
Submitted:
05 July 2023
Posted:
06 July 2023
You are already at the latest version
Abstract
The Shisa family is a type of single-transmembrane adaptor proteins containing an N-terminal cysteine-rich domain and a proline-rich C-terminal region. Nine Shisa subfamily genes have been proposed in most vertebrates; however, some of them might be species-specific. The number of Shisa genes present in zebrafish remains unclear. The purpose of this study was to investigate the evolutionary relationships among Shisa family genes in zebrafish (TU strain) using phylogenetic and syntenic analyses. The function of shisa-2 was preliminarily examined via CRISPR/Cas13d-mediated knockdown. Following identification in zebrafish,10 Shisa family genes, namely shisa-1, 2, 3, 4, 5, 6, 7, 8, 9a, and 9b, were classified into three main clades and six subclades. Their encoding proteins contained a cysteine-rich N-terminal domain and a proline-rich C-terminal region containing different motifs. A specific syntenic block containing atp8a2 and shisa-2 was observed to be conserved across all species. Furthermore, all these genes were found to be expressed during embryogenesis. shisa-2 was expressed in the presomitic mesoderm, somites, and so on. shisa-2 was identified as a regulator of the expression of the somite formation marker mesp-ab. Overall, our study provides new insights into the evolution of Shisa family genes and the control of shisa-2 over the convergent extension cells of somatic precursors in zebrafish.
Keywords:
gene expression
; knockdown
; CRISPR/Cas13d
; shisa -2
; mesp ab
; somite formation
1. Introduction
To maintain their development and growth, cells need to coordinate and integrate multiple functional modules, including controlling cytoskeleton remodeling, adapting to environmental alterations, and gene expression [1]. Functional modules are regulated by a series of signaling networks that communicate with each other via the interaction of their components with adaptor, docking, anchoring, or scaffold proteins [1,2,3]. Scaffold proteins contain several modular domains, including the SH3, WW, SH2, and PDZ domains, which mediate protein–protein interactions and/or protein–membrane associations [1,3].
The Shisa family proteins are recently discovered adaptor proteins [4]. The first gene of the Shisa family was discovered in 2005 and was named shisa because of its effect on the formation of African clawed frog (Xenopus laevis) heads [5]. As adaptor proteins, Shisa family proteins play various roles in different species and developmental processes, including cancer development and apoptosis [4]. Shisa-1 inhibits Wnt receptor Frizzled glycosylation and fibroblast growth factor receptor (FGFR) phosphorylation during X.laevis head formation [5]. During somitogenesis, X.laevis Shisa- 2 regulates segmental patterning by inhibiting Wnt receptor Frizzled glycosylation and FGFR phosphorylation [6]. During the growth and development of mouse commissural axons, Shisa- 2 only inhibits the glycosylation of Frizzled 3 [7]. In C2C12 cells, Shisa-2 regulates the rearrangement of the muscle cytoskeletal actin F-actin and subsequently promotes myoblast fusion [8]. Shisa- 3 serves as a tumor suppressor by accelerating catenin degradation [9]. Shisa- 4–6 and Shisa- 8 can degrade proteins by regulating the ubiquitination of target proteins [4]. Shisa 5 can degrade the nonstructural protein 5 (NS5A) of the hepatitis C virus (HCV) [10]. Its binding to Cyclin B results in DNA damage and degradation[11]. Shisa- 7 can transport γ-aminobutyric acid type A receptors (GABAA receptors) during brain neurodevelopment in mice [12]. Shisa- 6–9 are critical subunits of the α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid receptor during its biogenesis and function in the central nervous system [13]. Therefore, different Shisa proteins play different roles in various biological processes, even though the same protein plays different roles in various tissues or cells with different characteristics [14] and may have tissue- and cell-specific regulatory pathways.
All proteins in this family contain a predicted signal peptide, an N-terminal cysteine-rich domain (Cys), a transmembrane domain (TM), and a C-terminal proline-rich region. In the C-terminal proline region, other motifs and/or domains, such as the PY ([LP] PxY) motif, the PDZ (postsynaptic density protein) domain binding motif ([ST]x[VLI]), and the GRID domain (GABAA receptor binding domain), are present [4,12]. Different Shisa proteins play different roles in various species and developmental processes based on their domains and/or motifs [4]. In mice, Shisa- 7 binds with GABAA receptors through the GRID domain to control benzodiazepine actions [12]. The proline region of Shisa- 5 (Scotin) maintains its endoplasmic reticulum (ER) specific expression [15]. Shisa-5 induces apoptosis through the interaction of its cysteine domain with Cyclin B [11]. The interaction of the TM and proline region of Shisa-5 with HCV NS5A is required to control NS5A degradation [16].
To date, nine Shisa subfamily genes have been identified [4]. Most vertebrate genomes contain at least eight Shisa genes (Shisa- 2–9). However, shisa-1 is found only in zebrafish (Danio rerio) and X. laevis [17]. All of the subfamilies were predicted in zebrafish using the genome database and RNA sequencing; however, and shisa-8 was not found [4]. The zebrafish, a small freshwater teleost, is a widely used animal model in developmental biology, ecotoxicology, cancer, and neuroscience research. As a model organism, zebrafish have several advantages such as a short generation time, small size, high fecundity, transparent embryonic development, and high genetic homology to humans. However, the number of Shisa family genes present in zebrafish is still unknow. The aim of the present study was to examine the presence of Shisa family genes and their expression patterns in zebrafish and determine the potential functions of one of them during embryonic development. The findings of the presented study could provide a basis for the understanding the evolution of the Shisa family genes and the function of related genes.
2. Results
2.1. Nine Shisa Subfamilies Identified in Zebrafish
In the zebrafish genomic and transcriptomic databases ,eleven predicted Shisa candidates were found and named shisa 1bl, 2, 2a, 3, 4, 5, 6, 7l, 8b, 9a, and 9b(Table 1).All of them were cloned and sequenced to verify the original sequences and names. After sequencing, all the encoded amino acids were used for phylogenetic analysis (Figure 1). As a result, 10 of them were classified into nine subfamilies that were consistent with the previous subfamily classifications, including Shisa-1, Shisa-2, Shisa-3, Shisa-4, Shisa-5, Shisa-6, Shisa-7, Shisa-8, and Shisa-9. The predicted Shisa 2a, Shisa 7l, and Shisa-8b were determined to be Shisa-1, Shisa-7, and Shisha-8, respectively (Table 1). The nine subfamilies were divided into three clades and six subclades (Figure 1). Shisa-1 and Shisa-2, Shisa-6 and Shisa-7, and Shisa-8 and Shisa- 9 belonged to separate subclades.
2.2. Conserved Domain Among Shisa Proteins
The Shisa motif, TM, and low-complex proline-rich regions were predicted in all Shisa proteins (Figure 2). Signal peptides were present in zebrafish Shisa proteins except Shisa-9b. The proline-rich domain was only found in Shisa-4. The amino acid sequences were further aligned and analyzed (Figure 3). Shisa family genes shared a low average amino acid identity of 17.48%. All of them contained a cysteine-rich domain in the N-terminus with the following distinct pattern: C*C*CC*C*CC*C (“*” represents a series of amino acid residues). Several cysteine residues were present near the C-termini of the predicted TMs in zebrafish Shisa-1–5, which were not found in Shisa-6–9 (Figure 3). Compared with other Shisa subfamilies, several sequence features present in Shisa- 1–3; for example , long insertions with a “PEXXDXXDA” signature were observed between the second and third conserved cysteines in the cysteines-rich domain, and a conserved sequence “PxxxP” was found at the beginning of their predicted TMs (Figure 3) [4].
A typical conserved domain ( "C * C * CC * C * CC * C", shaded yellow) of Shisa protein and TM domain was observed. The number in the sequence represents the amino acid number between two amino-acids. The predicted transmembrane region is framed with a blue border (predicted by DNAMAN). Prolines and glycines are shown in red letters. The noncharged residues at the main hydrophobic residue positions are colored blue. The predicted cysteine residues within and after the transmembrane segment are marked in gray. The PY motif ([LP]PxY) are marked in green[4]. The C-terminal PDZ-binding motifs([TSVYF]x[VIL])are marked in pink [4].PY like motif, ‘ ’; C-terminal PDZ-binding like motif, ‘
’; C-terminal PDZ-binding like motif, ‘ ’ .The GRID domain [12] is shown in green letters.
’ .The GRID domain [12] is shown in green letters.
’; C-terminal PDZ-binding like motif, ‘’ .The GRID domain [12] is shown in green letters.2.3. Shisa Genes Localization on Chromosomes
The chromosomal locations of the Zebrafish Shisa family genes are shown in Figure 4. Most genes were located on chromosomes 3, 12, 14, 22, 24, and 25; however, shisa-1 (previously predicted to be shisa 2a), shisa-4, shisa-7, and shisa-8b were not located on those chromosomes based on the present genomics data.
2.4. Conserved Syntenic Block Containing shisa-2
Collinearity analysis was performed by comparing shisa-2 chromosome distribution among Homo sapiens, Mus musculus, Gallus gallus, X. tropicalis, and Danio rerio. We found that a specific syntenic block containing atp8a2 (ATPase phospholipid transporting 8A2) and shisa-2, arranged in reverse order, was conserved across all species (Figure 5).
2.5. Different Expression Patterns of Shisa Family Genes During Embryonic Development
The temporal expression profiles of Shisa family genes in zebrafish during embryonic development were determined using semi-quantitative PCR (Figure 6). The results indicated that most shisa genes were expressed during zebrafish embryogenesis and their expression was upregulated during embryonic development. Most of them were expressed at 0.2hours post fertilization (hpf), except shisa-9b.The expression of shisa-3 was weak at 8 hpf, then increased. After 16 hpf, the expression decreased. Shisa-9b was expressed from 12 hpf. The transcripts of shisa-1, shisa-2, shisa-4, shisa-5, shisa-6,shisa-7, shisa-8 and shisa-9a were high or weak in all test stages.
The spatiotemporal expression map of shisa-2 was analyzed using in situ hybridization in zebrafish. At 11 hpf, shisa-2 was expressed in the presomitic mesoderm, both in the medial and lateral regions of the somites and at the base of the eye vesicle (Figure 7A and B). At 16 hpf, shisa-2 expression was strong in the somites, optic vesicles, gill arches, and head regions (Figure 7C). At 20 hpf, shisa-2 expression in the differentiated somites gradually decreased but remained strong in the newly formed somites and presomitic cells in the tail region and optic vesicles (Figure 7D and E).
