
Submitted:
20 July 2023
Posted:
21 July 2023
You are already at the latest version
Abstract
Background: Alkylresorcinols (ARs) are polyphenolic compounds with a wide spectrum of biological activities and potentially implicated in the regulation of host metabolism. The present study is aimed to establish whether ARs can be produced by human gut microbiota and to evaluate alterations in the content of ARs in stool samples of C57BL and db/db and LDLR (-/-) mice in dependence on diet specifications and olivetol (5-n-pentylresorcinol) supplementation in order to estimate a regulatory potential of ARs. Methods: The quantita-tive analysis of ARs levels in mice stool samples was performed using gas chromatography with mass spectrometric detection; fecal microbiota transplantation (FMT) from human donor to germ-free mice was performed to explore whether the intestinal microbiota could produce ARs molecules. Results: A significant increase in the amounts of individual mem-bers of ARs homologues in stool samples was revealed in 14 days after FMT. 5-n-Pentylresorcinol supplementation to a regular-chow diet influences the amounts of sev-eral ARs in stool of C57BL/6 and LDLR (-/-), but not db/db mice. Conclusions: For the first time, we have shown that several ARs can be produced by the intestinal microbiota. Taking into account, that AR concentrations in human serum are correlated with LPS levels and with microbiota diversity indexes in human stool samples, it can be assumed that ARs may serve as quorum-sensing molecules, influencing gut microbiota composition and host me-tabolism as well.
Keywords:
alkylresorcinols
; olivetol
; gut microbiota
; fecal microbiota transplantation
; endocannabinoid system
1. Introduction
In recent years, we observe an increasing interest in 'host – gut microbiota' relationship research. The microbiota is a collection of various microorganisms (mainly bacteria) that normally reside in host tissues [1]. The major microbiota community inhabits the gastrointestinal tract, where it plays a barrier role, as well as acts as a regulator of host immunity and metabolism, that is confirmed by the strong correlations between the intestinal dysbiosis and certain inflammatory and metabolic diseases, such as obesity, diabetes mellitus, inflammatory bowel disease, etc.[2]. A large variety of bacterial metabolites are implicated in host-microbiota interactions through their signaling and regulatory capabilities. Alkylresorcinols (ARs) are highly lipophilic polyphenols synthesized by bacteria, fungi, some animals, and higher plants [3] through the activity of type III polyketide synthase (PKS) [4].
Humans receive ARs mainly from grain meal, as well as components of food preservatives, drugs, or cosmetics [5]. However, it should be considered that ARs potentially can be synthesized by many Bacteria, including those that constitute human gut microbiota although, to our knowledge, there is no strict evidence of bacterial origin of ARs in humans. According to the results of the investigation of the 'sequence-function relationship', 45% of the Bacteria phyla possess at least one type III PKS gene [4]. In the study of Funabashi M. [6] srs-like operon structures coding for type III PKS were found among many Gram-positive and Gram-negative bacteria, indicating that ARs production is a common feature of different prokaryotes. Experimentally it has been characterized seventeen bacterial type III PKSs from Actinobacteria, Proteobacteria, Firmicutes, and Cyanobacteria [4]. In bacteria, phenolic lipids serve as precursors for antibiotics, substances that confer antibiotic resistance, or regulators of antibiotic production [7], as well as UV-protective pigments [8], and alternative electron carriers [9]. However, entering the human body ARs possess many regulatory functions including anticancer, anti-inflammatory, antimicrobial, antiparasitics, antioxidant, genotoxic activities[3,10,11]. In this context, alkylresorcinols (ARs) can be considered as a connecting link between diet, gut microbiome, and host metabolism, as well as potential quorum sensing (QS) molecules.
Furthermore, according to published data, most dietary polyphenols are transformed into the colon by the intestinal microbiota before absorption [12]. This conversion is often essential for nutrients absorption and modulates the biological activity of these dietary compounds [13]. Gut bacteria can hydrolyse glycosides, glucuronides, sulphates, amides, esters, and lactones [12]. They also carry out ring cleavage, reduction, decarboxylation, demethylation, and dehydroxylation reactions [14,15]. Selma et al. perfectly reviewed the variety of species of gut microbiome, including Bifidobacterium sp., Lactobacillus sp., Clostridium sp., Eubacterium sp., etc., and observed their ability to metabolize different polyphenolic structures, converting them to low-absorbable and biologically active compounds [16]. It seems possible that ARs of complex structure ingested with food or originating in the gut might be converted to metabolites with a simpler structure, which are then absorbed in host circulation and influence its metabolism. On the other hand, phenolic compounds also have antimicrobial properties and can interact with the gut microbiota, thus modulating the microbial population of the gastrointestinal tract (GI), affecting both the health of the GI tract and the metabolism of dietary phenolic compounds. For example, in the study of Nikolaev et al., AR (in particular, 4-hexylresorcinol) in combination with antibiotics was shown to significantly decrease the number of germinating B. cereus spores in liquid medium, as well as in agar medium, in contrast to treatment with antibiotics alone [17]. This observation coincides with other studies showing the autoregulation potential of ARs [18]. As reported by Oishi et al. [19], ARs significantly increased the amount of Prevotella and reduced the amount of Enterococcus in the fecal microbiota of mice. Besides, it is known that some polyphenols from beverages, fruits, and vegetables can either promote or inhibit gut bacterial growth [20].
ARs regulatory activity may be mediated by its binding with endocannabinoid CB1 and CB2 receptors [21]. The CB1 receptors are expressed primarily in the central nervous system, neuronal tissues, and gastrointestinal (GI) tract, while the CB2 receptors are present predominantly in peripheral immune cells, such as B lymphocytes, macrophages, mast cells, natural killer cells, lymphatic organs, spleen, tonsils, and thymus [22]. Notable that almost all tissues can express endocannabinoid receptors, therefore endocannabinoid signaling system (or endocannabinoidome) is a critical homeostasis supportive system. The endocannabinoid (eCB) system of the GI tract is of particular importance in the context of the 'host-microbiota-environment' interrelationship. Extensive studying of eCB system functioning has led to the discovery of a bunch of lipophilic molecules – ligands of CB receptors with different and often antagonistic properties [21,23]. It has become clear that eCB plays a principal role in the maintenance of gut homeostasis, and its barrier function [24] and also is implicated in modulation of gut microbiota composition [25,26]. Olivetol (5-n-pentylresorcinol) is a resorcinol with a short-length alkyl chain and is one of the end-products of type III PKS that is responsible for cannabinoids synthesis in plants of Cannabis genus [27]. Olivetol and its derivatives are known to exhibit properties of antagonists (or competitive inhibitors) of CB1 and CB2 receptors [21,28,29]. In recent years, many intriguing effects of olivetol were revealed, such as anticancer activity in breast adenocarcinoma cells [30], and obesoprotective capabilities in diet-induced obesity in mice [31]. The authors claim that the effects of 5-pentylresorcinol are associated with specifically activated PGC-1α deacetylation, increased mitochondrial numbers, and accelerated fat burning through the induction of olivetol-related lipid metabolism genes [31].
In the way of studying ARs potential as signaling and regulatory molecules we hypothesized the existence of ARs synthesis in human gut by microbiota. Thus, the objectives of our study were 1) to show the possibility of synthesis of ARs by bacteria in the human gut and 2) to estimate the influence of olivetol supplementation on the ARs profile in the gut of mice in dependence on diet or genetic background to evaluate the regulatory potential of ARs and their contribution to the QS process of the microbiome.
