wang, W. , Wang, A., Yang, Y., Wang, F., Liu, Y., Zhang, Y., Sharshov, K., Gui, L. Composition, diversity and function of gastrointestinal microbiota in wild red-billed choughs (Pyrrhocorax pyrrhocorax). Int microbiol 2019, 22, 491–500. [Google Scholar] [CrossRef]
- Escallón, C.; Belden, L.K.; Moore, I.T. The Cloacal Microbiome Changes with the Breeding Season in a Wild Bird. Integr Org Biol 2019, 1, oby009. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, J.; Hucul, C.; Reasor, E.; Smith, T.; McGlothlin, J.W.; Haak, D.C.; Belden, L.K.; Moore, I.T. Assessing age, breeding stage, and mating activity as drivers of variation in the reproductive microbiome of female tree swallows. Ecol Evol 2021, 11, 11398–11413. [Google Scholar] [CrossRef] [PubMed]
- Costantini, L.; Molinari, R.; Farinon, B.; Merendino, N. Impact of Omega-3 Fatty Acids on the Gut Microbiota. Int J Mol Sci. 2017, 18, 2645. [Google Scholar] [CrossRef]
- Fernandes, J.; Su, W.; Rahat-Rozenbloom, S.; Wolever, T.M.; Comelli, E.M. Adiposity, gut microbiota and faecal short chain fatty acids are linked in adult humans. Nutr Diabetes 2014, 4, e121. [Google Scholar] [CrossRef] [PubMed]
- Geier, M.S.; Torok, V.A.; Allison, G.E.; Ophel-Keller, K.; Gibson, R.A.; Munday, C.; Hughes, R.J. Dietary omega-3 polyunsaturated fatty acid does not influence the intestinal microbial communities of broiler chickens. Poult sci 2009, 88, 2399–2405. [Google Scholar] [CrossRef]