2.6. Abnormal Expression of mesp-ab by shisa-2 Knockdown
The phenotype of embryos and larvae injected with Cas13d mRNA and gRNA was assessed at 48 hpf (Table 2). Approximately 70% of the embryos co-injected with Cas13d mRNA and gRNAs at least three times had morphological malformations with curved and shrunken trunks (Figure 8) . No morphological change was found in the embryos injected with Cas13d mRNA or gRNAs alone. As a result, the observed morphological changes were likely a specific phenotype of shisa-2 knockdown (Figure 8). To confirm the specific phenotype, a rescue experiment using flounder shisa-2 mRNA was performed. In that experiment, the percentage of embryos with severely curved trunk decreased, and also some rescued embryos displayed a weakly cured trunk (supplementary picture S1- S2).
Because the embryonic trunk was abnormal after knockdown, somite formation was analyzed using the mesp-ab expression-somite formation marker gene. The results of in situ hybridization at 11 hpf demonstrated that the signal of mesp-ab expression was scattered and not clustered as a stripe (Figure 9) in the newly formed somites in the knockdown group. In contrast, two stripes were observed in the control groups. In addition, the space between the two mesp-ab positive group cells was wider than that of the control groups (Figure 9), indicating that the convergent extension of somatic precursors was delayed.
3. Discussion
As adaptor proteins, Shisa family proteins play important roles during animal development. However, the number of Shisa family genes present in zebrafish remains unclear. Additionally, their expression patterns and roles during zebrafish embryonic development are still unknown. In this study, 10 shisa family genes were identified in zebrafish and classified into the following nine subfamilies: shisa-1 (previously named shisa 2a), shisa- 2 (shisa 2), shisa- 3 (shisa 3), shisa- 4 (shisa 4), shisa- 5 (shisa 5), shisa- 6 (shisa 6), shisa- 7 (previously named shisa 7 like), shisa 8 (predicted name shisa-8b), and shisa- 9 (shisa 9a and shisa 9b). A specific syntenic block containing atp8a2 and shisa-2 was found to be conserved across all species. We also found that shisa-2 was a regulator of the convergent extension cell movement of the somatic precursors during embryonic development.
3.1. Different Evolutionary Processes for Shisa Subfamily Genes
In this study, shisa 1 was first confirmed in zebrafish. Vertebrates have at least nine shisa genes. shisa- 2–9 were discovered in all tested species. shisa-1 was not presented in amniotes and has been reported only in X.laevis, zebrafish and Oncorhynchus mykiss [4,17]. shisa-1 and shisa-2 might be produced from genome duplication as they were in the same subclade. African clawed frog and fish are known to have gone through tetraploidization or whole genome duplication [17]. Others studies [4,17] and Our finds indicated that Shisa-1 subfamily was separated from subfamily Shisa-2,which implied that Shisa-1 was not originated from tetraploidization or whole genome duplication. However, putative Shisa-1 genes in birds and the elephant shark Callorhinchus milii had been predicted in their genomic sequences [4]. It is more likely that shisa-1 gene has degenerated in mammal. As a result, the occurrence of shisa-1 and shisa-2 genes might be due toa gene duplication – complementation – degeneration model [17]. Additionally, we found a specific syntenic block containing atp8a2 and shisa-2 conserved across all species. Theatp8a2 is considered an evolutionarily conserved gene [19];therefore, shisa-2 might be the original subclade gene of the shisa-1 and shisa-2 subfamilies.
Shisa-8 is considered as a mammalian-specific gene duplication of shisa-9 [4]; however,wehave isolated it in zebrafish for the first time.. Our findings further suggest that shisa- 8 and shisa- 9 might have originated from genome duplication. In zebrafish, subfamily-specific gene duplications have been observed in two shisa-9s (shisa-9a and shisa-9b). In fish, many genes are found with duplicated gene phenomena, including pax3a/3b and pax7a/7b in zebrafish, medaka (Oryzias latipes), tetraodon (Tetraodon nigroviridis), fugu (Takifugu rubripes), and olive flounder (Paralichthys olivace) [20,21,22,23]. An additional specific genome duplication (FSGD) occurred in the teleost lineage before the beginning of teleost radiation [24]. Two different shisa- 9 genes are more likely to originate from fish-specific genome duplication.
3.2. Conserved Domains and Divergent C-terminal Regions of Shisa Proteins
Apart from the conserved cysteine-rich domain of Shisa family proteins in the N-terminus,everal cysteine residues were present near the C-termini of the predicted TMs (Figure 2) of zebrafish Shisa-1–5, which have been proposed as potential sites for lipid modifications such as palmitoylation [25]. Such modifications stabilize the protein and transport it to specific membranes [25].Therefor, zebrafish Shisa-1–5 might be involved in cellular signaling or membrane trafficking through the modification.Subfamily Shisa-6–9 might not have a specific function for there were no cysteine residues for lipid modification near the C-termini of their predicted TMs. Through further analysis, we found that these cysteine residues are regard as the subfamily signatures, with “CCC[KQ]C,” “CCCRC,” and “YCCTC” motifs [4] in zebrafish Shisa-1, Shisa-2 and Shisa-3, respectively (Figure 2),although these cysteine residues were arranged as “CC[FR]*CSCC” and “C**C[SP]CC*****C” in Shisa-4 and Shisa-5, respectively (Figure 2) [4]. In the phylogenetic tree, Shisa-1, Shisa-2, and Shisa-3 were in same clade (Figure 1), while Shisa-4 and Shisa-5 were in same clade (Figure 1). Therefore, “[C/Y]CC[KQT]C” might be the clade signature for Shisa-1–3, while “*C**C[SP]CC” might be the clade signature for Shisa-4 and Shisa-5. X. laevis Shisa-2 and Shisa-3 were ER-specific proteins [6], while Shisa-1 was expressed as both an ER-residing and a secreted form [5]. Previous studies suggested that there was no ER retention signal in these three shisa subfamily proteins [5,6], and their location on ER was the key to their function in mediating cellular signaling [5,6,7]. Both the amino half of Shisa-1, which includes these cysteine residues and the conserved cysteine-rich domain, and the left carboxy half were important mediators of the cellular signaling in X.laevis [5]. Thus, the ER retention signals in these three Shisa subfamily proteins might scatter at different locations not only in the amino half but also in the carboxy half. Comparative analysis of Shisa-1 and Shisa-2/3 protein sequences might give us a clue. However, the conserved cysteine-rich domain of mouse SHISA-5 was required for its interaction with other proteins and the proline-rich region in the carboxy half was for its ER-specific location [15]. This suggested that both the amino half and the carboxy half were essential for its effective functioning in the correct position, which could also be why only half of X.laevis shisa-1 did not correctly mediate its signal transmission function alone [5]. But, we cannot exclude the cysteine residues of clade feature from the interaction between these Shisa proteins and other proteins or the ER-specific expression, because they were retained in the process of protein-protein interactions and ER-specific expression analysis. In zebrafish Shisa-1–3, a region with multiple-prolines that was not accumulated as like mouse SHISA-5 was observed. It needs to be proven in the future whether these proline regions play the same roles as in mouse SHISA-5.An analysis of these conserved sequences might be help to explain the functional conservation of these genes among different species as well as the similarities and differences between them and other subfamily genes.
Functional divergence after gene duplication for Shisa family proteins in vertebrates has divergent C-terminal regions [4]. In zebrafish, PY motifs were presented in their C-terminal regions of Shisa- 3, 4, 5, and 7, and a PY-like motif, “PLSY” and “PTGY” in Shisa-6 and 8, respectively (Figure 2). As the PY motif can interact with NEDD4 family proteins, zebrafish Shisa- 3, 4, 5, and 7 may be adaptor proteins that regulate the ubiquitination and degradation of other target proteins [4]. Future studies should analyze whether the function of the PY-like motif is the same as that of PY-motif. The PDZ-domain interacting motif ([ST]x[VLI]) “VTV” or “VTI” is present at the C-terminus of zebrafish Shisa-1 and Shisa-7–9, which is a “VTM” in Shisa-2. Many proteins containing PDZ-domain play a central role in scaffolding macromolecular complexes, which is critical to signaling and trafficking pathways [26,27]. As a result, the function of zebrafish Shisa-1 and Shisa- 7–9 might be carried out through their PDZ-domain interacting motif. Whether zebrafish Shisa-2 functions through the “VTM” needs to be studied further. In zebrafish Shisa-6, no PDZ-domain interacting motif is present, which is different from that in other species [4]. The function of zebrafish Shisa-6 may differ from that of other species, or the PDZ-domain interacting motif may not play a role in Shisa-6 functional implementation. In zebrafish Shisa-7, a GRID domain is present, which has been proven to be critical for the interaction between Shisa 7 and the GABAA receptor, further promoting GABAA receptor trafficking to the cell surface in the mouse brain [12].Zebrafish Shisa-7 might therefor be important for the neurological development.
3.3. Extensive Participation of Shisa Family Genes During Zebrafish Embryonic Development
Shisa plays various roles in developmental processes [4]. The expression patterns of shisa family genes suggest they might play different roles during embryonic development. shisa-3was expressed highly at 0.2 hpf, the first cellstage , and 16 hpf. At first cell stage, cytoplasmic movements begin [28] and the first cell appear. At 16 hpf, the peripheral and central sensory axons extend to their destination location [28]; therefore, shisa-3 might be critical to these developmental processes. Notably, shisa-9b, which might originate from fish-specific genome duplication, was expressed from12hpf. The Kupffer's-vesicle appeared from at approximately this time (11.7 hpf) [28]. Whether this gene represents the formation of Kupffer's-vesicle still needs to be investigated. The semi-quantitative PCR results suggested zebrafish shisa family genes might be involved different development processes; however, the actual participation process should be determined through spatial expression and functional analysis in the future.
3.4. Shisa-2 Regulating the Convergent Extension Cell Movement of the Somatic Precursors in Zebrafish
The results of in situ hybridization demonstrated that zebrafish shisa-2 was expressed in the presomitic mesoderm (PSM), somites, optic vesicles, gill arches, and head regions. The expression almost like that of X. laevis, chicken, and mouse Shisa2 [6,14,17,30]. The conserved expression in somite and PSM suggested that shisa-2 might be involved in the somite formation. Indeed, the convergent extension cell movement of somatic precursors and the maturation of somitic precursors are affected by shisa2 in X.laevis [6]. The effect of zebrafish shisa-2 on the somite formation was preliminary analyzed through knockdown. In wild type zebrafish, mesp- ab is expressed as one or two stripes in the front compartment of potential somites in the anterior presomitic mesoderm [31]. In shisa-2 knockdown zebrafish, the space between the left and right stripes of mesp- ab expression was wider than that in the control, and the expression was scattered and not clustered as a stripe. This result indicated that the convergent extension cell movement of somatic precursors was delayed after shisa-2 expression was downregulated. Therefore, shisa-2 may play a crucial role in the convergent extension cell movement of somatic precursors in zebrafish. The function of shisa2 might be conserved between zebrafish and African clawed frog. In X.laevis, shisa2 regulated proper segmental patterning through individual inhibition of Wnt and FGF signaling [6],while in C2C12 myoblast, shisa2 promoted myoblast fusion via Rac1/Cdc42-mediated cytoskeletal F-actin remodeling [8].The exact in vivo function and mechanism of shisa-2 in zebrafish need be explored in the future.
