
Submitted:
30 June 2023
Posted:
05 July 2023
You are already at the latest version
Abstract
Human disturbance, such as trampling, is an integral component of global change, yet we lack a comprehensive understanding of its effects on alpine ecosystems. Many alpine systems are seeing a rapid increase in recreation, and in understudied regions, such as the Coast Mountains of British Columbia, disturbance impacts on alpine plants remain unclear. We surveyed disturbed (trail-side) and undisturbed (off-trail) transects along elevational gradients of popular hiking trails in the T’aḵ’t’aḵ’múy̓in tl’a In̓inyáx̱a7n region (Garibaldi Provincial Park), Canada, focusing on dominant shrubs (Phyllodoce empetriformis, Cassiope mertensiana, Vaccinium ovalifolium) and graminoids (Carex spp). We used a hierarchical Bayesian framework to test for disturbance by elevation effects on total plant percent cover, maximum plant height and diameter (growth proxies), and buds, flowers, and fruits (reproduction proxies). We found that trampling reduces plant cover and impacts all species, but that effects vary by species and trait, and disturbance effects only vary with elevation for one species’ trait. Growth traits are more sensitive to trampling than reproductive traits, which may lead to differential impacts on population persistence and species level fitness outcomes. Our study highlights that disturbance responses are species-specific, and this knowledge can help land managers minimize disturbance impacts on sensitive vegetation types.
Keywords:
Alpine plants
; human trampling disturbance
; global change
; recreation
; plant traits
; trail planning
1. Introduction
Subalpine and alpine plants are sensitive to disturbances from trampling due to their perennial nature, short growing seasons, and shallow soils [1,2]. Species’ responses to trampling vary, with different effects seen in growth and reproduction. These species-specific responses may be related to differences in life history. Grime [3] defined combinations of different life history traits that make certain groups of species more successful along gradients of disturbance, stress, and competition. Ruderals, such as grasses and sedges, are most common in areas with high disturbance, whereas slower growing, competitive species do not tolerate disturbance well. For example, studies have found that Phyllodoce spp. (slow growing subalpine heathers) are vulnerable to trampling disturbances with reduced heights, whereas Carex spp. (sedges) are more resilient [4,5]. In contrast, another study found higher percent cover of Cassiope mertensiana (a slow growing, evergreen subalpine heather) at a trampled site compared to undisturbed sites in the North Cascades of Washington, USA [6].
In addition to reducing plant size, trampling can also reduce reproductive output, interrupt succession, and lower species richness [7,8,9,10,11,12,13,14,15]. Disturbance can also decrease total plant cover, but the severity of effects differ between ecosystems [14,16,17]. Reduced size, reproduction, and survival due to trampling can scale up to negatively affect overall population performance and potentially reduce long term population viability, especially for hiking trails within small, endangered community types [18]. Population performance may be less sensitive to trampling when this disturbance affects reproduction more than survival [19], and thus the plant tissue that is most affected by trampling will likely influence how trampling impacts overall population performance. However, the extent that trampling might differently affect vegetative growth versus reproduction, and whether this effect is uniform across species, is understudied.
One of the main mechanical effects of trampling on plant habitat is that trampling disturbance induces soil compaction. Increased soil compaction has been found to reduce pore connectivity, which in turn decreases its hydro-conductivity and root permeability [20]. Soil compaction also reduces the relative abundance of soil microbes and mycorrhizal fungi [21]. Such conditions can make trampled habitat inhospitable for many plant species. Elevation may also play an important role in how plants and soils respond to disturbance [22]. Climate, including precipitation and temperature, rates of soil development and hydrology, relative plant canopy cover, and numerous other factors vary consistently across elevation gradients, and thus the response of vegetation to the same intensity of disturbance may vary greatly at different elevation zones [22,23,24]. Furthermore, species diversity in a plant community can greatly influence sensitivity to human trampling, with mixed communities of subalpine plant species having three times higher survival than the same species in pure stands [25].
The synergistic effects of disturbance and climate further complicate our understanding of trampling impacts on plant communities. Climatic conditions can have varying effects on alpine communities depending on region, and regional climate changes can negatively impact the rate of plant regeneration post-disturbance [2,26]. In other cases, increased precipitation and warmer summers may even increase the richness of alpine plant species in general, although this response is individualistic and regional [27]. These changes could mediate or exacerbate the effects of trampling in unique ways with contrasting responses to trampling across different climatic zones [28,29]. Taken together, the complex interplays between anthropogenic disturbances and climate change emphasise the need to study these effects in alpine regions experiencing rapid changes in climate and human recreation.
One region experiencing rapid changes in climate and human recreation is British Columbia’s (BC) Coast Mountains. These ocean-proximate mountains are unique at a global scale, as they are among the most southerly glaciated mountains of the northern hemisphere and receive high levels of precipitation year-round [30]. Glacial landscapes and alpine ecosystems, such as those in this region, experience greater warming than other regions in the globe and have seen increasingly rapid glacial retreat in recent years [31]. They are also near Vancouver, Canada, a major population centre of 2.64 million people [32]. This large and growing population has led to a rapidly increasing number of outdoor enthusiasts and new trails are being built into the most accessible mountain terrain. For example, visits to Garibaldi Provincial Park within this region increased 62% (3-year average) between the periods 2006-09 and 2015-18 [33,34,35]. Understanding how these BC ecosystems will respond to warming and increasing trampling due to recreation will help park management determine the least sensitive areas for infrastructure development. However, our understanding of the effects of human trampling disturbance on alpine plant communities in the Coast Mountains of BC is surprisingly limited.
To our knowledge, only two studies have examined the effects of trampling in BC. In a managed subalpine/montane forest stand, Newman and Powell (1997) found negative effects of trampling by livestock (cattle) on lodgepole pine tree plantations primarily in the first 2-3 years of planting in the Kamloops and Merritt Forest Districts. Nepal & Way (2007) found that disturbance reduces plant percent cover in montane to alpine systems, but does not affect species richness in Mount Robson Provincial Park. Studies in nearby alpine ecosystems in the Colorado Rockies have documented immediate and long term impacts of trampling disturbance, including decreasing plant cover, lower reproductive output, soil erosion, and changed plant community structure [2,29,36]. However, we expect these findings to differ from trampling responses in the BC Coast Mountains, where a much deeper winter snowpack may buffer trampling impacts through winter but relatively high precipitation and snowmelt in summer may increase mud, resulting in trail widening by hikers.
Given the lack of information on disturbance impacts in the alpine systems of the BC Coastal Mountains, here we study how human trampling disturbance affects characteristic plant species in this region. Our findings will increase our understanding of how increased human recreation might affect alpine plant communities found across the globe and help to inform future trail designs to minimize disruption locally. We surveyed dominant and common plant communities on- and off- popular hiking trails in the T’aḵ’t’aḵ’múy̓in tl’a In̓inyáx̱a7n (“Landing Place of the Thunderbird”) region (Garibaldi Provincial Park, British Columbia, Canada) in the Squamish and Lil’wat territories. We quantified the impacts of trampling at community (plant percent cover) and species (growth and reproductive traits) levels over elevational gradients to ask:
- How does human trampling disturbance affect alpine plant communities and proxy traits for plant growth and reproduction, and does this effect vary with elevation?
- Which plant species and functional types (evergreen shrubs, deciduous shrubs, sedges) are most sensitive to human trampling disturbance?
