
Submitted:
30 May 2023
Posted:
31 May 2023
You are already at the latest version
Abstract
Fragile X Syndrome (FXS) is a pervasive developmental disorder and the most common monogenic cause of autism spectrum disorder (ASD). Female heterozygous (HET) carriers play a major role in the transmission of the pathology and present several FXS- and ASD-like behavioral alterations. Despite their clear genetic origins, FXS symptoms are known to be modulated by environmental factors, e.g., the exposure to chronic stress, especially during critical life periods, such as pregnancy. Pregnancy, together with pups’ care, constitutes maternal experience, i.e., another powerful environmental factor affecting several neurobehavioral functions in females. Here we investigated the impact of maternal experience on the long-term effects of stress in Fmr1-HET female mice. Our findings demonstrated that the behavioral abnormalities of HET females, i.e., hyperactivity and memory deficits, were unaffected by stress or maternal experience. In contrast, stress, independently of maternal experience, induced the appearance of cognitive deficits in WT mice. Maternal experience increased anxiety levels in all mice and enhanced their corticosterone levels, concomitantly promoting the effects of stress on social communication and adrenal glands. Our results advance our understanding of the environmental modulation of the behavioral alterations observed in FXS mice and highlight the long-term impact of maternal experience and its interactions with chronic stress.
Keywords:
Neurodevelopmental disorders
; unpredictable stress
; ultrasonic communication
; Fmr1
; maternal care
1. Introduction
Fragile X syndrome (FXS) is a major neurodevelopmental disorder (NDD), caused by the absence of fragile X mental retardation protein (FMRP), coded by the FMR1 X-linked gene and involved in synaptic functionality [1]. FXS patients are characterized by a variety of behavioral symptoms, including anxiety, hyperactivity, cognitive and social deficits [2]. These also include autistic-like symptoms, e.g., reduced social interest and altered communication; FXS indeed represents the most common monogenic cause of autism spectrum disorder (ASD) [3,4,5,6,7]. These FXS- and ASD-like behavioral phenotypes have been successfully recapitulated by the Fmr1-KO mouse model that has been extensively employed to investigate the etiopathology and pharmacology of these NDDs. However, the great majority of the existing studies on Fmr1 mutant mice have focused on males, because of the higher prevalence of FXS and the more marked severity of its symptoms in boys [8,9].
Nonetheless, FXS females are of great interest for human research, as they also show several behavioral alterations, including hyperactivity [10], mild cognitive impairments [11,12] and autistic symptoms [5]. Furthermore, female FXS subjects, in particular heterozygous female carriers, play a central role in the progression of this pathology, since they are responsible for the affected offspring [13], as affected males are incapable to pass the full mutation to their daughters [14]. Moreover, heterozygous females constitute the majority of FXS women, since homozygous FMR1 mutations are rarely found in the human population [15]. The behavioral investigation of heterozygous females in pre-clinical studies has therefore a high translational value; several studies have described the presence of FXS and ASD-like behavioral abnormalities in Fmr1 mutant females [16,17,18,19,20,21,22]. These include hyperactivity, anxiety, deficits in spatial memory, alterations in social behaviors and ultrasonic communication [16,17,18,19,20,21,22,23]; these behavioral alterations started to emerge in Fmr1 heterozygous females during the juvenile age and were mostly confirmed at adulthood [21,22].
Despite their clear and well-defined genetic origins, the pathological phenotypes of FXS can be strongly modulated by environmental factors, such as environmental stimulation or adversity. Behavioral stimulation is known to be able to induce therapeutic effects in FXS and ASD patients [e.g., [24]], as confirmed by pre-clinical studies demonstrating that environmental enrichment can rescue several neurobehavioral FXS- and ASD-like alterations in Fmr1-KO mice [25,26]. The chronic exposure to stressful events is instead known to exacerbate the behavioral symptoms of FXS patients [27,28], while the prenatal experience of stressful life events [29],[30], [31,32] have been shown to be associated with a more elevated risk for ASD in humans [30,33]. Similarly, the exposure to a chronic stress has shown to affect the behavioral phenotype of Fmr1-KO mice [34,35] and to anticipate its appearance in male and female mutants [22]. Overall, the reactivity to stress of Fmr1-KO mice is altered compared to their WT littermates, in line with human data from FXS patients, suggesting a role of gene-environment interactions in the etiopathology of FXS. Nonetheless, the early timing of stress exposure seems to be crucial in determining its behavioral effects in Fmr1-KO mice; pre-natal stressful experiences [22] induced a more marked impact than post-natal stress [34,35] with more pronounced and varied behavioral effects. Nonetheless, these studies have analysed the effects of gestational stress exclusively on the behavioral phenotypes of the Fmr1 offspring; therefore, no data are available on the effects of stress during pregnancy on the FXS-like phenotype of Fmr1 heterozygous dams.
The gestational phase represents a critical neurobehavioral period, marked by physiological and neurobehavioral changes in the future mother, underpinned by neuroendocrine modifications that have the role of ensuring the proper development of the offspring during embryonic and post-natal life. These changes persist during the early post-partum phases, which together with pregnancy constitute maternal experience; these modifications also involve the activity of the hypothalamic-pituitary-adrenal axis (HPA) [36,37]. Hence, maternal experience may markedly affect the female vulnerability to stress: indeed it has been shown that the exposure to a deleterious/stressful environment during the gestational period may promote the appearance of behavioral disorders in the mother, the risk being greater if associated with a genetic predisposition [37].
Although maternal experience may modulate the vulnerability to stress, for example by magnifying its neurobehavioral and endocrine effects, it is also possible that stress may alter the effects of maternal experience itself. Several behavioral differences, e.g., on anxiety and cognition, have been described between primiparous and nulliparous rodents (especially rats) [review, [36,38])]. An improvement in spatial memory has been for example described in rats with maternal experience [39], probably to allow mothers to better care for their pups by reducing the time to find their nests [38]. Interestingly, this cognitive improvement of primiparous versus nulliparous females disappeared following exposure to gestational stress, thus suggesting that stress during pregnancy may eliminate the beneficial behavioral effects of motherhood [40,41]. This hypothesis is further supported by the marked effects of gestational stress on maternal care, since previous studies have demonstrated that stressed dams performed less pup-oriented behaviors (e.g., licking/grooming, nursing postures) than non-stressed ones [42,43,44] . It is therefore possible that these effects of stress on maternal care may, in turn, have an impact on other behaviors of the mothers and mediate the stress effects on maternal experience.
Since most of the studies investigating the behavioral effects of maternal experience and of its interactions with stress have been conducted in rats, little is known about this issue in the mouse species, which is essential to the study of most genetic models of human diseases. Hence, the impact of stress x maternal experience interactions on the genetic predisposition to develop NDDs has not been studied yet in mice. In the present study, we therefore investigated the relative impact of stress and maternal experience on the FXS- and ASD-like behavioral and endocrine phenotypes of Fmr1 mutant female mice, following the experimental design illustrated in Figure 6. Fmr1-heterozygous (HET) and wild-type (WT) females, either pregnant or not (primiparous or nulliparous groups), were exposed to one week of unpredictable chronic stress (based on a protocol used in several previous studies on prenatal stress, e.g., [22,45,46,47,48]) or left undisturbed (stressed or no-stressed groups). The behavior of all females was evaluated 5 weeks after the end of the differential stress exposure through tests tailoring anxiety and exploration (elevated plus maze and open field), spatial memory (Y-maze), social interaction and ultrasonic communication, i.e., all tests used in previous studies on Fmr1 females [21,22]. Furthermore, the corticosterone blood levels and adrenal gland weight were assessed one week after the end of behavioral testing. In order to assess the potential role of changes in maternal care in mediating the effects of gestational stress, the maternal behavior of primiparous dams was also assessed during the first week after parturition. Our overall hypothesis was that Fmr1-HET females would be more sensitive to the behavioral and endocrine effects of stress and that this sensitivity would be further accentuated by pregnancy/maternal experience.
2. Results
Maternal Behavior
Stress during pregnancy reduced maternal care and this effect was slightly more pronounced in WT than in HET dams (Figure 1). Indeed, WT stressed dams performed less arched back postures than controls, an effect that was absent in HET females [interaction genotype x stress: F(1,36)= 4.99, p=0.03, post-hoc: WT-stress versus WT-no stress, p<0.05; Figure 1-A]. Stress decreased the expression of another nursing posture, i.e., blanket, as well as of licking/grooming of the pups, but this effect was observed in both WT and HET dams [stress effect, respectively: F(1,36)= 8.61 and 45.12, p=0.006 and p<0.0001; Figure 1-B and C]. No statistically significant effect was detected on non-nursing postures [genotype effect, stress effect and their interaction: F(1,36)=0.01, 2.45 and 2.02; ns; Figure 1-D].
Elevated Plus Maze
Maternal experience increased the anxiety levels of all mice, independently of their genotype and stress condition: primiparous females spent less percent time in the open arms of the maze than nulliparous mice [maternal experience effect: F(1,70)=6.82, p=0.01; Figure 2-A].
Open Field
As observed in the elevated plus maze, maternal experience increased the anxiety levels of all mice, independently of their genotype and stress condition: primiparous females spent less time in the central area of the open field than nulliparous mice [maternal experience effect: F(1,70)=6.16, p=0.02; Figure 2-B]. Mutant (HET) mice were more active than their WT littermates in the open field, independently of their stress and maternal conditions [Genotype effect on the distance travelled: F(1,71)=24.81, p<0.0001; Figure 2-C].
Y maze
Only WT no-stress females (both nulliparous and primiparous) showed a significant preference for the novel arm (t-test versus the chance level in nulliparous wt-no stress nulliparous and wt-no stress primiparous: t(5)=3.26 and t(10)=2.35, p=0.02 and 0.04; Figure 2-D), while all other groups displayed levels of spontaneous alternation that were undistinguishable from chance. Hence, both the mutant HET genotype and stress exposure induced a cognitive deficit in the Y maze test.
Social interaction and ultrasonic communication
Mutant genotype, exposure to stress or maternal experience did not affect the time spent in affiliative behaviors with an adult virgin female [all effects and interaction, ns; Figure 3-A]. No difference among experimental groups was found on non-social behaviors (data not shown). HET females also did not display quantitative differences in ultrasonic communication from their WT littermates, either in the USV number or mean duration [main effects of genotype, ns; Figure 3-B and C]. Stress increased instead the number of USVs, but only in primiparous females [stress x maternal experience interaction: F(1,63)=4.29, p=0.04; Figure 3-B]. The qualitative analysis of call types revealed a highly comparable distribution of call types in all groups [all main effects and their interactions, ns; Figure 4].