2. Results
2.1. Estimation of AR content in germ-free mouse faeces after FMT
To investigate whether ARs have gut microbiota’s origin or come from a diet we carried out fecal microbiota transplantation (FMT) from three human adult donors to germ-free mice of Balb/c lineage. A significant increase in the amounts of individual members of the homologous series of ARs in stool samples was revealed in 14 days after FMT (Figure 1).
Taking into account, that the animals were on the same diet and consumed almost the same amount of food, and that mouse cells are not able to synthesize any ARs molecules, a significant increase in the content of C3, C12, and C15 (p≤ 0,01) in the feces indicates a high probability of synthesis of these ARs by the human gut microbiota representatives transplanted into gnotobiotic mice.
2.2. Estimation of ARs content in C57BL/6 and db/db and Ldlr (-/-) mice’s feces in dependence on diet content
The fact that ARs can be synthesized in the gut led us to investigation of regulatory properties of one of the short alkyl chain-length resorcinol – olivetol (C5), which is known to possess inhibitory effects on some enzymes [30] and receptors [21] as well as acts as a regulator of gene transcription [31]. Mice of C57BL/6 line were feeding regular chow diet (RCD) with or without C5 supplementation and high fat diet (HFD) with or without C5 supplementation, whereas LDLR (-/-) and db/db mice (that were used for modulating of atherosclerosis and obesity) were feeding RCD with or without C5 supplementation only (Figure 2).
We detected significant differences (p<0.05) in the ARs profile in stool samples from C57BL/6 mice that received RCD in dependence on C5 supplementation of C5 (Figure 3).
The amounts of C1, C2, C3, C12 and C15 in stool samples of mice nourished with C5 were significantly lower than those of mice nourished without C5, whereas the amount of C0 in contrast was notably increased after C5 supplementation.
We also have compared stool ARs profiles of C57BL/6 mice nourished by HFD and HFD with C5 and established that mice received high fat chow had lower levels of C5 and C15 and higher levels of C0 (Figure 4). Multiple tests also showed significant (p<0,05) differences between C0, C1, C12 and C15 amounts in mice on HFD (w or w/o C5) and RCD (w or w/o C5) (Figure 4).
It should be noted that only the amount of C12 increased after HFD supplementation with C5, while in other cases the amounts of ARs decreased in the HFD + C5 diet.
Analysis of db/db mice stool samples haven’t revealed any significant differences between ARs content in stool after RCD or RCD+C5 nutrition. However, LDLR (-/-) mice showed significant differences in content of C6, C12 and C15. Again, amounts of these ARs in stool decreased after using C5 supplementation (Figure 5).
3. Discussion
During the last decades, interest to phenolic lipids studied significantly increased. Polyphenols, including ARs, constitute a large group of bioactive molecules that are known to confer health benefits, as well as may have an impact on the composition and functioning of the gut microbiome [32]. Several homologues of the AR family have been shown to possess anticancerogenic, obesoprotective, anti-inflammatory, antioxidant, and some other effects [10,11,32,33]. For example, 5-n-pentylresorcinol may be involved in the induction of genes for lipid metabolism, thus accelerating fat burning and protecting against diet-induced obesity [31]. However, ARs as metabolites of bacteria may be synthesised in the human gut, thus influencing the metabolic activity of the gut microbiota, and acting as signaling molecules for the host organism as well.
For the first we have shown the possibility of ARs synthesis in human gut using FMT from human donors to germ-free mice. According to our investigation, the human microbiota can produce 5-n-propyl-, 5-n-dodecyl-, and 5-n-pentadecylresorcinol among examined ones (Figure 1). Notably, indeed these types of ARs were found to positively correlate with LPS concentrations in blood serum in humans as we have shown in previous study [34].
Considering, that C3, C12 and C15 have bacterial origin, it is especially important to understand why their amounts decreased after C5 supplementation except of C12, which amount in contrast increased after C5 supplementation in conditions of HFD.
We can provide several explanations of the observed results. At first, it can be hypothesised that olivetol, as an end-product in a pathway of AR synthesis, may serve as an allosteric inhibitor of type III PKS, thus increasing C5 concentration slows the synthesis of ARs by the microbiota. Second, high concentrations of C5 in the intestine can lead to increased solubility of other lipophilic compounds, thus improving their absorption in enterocytes. At the same time, an increase in C0 amount may be associated with ARs metabolism. Potentially, resorcinol C0 can accumulate during AR degradation under bacterial enzyme activity. Bacteria are known to have a wide spectrum of enzymes for metabolising endogenic or exogenic compounds, including polyphenols and other phenolic lipids, making them more soluble or increasing their biological activity [35,36].
Thirdly, we consider olivetol may influence gut microbiota composition and its capability to synthesize ARs molecules. Due to its antagonistic or inhibitory effects on CB1 or CB2 receptors olivetol influences cannabinoidome functioning thus inducing gut microbiota alterations. In fact, CB1 blockade led to an increase in the relative abundance of Akkermansia muciniphila and a decrease in Lanchnospiraceae and Erysipelotrichaceae in the gut, as has been shown in the study [26]. In addition, the authors observed that CB1 blocking improves diet-induced obesity by decreasing macrophage-mediated inflammatory response, which was associated with lower intestinal permeability and decreased endotoxemia [26]. In another study some important associations of specific bacterial genera (i.e., Barnesiella, Adlercreutzia, Parasutterella, Propionibacterium, Enterococcus, and Methylobacterium) with local and circulating levels of mediators eCB system (i.e., anandamide and N-docosahexaenoylethanolamine) independently of changes in body weight were found [25]. Numerous investigations showed that an increase in blood eCB levels, as well as a modulation of cannabinoid CB1 receptors, occurs in obese individuals [26,37]. In turn, changes in gut eCB system are associated with dysregulation of lipopolysaccharide (LPS) level, increased gut permeability, chronic inflammation, and dysbiosis of gut microbiota [38]. Remarcable, that previously we also demonstrated the existence of correlations between amounts of C0, C3, C5, and C15 and indexes of microbiota’s diversity in human stool samples (see Table 1) [34].
On the other hand, we have demonstrated the absence of influence of C5 supplementation on AR content in stool samples from db/db mice and the decrease of C6, C12 and C15 levels in stool samples from LDLR(-/-) mice after C5 supplementation. We use db/db mice as a model of genetically induced obesity and LDLR(-/-) mice as a model of atherosclerosis to investigate whether ARs are implicated in lipid metabolism and the development of such lesions. It is known that CB1 are implicated in the development of obesity, metabolic syndrome, and type 2 diabetes [39]. However, due to unknown manner C5 influences the levels of ARs in LDLR(-/-) mice only.
Considering, that ARs C3, C12 and C15 can be synthesised by the human gut microbiota, their serum concentrations are correlated with LPS levels and with microbiota diversity indexes in human stool samples, it can be assumed that ARs may serve as QS molecules, influencing gut microbiota composition and host metabolism.
The ability of C5 to act as CB1/CB2 ligands suggests the connection of ARs regulatory activity and the functioning of the eCB system. Importantly, the length of the alkyl chain is directly correlated with the affinity of phenolic lipids to the CB1/2 receptors. In a study conducted in 2011 it was demonstrated that receptor affinity of different tetrahydrocannabinols analogues decreases linearly with the shortening of the alkyl chain [40]. That may explain the dependence of gut microbiota’s diversity and serum LPS levels mainly from C12 and C15 concentartions, which are presumably regulate CB-dependent signaling pathways in a greater extent compared to other ARs investigated.