- Fouhy, F.; Deane, J.; Rea, M.C.; O'Sullivan, Ó.; Ross, R.P.; O'Callaghan, G.; Plant, B.J.; Stanton, C. The effects of freezing on faecal microbiota as determined using MiSeq sequencing and culture-based investigations. PLoS ONE 2015, 10, e0119355. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Jin, G.; Wang, G.; Liu, T.; Liu, X.; Wang, B.; Cao, H. Current Sampling Methods for Gut Microbiota: A Call for More Precise Devices. Front Cell Infect Microbiol 2020, 10, 151. [Google Scholar] [CrossRef] [PubMed]
- Hadley, T.L. Disorders of the psittacine gastrointestinal tract. Vet Clin North Am Exot Anim Pract 2005, 8, 329–349. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Gut bacterial communities of canaries’ feces at the order level. Barplot showing the main bacterial composition of the female canaries’ fecal community during parental care (A), molting (B), and resting phase (C) at the order level. Only orders with relative abundance >1 are shown singularly.
Figure 1.
Gut bacterial communities of canaries’ feces at the order level. Barplot showing the main bacterial composition of the female canaries’ fecal community during parental care (A), molting (B), and resting phase (C) at the order level. Only orders with relative abundance >1 are shown singularly.
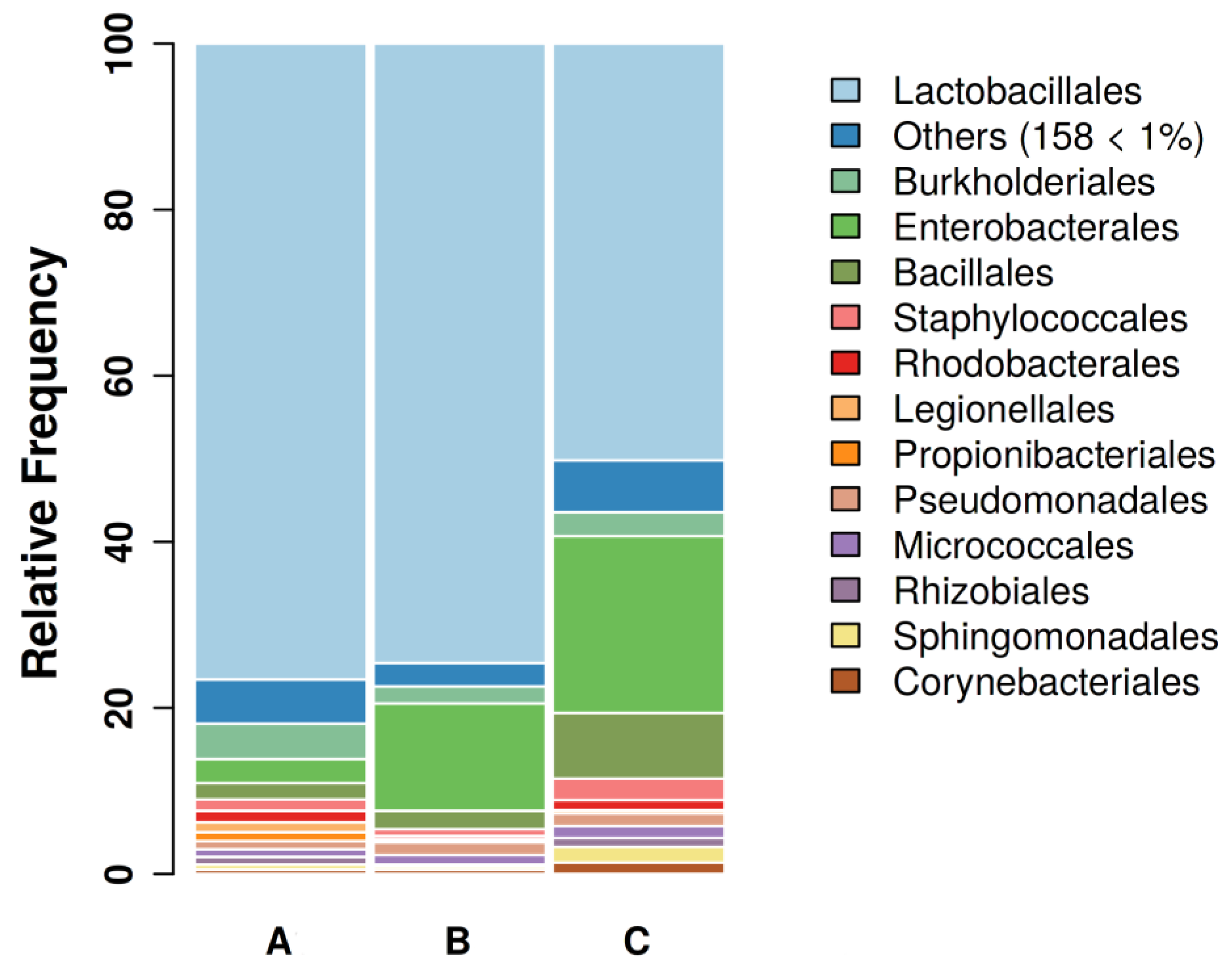
Figure 2.
