
Submitted:
08 May 2023
Posted:
09 May 2023
You are already at the latest version
Abstract
Citation: To be added by editorial staff during production.
Keywords:
Nanotechnology
; Pest management
; Biosynthesized silver nanoparticles
; Entomopathogenic fungi
; Toxicity mechanism
1. Introduction
Globally agriculture industry is facing a major issue of loss of crop yield due to several abiotic and biotic factors. But insect pests are causing a major decline in agricultural production. Insects can feed on a wide variety of plants, including crops, weeds, forest trees, and medicinal plants. They also infest food and other stored products in bins, godowns, and packages, causing massive food loss and deterioration of food quality. Insects that produce more than 10% harm are classified as major pests whereas those which cause damage between 5 to 10% are classified as minor pests [1]. Globally insect pests are causing an 18-20% loss in crop production and the annual cost of crop loss is approximately US $ 470 billion [2].Therefore, a variety of chemicals, particularly insecticides, are utilized to protect the valuable crops that are harmed by pests. Chlorantraniliprole, cyantraniliprole, novaluron, neonicotinoids, fipronil, farnesyl acetate, emamectin benzoate, phoxim, and pyrethroid are a few examples of pesticides used to control pests. Methoprene, fenoxycarb, and pyriproxyfen are a few examples of juvenile hormone analogues used as insecticides, but all of these pesticides harm the ecosystem, and many pests have acquired resistance to them [3,4,5]. To reduce the damage of pests in fields, many chemical pesticides like chlorantraniliprole and cyantraniliprole are used, but extreme usage of these chemical pesticides has led to the formation of insect resistance, pest resurgence, and various environmental hazards. The deteriorating activity of chemical pesticides has diverted researchers to focus on ecofriendly control tactics [6]. An entomopathogenic fungus is a microorganism with the ability to infect, parasitize and kill arthropod pests is the entomopathogenic fungi. Generally used as an alternative to conventional chemical pesticides in organic farming, also have their roots in the fields of biotechnological processing and Chinese medicine. These EPF don’t belong to a single monophyletic group but instead, there are known about 12 species from Oomycetes, 399 species from Microsporidia, 65 species from Chytridiomycota, 474 species from Entomophtoromycota, 283 species from Basidiomycota and a total of 476 species from Ascomycota. Out of all these the ones belonging to phylum Ascomycota and phylum Entomophtoromycota are found more frequently in nature. The highest entomopathogenic efficacy among them was observed from the ones belonging to phylum Entomophtorales (like Conidiobolus, Furia, Erynia, and Entomophaga) but due to lack of proper breeding technologies in laboratories, they are avoided as components for bio-preparation. However, the saprophytes belonging to the phylum Ascomycota are commonly utilized. Other species of entomopathogenic fungi which are utilized as biopesticides are from the genera Beauveria, Paecilomyces, Metarhizium, Lecanicillium,and Isaria. Since these fungi have an array of applications, they can be utilized to eradicate a wide spectrum of insect pests [7,8,9,10,11,12,13,14]. Entomopathogenic fungi directly penetrate the outer integument of an insect’s body to infect it. The process of infection starts with spore attachment to the exoskeleton of the arthropod due to electrostatic and hydrophobic forces, along with the activation of the lytic enzymes and secondary metabolites [15]. The fungal hyphae begin to develop after invading the insect’s body cavity. Some EPF produces blastoconidium or spores that invade the insect’s haemolymph and develop secondary hyphae that remain in the tissues and impair the physiological processes. The body of the insect, which is primarily soft, stiffens as the infection progresses due to fluid absorption by the pathogenic fungus [16]. However, the effectiveness of EPF is restricted by limitations such as a short shelf life, slow action, high application cost a lack of persistence, and a poor infection rate in harsh environmental conditions. Frequently slow-acting and necessitate a high application rate and extensive spray coverage. Therefore, one way to enhance the efficacy of these fungi is by using nanoparticles [17].
Nanotechnology is a novel approach to agricultural research that helps to reach the goal of environmentally benign, cost-effective crop protection and production. It provides a way to synthesize nanoparticles (1–100 nm) that may be used as nanopesticides to eradicate various agricultural pests. Nanoparticles are the primary building blocks of nanotechnology. Nanoparticles can be characterized based on of properties such as morphology, shape, size, etc [18]. When nanoparticles enter an insect’s intracellular area after penetrating the exoskeleton, they attach to phosphorus or sulfur in proteins or DNA, which leads to the denaturant of organelles and enzymes. Nanoparticles can reduce membrane permeability, resulting in cellular function loss and cell death. Nanoparticles can be administered judiciously and are swiftly taken up by cells, which slows the emergence of resistance in pests. Nanoparticles can be used in a variety of ways for effective pest management, such as a combination formulation of metal and another material that is virulent against a certain insect [19,20]. Among all nanoparticles, silver nanoparticles have prospective applications in different fields of science, such as life sciences, agriculture, forensic science, and food chemistry. Nowadays, silver nanoparticles are gaining a lot of popularity due to their insecticidal properties. Numerous studies have demonstrated that silver nanoparticles are quite effective against a range of insect species. Additionally, the use of biosynthesized nanosilver, more specifically silver nanoparticles produced by the entomopathogenic fungus, has grown in favour. EPF is becoming a desirable candidate for the synthesis of nanoparticles because of their high yields, ability to produce large quantities of proteins, ease of handling, and low toxicity of the residues [21,22,23,24,25]. In this regard, the main aim of this review is to highlight EPF as an emerging candidate for the synthesis of nanoparticles. The review also spotlights the efficacy of biosynthesized nanoparticles against pest species. Finally, the review concludes that there is a need for further research as very little is known regarding the mode of action of biosynthesized nanoparticles in insect pests.
2. What Are Entomopathogenic Fungi?
Generally, the term entomopathogens refers to the organisms that are pathogenic to insect pests. Entomopathogens can be naturally occurring such as certain bacteria, fungi, viruses, and nematodes which will infect a certain number of insect pest species and will help us to manage crop growth. Entomopathogenic fungi were first among the microorganisms used as biocontrol agents against pests. The first successful use of biocontrol methods to eradicate pests was initiated in 1762 to control the population of sugarcane red locust Nomadacris septemfasciata through the introduction of the mynah bird Acridotheres tristis from India into Mauritius. However, the use of EPF, Beauveria bassiana as a biocontrol agent first occurred in 1956 in China against the sweet potato leaf weevil Cylas formicarius in 1959 [26,27,28]. EPF is recognized as a class of bioinsecticides that can be used to eradicate insect pests. They are regarded as effective biological insect control agents. Throughout a significant portion of its life cycle, EPF is mostly harbored in natural or cultivable soils of the terrestrial ecosystem. These organisms are more abundant in soils with more organic matter and acidity [29,30]. They are phylogenetically different, unicellular or multicellular, eukaryotic microorganisms that have the potential to irradicate the pest by attacking them, causing infection, using them as hosts, and ultimately killing the pest to keep their population below the economic injury level. They are facultative or obligatory pathogens that have a high survivalship rate and reproduction capacity. Generally, these fungi are categorized into 6 major Phylum: Basidiomycota, Ascomycota, Oomycetes, Microsporidia , Entomophtoromycota, and Chytridiomycota [9,31,32,33].
Entomopathogens have advantages over chemically synthesized insecticides as they are eco-friendly, precise, and safe to use. The term entomopathogen was first coined in a study. It described the microbes which aided in the insect population control and maintain them within limits that caused no economic damage to the crop plants [34]. EPF has a larger host range compared to entomopathogenic bacteria and viruses as they are host specific, hence they can infect both the soil-dwelling as well as above-ground insect pests. Pest directly affects crop production by feeding on the crops, therefore, reducing the yield and quality of the crops. Even more devastating consequences are faced by farmers if the population of a particular pest is huge as many pests are believed to be found in millions globally. The impact of agricultural pests is so detrimental that globally it has resulted in a 40% reduction of the total agricultural output. It is also estimated that almost 10.80% of the agricultural losses worldwide are caused by insect pests only after the green revolution [1,35]. Conventionally chemical pesticides were a reliable method of insect pest control because they required low effort for application, showed good efficacy, and were expedient. However, in 80% of cases, overuse of chemical pesticides has resulted in the development of resistance to more than one class of insecticide [36,37]. Entomopathogenic fungi are natural insect population control agents as they are naturally pathogenic to a range of insect pests and also since they are derived from nature hence show little to no adverse effect on the environment. Different EPFs have different host choices and hence can be specialists or they can infect a variety of insect hosts making them a generalist pathogen. Other pathogenic organisms like protozoans, viruses, and bacteria needed to be ingested or enter inside the host body in some way to cause the infection, but entomopathogenic fungi can utilize the nutrients present in the body cuticle of the insect pest to colonize the host body hence making them a good entomopathogenic agent to utilize [38].
3. Effect of Different Entomopathogenic Fungi against Pest
Entomopathogenic fungi are parasitic microorganisms that are capable of infecting and killing a wide range of insects. In organic farming, these fungi are used as a biocontrol agent and serve as a safer, more effective, and eco-friendly method than chemical insecticides [8,11,13]. EPFs such as Metarhizium, Beauveria, and Trichoderma are parasitic microbes that can infect and kill a variety of arthropods. They directly penetrate the outer integument of the insect’s body to infect and kill them [15].
In tropical countries, one of the major fruit crops is the papaya (Carica papaya Linn.). Papaya is well known for its nutritional, therapeutic, and prophylactic values worldwide. Papaya Ring Spot Virus (PRSV) can cause crop loss from 10% up to 100% every year [39]. The vector for the PRSV is the aphid Aphis gossypii and they tend to transmit this virus in a very non-persistent manner [40]. The utilization of chemical pesticides is detrimental to the environment as well as to human health. The experiment was performed to study the efficacy of indigenous EPF which was collected from the agriculture fields of North 24 Parganas during the pre-monsoon time. From the soil, a range of fungi was isolated including 11 isolates of Trichoderma, 9 isolates of Aspergillus, and 8 isolates of Penicillium. A total of 20 isolates out of the 40 isolates extracted showed entomopathogenic tendencies against the 3rd and 4th instar larva of the aphids of papaya. However only 3 isolates out of these 20 showed proper entomopathogenic activities and those isolates were identified as Penicillium sp. (Nlg 1), Fusarium sp. (khr4), and Beauveria sp. (deb4). The efficacy test of these fungi on 10 aphids suggested that the efficiency of B.bassiana was greater than P.verrucosum followed by F. equiseti. These entomopathogenic fungi produce spores that germinate on the surface of the cuticle of the aphis, killing as a result of penetration of the cuticle [41].
Whitefly (Bemisiatabaci), a hemipteran insect having several genetically distinct species, is one of the most economically important insect pests of ornamental and vegetable plants throughout the world’s tropical and subtropical climates. The major threat to crops is the nymphs and the adult whiteflies as they feed directly on the plants as well as act as vectors for various plant viruses. Whiteflies produce honeydew while feeding, which reduces the rate of photosynthesis in plants. Chemical pesticides are commonly used to control whitefly populations, but they harm the environment and endanger the safety of non-target organisms. It has been discovered that insects are becoming resistant to these chemicals. EPF can be used as an alternative to chemical pesticides because it is derived from the environment and is a natural enemy of whiteflies. The majority of entomopathogenic fungi are found in the Zygomycetes (phylum-Zygomycota), Hyphomycetes (phylum-Deuteromycota), and Laboulbeniales and Pyrenomycetes (phylum-Ascomycota).