4. Materials and Methods
4.1. Experimental animals and study design
C57BL/6SPF mice (n=40, males) (were bred at the Nursery of Laboratory Animals in Puschino, Russia) and LDLR (-/-) (n=20, males) and db/db (n=20, males) mice (were bred at JAX-East and JAX-West Nurseries of Laboratory Animals, USA) housed under a 12-h light-dark cycle in a specific-pathogen-free (SPF) animal facility. All mice were fed with sterile food (Altromin 1324 FORTI, Germany) and water ad libitum. The alimentary obesity model was reproduced by feeding laboratory animals a high-fat diet enriched with animal-derived triglycerides and providing up to 30% of total calories (Altromin C 1090-30, Germany).
For the experiment only clinically healthy prequarantined animals during seven days were chosen. At the start of the investigation the age of the mice was 8 weeks, mean body weight was 19±2g. The administration of 5-n-pentylresorcinol (C5) (Hangzhou ROYAL Import & Export Co.,Ltd., China) was carried out through an atraumatic intragastric tube at a dosage of 2 mg/day per mouse for 90 days. The control group of animals was given a placebo according to a similar scheme. Stool samples were collected from each mouse immediately after excretion, frozen, and stored in liquid nitrogen.
All animal experiments were approved by the Ethics Committee for Animal Research, I.M. Sechenov First Moscow State Medical University, Russia (protocol number 96 from 02.09.2021).
4.2. Fecal microbiota transplantation
To explore whether the intestinal microbiota could produce AR molecules, we colonised germ-free Balb/c mice (8 to 10 weeks old) obtained from «Taconic Biosciences», New York, USA (n = 40) with the fecal microbiota of adult human donors (n = 3) (see Table 2).
Mice of both sexes were randomly selected for bacterial colonisation; each group was evenly balanced for male/female ratio. The mice were gavaged with 100 µl of fecal microbiota suspension obtained from human donors three times a day (100 µl/day). To prepare fecal microbiota samples for transplantation, a portion of frozen faeces of a donor was homogenised in a ratio of 0.1 g of stool to 1000 μl of saline solution and then filtered through a paper filter. The filtrate was collected for subsequent administration to mice. Mice in the control group were gavaged with 100 μl of 0.9% NaCl solution three times a day (100 µl/day). A period of acclimatisation of the animals was at least 4 days. The mice were housed for 2 weeks after the transplantation in sterilized ventilated cages in the SPF area of the vivarium of the Testing Laboratory Center on a 12-hour light/12-hour dark cycle with free access to food and water at a temperature of 20–23 °C and a humidity of 35–75%, with a circulation of purified air of 10–15 l/h, in groups of 5 animals per cage. Before the start of the experiment, a clinical examination and weighing of the animals were carried out. All experiments were approved by Ethics Committee for Animal Research, I.M. Sechenov First Moscow State Medical University, Russia (protocol number 96 from 02.09.2021).
The criteria for inclusion of human donors in the study were the absence of antibiotics, prebiotic and probiotic drug uptake for 3 months prior to the study, and a signed informed consent to participate in the survey. Exclusion criteria for healthy participants were severe somatic diseases, any disease of the gastrointestinal tract, any acute conditions, depression, alcoholism, smoking and pregnancy. The study was carried out in accordance with the Declaration of Helsinki and approved by the Ethics Council at the National Medical Research Centre for Oncology (protocol code № 44, date of approval 20.12.2019). For each of the individuals included in the study, a clinical examination, an anthropometric assessment, and a questionnaire survey were conducted, as well as stool samples were taken for laboratory research. Stool samples were freshly collected from each participant within 15 minutes after excretion, were immediately frozen and stored in liquid nitrogen.
4.3. Quantitative analysis of ARs
The quantitative analysis of ARs levels in stool samples was performed using gas chromatography with mass spectrometric detection (GC-MS). For quantitative determination of the compounds, a calibration was performed using an artificial matrix containing bovine serum albumin and sodium chloride. The calibration was based on the response of the internal standard (4-(benzyloxy)-phenol) introduced into the test samples in order to normalize the derivatization processes and smooth out the errors in determining the concentrations of the test substances. The study was carried out using an Agilent 6890 gas chromatograph (Agilent Inc., United States) with an automatic sample introduction system coupled to an Agilent 5850 mass spectrometric detector (Agilent Inc., United States) with electron impact ionisation. Chromatography was performed using the Restek Rtx 5 Sil-MS column (Restek Corporation, Bellefonte, USA) with the following column parameters: length - 30 m, cross-sectional diameter - 250 µm, particle size - 0.25 µm. Chromatography parameters were as follows: sample injection with a flow division of 5:1, carrier gas - helium, gas supply mode - constant flow, flow rate - 1 ml/min, inlet temperature - 290°C. Thermostat was set at the initial temperature of 80°C, holding time - 1 min, then - rising temperature to 320°C at a rate of 25°C/min, holding time - 4 min.
The stool samples were lyophilised, then the internal standard (4- (benzyloxy)-phenol) and 300 μL of diethyl ether were added to approximately 5 mg of the sample, the mixture was stirred on a shaker and then centrifuged at 2000 rpm. Liquid-liquid extraction was repeated twice. The organic phase was transferred to disposable tubes and evaporated in a stream of nitrogen, following by derivatization carried out with a silylating agent, methyl-silyl-trifluoroacetamide (MSTFA), for 30 min at 60°C. After the derivatization procedure, the samples were analysed by GC-MS. The technique was validated in terms of selectivity, linearity, accuracy, reproducibility, matrix effect, and analyte stability. Validation was carried out in accordance with the FDA Bioanalytical Method Validation Guidelines.
Feces samples were collected in ethanol and stored in Biobank at -80oC until testing.
4.4. Statistical data analysis
Statistical processing of the data was carried out using the method of nonparametric statistics using the Prism 9 statistical software package. The statistical significance of the differences in the mean values of independent samples was assessed using nonparametric analysis after checking the distribution of data for normality. Correlation analysis according to Spearman was carried out with an assessment of the statistical significance of the correlation coefficient.
5. Conclusions
For the first time, we have shown that such ARs as 5-n-propylresorcinol, 5-n-dodecylresorcinol and 5-n-pentadecylresorcinol can be produced by the intestinal microbiota in human gut. We have also investigated the influence of 5-n-pentylresorcinol dietary supplementation on ARs profiles in C57BL/6, LDLR (-/-), and db/db mice. According to our study olivetol supplementation led to changing in ARs content in stool samples of C57BL/6 and LDLR (-/-) mice, but not db/db mice. We hypothesise that olivetol's effects can be mediated through its interactions with intestinal endocannabinoid signaling system. Taking into account, that AR concentrations in human serum are correlated with LPS levels and with microbiota diversity indexes in human stool samples, it can be assumed that ARs may serve as quorum-sensing molecules, influencing gut microbiota composition and host metabolism as well.