We computed plant percent cover from standardized photographs, and measured traits on the dominant species in the heath, blueberry and sedge meadows found frequently throughout this region, encompassing evergreen and deciduous perennial shrubs and graminoids. Our focal species represent common subalpine and alpine plant communities found across arctic and alpine regions worldwide, making our results relevant to other regions around the world. Because of their frequency and broad distribution in the study area, we selected Phyllodoce empetriformis (Sm.) D. Don (Ericaceae) and Cassiope mertensiana (Bong.) G. Don (Ericaceae) from the evergreen heather community, Vaccinium ovalifolium Sm. (Ericaceae) from the deciduous blueberry community, and Carex spp. (Cyperaceae) from the graminoid sedge community (Figure 1). We included a berry species due to their importance to the local Skxwú7mesh (Squamish) and Lil’wat peoples [30,37]. These species provide other important ecosystem services including forage and habitat for wildlife [38], and soil stabilization and high soil carbon storage through fungal mycelium [39]. In addition, the local park authority, BC Parks, aims to better understand how recreation affects common plant communities to help plan for needed infrastructure in areas experiencing an immense increase in recreational traffic, making our study of particular interest from a management perspective.
At a community level, we predict that disturbance will reduce plant percent cover. At a species level, we predict that graminoids (with multiple ramets) will be the least sensitive to disturbance based on their quick recolonization of and high survival in disturbed areas [40]. We further predict that the deciduous (V. ovalifolium) or evergreen (P. empetriformis, C. mertensiana) brittle shrubs will be more sensitive to trampling, with the slow growing evergreen shrubs most affected by multi-year repeated disturbances [40]. Regarding the effects of trampling on plant size, we hypothesized that trampling would break branches and thus decrease plant size. We expected slow growing species (P. empetriformis, C. mertensiana and V. ovalifolium) to be most sensitive to this. With regard to reproduction, we expected that trampling will either i) decrease fruit numbers due to smaller plant biomass, but not change the relative number of reproductive structures per biomass of the plant [41], or ii) increase the density of reproductive structures on trampled plants in response to higher stress [42].
We used Bayesian hierarchical models to test for the effects of disturbance, elevation, and their interaction on plant percent cover and species traits to find human trampling disturbance impacts both at the community and species level, with species-specific responses to disturbance. Disturbance reduced total community plant percent cover, growth proxies for the three out of four species, and reproductive proxies for one out of three species. Unexpectedly, we found that disturbance effects changed with elevation for only one species’ trait and we did not find an effect of elevation on plant cover or other species traits. Surprisingly, Carex spp. was more sensitive to disturbance that we predicted. Our results suggest that species have different sensitivities to human trampling, with V. ovalifolium being the most sensitive in the T’aḵ’t’aḵ’múy̓in tl’a In̓inyáx̱a7n region, and elevation might not be as important as widely understood. Future studies will be needed to determine if the responses to trampling that we observed are species-specific in this region or can be generalized for broader groups (e.g. other graminoids, evergreen shrubs, or deciduous shrubs [40]).
2. Results
In total, we sampled 1772 plants from 280 quadrats along 14 paired trail-off trail transects across a 400 m elevational gradient between 1580 - 1980 m a.s.l.. These plants comprised 500 P. empetriformis individuals, 429 C. mertensiana individuals, 634 V. ovalifolium individuals, and 209 Carex spp. individuals. We checked each model for fit, skew, and dispersion, and models generally fit well with some exceptions. Skew and dispersion were moderately well to poorly modeled depending on the species and trait modeled (Table S1). Some models also had one observation where Pareto K values > 0.7, which slightly decreases the reliability of the Monte Carlo error estimates from Bayesian models.
At the community level, we found that disturbance reduced plant percent cover in quadrats located on trails compared to those off trails (Table 1, Figure 2). At the species level, we found that trampling disturbance reduced size in three out of four species and reproduction in one species, although the intensity of this effect varied by species and some species’ traits were not affected by disturbance. In particular, disturbance reduced both maximum height and maximum diameter in the deciduous shrub (V. ovalifolium), making this the species with highest sensitivity to disturbance. However, disturbance only reduced maximum diameter in one evergreen shrub (P. empetriformis) but had no effect on growth proxies of the other evergreen shrub (C. mertensiana). For the grasses (Carex spp.), disturbance only reduced maximum height and had no effect on diameter (Table 1, Figure 2).
Of the three shrub species for which we quantified relative reproductive output (defined as summed buds, flowers and fruit per plant area, relative to maximum reproductive output for that species), disturbance only affected the reproductive output of the evergreen shrub C. mertensiana (Table 1, Figure 2). Cassiope mertensiana individuals growing on the trail had a significantly lower reproductive output (per plant area) than those growing off the trail. Individuals of P. empetriformis and V. ovalifolium showed no difference in reproductive output whether growing on or off the trail. However, total mean and median reproduction was lower on the trails, and since larger plants produce more reproductive structures (Generalized Linear Mixed Model with random effects of species and transect: estimate = 0.019, standard error = 0.0009, p-value < 0.001), this is likely due to smaller plant sizes on trails. The negative effects of trampling increased with elevation on only Carex spp. height. However, the effects of trampling did not change with elevation for plant percent cover or other species traits. Elevation alone, within the narrow range we studied, did not affect plant percent cover nor any of the species traits that we measured (Table 1, Figure S1).
3. Discussion
This study is one of only a few to document human trampling disturbance in BC’s Coastal Mountains. We focused on the dominant plant species in heather, blueberry, and sedge communities on- and off-trails in a popular subalpine and alpine recreation region, and quantified community plant percent cover and measured proxies of plant growth and reproduction. We provide detailed measurements for nearly 1800 individuals of 4 focal plant species that are of particular regional importance, both culturally, in terms of traditional uses (berries), and ecologically, in terms of their importance within the flora (heather and sedge meadows). Disturbance reduced plant percent cover and decreased proxy traits for growth and reproduction in some species, although the intensity of these effects varied considerably by species. Unexpectedly, disturbance effects increased with elevation for only Carex spp. height, whereas disturbance effects on plant percent cover and species traits stayed constant with elevation. Elevation alone did not have an effect at the community or species level.
Species-Specific Responses
Heath (C. mertensiana, P. empetriformis), blueberry (V. ovalifolium), and sedge (Carex spp.) communities were sensitive to trampling. Human trampling disturbance reduced size in three out of our four study species (P. empetriformis, V. ovalifolium, Carex spp.) and reduced reproductive output in just one (C. mertensiana). Our empirical findings match those for another study in the nearby Washington Cascades [4], which found reduced height among experimentally trampled, multi-species communities dominated by P. empetriformis. Their study further found that Carex spp., a taxa that did not experience trampling effects on diameter in our study, had greater resilience to disturbances, such as trampling [5,17,40]. Another study in this region found that sedge communities are the most resilient to trampling, and heaths the least resilient [38]. The findings of resilience among sedges and other graminoids may be explained by the presence of adaptations, such as multiple ramets and effective root storage, that buffer against tissue damage [40]. We found Carex spp. height, but not diameter, to be sensitive to disturbance impacts, highlighting the need to measure multiple species traits for a better understanding of disturbance effects. While disturbance reduced the diameter of one our heath species (P. empetriformis) and reproductive output in the other heath species (C. mertensiana), blueberry (V. ovalifolium) was most sensitive to disturbance with reduction in both growth proxies (height and diameter).
Other studies have found differences in disturbance impacts between species and traits [40,43]. Variable responses of flower production to trampling across species might be related to both differences in the sensitivity of flowers to direct trampling and plant response to reduced (i.e. trampled) plant size. Reduced growth from trampling might limit the number of resources available for reproduction, an energetically costly process [44]. Meanwhile, species-specific fruit production responses to disturbance could be related to the effect that disturbance might have on pollinators. For example, insects, which might be negatively affected by human trampling [45], could reduce their pollination services for insect pollinated species near trails. While it is unclear how negative effects on growth may scale up to influence population persistence in perennial species, decreases in reproduction are more closely tied to plant fitness. However, some work has shown that population performance may be less sensitive to trampling affecting reproduction than survival [19].
