
Submitted:
23 May 2023
Posted:
25 May 2023
You are already at the latest version
Abstract
The taxonomic identity and phylogenetic relationships of several southern African perennial taxa related to Frankenia repens are discussed. In particular, F. nodiflora Lam., a misunderstood species described from the Cape region and synonymised to F. pulverulenta, is restored for plants endemic to salt-pans and riverbeds in the coastal lowlands across the Cape plains (Western Cape Province, South Africa). Further, a revision of morphologically close plants, usually identified as F. pulverulenta or F. repens, also occurring in similar saline ecosystems of the inland western South Africa revealed the existence of two distinct new entities not matching any described taxa of the genus. Molecular analyses of nuclear ribosomal (ITS1-5.8S-ITS2 region) DNA sequence data together with morphological divergence allow recognition of those taxa at species rank, within an independent lineage close to F. repens. In consequence, two new species are described in the so-called “F. repens group”: F. nummularia from the Nama-Karoo Biome (Western Cape and Northern Cape Provinces), and F. namaquana from the Succulent Karoo Biome (Northern Cape Province). Full morphological description and type designation are reported for each accepted species, as well as data on ecology, habitat, distribution and taxonomic relationships to other close relatives are given. Further, an identification key is presented to facilitate recognition of the southern African taxa of Frankenia.
Keywords:
Frankenia
; Frankeniaceae
; nomenclature
; ITS phylogeny
; plant endemics
; plant morphology
; southern African flora
; taxonomy
1. Introduction
Frankeniaceae Desv., nom. cons., is a family of mostly halophytic herbs and shrubs, which has been usually accepted to include two to five genera [1,2,3,4]. However, available molecular phylogenies [5–7, and references therein] recovered members of all those genera embedded in a single clade among species of Frankenia L., and therefore the latter is currently accepted to be the only genus in the family.
So treated, Frankenia s.l. includes about 80–90 species, mostly occurring in saline, gypsum- and carbonate-rich substrates in Mediterranean-type regions around the world, but with ca. 40 species found in Australia [8,9]. Members of that genus often occur in coastal and inland saltmarshes, commonly experiencing semi-arid to arid climates, usually having prevalent winter rainfall [10].
In southern Africa, four species are currently recognised occurring [10,11]. Three of them, Frankenia repens (P.J.Bergius) Fourc., F. pomonensis Pohnert and F. fruticosa J.C.Manning & Helme, are woody perennial or suffruticose endemics. The two former taxa occur in salt-marshes of South Africa and/or Namibia, while the latter is a very rare dwarf shrub limited exclusively to quartz outcrops of southern Knersvlakte in Namaqualand (South Africa). The fourth species, F. pulverulenta L., is a subcosmopolitan annual herb, commonly linked to human disturbance, and it is also widespread in most of South Africa.
Recent taxonomic research conducted in the southern part of Africa revealed the existence of peculiar suffruticose plants occurring on saline soils of the Northern Cape and Western Cape regions (South Africa), clearly differing from other species of the genus. These plants were usually identified as Frankenia capitata Webb & Berth. or F. repens (incl. F. kreibsii Cham. & Schltdl.), but more often as F. pulverulenta L. (subsp. pulverulenta), a name having F. nodiflora Lam. in synonymy [4,12] https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:77229962-1. However, Lamarck [13] described his F. nodiflora based on material gathered in an undetermined place in the lowlands surrounding Cape Town (South Africa), and it was said to be distinct from the Linnaean species. In fact, a close examination of Lamarck’s original material reveals a unique combination of morphological characters missing in any of the known South African taxa of the genus. Furthermore, plants akin to F. nodiflora occurring in the inland dry areas of Namaqualand and Karoo (western South Africa) also exhibit morphological peculiarities not fitting with the latter which allow easy recognition when compared to other South African congeners.
In the present contribution, the name Frankenia nodiflora is therefore restored for a misunderstood species, endemic to the Cape Town lowlands classified in the Fynbos (F) biome. Also, two new related species are described from the Nama-Karoo (NK) and Succulent-Karoo (SK) biomes [14] in western South Africa. These are named here F. nummularia and F. namaquana. Morphological, distributional, ecological and molecular-phylogenetic data are reported supporting their recognition at specific rank. Types are also designated for the concerned taxa, and a tentative taxonomic key is presented for identification of the known southern African species of Frankenia.
2. Materials and Methods
2.1. Morphological and habitat studies
Detailed morphological studies were undertaken on both living plants from wild populations and dried herbarium specimens sourced from the herbaria ABH, BOL, G-DC, K, M, HBG, NBG, LE, LINN, M, MW, P, PRE, SBT, W and WU (acronyms according to Thiers [15]), using an OLYMPUS SZX7 binocular. Digital images of Frankenia from iNaturalist (https://www.inaturalist.org/observations/?place_id=any&taxon_id=58170) were also checked and considered to fulfil distributions of the concerned species. Scanning Electron Microscope (SEM) micrographs of seeds were taken with a JEOL JSM-IT500HR operating at 15 kV. No special treatment of the material was required prior to observation. At least 10 mature seeds from different individuals of the studied species (Table 1) were observed in detail. Samples were glued directly on metallic stubs and then coated with 10 nm platinum in a QUORUM Q150T ES Plus sputter coater. The ImageJ software [16] was used for measurements on SEM micrographs.

Table 1.
Seed samples of Frankenia for SEM studies, with provenance and herbarium vouchers.
| Taxon | Locality | Herbarium voucher |
|---|---|---|
| F. nodiflora | South Africa: Cape Town | P05038804 |
| F. nummularia | South Africa: Kookfontein | ABH83290 |
| F. namaquana | South Africa: Klipfontein | ABH76891 |
| F. repens | South Africa: Port Nolloth | ABH76882 |
Authors of the taxa cited in the text follow IPNI [17]. Nomenclatural issues follow Turland et al [18], and orthography of geographical names and grid-number system accord with Leistner and Morris [19], and the grid-number system follows the National Geospatial Information (2020). Bioclimate, bioregion, and vegetation classification agrees with Rutherford and Mucina [14].
2.2. Molecular analyses
Herbarium vouchers and silica gel-dried material were used for total DNA extraction employing a modified 2 × cetyltrimethylammonium bromide (CTAB) protocol [20]. Total DNA was purified using MOBIO minicolumns and mostly kept in 0.1 × TE buffer [10 mM Tris-HCl, 1 mM ethylenediaminetetraacetic acid (EDTA), pH 8.0]. The internal transcribed spacer –ITS– region (ITS1 spacer, 5.8S gene, ITS2 spacer) of nuclear ribosomal DNA (nrDNA) was amplified using the ITS5 and ITS4 primers [21]. Amplifications were performed on a reaction volume of 25 μl containing 22 μl of ABGene 1.1 × Master Mix, 2.5 mM MgCl2 (Thermo Scientific Waltham, MA, USA), 0.5 μl of 0.4% bovine serum albumin (BSA), 0.5 μl of dimethyl sulfoxide (DMSO), 0.5 μl of each primer (10 pmol/μl) and 1 μl of template DNA on a 9700 GeneAmpl thermocycler (Applied Biosystems). The PCR program for ITS was: 2 min at 95 °C, followed by 30 cycles of 95 °C for 1 min, 53 °C for 1 min, 72 °C for 2 min and a final extension of 72 °C for 5 min.
Sequencer 4.1 (Gene Codes Corp., Ann Arbor, MI, USA) was used to assemble complementary strands and verify software base-calling. Sequence alignment was performed using MUSCLE [22] conducted in MEGA X v.10.2.6 [23] with minor manual adjustments to get the final aligned matrix. Twenty-four ITS sequence data belonging to 15 species of Frankenia, which cover most of the African and Mediterranean groups accepted in the genus, were used in the analyses, including as outgroups GenBank accessions of Tamarix gallica L. (code MH626294), Myricaria germanica L. (code KJ808607) and Reaumuria alternifolia (Labill.) Britten (code KJ729627); it is worth mentioning that the name R. alternifolia is used here instead of its superfluous synonym R. hypericoides Willd. that is applied in GenBank to the sequence used. All Frankenia accessions were obtained specifically for this study (Table 2).
Phylogenetic analyses of the ITS region were obtained using Maximum Parsimony (MP), Maximum Likelihood (ML) and Neighbour Joining (NJ) methods. MP analysis was conducted in both PAUP (using Heuristic search options with the tree searching strategy based on Nearest Neighbour Interchange, NNI) and MEGA (using Heuristic search options with the tree searching strategy based on Subtree-Pruning-Regrafting –SPR– with search level 1 [24]) for result comparison, with 10,000 replicates. ML [25] and NJ [26] analyses were also performed in MEGA, as well as the selection of the best model of DNA substitutions for each method using the Akaike Information Criterion (AIC; [27]); models with the lowest BIC (Bayesian Information Criterion) scores were considered to best describe the substitution pattern for the ML and NJ analyses. Phylogenetic reconstructions for ML and evolutionary distances for NJ were estimated using the K2 model (2-parameter method of Kimura [28]) and considering all sites; the rate variation model allowed for some sites to be evolutionarily invariable (+I, 29.80% sites). For comparison purposes, remotion of all ambiguous positions for each sequence pair (Pairwise Deletion option) was also performed and no significant differences (only affecting BP values in a few branches) were observed in the obtained phylogenies.
Furthermore, Bayesian inference (BI) analyses were conducted with MrBayes 3.2 [29], in which the Markov Chain Monte Carlo (MCMC) algorithm was run for 10 million generations and sampled every 1000 generations. Two runs were performed. The general time reversible (GTR) + proportion of invariant sites (I) + gamma distribution (G) model was used in the analyses (set nst = 6 rates = invgamma), according to the results obtained with jModelTest 2.1.10 [30] under AIC. The first 25% generations (burninfrac = 0.25) were excluded, and the remaining trees were used to compile a posterior probability (PP) distribution using a 50% majority-rule consensus. For all methods, support was assessed by the bootstrap [31] with 10,000 replicates but holding only 10 trees per replicate. Clades showing bootstrap percentage (BP) values of 50–74% were considered as weakly supported, 75–89% moderately supported and 90–100% strongly supported.
Table 2.
List of outgroups and Frankenia accessions used for the ITS phylogenetic analyses.
| Taxon | Provenance (herbarium voucher) | Source | GenBank accession |
|---|---|---|---|
| Tamarix gallica L. | France: Saintes Maries de la Mer (ABH57865) | Villar et al. [32] | MH626294 |
| Myricaria germanica L. | Kazakhstan: Zajsanskaya depression (LE) | Zhang et al. [33] | KJ808607 |
| Reaumuria alternifolia (Labill.) Britten1 | Azerbaijan: Caucasus (MW) | Zhang et al. [34] | KJ729627 |
| Frankenia boissieri Reut. ex Boiss. | Spain: Huelva, Ayamonte, Is. Canela (ABH83543) | This paper | |
| F. capitata Webb & Berthel. | Spain: Canary Is., Lanzarote, Yaiza (ABH57588) | This paper | |
| F. cf. chevalieri Maire | Morocco: Sidi Ifni to Oued Noun (MA758515) | This paper | |
| F. composita Pau & Font Quer | Morocco: Al Hoceïma, Cala Iris (ABH81590) | This paper | |
| F. corymbosa Desf. | Spain: Alicante, Santa Pola (ABH79956) | This paper | |
| Morocco: Nador, Punta Charrana (ABH54294) | This paper | ||
| Morocco: Al-Hoceïma (ABH54526) | This paper | ||
|
F. ericifolia C.Sm. ex DC, nom. cons. prop. |
Spain: Canary Isl., Tenerife (ABH79975) | This paper | |
| Spain: Canary Is., Lanzarote, Tinajo (ABH57566) | This paper | ||
| F. hirsuta L. | Turkyie: Tuz Gölii, salty lagoon (ABH45933) | This paper | |
| F. laevis L. | Italy: Sardinia (ABH70072) | This paper | |
| France: Corsica (ABH70584) | This paper | ||
| F. namaquana M.B.Crespo & al. | South Africa: Klipfontein (ABH76891) | This paper | |
| South Africa: Skoverfontein (ABH83196) | This paper | ||
| F. nummularia M.B.Crespo & al. | South Africa: Kookfontein River (ABH83290) | This paper | |
| South Africa: Tankwa Karoo (ABH83295) | This paper | ||
| F. pseudoericifolia Rivas Mart. & al. | Portugal: Cape Verde, São Antão (MA0906845) | This paper | |
| F. pulverulenta L. | South Africa: Redelinghuis (ABH77205) | This paper | |
| South Africa: Skoverfontein (ABH83195) | This paper | ||
| F. repens (P.J.Bergius) Fourc. | South Africa: S of Hondeklipbaai (ABH76862) | This paper | |
| South Africa: S of Groenrivier (ABH76868) | This paper | ||
| F. thymifolia Desf. | Algeria: Bougtob, Chott Cherguí (ABH59344) | This paper | |