Author Contributions
Conceptualization, A.A.Z and A.V.S; methodology, A.M.G., E.Yu.Z. S.A.A.; validation, P.A.M.; formal analysis, A.A.Z; investigation, A.M.G.; resources, A.Yu.M., A.S.G., I.A.N.; data curation, S.A.R.; writing—original draft preparation, A.A.Z.; writing—review and editing, A.V.S.; visualization, A.A.Z.; supervision, A.V.S.; project administration, A.V.S.; funding acquisition, A.V.S. and S.A.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Institutional Review Board (or Ethics Committee) the Ethics Council at the National Medical Research Centre for Oncology (protocol code № 44, date of approval 20.12.2019). The animal study protocol was approved by the Institutional Review Board (or Ethics Committee) of the Ethics Committee for Animal Research, I.M. Sechenov First Moscow State Medical University, Russia (protocol number 96 from 02.09.2021).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gomaa, E.Z. Human Gut Microbiota/Microbiome in Health and Diseases: A Review. Antonie Van Leeuwenhoek 2020, 113, 2019–2040. [Google Scholar] [CrossRef] [PubMed]
- Saad, M.J.A.; Santos, A.; Prada, P.O. Linking Gut Microbiota and Inflammation to Obesity and Insulin Resistance. Physiology 2016, 31, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Zabolotneva, A.A.; Shatova, O.P.; Sadova, A.A.; Shestopalov, A.V.; Roumiantsev, S.A. An Overview of Alkylresorcinols Biological Properties and Effects. J Nutr Metab 2022, 2022, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, Y.; Ogata, H.; Goto, S. Type III Polyketide Synthases: Functional Classification and Phylogenomics. ChemBioChem 2017, 18, 50–65. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Kim, D.W.; Kim, S.-G.; Lee, S.K. 4-Hexylresorcinol-Induced Protein Expression Changes in Human Umbilical Cord Vein Endothelial Cells as Determined by Immunoprecipitation High-Performance Liquid Chromatography. PLoS ONE 2020, 15, e0243975. [Google Scholar] [CrossRef]
- Funabashi, M.; Funa, N.; Horinouchi, S. Phenolic Lipids Synthesized by Type III Polyketide Synthase Confer Penicillin Resistance on Streptomyces Griseus. Journal of Biological Chemistry 2008, 283, 13983–13991. [Google Scholar] [CrossRef]
- Ridley, C.P.; Lee, H.Y.; Khosla, C. Evolution of Polyketide Synthases in Bacteria. Proceedings of the National Academy of Sciences 2008, 105, 4595–4600. [Google Scholar] [CrossRef]
- Austin, M.B.; Izumikawa, M.; Bowman, M.E.; Udwary, D.W.; Ferrer, J.-L.; Moore, B.S.; Noel, J.P. Crystal Structure of a Bacterial Type III Polyketide Synthase and Enzymatic Control of Reactive Polyketide Intermediates. Journal of Biological Chemistry 2004, 279, 45162–45174. [Google Scholar] [CrossRef]
- Anand, A.; Verma, P.; Singh, A.K.; Kaushik, S.; Pandey, R.; Shi, C.; Kaur, H.; Chawla, M.; Elechalawar, C.K.; Kumar, D.; et al. Polyketide Quinones Are Alternate Intermediate Electron Carriers during Mycobacterial Respiration in Oxygen-Deficient Niches. Mol Cell 2015, 60, 637–650. [Google Scholar] [CrossRef]
- Stasiuk, M.; Kozubek, A. Biological Activity of Phenolic Lipids. Cellular and Molecular Life Sciences 2010, 67, 841–860. [Google Scholar] [CrossRef]
- Kozubek, A.; Tyman, J.H.P. Resorcinolic Lipids, the Natural Non-Isoprenoid Phenolic Amphiphiles and Their Biological Activity. Chem Rev 1999, 99, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Aura, A.-M. Microbial Metabolism of Dietary Phenolic Compounds in the Colon. Phytochemistry Reviews 2008, 7, 407–429. [Google Scholar] [CrossRef]
- Duda-Chodak, A.; Tarko, T.; Satora, P.; Sroka, P. Interaction of Dietary Compounds, Especially Polyphenols, with the Intestinal Microbiota: A Review. Eur J Nutr 2015, 54. [Google Scholar] [CrossRef] [PubMed]
- LAMPE, J.; CHANG, J. Interindividual Differences in Phytochemical Metabolism and Disposition. Semin Cancer Biol 2007, 17, 347–353. [Google Scholar] [CrossRef]
- Rechner, A. Colonic Metabolism of Dietary Polyphenols: Influence of Structure on Microbial Fermentation Products. Free Radic Biol Med 2004, 36, 212–225. [Google Scholar] [CrossRef]
- Selma, M. v.; Espín, J.C.; Tomás-Barberán, F.A. Interaction between Phenolics and Gut Microbiota: Role in Human Health. J Agric Food Chem 2009, 57. [Google Scholar] [CrossRef]
- Nikolaev, Y.A.; Tutel’yan, A. v.; Loiko, N.G.; Buck, J.; Sidorenko, S. v.; Lazareva, I.; Gostev, V.; Manzen’yuk, O.Y.; Shemyakin, I.G.; Abramovich, R.A.; et al. The Use of 4-Hexylresorcinol as Antibiotic Adjuvant. PLoS ONE 2020, 15. [Google Scholar] [CrossRef]
- Bukharin, O. v; Perunova, N.B.; El’-Registan, G.I.; Nikolaev, I.A.; Iavnova, S. v; Molostov, E. v; Kirillov, D.A. [Influence of Chemical Analogue of Extracellular Microbial Autoregulators on Antilysozyme Activity of Bacteria]. Zh Mikrobiol Epidemiol Immunobiol.
- Oishi, K.; Yamamoto, S.; Itoh, N.; Nakao, R.; Yasumoto, Y.; Tanaka, K.; Kikuchi, Y.; Fukudome, S.; Okita, K.; Takano-Ishikawa, Y. Wheat Alkylresorcinols Suppress High-Fat, High-Sucrose Diet-Induced Obesity and Glucose Intolerance by Increasing Insulin Sensitivity and Cholesterol Excretion in Male Mice. J Nutr 2015, 145. [Google Scholar] [CrossRef]
- Etxeberria, U.; Fernández-Quintela, A.; Milagro, F.I.; Aguirre, L.; Martínez, J.A.; Portillo, M.P. Impact of Polyphenols and Polyphenol-Rich Dietary Sources on Gut Microbiota Composition. J Agric Food Chem 2013, 61. [Google Scholar] [CrossRef]
- Cascio, M.G.; Bisogno, T.; Palazzo, E.; Thomas, A.; van der Stelt, M.; Brizzi, A.; de Novellis, V.; Marabese, I.; Ross, R.; van de Doelen, T.; et al. In Vitro and in Vivo Pharmacology of Synthetic Olivetol- or Resorcinol-Derived Cannabinoid Receptor Ligands. Br J Pharmacol 2006, 149, 431–440. [Google Scholar] [CrossRef]
- Piomelli, D. The Molecular Logic of Endocannabinoid Signalling. Nat Rev Neurosci 2003, 4, 873–884. [Google Scholar] [CrossRef] [PubMed]
- Nevalainen, T. Recent Development of CB2 Selective and Peripheral CB1/CB2 Cannabinoid Receptor Ligands. Curr Med Chem 2013, 21, 187–203. [Google Scholar] [CrossRef] [PubMed]
- Cuddihey, H.; MacNaughton, W.K.; Sharkey, K.A. Role of the Endocannabinoid System in the Regulation of Intestinal Homeostasis. Cell Mol Gastroenterol Hepatol 2022, 14, 947–963. [Google Scholar] [CrossRef]
- Lacroix, S.; Pechereau, F.; Leblanc, N.; Boubertakh, B.; Houde, A.; Martin, C.; Flamand, N.; Silvestri, C.; Raymond, F.; Di Marzo, V.; et al. Rapid and Concomitant Gut Microbiota and Endocannabinoidome Response to Diet-Induced Obesity in Mice. mSystems 2019, 4. [Google Scholar] [CrossRef]
- Mehrpouya-Bahrami, P.; Chitrala, K.N.; Ganewatta, M.S.; Tang, C.; Murphy, E.A.; Enos, R.T.; Velazquez, K.T.; McCellan, J.; Nagarkatti, M.; Nagarkatti, P. Blockade of CB1 Cannabinoid Receptor Alters Gut Microbiota and Attenuates Inflammation and Diet-Induced Obesity. Sci Rep 2017, 7, 15645. [Google Scholar] [CrossRef] [PubMed]
- Tahir, M.N.; Shahbazi, F.; Rondeau-Gagné, S.; Trant, J.F. The Biosynthesis of the Cannabinoids. J Cannabis Res 2021, 3, 7. [Google Scholar] [CrossRef] [PubMed]
- Zagzoog, A.; Cabecinha, A.; Abramovici, H.; Laprairie, R.B. Modulation of Type 1 Cannabinoid Receptor Activity by Cannabinoid By-Products from Cannabis Sativa and Non-Cannabis Phytomolecules. Front Pharmacol 2022, 13. [Google Scholar] [CrossRef]
- James, J. Carberry Composition of Olivetol and Method of Use to Reduce or Inhibit the Effects of Tetrahydrocannabinol in the Human Body 2017.