3.5. Limitations
The expression of the Shisa family genes suggested that all of them might be crucial in embryonic development. We were unable to confirm whether they have a tissue- or cell-specific expression pattern and function. As a result, the spatiotemporal expression should be investigated using other methods, such as in situ hybridization, in the future. The function of shisa-2 was only preliminarily analyzed through the recently established Cas13d-mediated gene knockdown in the embryonic stage. The method still has some shortcomings, such as a lack of efficiency detection and specificity analysis methods. Thus, effective and specific detection methods should be discovered in the future. A homozygous mutant with a shisa-2 knockout should be used to analyze its function in the future as well.
4. Materials and Methods
4.1. Sample Collection
Wild-type zebrafish (TU strain) were cultured in a recirculation culture system at the institute aquarium (temperature: 28.5 ± 1 °C; light/dark cycle = 14 h/10 h). Fish were fed twice daily with commercial particulate food and once daily with brine shrimp. Fertilized eggs were obtained by mixing one male and two female fish in the morning. After washing with cycling water several times, fertilized eggs of the zebrafish were collected in a Petri dish and were ready for microinjection or sample collection.
For sample collection, the embryos were incubated in an incubator at 28.5 ± 1 °C, obtained at different developmental stages. Approximately 30 embryos were pooled for each sample, placed in a centrifugation tube, immediately frozen in liquid nitrogen, and stored at −80 °C until RNA extraction. The wild-type zebrafish embryos and the embryos injected with Cas13d mRNA and gRNA at 12 hpf were fixed in 4% paraformaldehyde in 1x phosphate-buffered saline (PBS) overnight at 4 °C. The embryos were dehydrated and stored in 100% methanol for in situ hybridization.
4.2. Total RNA Isolation and cDNA Synthesis
Each sample was placed in a centrifugation tube containing 200 μL of TRIzol Reagent (Toroivd Tech. Comp., Shanghai, China) and homogenized using a grinding rod. Total RNA was extracted according to the manufacturer’s instructions. The quality of isolated RNA was checked using 1.5% agarose gel electrophoresis, and its concentration and purity were determined using a NanoDrop 2000 spectrophotometer (Nanodrop Technologies, USA). For each sample, 1μg of RNA was transcribed into cDNA using the TransScript® One-step gDNA Removal and cDNA Synthesis SuperMix kit (TransGen Biotech, Beijing,China) with OligdT.
4.3. Cloning of Zebrafish Shisa Family Genes
All Shisa family genes were downloaded from the annotated zebrafish genome in the National Center for Biotechnology Information (NCBI) database (https://www.ncbi.nlm.nih.gov/). Primers were designed based on the predicted ORF sequences (Table 3). PCR was performed using GoldStar Taq DNA Polymerase (CWBIO, Ltd., China) and a mixed cDNA template of different developmental stages of zebrafish embryos. The PCR conditions were as follows: 5 min at 94 °C, 35 cycles of 5 s at 94 °C, 30 s at 50–60 °C, 2 min at 72 °C, and a final 5 min extension at 72 °C. All genes were cloned, ligated into TOPO vectors, and sequenced.
4.4. Bioinformatics Analysis
Phylogenetic analysis was performed using the amino acid sequences of the Shisa family genes from H. sapiens, M. musculus, G. gallus, X. laevis, X. tropicalis, and D. rerio (Table 1). A Maximum likelihood tree was constructed using MEGA 7.0 with 1000 bootstrap replicates [18]. All amino acid sequences of zebrafish shisa were aligned and the average amino acid identity was calculated using DNAMAN 11.0 (http://www.lynnon. com). For Synteny analysis, TBtools [32] was used to obtain the location of the target gene in H. sapiens, M. musculus, G. gallus, X. tropicalis, and D.rerio, and all of the gene information, such as gene type and location, were visualized using the ggplot2 tool [33] of R programing software. Based on the zebrafish genomic sequence annotation (GRCz11), the gene structure was revealed in TBtools [32]. The motif, including the signaling peptide, was predicted using the SMART online software (http://smart.embl-heidelberg.de/) combined with the Pfam, SignalP, and TMHMM databases. The predicted structural domains were visualized using the IBS software (Illustrator for Biological Sequences).
4.5. Knockdown of shisa 2 in Zebrafish Embryos Using Cas13d mRNA and gRNAs
Zebrafish shisa 2 was knockdown using Cas13d mRNA and gRNAs as described by Kushawah et al [34]. The Cas13d ORF fragment was cloned using primers (Table 4, Cas13D-F-psp64-T7/Cas13D-R-psp64-T7) with a T7 promoter in the 5’-end and the KOD enzyme. After purification, the PCR product was recombined into the Sal I and BamH I sites of the psp64 polyA vector using the EZ clone and named Cas13d/psp64. The Cas13d/psp64 was linearized using Pvu II (Takala, Dalian, China) and purified by phenol/chloroform as a template to synthesize the Cas13d mRNA using mMESSAGE mMACHINE™ T7 Transcription Kit (ThermoFisher Scientific, Waltham, MA, USA).. Four guide RNAs (gRNA 1–4) were designed using online software (https://cas13design.nygenome.org/) [35,36]. The gRNA DNA template was generated by PCR using pfu enzyme and primers (Table 4; Cas13d-Universal-F and Cas13D-zfshisa2- gRNA1–4). After purification with phenol/chloroform, the PCR products were used as templates to synthesize gRNA using a TranscriptAid T7 High Yield Transcription Kit (ThermoFisher Scientific, Waltham, MA, USA).. Following synthesis, the Cas13d mRNA and gRNAs were purified using SigmaSpin™ Post-Reaction Clean-Up Columns (Sigma Aldrich, St. Louis, MO, USA) .
At one cell stage, the optimized concentration of Cas13d mRNA (200 ng/μL) and gRNAs (total 800 ng/μL, 200 ng/μL for each) together, Cas13d mRNA (200 ng/μL), or gRNAs (total 800 ng/μL, 200 ng/μL for each ) in 0.2M KCl were microinjected into the cell of zebrafish embryo using a microinjector (Pli-100; Harvard Apparatus, Holliston, MA, USA).
At 48 hpf, after the chorion was removed, the embryos were observed in water under a microscope (DM LB2, Wetzlar, Germany) and photographed with a WeiTu HTC2000 digital camera, the number was counted under stereoscope (Leica 12.5, Wetzlar, Germany).
4.6. Rescue Using Flounder shisa-2 mRNA
Flounder shisa-2 was cloned using primers (Table 5, floundershisa2-F/R) and ligated into pEASY –T3, which was termed as flounder-shisa2/T3. After the sequence had been confirmed, the open read frame was cloned using primers (flounder-shisa2psp64-F/R) with the pfu enzyme and the flounder-shisa2/T3 plasmid as the template [37]. After purification, the PCR product was recombined into the Sal I sites of the psp64 polyA vector using the EZ clone and named floundershisa2/psp64. The flounder-shisa2/psp64 was linearized using Xba I (Takala, Dalian, China) and purified by phenol/chloroform as a template to synthesize the flounder shisa-2 RNA using mMESSAGE mMACHINE™ Sp6 Transcription Kit (ThermoFisher Scientific, Waltham, MA, USA).
At one cell stage, the optimized concentration of Cas13d mRNA (200 ng/μL), and gRNAs (total 800 ng/μL, 200 ng/μL for each) with or without flounder shisa-2 mRNA (24 ng/μL) in 0.2 M KCl were microinjected into the cells of zebrafish embryos.
At 48 hpf, the embryos were treated, observed, photographed, and counted as knockdown (section 4.5).
4.7. Semi-quantitative PCR
The primers (Table 6) used for semi-quantitative PCR of zebrafish shisa genes were designed using Primer3 (https://www.primer3plus.com/index.html) and Primer-BLAST of NCBI (https://www.ncbi.nlm.nih.gov/tools/primer-blast/).For the semi-quantitative PCR, ef-1a was used as an internal reference gene. The total PCR volume was 25 μL and included different amount cDNA templates, 12.5 μL of 2 × Taq Master Mix (Vazyme, China), 1 μL of forward primer, 1 μL of reverse primer, and 9.5 μL of ddH2O. The amount of cDNA templates in the other gene response systems is 2 μL, except for shisa-3, shisa-4 and shisa-9b, which had an amount of 4 μL because their expressions were very weak. The PCR protocol was as follows: 95 °C for 3 min, 94 °C for 15 s, 55 °C for 15 s, 72 °C for 15 s, 35 cycles, and finally 72 °C for 5 min. All RT–PCR experiments were performed in triplicate. The corresponding plasmids, which were obtained during gene isolation (section4.3) ,were used as positive control.
4.8. Whole Mount in situ Hybridization.
Whole mount in situ hybridization performed by modifying the method described by Du and Dienhart [38]. Briefly, digoxigenin-labeled RNA probes were synthesized against shisa 2 and mesp-ab mRNA. Subsequently, the fixed embryos were dechorionated, rehydrated in 50% methanol in PBST for 5 min, and washed with PBST (1x PBS pH 7.4 + 0.1% Tween 20) for 5 min. The embryos were soaked in the prehybridization buffer for 4 h at 65 °C for prehybridization and incubated in hybridization buffer with each RNA probe (100–200 ng) overnight. An NBT(nitroblue tetrazolium chloride)/BCIP(5-bromo-4-chloro-3-indolyl phosphate) substrate was added to detect alkaline phosphatase until a color developed, and the reaction was stopped by rinsing with PBST. Embryos were photographed in glycerol under a microscope (Leica DM LB2, Wetzlar, Germany) with a WeiTu HTC2000 digital camera.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Figure S1-S2.
Author Contributions
Conceptualization, X.T.; methodology, Y.L., N.D., B. Q., C.Z. and X.T.; investigation, Y.L., N.D., B. Q., C.Z., Y.Z.,L.W., F.X., S. Q., and X.T.; data curation, Y.L. and X.T.; writing-original draft preparation, Y.L. and X.T.; writing-review and editing, X.T.; supervision F.Y.; funding acquisition, X.T. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by Shandong Provincial Natural Science Foundation (Project No. ZR2022MC129) and the National Marine Genetic Resource Center.
Institutional Review Board Statement
The study was approved by the Animal Care and Use Committee of institute of Oceanology, Chinese Academy of Sciences.