| Spain: Zaragoza: Bujaraloz (ABH75454) | This paper | ||
| F. velutina Brouss. ex DC. | Morocco: Essaouira (ABH79929) | This paper |
1 Filed in GenBank as the synonym name R. hypericoides Willd.
3. Results
3.1. A brief story and typification of Frankenia nodiflora
Lamarck (1788) included four species in his treatment of Frankenia for the Encyclopaedia. Among them, he described Frankenia nodiflora (numbered 3) as new, after F. hirsuta L. (numbered 2) and before F. pulverulenta L. (numbered 4). No syntypes or illustrations were cited in the protologue, though the new species was described from dried material as follows: “3. FRANQUENNE nodiflore, Frankenia nodiflora, Frankenia caulibus simplicibus filiformibus subglabris, fasciculis florum lateralibus axillaribus & oppositis. N[obis]. / Ses tiges sont longues de six à huit pouces, simples ou presque simples, filiformes, feuillées, & presque glabres. Leurs feuilles sont opposées, pétiolées, ovales, glabres, à bords réfléchis en dessous, & longues de deux lignes & demie. Dans leurs aisselles, on observe sur toute la longueur des tiges, des rameaux non développés, opposes, plus courts que les entre-noeuds, feuillés & fleuris, & qui sont paroître les tiges entrecoupées dans leur longueur par des touffes ou paquets de feuilles & de fleurs biens sépares les uns des autres. Les fleurs ont leur calice oblong, légèrement anguleux, glabre, & naissent comme en faisceau entre les feuilles, aux noeuds des tiges. / Cette plante croît naturellement au Cap de Bonne-Espérance, & a l’aspect d’une espèce de Salicaire. (v. s.).” Furthermore, in the comment for F. hirsuta he clearly indicated the origin and collector of the material used for the description of F. nodiflora: “La plante β [of the intended “F. hirsuta”, probably corresponding to F. repens] a ses tiges presqu’entièrement glabres, ainsi que ses calices; elle croît au Cap de Bonne-Espérance, & nous a été communiquée avec la suivante [F. nodiflora] par M. Sonnerat.” In fact, the French naturalist and explorer Pierre Sonnerat (1748−1814) gathered that material most probably in the surroundings of Cape Town during his travels to the East Indies and China in 1774−1781. The results of those expeditions were published later in a two-volume work [35], which includes observations on the Cape area in the second volume that covers his visits to Cape Town, Madagascar, the Maldives, Mauritius, Ceylon (Sri Lanka), Indonesia, Burma, China, and the Philippines.
Frankenia nodiflora was later depicted in Tab. 262 of Lamarck’s Tableau [36], though the details are poor (Figure 1). He also added a brief diagnosis in French and Latin languages, with slight changes with regard to the protologue in the branching pattern of the stem: “4. FRANQUENNE nodiflore. Dict. n° 3. / F. tiges simples, filiformes, presque glabres; fascicules des fleurs latéraux, axillaires et opposés. [FRANKENIA nodiflora. / F. caulibus simplicibus, filiformibus, subglabris; fasciculis florum lateralibus, axillaribus et oppositis.].”
Details in the protologue of F. nodiflora on the length and branching pattern of the type material (“Ses tiges sont longues de six à huit pouces, simples ou presque simples…”) point out to the existence of various vouchers on which the description was prepared. In the herbarium P, we have traced two specimens matching the protologue, which are relevant for typification. First, the voucher P00287094 in Lamarck’s herbarium is labelled “Frankenia nodiflora Lam. / dict. / e cap[ut]. b[onae]. Sp[ei].” by Lamarck himself and includes a single unbranched fragment of ca. 15 cm long (ca. 6 inches). Secondly, the voucher P05038792, which is marked as part of Maire’s herbarium, bears a label reading “Frankenia nodiflora Lam. / (ego.) / Cap de B[onne]. Espér[ance]” in a calligraphy not incompatible with Lamarck’s handwriting and includes two unequal fragments: one of ca. 19 cm long (ca. 7.6 inches) with a short lateral branch in the upper part, similar to that illustrated in Lamarck [36], and the other of ca. 4 cm long (ca. 1.6 inches). Both specimens bear fragments very similar to each other, more likely coming from a single collection, and therefore they might be regarded as duplicates belonging to original material of that name. Because the specimen P00287094 is in Lamarck’s collection, it is designated below as lectotype for F. nodiflora; the specimen P05038792, despite some doubts about the handwriting on its label being Lamarck’s (C. Aupic pers. comm.) and why it was placed among Maire’s material, is here regarded as isolectotype.
Among the diagnostic characters in the protologue summarised by Lamarck [13,36], the long internodes, the condensed glomerular inflorescences, the ovate petiolate and entirely glabrous leaves, and the angulose and glabrous calyces are differential for F. nodiflora. The revision of herbarium material at K and P revealed the existence of plants occurring in the surroundings of Cape Town, which perfectly match Lamarck’s type material. They are perennial shrublets producing suffruticose procumbent, non-rooting stems with long internodes; leaves entirely glabrous, mostly concolorous (bright green on both sides), broadly elliptic to oblong, flattened and only folded downwards on margins (at least on the upper third), mostly falcate upwards, subacute to minutely mucronate at apex, fleshy, with conspicuous petiole 0.5–0.8 mm long, sometimes glabrous; bracteoles broad and flat, about half to two thirds the length of the calyx; flowers mostly disposed in crowded dichasial glomerules on lateral short branches; calyxes often glabrous, with teeth cucullate bearing a notable subapical mucro ca. 0.5 mm long (diagnostic character not present in other African taxa of the genus); and seeds ca. 1 mm long, covered with unequal medium-sized papillae 12–33 μm long, globose to conical-obtuse, more densely disposed on the distal part, among other characters.
Frankenia nodiflora has been treated in quite different ways. Candolle [37] accepted it as a distinct species, but Harvey [38], who did not know the Lamarck’s specimens, suggested its probable inclusion in F. pulverulenta perhaps as a variety; in the same work he surprisingly also regarded the South African “F. pulverulenta & F. nodiflora, of Drège’s Coll.”, which included gatherings of the true F. nodiflora (conserved in different European herbaria; see below), to belong to F. pulverulenta. Similarly, F. nodiflora was often considered to be merely a synonym of F. pulverulenta [1,4,39], and subsequently recent works did not accept the Lamarck’s species in the Southern African floras [10,11,40,41], probably assuming implicit synonymisation with F. pulverulenta. That synonymic treatment is currently accepted in POWO [12] under the name F. pulverulenta subsp. pulverulenta (https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:77229962-1#synonyms).
Figure 1.
Relevant material of F. nodiflora Lam. (a) Lectotype here designated from Lamarck’s collection (P00287094) with a close-up detail of inflorescences (scale bar: 5 mm); reproduced with permission (© Muséum National d’Histoire Naturelle, Herbarium, Paris); (b) Comparative illustrations of F. pulverulenta (left; num. 3) and F. nodiflora (right; num. 4), according to Tableau encyclopédique et méthodique of Lamarck [13], plate 262 [partially modified]).
Figure 1.
Relevant material of F. nodiflora Lam. (a) Lectotype here designated from Lamarck’s collection (P00287094) with a close-up detail of inflorescences (scale bar: 5 mm); reproduced with permission (© Muséum National d’Histoire Naturelle, Herbarium, Paris); (b) Comparative illustrations of F. pulverulenta (left; num. 3) and F. nodiflora (right; num. 4), according to Tableau encyclopédique et méthodique of Lamarck [13], plate 262 [partially modified]).

Nevertheless, in our view the Cape plants matching the Lamarckian concept of F. nodiflora are morphologically quite distinct from the Linnaean F. pulverulenta, showing a unique combination of characters unknown in the other Southern African species of Frankenia, this supporting acceptance at specific rank. Other similar populations growing in the northern and western inland areas of the Nama-Karoo and Succulent-Karoo biomes that show some resemblances to F. nodiflora and were often misidentified as F. pulverulenta, F. capitata or F. repens (incl. F. krebsii), are here re-evaluated in the light of new morphological and molecular data.
3.2. Phylogenetic relationships
The aligned ITS dataset was 765 bp, 168 of which (21.96%) were potentially parsimony informative. Analyses of this dataset, using NJ, MP, ML, and BI methods, yielded trees with similar topologies and similar bootstrap and branch length values. In the MP analysis (not shown), seven most parsimonious trees were obtained with a tree length (TL) of 477 steps, a consistency index (CI) of 0.781 and a retention index (RI) of 0.859. The phylogenetic relationships of taxa are shown in Figure 2 as recovered in our ML tree (after 10,000 replicates), in which PP values (from the BI analysis) are placed above branches, whereas BP values (from the ML/MP analyses) below branches.
Figure 2.
Maximum Likelihood (ML) phylogenetic tree of Frankenia accessions from ITS nuclear DNA sequences, in which the position of taxa in the Frankenia repens group are highlighted in Clade A. Numbers above branches indicate posterior probabilities (PP) from the Bayesian analysis; numbers below branches indicate bootstrap percentage (BP) values obtained in both the Maximum Likelihood (ML) tree and the Maximum Parsimony (MP) strict consensus tree (50% majority-rule). GenBank codes are shown after each taxon/accession name.
Figure 2.
Maximum Likelihood (ML) phylogenetic tree of Frankenia accessions from ITS nuclear DNA sequences, in which the position of taxa in the Frankenia repens group are highlighted in Clade A. Numbers above branches indicate posterior probabilities (PP) from the Bayesian analysis; numbers below branches indicate bootstrap percentage (BP) values obtained in both the Maximum Likelihood (ML) tree and the Maximum Parsimony (MP) strict consensus tree (50% majority-rule). GenBank codes are shown after each taxon/accession name.