- Yang, X.; Zhao, Z.; Zhao, C.; Li, Y.; El-kott, A.F.; Bani-Fwaz, M.Z. Anti-Breast Adenocarcinoma and Anti-Urease Anti-Tyrosinase Properties of 5-Pentylresorcinol as Natural Compound with Molecular Docking Studies. J Oleo Sci 2022, 71, ess22024. [Google Scholar] [CrossRef]
- Imai, S.; Ohama, M.; Suzuki, M.; Katayanagi, Y.; Kayashima, Y.; Tezuka, H. Research Article Short Alkyl Chain-Length Resorcinol Olivetol Protects Against Obesity with Mitochondrial Activation by Pgc-1α Deacetylation. SSRN Electronic Journal 2021. [Google Scholar] [CrossRef]
- Fraga, C.G.; Croft, K.D.; Kennedy, D.O.; Tomás-Barberán, F.A. The Effects of Polyphenols and Other Bioactives on Human Health. Food Funct 2019, 10, 514–528. [Google Scholar] [CrossRef] [PubMed]
- Rejman, J.; Kozubek, A. Long-Chain Orcinol Homologs from Cereal Bran Are Effective Inhibitors of Glycerophosphate Dehydrogenase. Cellular and Molecular Biology Letters 1997, 2, 411–419. [Google Scholar]
- Shestopalov, A.V.; Gaponov, A.M.; Zabolotneva, A.A.; Appolonova, S.A.; Markin, P.A.; Borisenko, O.V.; Tutelyan, A.V.; Rumyantsev, A.G.; Teplyakova, E.D.; Shin, V.F.; et al. Alkylresorcinols: New Potential Bioregulators in the Superorganism System (Human–Microbiota). Biology Bulletin 2022, 49, 150–159. [Google Scholar] [CrossRef]
- Corrêa, T.A.F.; Rogero, M.M.; Hassimotto, N.M.A.; Lajolo, F.M. The Two-Way Polyphenols-Microbiota Interactions and Their Effects on Obesity and Related Metabolic Diseases. Front Nutr 2019, 6. [Google Scholar] [CrossRef] [PubMed]
- Makarewicz, M.; Drożdż, I.; Tarko, T.; Duda-Chodak, A. The Interactions between Polyphenols and Microorganisms, Especially Gut Microbiota. Antioxidants 2021, 10, 188. [Google Scholar] [CrossRef]
- Engeli, S.; Böhnke, J.; Feldpausch, M.; Gorzelniak, K.; Janke, J.; Bátkai, S.; Pacher, P.; Harvey-White, J.; Luft, F.C.; Sharma, A.M.; et al. Activation of the Peripheral Endocannabinoid System in Human Obesity. Diabetes 2005, 54, 2838–2843. [Google Scholar] [CrossRef]
- Muccioli, G.G.; Naslain, D.; Bäckhed, F.; Reigstad, C.S.; Lambert, D.M.; Delzenne, N.M.; Cani, P.D. The Endocannabinoid System Links Gut Microbiota to Adipogenesis. Mol Syst Biol 2010, 6, 392. [Google Scholar] [CrossRef]
- Lu, D.; Dopart, R.; Kendall, D.A. Controlled Downregulation of the Cannabinoid CB1 Receptor Provides a Promising Approach for the Treatment of Obesity and Obesity-Derived Type 2 Diabetes. Cell Stress Chaperones 2016, 21, 1–7. [Google Scholar] [CrossRef]
- Andersson, D.A.; Gentry, C.; Alenmyr, L.; Killander, D.; Lewis, S.E.; Andersson, A.; Bucher, B.; Galzi, J.-L.; Sterner, O.; Bevan, S.; et al. TRPA1 Mediates Spinal Antinociception Induced by Acetaminophen and the Cannabinoid Δ9-Tetrahydrocannabiorcol. Nat Commun 2011, 2, 551. [Google Scholar] [CrossRef]
Figure 1.
Amounts (nmol/g) of (a) propylresorcinol (C3), (b) dodecylresorcinol (C12) and (c) pentadecylresorcinol (C15) in stool samples from mice after the fecal microbiota transplantation from human donors (Donor 1 - Donor 3) compared to the control group (Control) (p value ≤ 0,05).
Figure 1.
Amounts (nmol/g) of (a) propylresorcinol (C3), (b) dodecylresorcinol (C12) and (c) pentadecylresorcinol (C15) in stool samples from mice after the fecal microbiota transplantation from human donors (Donor 1 - Donor 3) compared to the control group (Control) (p value ≤ 0,05).