Data Availability Statement
All data generated or analyzed during this study are included in this published article and the supplementary file.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pan, C.Q.; Sudol, M.; Sheetz, M.; Low, B.C. Modularity and functional plasticity of scaffold proteins as p(l)acemakers in cell signaling. Cell Signal 2012, 24, 2143–2165. [Google Scholar] [CrossRef] [PubMed]
- Zeke, A.; Lukacs, M.; Lim, W.A.; Remenyi, A. Scaffolds: interaction platforms for cellular signalling circuits. International Journal of Developmental Biology 2009, 19, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Langeberg, L.K.; Scott, J.D. Signalling scaffolds and local organization of cellular behavior. Nature Reviews Molecular Cell Biology 2015, 16, 232–244. [Google Scholar] [CrossRef] [PubMed]
- Pei, J.; Grishin, N.V. Unexpected diversity in Shisa-like proteins suggests the importance of their roles as transmembrane adaptors. Cellular Signalling 2012, 24, 758–769. [Google Scholar] [CrossRef]
- Yamamoto, A.; Nagano, T.; Takehara, S.; Hibi, M.; Aizawa, S. Shisa promotes head formation through the inhibition of receptor protein maturation for the caudalizing factors, Wnt and FGF. Cell 2005, 120, 223–235. [Google Scholar] [CrossRef]
- Nagano, T.; Takehara, S.; Takahashi, M.; Aizawa, S.; Yamamoto, A. Shisa2 promotes the maturation of somitic precursors and transition to the segmental fate in Xenopus embryos. Development 2006, 133, 4643–4654. [Google Scholar] [CrossRef]
- Onishi, K.; Zou, Y. Sonic Hedgehog switches on Wnt/planar cell polarity signaling in commissural axon growth cones by reducing levels of Shisa2. eLife 2017, 6, e25269. [Google Scholar] [CrossRef]
- Liu, Z.J.; Wang, C.; Liu, X.Q.; Kuang, S. Shisa2 regulates the fusion of muscle progenitors. Stem Cell Research 2018, 31, 31–41. [Google Scholar] [CrossRef]
- Chen, C.C.; Chen, H.Y.; Su, K.Y.; Hong, Q.S.; Yan, B.S.; Chen, C.H.; Pan, S.H.; Chang, Y.L.; Wang, C.J.; Hung, P.F.; Yuan, S.; Chang, G.C.; Chen, J.J.W.; Yang, P.C.; Yang, Y.C.; Yu, S.L. Shisa3 is associated with prolonged survival through promoting beta-catenin degradation in lung cancer. American Journal of Respiratory and Critical Care Medicine 2014, 190, 433–444. [Google Scholar] [CrossRef]
- von Engelhardt, J.; Mack, V.; Sprengel, R.; Kavenstock, N.; Li, K.W.; Stern-Bach, Y.; Smit, A.B.; Seeburg, P.H.; Monyer, H. CKAMP44: a brain-specific protein attenuating short-term synaptic plasticity in the dentate gyrus. Science 2010, 327, 1518–1522. [Google Scholar] [CrossRef]
- Gupta, R.K.; Tripathi, R.; Naidu, B.J.; Srinivas, U.; Shashidhara, L. Cell cycle regulation by the pro-apoptotic gene Scotin. Cell Cycle 2014, 7, 2401–2408. [Google Scholar] [CrossRef]
- Han, W.; Li, J.; Pelkey, K.A.; Pandey, S.; Chen, X.; Wang, Y.X.; Wu, K.W.; Ge, L.; Li, T.M.; Castellano, D. Shisa7 is a GABA(A) receptor auxiliary subunit controlling benzodiazepine actions. Science 2019, 366, 246–250. [Google Scholar] [CrossRef]
- Abdollahi Nejat, M.; Klaassen, R.V.; Spijker, S.; Smit, A.B. Auxiliary subunits of the AMPA receptor: The Shisa family of proteins. Current Opinion in Pharmacology 2021, 58, 52–61. [Google Scholar] [CrossRef]
- Hedge, T.A.; Mason, I. Expression of Shisa2, a modulator of both Wnt and Fgf signaling, in the chick embryo. The International Journal of Developmental Biology 2008, 52, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Bourdon, J.C.; Renzing, J.; Robertson, P.L.; Fernandes, K.N.; Lane, D.P. Scotin, a novel p53-inducible proapoptotic protein located in the ER and the nuclear membrane. Journal of Cell Biology 2002, 158, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Kim, M.J.; Sung, P.S.; Bae, Y.C.; Shin, E.C.; Yoo, J.Y. Interferon-inducible protein SCOTIN interferes with HCV replication through the autolysosomal degradation of NS5A. Nature Communication 2016, 7, 10631. [Google Scholar] [CrossRef] [PubMed]
- Furushima, K.; Yamamoto, A.; Nagano, T.; Shibata, M.; Miyachi, H.; Abe, T.; Ohshima, N.; Kiyonari, H.; Aizawa, S. Mouse homologues of Shisa antagonistic to Wnt and Fgf signalings. Developmental Biology 2007, 306, 480–492. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7. 0 for Bigger Datasets. Molecular Biology and Evolution 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Shams, F.; Dyer, F.; Thompson, R.; Duncan, R.P.; Thiem, J.D.; Kilian, A.; Ezaz, T. Application of DArT seq derived SNP tags for comparative genome analysis in fishes; An alternative pipeline using sequence data from a non-traditional model species, Macquaria ambigua. PLoS One 2019, 14(12), e0226365. [Google Scholar] [CrossRef]
- Minchin, J.E.; Hughes, S.M. Sequential actions of Pax3 and Pax7 drive xanthophore development in zebrafish neural crest. Developmental Biology 2008, 319, 530. [Google Scholar] [CrossRef]
- Minchin, J.E.N.; Williams, V.C.; Hinits, Y.; Low, S.; Tandon, P.; Fan, C.M.; Rawls, J.F.; Hughes, S.M. Oesophageal and sternohyal muscle fibres are novel Pax3-dependent migratory somite derivatives essential for ingestion (vol 140, pg 2972, 2013). Development 2013, 140, 4296. [Google Scholar] [CrossRef]
- Jiao, S.; Tan, X.G.; Wang, Q.; Li, M.J.; Du, S.J. The olive flounder (Paralichthys olivaceus) Pax3 homologues are highly conserved, encode multiple isoforms and show unique expression patterns. Comparative Biochemistry and Physiology B-Biochemistry & Molecular Miology 2015, 180, 7–15. [Google Scholar]
- Jiao, S.; Tan, X.G.; Li, M.J.; Sui, Y.L.; Du, S.J.; You, F. The duplicated paired box protein 7 (pax7) genes differentially transcribed during Japanese flounder (Paralichthys olivaceus) embryogenesis. Comparative Biochemistry and Physiology B-Biochemistry & Molecular Biology 2015, 189, 62–68. [Google Scholar]
- Venkatesh, B. Evolution and diversity of fish genomes. Current Opinion in Genetics & Development 2003, 13, 588–592. [Google Scholar]
- Linder, M.E.; Deschenes, R.J. Palmitoylation: policing protein stability and traffic. Nature Reviews Molecular Cell Biology 2007, 8, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Amacher, J.F.; Brooks, L.; Hampton, T.H.; Madden, D.R. Specificity in PDZ-peptide interaction networks: Computational analysis and review. J Struct Biol X 2020, 4, 100022. [Google Scholar] [CrossRef]
- Murciano-Calles, J. The Conformational Plasticity Vista of PDZ Domains. Life (Basel) 2020, 10(8), 123. [Google Scholar] [CrossRef]
- Kimmel, C,B; Ballard, W.W; Kimmel, S. R.; Ullmann, B.; Schilling,, T. F. Stages of embryonic development of the zebrafish. Dev Dyn 1995, 203(3), 253–310. [Google Scholar]
- Abdollahi Nejat, M.; Klaassen, R.V.; Spijker, S.; Smit, A.B. Auxiliary subunits of the AMPA receptor: The Shisa family of proteins. Current Opinion in Pharmacology. 2021, 58, 52–61. [Google Scholar] [CrossRef]
- Silva, A.C.; Filipe, M.; Vitorino, M.; Steinbeisser, H.; Belo, J.A. Developmental expression of Shisa-2 in Xenopus laevis. Int J Dev Biol 2006, 50(6), 575–579. [Google Scholar] [CrossRef]
- Cutty, S.; Fior, R.; Henriques, P.M.; Saude, L.; Wardle, F.C. Identification and expression analysis of two novel members of the Mesp family in zebrafish. International Journal of Developmental Biology 2012, 56, 285–294. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit De-veloped for Interactive Analyses of Big Biological Data. Molecular Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis. Springer 2016.
- Kushawah, G.; Hernandez-Huertas, L.; del Prado, J.A.N.; Martinez-Morales, J.R.; DeVore, M.L.; Hassan, H.; Moreno-Sanchez, I.; Tomas-Gallardo, L.; Diaz-Moscoso, A.; Monges, D.E.; Guelfo, J.R.; Theune, W.C.; Brannan, E.O.; Wang, W.; Corbin, T.J.; Moran, A.M.; Alvarado, A.S.; Malaga-Trillo, E.; Takacs, C.M.; Bazzini, A.A.; Moreno-Mateos, M.A. CRISPR-Cas13d Induces Efficient mRNA Knockdown in Animal Embryos. Developmental Cell 2020, 54, 805. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Rahman, J.A.; Wessels, H.H.; Méndez-Mancilla, A.; Haro, D.; Chen, X.; Sanjana, N.E. Transcriptome-wide Cas13 guide RNA design for model organisms and viral RNA pathogens. Cell Genom 2021, 1(1), 100001. [Google Scholar] [CrossRef] [PubMed]
- Wessels, H.H.; Méndez-Mancilla, A.; Guo, X.; Legut, M.; Daniloski, Z.; Sanjana, N.E. Massively parallel Cas13 screens reveal principles for guide RNA design. Nat Biotechnol 2020, 38, 722–727. [Google Scholar] [CrossRef] [PubMed]
- Du, S.J.; Dienhart, M. The zebrafish tiggy-winkle hedgehog promoter directs notochord and floor plate GFP expression in transgenic zebrafish embryos. Dev Dyn 2001, 222, 655–666. [Google Scholar] [CrossRef]
Figure 1.
Phylogenetic tree of Shisa proteins. The phylogenetic tree was constructed via MEGA7.0 [18] using the Maximum likelihood method with 1000 bootstrap replicates. The Genebank accession numbers for these genes are listed in Table 1. Notes: Confidence values higher than 70 are indicated in the tree.
Figure 1.
Phylogenetic tree of Shisa proteins. The phylogenetic tree was constructed via MEGA7.0 [18] using the Maximum likelihood method with 1000 bootstrap replicates. The Genebank accession numbers for these genes are listed in Table 1. Notes: Confidence values higher than 70 are indicated in the tree.