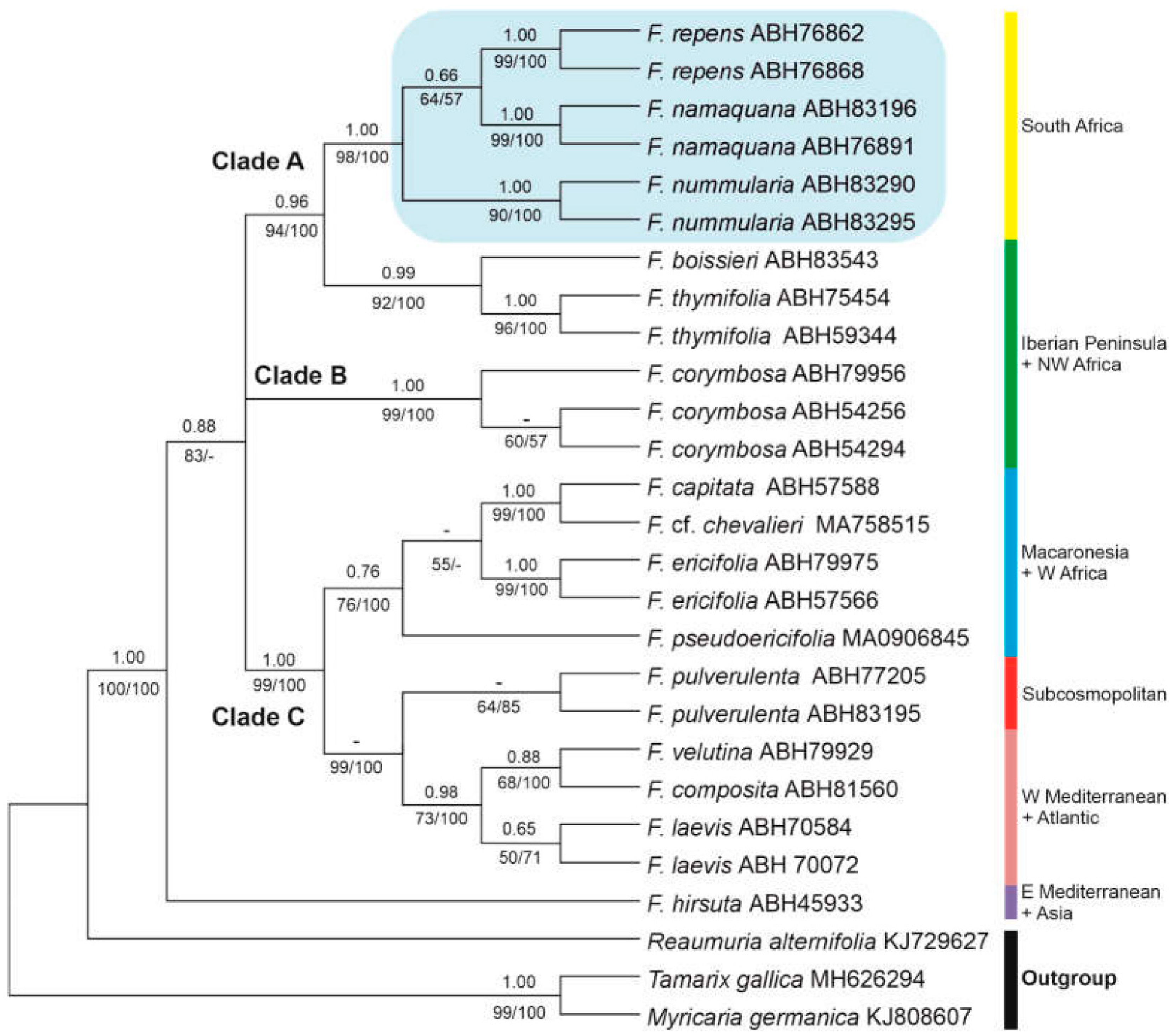
Species of Frankenia are arranged three strongly supported main groups, though not full resolved. Firstly, Clade A (0.96 PP, 94/100 BP) contains the South African members related to F. nodiflora nested in a strongly supported clade (1.00 PP, 98/100 % BP) together with F. repens (hereafter the “F. repens group”), in which three well to strongly supported lineages are recovered that correspond to the newly described F. nummularia (1.00 PP, 90/100 BS) and F. namaquana (1.00 PP, 99/100 BS) plus F. repens (1.00 PP, 99/100 BS). This clade is sister (0.99 PP, 92/100 BP) to a group including the E Iberian-W Algerian endemic F. thymifolia (1.00 PP, 96/100 BP) plus the SW Iberian-NW Moroccan endemic F. boissieri Reut. ex Boiss. Secondly, Clade B (1.00 PP, 99/100 BP) is formed by accessions of F. corymbosa Desf. from SE Iberian Peninsula and N Morocco, though the internal relationships of this clade are very weak. Finally, Clade C (1.00 PP, 99/100 BP) includes taxa with a broad Mediterranean, Macaronesian, Atlantic or Subcosmopolitan distributions. Samples of the annual F. pulverulenta from South Africa form only in the ML and MP analyses a weakly to moderately supported clade (- PP, 64/85 BP), which is strongly supported (0.98 PP, 73/100 BP) as sister to a clade including western Mediterranean-Atlantic (F. laevis L. and F. composita Pau & Font Quer) and northwestern Atlantic Moroccan (F. velutina Brouss. ex DC.) members of the genus. They all nest sister (0.76 PP, 76/100 BS) to a weakly supported clade (- PP, 55/-) including the Capeverdean endemic F. pseudoericifolia Rivas Mart. & al. as sister (0.76 PP, 76/100 BP) to the group formed by the Canarian F. ericifolia C.Sm. ex DC. (1.00 PP, 99/100 BP) plus the group (1.00 PP, 99/100 BP) of the also Canarian F. capitata and the W African F. cf. chevalieri Maire. The eastern Mediterranean species F. hirsuta L. (s.l.) falls sister to the rest of the studied taxa (Clade A+B), a relationship that is in need of further check.
3.3. Taxonomic treatment of Frankenia nodiflora and description of new related species
3.3.1. Frankenia nodiflora Lam., Encycl. 2(2): 543. 1788 ≡ Franca nodiflora (Lam.) Vis. in Mem. Reale Ist. Veneto Sci. 16: 158. 1871.
Type: [South Africa. Western Cape]. E Cap[ut]. B[onae]. Sp[ei]. [Sonnerat s.n.] (lecto. designated here: P00287094!; isolectotype: P05038792! (Cap de B[onne]. Espér[ance]. [Sonnerat s.n.]).
−F. pulverulenta auct. pl. atque F. krebsii auct. pl.
Description: Shrublet densely branched, tap-rooted, woody at the base with grey bark, mostly glabrous. Stems rooting at base, diffuse, usually creeping, suffruticose, up to 40 cm long, often divaricate branches 1–3 cm long; young branchlets with internodes up to 35 mm long, yellowish to reddish, glabrous or sometimes shortly and loosely puberulous. Leaves opposite-decussate, patent to erect-patent, bright-green or sometimes reddish, glabrous on both sides; petiole 0.6–1.2 × 0.2–0.3 mm, flattened, tapering distally; sheath extending along margins of petiole to blade, laxly ciliate (occasionally subglabrous), with 2–5 pairs of lateral cilia 0.2–0.8 mm long, unequal, whitish, flattened, obtuse or acute; leaf blade 2.5–3.5 × 1–2 mm, broadly ovate-elliptic to oblong, mostly falcate upwards, subacute to minutely mucronate at apex, fleshy, concolorous, sometimes slightly paler abaxially with minute glands bearing salt depositions, often cochleariform (spoon-like convex) to flattened, longitudinally folded downwards, with margins strongly to loosely and gradually revolute (at least on the upper third), somewhat thickened; midrib narrow, linear, tapering slightly towards apex, continuous with petiole below, somewhat raised abaxially, extending all along the blade length; young leaves on short shoots fasciculate, similar but smaller than those on long shoots and sometimes narrower. Flowers pentamerous, perfect, borne commonly in dichasial groups, usually condensed at stem nodes, glomerular or with short axillary branches up to 20 mm long (usually shorter); the central flower sessile, the lateral ones with pedicels up to 1 mm long. Floral bracts 2, 2–3 mm long, leaf-like, subpatent to erect-patent, connate at base, enveloping basal part of calyx for 0.5–0.7 mm. Bracteoles 2, 1–2 mm long, bract-like but smaller, about half to two thirds the length of calyx, adnate to the calyx base and alternating with bracts. Calyx 4–4.5 × 1–1.5 mm, tubular at anthesis, untwisted, straight, indurate, with 5 prominent thickened ribs, entirely glabrous; teeth 5, 0.9–1.2 mm long, triangular, narrowly membranous and shortly papillate on margins, often reddish, slightly recurved after anthesis, cucullate with a notable subapical acumen ca. 0.7 mm long, obtuse and slightly divergent. Petals 5, 5–7 × 1–2 mm, obovate-cuneate, pinkish-mauve but whitish below, overlapping only in the basal part; claw 2–3 × 0.5–0.7 mm, narrowly cuneate, imperceptibly tapering to blade, yellowish, hidden into the calyx tube; ligule 1.5–2.5 × 0.3–0.4 mm, narrowly oblanceolate, longitudinally adnate to claw, the free apex ca. 0.5 mm long, oblong-obtuse, entire; blade 2.3–3 × 1.5–2 mm, broadly obovate with apex rounded to truncate, slightly emarginate and irregularly sinuate (not erose-denticulate). Stamens 6, in 2 unequal whorls, usually long exserted, overtopping 1.5–2 mm the calyx teeth at anthesis; filaments 3–6 mm long, expanded ca. 0.5 mm wide in the lower half, but gradually tapering and filiform in the distal half, whitish; anthers 0.5–0.7 mm long, oblong-ellipsoid, versatile, yellowish. Ovary ellipsoid, subtrigonous, with 3 carpels; placentae 3, parietal-basal, extending up to the lower half to two thirds of carpel wall length, ventral traces moderately to highly branched; ovules 4–6 per placenta, attached along most of placenta by erect funiculi 0.1–0.2 mm long. Style 3–4.5 mm long, terete, somewhat sigmoid at base, exserted and elongating up to 8 mm after pollination, whitish; style branches 3, filiform, 0.5–0.7 mm long, whitish; stigmas slightly clavate. Capsule 2.5–3.5 × 0.6–2 mm, ovoid-ellipsoid, hidden into calyx tube, dark reddish-brown, early dehiscent. Seeds 9–12, 0.7–1.0 × 0.3–0.5 mm, sulcate on one side, ellipsoid, pale brown, darker at the funicular part, developing rapidly even before flower has completely withered; testa thin, not mucilaginous, with surface weakly and irregularly ornamented with a subrectangular-reticulate pattern, finely striate, covered with medium-sized papillae 12–33 μm long, heterogeneous, globose to conical-obtuse, more densely disposed on the distal part.
Etymology: The specific epithet (nodiflorus, –a, –um = with flowers at nodes) refers to the disposition of flowers and inflorescences, mostly crowded at stem nodes.
Phenology: Flowering in late October–early January (occasionally in July–August), fruiting in November–February (occasionally in August–September).
Figure 4.
Frankenia nummularia sp. nov. (a) Western Cape: Beukesfontein, H.H.W.Pearson 5005; Isotype: P05038725); available at: https://science.mnhn.fr/institution/mnhn/collection/p/item/p05038725 (© Muséum National d’Histoire Naturelle, Herbarium, Paris); (b) Northern Cape: Kookfontein river; living plants in habitat, with details of leaves and withered inflorescences.
Figure 4.
Frankenia nummularia sp. nov. (a) Western Cape: Beukesfontein, H.H.W.Pearson 5005; Isotype: P05038725); available at: https://science.mnhn.fr/institution/mnhn/collection/p/item/p05038725 (© Muséum National d’Histoire Naturelle, Herbarium, Paris); (b) Northern Cape: Kookfontein river; living plants in habitat, with details of leaves and withered inflorescences.

Habitat and distribution: Frankenia nodiflora occurs on saline, azonal soils of salt pans and saline riverbeds, in the coastal lowlands at elevations of 0–150 m above sea level. The known distribution of the species extends across the Cape plains into neighbouring areas northeast from Cape Town in the Western Cape Province (Figure 3), a territory broadly included in the Fynbos (F) biome (mostly the F07 Bioregion) sensu Mucina and Rutherford [14], where it specifically inhabits the so-called “Cape Inland Salt Pans” (code AZi 9). In the coastal lowlands of the Fynbos biome, the climate is mild and oceanic (ameliorated by the ocean influences), with average temperatures ranging about 7°C in winter to 30°C in summer (average annual temperature ca 17°C) and frosts being rare and occasional. The average annual precipitation amounts to ca 500–540 mm, though rather differently distributed, the rainfall occurring mostly during winter (May to August) with a peak in July [14].
Notes: In the past, F. nodiflora surely was much commoner than today, just before the severe changes of landscape caused by extensive agriculture and urbanisation. Nowadays, the known extant populations of this species are concealed to nature reserves where meadows and patches of halophytic vegetation are conserved, in a few sites between Durbanville to Paarl. The most important threats are related to invasion of the natural habitats by alien species such as Cynodon dactylon L. and Cenchrus clandestinus (Hochst. ex Chiov.) Morrone (Pennisetum cladestinum Hochst. ex Chiov.) (P. Winter pers. comm.). In this context, urgent field prospections are required to locate new populations of this rare endemism that likely remain unnoticed, and also active management of the natural sites is required to avoid decline and extinction of the scarce wild populations. Therefore, new data are necessary for an accurate conservational labelling of the species, including counts of the number of populations and individuals, as well as their evolution in time. In the meantime, the conservation status of F. nodiflora is suggested here as Data Deficient (DD), though very likely it might be assessed at least as Vulnerable (VU) according to IUCN [42].