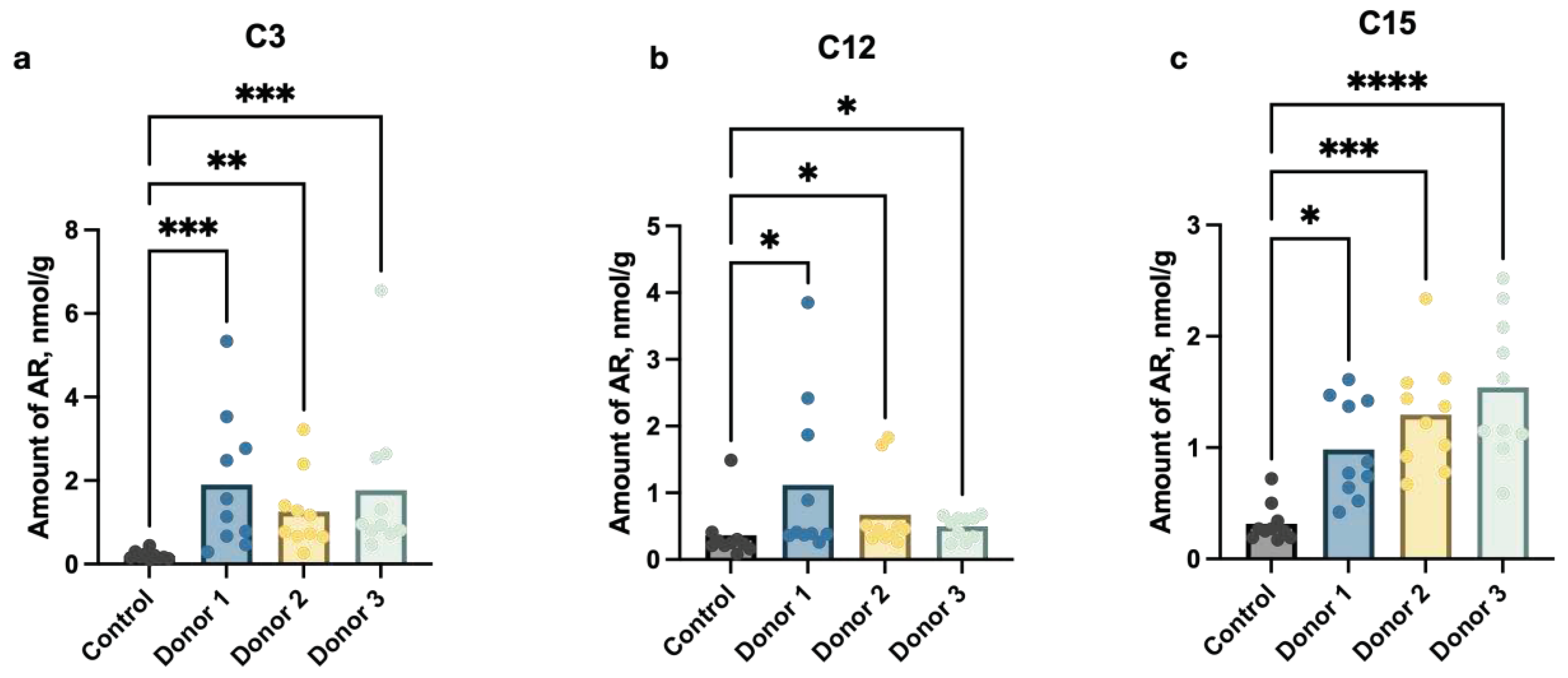
Figure 2.
Mice of three lines (C57BL/6, LDLR (-/-), db/db) were feeding by regular chow diet (RCD) or high fat diet (HFD) with or without olivetol (C5) supplementation. Stool samples from all mice were collected for the following GC-MS analysis of the ARs content.
Figure 2.
Mice of three lines (C57BL/6, LDLR (-/-), db/db) were feeding by regular chow diet (RCD) or high fat diet (HFD) with or without olivetol (C5) supplementation. Stool samples from all mice were collected for the following GC-MS analysis of the ARs content.
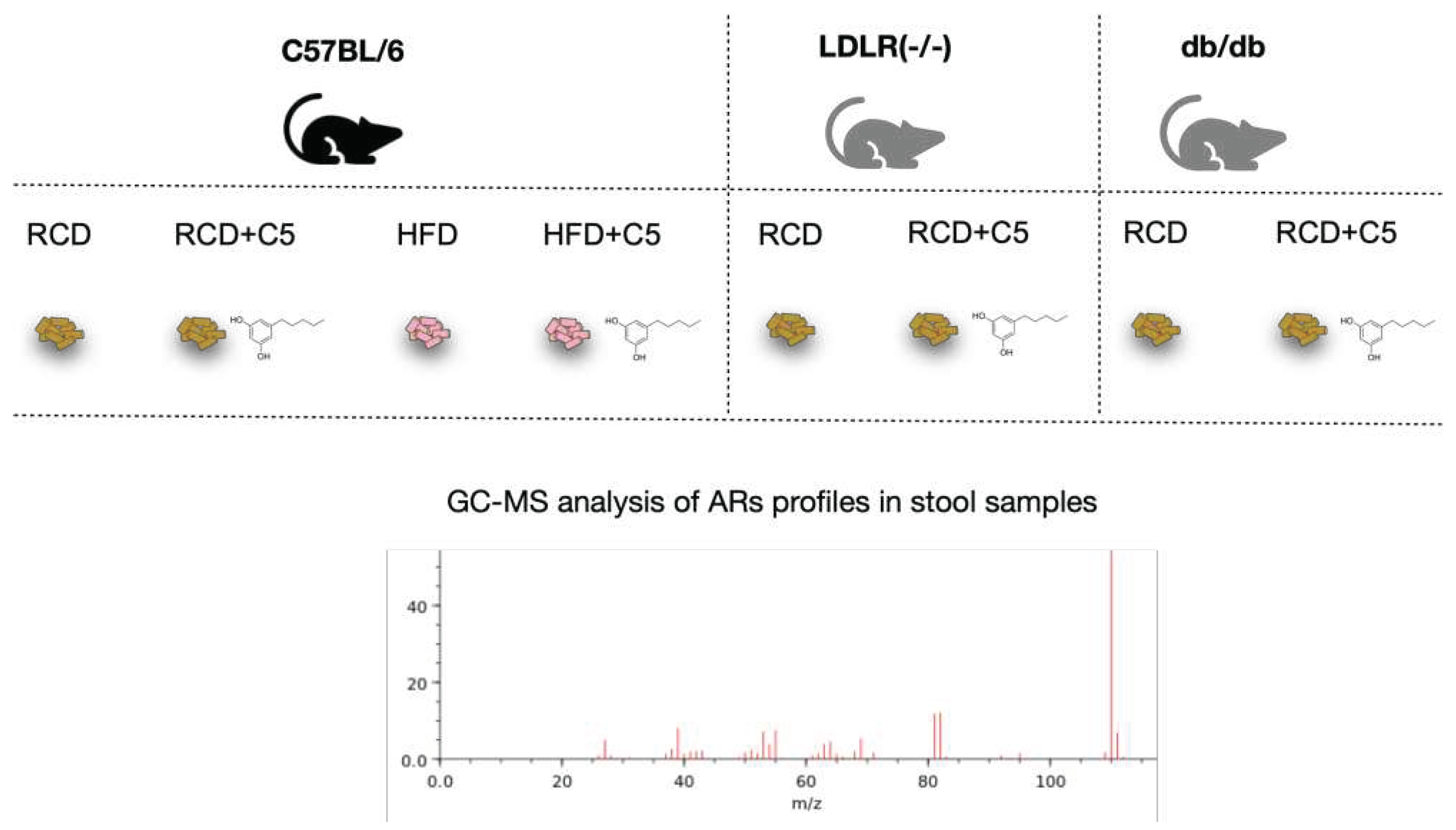
Figure 3.
Differences in ARs content in stool samples of C57BL/6 mice after feeding with regular diet without or with C5 (sign “+”) supplementation. C0 – resorcinol, C1 – methylresorcinol, C2 – ethylresorcinol, C3 – propylresorcinol, C12 – dodecylresorcinol, C15 – pentadecylresorcinol.
Figure 3.
Differences in ARs content in stool samples of C57BL/6 mice after feeding with regular diet without or with C5 (sign “+”) supplementation. C0 – resorcinol, C1 – methylresorcinol, C2 – ethylresorcinol, C3 – propylresorcinol, C12 – dodecylresorcinol, C15 – pentadecylresorcinol.