Corticosterone blood levels and adrenal gland weight
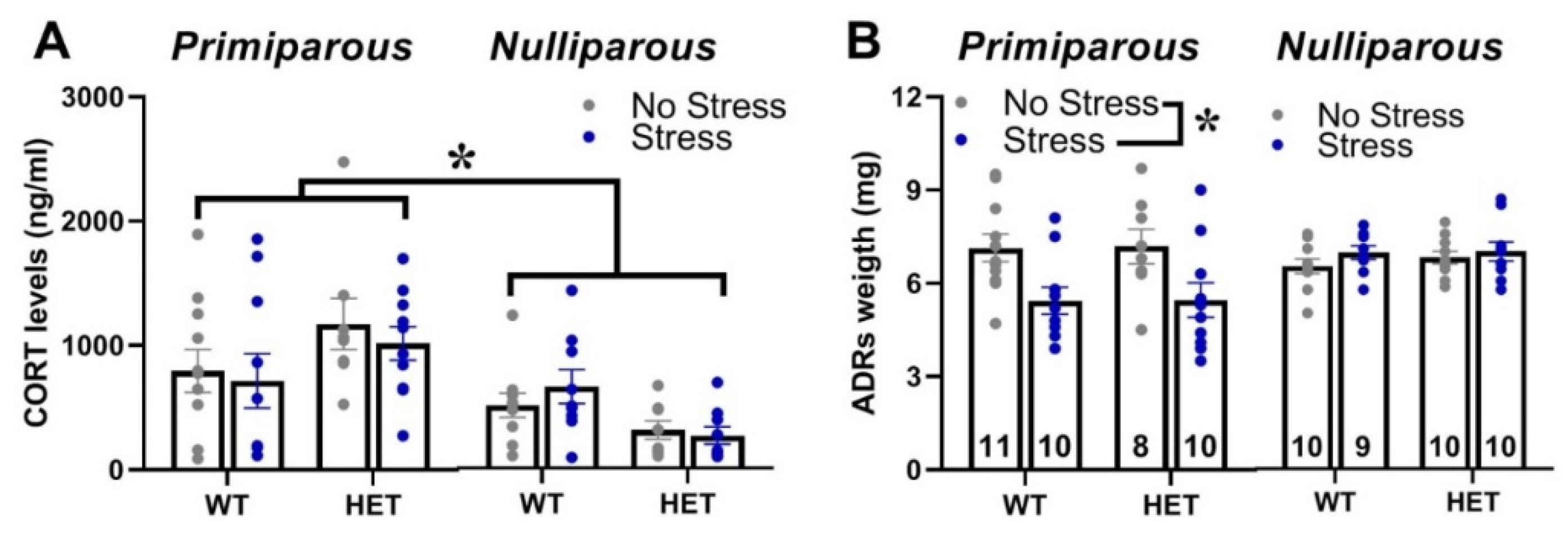
Maternal experience increased the basal blood levels of corticosterone of all females and this effect was observed in mice of both genotypes and stress conditions [main effect of maternal experience: F(1,69)=13.01, p=0.0006; Figure 5-A]. Maternal experience also promoted the effects of stress on adrenal gland weight, since stress reduced it only in primiparous females of both genotypes [interaction maternal experience x stress: F(1,70)=13.25, p=0.0005; Figure 5-B].
3. Discussion
In this study we investigated the relative impact of mild chronic stress and maternal experience on the behavioral and endocrine profile of Fmr1-HET females. Our overall hypothesis was that stress could exacerbate the severity of the FXS-like phenotypes of Fmr1-HET mice and these effects would be exaggerated by maternal experience, i.e., when stress was applied during pregnancy. Hence, we hypothesized that the Fmr1 mutation would enhance the vulnerability of female mice to chronic mild stress. Our findings demonstrated instead that the behavioral phenotypes of mutant females, i.e., hyperactivity and cognitive deficits, were unaffected by stress exposure even when applied during pregnancy. Furthermore, Fmr1-HET females were slightly less sensitive to the effects of stress than their WT littermates, since the effects of stress on maternal behavior and spatial memory in the Y maze tended to be more marked in WT mice. Our results also showed that certain effects of stress were promoted by maternal experience, as we expected, such as those detected on ultrasonic communication and adrenal gland weight, but others were instead independent, i.e., the cognitive effects in the Y maze. Furthermore, we demonstrated that maternal experience alone could enhance anxiety and basal corticosterone levels and these effects were equally observed in mice of both genotypes.
Concerning the altered phenotypes of Fmr1-HET females, our results here demonstrated mutant hyperactivity in the open field (Figure 2-C) and cognitive deficits in the Y maze (Figure 3-D), without alterations in emotionality (Figure 2-A and B), social interaction (Figure 3-A) and ultrasonic communication (Figure 3-B, C and Figure 4). This behavioral phenotype was mostly in agreement with previous reports, describing robust motor and cognitive alterations in mutant females, with inconsistent abnormalities in anxiety-like and social behaviors (e.g., [21,22,23]). Nonetheless, we previously described enhanced USV mean duration in HET females [21,22], associated with the emission of more complex ultrasonic calls compared to WT littermates [22]. It is possible that the lack of USV qualitative alterations we found here in HET females could be related to the particular social housing conditions we used (e.g., housing first with a male, followed by a single housing period before being housed in same-sex pairs, see also Figure 6) that were different from the standard social housing conditions used in our previous studies on Fmr1-HET mice [21,22]. Altogether, our results on the behavioral profile of Fmr1 mutant females confirmed that motor and cognitive alterations represent the most robust FXS-like phenotypes, as it has been suggested for Fmr1-KO males [23,56]. Furthermore, our findings add to the existing knowledge of the behavioral phenotypes of the Fmr1-heterozygous providing evidence of unaltered maternal care in Fmr1-HET females. Interestingly, Fmr1-HET mice were undistinguishable from their WT littermates in their levels of nursing postures as well as pups’ care (Figure 1). This result is highly relevant to research on the Fmr1 mouse line, since previous studies have demonstrated that certain behavioral phenotypes of Fmr1 males were associated to the genotype of their mothers [57,58,59], suggesting a role of alterations in maternal care, milk composition or immunological factors [59]. Our findings thus allow ruling out differences in maternal behaviors as the source of the effects of maternal HET mutation on the offspring behavioral phenotypes in the Fmr1 mouse model.
In contrast to our hypothesis of a higher vulnerability of the Fmr1 mutant genotype, the FXS-like behavioral phenotypes of HET females, i.e., hyperactivity and cognitive deficits, were unaltered by stress, independently of maternal experience. Interestingly, while stress did not affect locomotor activity in all animals, it impaired the Y-maze performance in WT mice, without exacerbating the cognitive deficit of HET females (Figure 2-D). Hence, in disagreement with our initial expectations, WT mice seemed more sensitive to the effects of stress, independently of maternal experience. This impression was further supported by the effects of stress on maternal behaviors (Figure 1), which were slightly more marked in WT dams, where it reduced not only blanket posture and pup grooming as in HET mice, but also arched-back posture. Although in contrast with our initial hypothesis, these effects of stress on Y-maze performance and maternal behavior were in line with previous findings from WT rodents, highlighting stress-induced deficits in spatial memory [60,61,62] and maternal care [42,43,44]. Furthermore, a reduced behavioral and endocrine sensitivity of Fmr1 mutant mice to chronic stress has been already suggested by previous studies (though they mostly focused on males) [22,34,35,54], and it could be interpreted as a deficit in the adaptive response to stressors [35]. This hypothesis, suggested also by previous authors [35], is supported by evidence showing that chronic stress induced changes in dendritic branching and morphology in brain areas crucial for behavioral control, i.e., the amygdala, in WT, but not in Fmr1-KO male mice [35]. Interestingly, this attenuated sensitivity to stress displayed by Fmr1 mutants was limited to the behavioral response, since no difference in corticosterone levels or adrenal gland weight was detected here in HET mice (Figure 5). Our results are in agreement with previous studies from Fmr1 mutant males, showing no difference in markers of the HPA axis functioning under basal conditions [35,63,64].
Overall, stress did not seem a highly powerful behavioral and endocrine modulator, at least when applied independently of maternal experience. Indeed, stress did not affect emotionality, locomotion, social interaction or any qualitative characteristic of ultrasonic communication either in WT or HET mice. Only when combined with maternal experience, stress increased the number of USVs and reduced the adrenal gland weight. The limited behavioral effects of stress may be surprising, considering the accumulating evidence from rodents describing several stress-induced effects on emotionality, locomotion and social behaviors [reviewed in [65,66]]. The lack of effects of stress on basal corticosterone levels is instead in line with a large body of data reviewed in [67,68], although discrepancies across studies exist, especially due to methodological differences [35,69]. Nonetheless, it should be noticed that here the long-term behavioral effects of stress were mostly investigated, i.e., between 5 and 7 weeks after the end of stress exposure (see Figure 6), while previous studies were rather focused on short-term effects.
Compared to stress, maternal experience was more efficacious in inducing behavioral and endocrine effects in mice of both genotypes. Primiparous WT and mutant females showed increased anxiety levels in both the elevated plus maze and open field tests than nulliparous mice together with enhanced basal corticosterone levels. These two effects are likely to be linked, as suggested by previous literature on corticosterone and emotionality [70,71,72,73]. The higher corticosterone levels of primiparous females could also explain the reduced weight of adrenal glands that we detected when maternal experience was combined with stress, possibly linked to a negative feedback mechanism on HPA axis. Overall, the effects of maternal experience we detected in the present study were mostly “detrimental”, inducing blunted emotionality and high chronic stress levels; this finding could seem surprising, considering the general “beneficial” effects of maternal experience previously described, especially on cognition, in rats [reviewed in [74]. Although an anxiogenic effect of maternal experience has already been described [36,38], comparisons with previous studies on maternal experience are complicated by differences in the experimental design. Among these, the most relevant are the inclusion of primiparous versus multiparous comparisons, differences in the interval from birth/weaning of the pups and mothers’ testing as well as the choice of control groups without a vasectomized male (that was instead employed in our study).