Gut bacterial communities of canaries’ feces at the family level. Barplot showing the main bacterial composition of the female canaries’ fecal community during parental care (A), molting (B), and resting phase (C) at the family level. Only families with relative abundance >1 are shown singularly.
Figure 2.
Gut bacterial communities of canaries’ feces at the family level. Barplot showing the main bacterial composition of the female canaries’ fecal community during parental care (A), molting (B), and resting phase (C) at the family level. Only families with relative abundance >1 are shown singularly.
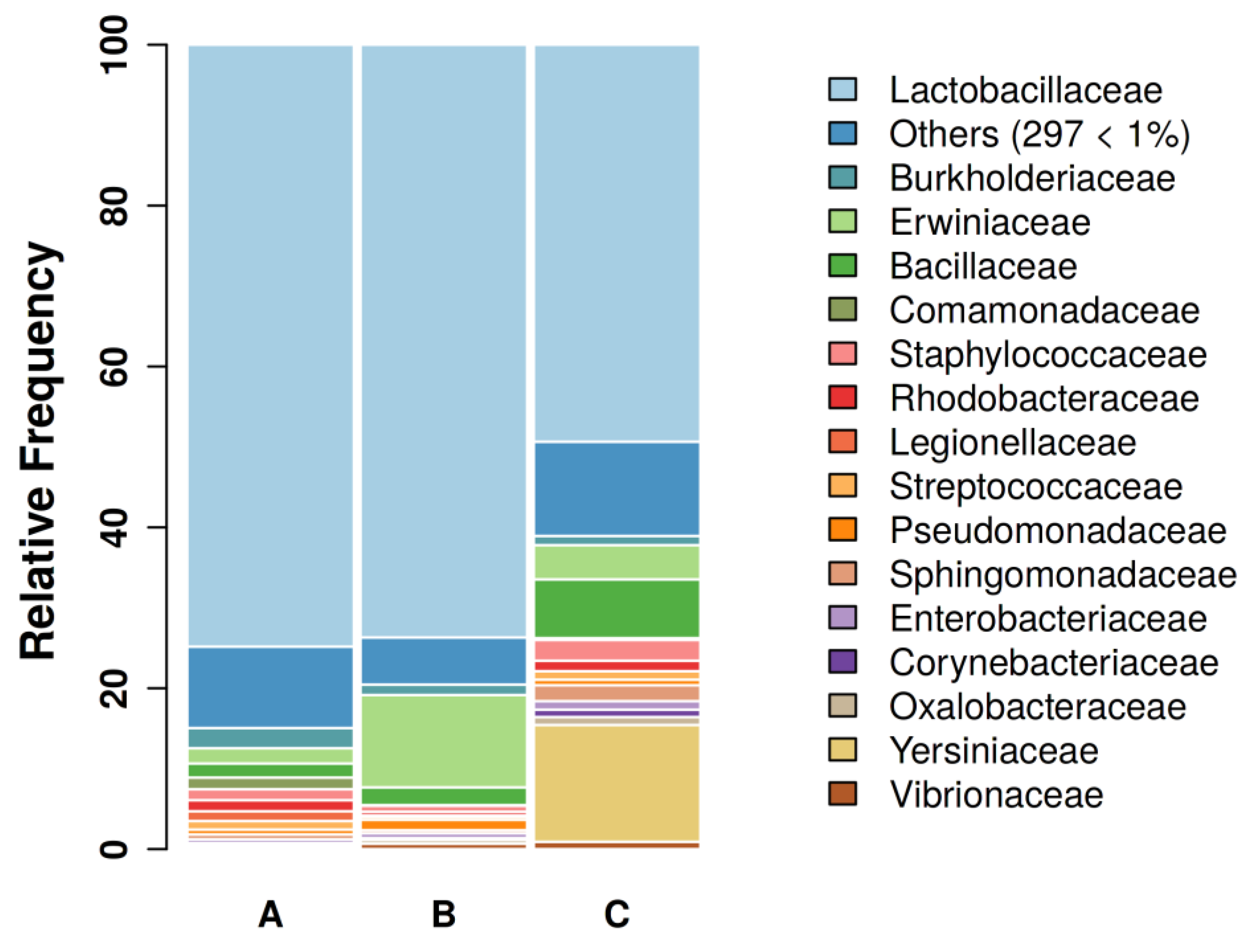
Figure 3.
Alpha diversity of gut bacterial communities of female canaries along one reproductive cycle is shown according to Chao1, Shannon and Simpson’s indexes. Observed species and microbiota diversity are represented in the Chao1 graphic, and combined species and abundance are shown in the Shannon and Simpson graphics. Data are divided according to reproductive phase, i.e., parental care (A), molting (B), and resting phase (C).
Figure 3.
Alpha diversity of gut bacterial communities of female canaries along one reproductive cycle is shown according to Chao1, Shannon and Simpson’s indexes. Observed species and microbiota diversity are represented in the Chao1 graphic, and combined species and abundance are shown in the Shannon and Simpson graphics. Data are divided according to reproductive phase, i.e., parental care (A), molting (B), and resting phase (C).