Entomopathogenic fungi are also easy to isolate because they can be extracted from cadavers of insects and soil, or grown in an artificial medium. Some of the EPF which are commonly produced commercially and have been documented to act as biological control agents are B. bassiana, M. anisopliae, and I. fumosorosea. Some of the genera of EPF which are known to kill the developmental stages of white flies are Aschersonia, Verticillium, and Isaria. Clonostachysrosea is another EPF that has been found to have pathogenic effects on the 4th instar nymph and adult stages of B. tabaci. An entomopathogenic fungi Aschersoniaaleyrodis could be used as a potential biological control agent for Bemisia. tabaci as it is documented to be able to efficiently parasitize the whiteflies with greater than 50% mortality within 7 days. The survival rate of the Bemisia tabaci 1st, 2nd, and 3rd instars was also significantly reduced after treatment with A. aleyrodis in the greenhouse [42].
Throughout Appalachia, one of the foundation tree species is the American Chestnut (Castanea dentate) which once numbered 4 billion in the early 1900s. However ever since the commercial value of chestnut wood increased to nearly USD 750,000 today, the chestnut forest saw a huge amount of deforestation. Following the deforestation, there was also the introduction of chestnut blight (Cryphonectria parasitic) which coupled with the deforestation led to the rapid decline in the chestnut population to the point where there was no chestnut tree canopy in the eastern forest. Fortunately, chestnut blight-resistant trees were introduced in the forest which was able to seize the spread of chestnut blight. Unfortunately, following this success was the resurgence of another mortal enemy of the chestnut trees known as the (Curculio sayi) Lesser Chestnut Weevil. C. sayi was a prominent pest of the chestnut trees with an infestation rate between 50-75% considered to be normal while some reached a 100% infestation rate also.
This infestation with the lesser chestnut weevil larvae and excreta also decreased the commercial value of the infested tree due to the quality of the wood being adversely affected. The C. sayi larvae can directly damage the chestnut wood physically and even if they are heat-killed post-harvest then also they reduce the nut meat substantially. Also, the larvae will facilitate the growth of Aspergillus fungus which will produce the diarrheagenic toxin emodin. Like with any other chemical pesticides, the pesticides administered to control the C.sayi population will eventually adversely affect the environment hence a greener and eco-friendly approach is required to control the lesser chestnut weevil population. Since the adult lesser chestnut weevil will climb the canopy of the chestnut trees during the spring season hence utilizing EPF as a biological control agent to control them seems like a favorable alternative to chemical pesticides. For the experiment, an entomopathogenic fungus called Beauveria bassiana was applied to each trunk of mature chestnut trees in a 0.5m band encircling the circumference of the chestnut trees and terminating approximately around 1 meter above the ground level. For measuring the efficacy of the EPF pyramid and trunk traps were used to monitor the adult lesser chestnut weevil population. After the application of the EPF on the trunk of the chestnut trees, the catch rate of the C.sayi adults in the trap was significantly reduced compared to the control. Furthermore, chestnuts treated with entomopathogenic fungus had a much lower chance of weevil infection, both as the major effect and when compared to the control. The reduction of the trap catch of C. sayi was attributed to two possibilities. The first possibility was that the application of the EPF on the trunks of the mature chestnut trees acted as a natural repellent for the adult lesser chestnut weevil to climb the top of the canopy and secondly, the application of the EPF showed a potentially lethal effect on the adult C. sayi as the unfortunate insects which came in contact with the EPF contracted the fungus which slowly colonized them ultimately killing them in the process in a matter of days via systematic fungus infections. Another EPF besides B. bassiana which was able to survive the harsh northern winters and also showed better efficacy in controlling the lesser chestnut weevil population was S. feltiae [43].
One of the economically important pests in commercial crops is from the order Hemiptera which are often referred to as true bugs. The name of the infamous bug is Elasmolomus pallens, it is a post-harvest pest insect and can be found worldwide in different climatic zones including subtropical, tropical, and temperate. The success of the bug is linked to its feeding choice of legume plants like Arachis hypogea L. in Sub-Saharan Africa. A significant chunk of the economic loss of peanuts is caused by the adults and the nymphs of the E. pallens during the harvest and storage of the peanuts. These bugs use their piercing mouth part called the Rostrum for feeding. The aftermath of the E. pallens infestation is that the seeds will be shriveled which will result in increased content of fatty acids and a rancid flavor in the oil made from those seeds. The entomopathogenic studied was A. flavus and the fungus was isolated from the colony of A. flavus. The bioassays used in this experiment included a dose-response bioassay, an enzyme bioassay, and a correlation between mortality rates and enzymatic activities. In the dose-response bioassay, the conidial concentration was directly proportional to the mortality rates. With the increase in the conidial concentration, the mortality rates also increased. However, the oil-formulated conidia were more virulent, with 100% combined mortality achieved after 8 days compared to 92% mortality achieved after 9 days of treatment with conidia formulated in the surfactant Tween 80. Both mortality rates were higher compared to the control which was approximately 10% after 10 days. In the enzyme assay, it was observed that the enzyme activity increased with the number of days of culture which decreased later. A. flavus showed maximum protease activity on the 8th day, chitinase activity on the 8th day, and lipase activity on the 6th day. The correlation between E. pallens mortality rates and enzyme activities of cuticle-degrading enzymes such as protease, lipase, and chitinase was found to be positive. As a result, the study confirmed that A. flavus fungal isolates were a potential biocontrol entomopathogenic fungal agent for controlling seed bug (E. pallens) populations. The efficacy of Aspergillus flavus on the mango seed weevil (Sternochetus mangiferae) was also investigated, and it was found to have an 80% mortality rate [38].
3.1. Secondary Metabolites of Entomopathogenic Fungi and Their Role in Pest Infection
It has been confirmed that entomopathogenic fungi secrete a wide variety of secondary metabolites which are low molecular weight organic compounds. By lowering the target species’ resistance level and ultimately causing nervous system damage, these secondary metabolites aid in the effective infection of pests and other target organisms. Secondary metabolites are classified chemically as amino acid derivatives, peptides, cyclic depsipeptides polyketides, terpenoids, and peptide hybrids [14,44].It has been stated that the secondary metabolites are synthesized through gene clusters such as non-ribosomal peptide synthetases (NRPSs), hybrid NRPS–PKS genes, etc. The presence of more secondary metabolites in the entomopathogenic fungal genome than in the genome of typical fungi has been demonstrated during the sequencing of fungi. When these secondary metabolites come into touch with insect tissues, they become active. Entomopathogenic fungi gain pathogenicity through secondary metabolites. However, the bulk of these secondary metabolites are dormant and need activators like host exposure or environmental stress.The generalist insect pathogens Metarhizium and Beauveria infect more than a thousand pest species. Both of these entomopathogenic fungi include more than 80% distinct secondary metabolite genes, according to genomic analyses. Beauvericin and bassianolide, both generated by Beauveria spp., as well as destruxins, which are mostly produced by Metarhizium, are the secondary metabolites that are present. These compounds are known to play a significant function in giving entomopathogenic fungus pathogenicity [45,46].
The effect of secondary metabolites secreted by Beauveria bassiana on the adult pest, Eurygaster integriceps was studied in a research [47]. According to the study, the metabolites inhibited phagocytic activity, which ultimately changed the way the immune system worked. Additionally, it was shown that the number of hemocytes rapidly decreased as metabolite concentration increased. Plutella xylostella is a cosmopolitan pest of economically important crops. The toxic crude metabolites extracted from Isaria fumosorosea in Czapek Dox liquid medium were used against the third larvae of P. xylostella. The highest mortality of 91 percent was recorded after 6 days of treatment which showed maximum mortality. Therefore, it was concluded thatthe active microbial metabolite compound can be extracted which could lead to the development of natural products for plant protection [48]. The effects of secondary metabolites from Isaria fumosorosea, Beauveria bassiana, and Paecilomyces variotii on feeding, growth, fecundity, and hatchability of Spodoptera litura was also investigated. The secondary metabolites were extracted through the solvent extraction method. It was observed that the metabolites had an immense impact on the fecundity and the hatchability of the pest [49].
The most effective entomopathogenic fungus against insect pests is Cladosporium cladosporioides. Tests on cotton aphid nymphs and adults were conducted on the extract of this fungus’ secondary metabolites. At concentrations of 128 ppm and 154 ppm, the C. cladosporioides extract was shown to be most efficient against both nymphs and adults. The 3-(4-hydroxy-6-pyranonyl)-5-isopropylpyrrolidin-2-one) and 3-(4-hydroxy-6-pyranonyl)-6-pyranonyl)-pyrrolidin-2-one were also found in the fungal extract after GCMS analysis. The insecticidal property was thought to be caused by both compounds [50]. Different secondary metabolites produced by entomopathogenic fungi and acting as insect growth regulators were examined in a study. The fungi were isolated from the soil to investigate their insecticidal property. The culture was identified as Lecanicillium attenuatum which showed maximum insecticidal activity against the pest Plutella xylostella. The fungi strain showed a high level of juvenile hormone antagonists (JHANs) that disrupted the juvenile hormone receptor complex which led to the larval death due to the disruption in insect physiology. Therefore it was concluded that entomopathogenic fungi L.attenuatum secretes different kinds of secondary metabolites exhibiting JHANs activity [51]. GC-MS analysis revealed various bioactive chemicals, including propanoic acid, ethyl ester, acetic acid, and propyl ester, among others, in the ethyl acetate extract of Penicillium sp. for characterization. After 48 hours of treatment, Spodoptera litura larvae exposed to Penicillium sp. ethyl acetate extract demonstrated considerable larvicidal activity, with LC50: 72.205 mg/ml and LC90: 282.783 mg/ml. After 48 hours of exposure to crude extract, high antifeedant activity was seen in 300 g/ml. According to the results of this study, secondary metabolites from Penicillium species are useful for controlling Spodoptera litura larvae [52].