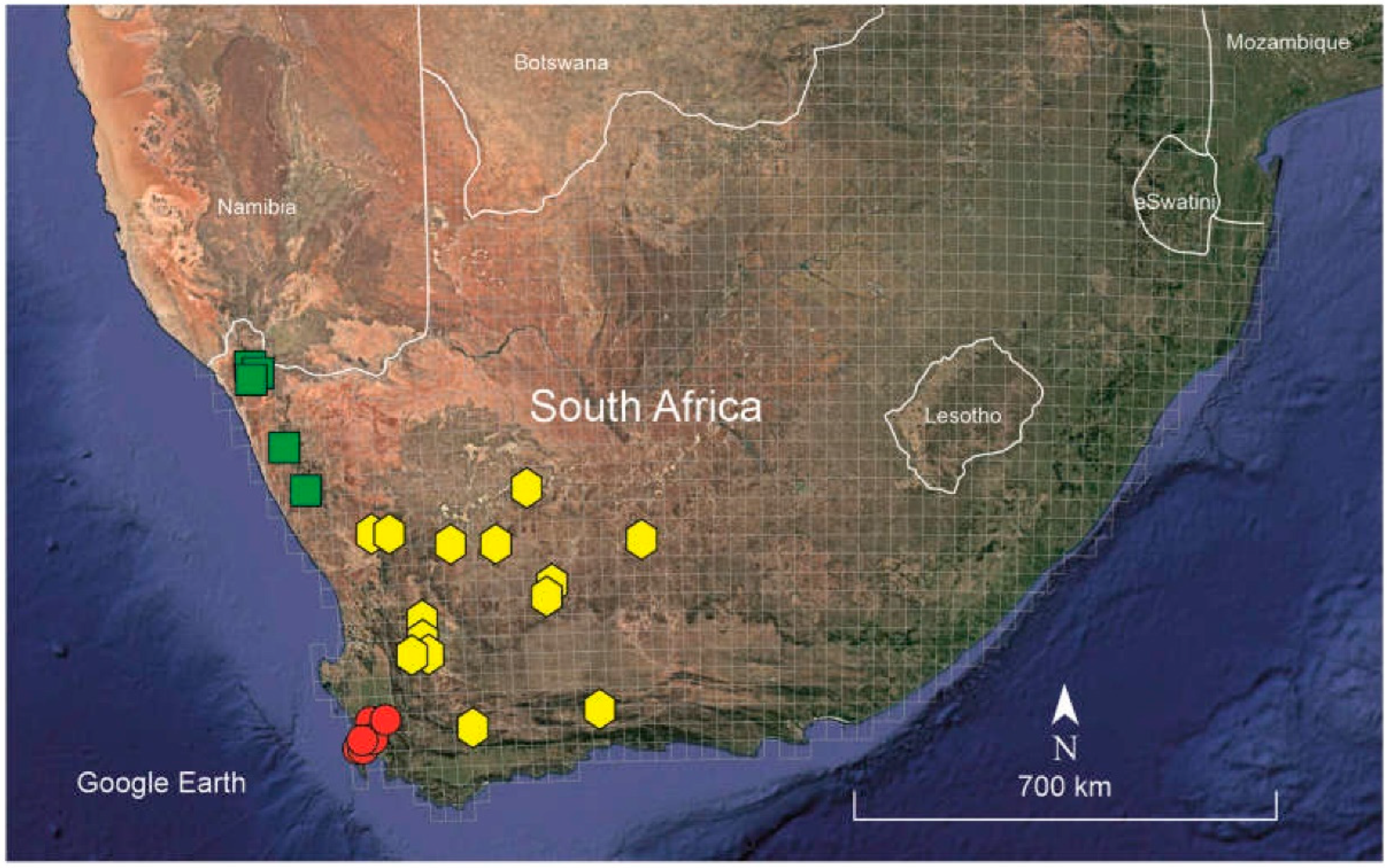
Figure 3.
Distribution map of the studied material (both herbarium vouchers and iNaturalist data) of Frankenia nodiflora (red circles), F. nummularia (yellow hexagons) and F. namaquana (green squares).
Figure 3.
Distribution map of the studied material (both herbarium vouchers and iNaturalist data) of Frankenia nodiflora (red circles), F. nummularia (yellow hexagons) and F. namaquana (green squares).
Other studied materials: South Africa. Western Cape Province: 3318 (Cape Town): Cape of Good Hope, Table Mountain (-CD), December 1832, J.Mac Gillivray 584 (K!); 3318 (Cape Town): Cape Town, Green Point (-CD), November 1846, A.Prior s.n. (K!); 3318 (Cape Town): Cape Town, Mowbray (-CD), shores of vlei, August 1912, W.C.Worsdell s.n. (K!); 3318 (Cape Town): Cape Peninsula, Raapenberg Vley (-CD), 26 November 1896, A.H.Wooley 2110 (BOL !, K!, excl. fragment in the central part); 3318 (Cape Town): Cape Town, about Salt River near the Windmills (-CD), 14 November 1811, [Pl. Africae Australis Extratropicae,] Burchell 513 (G-DC G00211140!, K!, P05038790!). 3318 (Cape Town): Cape Town, Uitkamp Wetlands Nature Reserve (-DC), 33°48’59.7”S 18°38’26.0”E, 137 m elev., 7 November 2014, H. Stummer s.n. (ABH83529!).
Unidentified sites: South Africa. Habitat ad C[apum] B[onae] Spei (B-W06993). Cape, Dr. Pappe s.n. (K000232047 !). Cap de Bonne-Espérance, 1842 (MPU693060 !). Cap de Bonne-Espérance (P05038793!). C[aput] B[onae] S[pei], Mrs. Gilavray s.n. (P05038816!). Cap de Bone Espérance, ex herb. Pet. Thouare s.n. (P05038804!). [Cape] Pl. Capenses, Ecklon (P05038764!).
Digital iNaturalist images: South Africa. Western Cape Province. 3318 (Cape Town): Cape Town, Durbanville, Belleville, Uitkamp Wetlands Nature Reserve (-DC), 33°48’59.7”S 18°38’26.0”E, 137 m elev., 7 November 2014, M.Goets (v.v.): https://www.inaturalist.org/observations/11060535; 3318DA (Cape Town): Cape Town, Malmesbury Farms, Teleport Rd, 33°41’18.0”S 18°42’25.0”E, 88 m elev., 26 April 2019, I. Ebrahim (v.v.): https://www.inaturalist.org/observations/24159910; 3318 (Cape Town): Paarl, Cape Winelands District, Windmeul Farm, Langerug Private Nature Reserve (-BD), 33°39’37.14"S 18°54’1.62"E, 146 m elev., 10 November 2017, J. Wicht (v.v.): https://www.inaturalist.org/observations/148456049
3.3.2. Frankenia nummularia M.B.Crespo, M.Á.Alonso, Mart.-Azorín, J.L.Villar & Mucina, sp. nov.
Type: South Africa. Western Cape: Karoo, Beukesfontein, sandy river bed, 1420 ft elev., 1 December 1908, H.H.W.Pearson 5005 (holo.: BOL!; iso.: K!, P05038725!). Figure 4a.
−F. pulverulenta auct. pl. atque F. repens auct. pl.
Diagnosis: Planta speciosa a F. pulverulenta et F. nodiflora foliis latis, subplanis (vel ad margines parve revolutis), longe petiolatis, et caulibus prostratis vel ascendentibus accedenti, sed ab eas distinctissima et bene distinguenda. A priore insuper differt caulibus suffruticosis, perennibus (non herbaceis annuis); foliis glaberrimis (non subtus pilosis); bracteolis calycem multo brevioribus (non aequantibus); et seminibus minus numerosis (ad 20, non 45) minoribusque (0.7–1.1 mm long., non 0.5-0.7 mm long.). A posteriore insuper discrepat foliis discoloribus, subtus valde pallidioribus (non subconcoloribus); calycibus majoribus 4.5–6 mm long., dentibus acutis vel mucrone inconspicuo ad 0.2 mm long. (non calycibus 4–4.5 mm long., dentibus mucrone magno ad 0.7 mm long.); ; et praesertim seminum testa papillis brevioribus, 3.5–9 μm long., subhomogeneis, omnibus conico-obtusis (non papillis longioribus 12–33 μm long., heterogeneis, aliis globosis aliis conico-obtusis).
Description: Shrublet densely branched, tap-rooted, woody at the base with grey bark, glabrous to long hairy. Stems rooting at base, procumbent to ascending, suffruticose, up to 40 cm long, usually with erect, often divaricate branches 5–35 cm long; young branchlets with internodes up to 30 mm long, yellowish to reddish, glabrous to ± densely hairy covered on one side with heterogeneous indumentum of both curled minute trichomes (ca. 0.2 mm) and flexuous long pluricellular complanate trichomes (0.7–1 mm long), denser below nodes. Leaves opposite-decussate, patent to erect-patent, greyish- to bright-green, somewhat glaucescent, glabrous on both sides, sometimes with salt depositions; petiole 0.7–1.5 × 0.3–0.5 mm, flattened, tapering distally; sheath extending along margins of petiole almost to blade, densely ciliate (occasionally almost glabrous), with 2–5 pairs of lateral cilia 0.3–0.8 mm long, unequal, whitish; leaf blade 2–7 × 1.5–6 mm, mostly suborbicular or broadly elliptic, rounded to slightly pointed at apex, somewhat fleshy, often discolorous, abaxially paler with minute glands usually bearing salt depositions, often longitudinally folded, with margins loosely and gradually revolute, somewhat thickened; midrib narrow, linear, tapering slightly towards apex, continuous with petiole below, somewhat raised abaxially, extending about half the blade length; young leaves on short shoots fasciculate, similar but smaller than those on long shoots and sometimes narrower and incurved on margins at the upper part. Flowers pentamerous, perfect, borne commonly in loose dichasial groups, usually widely branched, with erect-patent branchlets up to 20 mm long; the lowermost flowers solitary, sessile, the uppermost dichasia often denser and subcorymbose (the central flower always sessile, the lateral ones with pedicels 1–2 mm long). Floral bracts 2, 2–7 mm long, leaf-like, subpatent to erect-patent, connate at base, enveloping basal part of calyx for 0.5–0.7 mm. Bracteoles 2, 1–2 mm long, bract-like but smaller, up to half the length of calyx, adnate to the calyx base and alternating with bracts. Calyx 4.5–6 × 1–1.5 mm, fusiform-tubular to gradually fusiform after anthesis, untwisted, straight to slightly curved, indurate, with 5 prominent thickened ribs, entirely glabrous or sparsely puberulous to hirtellous at base and on ribs, hairs up to 0.5 mm long, whitish, occasionally with scattered whitish depositions on the upper part; teeth 5, 0.7–0.9 mm long, triangular, acute to minutely mucronate (mucro up to 0.2 mm long), minutely papillate on margins, often yellowish, erect to slightly incurved after anthesis. Petals 5, 6–9 × 1–2 mm, long obovate cuneate, pinkish-mauve, overlapping in most of their length; claw 4–5 × 0.8–1 mm, narrowly cuneate, imperceptibly tapering to blade, whitish, hidden into the calyx tube; ligule 2–3 × 0.5–1 mm, narrowly oblanceolate, longitudinally adnate to claw, the free apex ca. 1 mm long, triangular-acute, entire; blade 2.3–4 × 1.5–2 mm, broadly obovate to suborbicular with apex rounded to truncate, irregularly erose-denticulate. Stamens 6, in 2 unequal whorls, exserted, overtopping 1.5–2 mm the calyx teeth at anthesis; filaments 6–8 mm long, expanded ca. 0.5 mm wide in the lower half, but gradually tapering and filiform in the distal half, pinkish to bluish-pink; anthers 0.8–1.2 mm long, ellipsoid, versatile, yellowish to reddish. Ovary ellipsoid, subtrigonous, with 3 carpels; placentae 3, parietal-basal, extending up to the lower half to two thirds of carpel wall length, ventral traces moderately to highly branched; ovules 6–10 per placenta, attached along most of placenta by erect funiculi 0.3–0.4 mm long. Style 6–9 mm long, terete, somewhat sigmoid at the base, exserted and elongating up to 11 mm after pollination, whitish; style branches 3, filiform, 0.5–0.7 mm long, pinkish-white to reddish; stigmas slightly clavate. Capsule 2.5–3.5 × 1–2 mm, ovoid-ellipsoid, hidden into calyx tube, dark reddish-brown, early dehiscent. Seeds 12–22, 0.7–0.9 × 0.3–0.5 mm, ellipsoid to ovoid-ellipsoid, sulcate on one side, pale brown, darker at the funicular part, developing rapidly even before flower has completely withered; testa thin, not mucilaginous, with surface weakly and irregularly ornamented with a subrectangular-reticulate pattern, finely striate, almost smooth, only sparsely covered on the funicular side with small papillae 3.5–9 μm long, subhomogeneous, conical-obtuse.