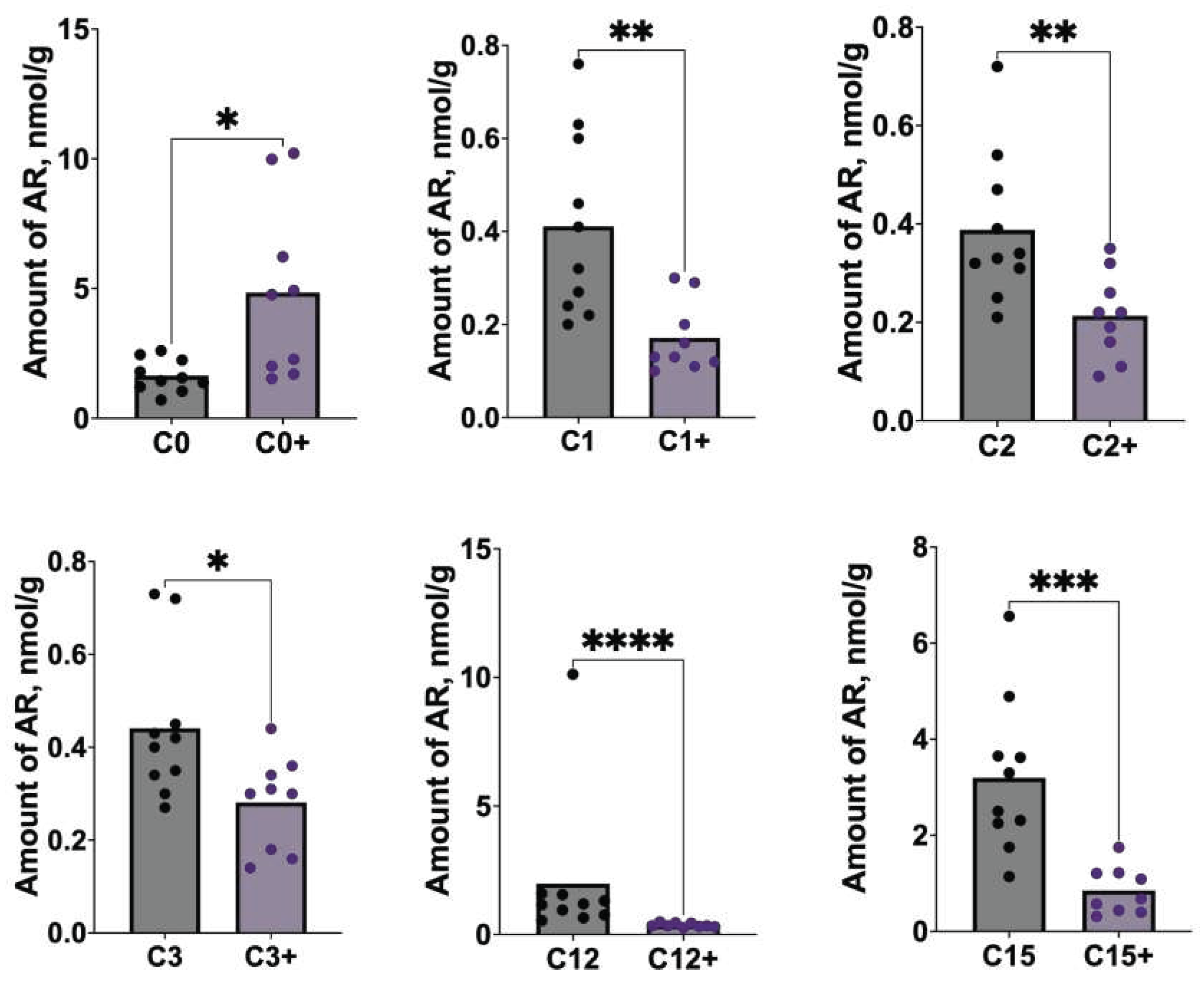
Figure 4.
Differences in ARs content (nmol/g) in stool samples of C57BL/6 mice after feeding with regular (RCD) or hig fat (FHD) diet without or with C5 (sign “+”) supplementation. (a) Comparison of stool samples after HFD and RCD w/o C5 supplementation; (b) multiple comparison of stool samples after HFD and RCD w and w/o C5 supplementation. C0 – resorcinol, C1 – 5-methylresorcinol, C5 – 5-n-pentylresorcinol, C12 – 5-n-dodecylresorcinol, C15 – 5-n-pentadecylresorcinol.
Figure 4.
Differences in ARs content (nmol/g) in stool samples of C57BL/6 mice after feeding with regular (RCD) or hig fat (FHD) diet without or with C5 (sign “+”) supplementation. (a) Comparison of stool samples after HFD and RCD w/o C5 supplementation; (b) multiple comparison of stool samples after HFD and RCD w and w/o C5 supplementation. C0 – resorcinol, C1 – 5-methylresorcinol, C5 – 5-n-pentylresorcinol, C12 – 5-n-dodecylresorcinol, C15 – 5-n-pentadecylresorcinol.
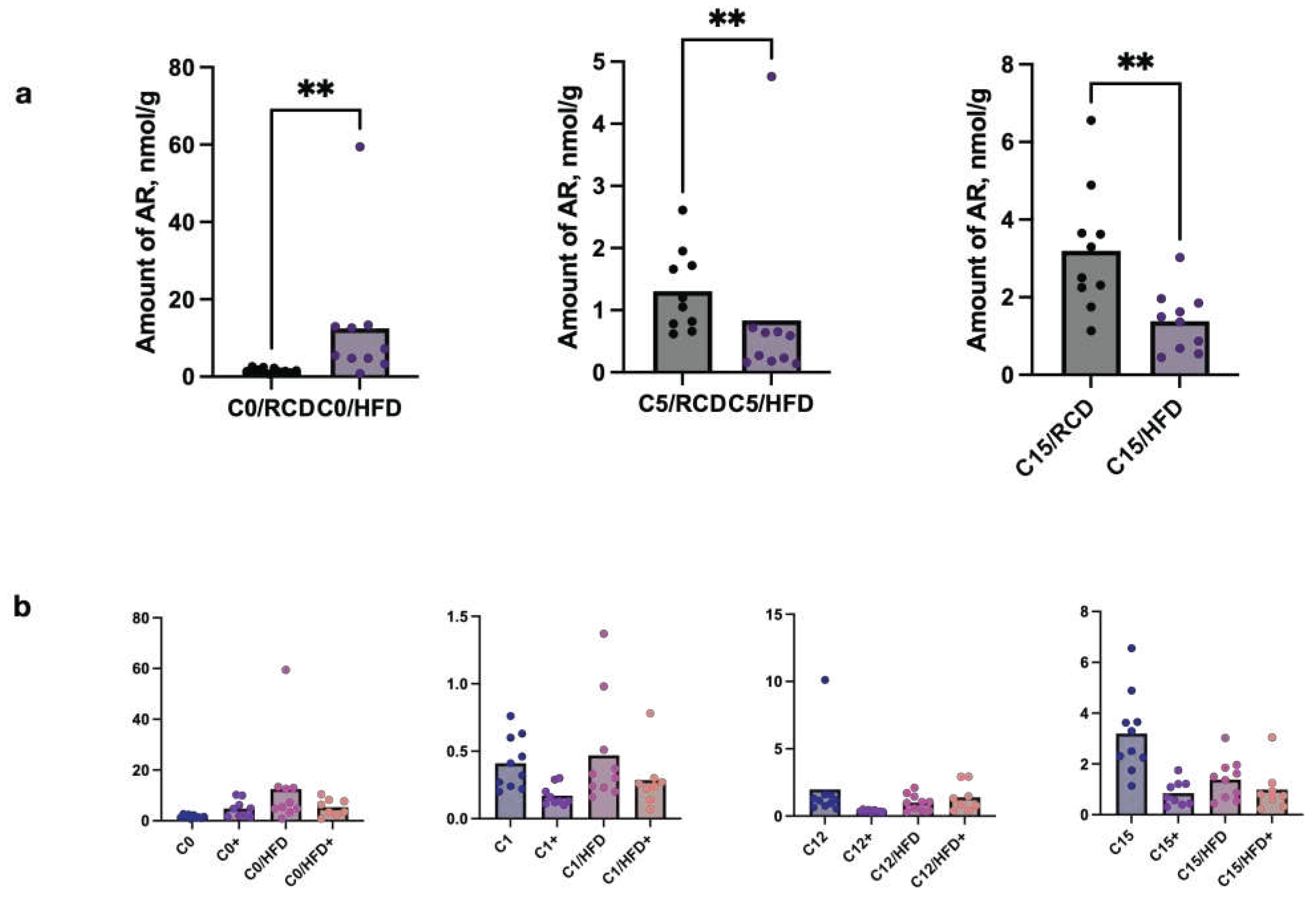
Figure 5.