In conclusion, our findings suggest the presence of negligible interactions between environmental factors, such as stress and maternal experience, and the mutant genotype of Fmr1 heterozygous female mice. Our results highlight instead the behavioral and endocrine impact of maternal experience in mice of both genotypes and its ability to interact with chronic mild stress. The present findings thus contribute to advancing our understanding of the complex environmental modulation of the pathological phenotypes of the Fmr1 preclinical model of FXS.
4.4. Materials and Methods
Breeding procedures
A total of 81 female mice were used for the study: 42 “primiparous” [22 WT and 20 HET (+/-), n=11/10 for each stress condition] and 39 “nulliparous” [19 WT and 20 HET (+/-), n=9/10 for each stress condition]. C57BL/6JFmr1tm1Cgr/Nwu (B6) female mice were originally obtained from Neuromice.org (Northwestern University) and maintained on the C57BL6/J background for more than 10 generations. Fmr1 heterozygous (+/-) females and their WT littermates used for this study were bred and genotyped in our animal facility at Bordeaux University as described in detail elsewhere [75]. They underwent the experimental procedures that are schematically summarized in Figure 6.
Forty-two adult (12±1 weeks-old) virgin females [20 Fmr1 heterozygous (+/-) and 22 WT littermates] and 21 C57BL/6J adult wild type intact males [16 weeks-old; purchased from Janvier (Le Genest St Isle, France)] were used for the « primiparous » experimental group. Thirty-nine adult (12±1 weeks-old) virgin females [20 Fmr1 heterozygous (+/-) and 19 WT littermates] and 20 C57BL/6J adult wild type vasectomized males [16 weeks-old; purchased from Janvier (Le Genest St Isle, France)] were used for the « nulliparous » group. Breeding trios (2 females, either WT or HET, and one WT male, either intact or vasectomized for the primiparous or the nulliparous condition, respectively) were formed. Pregnancy was determined by the presence of semen in vaginal smears and its detection identified the first day of gestation (GD0). For both primiparous and nulliparous groups the male was removed after two weeks from the formation of breeding trios (Figure 6). After removal of the male, pregnant and non-pregnant females were randomly assigned to either the no-stress or stressed group and they were individually housed in polycarbonate standard cages (37×21×15 cm in size; Tecniplast, Limonest, France), provided with sawdust bedding (SAFE, Augy, France) and a stainless steel wired lid. At 18 weeks of age, all females were housed in same-sex pairs and kept under these social housing conditions until the end of the study. Food chow (SAFE, Augy, France) and water were provided ad libitum. All animals were maintained in a temperature- (22°C) and humidity- (55%) controlled vivarium, under a 12:12 h light–dark cycle (lights on at 7 a.m.).
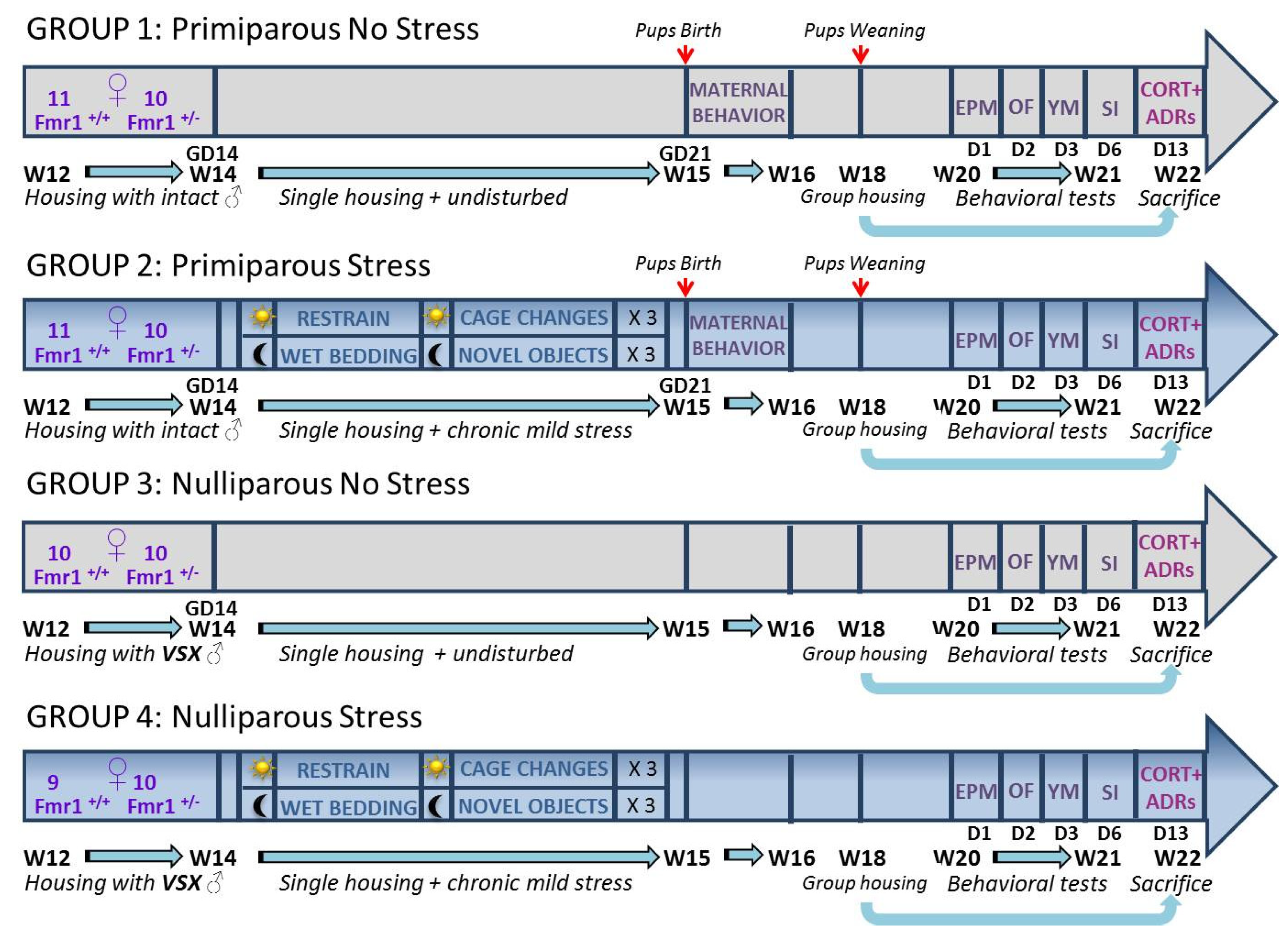
Figure 6.
Schematic representation of the experimental design of the study. Unpredictable mild stress (groups 2 and 4) consisted of a 2 day-sequence that was repeated 3 consecutive times: on day 1, 3 sessions of 30-min restrain stress during the light phase were followed by overnight housing with wet bedding; on day 2, 3 sessions of sawdust and cage changes during the light phase were followed by overnight housing with novel objects. Control no stress mice were instead left undisturbed. Group housing consisted in same-sex pairs. W=week, referring to the age of the subjects; VSX= vasectomized; GD=gestational day; D=day; EPM= elevated plus maze; OF=open field; YM=Y maze; SI=social interaction; CORT=corticosterone; ADRs= adrenal glands.
Figure 6.
Schematic representation of the experimental design of the study. Unpredictable mild stress (groups 2 and 4) consisted of a 2 day-sequence that was repeated 3 consecutive times: on day 1, 3 sessions of 30-min restrain stress during the light phase were followed by overnight housing with wet bedding; on day 2, 3 sessions of sawdust and cage changes during the light phase were followed by overnight housing with novel objects. Control no stress mice were instead left undisturbed. Group housing consisted in same-sex pairs. W=week, referring to the age of the subjects; VSX= vasectomized; GD=gestational day; D=day; EPM= elevated plus maze; OF=open field; YM=Y maze; SI=social interaction; CORT=corticosterone; ADRs= adrenal glands.
Stress procedure
While females from the no-stress group were kept undisturbed in their home-cage (Figure 6), mice from the stress group were exposed to the unpredictable stress procedure. This included the following 2-day sequence of events that was repeated three consecutive times:
-Day 1: 30 minutes of restrain stress (3 times each day during the light phase, with a 4h-interval) in perforated conical tubes (3cm in diameter, 11.5cm long; Becton Dickinson Labware Europe, France), followed by overnight housing with wet bedding (50ml of water were added to floor sawdust of the home cage at the beginning of the dark phase).
-Day 2: multiple sawdust and cage changes (3 times each day during the light phase, with a 4h-interval), followed by overnight housing with novel objects (12 glass black beads, 1.5cm in diameter were added in the home cage at the beginning of the dark phase).
Pregnant (primiparous group) and non-pregnant (nulliparous group) females were exposed to this sequence of events for 3 times during one week (for pregnant females during the last week before parturition): this procedure was based on previous studies [e.g., [22,45,46,47,48]] and it is known to limit the habituation to stressful stimuli without using pain or nutritional manipulations. All pregnant females used for the study gave birth within 48hs after the last day of exposure to stress procedure. No alteration in the general health status of stressed animals emerged at the end of the stress paradigm. The health measures were taken by the animal caretakers through the daily observation of the animals in their home cage in order to assess both behavioral and physical indicators of welfare [76]. These included hunched posture, dull or sluggish movements, reduced locomotion/immobility, altered nest building and stereotypic behaviors, excessive grooming, absence of feces, rough hair coat, squinted eyes and skin abrasions/lesions [76].
Behavioral testing procedures
For the primiparous groups, maternal behavior was observed in their home-cages twice a day for one hour (at 9.00 AM and at 5.00 PM) during the first 6 postnatal days of the pups, using an instantaneous sampling method (one sampling/2min, for a total of 30 sampling points/session). The following items were scored as absolute frequencies by an observer who was blind to the stress condition of the breeders [26,49,50]: (i) nursing postures, including arched-back nursing (the female is in an arched position over the nursing pups) and blanket nursing (the female is lying flat on top of the pups), (ii) non-nursing postures (the female is in contact with the pups, but not nursing, i.e., with no access to the nipples), (iii) licking/grooming of the pups.
For all subjects behavioral tests began at 20 weeks of age (i.e., 2 weeks after weaning of the pups for the primiparous group) and were conducted as follows (see also Figure 6). On day 1, an elevated plus maze test for anxiety was administered, followed on day 2 by an open field test for locomotion and exploration, on day 3 by a spontaneous alternation test in a Y-maze and on day 6 by a direct social interaction test and the females’ estrous cycle assessment. All behavioral tests were carried out during the light phase of the cycle (between 9 a.m. and 4 p.m.) by an experimenter who was blind to the group assignment of the subjects. All mice were habituated to the experimental room for at least 30 min before the beginning of each behavioral test.