Etymology: The specific epithet (nummularius, –a, –um = coin-bearing) refers to the shape of leaves and bracts, usually suborbicular and flattened, resembling coins.
Phenology: Flowering in late October–early January (occasionally in July–August), fruiting in November–February (occasionally in August–September).
Habitat and distribution: Frankenia nummularia grows in saline, well-drained sandy, azonal soils of salt pans, intermittent saline riverbeds and ravines in inland regions, at elevations of 250–1380 m above sea level (Figure 4b). The known distribution of the species extends through most of the SW part of the Karoo Region in western South Africa, ranging from Vanrhynsdorp and Riversdale District in the Western Cape to Calvinia and Victoria West District in the Northern Cape Province (Figure 3), a territory mostly included in the Nama-Karoo (NK) Biomes and reaching the southern Succulent-Karoo Biomes (mostly the SKk, SKt and SKv Bioregions) sensu Mucina and Rutherford [14], where it occurs concretely in to the so-called “Bushmanland vloere” (code AZi 5). In those regions, the climate is semiarid to arid to arid, continental (not or scarcely ameliorated by the ocean influences), with average temperatures ranging from –5°C in winter to 43°C in summer and frosts being usual in the higher areas. The average annual precipitation varies between 100 mm and 500 mm, though rather differently distributed, the rainfall occurring mostly during late summer (December to April) with a peak in March [14].
Notes: Wild populations of F. nummularia include numerous individuals covering a large territory in South Africa, and no special threats are known so far to occur that might lead to any inferred decline in either the number of populations or the number of individuals. Therefore, its conservation status is suggested here as Least Concern (LC) according to IUCN [42].
Other studied materials: South Africa. Northern Cape Province: 3021 (Vanwyksvlei): Rietspoort (-CB), (I, d1), 3000–4000 ft elev., 30 November 1826, J.F.Drège 2648 (765) (P05038732!); 3119 (Calvinia): Namaqualand, river bed W of Brandkop (-AC), 9 December 1946, F.M.Leighton 2441 (BOL!); 3119 (Calvinia): Zwart Doorn River, W of Brandkop (-AC), 9 December 1946, R.H.Compton 18893 (BOL!; NBG! excluding two annual plants); 3120 (Williston): An der Dualls Slangenfontein [Slangfontein] (-BD), (I, d1 d), 3000–4000 ft elev., 17 November 1826, J.F.Drège s.n. (P05038728!); 3120 (Williston): Sandwef [sic] on road to Brandvlei (-AC), 29 November 1986, G.Germishuizen 4011 (NBG!); 3120 (Williston): Hantam, Kookfontein farm, Kookfontein rivier at crossing R-354 (-CA), 31°43’35”S 20°14’07”E, 1082 m elev., in saline substrate of ravine, 25 August 2022, M.Martínez Azorín et al. s.n. (ABH83290!); 3121 (Fraserburg): Karoo Region, near Fraserburg (-DC), 4200 ft elev., January 1888, H.Bolus 10381 (NBG!); 3121 (Victoria West): Little Namaqualand, Bed of Brakrivier (-BD), 1600 ft elev., 11 December 1908, H.H.W.Pearson 4864 (K!); 3123 (Victoria West): Central Cape, Victoria West District, Hutchinson, Zeisiesfontein (-AC), 1260 m elev., E.M.Nortje 10 (NBG!); 3220 (Sutherland): Tankwa Karoo, between Middlepos and Ganaga Pass (-CB), 32°37’46.7”S 20°21’40.5”E, 573 m elev., 26 August 2022, M.Martínez Azorín et al. s.n. (ABH83295!); 3221 (Merweville): Fraserburg, Ratelfontein vel “Balmoral” (-BA), 4500 ft elev., January 1888, H.Bolus 10381 (BOL!, NBG!); 3221 (Merweville), Upper Region, Kopjies Kraal, river bed (-BA), 2000 ft elev., 12 December 1908, H.H.W.Pearson 4886 (K!). Capland: Boschjemanskarroo [probably near Bitterfontein], 3000–4000 ft elev., November, J.F.Drège s.n. (HBG516896!). Boschjemans-karroo oder Onderbokkeveld, [3000–4000 ft elev., November], J.F.Drège 6242 (P05038787!). Afrique austral, Herb. J. Hennecart, Drège, locum, 69 n° 1, [probably near Platberg], November 1838–1839, J.F.Drège (K!, P05038788!). Western Cape Province: 3118 (Vanrhynsdorp): Knersvlakte, Kalkgat farm (-BB), 255 m elev., 18 June 1987, C.Boucher 5175 (NBG!); 3219 (Wuppertal): SW Cape Region, foot of Katbakkies Pass, east side, at Skitterykloof picnic site (-DC), 1800 ft elev., 6 January 1976, H.C.Taylor 9049 (NBG!, K!); 3219 (Wuppertal): Karoo, Beukesfontein, sandy river bed (-CD), 1420 ft elev., 1 December 1908, H.H.W.Pearson 5005 (BOL!, K!, P05038725!); 3219 (Wuppertal), Central Karoo District: Pappekuil [Papekuil] (-BC), not far from river, in sand, 950 ft elev., 3 November 1908, H.H.W.Pearson 3985 (K!); 3219 (Wuppertal): Karoo: North of Gansfontein, river bed (-DA), 1200 ft elev., bush 1–1½ ft elev., 2 December 1908, H.H.W.Pearson 3984 (K!); 3319 (Worcester): Worcester, near Mowers station (-DA), 10 November 1964, Van Breda 1758 (NBG!); 3320 (Montagu): Laingsburg, Wittebergen near Matjiesfontein (-BA), October 1908, R.Marloth 11442 (NBG!); 3321 (Ladismith): Riversdale Div., Klein Karroo, damp places in river beds (-CC?), 1200 ft elev., October 1924, J.Muir 3546 (BOL!).
Unidentified sites: South Africa. Cap de Bonne Espérance, collection de Drège s.n. (HBG516896!, P05038791!). Cape, Drège s.n. (K!). Cap, 1838, Drège s.n. (P05144899!). Afr[ica]. austr[alis]., 1836, J.F.Drège s.n., sub F. nodiflora (K!, P05038789!).
Digital iNaturalist images:South Africa. Eastern Cape Province. 3323 (Willowmore): Graaff-Reinet, Dr Beyers Naudé Local Municipality, Timbila Nature reserve, Grootrivier bed (-BB), 33°11’14”S 23°53’16”E, 580 m elev., 29 September 2019, K.Jolliffe s.n. (v.v.): https://www.inaturalist.org/observations/33692903.
3.3.3. Frankenia namaquana M.B.Crespo, M.Á.Alonso, Mart.-Azorín, J.L.Villar & Mucina, sp. nov.
Type: South Africa. Northern Cape: Namaqualand, Steinkopf, 11 December 1897, Schlechter 40 (holo.: BOL!; iso.: P05038802!).
−F. capitata auct. pl. atque F. repens auct. pl.
Diagnosis: Planta speciosa a F. repenti caulibus valde lignosis et calycibus dense puberulis (interdum etiam costis hirsutis) remote accedenti, sed ab ea distinctissima et bene distinguenda caulibus erectis non radicantibus; petalis brevioribus 5.5.–6.5 mm long. (non 9–11 mm); calycibus minoribus 3-4 mm long. (non 6-8 mm); et praesertim seminum testa papillis brevioribus, 10–43 μm long, heterogeneis, aliis globosis aliis conico-obtusis (non papillis longioribus 55–110 μm, subhomogeneis, omnibus conico-obtusis).
Description: Shrub densely branched, tap-rooted, woody at the base with grey bark, glabrous to sparsely hairy. Stems non-rooting, erect to ascending, fruticose, usually with erect, often divaricate branches 15–45 cm long; young branchlets with internodes up to 20 mm long, yellowish to reddish, ± densely pubescent (rarely glabrescent), covered all around with homogeneous indumentum of minute claviform or hooked trichomes (ca. 0.1–0.2 mm) and longer complanate trichomes (up to 0.4 mm long), denser below nodes and somewhat retrorse to patent. Leaves opposite-decussate, patent to erect-patent, deep-green, somewhat glaucescent, glabrous on the upper side and ± densely papillate beneath, mostly with scattered whitish depositions; petiole 0.3–0.5 × 0.2–0.5 mm, flattened, tapering distally; sheath extending along margins of petiole almost to blade, densely ciliate, with 6–10 pairs of cilia 0.4–1.2 mm long, unequal, filiform, whitish; leaf blade 2–4 × 1–2 mm, broadly oblong to elliptic, rounded to subacute at apex, somewhat fleshy, often discolorous, abaxially paler with minute glands usually bearing salt depositions, margins strongly revolute hiding almost completely the abaxial side, somewhat thickened; midrib thickened, tapering slightly towards apex, continuous with petiole and raised abaxially, extending about half the blade length; young leaves on short shoots fasciculate, similar but smaller than those on long shoots and sometimes narrower. Flowers pentamerous, perfect, borne commonly in dichasial groups, usually branched, with erect branchlets up to 15 mm long, but often crowded in compact inflorescences; the lowermost flowers in reduced groups, briefly pedunculate, the uppermost dichasia often denser and subcorymbose (the central flower always sessile, the lateral ones with pedicels 0.5–1 mm long). Floral bracts 2, 2–3.5 mm long, leaf-like, erect-patent to erect, connate at base, enveloping basal part of calyx for ca. 0.5 mm. Bracteoles 2, 1–2 mm long, bract-like but smaller, about half the length of calyx, adnate to the calyx base and alternating with bracts. Calyx 3–4 × 0.8–1.5 mm, fusiform-tubular to gradually fusiform after anthesis, untwisted, straight, indurate, with 5 prominent thickened ribs, densely papillate (papillae whitish, minute, claviform or globose) between ribs but sparsely hirtellous on ribs (trichomes whitish, up to 0.2 mm long), sometimes with scattered whitish depositions on the upper part; teeth 5, 0.8–1.2 mm long, triangular, acute or briefly mucronulate (mucro ca. 0.2 mm long), minutely papillate, often yellowish, not recurved after anthesis. Petals 5, 5.5–6.5 × 0.9–1.3 mm, long obovate-cuneate, whitish to purplish; claw 4–5 × 0.8–1 mm, narrowly cuneate, imperceptibly tapering to blade, whitish, hidden into the calyx tube; ligule 2–3 × 0.3–0.5 mm, narrowly oblanceolate, longitudinally adnate to claw, the free apex ca. 0.5 mm long, ovate-acute to acuminate, entire to slightly denticulate on margins; blade 2.3–4 × 1.5–2 mm, broadly obovate to suborbicular with rounded to truncate, irregularly erose-denticulate apex. Stamens 6, in 2 unequal whorls, long exserted, overtopping 1.5–2.5 mm the calyx teeth at anthesis; filaments 6–8 mm long, expanded ca. 0.5 mm wide in the lower half, but gradually tapering and filiform in the distal half, pinkish to bluish-pink; anthers 0.4–0.6 mm long, ellipsoid, versatile, yellow. Ovary ellipsoid, subtrigonous, with 3 carpels; placentae 3, parietal-basal, extending up to the lower half to two thirds of carpel wall length, ventral traces moderately to highly branched; ovules 5–6 per placenta, attached along most of placenta by erect funiculi 0.2–0.4 mm long. Style 7–8 mm long, terete, somewhat sigmoid at the base, exserted and elongating up to 11 mm after pollination, whitish; style branches 3, filiform, 0.9–1.5 mm long, whitish; stigmas slightly clavate. Capsule 2.5–3.5 × 1–2 mm, ovoid-ellipsoid, hidden into calyx tube, dark reddish-brown, early dehiscent. Seeds 1–3, 0.7–1.1 × 0.3–0.4 mm, ellipsoid to ovoid-ellipsoid, sulcate on one side, pale brown, darker at the funicular part, developing rapidly even before flower has completely withered; testa thin, not mucilaginous, with surface weakly and irregularly ornamented with a subrectangular-reticulate pattern, finely striate, sparsely covered with medium-sized papillae 10–43 μm long, heterogeneous, globose to conical-obtuse, more densely disposed on the distal part.