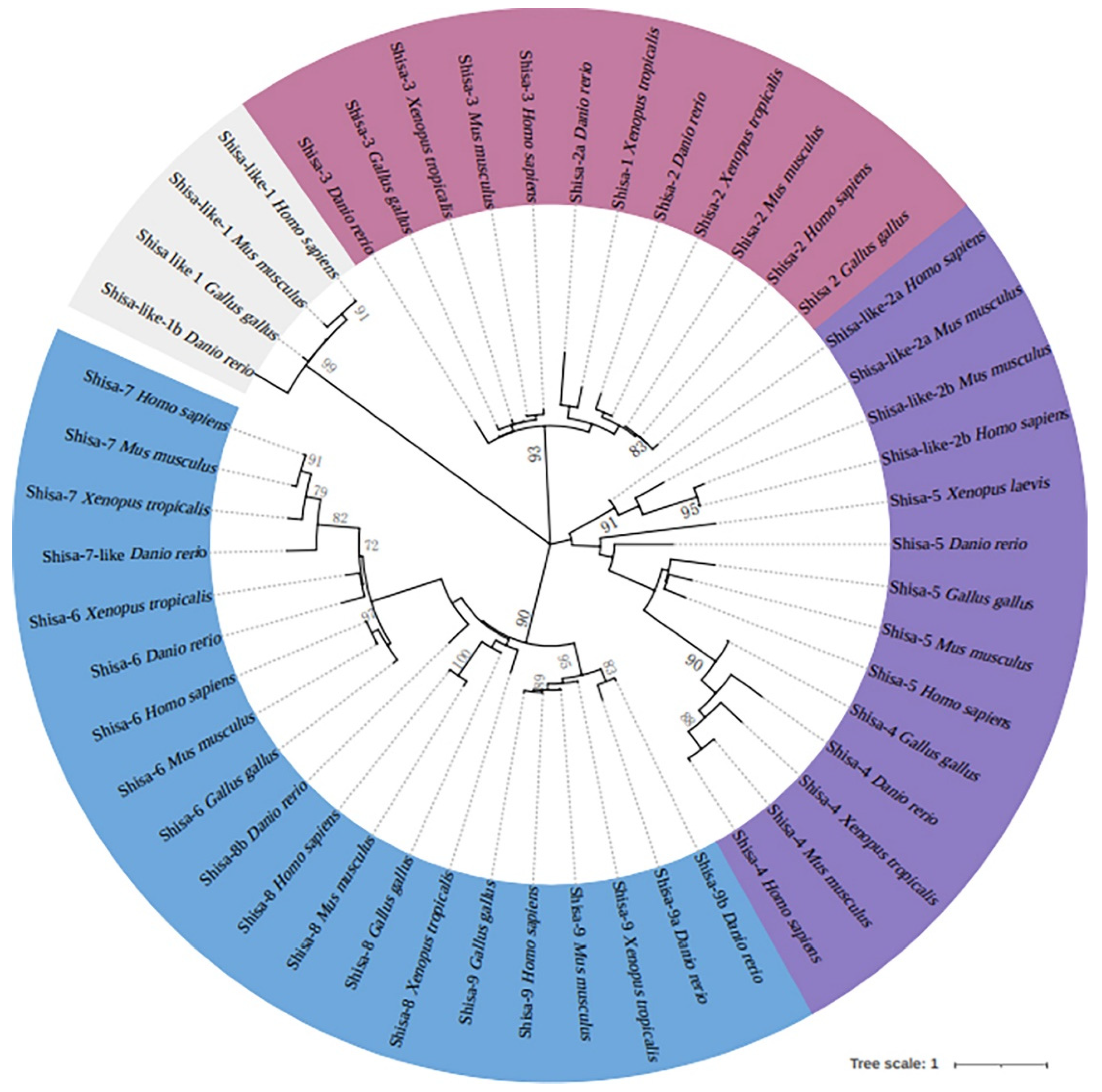
Figure 2.
Domain architecture of zebrafish Shisa proteins.
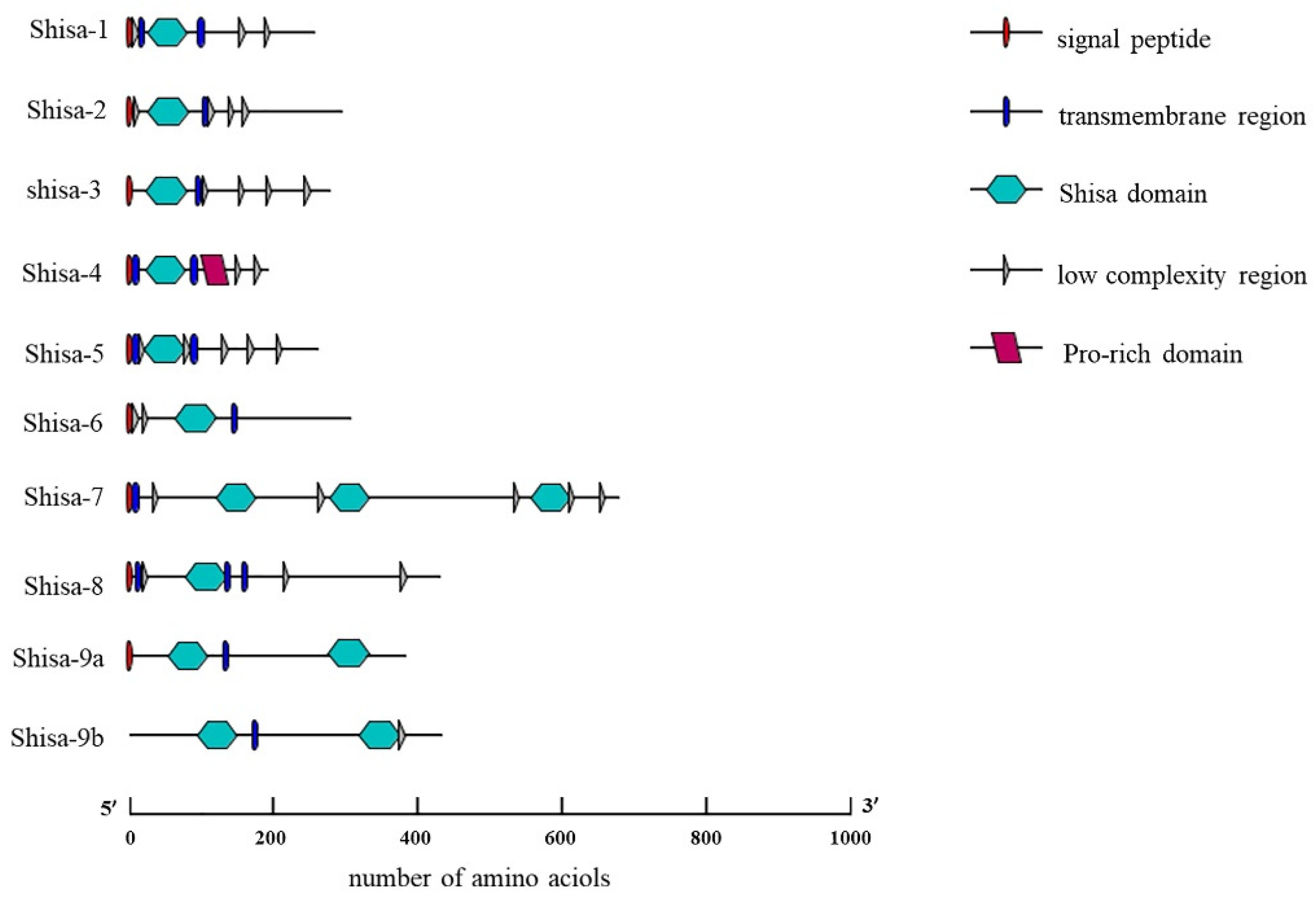
Figure 3.
Sequence alignment of zebrafish Shisa proteins.
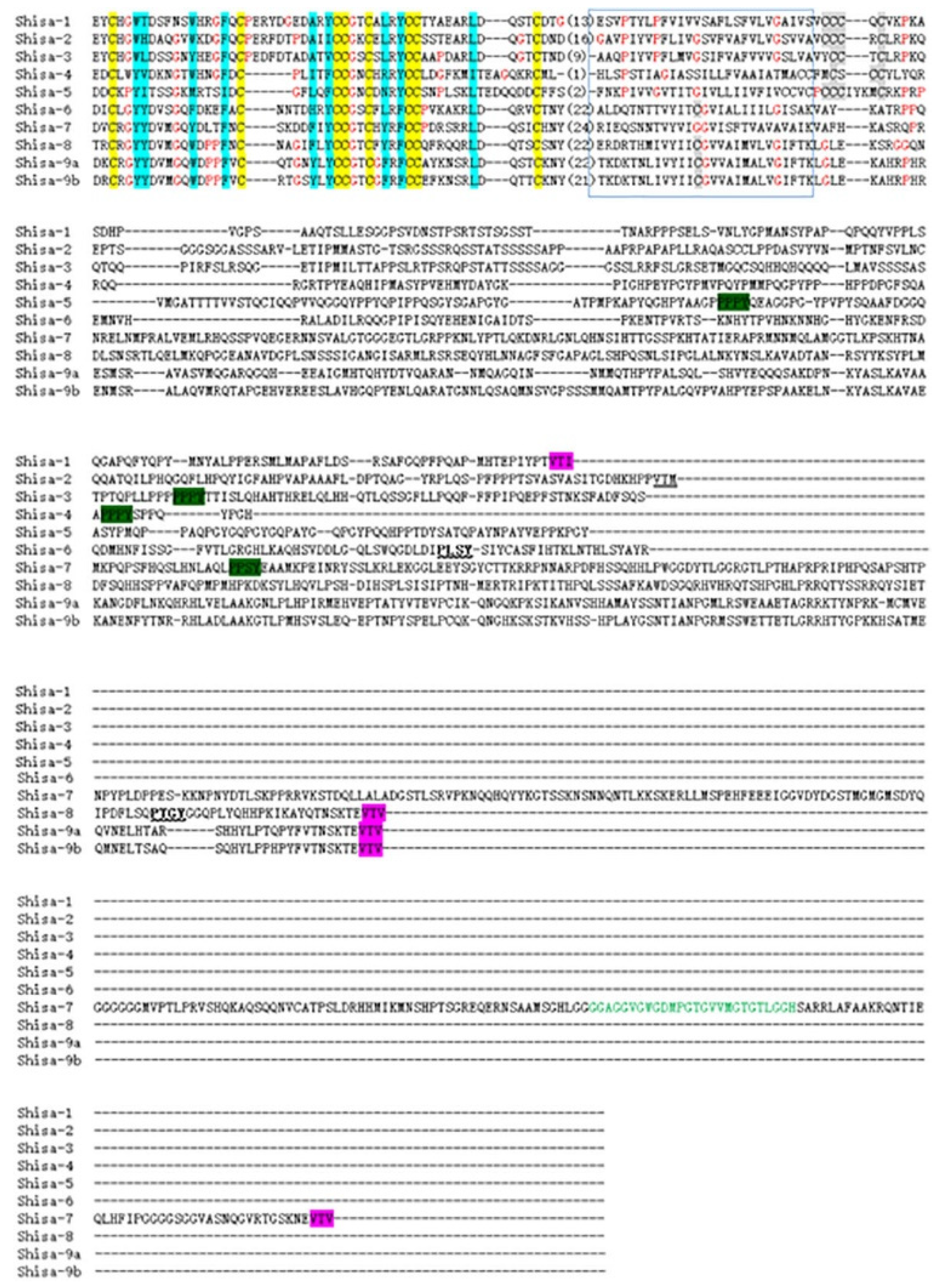
Figure 4.