No effect of Elevation on Species Traits
We found no effects of elevation on species traits in our study, and the effects of disturbance only increased with elevation for one species trait. This is surprising, since plant height and reproduction generally decrease with elevation, as higher areas are regionally cooler and less productive [46,47]. The 400 m elevation gradient of our survey may be too small for macroclimatic differences to override microclimatic effects [48]. Moreover, the particular climatic conditions that characterize this maritime-influenced region, particularly the deep, long-lasting snowpack [49], may protect flora against foot traffic in winter. By comparison, in the nearby drier Colorado Rockies there is evidence that vegetation is vulnerable to trampling on windswept and exposed substrates [2]. Snowpack also suppresses tree recruitment, lowering the elevational transition to alpine tundra by several hundred metres in this region relative to less maritime-influenced regions at similar latitudes [49]. As a result, environmental conditions well above treeline are likely more similar to those at lower elevations in many other alpine zones. In addition, the strong maritime climate moderates conditions here year round. Moreover, climate change is altering moisture patterns and producing a thinner snowpack here [49,50] and across western North America [51]. Trampling may thus interact with elevation in the future, amplifying the need for further study.
Future Research
Although we did not explicitly investigate species richness, other studies have found that decreased species richness occurred alongside growth and reproductive effects of trampling (e.g., [28]), including in heavily experimentally trampled P. empetriformis dominated communities (e.g., [4]). Our results are in line with previous work that found that disturbance reduces species richness and diversity in an alpine system [28]. Community level impacts from trampling are thus likely, but further study will be needed to evaluate these in the present system and we are conducting ongoing surveys at our transects to quantify the effect of disturbance on species richness and diversity. Higher levels of species richness have also been associated with higher resilience of subalpine plant communities to trampling [25], making this an important consideration of land management to promote highly mixed rather than single species dominated vegetation alongside trails.
Another major impact of human disturbance, including trampling, is promoting the survival and spread of invasive species in plant communities by unintentional introduction of seeds carried on clothes and shoes, as well as intentional introduction of non-natives for ornamental or other purposes [52,53]. Exacerbating this issue is that non-native species can outperform native species in trampled habitat [52,54,55,56]. However, there is limited data to corroborate such findings for alpine ecosystems. The few studies that have examined disturbance and invasibility find cold-climate ecosystems to be more prone to invasion [57]. While we did not investigate this issue, ongoing studies at our permanent transects examine both changes to species diversity and species invasions.
Implications for Trail Planning
With interest in outdoor recreation in BC rapidly increasing [34,35], there is a need for more trails to spread out disturbance impacts while still providing public access to outdoor spaces. Our findings suggest that some species are more affected by trampling than others and specific management approaches may need to be tailored toward particular species. These varying species-specific impacts should be considered in the planning of future trail and recreation infrastructure (i.e. avoid trails through regions with species that respond strongly to trampling disturbances and/or exist in very localized areas). We suggest that trails through graminoid and heath dominated meadows may be able to recover more quickly from regular disturbance than trails through deciduous shrubs based on our findings and those of previous studies [40]. However, it should be considered that sedge meadows often have significantly more moist soils and this can result in trail widening (when hikers avoid getting muddy, wet feet) affecting a much larger area and number of individuals (pers. obs.). Based on our results that blueberry (V. ovalifolium) was most affected by trampling out of the four species we measured, trails should avoid going through these berry ecosystems since it may decrease berry yields for cultural harvesting and for wildlife. We emphasize that our findings only reflect the four species along the elevational gradient that we studied, and future studies are needed on additional species and in other regions to make our findings generalizable.
Although we studied common species found in well established, large subalpine meadows, alpine ecosystems are also filled with less common, sometimes endangered, species found in small microhabitats [18]. Based on our findings that individuals on trails were smaller than those off trails, and sometimes resulting in less biomass for reproduction, we suggest that trails should avoid going through microhabitats hosting rare or endangered species for whom a decrease in local reproduction could have population level consequences. Trail shape and infrastructure (e.g., trail markers, boardwalks, etc.) likely also affect community level sensitivity to trampling. Small and rare communities are likely much more affected by trails that repeatedly go through them, especially if trampling disturbance impacts decrease in a gradient from the trail edge (e.g., switchbacks). Future studies should investigate how far from trail edges plant size and reproduction are affected. If effects are always directly next to trails, then small, rare plant communities should be avoided for any trail development.
Conclusions
Given that we monitored these plant communities and their individuals for only a single growing season, it is notable that we find such reduced vegetative growth and reproduction across species, even if this effect is variable. This suggests a need for long-term studies that include more taxa, and explore invasive species as well as species diversity. Human recreation in alpine regions will likely continue to grow, increasing the risk of plant trampling by hikers. Furthermore, continued climate change may further exacerbate species-specific plant vulnerability to trampling. For example, trampled trailing edge populations might be more prone to displacement via colonization from lower elevation species expanding their ranges [58]. Given the decreases in size, reproductive capacity, and plant cover observed in our study, it will be important to evaluate possible interactions between trampling and climate change in other systems where displacement by colonization of lower elevation species is expected.
4. Materials and Methods
Study Site
We conducted this study in the T’aḵ’t’aḵ’múy̓in tl’a In̓inyáx̱a7n (“Landing Place of the Thunderbird”) region (Taylor Meadows, Garibaldi Provincial Park, British Columbia, Canada) in the Squamish and Lil’wat territories (Figure 3a). The area is classified as Coast Mountain Heather alpine zone within the Biogeoclimatic Ecosystem Classification (BEC) system of BC, where a deep long-lasting winter snowpack limits tree recruitment, and the transition to alpine tundra begins at lower elevations than for other alpine zones at comparable latitudes towards the interior [49]. Vegetation is dominated by dwarf shrubs, including Cassiope spp. and Phyllodoce spp. (mountain heathers), Luetkea pectinata (partridgefoot), and herbaceous species of sedge, grass and forb. There are patches of stunted trees or “krummholz” at treeline, comprised mainly of Abies lasiocarpa (Subalpine fir), Picea engelmannii (Engelmann spruce), and Tsuga mertensiana (Mountain hemlock). Mean annual temperatures for the BC alpine have historically been between -4 and 0°C, with a short growing season and mean annual precipitation of 700-3000 mm, much of which falls as snow [59].
Approximately 100 km from the city of Vancouver, the study area is one of the most popular hiking destinations in southwestern British Columbia [60]. Garibaldi Park attendance has increased substantially, especially since the highway linking Vancouver with the park was upgraded for the 2010 Winter Olympics [33,34,35]. Popular trailheads (Diamond Head, Rubble Creek, Wedgemont Lake) are filled with hikers and backpackers in the summer season. In the last few years, new trail developments have been proposed (e.g. the Darling Lake trail and the hut developments around the Spearhead Range).
T’aḵ’t’aḵ’múy̓in tl’a In̓inyáx̱a7n (“Landing Place of the Thunderbird”) region (Garibaldi Provincial Park, British Columbia, Canada) in the Squamish and Lil’wat territories. We established transects (not all shown) along the popular Taylor Meadows, Black Tusk, and Panorama Ridge trails. (b) Along each trail (indicated in brown), we established transects (not all shown) at the edge of the trail (orange = disturbed) and 5 m perpendicular to the disturbed transect (grey = undisturbed) below and above the treeline. We recorded plant data in 100 cm by 50 cm quadrats (photo inserts) along each transect. (c) At each quadrat, we randomly selected five individuals of each of our focal species to measure maximum plant height and maximum plant diameter (indicated along picture, not to scale), and counted the number of buds, flowers, and fruits (white circles). We did not count reproductive metrics for Carex spp. as it was not possible to accurately distinguish between phenophases. (d) At each quadrat, we also took a standardized photograph from which to compute plant percent cover.