Etymology: The specific epithet (namaquanus, –a, –um = Namaqualander) refers to Namaqualand, a semi-desert region of NW South Africa to which the new species is native.
Phenology: Flowering in late October–early January (occasionally in July–August), fruiting in November–February (occasionally in August–September).
Habitat and distribution: Frankenia namaquana grows in saline, well-drained sandy, azonal soils of salt pans, waadis, intermittent riverbeds and ravines in inland regions, at elevations of 300–600 m above sea level (Figure 5). The known distribution of the species is restricted to the NW part of the Karoo Region in NW South Africa, ranging from Eksteenfontein and Steinkopf to Klipfontein in the Northern Cape Province (Figure 2), a territory included in the northern Succulent-Karoo (SK) biomes (mostly in the SKn, SKr and SKs bioregions) sensu Mucina and Rutherford [14], where it specifically inhabits the so-called “Namaqualand Salt Pans” (code AZi 2). In those areas, the climate is subdesert to arid and continental (not or scarcely ameliorated by the ocean influence), with average temperatures ranging from 5°C in winter to 30°C (or even more) in summer, and frosts being absent or scarce but much varying between years. The average annual precipitation ranges around 70–200 mm, though rather differently distributed, and with occasional local rains reaching about 300 mm; the rainfall occurs mostly during winter (May to September) with a peak in June, and episodic drought periods well below 100 mm per year being frequent [14].
Figure 5.
Frankenia namaquana sp. nov. (a) Plant in habitat, near Springbok (Northern Cape Province); (b) Details of the inflorescences at anthesis (photos A. le Roux, 1 November 2014).
Figure 5.
Frankenia namaquana sp. nov. (a) Plant in habitat, near Springbok (Northern Cape Province); (b) Details of the inflorescences at anthesis (photos A. le Roux, 1 November 2014).

Notes: Wild populations of F. namaquana include numerous individuals covering a large territory in NW South Africa, and no special threats are known so far to occur that might lead to any inferred decline in either the number of populations or the number of individuals. Therefore, its conservation status is suggested here as Least Concern (LC) according to IUCN [42].
Studied material:South Africa.Northern Cape Province: 2817 (Vioolsdrift): Skoverfontein, ca 12 km NW of Eksteenfontein (-CC), P15, 28°45’50"S 17°09’15"E, 448 m elev., 19 August 2022, M.Martínez Azorín et al. s.n. (ABH831912!); 2817 (Vioolsdrift): Skoverfontein, ca 12 km NW of Eksteenfontein (-CC), P16, 28°46’25"S 17°09’41"E, 460 m elev., 19 August 2022, M.Martínez Azorín et al. s.n. (ABH83196!); 2817 (Vioolsdrift): Skoverfontein, ca 12 km NW of Eksteenfontein (-CC), P17, 28°46’33"S 17°10’18"E, 481 m elev., 19 August 2022, M.Martínez Azorín et al. s.n. (ABH83198!); 2917 (Springbok): Namaqualand, Steinkopf (-BC), 11 December 1897, Schlechter 40 (BOL!, P05038802!); Namaland Minor [near Springbok], W.C.Scully 9 (BOL!, P05038765!); 3017 (Hondeklipbaai): Klipfontein, 2–3 km N of Klipfontein, S of Kersboshoek (-BD), 30°28’43.3"S 17°49’40.9”E, 309 m elev., 30 August 2017, M.Martínez Azorín et al. s.n. (ABH76891!).
Digital iNaturalist images: South Africa. Northern Cape Province. 2817 (Vioolsdrif): Namaqualand, Eksteenfontein, Sendelingsdrif (-CC), 28°46’29”S 17°09’45”E, 458 m, wadi bed, 12 October 2018, S.Swanepoel s.n. (v.v.): https://www.inaturalist.org/observations/17553648; 2917 (Springbok): Namaqualand, Komaggas area, NW Oubeep (-DC), 29°51’41.36”S 17°33’53.26”E, 581 m elev., river bed, 16 March 2017, N.Helme s.n. (v.v.): https://www.inaturalist.org/observations/11292132.
4. Discussion
The findings presented in this paper are part of an ongoing broader study evaluating the global generic and specific relationships in Frankeniaceae, currently focused on the Eurasian and African taxa. Our ITS tree, as shown in Figure 2, is still partial but, in combination with morphological studies and field observations, it brings a better scenario for more accurate interpretation of the diversity of Frankenia in Southern Africa.
The three main groups (Clades A−C) obtained with the studied Mediterranean and African (including Macaronesia) taxa are apparently not clearly supported by gross morphology, perhaps due to the fact that convergent evolution in saline ecosystems might model similar morphological traits in members of phylogenetically distant lineages [43]. However, individual species discrimination is feasible based on characters such as habit, indumentum, leaf morphology, inflorescence structure, petal and calyx size or colour, or seed features, among others [e.g., 39,41,43−46].
In the case of the Frankenia repens group (Clade A), the internal molecular relationships among the three lineages found are rather well resolved and also well-characterised morphologically and biogeographically, and hence they are treated here at species rank: F. nummularia, F. namaquana and F. repens.
All those species share a suffruticose to woody perennial habit, with flowers often disposed in dichasial, more or less condensed, inflorescences; calyx untwisted; ovary with 15–20 parietal ovules; and seeds large, 0.7–1.1 mm long. However, important differences exist concerning particularly seed features, which are diagnostic to support distinction (Figure 6). First, F. nummularia is recovered in our tree as strongly sister (1.00 PP, 90/100 % BS) to the pair F. repens-F. namaquana, from which it differs on account of its leaves long petiolate (0.7-1.5 mm), mostly suborbicular or broadly elliptic, rounded to slightly pointed at apex, often flattened or longitudinally folded, with margins loosely and gradually revolute; stems with internodes up to 30 mm long, much longer than leaves; calyx 4.5–6 mm long, with teeth acute to minutely mucronate (mucro up to 0.2 mm long); seed testa almost smooth, with papillae small, 3.5–9 μm long, only at base, homogeneous, conical-obtuse in shape (Figure 6c−d). Second, F. namaquana is weakly supported as sister (0.66 PP, 64/57 % BS) to F. repens in our tree, both being superficially closer due to its leaves shortly petiolate (0.3–0.5 mm), with allantoid blade strongly revolute on margins to hide almost completely the abaxial side; and seeds covered with much larger papillae, up to 110 μm long (Figure 6e−f). All this led to misidentification of F. namaquana with F. repens or its synonyms [47−48], or also with the Macaronesian endemic F. capitata [38]. However, the weak sister support of both lineages is congruent with their clear morphological differences, F. namaquana clearly differing from the typical F. repens (lectotype: SBT10193 [digital image!]) based on:
- i)
- ascendent to erect stems (vs. prostrate, usually rooting stems);
- ii)
- calyx smaller (3–4 mm vs. 6–8 mm);
- iii)
- distinct petal length (5.5–6.5 mm vs. 9–11 mm); and
- iv)
- seed testa with papillae smaller (10–43 μm vs. 55–110 μm long), heterogeneous in shape, globose to conical-obtuse (vs. subhomogeneous in shape, cylindrical-conical obtuse), and sparsely disposed but denser on the distal part (vs. more densely disposed on testa surface; Figure 6f−g).
Figure 6.
Seed morphology (left) and testa details (right) of: Frankenia nodiflora (a−b); F. nummularia (c−d); F. namaquana (e−f); and F. repens s.l. (g−h).
Figure 6.
Seed morphology (left) and testa details (right) of: Frankenia nodiflora (a−b); F. nummularia (c−d); F. namaquana (e−f); and F. repens s.l. (g−h).
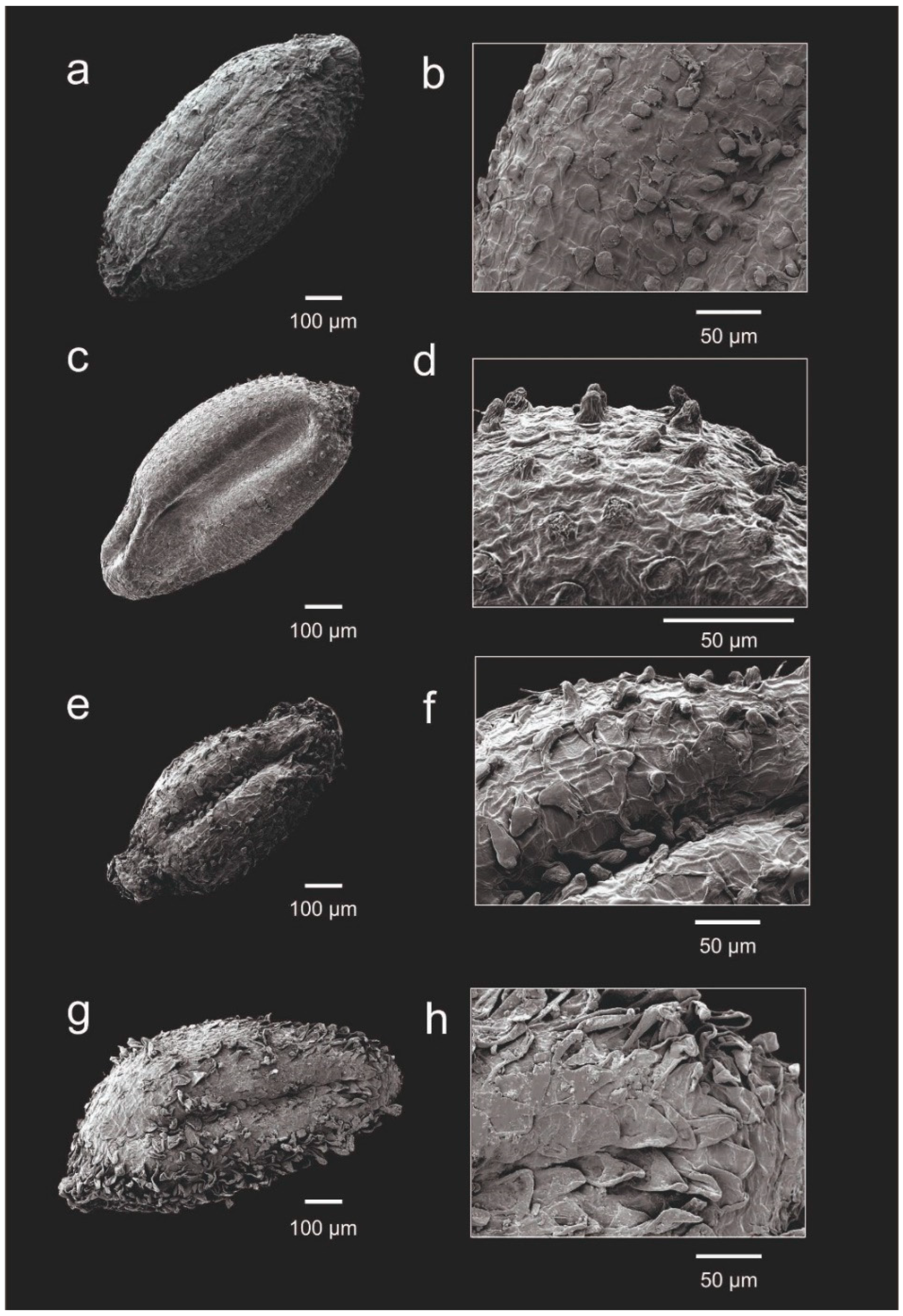
Furthermore, although no sequences were obtained for F. nodiflora, its morphological traits are distinct enough for an unequivocal and easy differentiation based on:
- i)
- leaves concolorous, bright green on both sides (or slightly paler beneath);
- ii)
- inflorescences mostly axillary and glomerular, borne on short lateral opposite branches;
- iii)
- calyx teeth conspicuously acuminate, with acumen up to 0.7 mm long;
- iv)
- ovary with 4–6 ovules per placenta; and
- v)
- seed testa sparsely covered with medium-sized papillae 12–33 μm long, heterogeneous, globose to conical-obtuse (Figure 6a−b).