Chromosomal location of the zebrafish Shisa family genes.
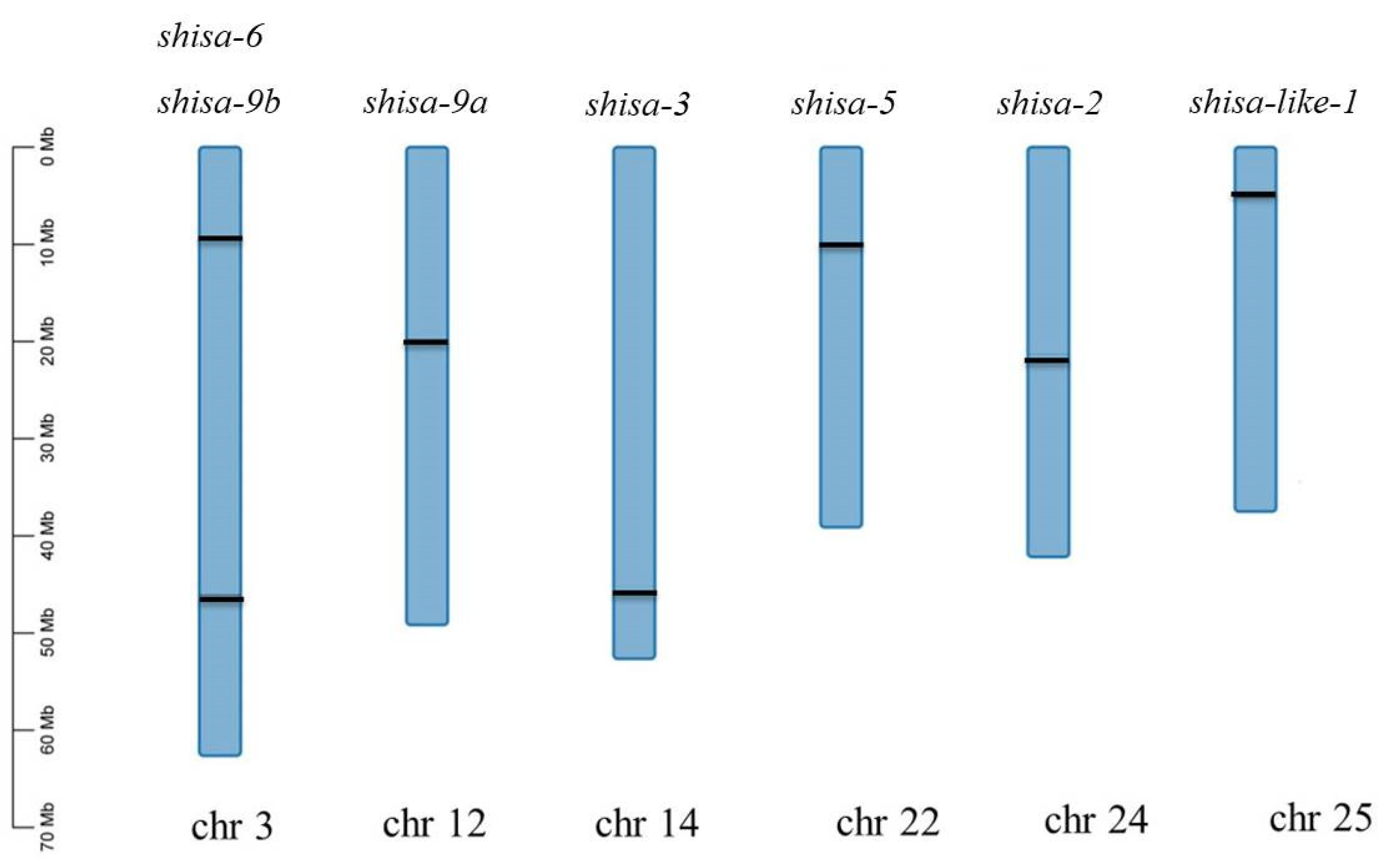
Figure 5.
Synteny analysis of shisa-2 neighbouring genes in different species that share the same ancestral species.
Figure 5.
Synteny analysis of shisa-2 neighbouring genes in different species that share the same ancestral species.
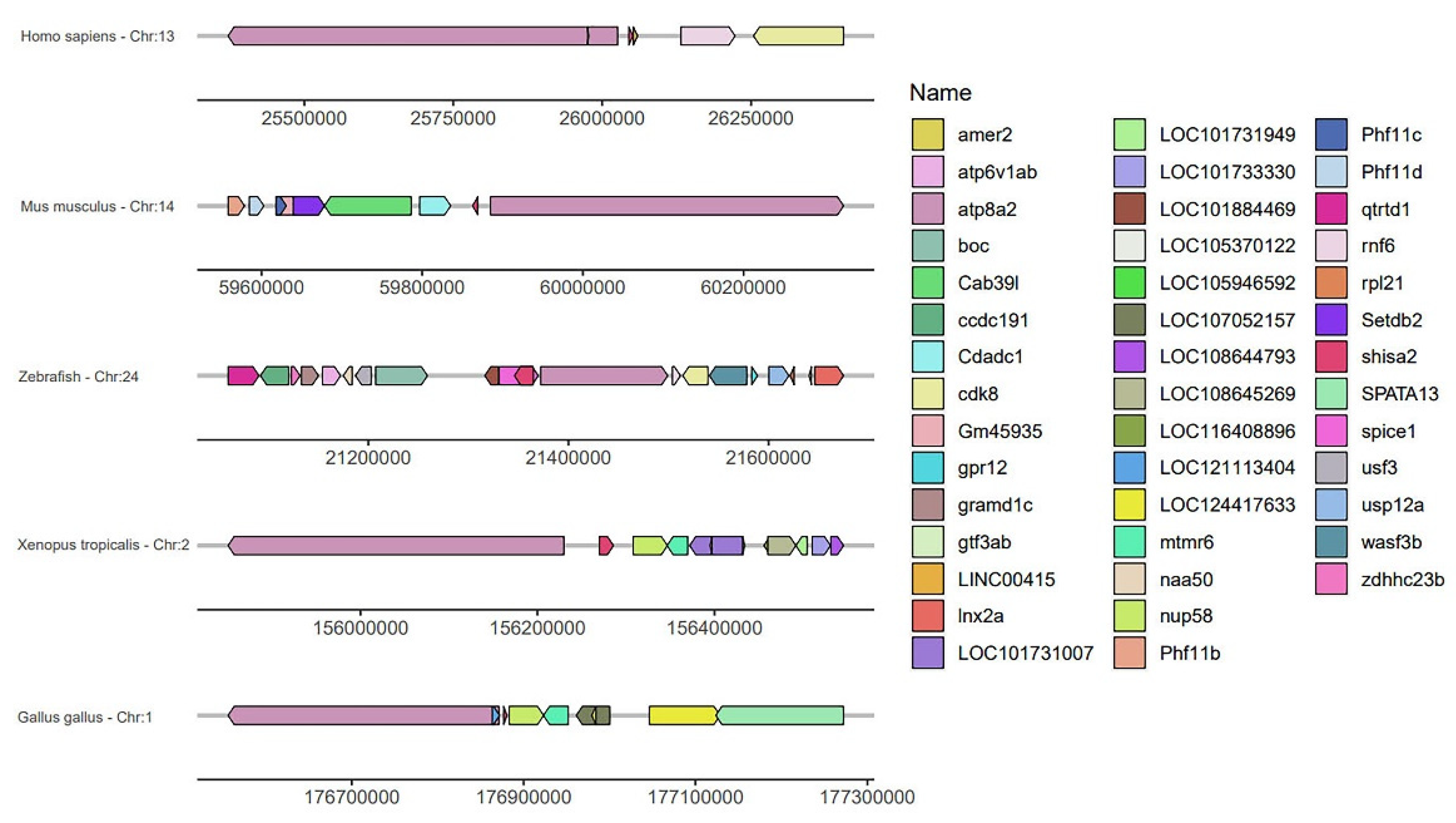
Figure 6.
Temporal expression patterns of zebrafish shisa genes during embryonic development. B, blank. PC, positive control.
Figure 6.
Temporal expression patterns of zebrafish shisa genes during embryonic development. B, blank. PC, positive control.
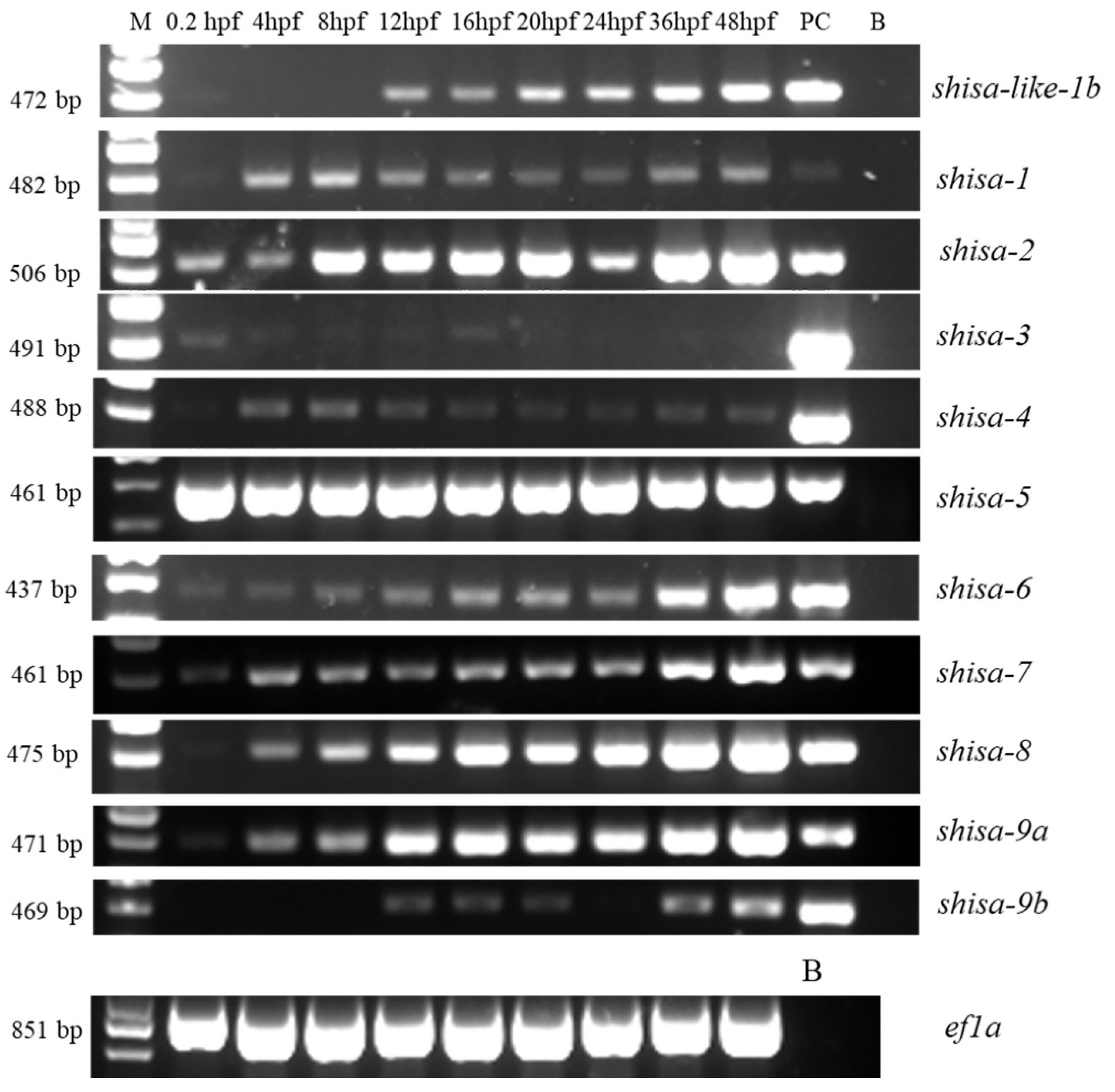
Figure 7.