Elevated plus maze
The maze described in detail elsewhere [75,77] was placed 55cm above floor level, in a quiet testing room with diffuse dim lighting (55 lux in the maze center). A digital camera was mounted above the maze, and images were transmitted to a PC running the Ethovision (Version 13, Noldus Technology, The Netherlands) tracking system. To begin a trial, the mouse was gently placed in the central square with its head facing one of the open arms and allowed to explore freely for 5 min. We measured the percent time in open arms as (time(open arms) / time(open + closed arms)) x 100.
Open field
The open field consisted of a white opaque plastic arena (42x26x15cm) under dim lighting conditions (55 lux). Each mouse was placed in the center of the arena and allowed to freely explore it for 10 minutes. Locomotor habituation, requiring longer testing sessions, was not assessed since it is known to be unaltered in Fmr1-KO adult mice [e.g., [75,78]; see also [23] for a review]. Automated tracking of the videos obtained from a camera mounted above the open field was performed by Ethovision to analyze the distance travelled.
Y maze
A grey plastic Y-maze (each arm measuring 8x42x15cm, 120° spaced) was placed on a table 80cm high, in a room with extramaze cues on the walls. For the habituation phase, each mouse was introduced to the end of one maze arm and allowed to explore two arms for 5 minutes, while the access to the third arm was blocked by a transparent plastic door. After an interval of 10 minutes in a waiting cage, the testing phase began: the door of the blocked arm was removed and the mouse was allowed to explore all three arms for 2 minutes. Allocations of the start and blocked arms were counterbalanced within experimental groups. Time spent in each arm during the habituation and testing phases was scored by Ethovision through automatic tracking of the videos collected from a camera mounted above the maze center. We measured the percent alternation rate as (time(novel arm) / time(all arms)) x 100.
Social interaction and ultrasonic communication
Stimulus mice used for the direct social interaction test were adult (12 weeks of age) female NMRI mice, as this strain is commonly employed in social studies [79,80], especially those using the Fmr1-KO mouse model [21,26,81]. This strain is often chosen since it is characterized by high levels of sociability, and it facilitates the behavioral analysis during social encounters with B6 mutants because of its albino phenotype. NMRI mice were purchased at 10 weeks of age from Janvier (Le Genest-Saint-Isle, France), housed in groups of 3-4 per cage and left undisturbed for 2 weeks before being used in behavioral tests. The choice of the age of stimulus mice was based on previous studies with females in the resident-intruder setting [80,82,83], all using adult stimulus females. Indeed, in this experimental context, adult stimulus females do not emit ultrasonic vocalizations (USVs) that are instead mostly uttered by the resident female [82,83], as demonstrated by alternately anesthetizing each pair member. The absence of “double calls”, i.e., overlapping in their timing, but with different, non-harmonic, characteristics (e.g., different peak and mean frequency, modulation) was also confirmed here by the inspection of all spectrograms.
Female experimental subjects (WT or Fmr1-HET mice) were isolated in the testing cage for 72hs, in order to induce a status of resident in experimental females and therefore promote the emission of ultrasonic vocalizations (USVs) towards an adult female intruder [80]. An unfamiliar NMRI stimulus female mouse was then introduced into the testing cage of either male or female subjects and left there for 3 min.
Testing sessions were recorded by a camera placed on the side of the cage and videos analyzed with Observer XT (Noldus, The Netherlands). One observer who was unaware of the genotype and sex of the animals scored the behavior of the test mice, quantifying the time spent performing affiliative behaviors [21,26,81], i.e., sniffing the head and the snout of the partner, its anogenital region, or any other part of the body; contact with the partner through traversing the partner’s body by crawling over/under from one side to the other or allogrooming. Nonsocial activities were also measured [26,81]: rearing (standing on the hind limbs sometimes with the forelimbs against the walls of the cage); digging; self-grooming (the animal licks and mouths its own fur).
An ultrasonic microphone UltraSoundGate Condenser Microphone CM 16 (Avisoft Bioacoustics, Berlin, Germany) was mounted 2 cm above the cover of the testing cage. Recordings were then analyzed through Sonotrack Software (version 1.4.7, Metris B.V., The Netherlands) This software fully automatically recognizes several different USV types and also calculates quantitative parameters including the total number and mean duration of the calls. Based on previous literature on call types [22,53,54,55,84,85,86], the following USV types were selected for automatic recognition in our dataset: Short, Flat, (Ramp) Up, (Ramp) Down, Chevron, Step-Up, Step-Down, Step-Double (Split), Complex-3, Complex-4, Complex-5, Complex-5+. Their characteristics are described in detail elsewhere [22,53,54,55] Definitions of call types were mutually exclusive. Overlap of components was removed when more than 70 % to prevent wrong call durations. Short gaps between components in both frequency (≤ 6 kHz) and time (≤ 5 ms) were interpolated (gaps can be caused by changes in microphone sensitivity or direction of vocalization). The calls classified as “complex 3 component” and “+3 component” were summed up into a “total complex” category, as in previous studies [22,53,54,55].
The estrus phase of female mice was assessed by analysis of vaginal smears [87] performed on the testing day in both the experimental subjects and NMRI stimulus mice. The evaluation of Fmr1 WT and HET (+/-) females used as experimental subjects was conducted after their testing, in order to minimize the potential stress effects of the manipulation necessary for determining the estrous phase. Stimulus NMRI females and experimental subjects were all in the diestrous phase at the moment of social testing.
Assessment of corticosterone blood levels
One week after the end of behavioral testing (Figure 6), mice were euthanized by cervical dislocation; adrenal glands were immediately dissected and weighted. Blood samples were obtained from trunk blood, collected in heparinized tubes and stored on ice. After centrifugation at 3000 r.p.m. for 10 min, the supernatant was stored at -80°C until the assay was performed. Corticosterone measures were assessed in duplicate by ELISA using a commercial kit (Cayman Chemical Company, Ann Arbor, Michigan, USA).
Statistical analysis
Except for data on maternal behavior (that were obviously obtained only from the primiparous group and therefore were analysed without the variable of maternal experience), all data were analysed through a 2x2x2 ANOVA with maternal experience, stress and genotype as the between-subject factors. Alternation rates from the Y-maze test were instead analysed with t-tests from the chance level (corresponding to the value of 33.33%), as in previous studies (e.g., [54,88]). Post-hoc comparisons were performed using Tuckey’s test when a significant interaction was detected.
Analyses were conducted using the software Statview and SPSS and α was set at 0.05. Results are expressed as mean ± SEM throughout the text. The exact number of mice is indicated in the legend of each figure; slight differences may be due to technical reasons (e.g., problems in behavioral scoring or recording) or to the exclusion of outliers (using Grubbs' ESD test adapted for small sample size).
Author Contributions
E.S. performed most of the experimental work and analysed the behavioral data; V.L. participated to the experimental work, data analysis and study design; V.P. analysed all the ultrasonic vocalizations and prepared all figures; S.P. performed the statistical analyses, supervised the behavioral data collection and wrote the manuscript. All authors reviewed and approved the manuscript.
Funding
This work was supported by Bordeaux University and CNRS. S. Pietropaolo and V. Petroni received funding from Association Autour de Williams and Fondation pour l’Audition (FPA-RD-2020-8).
Institutional Review Board Statement
All experimental procedures were in accordance with European Communities Council Directive 2010/63/EEC. Furthermore, they were approved by local ethical committee (“Comité d’Ethique pour l’expérimentation animale de Bordeaux”, CE 50) and the French Ministry (“Ministère de l’enseignement supérieur de la recherche et de l’innovation”).
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets used and analysed during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
The authors thank Elodie Poinama for animal care, Thierry Lafon for technical support and Marie-Laure Rousseau for administrative assistance.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Verkerk, A. J., M. Pieretti, J. S. Sutcliffe, Y. H. Fu, D. P. Kuhl, A. Pizzuti, O. Reiner, S. Richards, M. F. Victoria, F. P. Zhang, and et al. "Identification of a Gene (Fmr-1) Containing a Cgg Repeat Coincident with a Breakpoint Cluster Region Exhibiting Length Variation in Fragile X Syndrome." Cell 65, no. 5 (1991): 905-14. [CrossRef]
- Hagerman, Randi Jenssen, and Paul J Hagerman. Fragile X Syndrome: Diagnosis, Treatment, and Research: Taylor & Francis US, 2002.
- Bailey, D. B., Jr., D. D. Hatton, M. Skinner, and G. Mesibov. "Autistic Behavior, Fmr1 Protein, and Developmental Trajectories in Young Males with Fragile X Syndrome." J Autism Dev Disord 31, no. 2 (2001): 165-74. [CrossRef]
- Kerby, D. S., and B. L. Dawson. "Autistic Features, Personality, and Adaptive Behavior in Males with the Fragile X Syndrome and No Autism." Am J Ment Retard 98, no. 4 (1994): 455-62.