3.2. Mode of Action of Entomopathogenic Fungi against Insect Pest
The mode of action of any entity in the insect body can be through oral ingestion or contact but entomopathogenic fungi don’t need to be ingested to cause infection. entomopathogenic fungi can directly infect the insect by penetrating the insect exoskeleton. The infection process is mediated by several adhesions, secondary metabolites, and lytic enzymes [14]. The initial step of infection is a fungal invasion. The process of fungal invasion starts with the attachment of conidia of insect pathogenic fungi on the insect cuticle, which is the first barrier. The composition of an insect’s cuticle varies depending on its developmental stage. The outermost layer of the insect cuticle is the epicuticle, which is composed of cuticulin (lipoprotein), wax, and some organic compounds, and is followed by the procuticle. Endocuticle, mesocuticle, and exocuticle are the three types of procuticle. Procuticle majorly constitutes chitin and proteins. Exocuticle consists of protein such as melanin and carotin which provide hardness, impermeability, and dark color to the chitin. The endocuticle is the third innermost layer of the epidermis that surrounds and protects the internal structures of the insect [53,54,55]. The attachment of spores to the insect’s outer integument layer occurs by firstly using electrostatic and hydrophobic forces followed by activation of the lytic enzymes, low molecular weight proteins, and secondary metabolites [15]. During germination and swelling, conidium may attach to the cuticle or secrete mucus for adhesion. Some infective sessile spores have an adhesion drop at the spore end to aid in insect attachment [56,57]. B. bassiana and M. anisopliae both produce hydrophobic conidia with a surface rodlet layer composed of proteins known as hydrophobins which can easily interact with the hydrophobic outer lipid layer. Mad1 and Mad2 are adhesion genes found in M. anisopliae. These adhesion proteins contain signal peptides, threonine-proline-rich regions implicated in adhesion mediating, and putative glycosylphosphatidylinositol anchor sites that would localize the proteins to the plasma membrane. The Mad1 gene may not specifically target an adherence gene, and its loss results in low spore germination and adhesion, resulting in a reduction in blastospore number and virulence. These genes are responsible for passive adhesion and adsorption, whereas the Mad2 gene is a target-specific gene for the active stage. At least two hydrophobins (Hyd1 and Hyd2) are involved in rodlet layer assembly in B. bassiana, which contributes to virulence adhesion to hydrophobic surfaces and cell surface hydrophobicity [58]. After adhesion, the cuticle penetration of pre-germinated spores involves some structures and general processes, and the mechanisms of penetration may vary for each fungus. It is required for fungus to penetrate the cuticle in order to reach the nutrition and energy sources. Cuticle penetration requires the collaboration of mechanical, enzymatic, and physical activities. Fungus enters the cuticle through a germ tube or an appressorium that injects an infection peg into the cuticle [59,60]. Entomopathogenic fungi secrete many cuticle-degrading enzymes chitinases, esterases, proteases (chymoelastase, collagenase, trypsin, esterase, and chymotrypsin), and lipases. Also, the secretion of enzymes endoproteases and aminopeptidase is associated with appressorium formation. B. bassiana requires four catalases (catB, catP, catC, catD), a hydrocarbon carrier protein (Acyl-CoA oxidase), eight cytochromes P450 genes (CYP52-X1, CYP655-C1, CYP5337-A1, CYP52-G11, CYP539-B5, CYP617-N1, CYP53-A26, CYP584-Q1) and long-chain alcohol (3-oxoacyl carrier protein reductase) for digestion of lipids in the cuticle [61,62,63,64]. The fungal hyphae begin to develop after invading the arthropod’s body cavity. Some entomopathogenic fungi produce blastoconidium or spores that invade the insect’s hemolymph and develop secondary hyphae that remain in the tissues and impair physiological processes. In hemocoel of insect fungi release various mycotoxins such as M. anisoplia produce cytochalasins and Destruxins (depsipeptidesdestruxin B and desmethyldestruxin B). B.bassiana produce beauvericin, brassinolide, beauverolides, and isarolides [65,66]. The body of an insect, which was primarily soft, stiffens as the infection progresses due to fluid absorption by the pathogenic fungus and ultimately insects die due to inadequate nutrition, mycotoxin, and tissue impairment. But entomopathogenic Fungi require specific environmental conditions to germinate and cause infection, and their shelf life is limited. Also, preventive treatment is difficult because pests must be present before the pathogen can be usefully applied. They are slow acting and necessarily involve a relatively high rate of application and extensive spray coverage. Under adverse environmental conditions, there is a lack of persistence and a low rate of infection. So, to enhance its effect on pests mycosynthesized nanoparticles are being studied [17,44].
4. Different Routes of Synthesis of Nanoparticles
Physical and chemical techniques are used in the production of nanoparticles. Both bottom-up and top-down mechanisms can be used to obtain nanoparticles. The top-down approach entails mechanically grinding bulk metals, followed by the stabilization of the resulting nanoscale metal particles with colloidal stabilizers. The generated objects display a broad size variation with a mean diameter of roughly 100 nm. As a result, such a method is typically unsuitable for producing particles with clearly defined geometrical configurations. The bottom-up approach can be regarded as an approach that comes from the other side: There are numerous ways to break down a tiny precursor, which is typically an organometal complex or a salt: thermal breakdown, optical excitation, which separates the metal atom from the organic residue, or using a reducing agent. Narrow size distribution and particles with a diameter of 1–50 nm are produced by the nucleation of several metal atoms. Therefore, this technique is the most widely employed in NPs chemical synthesis [67,68]. The top-down approach includes physical methods like laser ablation, evaporation-condensation, and arc discharge method, and the bottom-up approach includes chemical methods such as microemulsion techniques, chemical reduction, and electrochemical synthetic techniques for the manufacture of NPs. Most chemical and physical methods of producing nanosilver are extremely expensive and involve the use of poisonous, dangerous substances that may offer possible environmental and biological concerns. [69,70,71].
4.1. The Greener Synthesis of Metal Nanoparticles
These constraints prompted the recent creation of NPs synthesis based on the concept of green nanotechnology, which was then successfully applied in the biosynthesis of nanoparticles. Whereas extracts from living animals are used, highlighting the usage of amino acids, polysaccharides, vitamins, and proteins/enzymes as well as other compounds with a coating capacity of metal nanoparticles. The reduction and stabilization operations are carried out in a single step during the biosynthesis process, making it possible to produce nanoparticles quickly [72,73,74,75]. It has been observed that several microorganisms, including bacteria, fungi, actinomycetes, and yeast, are capable of synthesizing nanoparticles, mineral crystals, and metallic nanoparticles either intracellularly or extracellularly. Two mechanisms—extracellular and intracellular—are involved, depending on where NPs are synthesized. Extracellular synthesis refers to the generation of NPs outside of the plant or microbial cell as a result of the presence of biomolecules such as enzymes, amino acids, and proteins. Metal Ion transition within the cell occurs as a result of electrostatic interaction between positively charged metal ions and negatively charged cell walls. Metal ions are converted into metal NPs by enzymes found in the cell wall. Since NPs are created inside the cells, intracellular synthesis is a very difficult process. The extracellular technique does away with the extra steps needed for the intracellularly produced NPs to be extracted. This is why the extracellular technique is typically chosen for the creation of nanoparticles [76,77,78].
In this review, the emphasis will be on developments in the synthesis mechanism and elements influencing nanoparticle properties. To comprehend the mechanisms of nanoparticle biosynthesis, microbes are still being used increasingly in recent years. Since bacteria and fungi have more advanced technology than actinomycetes and yeast, their usage in the creation of nanoparticles has attracted greater attention than that of actinomycetes and yeast. Fungi are appealing as reducing and stabilizing agents in the biogenic synthesis of silver nanoparticles due to the production of significant amounts of proteins, high productivity, simplicity in handling, and low toxicity of the residues. Furthermore, the nanoparticles are coated with fungus-derived proteins, which can increase stability and possibly provide biological activity [77,79]. Fungal cultures have advantages over bacterial cultures, such as strong biomass generation and the lack of additional procedures needed to collect the filtrate. Fungi mycelial mass is more resistant to pressure and agitation than plant mycelial mass, making them better suited for large-scale syntheses. Furthermore, it is possible to manipulate fungus metabolism to produce nanoparticles with specific properties, such as size and morphology, by adjusting culture factors such as time, temperature, biomass amount, and pH, among others. [80,81]. Figure 1 illustrates different methods for the synthesis of nanoparticles.
5. Synthesis of Metal Nanoparticles by Entomopathogenic Fungi
5.1. Selenium Nanoparticles
Fungi of the Trichoderma genus are widely utilized and studied as a biocontrol agent against pathogenic plant fungi as well as nematodes and bacteria. The hydrolytic enzymes present in Trichoderma tend to degrade the cell wall of pathogens. Trichoderma also have secondary metabolites for example isoprenoids and peptaibols which aid in the control of root and foliar pathogens. Studies related to the entomopathogenic effect of Trichoderma were scarce as generally the fungus interacts with subterranean organisms. However, it is described in certain places that Trichoderma species do have entomopathogenic properties, especially against the lepidopteran insects as they are known to contain chitinase enzyme that degrades the cuticle of the target insect and when suitable conditions are provided, will affect the peritrophic matrix in silkworms negatively. Aside from biological methods, physical and chemical approaches can be utilized to synthesize the selenium nanoparticles although these will require the use of a higher quantity of hazardous chemicals, higher thermal conditions, and more acidic pH due to which the biological application of these nanoparticles can become extremely lethal and dangerous. Acknowledging these side-effects, the synthesis of selenium nanoparticles via biological method is preferred as gives a safer, eco-friendly, and more non-toxic alternative than the conventional way. Additionally, those biologically synthesized SeNPs tend to be more stable as they have a natural coating of organic material on their surface which will prevent the aggregation of the nanoparticles over a long time [82,83,84,85,86,87,88,89,90].
A polyphagous insect that is responsible for widespread destruction of crops such as cauliflower, tea, rice, cotton, chili, groundnut, cabbage, capsicum, maize, and tobacco is Spodoptera litura. Conventionally, to control the S. litura population chemical pesticides were used however they adversely affected human health along with the environment and non-target beneficial arthropods. A study was performed to know the efficacy of mycosynthesized SeNP from Trichoderma and its effect on the S. litura larva. The synthesis of the nanoparticles was done utilizing the Trichoderma extract which was added dropwise into 20mM 100 mL selenium solution which was placed in a magnetic stirrer and then Na2SO3was added in the solution to maintain the pH at 8-12. The reaction mixture was kept in the dark condition for reduction and the temperature was maintained at 27O C ± 2O C also the mixture was kept in the orbital shaker for 24 hoursat 120 rpm and any color change was observed. After this, the nanoparticles synthesized were stored in dark conditions till a brick red-colored sedimentation was observed at the conical flask bottom. Following this, they were kept for 3-4 days after which they were subjected to centrifugation at 8000rpm and then they collected the pellet, dried them at room temperature, and stored them for further characterization.
The characterization results obtained indicated that the Trichoderma mycosynthesized SeNPs have a 259nm UV-Vis peak range. FTIR analysis also indicated the presence of many functional groups like phosphines, anhydrides, nitro groups, alcohols, alkanes, and also sulfonates. The morphology of the SeNP was confirmed to be spherical, while the EDaX analysis showed that 87.98% of the Selenium elemental compound was present. XRD confirmed the crystalline structure of selenium with an average size of 137 nm. DLS indicated the SeNP ranged between 40-100nm in size [90,91,92,93,94].