Expression pattern of zebrafish shisa-2 during early embryonic development. Shisa-2 transcript was detected in He (Head region),So (Somite),Ov ( Optic vesicles),Pm (Pre-somitic mesoderm), Ga (Gill arch). A,B, 11hpf. C, 16hpf. D, E, 20hpf. A, Head to Top. Dorsal View. B,E. Dorsum to Top. Front View. C, Head to left. Dorsal View. D, Head to left, side view.Bar, 50 μm
Figure 7.
Expression pattern of zebrafish shisa-2 during early embryonic development. Shisa-2 transcript was detected in He (Head region),So (Somite),Ov ( Optic vesicles),Pm (Pre-somitic mesoderm), Ga (Gill arch). A,B, 11hpf. C, 16hpf. D, E, 20hpf. A, Head to Top. Dorsal View. B,E. Dorsum to Top. Front View. C, Head to left. Dorsal View. D, Head to left, side view.Bar, 50 μm

Figure 8.
Phenotype of zebrafish embryos(48 hpf)after shisa-2 knockdown. Bar, 200 μm, Magnification, 4X.
Figure 8.
Phenotype of zebrafish embryos(48 hpf)after shisa-2 knockdown. Bar, 200 μm, Magnification, 4X.
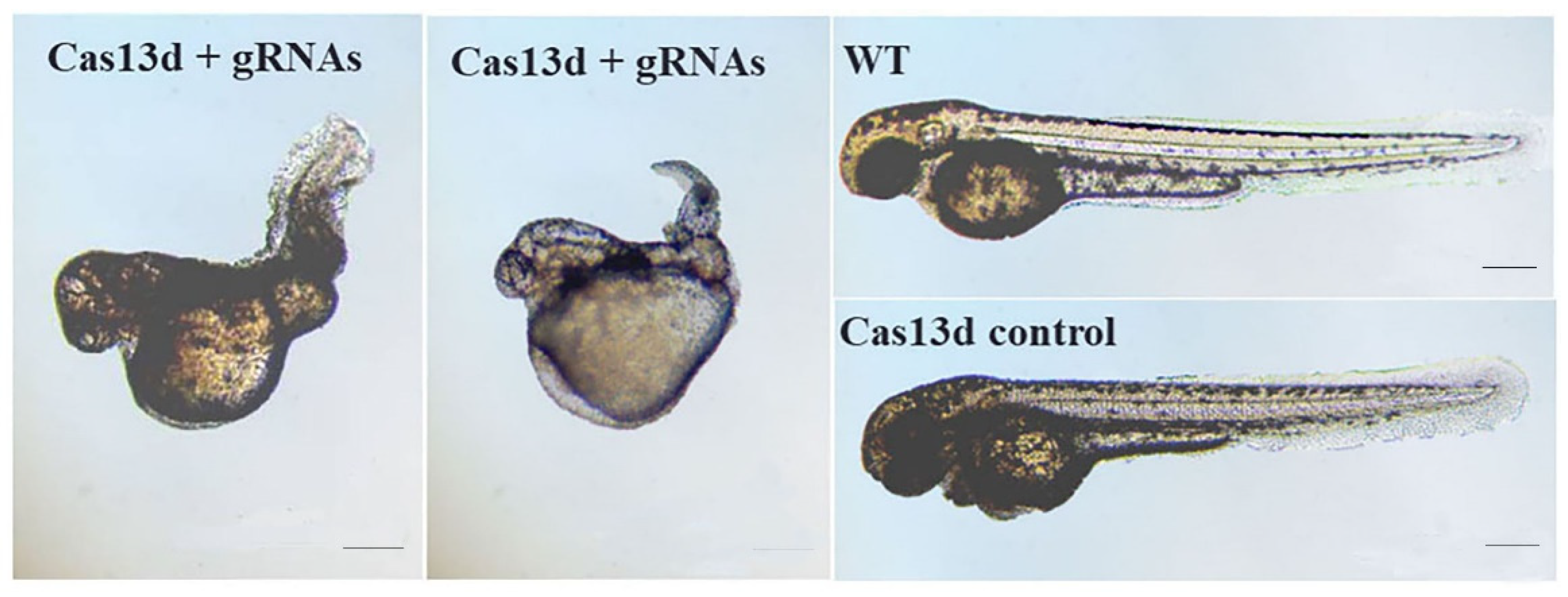
Figure 9.
Expression of mesp-ab in embryos injected with Cas13d mRNA +gRNA,Cas13d mRNA and no-injected embryos (wild type). The black arrows represent mesp-ab signal. Red arrows indicat the space between left and right stripes of mesp-ab. Head to Top, dorsal view. Bar, 100 μm.
Figure 9.
Expression of mesp-ab in embryos injected with Cas13d mRNA +gRNA,Cas13d mRNA and no-injected embryos (wild type). The black arrows represent mesp-ab signal. Red arrows indicat the space between left and right stripes of mesp-ab. Head to Top, dorsal view. Bar, 100 μm.
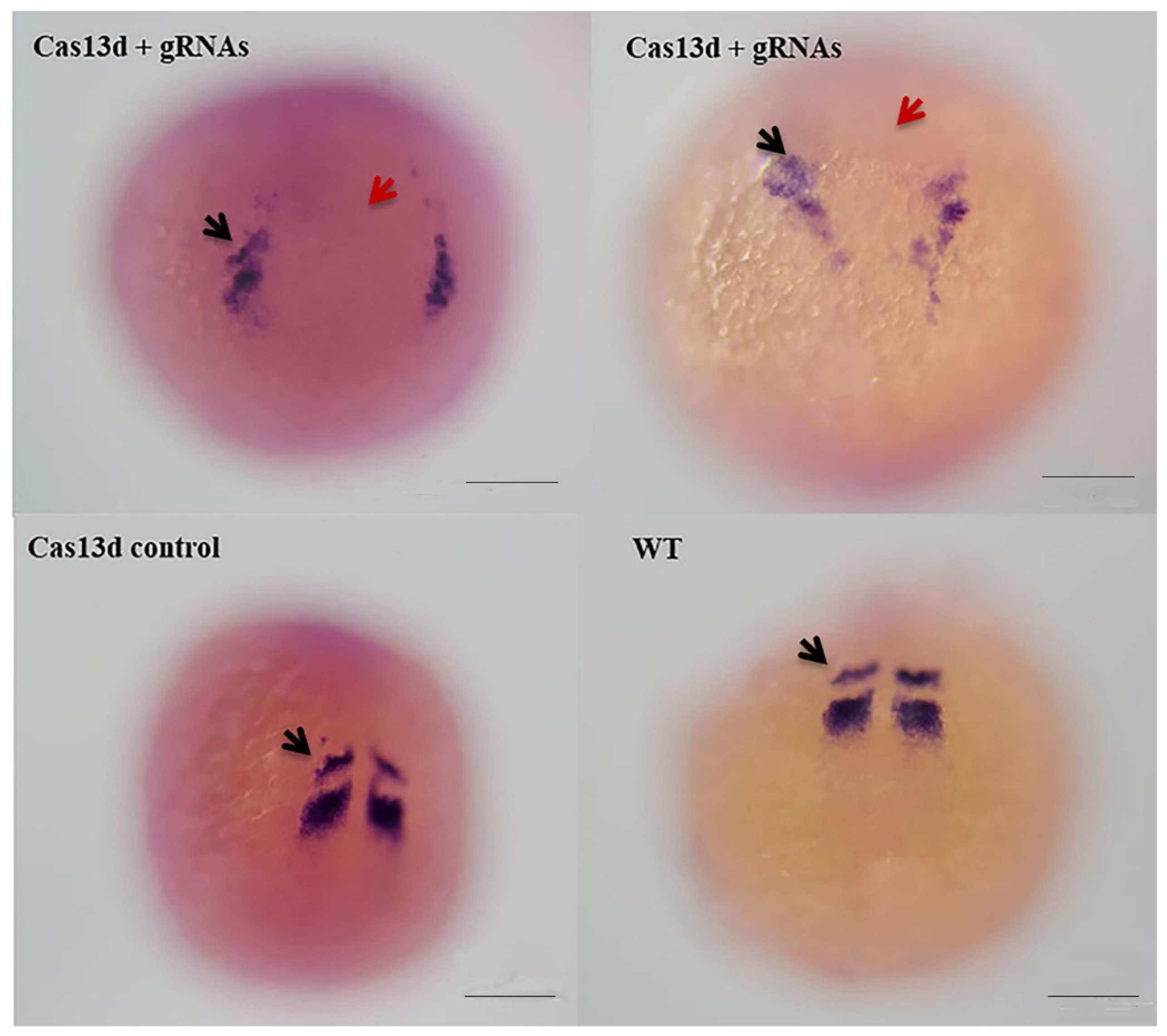

Table 1.