Accordingly, the name F. nodiflora is restored here as initially described by Lamarck [13] and applied by South African botanists in the first half of the 19th century [47]. Some superficial resemblance of both F. nodiflora and F. nummularia to the annual or short-lived perennial F. pulverulenta, such as the broad, flattened or slightly revolute on margins, long-petiolate, and almost glabrous leaves; and the relatively small flowers, usually led to misidentification and consequently to synonymisation to the latter. However, the typical individuals of F. pulverulenta (lectotype: LINN 457.6 [digital image!]) can be easily distinguished (vs. both F. nodiflora and F. nummularia) by many taxonomically relevant characters such as:
- i)
- slender annual habit (vs. suffruticose perennial), with stems and branches pubescent only on one side with minute curled hairs (vs. glabrous or covered all around with minute curled and long flexuous hairs up to 1.2 mm long);
- ii)
- leaf blade long hairy on the abaxial side (vs. glabrous on both sides);
- iii)
- flowers solitary and scattered along branches (vs. crowded in loose or dense terminal and/or axillary dichasia);
- iv)
- petals 3.5–5 mm long (vs. 5–9 mm);
- v)
- calyx 2.5–4(5) mm, densely puberulous on groves (vs. entirely glabrous to sparsely puberulous to hirtellous at base and on ribs), with 2 bracteoles oblong-linear, as long as or longer than calyx tube (vs. suborbicular to broadly ovate-oblong, much shorter than the calyx tube); and
- vi)
- seeds very numerous, up to 45 per capsule, ca. 0.5–0.7 mm long (vs. less numerous, up to 20 per capsule, ca. 0.7–1.1 mm long), with testa very sparsely covered with small papillae 4–17 μm long, homogeneous, conical-obtuse.
Obermeyer [4], as other previous authors, regarded F. nodiflora (treated in a very broad sense to include also F. nummularia and F. nodiflora) as a perennial form of F. pulverulenta occurring in the subdesert parts of South Africa: “[recorded] further inland in the drier areas, in saline surroundings such as salt pans and banks of brackish streams. /…/ Collections indicate that the plants may behave as annuals or occasionally persist as perennials when they become more woody and larger.” On this respect, annual plants belonging to F. pulverulenta s.l. can grow together with other perennial members of the genus and sometimes they have been collected in a single site. In fact, vouchers R.H.Compton 18893 (NBG!) and R.H.Compton 18894 (NBG!), both identified as F: pulverulenta and collected in Zwart Doorn River, W of Brandkop (Calvinia) on 9 December 1946, are good examples illustrating this matter. The former (R.H.Compton 18893) includes several fragments of F. nummularia plus two entire annual individuals (affixed on the upper-left side and the lower-right corner of the voucher) with leaves pubescent on the abaxial surface and bracteoles equalling the calyx length (P. Winter pers. comm.) clearly belonging to the true F. pulverulenta, whereas the latter (R.H.Compton 18894) bears several individuals of the typical F. pulverulenta. Similarly, perennial plants of F. nodiflora co-occur with annual plants of F. pulverulenta in the surroundings of Cape Town, as deduced from a single herbarium sheet at K (“Herbarium Zeyheri - Cape, Dr. Pappe”), which includes three smaller herbaceous fragments matching F. pulverulenta affixed on the upper part of the sheet and marked “(1)” in pencil (barcode number K000232048!) plus four larger fragments of F. nodiflora (identified as “Frankenia krebsii ? /…/ an a Frank. capitata Webb diversa? /…/ non Frankenia nothria Thbg,”) in the central and lower parts of the sheet (barcode number K000232047!) and marked “(2)” in pencil.
Probably, the occurrence of both annual and perennial individuals sharing some morphological characters might have led to mistakenly consider them as conspecific under the name F. pulverulenta, and hence neglecting the existence of other well-characterised entities deserving taxonomic recognition.
Regarding other woody perennial Southern African taxa, such as F. fruticosa and F. pomonensis, both can be easily separated by exclusive combinations of character not found in any of the newly described species. Firstly, F. fruticosa (holotype: N.A.Helme 7796, NBG!) is a gnarled erect shrub with stems thickened and strongly woody at base, many-branched; leaves allantoid, small (1.5–2.5 × 0.5–1 mm) and dense puberulent on both sides; flowers showing petals with claw imperceptibly tapering and scarcely narrower than the blade, stamens long exserted after anthesis and only 6 ovules per ovary (one pair near the base of each of three subbasal-parietal placentas). These distinctive morphological traits make F. fruticosa a unique species in Africa [10], which should be regarded an edaphic specialist evolved after ecological diversification and adaptation to the peculiar conditions provided by quartz patches to which it is endemic in Knersvlakte Bioregion, western South Africa [49]. Secondly, F. pomonensis (holotype: M0104482 [digital image!]) differs by being a sprawling shrublet, with procumbent to ascendent stems up to 1 m tall; leaves 3–6 mm long, minutely puberulous or papillate beneath; flowers scattered, in loose terminal dichasia; calyx 5.5–6.5 mm long, glabrous; petals 8–9 mm long, purplish; stamens much shorter than petals, scarcely protruding from the corolla throat [50]. This remarkable plant is only known from the coastal areas of the Namib desert in southern Namibia, where it occurs on brackish flats, dolomite outcrops facing the sea, and bordering saline lagoons [4].
5. Conclusions
According to our current data, given the molecular and morphological distinctiveness and their non-overlapping distribution areas, we suggest recognition of all the studied South African entities in the F. repens group, at species rank: F. nodiflora, F. nummularia, F. namaquana and F. repens. With the newly described and restored species, Frankenia is constituted by seven species, which can be identified using the dichotomous key in Appendix A.
Further research including field observations and molecular work is being carried out specifically on coastal populations of the F. repens group to clarify their taxonomic adscription. Similarly, the annual or short-lived perennials resembling F. pulverulenta which occur in the subdesert and desert areas of both northern and southern Africa are currently being evaluated to better determine its identity and relationships.
Author Contributions
The following individual contributions were made: conceptualization, M.B.C., M.Á.A. and M.M.A.; methodology, M.B.C., M.Á.A. and M.M.A.; formal analyses, M.B.C., M.Á.A., J.L.V. and M.M.A.; writing—original draft preparation, M.B.C. and M.Á.A.; writing—review and editing, M.B.C.; collecting permit acquisition, M.M.A. and L.M.; field work, M.B.C., M.Á.A., M.M.A., J.L.V. and L.M. All authors have contributed substantially to the general discussion of results and taxonomic treatment, and have also read and agreed to the published version of the manuscript.
Funding
This work was partly supported by H2020 Research and Innovation Staff Exchange Programme of the European Commission, project 645636: ‘Insect-plant relationships: insights into biodiversity and new applications’ (FlyHigh); and by the grants UAUSTI19-08, UAUSTI21-04, ACIE17-01, ACIE18-03 and ACIE21-01, from diverse programmes of Universidad de Alicante. L.M. acknowledges logistic support of the Iluka Chair in Vegetation Science and Biogeography at Murdoch University, Perth, Australia.
Data Availability Statement
DNA sequence data generated in the present research are available at GenBank (https://www.ncbi.nlm.nih.gov/genbank/).
Acknowledgments
We thank the curators and staff of the herbaria ABH, BOL, K, M, HBG, NBG, P PRE, and PRU for facilitating access or providing information on their collections of Frankenia. Especially, we are thankful to Cornelia Klak (BOL), Anthony Magee (NBG) and Erik van Wick (PRE) for their help during our visit to the South African herbaria. Pieter Winter (NBG) and Hedi Stummer (Cape Town) kindly provided some important data and material from Cape Town. Vanessa Rodríguez Invernón (P) is kindly thanked for inclusion of the image of Frankenia nummularia from Paris, and also Cécile Aupic and Veronique Andro (P) for the image of F. nodiflora from Lamarck’s herbarium and a sample of seeds for SEM studies, as well as for interesting comments on some historical material at Paris. Alberto Herrero and Leopoldo Medina (MA) provided with samples from some vouchers for DNA extraction. María Álvarez and Saba Rokni (K) kindly helped with African collections and some rare publications at Kew. José Luis Villar (Universidad de Alicante) and Michael Pinter (Karl-Franzens University Graz) kindly helped with the field work, and Josy Zarur de Matos, Alexandria de Rusha, Jordi Abad and Miguel Pascual (University of Alicante) with the laboratory work. All herbarium specimens were collected under permission provided by the Department of Environment and Nature Conservation of Northern Cape Province (permits FLORA0057/2017, FLORA0058/2017) and CapeNature of Western Cape Province of South Africa (permits AAA005-00080-0028, AAA008-00031-0028, 0027-AAA008-00699, 0028-AAA008-00203).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Appendix A. Identification key for southern African species of Frankenia.
- 1.
- Leaves on flowering stems about 3–10 times shorter than internodes; blade broad, suborbicular to broadly oblong or obovate, flat or longitudinally folded, sometimes recurved on margins and on the distal half; petiole distinct, 0.5–1.5 mm long 2
- -
- Leaves on flowering stems mostly longer or up to 3 times shorter than internodes; blade allantoid, linear to linear-lanceolate, strongly recurved on margins; petiole inconspicuous, up to 0.5 mm long 4
- 2.
- Plants annual, delicate; leaves and bracts hairy beneath. Flowers scattered and solitary in forks of branches. Bracteoles as long as or longer than calyx. Petals 3.5–5 mm long. Seeds up to 45, small, 0.5-0.7 mm long F. pulverulenta
- -
- Plants suffruticose perennial, shrubby. Leaves and bracts glabrous on both sides. Flowers in terminal dichasia. Bracteoles about half the length of calyx. Petals 6–9 mm long. Seeds up to 20, large, 0.7–1.1 mm long 3
- 3.
- Leaves concolorous, bright green on both sides. Calyx 4–4.5 mm long; teeth with a long, conspicuously acumen up to 0.7 mm long. Ovules 4–6 per placenta. Seed testa sparsely covered with medium-sized papillae 12–33 μm long, heterogeneous, globose to conical-obtuse F. nodiflora
- -
- Leaves markedly discolorous, paler abaxially. Calyx 4.5–6 mm long; teeth acute or with an inconspicuous mucro up to 0.2 mm long. Ovules 4–6 per placenta. Seed testa almost smooth, with small papillae 3.5–9 μm long only at base, homogeneous, conical-obtuse F. nummularia
- 4.
- Stems stout, tortuous, strongly woody at base. Leaves 1.5–2.5 mm long. Calyx ribs densely puberulous. Petal blade scarcely wider than claw. Ovules 6, subbasal-parietal F. fruticosa
- -
- Stems thin, suffruticose. Leaves 3–7 mm long. Calyx ribs glabrous or sparsely hairy. Petal blade distinctly wider than claw. Ovules 15–18, parietal … 5
- 5.