Data Collection
To answer how hiker trampling disturbance affects common subalpine and alpine plant communities (blueberry, heather and sedge meadows) along elevational gradients, we chose sites at multiple elevations (1580 - 1980 m a.s.l.) along three different trails (Figure 3b). We set up 14 permanent 10 m x 0.5 m paired trail-side (disturbed) and off-trail (undisturbed) transects in August 2022 along the heavily used Taylor Meadows-Helm Creek, Black Tusk, and Panorama Ridge trails. We established transects directly adjacent to the trail’s edge and at least a 5 m perpendicular distance away from the trail to compare trampled and untrampled plant communities, respectively (following [28,29]). To mark the trail’s edge, we identified sections of the trails disturbed predominantly by human trampling, but not by water run-off or trail cut construction. We recorded latitude and longitude in the field with the Gaia mobile application v2022 (WGS 84, ESPG: 4326) to obtain elevation per site (see below).
We recorded plant community data in ten 1 m x 0.5 m quadrats within each transect. In each quadrat, we randomly selected five individuals per focal species (Phyllodoce empetriformis, Cassiope mertensiana, Vaccinium ovalifolium, Carex spp.) and recorded maximum plant height (height perpendicular to the ground without stretching the plant) and maximum diameter of the plant within the quadrat (Figure 3c, Table 2). To quantify reproductive output of our focal shrub species, we counted the number of buds, flowers and fruits on each of the five plants randomly selected. We did not count reproductive metrics for Carex spp. as it was not possible to accurately distinguish between phenophases. For image analyses of plant percent cover, we took a standardized photograph 1.5 meters directly above each quadrat (Figure 3d).
Data Processing
We conducted all data processing and analyses in R version 4.2.3 [61]. We used field-recorded latitude and longitude data to obtain elevation per site using the USGS NED1 digital elevation model at 30 m resolution [62]. We used the ‘tidyverse’ [63] and ‘dplyr’ [64] packages to check for unusual patterns or outliers in our data. For an area-standardized reproductive output per individual plant, we first calculated a proxy for plant area by multiplying maximum plant height by maximum plant diameter. We then summed bud, flower, and fruit counts and divided this by plant area for an area-standardized summed reproductive output. Finally, we calculated relative reproduction for each individual by dividing summed reproductive output by that species’ maximum reproductive output (i.e. individual with highest reproductive output). While other studies have shown that reproductive output density increases with plant area [65,66], this is not the case in our study in either disturbed or undisturbed transects (tested with linear mixed model with package ‘lmerTest’; [67]; Figure S2).
To standardize our quadrat photos, we manually flattened and cropped all images to just the quadrat border using Google Drive’s photo editing software. To account for any shading in the plot, we increased image saturation to the point where changes in lighting negligibly affected color intensity. Logs and rocks found in quadrats (25.45% of photos) were counted as non-green coverage and moss was counted as green cover, although very little moss (8.24% of photos) was seen in the photos. We then processed the cropped images with a custom algorithm in Python 3.10 [68], which first boosts saturation by a factor of 30 to emphasize green vegetation. The fraction of green pixels out of the entire image is then calculated as an estimate of percent plant coverage in the quadrat.
Data Analyses
To test how trail disturbance, elevation, and their interaction affect plant percent cover and species traits, we fit Bayesian generalized non-linear mixed models. We fit separate models for plant percent cover and each species by trait to account for potential response differences. These models allow for non-linear relationships and account for non-independence of data within a transect pair (i.e. on- and off-trail transect pair) by using a random effect of transect pairs. Our model equation is response ~ disturbance * elevation + (1|transect pair), where ‘response’ is plant percent cover, maximum plant height, or maximum plant diameter.
To account for left-skew in the diameter and height trait data, we fit negative binomial models (link = ‘log’, link_shape = ‘log’) for these response variables and rounded measured diameter and height values to the nearest integer value for this distribution family. The Beta model outperformed the Zero-inflated Beta model (tested on P. empetriformis) as checked with leave-one-out cross validation (‘loo()’), so to best model our left-skewed proportional relative reproduction data, we fit Beta models (link = ‘log’, link_phi = ‘log’). We also modeled plant percent cover with the Beta distribution. We adjusted 0, 1 values by 0.0001 to keep our data within the Beta distribution interval [0,1]. All models and family distributions are described in Table S1.
We fit models with the ‘brms’ [69,70,71], ‘rjags’ [72], and ‘R2jags’ [73] packages. We fit models using 3 chains, 5000 iterations and 1000 warmup iterations on 4 cores on macOS Version 12.6.5 with a 2.2 GHz Quad-Core Intel Core i7 processor. We set ‘init = 0’ for plant percent cover and reproduction models. We inferred any parameter estimate’s credible interval that did not contain zero as evidence for an effect of the parameter. For each model, we assessed model fit by checking that i) Rhat > 1.1 and that Estimated Sample Size (Bulk_ESS and Tail_ESS) > 1000 (‘summary()’), and ii) all MCMC chains converged (‘plot()’). We assessed the prior distribution of each model by checking that priors do not overwhelm likelihood (‘powerscale_sensitivity()’). We assessed the posterior distribution by checking that i) predicted values are similar to posterior distribution with 1000 posterior draws (‘pp_check(ndraws = 1000)’), ii) scatterplot matrices of posterior parameter distributions are normally distributed (‘pairs()’), iii) skew is properly modeled using the Fisher-Pearson Skew function (‘ppc_stat()’), iv) Pareto k values < 0.5 (‘loo()’), and v) dispersion is properly modeled (‘ppc_loo_pit_overlay()’). The indicated functions are from the ‘bayesplot’ [74,75], ‘loo’ [76,77], and ‘priorsense’ [78] packages. See Table S1 for additional information on model fit.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Model fitting details; S1: Effects of disturbance and elevation; Figure S2: Reproductive structure density.
Author Contributions
Conceptualization, NIC, CE, CGC; methodology, NIC, CE, CGC; software, NIC, PS, AZ; formal analysis, NIC, CGC; investigation, NIC, CH, TM, BR; data curation, CH, NIC, PS; writing—original draft preparation, NIC, PS, CH, TM, BR, AZ, KG, CGC, NH, CE; writing—review and editing, NIC, PS, CH, TM, BR, AZ, KG, CGC, NH, CE; visualization, PS, BR, CH, NIC, CGC; supervision, NIC, CE, CGC, NH; project administration, NIC, CE, CGC; funding acquisition, CE, CGC, NIC. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by BC Parks Living Lab for Climate Change and Conservation, University of British Columbia’s Work Learn program and Biology Undergraduate Diversity in Research program, and Natural Sciences and Engineering Council of Canada Discovery Grant (to G. Henry). NIC was funded by a Swiss National Science Foundation Postdoc Mobility Fellowship (Grant ID: 194331). GCG was funded by a Biodiversity Research Centre Postdoctoral fellowship. The APC was funded by the Swiss National Science Foundation.
Data Availability Statement
The data supporting reported results and R code for analyses can be found on GitHub at https://github.com/ITEX-sites/Garibaldi.