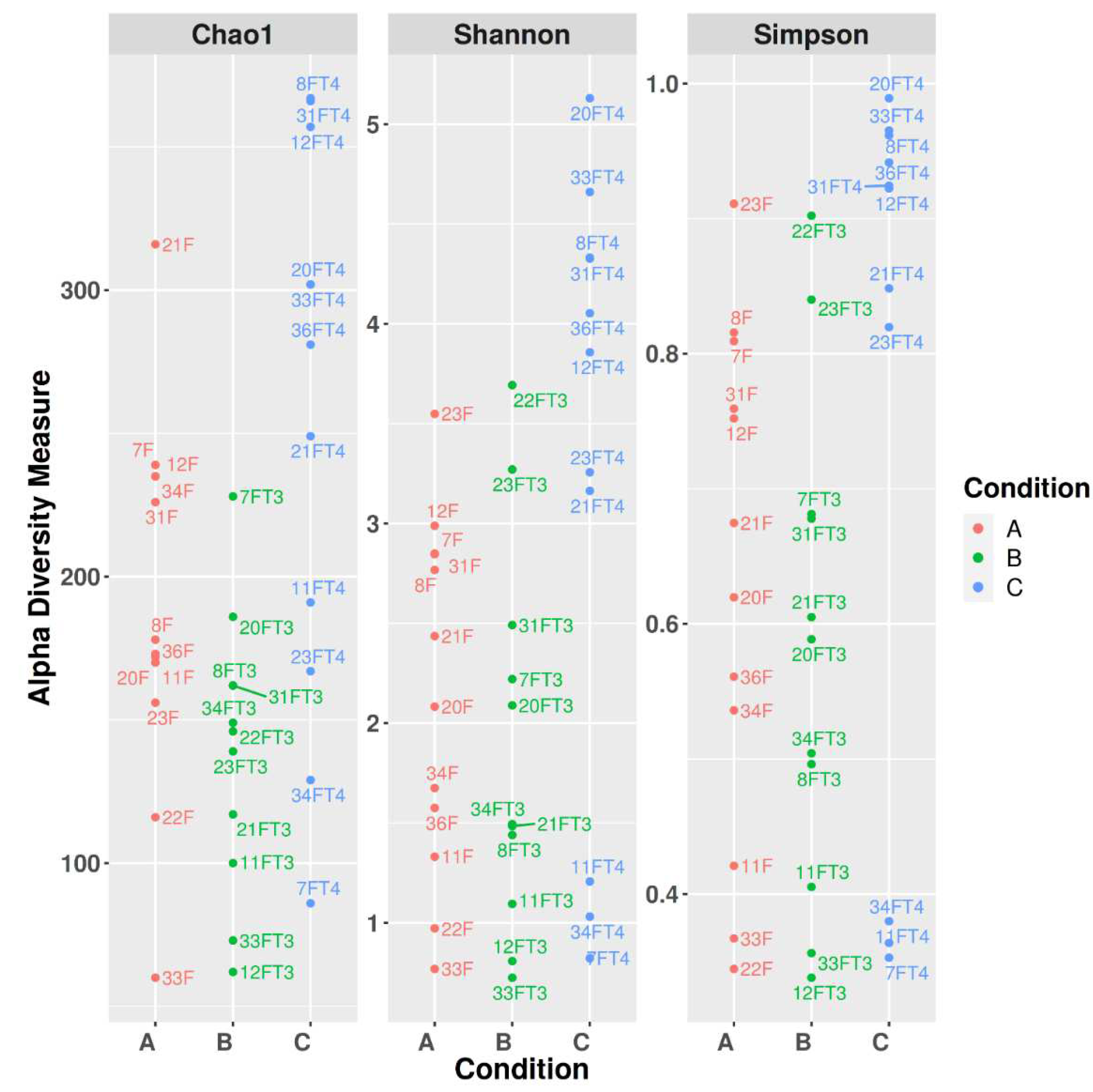
Figure 4.
Box and whiskers plots illustrating the Beta diversity based on Bray-Curtis and UnWeighted Unifrac diversity. Groups are represented in different colors, each point being the graphical representation of the distance comparison among the three groups A, B, and C. The lower whiskers represent the values from the minimum value up to the first quartile, and the upper whiskers the values ranging from the third quartile and the maximum value.
Figure 4.
Box and whiskers plots illustrating the Beta diversity based on Bray-Curtis and UnWeighted Unifrac diversity. Groups are represented in different colors, each point being the graphical representation of the distance comparison among the three groups A, B, and C. The lower whiskers represent the values from the minimum value up to the first quartile, and the upper whiskers the values ranging from the third quartile and the maximum value.