5.3. Zinc Nanoparticles
The biocompatible nature and the unique properties of the zinc nanoparticles make it the best candidate for a variety of applications including biomedical, anti-microbial, etc. However, the chemicals used in the conventional synthesis procedures render this ability of zinc nanoparticles ineffective. In order to combat this issue in recent times, green synthesis methods for zinc nanoparticles are being developed worldwide and are gaining their deserved spotlight with the scientific community. Green synthesis includes synthesizing the nanoparticles, may it be zinc, silver, titanium, silica, or any other, using biological entities like plants (phytosynthesis) or fungi (mycosynthesis). Fungi can be found in a variety of locations but the most common approach to collecting fungi for mycosynthesis is from the soil or insect cadaver [98]. In the study [99], the zinc nanoparticles were synthesized using the mushrooms of Daedalea species. In the study 10 grams of the mushroom culture was added into 100ml of distilled water to prepare the extract which was then slowly mixed in the 100ml zinc solution. The zinc solution was prepared by dissolving 1.834 grams of zinc acetate dehydrate in 100ml of distilled water. This solution was held on the hot plate for 6 hours at 100 degrees Celsius, with the magnetic stirrer set to 250 revolutions per minute. The colour of the zinc oxide solution was conserved, and the pH was determined to be 1.9. That sample was then sentenced to centrifugation for 15 min at 15,000 rpm. The pellet was collected while the supernatant was discarded. It was then washed and transferred the nanoparticles into a petri dish, which was heated on a hot plate at 300º C. The resulting dry particles were crushed to a fine powder and properly kept at room temperature until further usage. The UV Vis spectrophotometric analysis showed the range of wavelength to be between 360nm to 380nm at room temperature. XRD analysis was used to determine the crystallinity of the nanoparticles. The structural characteristics of the ZnO nanoparticles were determined using the XRD spectrum. The crystals are just normal atom arrays. The XRD patterns validated the phase identity, purity, and crystallite size. The spectra of ZnO NPs show distinct diffraction, with peak values of (100), (002), (101), (102), (110), (103), (200), (112), (201), and (202), respectively. They have a hexagonal crystalline structure. The zinc nanoparticles had an average crystal size of 14.53 nm. The SEM study showed that the nanoparticle’s shapes were irregular and that they were aggregated and agglomerated which might be credited to the use of extract made from wild sticky mushrooms that acted as a stabilizing and capping agent. The EDX analysis indicated that in the mycosynthesized nanoparticles, the amount of zinc was 58.34 % and oxygen was 13.28%. The mycosynthesis of zinc nanoparticles utilizing species of Trichoderma namely, T. reesei, T. harzianum, and T. reesei, and the co-culture of Trichoderma spp. 1g of zinc nitrate hexahydrate was dissolved in 10 ml of double distilled water. Then 2 milliliters of the fungal extract of T. reesei, T. harzianum, and T. reesei and the co-culture of Trichoderma spp. was added to the solution and stirred in a magnetic stirrer for 5-10 minutes approx. Following this the mixture thus obtained was heated in a preheated muffle furnace that was maintained at 400 ± 10O C in which the mixture boiled, frothed, and the heat-forming foam dehydrated within 3 minutes. The sample obtained was calcinated at 700O C for 2 hours and the final product thus obtained was stored till further use. When the UV visible spectrophotometric was performed, the absorption spectrum was observed within 372 nm- 374nm. The FTIR analysis spectrum was observed between the range 400 cm-1 to 600 cm-1. EDX analysis showed that the proportion of zinc was 50.36% and that of oxygen was 49.64%. TEM images indicated the presence of small zinc nanoparticles which were agglomerated [100].
5.4. Biogenic Synthesis of Silver Nanoparticles Mediated by Fungi
Fungi’s ability to produce a diverse range of compounds useful in a variety of applications is very promising. Ascomycetes and imperfect fungi, which are microscopic filamentous fungi, as well as other fungi species, are known to produce approximately 6,400 bioactive compounds. Due to their tolerance for heavy metals and ability to internalize and bioaccumulate metals, these organisms are frequently utilized as reducing and stabilizing agents. Additionally, the fungus may readily be grown on an industrial scale to manufacture nanoparticles with precise size and morphology (“nano factories”) [79,109]. Beauveria and Metarhizium, two genera of the entomopathogenic fungus, are frequently isolated from soil. They offer benefits like strong enzymatic activity, secondary metabolite synthesis, and insecticidal qualities. Consequently, EPF is viewed as the best substitute for chemical and physical synthesis [14,110].
To date, different fungi have been found to synthesize silver nanoparticles. Silver nanoparticles were synthesized extracellularly in a study using Beauveria bassaina. Freshly inoculated B. bassiana pure culture was placed in an Erlenmeyer flask on a liquid medium that contained KH2PO4, K2HPO4, MgSO4, H2O, (NH4)2SO4, yeast extract, and glucose. For 72 hours, the flask containing medium was incubated in an orbital shaker at 150 rpm at 25±2 °C. The biomass was harvested after 72 h of growth by sieving through Whatman No. 1 filter paper, followed by an extensive wash with distilled water to remove any medium components from the biomass. 200 mL milli-Q water solution and 20 grams of fresh, clean biomass were placed in an Erlenmeyer flask, which was then incubated at 25 ºC for 72 hours while being stirred in the manner previously described. After incubation, the cell filtrates were obtained by filtering them through Whatman No. 1 filter paper. A 250 mL Erlenmeyer flask was filled with cell filtrate (50 mL), and the final concentration of 1 mM AgNO3 (0.017 g/100 mL) was added. The flasks were stirred and incubated in the same manner as previously stated for 120 hours at 25 °C in the dark. The nano silver solution changed color to a brownish-yellow tint and was stored at room temperature in screw-capped vials [111]. Similarly, Metarhizium spores were used to create silver nanoparticles. Metarhizium anisopliae spores were collected by washing with sterile distilled water and gentle brushing with a paintbrush without affecting mycelia growth. After that, sterile filters were used to filter the spore suspension. 105 conidia/mL concentration was adjusted as a standard inoculum. In the studies, 105 spores/ml of medium were inoculated at 32 °C and agitated at 120 rpm for 72 hours in 250 ml Erlenmeyer flasks containing 100 ml of potato dextrose broth. The fungal biomass was cleaned before being transferred to a 250 ml Erlenmeyer flask with 100 ml of double-distilled water and incubated for 72 hours in an orbital shaker at 100 rpm (28±4 °C). Following incubation, Whatman No. 1 filter paper was used to separate the fungal filtrate. 100 ml of fungal extract was mixed with 1 mM silver nitrate solution (0.16987 mg/ml,1 mM) and kept in an orbital shaker at 100 rpm (28±4 °C). A brownish-yellow solution indicated the presence of silver nanoparticles [105]. The synthesis of nanosilver from entomopathogenic fungi has been shown in Figure 2.
Silver nanoparticles have also been synthesized using Trichoderma longibrachiatum. The optimum conditions were 40°C temperature, 24 hours time period, pH 12, 3 g of biomass, 4 millimolar of AgNO3, and 150 rev/min, for the mycosynthesis AgNPs, which were subsequently purified by lyophilization. These ideal circumstances produced 3 g of AgNPs, which were then subjected to characterization techniques. The average size of the synthesized AgNPs and their respective zeta potential values of 1775 nm and 268 mV were discovered using dynamic light scattering (DLS) which demonstrated their stability. According to the X-ray diffraction (XRD) pattern, the mycosynthesized AgNPs with an average size of 61 nm were crystallin. The field emission scanning electron microscope (FESEM) and high-resolution transmission electron microscope (HRTEM) revealed non-agglomerated cuboidal, spherical, and triangular AgNPs with sizes ranging from 5 to 11.05 nm (HRTEM). Fourier transform infrared spectroscopy (FTIR) confirmed the presence of mycelial cell-free filtrate as a reducing and capping agent [112]. Penicillium oxalicum, a fungal endophyte found on the leaf of the Amoorarohituka plant, has also been utilized for the biofabrication of nanosilver. After the synthesis the surface plasmon resonance of AgNPs and the reduction of silver salt caused sharp UV-visible spectra to arise at 420 nm. The presence of functional groups in P. oxalicum, which were responsible for the reduction of silver salt into silver nanoparticles, was confirmed by FTIR analysis. Through XRD examination, a high level of crystallinity was discovered, and microscopy-based characterizations like AFM, TEM, and FESEM indicated evenly distributed, spherically formed nanoparticles [113]. Finally, Penicillium notatum was employed to generate silver nanoparticles in a recent investigative work. P. notatum produces the most nanoparticles after 48 hours of natural biogenesis; when the microwave-aided biogenesis method is utilised (20 minutes), this output is inhibited. The average size of the nanoparticles, revealed by SEM, XRD, and EDX peaks, was 55-65 nm, indicating a face-centered cubic shape. Furthermore, it has been established that the stability of AgNPs has improved with time. Table 1 depicts different entomopathogenic fungi employed fo the synthesis of nanosilver
6. Effects of Mycosynthesized Silver Nanoparticles: A Greener Approach in Controlling Pest Species
Nowadays entomopathogenic-based silver nanoparticles can be used as biopesticides to kill the pest. AgNPs are frequently used in the field of medicine and agri-food applications due to their versatile nature. AgNPs synthesized by mycogenic processes are used to control pest population as it has the potential to be antifeedant, larvicidal, and environmentally friendly. NPs treatment slowed insect growth and prolonged the larval period by up to 4 days compared to larvae fed with fungus cell-free extract and untreated castor leaves. Trichoderma harzianum, an entomopathogenic fungus, produced Ag nanoparticles is reported to show the highest mortality rates against Aedes aegypti which is a vector of dengue 92% for the 1st and 96% for 2nd instars and 100% for the 3rd and 4th instars, as well as pupa respectively [114]. The effect of silver nanoparticles produced by the insect pathogenic fungus Beauveria bassiana on mustard aphids was investigated in a study. The bioefficacy of 25 B. bassiana isolates against mustard aphids was tested. The isolates were divided into two groups based on the mortality rate of the mustard aphid: Group I (0-50%; 20 isolates) and Group II (>50%; 20 isolates) (51-100 percent; 5 isolates). This study, which is possibly the first report on the synthesis of nanosilver using the entomopathogenic fungus B. bassiana and its effectiveness against (L. erysimi) mustard aphid, suggests the potential application of silver nanoparticles for insect management in agriculture [106].
At 100 ppm concentration, nanoparticles demonstrated 100% antifeedant action and pupicidal activity against 1, 2, and 3 instar larvae in Helicoverpaarmigera treated with Trichoderma viride-mediated nanoparticles. After exposure to nanoparticles, glutathione-S-transferase activity increased, and -the glucosidase enzyme level was reduced in third-instar larval H. armigera [115]. Silver nanoparticles-loaded fungal metabolites nanoconjugate exhibit high mortality against Spodoptera litura. Nanoconjugate shows 100%, 100%, 91.2%,84.3%, 78.4%, and 71.3% mortality against first, second, third, fourth, fifth, and sixth instar larvae respectively [116]. Nanoparticles produced by Metarhizium anisopliae have insecticidal properties as well. Against G. mellonella, nanoparticles displayed a 22 percent larvicidal activity, but a synergistic interaction between the nanoparticles and gamma-irradiated M. anisopliae (which had greater amylase, nitrate reductase, protease, and lipase activities) resulted in larval mortality rate 82% [117].