Acknowledgments
We are guests on the traditional, ancestral, and unceded territory of the xwməθkwəy̓əm (Musqueam), Skwxwú7mesh (Squamish), Stó:lō, and Səl̓ílwətaʔ/Selilwitulh (Tsleil Waututh) and Líl̓wat Nations. As such we are communicating and learning from these Nations as much as possible during our studies. This research was supported in part through computational resources and services provided by Advanced Research Computing at the University of British Columbia. We thank the International Tundra Experiment, and Blackcomb Helicopters for logistical support. Fieldwork was completed by Nathalie Isabelle Chardon, Mackenzie Urquhart-Cronish, Christian Lauber, Carolyn Chong, Teagan Maclachlan, Carly Hilbert, Brianna Ragsdale, and Vicky Lee.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Billings, W.D. Arctic and Alpine Vegetations: Similarities, Differences, and Susceptibility to Disturbance. BioScience 1973, 23, 697–704. [Google Scholar] [CrossRef]
- Willard, B.E.; Cooper, D.J.; Forbes, B.C. Natural Regeneration of Alpine Tundra Vegetation after Human Trampling: A 42-Year Data Set from Rocky Mountain National Park, Colorado, U.S.A. Arct. Antarct. Alp. Res. 2007, 39, 177–183. [Google Scholar] [CrossRef]
- Grime, J.P. Vegetation Classification by Reference to Strategies. Nature 1974, 250, 26–31. [Google Scholar] [CrossRef]
- Cole, D.N.; Intermountain Research Station (Ogden, U. Trampling Effects on Mountain Vegetation in Washington, Colorado, New Hampshire, and North Carolina; U.S. Dept. of Agriculture, Forest Service, Intermountain Research Station: Ogden, UT, 1993. [Google Scholar]
- Crisfield, V.E.; Macdonald, S.E.; Gould, A.J. Effects of Recreational Traffic on Alpine Plant Communities in the Northern Canadian Rockies. Arct. Antarct. Alp. Res. 2012, 44, 277–287. [Google Scholar] [CrossRef]
- de Gouvenain, R.C. Indirect Impacts of Soil Trampling on Tree Growth and Plant Succession in the North Cascade Mountains of Washington. Biol. Conserv. 1996, 75, 279–287. [Google Scholar] [CrossRef]
- Pickering, C.M.; Hill, W.; Newsome, D.; Leung, Y.-F. Comparing Hiking, Mountain Biking and Horse Riding Impacts on Vegetation and Soils in Australia and the United States of America. J. Environ. Manage. 2010, 91, 551–562. [Google Scholar] [CrossRef]
- Li, W.; He, S.; Cheng, X.; Zhang, M. Functional Diversity Outperforms Taxonomic Diversity in Revealing Short-Term Trampling Effects. Sci. Rep. 2021, 11, 18889. [Google Scholar] [CrossRef]
- Barros, A.; Pickering, C.M. Impacts of Experimental Trampling by Hikers and Pack Animals on a High-Altitude Alpine Sedge Meadow in the Andes. Plant Ecol. Divers. 2015, 8, 265–276. [Google Scholar] [CrossRef]
- Bisht, V.K.; Kuniyal, C.P.; Nautiyal, B.P.; Prasad, P. Integrated Analysis of the Trees and Associated Under-Canopy Species in a Subalpine Forest of Western Himalaya, Uttarakhand, India. J. Mt. Sci. 2015, 12, 154–165. [Google Scholar] [CrossRef]
- Pickering, C.M.; Hill, W. Impacts of Recreation and Tourism on Plant Biodiversity and Vegetation in Protected Areas in Australia. J. Environ. Manage. 2007, 85, 791–800. [Google Scholar] [CrossRef]
- Rossi, G.; Parolo, G.; Ulian, T. Human Trampling as a Threat Factor for the Conservation of Peripheral Plant Populations. Plant Biosyst. - Int. J. Deal. Asp. Plant Biol. 2009, 143, 104–113. [Google Scholar] [CrossRef]
- Ballantyne, M.; Pickering, C.M.; McDougall, K.L.; Wright, G.T. Sustained Impacts of a Hiking Trail on Changing Windswept Feldmark Vegetation in the Australian Alps. Aust. J. Bot. 2014, 62, 263. [Google Scholar] [CrossRef]
- Barros, A.; Gonnet, J.; Pickering, C. Impacts of Informal Trails on Vegetation and Soils in the Highest Protected Area in the Southern Hemisphere. J. Environ. Manage. 2013, 127, 50–60. [Google Scholar] [CrossRef]
- Jägerbrand, A.K.; Alatalo, J.M. Effects of Human Trampling on Abundance and Diversity of Vascular Plants, Bryophytes and Lichens in Alpine Heath Vegetation, Northern Sweden. SpringerPlus 2015, 4, 95. [Google Scholar] [CrossRef]
- Yang, M.; Hens, L.; Ou, X.; Wulf, R.D. Impacts of Recreational Trampling on Sub-Alpine Vegetation and Soils in Northwest Yunnan, China. Acta Ecol. Sin. 2009, 29, 171–175. [Google Scholar] [CrossRef]
- Cole, D.N. Experimental Trampling of Vegetation. I. Relationship Between Trampling Intensity and Vegetation Response. J. Appl. Ecol. 1995, 32, 203–214. [Google Scholar] [CrossRef]
- Maschinski, J.; Frye, R.; Rutman, S. Demography and Population Viability of an Endangered Plant Species before and after Protection from Trampling. Demografia y Viabilidad Poblacional de Un Especie de Planta En Peligro de Extincion Antes y Despues de Protegerla Contra Pisoteo. Conserv. Biol. 2003, 11, 990–999. [Google Scholar] [CrossRef]
- Pfister, C.A. Patterns of Variance in Stage-Structured Populations: Evolutionary Predictions and Ecological Implications. Proc. Natl. Acad. Sci. 1998, 95, 213–218. [Google Scholar] [CrossRef]
- Quantification of the Effects of Soil Compaction on Water Flow Using Dye Tracers and Image Analysis - Mooney - 2003 - Soil Use and Management - Wiley Online Library Available online:. Available online: https://bsssjournals.onlinelibrary.wiley.com/doi/abs/10.1111/j.1475-2743.2003.tb00326.x (accessed on 10 June 2023).
- Liu, Q.; Li, W.; Nie, H.; Sun, X.; Dong, L.; Xiang, L.; Zhang, J.; Liu, X. The Effect of Human Trampling Activity on a Soil Microbial Community at the Urban Forest Park. Forests 2023, 14, 692. [Google Scholar] [CrossRef]
- Kuss, F.R. A Review of Major Factors Influencing Plant Responses to Recreation Impacts. Environ. Manage. 1986, 10, 637–650. [Google Scholar] [CrossRef]
- Pearcy, R.W.; Ward, R.T. Phenology and Growth of Rocky Mountain Populations of Deschampsia Caespitosa at Three Elevations in Colorado. Ecology 1972, 53, 1171–1178. [Google Scholar] [CrossRef]
- ECOLOGICAL IMPACT OF CAMPING UPON THE SOUTHERN SIERRA NEVADA - ProQuest. Available online: https://www.proquest.com/openview/8bdb0fa22b4747cbafa2b8f68647dfef/1.pdf?pq-origsite=gscholar&cbl=18750&diss=y (accessed on 29 June 2023).
- Holmes, D.O.; Dobson, H.E.M. The Effects of Human Trampling and Urine on Subalpine Vegetation, a Survey of Past and Present Backcountry Use, and the Ecological Carrying Capacity of Wilderness: Final Report, Ecological Carrying Capacity Research Contract CX8000-4-0026. 1976. [Google Scholar]
- Kycko, M.; Zagajewski, B.; Lavender, S.; Romanowska, E.; Zwijacz-Kozica, M. The Impact of Tourist Traffic on the Condition and Cell Structures of Alpine Swards. Remote Sens. 2018, 10, 220. [Google Scholar] [CrossRef]
- Moen, J.; Lagerström, A. High Species Turnover and Decreasing Plant Species Richness on Mountain Summits in Sweden: Reindeer Grazing Overrides Climate Change. Arct. Antarct. Alp. Res. 2008, 40, 382–395. [Google Scholar] [CrossRef]
- Chardon, N.I.; Wipf, S.; Rixen, C.; Beilstein, A.; Doak, D.F. Local Trampling Disturbance Effects on Alpine Plant Populations and Communities: Negative Implications for Climate Change Vulnerability. Ecol. Evol. 2018, 8, 7921–7935. [Google Scholar] [CrossRef]
- Chardon, N.I.; Rixen, C.; Wipf, S.; Doak, D.F. Human Trampling Disturbance Exerts Different Ecological Effects at Contrasting Elevational Range Limits. J. Appl. Ecol. 2019, 56, 1389–1399. [Google Scholar] [CrossRef]
- Evans, I.S. Climatic Effects on Glacier Distribution Across the Southern Coast Mountains. B.C., Canada. Ann. Glaciol. 1990, 14, 58–64. [Google Scholar] [CrossRef]
- Bevington, A.R.; Menounos, B. Accelerated Change in the Glaciated Environments of Western Canada Revealed through Trend Analysis of Optical Satellite Imagery. Remote Sens. Environ. 2022, 270, 112862. [Google Scholar] [CrossRef]
- Statistics Canada Census of Population 2021. 2023.