Shisa genes used in Maximum likelihood tree construction.
| Gene Name (NCBI name) | Species | GenBank accession number | |
|---|---|---|---|
| shisa-like 1b | Danio rerio | XM_002667571.6 | |
| shisa-1 ( shisa 2a) | Danio rerio | NM_001003631.1 | |
| shisa-2 | Danio rerio | XM_003201303.5 | |
| shisa-3 | Danio rerio | NM_001080662.2 | |
| shisa-4 | Danio rerio | NM_001017869.1 | |
| shisa-5 | Danio rerio | NM_001044870.1 | |
| shisa -6 | Danio rerio | XM_002667291.5 | |
| shisa-7( shisa like 7) | Danio rerio | XM_021472968.1 | |
| shisa-8 | Danio rerio | XM_021480293.1 | |
| shisa-9a | Danio rerio | NM_001013509.1 | |
| shisa-9b | Danio rerio | NM_001135975.2 | |
| shisa-like 1 | Gallus gallus | XM_015291144.4 | |
| Shisa-2 | Gallus gallus | NM_204501.2 | |
| Shisa-3 | Gallus gallus | XM_040700477.2 | |
| Shisa-4 | Gallus gallus | XM_015298798.4 | |
| Shisa-5 | Gallus gallus | NM_001030591.3 | |
| Shisa-6 | Gallus gallus | XM_025141681.2 | |
| Shisa-8 | Gallus gallus | XM_003640380.6 | |
| Shisa-9 | Gallus gallus | XM_040647667.2 | |
| Shisa-2 | Homo sapiens | NM_001007538.2 | |
| Shisa-3 | Homo sapiens | NM_001080505.3 | |
| Shisa-4 | Homo sapiens | NM_198149.3 | |
| Shisa-5 | Homo sapiens | NM_001272065.3 | |
| Shisa-6 | Homo sapiens | NM_001173461.2 | |
| Shisa-7 | Homo sapiens | NM_001145176.2 | |
| Shisa-8 | Homo sapiens | NM_001207020.3 | |
| Shisa-9 | Homo sapiens | NM_001145204.3 | |
| Shisa-like 1 | Homo sapiens | NM_001099294.2 | |
| Shisa-like 2a | Homo sapiens | NM_001042693.3 | |
| Shisa-like 2b | Homo sapiens | NM_001164442.2 | |
| Shisa-2 | Mus musculus | NM_145463.5 | |
| Shisa-3 | Mus musculus | NM_001033415.3 | |
| Shisa-4 | Mus musculus | NM_175259.5 | |
| Shisa-5 | Mus musculus | NM_001284332.1 | |
| Shisa-6 | Mus musculus | NM_001034874.4 | |
| Shisa-7 | Mus musculus | NM_001290291.1 | |
| Shisa-8 | Mus musculus | NM_001207021.2 | |
| Shisa-9 | Mus musculus | NM_001174086.1 | |
| Shisa-like 1 | Mus musculus | NM_001163145.2 | |
| Shisa-like 2a | Mus musculus | NM_001099303.2 | |
| Shisa-like 2b | Mus musculus | NM_029984.1 | |
| Shisa-1 | Xenopus tropicalis | XM_004915754.4 | |
| Shisa-2 | Xenopus tropicalis | XM_002940554.3 | |
| Shisa-3 | Xenopus tropicalis | XM_002933451.5 | |
| Shisa-4 | Xenopus tropicalis | XM_018091360.2 | |
| Shisa-5 | Xenopus laevis | XM_018261282.2 | |
| Shisa-6 | Xenopus tropicalis | XM_031894479.1 | |
| Shisa-7 | Xenopus tropicalis | XM_002939964.4 | |
| Shisa-8 | Xenopus tropicalis | XM_002934724.5 | |
| Shisa-9 | Xenopus tropicalis | NM_001112925.1 | |
Table 2.
Percentage of phenotype at 48hpf after shisa-2 knockdown.
| Group | Phenotype (%)(n/N) | ||
|---|---|---|---|
| First | Second | Third | |
| Wildtype | 0 (0/59) |
0 (0/80) |
0 (0/102) |
| Cas13d mRNA Control | 0 (0/103) |
8.0 (2/25) |
0 (0/48) |
| gRNAs Control | 0 (0/83) |
4.4 (2/45) |
2.5 (1/40) |
| Cas13d mRNA+gRNAs | 72.9 (70/96) |
69.4 (34/49) |
90.5 (19/21) |
Table 3.
Primers used in the shisa gene cloning.
| Gene Name(NCBI name) | Primer Name | Sequence (5′-3′) | GenBank number |
|---|---|---|---|
| shisa- like 1 | shisa like-1-F | CTGATGGAGGACAAGAAGATG | XM_002667571.6 |
| shisa like-1-R | CTATGGTCAGTCTCAGGCT | ||
| shisa-1(Shisa 2a) | shisa-2a-F | AAGATGAAGTCATCGGCATC | NM_001003631.1 |
| shisa-2a-R | AATAATCCATGTGTAGTCC | ||
| shisa-2 | shisa-2-F | GTGGTTTGTGACACGATG | XM_003201303.5 |
| shisa-2-R | CATTGGGTTTCACATGGT | ||
| shisa-3 | shisa-3-F | AATTCAAGTTTGTCGGCGAG | NM_001080662.2 |
| shisa-3-R | GAGGTCACAGGTCAGCTCTG | ||
| shisa-4 | shisa-4-F | GATGTCCTTCTACGCTGTC | NM_001017869.1 |
| shisa-4-R | GTTATCTTCTCCTCGCAGAG | ||
| shisa-5 | shisa-5-F | GCGAGAGAGCAGCGCTATG | NM_001044870.1 |
| shisa-5-R | AAATGAACCATCCAGCTTGT | ||
| shisa-6 | shisa-6-F | GAAACACACCCTGAAGCCAT | XM_002667291.5 |
| shisa-6-R | TCCAGAGCATCCAAACAGC | ||
| shisa-7( shisa-like-7) | shisa-like-7-F | CATGTAAAGATGATGCCCACC | XM_021472968.1 |
| shisa-like-7-R | CCTCTACCATCCTCCAACTC | ||
| shisa-8b | shisa-8b-F | ATTTCTGGACAGGACCAGAG | XM_021480293.1 |
| shisa-8b-R | TGCATACAGTTATCTGAGTC | ||
| shisa-9a | shisa-9a-F | CCAGGAGACTACAGGATGA | NM_001013509.1 |
| shisa-9a-R | TCCCGCTCTCAGCTGCTTC | ||
| shisa-9b | shisa-9b-F | CCTCAAACATGAGCAGCATC | NM_001135975.2 |
| shisa-9b-R | CCACGTTCACACAGTCACC |
Table 4.
Primers used for construction of Cas13d and synthesis of gRNA.
| Primer Name | Sequence (5′-3′) | Purpose |
|---|---|---|
| Cas13D-F-psp64-T7 | AAGCTTGGGCTGCAGGTCGACTAATACGACTCACTATAGGGAGCCACCATGAGCGAGGCCAGCATCGAAAAAAAAAAG | construction of Cas13d |
| Cas13D-R-psp64-T7 | TGGGAGCTCGCCCGGGGATCCTTAAGCGTAATCTGGAACATCGTATGGGTAAGCGGCCGCTCCGGATCCGGAATTGCCG | construction of Cas13d |
| Cas13d-Universal-F | TAATACGACTCACTATAGGAACCCCTACCAACTGGTCGGGGTTTGAAAC | synthesis of gRNA |
| Cas13D-zfshisa2-gRNA1 | ATCGTCGGCTCAGTTTTTGTGGCGTTTCAAACCCCGACCAGTTGGTAGGGGTT | synthesis of gRNA |
| Cas13D-zfshisa2-gRNA2 | TCGTCGGCTCAGTTTTTGTGGCAGTTTCAAACCCCGACCAGTTGGTAGGGGTT | synthesis of gRNA |
| Cas13D-zfshisa2-gRNA3 | CGTCGGTTCAGTTTTTGTGGCATGTTTCAAACCCCGACCAGTTGGTAGGGGTT | synthesis of gRNA |
| Cas13D-zfshisa2-gRNA4 | TGGGCTCTGTTGTTGCTGTATGCGTTTCAAACCCCGACCAGTTGGTAGGGGTT | synthesis of gRNA |
Table 5.
Primers used for flounder shisa-2 cloning and mRNA synthesis.
| Primer | Sequence (5′-3′) | Purpose |
|---|---|---|
| flounder- shisa-2-F | TGGTCGAGGATGTGGGGCGG | flounder shisa-2 cloning |
| flounder-shisa-2-R | GTGGCAGAGTGGACTACATG | flounder shisa-2 cloning |
| flounder-shisa2psp64-F | AAGCTTGGGCTGCAGGTCGACATGTGGGGCGGAGGTTTCCC | construction of flounder shisa-2 mRNA expression vector |
| flounder-shisa2psp64-R | TGGGAGCTCGCCCGGGGATCCGTGGCAGAGTGGACTACATG | construction of flounder shisa-2 mRNA expression vector |
Table 6.
Primers used for semi-quantitativePCR.
| Gene Name | Primer Name | Sequence (5′-3′) |
|---|---|---|
| shisa- like 1 | shisa-like-1-RT-F | ACTCTCGGACAACAAGACGT |
| shisa-like-1-RT-R | CTATGGTCAGTCTCAGGCT | |
| shisa-1 | shisa-1-RT-F | CGGTGCGATTGTATCTGTCTG |
| shisa-1-RT-R | AATAATCCATGTGTAGTCC | |
| shisa-2 | shisa-2-RT-F | AGTACCCATCTACGTGCCCT |
| shisa-2-RT-R | GAGACTGTAACGGCCGGTAG | |
| shisa-3 | shisa-3-RT-F | CTGGACAGCAGTGGGAATTAC |
| shisa-3-RT-R | TGTGAACATTGACCCATCGT | |
| shisa-4 | shisa-4-RT-F | GATGTCCTTCTACGCTGTC |
| shisa-4-RT-R | TCATCGGATACTGAGGCACC | |
| shisa-5 | shisa-5-RT-F | GCGAGAGAGCAGCGCTATG |
| shisa-5-RT-R | TGGGCTGATATGGTGGGTAC | |
| shisa-6 | shisa-6-RT-F | GAAACACACCCTGAAGCCAT |
| shisa-6-RT-R | AGAGCAGGGTCATACGTGTC | |
| shisa-7 | shisa-7-RT-F | CATGTAAAGATGATGCCCACC |
| shisa-7-RT-R | CAGGTCCCACAGCAGTAGAT | |
| shisa-8 | shisa-8-RT-F | TGCAAACCGGAGCTACTACA |
| shisa-8-RT-R | TGCATACAGTTATCTGAGTC | |
| shisa-9a | shisa-9a-RT-F | CCAGGAGACTACAGGATGA |
| shisa-9a-RT-R | TATCCCAACCAGTGCCATGA | |
| shisa-9b | shisa-9b-RT-F | TCACCCCTATGAGCCGTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