Differences in ARs content in stool samples of LDLR (-/-) mice after feeding with RCD without or with C5 (sign '+') supplementation. C6 – hexylresorcinol, C12 – dodecylresorcinol, C15 – pentadecylresorcinol.
Figure 5.
Differences in ARs content in stool samples of LDLR (-/-) mice after feeding with RCD without or with C5 (sign '+') supplementation. C6 – hexylresorcinol, C12 – dodecylresorcinol, C15 – pentadecylresorcinol.
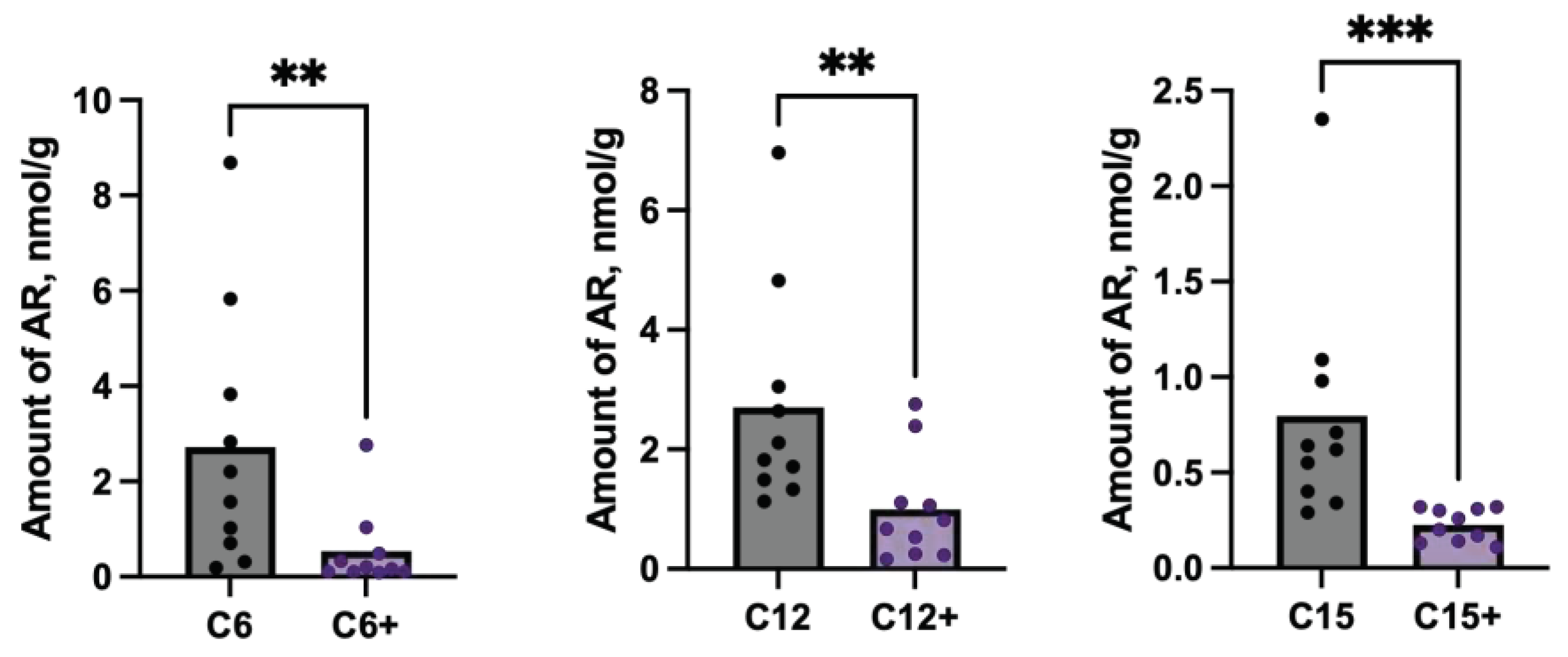

Table 1.
Spearmen correlation coefficients estimated between amounts (nmol/g) of ARs in stool samples from adults and children and indexes of the diversity of the microbiota according to the study [34].
Table 1.
Spearmen correlation coefficients estimated between amounts (nmol/g) of ARs in stool samples from adults and children and indexes of the diversity of the microbiota according to the study [34].
| Index | AR | p_value | Spearmen correlation |
|---|---|---|---|
| Stool samples of adults | |||
| Berger-Parker Dominance | C15 | 0.015 | 0.26 |
| Inverse Simpson Index | C15 | 0.045 | 0.21 |
| Gini–Simpson Index | C15 | 0.045 | 0.21 |
| Distinct OTUs | C0 | 0.019 | 0.26 |
| Chao1 Richness | C0 | 0.021 | 0.25 |
| Distinct OTUs | C3 | 0.030 | 0.24 |
| Chao1 Richness | C3 | 0.042 | 0.22 |
| Stool samples of children | |||
| Inverse Simpson Index | C5 | 0.033 | 0.26 |
| Gini–Simpson Index | C5 | 0.033 | 0.26 |
Table 2.
Characteristics of groups of germ-free mice and donors of stool samples for fecal microbiota transplantation. N/A - not applicable, yo - years old.
Table 2.
Characteristics of groups of germ-free mice and donors of stool samples for fecal microbiota transplantation. N/A - not applicable, yo - years old.
| Group of animals | Number of animals | Material for transplantation | Route of administration | Microbiota donor |
|---|---|---|---|---|
| 1 (control) | 10 | 0,9% NaCl solution | Intragastric | N/A |
| 2 | 10 | Fecal microbiota sample from Donor 1 | Intragastric | Donor 1. Male, 42 yo |
| 3 | 10 | Fecal microbiota sample from Donor 2 | Intragastric | Donor 2. Male, 36 yo |
| 4 | 10 | Fecal microbiota sample from Donor 3 | Intragastric | Donor 3. Female, 28 yo |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