Another method for producing nanoparticles with an insecticidal activity using these microorganisms is to use fungal extracellular enzymes to synthesize silver nanoparticles (AgNPs). In a study, entomopathogenic fungal isolates were used to assess biomass production and the ability to synthesize silver nanoparticles. The study used sixteen isolates of entomopathogenic fungus. An insect model, Tenebrio molitor, was used to test the pathogenicity and virulence of the fungi at a concentration of 5×106 spores/mL. The study confirmed that B. bassiana, M. anisopliae, and I. fumosorosea isolates are effective anti-T.molitor candidates. [118]. Lastly, using Fusarium pallidoroseum biomass, mycogenic AgNPs were synthesized and tested for efficacy against third-instar white grubs (Holotrichia sp), a significant sugarcane pest in western Uttar Pradesh (India). AgNPs were utilised to treat third-instar white grub larvae in vitro, and the lethal dosage (LD50) was calculated using Probit analysis. The chi-square test was used to further evaluate the results and determined that the LD50 was significant at 0.05 levels. The purpose of the study was to demonstrate how AgNPs have a larvicidal effect on white grubs. This study strongly supports the effectiveness of AgNPs as white grub control agents, which may encourage the future replacement of dangerous chemical pesticides [119]. The fungal-mediated SeNPs showed high larvicidal efficacy against the larva of S.lituria. The mortality percentage was found to be directly proportional to the concentration of the selenium nanoparticles starting from 25ppm – 100 ppm. Remarkable high mortality of 78.49% was observed at the concentration of 100ppm against the larvae of S.lituria compared to diet treated with the fungal extract which only gave a moderate mortality of 45.72% and 51.05% at the concentration of 100 ppm. Additionally, during the period of larvicidal activity testing, SeNP interference affects the growth of the larva with them progressively smaller in size along with a variety of anomalies being observed. The treatment also rendered the larva unable to reach the further stages of growth development of their lifecycle [94,120]. Table 2 explains the efficacy of entomopathogenic fungi against wide variety of pest species
6.1. Mode of Action of Mycosynthesized Nanoparticles
Nanoparticles are nano-sized metal oxides ranging from 1nm to 100 nm. these particles are well known for their insecticidal properties because of their shape, size, greater strength, depth qualities, optical properties, high electrical conductivity, and high concoction reactivity [126]. But even after learning about its effective insecticidal properties, information on its mode of entry, penetration, and action against insects is restricted. Few research papers related to cytotoxicity and genotoxicity have mentioned that the uptake of nanoparticles or coated nanoparticles is size dependent. according to their size nanoparticles follow different pathways to enter the cell. Smaller particles are taken up via the endocytic pathway, large particles of size 500 nm are processed by the phagocytic pathway, and large aggregates are taken up via micropinocytosis [127]. In order to protect crops from pests many eco-friendly control methods and biocontrol agents are being employed in agriculture but the efficacy rate of these biocontrol agents (bacteria, fungus, plant extract, and virus) is less. To enhance their effect these biocontrol agents are being combined with nanoparticles, by implementing nano-encapsulation efficiently [20]. Whether it is a metal ion or active bio ingredient that is carried by nanoparticles is causing penetration and toxicity is still not known. The exposed biosynthesized nanoparticle can enter insects through contact, inhalation, or oral route [128]. It has been mentioned that nanoparticles can penetrate insects through the skin’s outer layer and move through spiracles, mouth openings, anal prologs, abdominal prolegs, and setae [129]. While nanoparticles enter into cuticles adversely affect the physiology and morphology of insect. Nanostructured alumina sorb the wax layer by surface effect and using triboelectric forces leading to cuticle dehydration [130]. P. anisum essential oil-based nanoemulsion showed pigmentation changes in the cuticle, muscular destruction, cellular detritus, thickness, and necrosis in the epidermis, and lost the distinction between the exocuticle and endocuticle in T. castaneum [131]. It was also demonstrated that nanosilver causes melanin cuticular pigment loss and downregulation of Cu-dependent enzymes (tyrosinase and Cu-Zn superoxide dismutase) and that nanosilver combined with membrane-bound Cu transporter proteins causes Cu sequestration, simulating starvation of Cu and vertical flight ability reduction in D.melanogaster [132]. Carbon-silver nanohybrids bind to phosphorus and/or sulfur in biological structures such as DNA and proteins leading to blackened heads and disrupted cell, cuticle membrane organization, and guts in Culex quinquefasciatus and Anopheles stephensi [133]. It was also observed that when Chironomus riparius is exposed to commercial nanosilver it causes abnormality in the expression level of glutathione S-transferase (GST) genes which is related to oxidative stress and nanosilver toxicity also resulted in the downregulation of CrL15, a ribosomal protein gene, which controls the arrangement of ribosomes and hence protein formation. Moreover, CrGnRH1, a gonadotrophin-releasing hormone gene, and CrBR2.2, a Balbiani ring protein gene were upregulated, indicating the initiation of insect’s defense mechanism against silver nanoparticles, gonadotrophin-releasing hormone induced signal transduction pathways and reproductive failure [134]. AgNPs ingestion in early larval stages has been linked to impaired crawling and climbing abilities in later larval and adult stages. Increased AgNPs oral dosage during the larval stage causes metabolic changes in protein, carbohydrate, and lipid levels, as well as a reduction in the presence of lipid droplets and an increase in ROS in the larval tissue [135]. AgNPs increase oxidative stress in D. melanogaster midgut cells and increase the expression of hsp70 and hsp22, which are proteins found in the endoplasmic reticulum and mitochondria, respectively, and are also in charge of activating caspases, which regulate programmed cell death, membrane destabilization, and potential mitochondrial membrane loss [136]. Figure 3 illustrates the mode of entry of EPF-based nanoparticles through spiracles and its toxicity in insects.
7. Conclusions
Agriculture pests can substantially reduce crop yield and quality. Chemical pesticides have been heavily applied as the main strategy for pest control since the 1960s. Although that pesticides have been effective at reducing pest populations, it is generally known that their use can have adverse impacts on the environment and the crops themselves. For sustainable crop production, eco-friendly management is therefore required. As they have been proven to be successful in pest management and the production of sustainable agricultural goods, biopesticides can be used as an alternative to chemical pesticides. However, up to now, their efficiency has been constrained by technical difficulties. This review article emphasizes the importance of entomopathogenic fungi in controlling the pest population. In this case, an emerging topic of study called effective nano pesticides for plant protection and pest control offers fresh approaches to designing active chemicals at the nanoscale. Previous studies confirmed that metal nanoparticles are effective against a wide variety of insects and pests. Out of all the metal nanoparticles, silver nanoparticles have been considered an excellent insecticidal agent. Nowadays biologically synthesized silver nanoparticles are gaining a lot of popularity. Biosynthesized nanoparticles are an ecologically safe method of managing insect pest populations. At very low concentrations, entomopathogenic fungi-based nanosilver is particularly efficient against a wide range of pest species. Although still not much research has been conducted on this topic. The major goal of this review is to emphasize the rapidly developing field of nanotechnology by emphasizing its potential to offer environmentally friendly and effective options for the management of insect pests in agriculture without endangering the environment. The traditional methods of controlling insect pests, the drawbacks of chemical pesticides, and the potential of EPF-nanosilver as a novel candidate for pest control were also covered in this paper. Finally, The review necessitates the need for further studies so that the exact mode of action of EPF-nanosilver could be studied in detail which would also help in their field application.
Author Contributions
A.N.B.;K.A.A-E; J.M.A.-K. and P.N. conceived of this review. F.K.A-E., K.A.A-E.M.A.-K. helped in funding acquisition and the revision of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by The Deanship of Scientific Research, Vice Presidency for Graduate Studies and Scientific Research, King Faisal University, Saudi Arabia.
References
- Dhaliwal, G.S.; Jindal, V; Dhawan, A. K. Insect pest problems and crop losses: changing trends. Indian J Ecol. 2007, 37, 1–7. [Google Scholar]
- Sharma, S.; Kooner, R.; Arora, R. Insect pests and crop losses. In Breed Insect Resistant Crops Sustain Agri, 2017, 45-66.
- Lawler, S. P. Environmental safety review of methoprene and bacterially-derived pesticides commonly used for sustained mosquito control. Ecotoxicol. Environ. Saf. 2017, 139, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Rust, M.K. ., Lance, W.; Hemsarth, H. Synergism of the IGRs methoprene and pyriproxyfen against larval cat fleas (Siphonaptera: Pulicidae). J. Med. Entomol. 2016, 53, 629–633. [Google Scholar] [CrossRef] [PubMed]
- Tunaz, H.; Uygun, N. Insect growth regulators for insect pest control. Turk J Agric For. 2004, 28, 377–387. [Google Scholar]
- Madhu, S. K.; Shaukath, A. K.; Vijayan, V. A. Efficacy of bioactive compounds from Curcuma aromatica against mosquito larvae. Acta tropica. 2010, 113, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Guo, L.; Maimaiti, Y.; et al. Entomopathogenic fungi as microbial biocontrol agent. Mol Plant Breed. 2012. [CrossRef]
- Jaihan, P.; Sangdee, K.; Sangdee, A. Selection of entomopathogenic fungus for biological control of chili anthracnose disease caused by Colletotrichum spp. Eur J Plant Pathol. 2016, 146, 551–564. [Google Scholar] [CrossRef]
- Araújo, J.P.; Hughes, D. P. Diversity of entomopathogenic fungi: which groups conquered the insect body? Adv Genet. 2016, 94, 1–39. [Google Scholar]
- Castro, T.; Mayerhofer, J.; Enkerli, J. Persistence of Brazilian isolates of the entomopathogenic fungi Metarhiziumanisopliae and M. robertsii in strawberry crop soil after soil drench application. Agric Ecosyst Environ. 2016, 233, 361–369. [Google Scholar] [CrossRef]
- Rı’os-Moreno, A.; Garrido-Jurado, I.; Resquı´n-Romero, G et al. Destruxin A production by Metarhiziumbrunneum strains during transient endophytic colonisation of Solanum tuberosum. Biocontrol Sci Technol. 2016, 26, 1574–15. [Google Scholar] [CrossRef]
- Mascarin, GM.; Jaronski, ST. The production and uses of Beauveriabassiana as a microbial insecticide. World J Microbiol Biotechnol. 2016, 32, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Lovett, B. ; Leger, RJS (2017) The insect pathogens. Microbiol Spectr. 2017, 5, 2–5. [Google Scholar] [CrossRef] [PubMed]
- Litwin, A.; Nowak, M.; Różalska, S. Entomopathogenic fungi: unconventional applications. Rev Environ Sci Bio/Technol. 2020, 19, 23–42. [Google Scholar] [CrossRef]
- Skinner, M., Parker, BL.; Kim, JS. Role of entomopathogenic fungi. In: Abrol DP (ed) Integrated pest management, 2014, Academic Press, Cambridge, pp 169–191.
- Donzelli, BGG. ; Krasnoff, SB.; Churchill, ACL. Identification of a hybrid PKS–NRPS required for the biosynthesis of NG-391 in Metarhiziumrobertsii. Curr Genet. 2010, 56, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Maina, U. M.; Galadima, I. B.; Gambo, F. M.; Zakaria, D. A review on the use of entomopathogenic fungi in the management of insect pests of field crops. J. Entomol. Zool. Stud. 2018, 6, 27–32. [Google Scholar]
- Smith, A. M.; Duan, H.; Rhyner, M. N.; Ruan, G.; Nie, S. A systematic examination of surface coatings on the optical and chemical properties of semiconductor quantum dots. Phys Chem Chem Phys. 2006, 8, 3895–3903. [Google Scholar] [CrossRef]
- Goswami, A.; Roy, I.; Sengupta, S.; Debnath, N. Novel applications of solid and liquid formulations of nanoparticles against insect pests and pathogens. Thin Solid Films. 2010, 519, 1252–1257. [Google Scholar] [CrossRef]
- Bhattacharyya A, Duraisamy P, Govindarajan M, Buhroo AA, and Prasad R, Nano biofungicides: emerging trend in insect pest control. In Advances and applications through fungal nanobiotechnology, ed. by Prasad R. Springer International Publishing, Switzerland, 2016, 307–319.