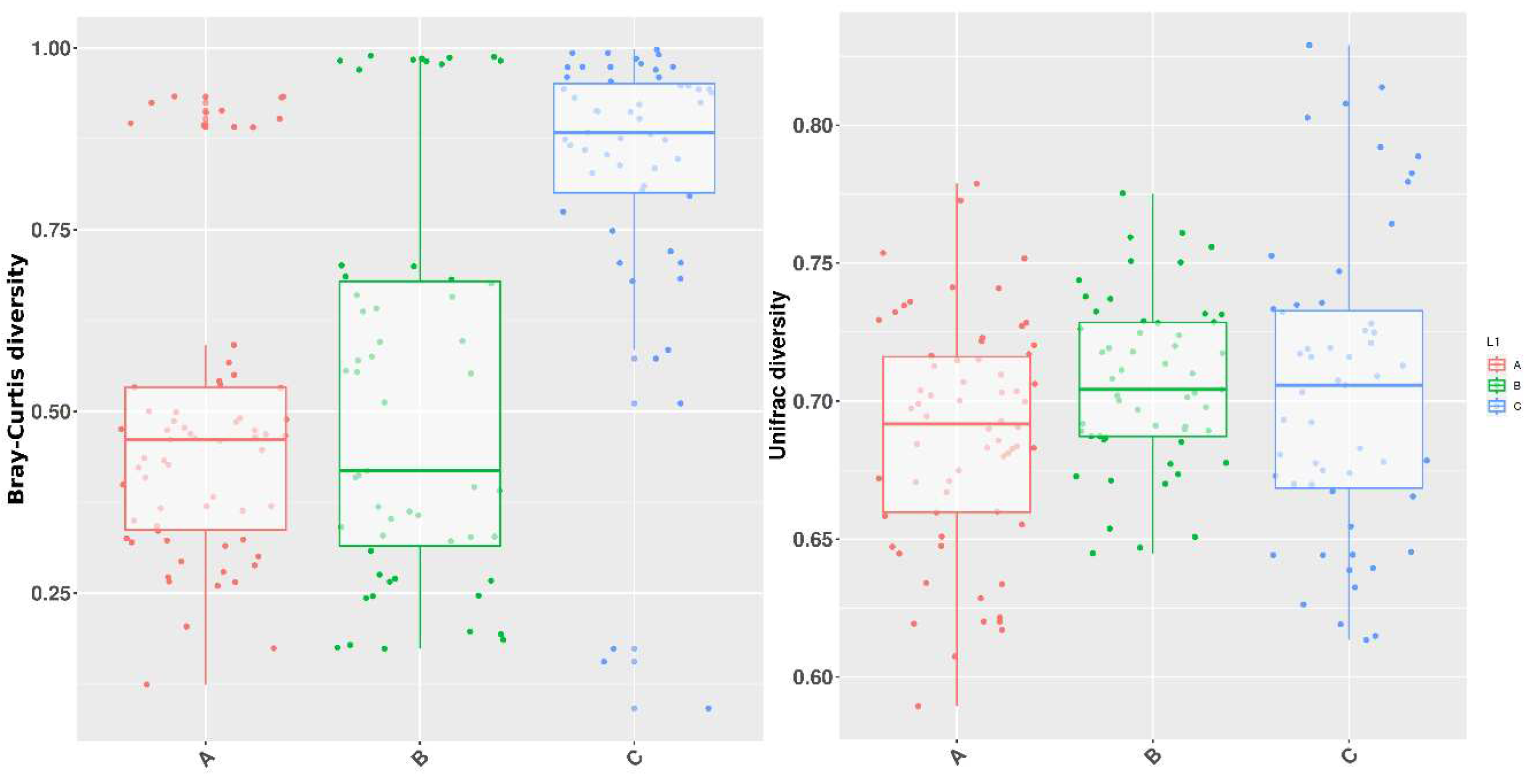

Table 1.
The number of samples taken for each phase of the reproductive cycle is shown.
| Groups | A | B | C |
|---|---|---|---|
| Reproductive phase | Parental care | Molting | Rest |
| N. of samples | 12 | 12 | 11 |
Table 2.
Alpha diversity indexes comparisons between groups. P value for each comparison (A vs B, A vs C and B vs C) is reported for each Alpha diversity index. Values of p<0.05 are shown in bold.
Table 2.
Alpha diversity indexes comparisons between groups. P value for each comparison (A vs B, A vs C and B vs C) is reported for each Alpha diversity index. Values of p<0.05 are shown in bold.
| A vs B | A vs C | B vs C | |
|---|---|---|---|
| Pielou’s Evenness | 0.622461 | 0.048900 | 0.122800 |
| Faith phylogenetic diversity | 0.026716 | 0.218355 | 0.009493 |
| Observed Features | 0.022741 | 0.056219 | 0.009453 |
| Shannon | 0.423656 | 0.042254 | 0.045201 |
Table 3.
Beta diversity indexes comparisons between groups. P value for each comparison (A vs B, A vs C and B vs C) is reported for each Alpha diversity index. Values of p<0.05 are shown in bold. .
Table 3.
Beta diversity indexes comparisons between groups. P value for each comparison (A vs B, A vs C and B vs C) is reported for each Alpha diversity index. Values of p<0.05 are shown in bold. .
| A vs B | A vs C | B vs C | |
|---|---|---|---|
| Bray-Curtis dissimilarity | 0.863137 | 0.002997 | 0.011988 |
| UnWeighted Unifrac | 0.001998 | 0.005994 | 0.003996 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