- Mazzocco, M. M., W. R. Kates, T. L. Baumgardner, L. S. Freund, and A. L. Reiss. "Autistic Behaviors among Girls with Fragile X Syndrome." J Autism Dev Disord 27, no. 4 (1997): 415-35. [CrossRef]
- Mazzocco, M. M., M. Pulsifer, A. Fiumara, M. Cocuzza, F. Nigro, G. Incorpora, and R. Barone. "Brief Report: Autistic Behaviors among Children with Fragile X or Rett Syndrome: Implications for the Classification of Pervasive Developmental Disorder." J Autism Dev Disord 28, no. 4 (1998): 321-8. [CrossRef]
- Reiss, A. L., and L. Freund. "Behavioral Phenotype of Fragile X Syndrome: Dsm-Iii-R Autistic Behavior in Male Children." Am J Med Genet 43, no. 1-2 (1992): 35-46. [CrossRef]
- Hatton, D. D., J. Sideris, M. Skinner, J. Mankowski, D. B. Bailey, Jr., J. Roberts, and P. Mirrett. "Autistic Behavior in Children with Fragile X Syndrome: Prevalence, Stability, and the Impact of Fmrp." Am J Med Genet A 140A, no. 17 (2006): 1804-13. [CrossRef]
- Turner, G., T. Webb, S. Wake, and H. Robinson. "Prevalence of Fragile X Syndrome." Am J Med Genet 64, no. 1 (1996): 196-7. [CrossRef]
- Wheeler, A., M. Raspa, C. Bann, E. Bishop, D. Hessl, P. Sacco, and D. B. Bailey, Jr. "Anxiety, Attention Problems, Hyperactivity, and the Aberrant Behavior Checklist in Fragile X Syndrome." Am J Med Genet A 164A, no. 1 (2014): 141-55. [CrossRef]
- Loesch, D. Z., and D. A. Hay. "Clinical Features and Reproductive Patterns in Fragile X Female Heterozygotes." J Med Genet 25, no. 6 (1988): 407-14. [CrossRef]
- Loesch, D. Z., Q. M. Bui, J. Grigsby, E. Butler, J. Epstein, R. M. Huggins, A. K. Taylor, and R. J. Hagerman. "Effect of the Fragile X Status Categories and the Fragile X Mental Retardation Protein Levels on Executive Functioning in Males and Females with Fragile X." Neuropsychology 17, no. 4 (2003): 646-57. [CrossRef]
- Nolin, S. L., F. A. Lewis, 3rd, L. L. Ye, G. E. Houck, Jr., A. E. Glicksman, P. Limprasert, S. Y. Li, N. Zhong, A. E. Ashley, E. Feingold, S. L. Sherman, and W. T. Brown. "Familial Transmission of the Fmr1 Cgg Repeat." Am J Hum Genet 59, no. 6 (1996): 1252-61.
- Reyniers, E., L. Vits, K. De Boulle, B. Van Roy, D. Van Velzen, E. de Graaff, A. J. Verkerk, H. Z. Jorens, J. K. Darby, B. Oostra, and et al. "The Full Mutation in the Fmr-1 Gene of Male Fragile X Patients Is Absent in Their Sperm." Nat Genet 4, no. 2 (1993): 143-6. [CrossRef]
- Mazzocco, M. M., and J. J. Holden. "Neuropsychological Profiles of Three Sisters Homozygous for the Fragile X Premutation." Am J Med Genet 64, no. 2 (1996): 323-8. [CrossRef]
- Baker, K. B., S. P. Wray, R. Ritter, S. Mason, T. H. Lanthorn, and K. V. Savelieva. "Male and Female Fmr1 Knockout Mice on C57 Albino Background Exhibit Spatial Learning and Memory Impairments." Genes Brain Behav 9, no. 6 (2010): 562-74. [CrossRef]
- Ding, Q., F. Sethna, and H. Wang. "Behavioral Analysis of Male and Female Fmr1 Knockout Mice on C57bl/6 Background." Behav Brain Res 271 (2014): 72-8. [CrossRef]
- Musumeci, S. A., P. Bosco, G. Calabrese, C. Bakker, G. B. De Sarro, M. Elia, R. Ferri, and B. A. Oostra. "Audiogenic Seizures Susceptibility in Transgenic Mice with Fragile X Syndrome." Epilepsia 41, no. 1 (2000): 19-23. [CrossRef]
- Nguy, S., and M. V. Tejada-Simon. "Phenotype Analysis and Rescue on Female Fvb.129-Fmr1 Knockout Mice." Front Biol 11, no. 1 (2016): 43-52. [CrossRef]
- Qin, M., J. Kang, and C. B. Smith. "A Null Mutation for Fmr1 in Female Mice: Effects on Regional Cerebral Metabolic Rate for Glucose and Relationship to Behavior." Neuroscience 135, no. 3 (2005): 999-1009. [CrossRef]
- Gauducheau, M., V. Lemaire-Mayo, F. R. D'Amato, D. Oddi, W. E. Crusio, and S. Pietropaolo. "Age-Specific Autistic-Like Behaviors in Heterozygous Fmr1-Ko Female Mice." Autism Res 10, no. 6 (2017): 1067-78. [CrossRef]
- Petroni, V., E. Subashi, M. Premoli, M. Wohr, W. E. Crusio, V. Lemaire, and S. Pietropaolo. "Autistic-Like Behavioral Effects of Prenatal Stress in Juvenile Fmr1 Mice: The Relevance of Sex Differences and Gene-Environment Interactions." Sci Rep 12, no. 1 (2022): 7269. [CrossRef]
- Pietropaolo, S., and E. Subashi. "Mouse Models of Fragile X Syndrome." In Behavioral Genetics of the Mouse, edited by S. Pietropaolo, F. Sluyter and W.E. Crusio, 146-63. Cambridge: Cambridge University Press, 2014.
- Dawson, G., S. Webb, G. D. Schellenberg, S. Dager, S. Friedman, E. Aylward, and T. Richards. "Defining the Broader Phenotype of Autism: Genetic, Brain, and Behavioral Perspectives." Dev Psychopathol 14, no. 3 (2002): 581-611. [CrossRef]
- Restivo, L., F. Ferrari, E. Passino, C. Sgobio, J. Bock, B. A. Oostra, C. Bagni, and M. Ammassari-Teule. "Enriched Environment Promotes Behavioral and Morphological Recovery in a Mouse Model for the Fragile X Syndrome." Proc Natl Acad Sci U S A 102, no. 32 (2005): 11557-62. [CrossRef]
- Oddi, D., E. Subashi, S. Middei, L. Bellocchio, V. Lemaire-Mayo, M. Guzman, W. E. Crusio, F. R. D'Amato, and S. Pietropaolo. "Early Social Enrichment Rescues Adult Behavioral and Brain Abnormalities in a Mouse Model of Fragile X Syndrome." Neuropsychopharmacology 40, no. 5 (2015): 1113-22. [CrossRef]
- Dyer-Friedman, J., B. Glaser, D. Hessl, C. Johnston, L. C. Huffman, A. Taylor, J. Wisbeck, and A. L. Reiss. "Genetic and Environmental Influences on the Cognitive Outcomes of Children with Fragile X Syndrome." J Am Acad Child Adolesc Psychiatry 41, no. 3 (2002): 237-44. [CrossRef]
- Hessl, D., J. Dyer-Friedman, B. Glaser, J. Wisbeck, R. G. Barajas, A. Taylor, and A. L. Reiss. "The Influence of Environmental and Genetic Factors on Behavior Problems and Autistic Symptoms in Boys and Girls with Fragile X Syndrome." Pediatrics 108, no. 5 (2001): E88. [CrossRef]
- Ward, A. J. "A Comparison and Analysis of the Presence of Family Problems During Pregnancy of Mothers of "Autistic" Children and Mothers of Normal Children." Child Psychiatry Hum Dev 20, no. 4 (1990): 279-88. [CrossRef]
- Beversdorf, D. Q., S. E. Manning, A. Hillier, S. L. Anderson, R. E. Nordgren, S. E. Walters, H. N. Nagaraja, W. C. Cooley, S. E. Gaelic, and M. L. Bauman. "Timing of Prenatal Stressors and Autism." J Autism Dev Disord 35, no. 4 (2005): 471-8. [CrossRef]
- Kinney, D. K., A. M. Miller, D. J. Crowley, E. Huang, and E. Gerber. "Autism Prevalence Following Prenatal Exposure to Hurricanes and Tropical Storms in Louisiana." J Autism Dev Disord 38, no. 3 (2008): 481-8. [CrossRef]
- Kinney, D. K., K. M. Munir, D. J. Crowley, and A. M. Miller. "Prenatal Stress and Risk for Autism." Neurosci Biobehav Rev 32, no. 8 (2008): 1519-32. [CrossRef]
- Beversdorf, D. Q., H. E. Stevens, K. G. Margolis, and J. Van de Water. "Prenatal Stress and Maternal Immune Dysregulation in Autism Spectrum Disorders: Potential Points for Intervention." Curr Pharm Des 25, no. 41 (2019): 4331-43. [CrossRef]
- Lemaire-Mayo, V., E. Subashi, N. Henkous, D. Beracochea, and S. Pietropaolo. "Behavioral Effects of Chronic Stress in the Fmr1 Mouse Model for Fragile X Syndrome." Behav Brain Res 320 (2017): 128-35. [CrossRef]
- Qin, M., Z. Xia, T. Huang, and C. B. Smith. "Effects of Chronic Immobilization Stress on Anxiety-Like Behavior and Basolateral Amygdala Morphology in Fmr1 Knockout Mice." Neuroscience 194 (2011): 282-90. [CrossRef]
- Macbeth, A. H., and V. N. Luine. "Changes in Anxiety and Cognition Due to Reproductive Experience: A Review of Data from Rodent and Human Mothers." Neurosci Biobehav Rev 34, no. 3 (2010): 452-67. [CrossRef]
- Brunton, P. J., and J. A. Russell. "The Expectant Brain: Adapting for Motherhood." Nat Rev Neurosci 9, no. 1 (2008): 11-25. [CrossRef]
- Kinsley, C. H. "The Neuroplastic Maternal Brain." Horm Behav 54, no. 1 (2008): 1-4. [CrossRef]
- Darnaudery, M., M. Perez-Martin, F. Del Favero, C. Gomez-Roldan, L. M. Garcia-Segura, and S. Maccari. "Early Motherhood in Rats Is Associated with a Modification of Hippocampal Function." Psychoneuroendocrinology 32, no. 7 (2007): 803-12. [CrossRef]
- Lemaire, V., J. M. Billard, P. Dutar, O. George, P. V. Piazza, J. Epelbaum, M. Le Moal, and W. Mayo. "Motherhood-Induced Memory Improvement Persists across Lifespan in Rats but Is Abolished by a Gestational Stress." Eur J Neurosci 23, no. 12 (2006): 3368-74. [CrossRef]
- Lemaire, V., S. Lamarque, M. Le Moal, P. V. Piazza, and D. N. Abrous. "Postnatal Stimulation of the Pups Counteracts Prenatal Stress-Induced Deficits in Hippocampal Neurogenesis." Biol Psychiatry 59, no. 9 (2006): 786-92. [CrossRef]
- Muir, J. L., H. P. Pfister, and A. Ivinskis. "Effects of Prepartum Stress and Postpartum Enrichment on Mother-Infant Interaction and Offspring Problem Solving Ability in Rattus Norvegicus." Journal of Comparative Psychology 99, no. 4 (1985): 468-78.