- Cantu, A.A. Nanoparticles in forensic science. In Optics and Photonics for Counterterrorism and Crime Fighting IV, 2008, 7119, 71190F. [Google Scholar]
- Li, H.; Li, F.; Wang, L.; Sheng, J.; Xin, Z.; Zhao, L. . & Hu, Q. Effect of nano-packing on preservation quality of Chinese jujube (Ziziphus jujuba Mill. var. inermis (Bunge) Rehd). Food Chem. 2009, 114, 547–552. [Google Scholar]
- Yokesh Babu, M.; Janaki Devi, V.; Ramakritinan, C.M.; Umarani, R.; Taredahalli, N.; Kumaraguru, A.K. Application of biosynthesized silver nanoparticles in agricultural and marine pest control. Curr Nanosci. 2014, 10, 374–381. [Google Scholar] [CrossRef]
- Guilger-Casagrande, M.; Lima, R.D. Synthesis of silver nanoparticles mediated by fungi: a review. Front Bioeng Biotechnol. 2019, 7, 287. [Google Scholar] [CrossRef] [PubMed]
- Hazaa, M.; Alm-Eldin, M.; Ibrahim, A.E.; Elbarky, N.; Salama, M.; Sayed, R.; Sayed, W. Biosynthesis of Silver Nanoparticles using Borago officinslis leaf extract, characterization and larvicidal activity against cotton leaf worm, Spodoptera littoralis (Bosid). Int J Trop Insect Sci. 2021, 41, 145–156. [Google Scholar] [CrossRef]
- Sinha, K.K.; Choudhary, A.K.; Kumari, P. Entomopathogenic fungi. In Ecofriendly pest management for food security, 2016, 475–505.
- Lin, B.X. ‘Use of Beauveria bassiana against the Sweet Potato Weevil’, ActaEntomologicaSinica. 1956, 6, 539540 (in Chinese).
- Xu, Q.F. ‘Study on Soybean Moth Control by Beauveria bassiana’, ActaEntomologicaSinica. 1959, 9, 203215 (in Chinese).
- Quesada-Moraga, E.; Carrasco-Diaz, J.A.; Santiago-Á, lvarez C. Insecticidal and antifeedant activities of proteins secreted by entomopathogenic fungi against Spodoptera littoralis (Lep., Noctuidae). J Appl Entomol. 2006, 130, 442–452. [Google Scholar] [CrossRef]
- Vega, F.E.; Infante, F.; Castillo, A.; Jaramillo, J. The coffee berry borer, Hypothenemushampei (Ferrari)(Coleoptera: Curculionidae): a short review, with recent findings and future research directions. 2009. [Google Scholar]
- BADII, M.H.; ABREU, J.L. Control biológicouna forma sustentable de control de plagas. Int J Good Consci. 2006, 1, 82–89. [Google Scholar]
- Behie, SW.; Bidochka, MJ. Ubiquity of insect-derived nitrogen transfer to plants by endophytic insect-pathogenic fungi: an additional branch of the soil nitrogen cycle. Appl Environ Microbiol. 2014, 80, 1553–1560. [Google Scholar] [CrossRef]
- DELGADO, P.A.M.; MURCIA, O.P. Hongosentomopatógenos: umaalternativa para la obtención de Biopesticidas. Ambi-Agua. 2011, 6, 77–90. [Google Scholar] [CrossRef]
- Tanzini, M.; Alves, S.; Setten, A.; Augusto, N. Compatibilidad de agent estensoactivos com Beauveria bassiana y Metarhizium anisopliae. ManejoIntegrado De Plagas. 2001, 59, 15–18. [Google Scholar]
- Mantzoukas, S.; Eliopoulos, P.A. Endophytic entomopathogenic fungi: A valuable biological control tool against plant pests. Appl Sci. 2020, 10, 360. [Google Scholar] [CrossRef]
- Sharma, A.; Sood, K.; Kaur, J.; Khatri, M. Agrochemical loaded biocompatible chitosan nanoparticles for insect pest management. Biocatal Agri Biotechnol. 2019, 18, 101079. [Google Scholar] [CrossRef]
- Deka, B.; Baruah, C.; Babu, A. Entomopathogenic microorganisms: their role in insect pest management. Egypt J Biol Pest Control. 2021, 31, 1–8. [Google Scholar] [CrossRef]
- Umaru, F.F.; Simarani, K. Efficacy of Entomopathogenic Fungal Formulations against Elasmolomuspallens (Dallas) (Hemiptera: Rhyparochromidae) and Their Extracellular Enzymatic Activities. Toxins. 2022, 14, 584. [Google Scholar] [CrossRef] [PubMed]
- Tennant, P.F.; Fermin, G.A.; Roye, M.E. Viruses infecting papaya (Carica papaya L.): etiology, pathogenesis, and molecular biology. Plant Viruses. 2007, 1, 178–188. [Google Scholar]
- Chakrabarti, S.; Raychaudhuri, D.N. New and little known aphids (Homoptera: Aphididae) from Kumaon Himalaya, India. Entomon. 1978, 3, 95–103. [Google Scholar]
- Mukherjee, A. , Debnath, P., Ghosh, S.K.; Medda, P.K. Biological control of papaya aphid (Aphis gossypii Glover) using entomopathogenic fungi. Vegetos. 2020, 33, 1–10. [Google Scholar] [CrossRef]
- Sani, I.; Ismail, S.I.; Abdullah, S.; Jalinas, J.; Jamian, S.; Saad, N. A review of the biology and control of whitefly, Bemisiatabaci (Hemiptera: Aleyrodidae), with special reference to biological control using entomopathogenic fungi. Insects. 2020, 11, 619. [Google Scholar] [CrossRef]
- Filgueiras, C.C.; Willett, D.S. The Lesser Chestnut Weevil (Curculio sayi): Damage and Management with Biological Control Using Entomopathogenic Fungi and Entomopathogenic Nematodes. Insects. 2022, 13, 1097. [Google Scholar] [CrossRef]
- Donzelli, BGG.; Krasnoff, SB. Molecular genetics of secondary chemistry in Metarhizium Fungi. In Lovett B, Leger RJS (eds) Advances in genetics. 2016, 94, 365–436.
- Gibson, D.M.; Donzelli, B.G.; Krasnoff, S.B.; Keyhani, N.O. Discovering the secondary metabolite potential encoded within entomopathogenic fungi. Nat Prod Rep. 2014, 31, 1287–1305. [Google Scholar] [CrossRef]
- Pedrini, N. The entomopathogenic fungus Beauveria bassiana shows its toxic side within insects: expression of genes encoding secondary metabolites during pathogenesis. J Fungi. 2022, 8, 488. [Google Scholar] [CrossRef] [PubMed]
- Zibaee, A.; Bandani, A.R.; Talaei-Hassanlouei, R.; Malagoli, D. Cellular immune reactions of the sunn pest, Eurygasterintegriceps, to the entomopathogenic fungus, Beauveria bassiana and its secondary metabolites. J Insect Sci. 2011, 11, 138. [Google Scholar] [CrossRef] [PubMed]
- Freed, S.; Feng-Liang, J.; Naeem, M.; Shun-Xiang, R.; Hussian, M. Toxicity of proteins secreted by entomopathogenic fungi against Plutellaxylostella (Lepidoptera: Plutellidae). Int J Agri Biol. 2012, 14. [Google Scholar]
- Vinayaga Moorthi, P.; Balasubramanian, C.; Selvarani, S.; Radha, A. Efficacy of sub lethal concentration of entomopathogenic fungi on the feeding and reproduction of Spodoptera litura. Springerplus. 2015, 4, 1–12. [Google Scholar] [CrossRef] [PubMed]
- El-Sawy, M.; Mostafa, E.H.; Ismail, N.A.E.R. Secondary metabolites of the entomopathogenic fungus, Cladosporium cladosporioides and its relation to toxicity of cotton aphid, Aphis gossypii (Glov. ). Int J. 2019, 5, 115–120. [Google Scholar]
- Woo, R.M.; Park, M.G.; Choi, J.Y.; Park, D.H.; Kim, J.Y.; Wang, M.; Kim, H.J.; Woo, S.D.; Kim, J.S.; Je, Y.H. Insecticidal and insect growth regulatory activities of secondary metabolites from entomopathogenic fungi, Lecanicilliumattenuatum. J Appl Entomol. 2020, 144, 655–663. [Google Scholar] [CrossRef]
- Arunthirumeni, M.; Vinitha, G.; Shivakumar, M.S. Antifeedant and larvicidal activity of bioactive compounds isolated from entomopathogenic fungi Penicillium sp. for the control of agricultural and medically important insect pest (Spodoptera litura and Culex quinquefasciatus). Parasitol Int. 2023, 92, 102688. [Google Scholar] [CrossRef]
- Sevim, A.; Sevim, E.; Demirbağ, Z. General biology of entomopathogenic fungi and their potential to control pest species in Turkey (EntomopatojenikfunguslarıngenelbiyolojileriveTürkiye’dezararlıböceklerinmücadelesindekullanılmapotansiyelleri). Erzincan Üniversitesi Fen BilimleriEnstitüsüDergisi. 2015, 8, 115–147. [Google Scholar] [CrossRef]
- S. O. Andersen, Biochemistry of insect cuticle. Annu. Rev. Entomol. 1979, 29–59. [Google Scholar]
- Ross, H. H. A textbook of entomology. A Textbook of Entomology. 1948. [Google Scholar]
- Glare, T. 11 Entomopathogenic Fungi and their Role in Regulation of Insect Populations. Insect Control. 2010. [Google Scholar]
- Hajek, A. E. ; St. Leger, R. J. Interactions between fungal pathogens and insect hosts. AnnRev Entomol. 1994, 39, 293–322. [Google Scholar]
- Ortiz-Urquiza, A. and Keyhani, N.O. Action on the surface: entomopathogenic fungi versus the insect cuticle. Insects. 2013, 4, 357–374. [Google Scholar] [CrossRef] [PubMed]
- Islam, W.; Adnan, M.; Shabbir, A.; Naveed, H.; Abubakar, Y.S.; Qasim, M.; Tayyab, M.; Noman, A.; Nisar, M.S.; Khan, K.A.; Ali, H. Insect-fungal-interactions: A detailed review on entomopathogenic fungi pathogenicity to combat insect pests. Microb. Pathog. 2021, 159, 105122. [Google Scholar] [CrossRef] [PubMed]
- Zacharuk, R. Y. Electron-microscope studies of the histopathology of fungal infections by Metarrhizium anisopliae. Misc. Publ. Entomol. Soc. Am. 1973, 9, 112–119. [Google Scholar]
- J. Moty ´ an, ´ F. Toth, ´ J. Tozs ˝ ´er, Research applications of proteolytic enzymes in molecular biology. Biomolecules 2013, 3, 923–942. [CrossRef] [PubMed]
- St Leger, R.J. The role of cuticle-degrading proteases in fungal pathogenesis of insects, Can. J. Bot. 1995, 73, 1119–1125. [Google Scholar]
- Pedrini, N.; Ortiz-Urquiza, A.; Huarte-Bonnet, C.; Zhang, S.; Keyhani, N.O. Targeting of insect epicuticular lipids by the entomopathogenic fungus Beauveria bassiana: hydrocarbon oxidation within the context of a host-pathogen interaction. Front. Microbiol. 2013, 4. [Google Scholar] [CrossRef]
- Pedrini, N.; Zhang, S.; Ju´ arez, M.P.; Keyhani, N.O. Molecular characterization and expression analysis of a suite of cytochrome P450 enzymes implicated in insect hydrocarbon degradation in the entomopathogenic fungus Beauveria bassiana, Microbiol. 2010, 156, 2549–2557.