- BC Parks. 2008/09 BC Parks Year End Report; Ministry of Environment: Victoria, 2009; p. 12. [Google Scholar]
- BC Parks. BC Parks 2015/16 Statistics Report; Ministry of Environment: Victoria, BC, 2016; p. 43. [Google Scholar]
- BC Parks. BC Parks 2017/18 Statistics Report; Ministry of Environment: Victoria, BC, 2018; p. 44. [Google Scholar]
- Curtin, C.G. Can Montane Landscapes Recover from Human Disturbance? Long-Term Evidence from Disturbed Subalpine Communities. Biol. Conserv. 1995, 74, 49–55. [Google Scholar] [CrossRef]
- Ross, E. Squamish Nation Culturally Significant Vegetation n.d.
- Martin, K. D.H. Johnson and T. A. O’Neil (Managing Directors). Wildlife-Habitat Relationships in Oregon and Washington; Oregon State University Press: Corvallis, 2001; Volume 2001, p. 2001. [Google Scholar]
- Vowles, T.; Björk, R.G. Implications of Evergreen Shrub Expansion in the Arctic. J. Ecol. 2019, 107, 650–655. [Google Scholar] [CrossRef]
- Cole, D.N. Experimental Trampling of Vegetation. II. Predictors of Resistance and Resilience. J. Appl. Ecol. 1995, 32, 215–224. [Google Scholar] [CrossRef]
- Fournier, M.-P.; Paré, M.C.; Buttò, V.; Delagrange, S.; Lafond, J.; Deslauriers, A. How Plant Allometry Influences Bud Phenology and Fruit Yield in Two Vaccinium Species. Ann. Bot. 2020, 126, 825–835. [Google Scholar] [CrossRef] [PubMed]
- Wada, K.C.; Takeno, K. Stress-Induced Flowering. Plant Signal. Behav. 2010, 5, 944–947. [Google Scholar] [CrossRef] [PubMed]
- Evju, M.; Hagen, D.; Hofgaard, A. Effects of Disturbance on Plant Regrowth along Snow Pack Gradients in Alpine Habitats. Plant Ecol. 2012, 213, 1345–1355. [Google Scholar] [CrossRef]
- Lee, T.D. Patterns of Fruit and Seed Production. In Plant reproductive ecology; Oxford University Press: New York, U.S.A, 1988; pp. 179–202. [Google Scholar]
- Ciach, M.; Maślanka, B.; Krzus, A.; Wojas, T. Watch Your Step: Insect Mortality on Hiking Trails. Insect Conserv. Divers. 2017, 10, 129–140. [Google Scholar] [CrossRef]
- Macek, P.; Lepš, J. Environmental Correlates of Growth Traits of the Stoloniferous Plant Potentilla Palustris. Evol. Ecol. 2008, 22, 419–435. [Google Scholar] [CrossRef]
- Mao, L.; Chen, S.; Zhang, J.; Zhou, G. Altitudinal Patterns of Maximum Plant Height on the Tibetan Plateau. J. Plant Ecol. 2018, 11, 85–91. [Google Scholar] [CrossRef]
- Ford, K.R.; Ettinger, A.K.; Lundquist, J.D.; Raleigh, M.S.; Hille Ris Lambers, J. Spatial Heterogeneity in Ecologically Important Climate Variables at Coarse and Fine Scales in a High-Snow Mountain Landscape. PLoS ONE 2013, 8, e65008. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, W. The Ecology of the Alpine Zones; BC Ministry of Forests and Range Research: Victoria, BC, 2006. [Google Scholar]
- Mote, P.W.; Hamlet, A.F.; Clark, M.P.; Lettenmaier, D.P. DECLINING MOUNTAIN SNOWPACK IN WESTERN NORTH AMERICA*. Bull. Am. Meteorol. Soc. 2005, 86, 39–50. [Google Scholar] [CrossRef]
- Lute, A.C.; Abatzoglou, J.T.; Hegewisch, K.C. Projected Changes in Snowfall Extremes and Interannual Variability of Snowfall in the Western United States. Water Resour. Res. 2015, 51, 960–972. [Google Scholar] [CrossRef]
- Barros, A.; Aschero, V.; Mazzolari, A.; Cavieres, L.A.; Pickering, C.M. Going off Trails: How Dispersed Visitor Use Affects Alpine Vegetation. J. Environ. Manage. 2020, 267, 110546. [Google Scholar] [CrossRef]
- Hulme, P.E.; Pyšek, P.; Pergl, J.; Jarošík, V.; Schaffner, U.; Vilà, M. Greater Focus Needed on Alien Plant Impacts in Protected Areas. Conserv. Lett. 2014, 7, 459–466. [Google Scholar] [CrossRef]
- Luong, J.C.; Villanueva, E.M.; Bauman, T.A. Native and Invasive Bunchgrasses Have Different Responses to Trail Disturbance on California Coastal Prairies. 2022. [Google Scholar]
- Urbina, J.C.; Benavides, J.C. Simulated Small Scale Disturbances Increase Decomposition Rates and Facilitates Invasive Species Encroachment in a High Elevation Tropical Andean Peatland. Biotropica 2015, 47, 143–151. [Google Scholar] [CrossRef]
- Bella, E.M. Invasion Prediction on Alaska Trails: Distribution, Habitat, and Trail Use. Invasive Plant Sci. Manag. 2011, 4, 296–305. [Google Scholar] [CrossRef]
- Lembrechts, J.J.; Milbau, A.; Nijs, I. Alien Roadside Species More Easily Invade Alpine than Lowland Plant Communities in a Subarctic Mountain Ecosystem. PLoS ONE 2014, 9, e89664–e89664. [Google Scholar] [CrossRef]
- Parmesan, C. Ecological and Evolutionary Responses to Recent Climate Change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef]
- Alpine Tundra Zone. Ecosystems of British Columbia; Meidinger, D.V., Pojar, J., Eds.; Special report series; Research Branch, Ministry of Forests: Victoria, B.C, 1991; pp. 264–274. ISBN 978-0-7718-8997-4. [Google Scholar]
- Bryceland, J.; Macaree, D.; Macaree, M. 103 Hikes in Southwestern British Columbia; Greystone Books: Canada, 2008; ISBN 978-1-926685-02-1. [Google Scholar]
- R Core Team. 2023.
- GPS Visualizer. Available online: https://www.gpsvisualizer.com/ (accessed on 11 November 2023).
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Wickham, H.; François, R.; Henry, L.; Müller, K.; Vaughan, D. Dplyr: A Grammar of Data Manipulation. 2023. [Google Scholar]
- Bonser, S.P. Size and Phenology Control Plant Reproduction and Agricultural Production. A Commentary on: ‘How Plant Allometry Influences Bud Phenology and Fruit Yield in Two Vaccinium Species.’. Ann. Bot. 2020, 126, vi–vii. [Google Scholar] [CrossRef]
- Weiner, J.; Rosenmeier, L.; Massoni, E.S.; Vera, J.N.; Plaza, E.H.; Sebastià, M.-T. Is Reproductive Allocation in Senecio Vulgaris Plastic? Botany 2009, 87, 475–481. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. LmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- Python Software Foundation. 2021.