- Maccari, S., P. V. Piazza, M. Kabbaj, A. Barbazanges, H. Simon, and M. Le Moal. "Adoption Reverses the Long-Term Impairment in Glucocorticoid Feedback Induced by Prenatal Stress." J Neurosci 15, no. 1 Pt 1 (1995): 110-6. [CrossRef]
- Smith, J. W., J. R. Seckl, A. T. Evans, B. Costall, and J. W. Smythe. "Gestational Stress Induces Post-Partum Depression-Like Behaviour and Alters Maternal Care in Rats." Psychoneuroendocrinology 29, no. 2 (2004): 227-44. [CrossRef]
- Misdrahi, D., M. C. Pardon, F. Perez-Diaz, N. Hanoun, and C. Cohen-Salmon. "Prepartum Chronic Ultramild Stress Increases Corticosterone and Estradiol Levels in Gestating Mice: Implications for Postpartum Depressive Disorders." Psychiatry Res 137, no. 1-2 (2005): 123-30. [CrossRef]
- Negroni, J., P. Venault, M. C. Pardon, F. Perez-Diaz, G. Chapouthier, and C. Cohen-Salmon. "Chronic Ultra-Mild Stress Improves Locomotor Performance of B6d2f1 Mice in a Motor Risk Situation." Behav Brain Res 155, no. 2 (2004): 265-73. [CrossRef]
- Pardon, M., F. Perez-Diaz, C. Joubert, and C. Cohen-Salmon. "Age-Dependent Effects of a Chronic Ultramild Stress Procedure on Open-Field Behaviour in B6d2f1 Female Mice." Physiol Behav 70, no. 1-2 (2000): 7-13. [CrossRef]
- Pardon, M. C., F. Perez-Diaz, C. Joubert, and C. Cohen-Salmon. "Influence of a Chronic Ultramild Stress Procedure on Decision-Making in Mice." J Psychiatry Neurosci 25, no. 2 (2000): 167-77.
- Champagne, F. A., J. P. Curley, E. B. Keverne, and P. P. Bateson. "Natural Variations in Postpartum Maternal Care in Inbred and Outbred Mice." Physiol Behav 91, no. 2-3 (2007): 325-34. [CrossRef]
- Curley, J. P., C. L. Jensen, B. Franks, and F. A. Champagne. "Variation in Maternal and Anxiety-Like Behavior Associated with Discrete Patterns of Oxytocin and Vasopressin 1a Receptor Density in the Lateral Septum." Horm Behav 61, no. 3 (2012): 454-61. [CrossRef]
- Maggio, John C, and Glayde Whitney. "Ultrasonic Vocalizing by Adult Female Mice (Mus Musculus)." Journal of Comparative Psychology 99, no. 4 (1985): 420.
- Whitney, G., J. R. Coble, M. D. Stockton, and E. F. Tilson. "Ultrasonic Emissions: Do They Facilitate Courtship of Mice." J Comp Physiol Psychol 84, no. 3 (1973): 445-52. [CrossRef]
- Giannoccaro, S., C. Ferraguto, V. Petroni, C. Marcelly, X. Nogues, V. Campuzano, and S. Pietropaolo. "Early Neurobehavioral Characterization of the Cd Mouse Model of Williams-Beuren Syndrome." Cells 12, no. 3 (2023). [CrossRef]
- Petroni, V., E. Subashi, M. Premoli, M. Memo, V. Lemaire, and S. Pietropaolo. "Long-Term Behavioral Effects of Prenatal Stress in the Fmr1-Knock-out Mouse Model for Fragile X Syndrome." Front Cell Neurosci 16 (2022): 917183. [CrossRef]
- Premoli, M., V. Petroni, R. Bulthuis, S. A. Bonini, and S. Pietropaolo. "Ultrasonic Vocalizations in Adult C57bl/6j Mice: The Role of Sex Differences and Repeated Testing." Front Behav Neurosci 16 (2022): 883353. [CrossRef]
- Kat, R., M. Arroyo-Araujo, R. B. M. de Vries, M. A. Koopmans, S. F. de Boer, and M. J. H. Kas. "Translational Validity and Methodological Underreporting in Animal Research: A Systematic Review and Meta-Analysis of the Fragile X Syndrome (Fmr1 Ko) Rodent Model." Neurosci Biobehav Rev 139 (2022): 104722. [CrossRef]
- Zupan, B., A. Sharma, A. Frazier, S. Klein, and M. Toth. "Programming Social Behavior by the Maternal Fragile X Protein." Genes Brain Behav 15, no. 6 (2016): 578-87. [CrossRef]
- Zupan, B., and M. Toth. "Wild-Type Male Offspring of Fmr-1+/- Mothers Exhibit Characteristics of the Fragile X Phenotype." Neuropsychopharmacology 33, no. 11 (2008): 2667-75. [CrossRef]
- ———. "Fmr-1 as an Offspring Genetic and a Maternal Environmental Factor in Neurodevelopmental Disease." Results Probl Cell Differ 54 (2012): 243-53. [CrossRef]
- Bowman, R., M. Frankfurt, and V. Luine. "Sex Differences in Cognition Following Variations in Endocrine Status." Learn Mem 29, no. 9 (2022): 234-45. [CrossRef]
- Bowman, R. E. "Stress-Induced Changes in Spatial Memory Are Sexually Differentiated and Vary across the Lifespan." J Neuroendocrinol 17, no. 8 (2005): 526-35. [CrossRef]
- Bowman, R. E., K. D. Beck, and V. N. Luine. "Chronic Stress Effects on Memory: Sex Differences in Performance and Monoaminergic Activity." Horm Behav 43, no. 1 (2003): 48-59. [CrossRef]
- Nielsen, D. M., J. J. Evans, W. J. Derber, K. A. Johnston, M. L. Laudenslager, L. S. Crnic, and K. N. Maclean. "Mouse Model of Fragile X Syndrome: Behavioral and Hormonal Response to Stressors." Behav Neurosci 123, no. 3 (2009): 677-86. [CrossRef]
- Qin, M., and C. B. Smith. "Unaltered Hormonal Response to Stress in a Mouse Model of Fragile X Syndrome." Psychoneuroendocrinology 33, no. 6 (2008): 883-9. [CrossRef]
- Sandi, C., and J. Haller. "Stress and the Social Brain: Behavioural Effects and Neurobiological Mechanisms." Nat Rev Neurosci 16, no. 5 (2015): 290-304. [CrossRef]
- Weinstock, M. "The Long-Term Behavioural Consequences of Prenatal Stress." Neurosci Biobehav Rev 32, no. 6 (2008): 1073-86. [CrossRef]
- Laviola, G., W. Adriani, S. Morley-Fletcher, and M. L. Terranova. "Peculiar Response of Adolescent Mice to Acute and Chronic Stress and to Amphetamine: Evidence of Sex Differences." Behav Brain Res 130, no. 1-2 (2002): 117-25. [CrossRef]
- Dallman, M. F., S. F. Akana, A. M. Strack, K. S. Scribner, N. Pecoraro, S. E. La Fleur, H. Houshyar, and F. Gomez. "Chronic Stress-Induced Effects of Corticosterone on Brain: Direct and Indirect." Ann N Y Acad Sci 1018 (2004): 141-50. [CrossRef]
- Nair, B. B., Z. Khant Aung, R. Porteous, M. Prescott, K. A. Glendining, D. E. Jenkins, R. A. Augustine, M. S. B. Silva, S. H. Yip, G. T. Bouwer, C. H. Brown, C. L. Jasoni, R. E. Campbell, S. J. Bunn, G. M. Anderson, D. R. Grattan, A. E. Herbison, and K. J. Iremonger. "Impact of Chronic Variable Stress on Neuroendocrine Hypothalamus and Pituitary in Male and Female C57bl/6j Mice." J Neuroendocrinol 33, no. 5 (2021): e12972. [CrossRef]
- Marchette, R. C. N., M. A. Bicca, Ecds Santos, and T. C. M. de Lima. "Distinctive Stress Sensitivity and Anxiety-Like Behavior in Female Mice: Strain Differences Matter." Neurobiol Stress 9 (2018): 55-63. [CrossRef]
- Safari, M. A., M. Koushkie Jahromi, R. Rezaei, H. Aligholi, and S. Brand. "The Effect of Swimming on Anxiety-Like Behaviors and Corticosterone in Stressed and Unstressed Rats." Int J Environ Res Public Health 17, no. 18 (2020). [CrossRef]
- Lupien, S. J., B. S. McEwen, M. R. Gunnar, and C. Heim. "Effects of Stress Throughout the Lifespan on the Brain, Behaviour and Cognition." Nat Rev Neurosci 10, no. 6 (2009): 434-45. [CrossRef]
- Juruena, M. F., F. Eror, A. J. Cleare, and A. H. Young. "The Role of Early Life Stress in Hpa Axis and Anxiety." Adv Exp Med Biol 1191 (2020): 141-53. [CrossRef]
- Pawluski, J. L., K. G. Lambert, and C. H. Kinsley. "Neuroplasticity in the Maternal Hippocampus: Relation to Cognition and Effects of Repeated Stress." Horm Behav 77 (2016): 86-97. [CrossRef]
- Pietropaolo, S., A. Guilleminot, B. Martin, F. R. D'Amato, and W. E. Crusio. "Genetic-Background Modulation of Core and Variable Autistic-Like Symptoms in Fmr1 Knock-out Mice." PLoS ONE 6, no. 2 (2011): e17073. [CrossRef]
- Burkholder, T., C. Foltz, E. Karlsson, C. G. Linton, and J. M. Smith. "Health Evaluation of Experimental Laboratory Mice." Curr Protoc Mouse Biol 2 (2012): 145-65. [CrossRef]
- Pietropaolo, S., and W. E. Crusio. "Strain-Dependent Changes in Acoustic Startle Response and Its Plasticity across Adolescence in Mice." Behav Genet (2009). [CrossRef]
- Pietropaolo, S., M. G. Goubran, C. Joffre, A. Aubert, V. Lemaire-Mayo, W. E. Crusio, and S. Laye. "Dietary Supplementation of Omega-3 Fatty Acids Rescues Fragile X Phenotypes in Fmr1-Ko Mice." Psychoneuroendocrinology 49 (2014): 119-29. [CrossRef]
- Moles, A., and R. D'Amato F. "Ultrasonic Vocalization by Female Mice in the Presence of a Conspecific Carrying Food Cues." Anim Behav 60, no. 5 (2000): 689-94. [CrossRef]
- Moles, A., F. Costantini, L. Garbugino, C. Zanettini, and F. R. D'Amato. "Ultrasonic Vocalizations Emitted During Dyadic Interactions in Female Mice: A Possible Index of Sociability?" Behav Brain Res 182, no. 2 (2007): 223-30. [CrossRef]
- Gaudissard, J., M. Ginger, M. Premoli, M. Memo, A. Frick, and S. Pietropaolo. "Behavioral Abnormalities in the Fmr1-Ko2 Mouse Model of Fragile X Syndrome: The Relevance of Early Life Phases." Autism Res 10, no. 10 (2017): 1584-96. [CrossRef]
- Maggio, J. C., and G. Whitney. "Ultrasonic Vocalizing by Adult Female Mice (Mus Musculus)." J Comp Psychol 99, no. 4 (1985): 420-36.