- Suzuki, A.; Kawakami, K.; Tamura, S. Detection of destruxins in silkworm larvae infected with Metarrhizium anisopliae. Agric. Biol. Chem. 1971, 35, 1641–1643. [Google Scholar]
- Eyal, J.; Mabud, M.A.; Fischbein, K.L.; Walter, J.F.; Osborne, L.S.; Landa, Z. Assessment of Beauveria bassiana Nov. EO-1 strain, which produces a red pigment for microbial control, Appl. Biochem. Biotechnol. 1994, 44, 65–80. [Google Scholar] [CrossRef]
- Sweet, M.J.; Chessher, A.; Singleton, I. Metal-based nanoparticles; size, function, and areas for advancement in applied microbiology. Adv. Appl. Microbiol. 2012, 80, 113–142. [Google Scholar]
- Beltrán Pineda, M.E.; Lizarazo Forero, L.M.; Sierra, Y.C.A. Mycosynthesis of silver nanoparticles: a review. BioMetals. 2022, 1–32. [Google Scholar] [CrossRef]
- Iravani, S.; Korbekandi, H.; Mirmohammadi, SV.; Zolfaghari, B. Synthesis of silver nanoparticles: chemical, physical and biological methods. Res. Pharma. Sci. 2014, 9, 385. [Google Scholar]
- Kumar, S.; Bhushan, P.; Bhattacharya, S. Fabrication of nanostructures with bottom-up approach and their utility in diagnostics, therapeutics, and others. In: Environmental Chemical and Medical Sensors, Springer: Singapore, 2018; pp 167-198.
- Prabhu, S.; Poulose, E.K. Silver nanoparticles: mechanism of antimicrobial action, synthesis, medical applications, and toxicity effects. Int. nano lett. 2012, 2, 1–10. [Google Scholar] [CrossRef]
- Bhattacharya, D.; Gupta, R. Nanotechnology and potential of microorganisms. Crit. Rev. Biotechnol. 2005, 25, 25,199–201. [Google Scholar] [CrossRef]
- Thakkar, K.N.; Mhatre, S.S.; Parikh, R.Y. Biological synthesis of metallic nanoparticles. Nanomed: NanotechnolBiol Med. 2010, 6, 257–262. [Google Scholar] [CrossRef]
- Abbasi, E.; Milani, M.; Fekri, S.; Kouhi, M.; Akbarzadeh, A.; Nasrabadi, H.; Nikasa, P.; Joo, S.; Hanifehpour, Y.; Nejati-Koshki, K.; Samiei, M. International nano letters Silver nanoparticles: synthesis methods, bioapplications and properties. Crit Rev Microbiol. 2014, 42, 173–180. [Google Scholar]
- Keat, C.; Aziz, A. Eid A, Elmarzugi A Biosynthesis of nanoparticles and silver nanoparticle. Bioresour. Bioprocess. 2015. [Google Scholar] [CrossRef]
- Bhainsa, K.C.; D’Souza, S.F. Extracellular biosynthesis of silver nanoparticles using the fungus Aspergillus fumigatus. Colloids Surf B Biointerfaces. 2006, 47, 160–164. [Google Scholar] [CrossRef]
- Zhang, X. , Yan, S., Tyagi, R.D., Surampalli, R.Y. Synthesis of nanoparticles by microorganisms and their application in enhancing microbiological reaction rates. Chemosphere. 2011, 82, 489–494. [Google Scholar] [CrossRef]
- Kudesia, N. , Najitha Banu, A., Raut, A.M. and Wahengbam, J., Biofabricated Nanoparticles: A Greener Approach Towards Insect Control. In Advances in Integrated Pest Management Technology: Innovative and Applied Aspects; Cham: Springer International Publishing. 2019, 391–419.
- Guilger-Casagrande, M.; Lima, R.D. Synthesis of silver nanoparticles mediated by fungi: a review. Front. Bioeng. Biotechnol. 2019, 7, 287. [Google Scholar] [CrossRef]
- Gade, A. K.; Bonde, P.; Ingle, A. P.; Marcato, P. D.; Durán, N.; Rai, M. K. Exploitation of Aspergillus niger for synthesis of silver nanoparticles. J. Biobased Mater. Bioenergy. 2008, 2, 243–247. [Google Scholar] [CrossRef]
- Velusamy, P.; Kumar, G.V.; Jeyanthi, V.; Das, J.; Pachaiappan, R. Bio-inspired green nanoparticles: synthesis, mechanism, and antibacterial application. Toxicol. Res. 2016, 32, 95–102. [Google Scholar] [CrossRef]
- Park, Y.; Hong, Y.N.; Weyers, A.; Kim, Y.S.; Linhardt, R. J. Polysaccharides and phytochemicals: a natural reservoir for the green synthesis of gold and silver nanoparticles. IET Nanobiotechnol. 2011, 5, 69–78. [Google Scholar] [CrossRef]
- Vela´zquez-Robledo, R.; Contreras-Cornejo, H.A.; Macı´asRodrı´guez, L.; Herna´ndez-Morales, A.; Aguirre, J.; Casas-Flores, S.; Herrera-Estrella, A. Role of the 4-phosphopantetheinyl transferase of Trichoderma virens in secondary metabolism and induction of plant defense responses. Mol. Plant Microbe Interact. 2011, 24, 1459–1471. [Google Scholar] [CrossRef]
- Iranifam, M.; Fathinia, M.; Rad, T. S.; Hanifehpour, Y.; Khataee, A. R.; Joo, S. W. Talanta. A novel selenium nanoparticles-enhanced chemiluminescence system for determination of dinitrobutylphenol. Talanta 2013, 107, 263–269. [Google Scholar] [CrossRef]
- Contreras-Cornejo, H. A.; Macı´as-Rodrı´guez, L.; HerreraEstrella, A.; Lo´pez-Bucio, J. The 4-phosphopantetheinyl transferase of Trichoderma virens plays a role in plant protection against Botrytis cinerea through volatile organic compound emission. Plant Soil. 2014, 379, 261–274. [Google Scholar] [CrossRef]
- Berini, F.; Caccia, S.; Franzetti, E.; Congiu, T.; Marinelli, F.; Casartelli, M.; Tettamanti, G. Effects of Trichoderma viride chitinases on the peritrophic matrix of Lepidoptera. Pest Manag. Sci. 2016, 72, 980–989. [Google Scholar] [CrossRef]
- Wadhwani, S.A.; Shedbalkar, U.U.; Singh, R.; Chopade, B. A. Biogenic selenium nanoparticles: current status and future prospects. Appl. Microbiol. Biotechnol. 2016, 100, 2555–2566. [Google Scholar] [CrossRef]
- Saravanakumar, K.; Li, Y.; Yu, C.; Wang, Q.Q.C.; Wang, M.; Sun, J.; Chen. Effect of Trichoderma harzianum on maize rhizosphere microbiome and biocontrol of Fusarium Stalk rot. J. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Martı´nez-Medina, Appels, F. V.; van Wees, S. C. Impact of salicylic acid-and jasmonic acid-regulated defences on root colonization by Trichoderma harzianum T-78. Plant Signal. Behav. 2017, 12, e1345404. [Google Scholar]
- Arunthirumeni, M. , Veerammal, V., & Shivakumar, M. S. Biocontrol efficacy of mycosynthesized selenium nanoparticle using Trichoderma sp. on insect pest Spodoptera litura. J. Clust. Sci. 2022, 33, 1645–1653. [Google Scholar]
- Pimentel, D. J. Amounts of Pesticides Reaching Target Pests: Environmental Impacts and Ethics. Agric. Environ. Ethics 1995, 8, 17–29. [Google Scholar] [CrossRef]
- Sengottayan, S. N. [HTML] Physiological and biochemical effect of neem and other Meliaceae plants secondary metabolites against Lepidopteran insects. Front. Physiol. 2013, 4, 359. [Google Scholar]
- Senthil-Nathan. A Review of Biopesticides and Their Mode of Action Against Insect Pests. A Review of Biopesticides and Their Mode of Action Against Insect Pests. Environ. Sustain. 2015, 49–63. [Google Scholar]
- Dhivya, K.; Vengateswari, G.; Arunthirumeni, M.; Karthi, S.; Senthil-Nathan, S.; Shivakumar, M. S. Bioprospecting of Prosopis juliflora (Sw.) DC seed pod extract effect on antioxidant and immune system of Spodoptera litura (Lepidoptera: Noctuidae). Physiol. Mol. Plant Pathol. 2018, 101, 45–53. [Google Scholar] [CrossRef]
- Tamayo, L.; Azócar, M.; Kogan, M.; Riveros, A.; Páez, M. Copper-polymer nanocomposites: An excellent and cost-effective biocide for use on antibacterial surfaces. Mater. Sci. Eng. 2016, 69, 1391–1409. [Google Scholar] [CrossRef]
- Kora, A.J. Copper-based nanopesticides. Copper Nanostructures: Next-Generation of Agrochemicals for Sustainable Agroecosystems. 2022, 133–153. [Google Scholar]
- Vivekanandhan, P.; Swathy, K.; Thomas, A.; Kweka, E. J.; Rahman, A.; Pittarate, S.; Krutmuang, P. Insecticidal efficacy of microbial-mediated synthesized copper nano-pesticide against insect pests and non-target organisms. Int J Environ Res Public Health. 2021, 18, 10536. [Google Scholar] [CrossRef]
- Rafeeq, C. M.; Paul, E.; Saagar, E. V.; Ali, P. M. Mycosynthesis of zinc oxide nanoparticles using Pleurotus floridanus and optimization of process parameters. Ceram. Int. 2021, 47, 12375–12380. [Google Scholar] [CrossRef]
- Kamal, A.; Saba, M.; Ullah, K.; Almutairi, S. M.; AlMunqedhi, B. M.; Ragab abdelGawwad, M. Mycosynthesis, Characterization of Zinc Oxide Nanoparticles, and Its Assessment in Various Biological Activities. Crystals. 2023, 13, 171. [Google Scholar] [CrossRef]
- Shobha, B.; Lakshmeesha, T. R.; Ansari, M. A.; Almatroudi, A.; Alzohairy, M. A.; Basavaraju, S.; Chowdappa, S. Mycosynthesis of ZnO nanoparticles using Trichoderma spp. isolated from rhizosphere soils and its synergistic antibacterial effect against Xanthomonas oryzaepv. oryzae. J. Fungi. 2020, 6, 181. [Google Scholar] [CrossRef]
- Rodrigues, A.G.; Ping, L.Y.; Marcato, P.D.; Alves, O.L.; Silva, M.C.; Ruiz, R.C.; Melo, I.S.; Tasic, L.; De Souza, A.O. ; Biogenic antimicrobial silver nanoparticles produced by fungi. Appl.Microbial. Biotechnol. 2013, 97, 775–782. [Google Scholar] [CrossRef]