- Bürkner, P.-C. Brms: An R Package for Bayesian Multilevel Models Using Stan. J. Stat. Softw. 2017, 80, 1–28. [Google Scholar] [CrossRef]
- Bürkner, P.-C. Advanced Bayesian Multilevel Modeling with the R Package Brms. R J. 2018, 10, 395–411. [Google Scholar] [CrossRef]
- Bürkner, P.-C. Bayesian Item Response Modeling in R with Brms and Stan. J. Stat. Softw. 2021, 100, 1–54. [Google Scholar] [CrossRef]
- Plummer, M.; Stukalov, A.; Denwood, M. Rjags: Bayesian Graphical Models Using MCMC. 2023. [Google Scholar]
- Su, Y.-S.; Yajima, M. R2jags: Using R to Run “JAGS”. 2021. [Google Scholar]
- Gabry, J.; Simpson, D.; Vehtari, A.; Betancourt, M.; Gelman, A. Visualization in Bayesian Workflow. J. R. Stat. Soc. Ser. A Stat. Soc. 2019, 182, 389–402. [Google Scholar] [CrossRef]
- Gabry, J.; Mahr, T. Bayesplot: Plotting for Bayesian Models. 2022. [Google Scholar]
- Vehtari, A.; Gelman, A.; Gabry, J. Practical Bayesian Model Evaluation Using Leave-One-out Cross-Validation and WAIC. Stat. Comput. 2017, 27, 1413–1432. [Google Scholar] [CrossRef]
- Vehtari, A.; Gabry, J.; Magnusson, M.; Bürkner, P.-C.; Paananen, T.; Gelman, A. Loo: Efficient-Leave-One-out Cross-Validation and WAIC for Bayesian Models. 2022. [Google Scholar]
- Kallioinen, N.; Bürkner, P.-C.; Paananen, T.; Vehtari, A. Priorsense: Prior Diagnostics and Sensitivity Analysis. 2023. [Google Scholar]
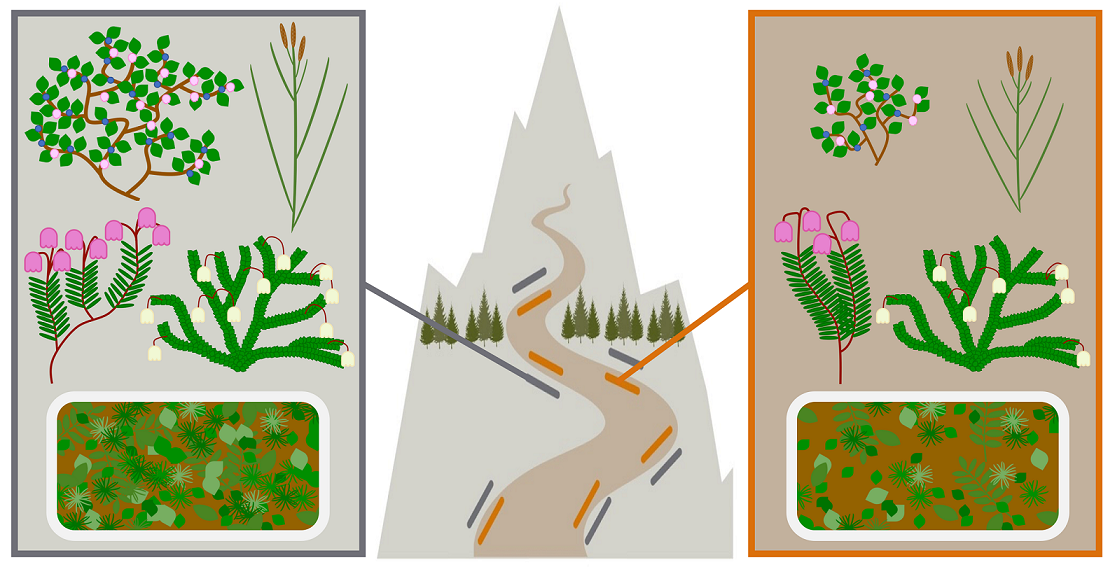
Figure 1.
Study species. The four focal species in our studies are common and dominant species of the heather, blueberry, and sedge communities in the T’aḵ’t’aḵ’múy̓in tl’a In̓inyáx̱a7n region (Garibaldi Provincial Park, British Columbia, Canada). They are of cultural importance to the Squamish and Lil’wat Nations and of management concern to the park authority, BC Parks.
Figure 1.
Study species. The four focal species in our studies are common and dominant species of the heather, blueberry, and sedge communities in the T’aḵ’t’aḵ’múy̓in tl’a In̓inyáx̱a7n region (Garibaldi Provincial Park, British Columbia, Canada). They are of cultural importance to the Squamish and Lil’wat Nations and of management concern to the park authority, BC Parks.

Figure 2.
Effects of disturbance on plant traits and plant percent cover. Responses to human trampling disturbance are species-specific, with variable effects on maximum plant height, maximum plant diameter, and relative reproductive output (summed buds, flowers, fruits by total plant area, relative to the maximum per species). Shown are back-transformed parameter estimates (circles) of disturbance with their credible intervals (bars) from Bayesian hierarchical non-linear mixed models. Parameter estimates of disturbance and elevation are shown in Figure S1. Asterisks (*) indicate parameter estimates whose credible intervals do not encompass 0 and can be interpreted as having an effect. Legend for all plots is as in (a). Note that y-axes are on different scales.
Figure 2.
Effects of disturbance on plant traits and plant percent cover. Responses to human trampling disturbance are species-specific, with variable effects on maximum plant height, maximum plant diameter, and relative reproductive output (summed buds, flowers, fruits by total plant area, relative to the maximum per species). Shown are back-transformed parameter estimates (circles) of disturbance with their credible intervals (bars) from Bayesian hierarchical non-linear mixed models. Parameter estimates of disturbance and elevation are shown in Figure S1. Asterisks (*) indicate parameter estimates whose credible intervals do not encompass 0 and can be interpreted as having an effect. Legend for all plots is as in (a). Note that y-axes are on different scales.