- D'Amato, F. R., and A. Moles. "Ultrasonic Vocalizations as an Index of Social Memory in Female Mice." Behav Neurosci 115, no. 4 (2001): 834-40. [CrossRef]
- Premoli, M., S. A. Bonini, A. Mastinu, G. Maccarinelli, F. Aria, G. Paiardi, and M. Memo. "Specific Profile of Ultrasonic Communication in a Mouse Model of Neurodevelopmental Disorders." Sci Rep 9, no. 1 (2019): 15912. [CrossRef]
- Scattoni, M. L., L. Ricceri, and J. N. Crawley. "Unusual Repertoire of Vocalizations in Adult Btbr T+Tf/J Mice During Three Types of Social Encounters." Genes Brain Behav 10, no. 1 (2011): 44-56. [CrossRef]
- Caruso, A., L. Ricceri, and M. L. Scattoni. "Ultrasonic Vocalizations as a Fundamental Tool for Early and Adult Behavioral Phenotyping of Autism Spectrum Disorder Rodent Models." Neurosci Biobehav Rev 116 (2020): 31-43. [CrossRef]
- Caligioni, C. S. "Assessing Reproductive Status/Stages in Mice." Curr Protoc Neurosci Appendix 4 (2009): Appendix 4I. [CrossRef]
- Vandesquille, M., M. Baudonnat, L. Decorte, C. Louis, P. Lestage, and D. Beracochea. "Working Memory Deficits and Related Disinhibition of the Camp/Pka/Creb Are Alleviated by Prefrontal Alpha4beta2*-Nachrs Stimulation in Aged Mice." Neurobiol Aging 34, no. 6 (2013): 1599-609. [CrossRef]
Figure 1.
Maternal behaviors in primiparous dams. The maternal behavior of all females of the primiparous group (n=8-11 for each genotype x stress condition, as detailed in A) was observed in their home-cages twice a day for one hour (at 9.00 AM and at 5.00 PM) from PND 1 to PND 6, using an instantaneous sampling method (one sampling/2min). The following items were scored as absolute frequencies by an observer who was blind to the experimental conditions of the breeders [26,49,50]: (i) nursing postures, including arched-back nursing (the female is in an arched position over the nursing pups, A) and blanket (the female is lying flat on top of the pups, B), (ii) licking/grooming of the pups (C), (iii) non-nursing postures (the female is in contact with the pups, but not nursing, i.e., with no access to the nipples, D). All behaviors were summed up across the 6 days of scoring. Data are expressed as mean±SEM. * p<0.05.
Figure 1.
Maternal behaviors in primiparous dams. The maternal behavior of all females of the primiparous group (n=8-11 for each genotype x stress condition, as detailed in A) was observed in their home-cages twice a day for one hour (at 9.00 AM and at 5.00 PM) from PND 1 to PND 6, using an instantaneous sampling method (one sampling/2min). The following items were scored as absolute frequencies by an observer who was blind to the experimental conditions of the breeders [26,49,50]: (i) nursing postures, including arched-back nursing (the female is in an arched position over the nursing pups, A) and blanket (the female is lying flat on top of the pups, B), (ii) licking/grooming of the pups (C), (iii) non-nursing postures (the female is in contact with the pups, but not nursing, i.e., with no access to the nipples, D). All behaviors were summed up across the 6 days of scoring. Data are expressed as mean±SEM. * p<0.05.
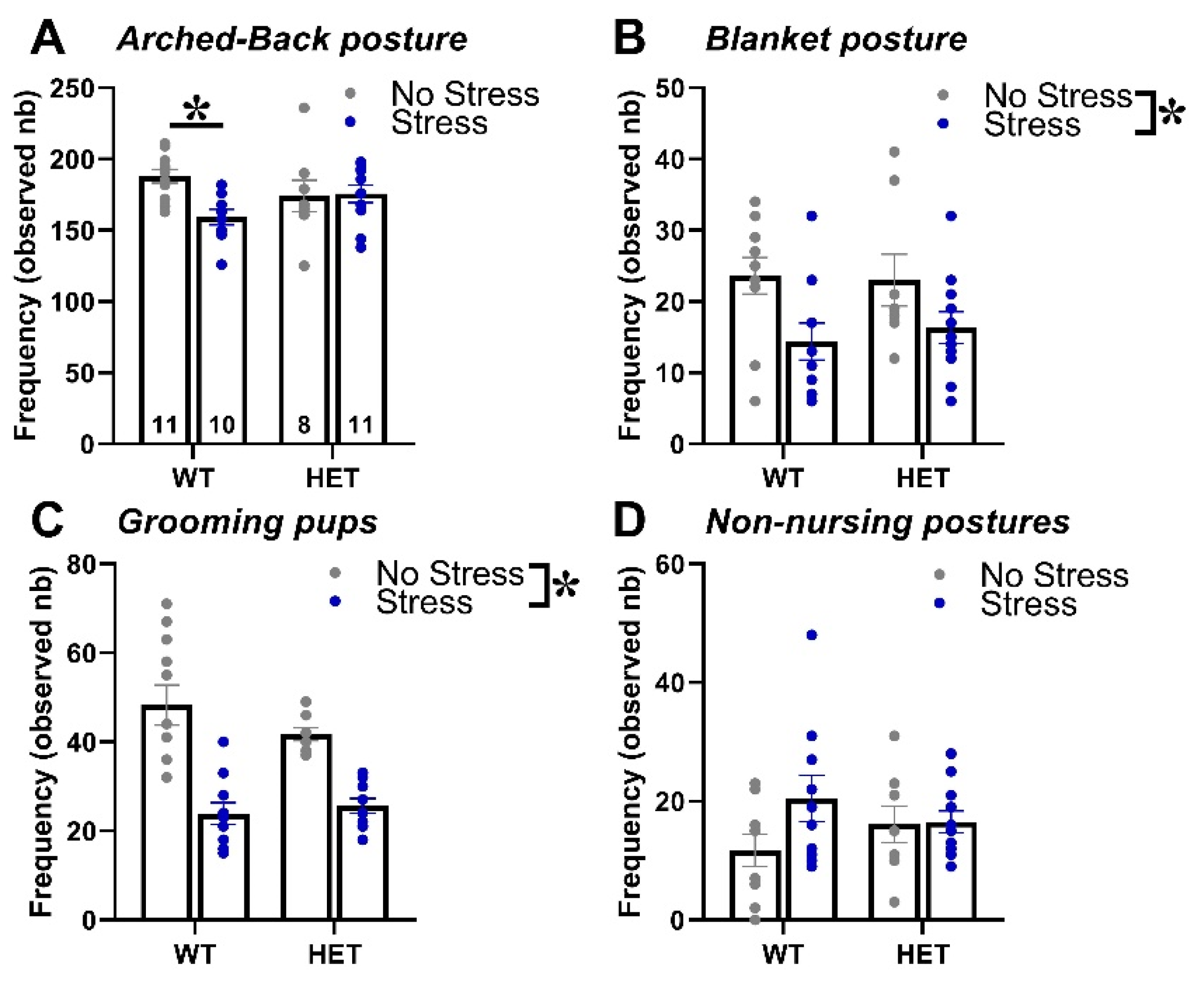
Figure 2.
Anxiety levels, exploratory behaviors and spontaneous alternation. Anxiety levels were investigated in the elevated plus maze test (A) and the open field test (B), where locomotion was also assessed (C). Spontaneous alternation was evaluated in the Y maze (D). * p<0.05; # p<0.05 versus chance level (indicated by the dotted line). N for each group is indicated in C. Data are expressed as mean±SEM.
Figure 2.
Anxiety levels, exploratory behaviors and spontaneous alternation. Anxiety levels were investigated in the elevated plus maze test (A) and the open field test (B), where locomotion was also assessed (C). Spontaneous alternation was evaluated in the Y maze (D). * p<0.05; # p<0.05 versus chance level (indicated by the dotted line). N for each group is indicated in C. Data are expressed as mean±SEM.