- Plant sprawling, up to 1 m tall. Calyx 5.5–6.5 mm long, glabrous. Stamens much shorter than petals, scarcely protruding from the corolla throat F. pomonensis
- -
- Plant compact, erect to prostrate perennial, up to 45 cm tall. Calyx shorter (3–4 mm) or longer (6–8 mm), densely puberulent in grooves and often with coarse whitish hairs on ribs. Stamens much protruding from the corolla throat 6
- 6.
- Stems erect or ascendant, non-rooting, covered with minute hooked trichomes and longer complanate trichomes. Petals 5.5–6.5 mm long. Calyx 3–4 mm long. Seeds sparsely covered with medium-sized papillae 10–43 μm long, heterogeneous in shape, globose to conical-obtuse F. namaquana
- -
- Stems prostrate, often rooting at nodes, covered with short curly hairs. Petals 9–11 mm long. Calyx 6–8 mm long. Seeds densely covered with large papillae 55–110 μm long, subhomogeneous in shape, cylindrical-conical obtuse F. repens
References
- Niedenzu, F. Frankeniaceae. In Die Natürlichen Pflanzenfamilien, 2nd ed.; Engler, H.G., Prantl, K.A., Eds.; W. Engelmann, Leipzig, 1925; Volume 21, pp. 276–281.
- Correa, M.N. Las Frankeniaceae argentinas. Darwiniana 1966, 14, 68–94.
- Kubitzki, K. Frankeniaceae. In The families and genera of vascular plants 5. Malvales, Capparales and non-betalain Caryophyllales; Kubitzki, K., Bayer, C. Eds.; Springer: Berlin, Germany, 2003; Volume 5, pp. 209–212.
- Obermeyer, A.A. Frankeniaceae. Flora of southern Africa; Ross, J.H, Ed.; Botanical Research Institute: Pretoria, South Africa, 1976; Volume 22, pp. 32–36.
- Cuénoud, P., Savolainen, V., Chatrou, L.W., Powell, M., Grayer, R.J., Chase, M.W. Molecular phylogenetics of Caryophyllales based on nuclear 18S rDNA and plastid rbcL, atpB, and matK DNA sequences. Amer. J. Bot. 2002, 89, 132–144. https://doi.org/10.3732/ajb.89.1.132. [CrossRef]
- Olson, M.E., Gaskin, J.F., Ghahremani-Nejad, F. Stem anatomy is congruent with molecular phylogenies placing Hypericopsis persica in Frankenia (Frankeniaceae): comments on vasicentric tracheids. Taxon, 2003, 52, pp. 525–532. https://doi.org/10.2307/3647451. [CrossRef]
- Gaskin, J.F., Ghahremani-Nejad, F., Zhang, D.-Y., Londo, J.P. A systematic overview of Frankeniaceae and Tamaricaceae from nuclear rDNA and plastid sequence data. Ann. Missouri Bot. Gard. 2004, 91, 401–409.
- Summerhayes, V.S. A revision of the Australian species of Frankenia. Bot. J. Linn. Soc. 1930, 48, 337–387. https://doi.org/10.1111/j.1095-8339.1930.tb02580.x. [CrossRef]
- Heywood, V.H., Brummitt, R.K., Culham, A., Seberg, O. Flowering plant families of the world. Royal Botanic Gardens: Kew, UK, 2007.
- Manning, J.C., Helme, N.A. Frankenia fruticosa (Frankeniaceae), a new dwarf shrub from the Knersvlakte, Western Cape. S. African J. Bot. 2014, 91, 84–87. https://doi.org/10.1016/j.sajb.2014.01.001. [CrossRef]
- Bredenkamp, C.L. Frankeniaceae. In Plants of southern Africa: an annotated checklist; Geminshuizen, G., Meyer, N.L., Eds.; National Botanic Institute: Pretoria, South Africa, 2003; [Strelitzia 14], p. 561.
- POWO. Plants of the World Online (continuously updated). Available online: https://powo.science.kew.org/ (accessed on 3 April 2023).
- Lamarck, J.-B.P.A.M. de. Encyclopédie méthodique. Botanique. Panckoucke: Paris, France, 1788; Volume 2(2), pp. 543–544. https://www.biodiversitylibrary.org/page/33447031.
- Mucina, L., Rutherford, M.C., Eds. The vegetation of South Africa, Lesotho and Swaziland. South African National Biodiversity Institute: Pretoria, South Africa, 2006. [Strelitzia 19].
- Thiers, B. Index Herbariorum: a global directory of public herbaria and associated staff (continuously updated). Available online: http://sweetgum.nybg.org/ih/ (accessed on 1 Aug 2018 2023).
- Rasband, W.S. ImageJ. U.S. National Institutes of Health, Bethesda, Maryland. Available online: https://imagej.nih.gov/ij/(accessed on 19 March 2020).
- IPNI. The International Plant Names Index. Available online: http://www.ipni.org (accessed on 3 April 2023).
- Turland, N.J., Wiersema, J.H., Barrie, F.R., Greuter, W., Hawksworth, D.L., Herendeen, P.S., Knapp, S., Kusber, W.-H., Li, D.-Z., Marhold, K., May, T.W., McNeill, J., Monro, A.M., Prado, J., Price, M.J., Smith G.F., Eds.; International Code of Nomenclature for algae, fungi, and plants (Shenzhen Code) adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017. − Koeltz Scientific Books: Königstein, Germany, 2018; [Regnum Veg. 159]. https://doi.org/10.12705/Code.2018. [CrossRef]
- Leistner, O.A., Morris, J.W. Southern African place names. Ann. Cape Prov. Mus. Nat. Hist. 1976, 12, 1–565. [CrossRef]
- Doyle, J.J., Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15.
- White, T.J., Bruns, T.D., Lee, S.B., Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR protocols: a guide to methods and applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, USA, 1990; pp 315–322.
- Edgar, R.C. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucl. Acids Res. 2004, 32, 1792–1797. https://doi.org/10.1093/nar/gkh340. [CrossRef]
- Kumar, S., Stecher, G., Li, M., Knyaz, C., Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. https://doi.org/10.1093/molbev/msy096. [CrossRef]
- Nei, M., Kumar, S. Molecular evolution and phylogenetics. Oxford University Press: New York, USA, 2000.
- Felsenstein, J. Evolutionary trees from DNA sequences: a maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. https://doi.org/10.1007/BF01734359. [CrossRef]
- Saitou, N., Nei, M. The neighbour-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. https://doi.org/10.1093/oxfordjournals.molbev.a040454. [CrossRef]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control. 1874, 19, 716–723. https://doi.org/10.1109/TAC.1974.11007051974. [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. https://doi.org/10.1007/BF01731581. [CrossRef]
- Ronquist, F., Teslenko, M., van der Mark, P., Ayres, D.L., Darling, A., Hohna, S., Larget, B., Liu, L., Suchard, M.A., Huelsenbeck, J.P. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. https://doi.org/10.1093/sysbio/sys029. [CrossRef]
- Darriba, D., Taboada, G.L., Doallo, R., Posada, D. jModelTest 2: more models, new heuristics and parallel computing. Nat. Method. 2012, 9(8), 772. https://doi.org/10.1038/nmeth.2109. [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution 1985, 39, 783–791. https://doi.org/10.2307/2408678. [CrossRef]
- Villar, J.L., Alonso, M.Á., Juan, A., Gaskin, J., Crespo, M.B. Out of the Middle East: new phylogenetic insights in the genus Tamarix (Tamaricaceae). J. Syst. Evol. 2019, 57, 488–507. https://doi.org/10.1111/jse.12478. [CrossRef]
- Zhang, M.-L., Meng, H.-H., Zhang, H.-X., Vyacheslav, B.V., Sanderson, S.C. Himalayan origin and evolution of Myricaria (Tamaricaeae) in the Neogene. PLoS One 2014, 9(6), e97582. https://doi.org/10.1371/journal.pone.0097582. [CrossRef]
- Zhang, M.-L., Hao, X.-L., Sanderson, S.C., Vyacheslav, B.V., Sukhorukov, A.P., Zhang, H.-X. Spatiotemporal evolution of Reaumuria (Tamaricaceae) in Central Asia: insights from molecular biogeography. Phytotaxa 2014, 167, 89–103. http://dx.doi.org/10.11646/phytotaxa.167.1.5. [CrossRef]
- Sonnerat, P. Voyage aux Indes orientales et a la Chine, Fait par ordre du Roi, depuis 1774 jusqu’en 1781, 2 vols; l’auteur: Paris, France, 1782.
- Lamarck J.-B.P.A.M. de. Tableau encyclopédique et méthodique de trois règnes de la Nature. Botanique. Premiere livraison. Panckoucke: Paris, France, 1792; Volume 2(1); pp. 395–396, tab. 262.
- Candolle, A.P. de. Prodromus systematis naturalis regni vegetabilis. Treuttel et Würtz: Paris, France, 1824; Volume 1, pp. 349–350. https://www.biodiversitylibrary.org/page/153953.
- Harvey, W.H. Order XIV Frankeniaceae St. Hil. In: Flora Capensis; being a systematic description of the plants of the Cape Colony, Caffraria, & Port Natal; Harvey, W.H., Sonder, O.W., Eds.; Hodges, Smith & Co.: Dublin, Ireland, 1860; Volume 1, pp. 114–115.
- Roessler, H., Schreiber, A. Frankeniaceae. In Prodromus einer Flora von Südwestafrika; Merxmüller, H., Ed.; J. Cramer: Lehre, Germany, 1968; Volume 91, pp. 1–2.
- Manning, J.C., Goldblatt, P., Eds. Plants of the Greater Cape Floristic Region 1: The Core Cape Flora. South African National Biodiversity Institute: Pretoria, South Africa, 2012. [Strelitzia 29]. Volume 1, p. 583.
- Snijman, D.A. Frankeniaceae. In Plants of the Greater Cape Floristic Region 2: The extra Cape Flora; Snijman, D.A., Ed.; South African National Biodiversity Institute: Pretoria, South Africa, 2013; [Strelitzia 30]; Volume 2, pp. 399–400.
- IUCN. IUCN Red List Categories and Criteria: Version 3.1, 2nd ed.; UCN Species Survival Commission: Gland, Switzerland, and Cambridge, UK, 2012.
- Grigore, M.N., Flowers, T.J. Evolution in gngiosperm halophytes. In Handbook of halophytes; Grigore, M.N. Eds.; Springer: Cham, Switzerland, 2021; pp. 2117–2146. https://doi.org/10.1007/978-3-030-57635-6_86. [CrossRef]
- Nègre, R. Les Frankenia du Maroc. Trav. Inst. Sci. Chérifien, Sér. Bot. 1957, 12, 5–56.
- Chater, A.O. Frankenia. In Flora europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, Eds.; Cambridge University Press: Cambridge, UK, 1968; Volume 2, pp. 294–295.
- Santos, A. Frankenia. In Flora iberica 3, Plumbaginaceae (partim)-Capparaceae; Castroviejo, S., Aedo, C., Cirujano, S., Laínz, M., Montserrat, P., Morales, R., Muñoz Garmendia, F., Navarro, C., Paiva, J., Soriano, C., Eds.; Real Jardín Botánico, CSIC: Madrid, Spain, 1993; Volume 3, pp. 446–453.
- Drège, J.F.: Zwei planzengeographische Documente. Flora 1843, 26(Besondere Beigabe), 1–230.
- Le Roux, A. Wild flowers of Namaqualand, 4th rev. ed. Struik Nature: Cape Town, South Africa, 2015.
- Alonso, M.Á., Crespo, M.B., Martínez-Azorín, M., Mucina L. Morphological and molecular data support recognition of Spergularia quartzicola (Caryophyllaceae) as a new species endemic to South Africa. Plant Biosyst. 2022, 156, 506–514. https://doi.org/10.1080/11263504.2021.1887955. [CrossRef]
- Pohnert, H. Neue Arten aus Südwest-Afrika. Frankeniaceae. Mitt. Bot. Staatssamml. München 1954, 1, pp. 446–448.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