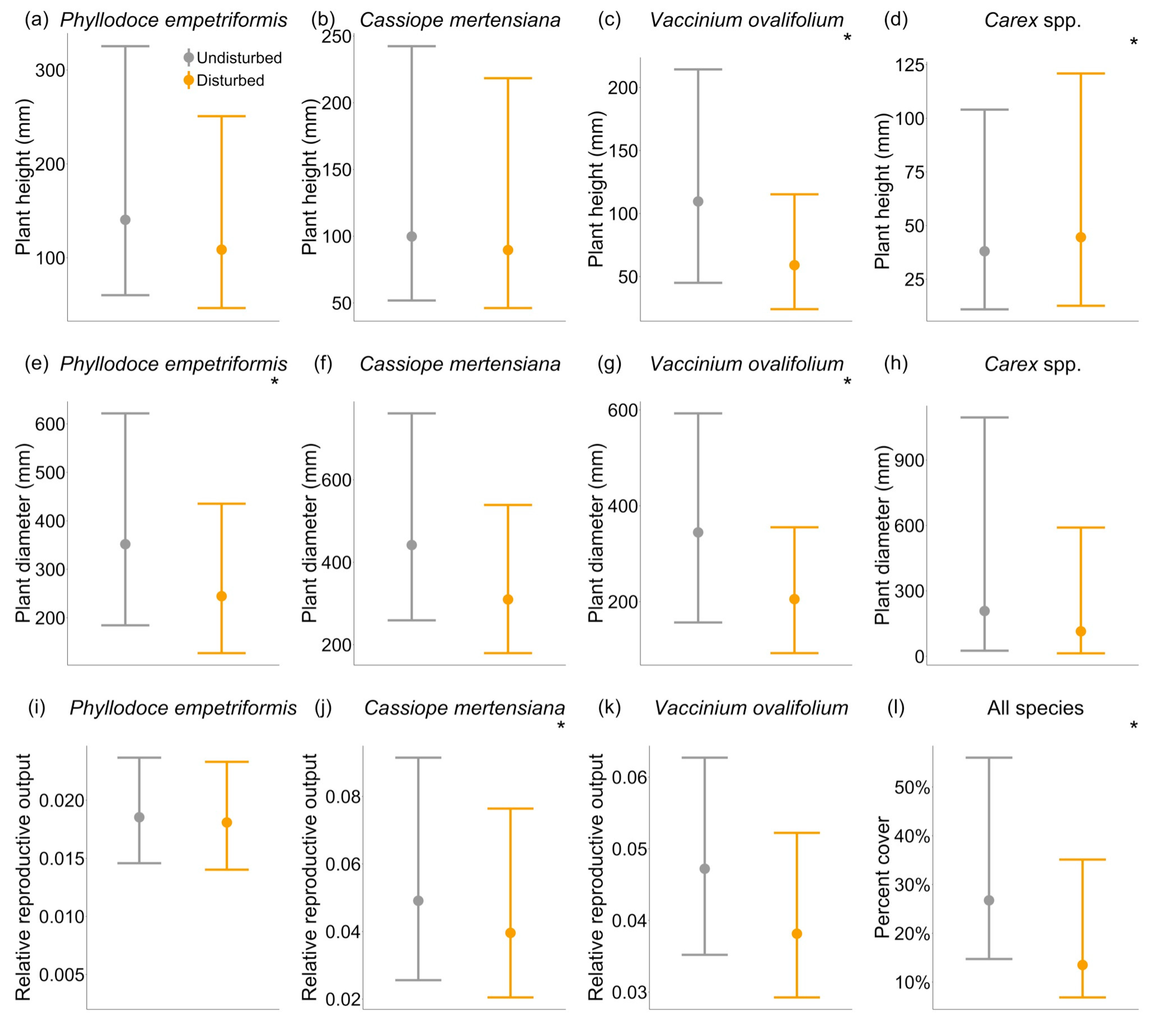
Figure 3.
Methods schema. (a) We quantified the effects of disturbance along elevational gradients on four dominant subalpine and alpine plant species along disturbed (orange) and undisturbed (grey) transects in the.
Figure 3.
Methods schema. (a) We quantified the effects of disturbance along elevational gradients on four dominant subalpine and alpine plant species along disturbed (orange) and undisturbed (grey) transects in the.


Table 1.
Estimated parameters of hierarchical models. Sample size (N), parameter estimates (and credible intervals) from Bayesian generalized non-linear mixed models testing the effects of disturbance, elevation, and their interaction (Dist*Elev) on total plant percent cover, and maximum height, maximum diameter, and relative reproductive output of Phyllodoce empetriformis, Cassiope mertensiana, Vaccinium ovalifolium, and Carex spp. Estimated parameters whose 95% credible intervals do not overlap zero are in bold, and can be interpreted as having an effect. We fit models with a random intercept of transect pair and fit separate models for plant percent cover and each species by trait combination to account for differences responses. We did not measure reproductive output for Carex spp. Additional model fitting information is given in Table S1.
Table 1.
Estimated parameters of hierarchical models. Sample size (N), parameter estimates (and credible intervals) from Bayesian generalized non-linear mixed models testing the effects of disturbance, elevation, and their interaction (Dist*Elev) on total plant percent cover, and maximum height, maximum diameter, and relative reproductive output of Phyllodoce empetriformis, Cassiope mertensiana, Vaccinium ovalifolium, and Carex spp. Estimated parameters whose 95% credible intervals do not overlap zero are in bold, and can be interpreted as having an effect. We fit models with a random intercept of transect pair and fit separate models for plant percent cover and each species by trait combination to account for differences responses. We did not measure reproductive output for Carex spp. Additional model fitting information is given in Table S1.
| Species | Trait | N | Intercept | Disturbance | Elevation | Dist*Elev |
|---|---|---|---|---|---|---|
| [All Plants] | Percent Cover | 267 | 6.58 ( 0.34 , 12.61 ) | -8.38 ( -12.56 , -4.4 ) | 0 ( -0.01 , 0 ) | 0 ( 0 , 0.01 ) |
| P. empetriformis | Height | 500 | 7.04 ( 2.27 , 11.87 ) | -0.44 ( -3.05 , 2.11 ) | 0 ( 0 , 0 ) | 0 ( 0 , 0 ) |
| Diameter | 500 | 10.48 ( 6.66 , 14.16 ) | -7.15 ( -10.53 , -3.85 ) | 0 ( 0 , 0 ) | 0 ( 0 , 0.01 ) | |
| Reproduction | 500 | -4.77 ( -7.78 , -1.76 ) | 0.39 ( -3.47 , 4.22 ) | 0 ( 0 , 0 ) | 0 ( 0 , 0 ) | |
| C. mertensiana | Height | 429 | 7.84 ( 3.73 , 12.04 ) | 0.66 ( -1.27 , 2.59 ) | 0 ( 0 , 0 ) | 0 ( 0 , 0 ) |
| Diameter | 429 | 9.44 ( 5.9 , 12.98 ) | -2.51 ( -5.13 , 0.17 ) | 0 ( 0 , 0 ) | 0 ( 0 , 0 ) | |
| Reproduction | 429 | 0.66 ( -4.18 , 5.38 ) | -4.28 ( -8.13 , -0.52 ) | 0 ( 0 , 0 ) | 0 ( 0 , 0 ) | |
| V. ovalifolium | Height | 634 | 2.1 ( -4.54 , 8.75 ) | -4.6 ( -7.41 , -1.84 ) | 0 ( 0 , 0.01 ) | 0 ( 0 , 0 ) |
| Diameter | 634 | 6.39 ( -0.05 , 12.96 ) | -9.18 ( -12.34 , -5.99 ) | 0 ( 0 , 0 ) | 0.01 ( 0 , 0.01 ) | |
| Reproduction | 634 | -4.5 ( -7.93 , -0.73 ) | -3.76 ( -7.99 , 0.33 ) | 0 ( 0 , 0 ) | 0 ( 0 , 0 ) | |
| Carex spp. | Height | 209 | 22.85 ( 5.06 , 39.25 ) | -20.04 ( -30.96 , -9.37 ) | -0.01 ( -0.02 , 0 ) | 0.01 ( 0.01 , 0.02 ) |
| Diameter | 209 | -6.29 ( -29.7 , 16.07 ) | 11.98 ( -0.5 , 23.86 ) | 0.01 ( -0.01 , 0.02 ) | -0.01 ( -0.01 , 0 ) |
Table 2.
Measured parameters. All of the parameters used in this study, and the methodology, unit, and spatial scale of measurement
Table 2.
Measured parameters. All of the parameters used in this study, and the methodology, unit, and spatial scale of measurement
| Parameter | Method | Unit of Measurement | Spatial Scale |
|---|---|---|---|
| Maximum plant height | Field measurement | Plant | 0.0001 - 1 m |
| Maximum plant diameter | Field measurement | Plant | 0.002 - 1.5 m |
| Reproductive Structures(buds + flowers + fruits) | Field counts | Plant | 0.0001 - 1.5 m |
| Plant Percent Cover | Computed from field photos | Quadrat | 1 x 0.5 m |
| Elevation | Computed from field lat/long | Transect | 30 m |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