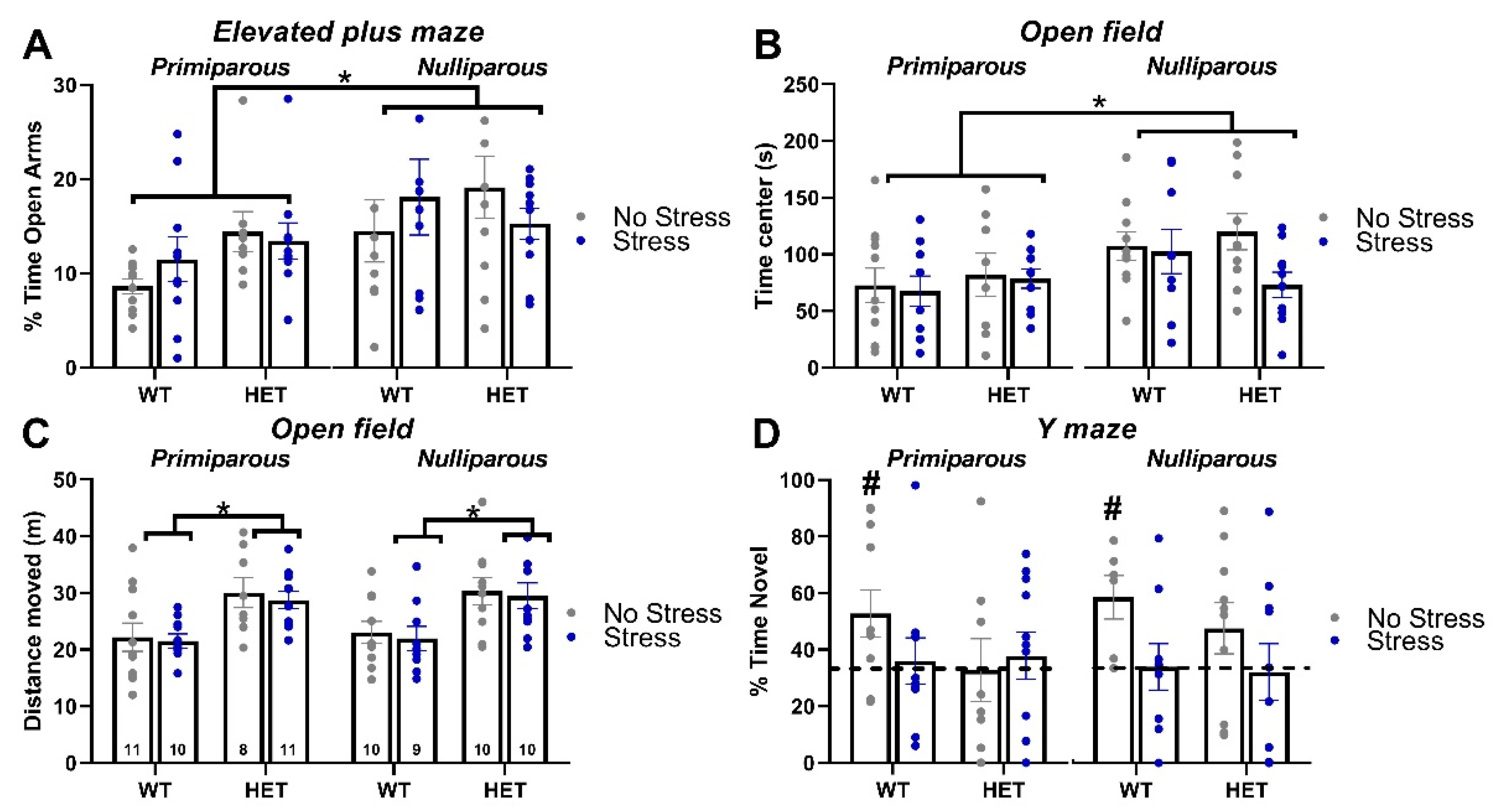
Figure 3.
Quantitative analysis of the social interaction and ultrasonic vocalizations (USVs) emitted during the social interaction test. Social interaction was measured during a 3-min encounter with an adult NMRI WT female, computing the time spent in affiliative behaviors towards the stimulus (A). USVs were recorded during the direct social interaction test. Female experimental subjects were single housed in the testing cage for 72 hs prior to testing; an unfamiliar adult stimulus female mouse was then introduced and left there for 3 min. Previous studies have shown that in these experimental settings USVs are emitted only by the female intruder [51,52]. The number (B) and mean duration (C) of USVs were automatically measured using the software Sonotrack. Data are expressed as mean±SEM. * p<0.05.
Figure 3.
Quantitative analysis of the social interaction and ultrasonic vocalizations (USVs) emitted during the social interaction test. Social interaction was measured during a 3-min encounter with an adult NMRI WT female, computing the time spent in affiliative behaviors towards the stimulus (A). USVs were recorded during the direct social interaction test. Female experimental subjects were single housed in the testing cage for 72 hs prior to testing; an unfamiliar adult stimulus female mouse was then introduced and left there for 3 min. Previous studies have shown that in these experimental settings USVs are emitted only by the female intruder [51,52]. The number (B) and mean duration (C) of USVs were automatically measured using the software Sonotrack. Data are expressed as mean±SEM. * p<0.05.
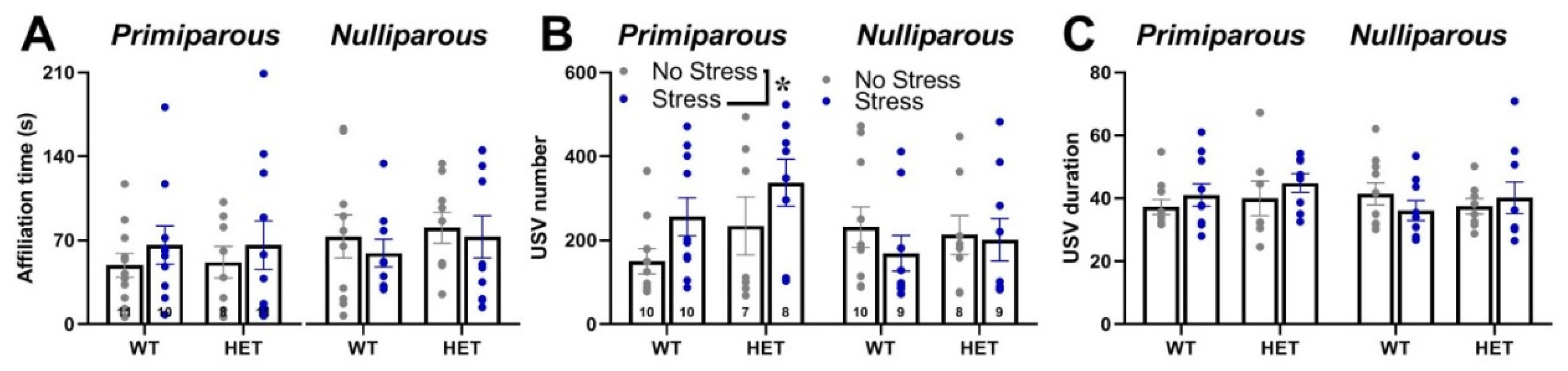
Figure 4.
Qualitative analysis of the ultrasonic vocalizations (USVs) emitted during the social interaction test. Ultrasonic calls were automatically categorized as described before [22,53,54,55]. The “complex tot” category included all complex calls with more than 3 components. Data are expressed as mean percentages over the total number of USVs for each group.
Figure 4.
Qualitative analysis of the ultrasonic vocalizations (USVs) emitted during the social interaction test. Ultrasonic calls were automatically categorized as described before [22,53,54,55]. The “complex tot” category included all complex calls with more than 3 components. Data are expressed as mean percentages over the total number of USVs for each group.
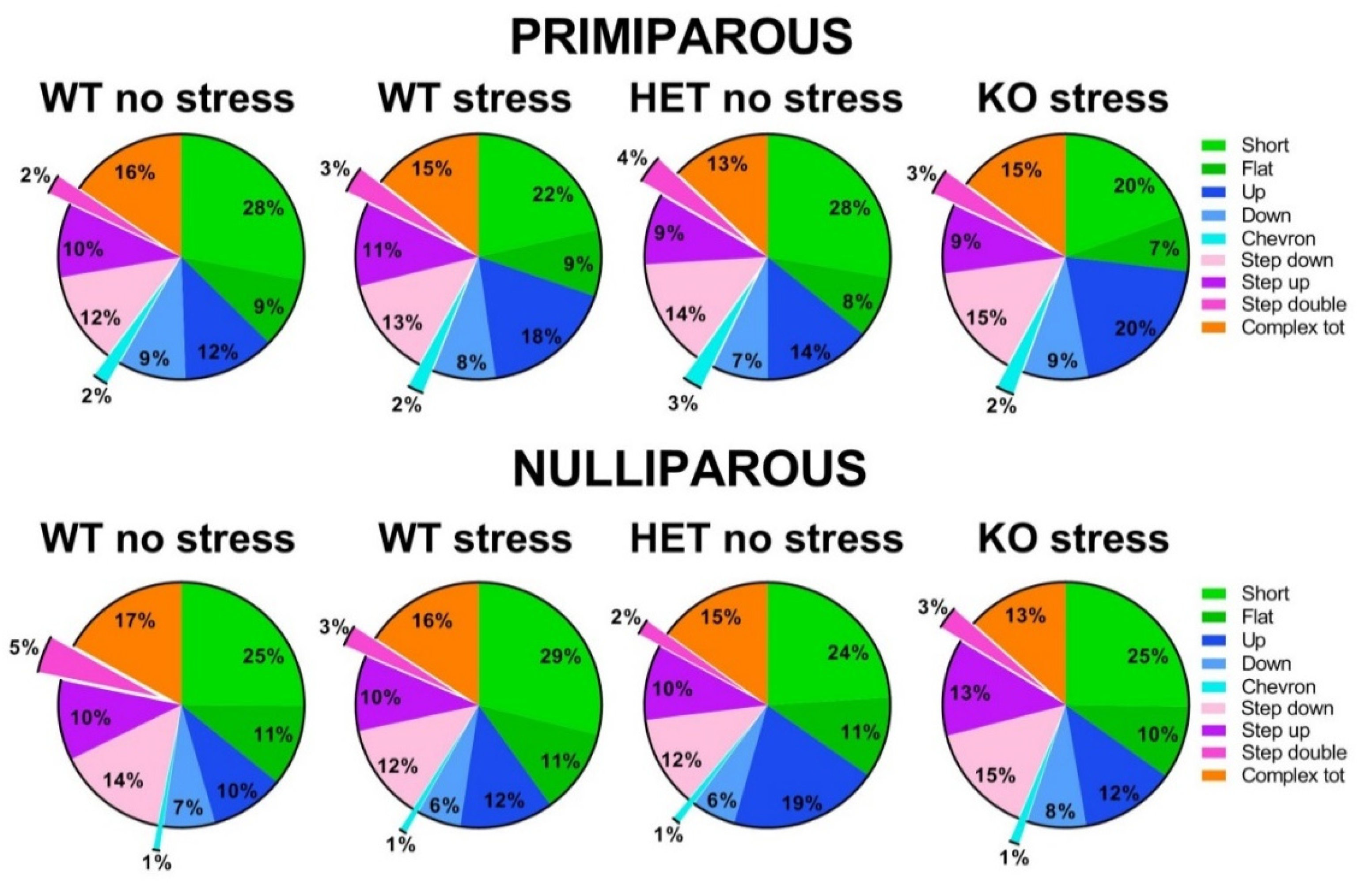
Figure 5.
Basal corticosterone blood levels and adrenal gland weight. Basal corticosterone levels (A) and adrenal gland weight (B) were assessed one week after the end of behavioral testing, i.e., when mice were euthanatized (see also Figure 1). * p<0.05; N= 8-11 as indicated in (B). Data are expressed as mean±SEM.
Figure 5.
Basal corticosterone blood levels and adrenal gland weight. Basal corticosterone levels (A) and adrenal gland weight (B) were assessed one week after the end of behavioral testing, i.e., when mice were euthanatized (see also Figure 1). * p<0.05; N= 8-11 as indicated in (B). Data are expressed as mean±SEM.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