- Honary, S.; Barabadi, H.; Gharaei-Fathabad, E.; Naghibi, F. Green synthesis of silver nanoparticles induced by the fungus Penicillium citrinum. Trop. J. Pharm. Res. 2013, 12, 7–11. [Google Scholar] [CrossRef]
- Roy, S.; Mukherjee, T.; Chakraborty, S.; Das, T.K. Biosynthesis, characterisation & antifungal activity of silver nanoparticles synthesized by the fungus Aspergillus foetidus MTCC8876. Dig. J. Nanomater. Biostruct. 2013, 8, 197–205. [Google Scholar]
- Banu, A.N.; Balasubramanian, C. Optimization and synthesis of silver nanoparticles using Isaria fumosorosea against human vector mosquitoes. Parasitol. Res. 2014, 113, 3843–3851. [Google Scholar] [CrossRef]
- Amerasan, D.; Nataraj, T.; Murugan, K.; Panneerselvam, C.; Madhiyazhagan, P.; Nicoletti, M.; Benelli, G. Myco-synthesis of silver nanoparticles using Metarhizium anisopliae against the rural malaria vector Anopheles culicifacies Giles (Diptera: Culicidae). J. pest sci. 2016, 89, 249–256. [Google Scholar] [CrossRef]
- Kamil, D.; Prameeladevi, T.; Ganesh, S.; Prabhakaran, N.; Nareshkumar, R.; Thomas, S. P. Green synthesis of silver nanoparticles by entomopathogenic fungus Beauveria bassiana and their bioefficacy against mustard aphid (Lipaphiserysimi Kalt. ). 2017. [Google Scholar]
- Elamawi, R.M.; Al-Harbi, R.E.; Hendi, A.A. Biosynthesis and characterization of silver nanoparticles using Trichoderma longibrachiatum and their effect on phytopathogenic fungi. Egypt. j. biological pest control, 2018, 28, 1–11. [Google Scholar] [CrossRef]
- Hayat, P.; Khan, I.; Rehman, A.; Jamil, T.; Hayat, A.; Rehman, M.U.; Ullah, N.; Sarwar, A.; Alharbi, A.A.; Dablool, A.S.; Daudzai, Z. ; Myogenesis and Analysis of Antimicrobial Potential of Silver Nanoparticles (AgNPs) against Pathogenic Bacteria. Molecules, 2023, 28, 637. [Google Scholar] [CrossRef]
- Bérdy, J. Bioactive Microbial Metabolites. J Antibiot. 2005, 58, 1–26. [Google Scholar] [CrossRef]
- Banu, A.N.; Kudesia, N.; Raut, A.M.; Pakrudheen, I.; Wahengbam, J. Toxicity, bioaccumulation, and transformation of silver nanoparticles in aqua biota: A review. Environ. Chem. Letters. 2021, 19, 4275–4296. [Google Scholar] [CrossRef]
- Banu, A.N.; Balasubramanian, C. Myco-synthesis of silver nanoparticles using Beauveria bassiana against dengue vector, Aedes aegypti (Diptera: Culicidae). Parasitol. Res. 2014, 113, 2869–2877. [Google Scholar] [CrossRef]
- Omran, B.A.; Nassar, H.N.; Younis, S.A.; Fatthallah, N.A.; Hamdy, A.; El-Shatoury, E.H.; El-Gendy, N.S. ; Physiochemical properties of Trichoderma longibrachiatum DSMZ 16517-synthesized silver nanoparticles for the mitigation of halotolerant sulphate-reducing bacteria. J. appl.microbio. 2019, 126, 138–154. [Google Scholar] [CrossRef]
- Gupta, P.; Rai, N.; Verma, A.; Saikia, D.; Singh, S.P.; Kumar, R.; Singh, S.K.; Kumar, D.; Gautam, V. Green-Based Approach to Synthesize Silver Nanoparticles Using the Fungal Endophyte Penicillium oxalicum and Their Antimicrobial, Antioxidant, and In Vitro Anticancer Potential. ACS omega. 2022. [Google Scholar] [CrossRef]
- Sundaravadivelan, C.; Padmanabhan, M. N. Effect of mycosynthesized silver nanoparticles from filtrate of Trichoderma harzianum against larvae and pupa of dengue vector Aedes aegypti L. Environ. Sci. Pollut. Res. 2014, 21, 4624–4633. [Google Scholar] [CrossRef]
- Chinnaperumal, K.; Govindasamy, B.; Paramasivam, D.; Dilipkumar, A.; Dhayalan, A.; Vadivel, A. , Pachiappan, P. Bio-pesticidal effects of Trichoderma viride formulated titanium dioxide nanoparticle and their physiological and biochemical changes on Helicoverpa armigera (Hub. ). Pestic. Biochem. physiol. 2018, 149, 26–36. [Google Scholar] [CrossRef]
- Karthick Raja Namasivayam, S. , Arvind Bharani, R. S. Biocompatible silver nanoparticles-loaded fungal metabolites nanoconjugate (agnp–fm) preparation for the noteworthy pesticidal activity. Natl. Acad. Sci. Lett. 2021, 44, 511–517. [Google Scholar] [CrossRef]
- Yosri, M.; Abdel-Aziz, M.M.; Sayed, R.M. Larvicidal potential of irradiated myco-insecticide from Metarhizium anisopliae and larvicidal synergistic effect with its mycosynthesized titanium nanoparticles (TiNPs). J Radiat Res Appl Sci. 2018, 11, 328–334. [Google Scholar] [CrossRef]
- Santos, T.S.; Passos, E.M.D.; Seabra, M.G.D.J.; Souto, E.B.; Severino, P.; Mendonça, M.D.C. Entomopathogenic fungi biomass production and extracellular biosynthesis of silver nanoparticles for bioinsecticide action. Appl. Sci. 2021, 11, 2465. [Google Scholar] [CrossRef]
- Shukla, G.; Gaurav, S.S.; Singh, A.; Rani, P. Synthesis of mycogenic silver nanoparticles by Fusarium pallidoroseum and evaluation of its larvicidal effect against white grubs (Holotrichia sp. ). Mater. Today: Proc. 2022, 49, 3517–3527. [Google Scholar] [CrossRef]
- Kamalakannan, S.; Gobinath, C.; Ananth, S. Synthesis and characterization of fungus mediated silver nanoparticle for toxicity on filarial vector, Culex quinquefasciatus. Int. J. Pharm. Sci. Rev. Res. 2014, 24, 124–132. [Google Scholar]
- Salunkhe, R.B.; Patil, S.V.; Patil, C.D.; et al. Larvicidal potential of silver nanoparticles synthesized using fungus Cochliobolus lunatus against Aedes aegypti (Linnaeus, 1762) and Anopheles stephensi Liston (Diptera; Culicidae). Parasitol. Res. 2011, 109, 823–831. [Google Scholar] [CrossRef]
- Salunkhe, R.B.; Patil, S.V.; Patil, C.D.; Salunke, B.K. ; Larvicidal potential of silver nanoparticles synthesized using fungus Cochliobolus lunatus against Aedes aegypti (Linnaeus, 1762) and Anopheles stephensi Liston (Diptera; Culicidae). Parasitol. Res. 2011, 109, 823–831. [Google Scholar] [CrossRef]
- KHOOSHE-BAST, Z.; Sahebzadeh, N.; GHAFFARI-MOGHADDAM, M.; MIRSHEKAR, A. Insecticidal effects of zinc oxide nanoparticles and Beauveria bassiana TS11 on Trialeurodes vaporariorum (Westwood, 1856)(Hemiptera: Aleyrodidae). Acta AgriculturaeSlovenica. 2016, 107, 299–309. [Google Scholar] [CrossRef]
- Wang, X.; Xu, J.; Wang, X.; Qiu, B.; Cuthbertson, A.G.; Du, C.; Wu, J.; Ali, S.; Isaria fumosorosea-based zero-valent iron nanoparticles affect the growth and survival of sweet potato whitefly, Bemisia tabaci (Gennadius). Pest manag. Sci. 2019, 75, 2174–2181. [Google Scholar]
- Xu, J.; Zhang, K.; Cuthbertson, A. G.; Du, C.; Ali, S. Toxicity and biological effects of Beauveria brongniartii Fe0 nanoparticles against Spodoptera litura (Fabricius). Insects. 2020, 11, 895. [Google Scholar] [CrossRef]
- Sahayaraj, K.; Madasamy, M.; Radhika, S.A. Insecticidal activity of bio-silver and gold nanoparticles against PericalliariciniFab.(Lepidaptera: Archidae). J. Biopestic. 2016, 9, 63. [Google Scholar] [CrossRef]
- Foldbjerg, R.; Jiang, X; Miclăus, T. ; Chunying, C.; Autrup, H.; Beer, C. Silver nanoparticles—wolves in sheep’s clothing? Toxicol. Res. 2015, 4, 563–575. [Google Scholar]
- Shahzad, K.; Manzoor, F. Nanoformulations and their mode of action in insects: a review of biological interactions. Drug chem. toxicol. 2021, 44, 1–11. [Google Scholar] [CrossRef]
- Lade, B.D.; Gogle, D.P. Nano-biopesticides: Synthesis and applications in plant safety. Nanobiotechnol. Application Plant Prot. 2019, 2, 169–189. [Google Scholar]
- Stadler, T. et al., Nanostructured alumina: biocidal properties and mechanism of action of a novel insecticide powder. Bull. Insectology. 2017, 70, 17–25. [Google Scholar]
- Hashem, A.S.; et al. Pimpinella anisum essential oil nanoemulsions against Tribolium castaneum-insecticidal activity and mode of action. Environ. Sci. Pollut. Res. Int. 2018. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, N.; Ramamoorthy, M.; Lyon, D.; Jones, K.; Duttaroy, A. Mechanism of silver nanoparticles action on insect pigmentation reveals intervention of copper homeostasis. PLoS One 2013, 8, e53186. [Google Scholar] [CrossRef] [PubMed]
- Sultana, N.; et al. Nanoweapon: control of mosquito breeding using carbon-dot-silver nanohybrid as a biolarvicide. Environmental Chemistry Letters. 2018. [CrossRef]
- Nair, P.M.G.; Choi, J. Identification, characterization and expression profiles of Chironomus riparius glutathione S-transferase (GST) genes in response to cadmium and silver nanoparticles exposure. AquatToxicol. 2011, 101, 550–560. [Google Scholar] [CrossRef]
- Raj, A.; Shah, P.; Agrawal, N. ; Sedentary behavior and altered metabolic activity by AgNPs ingestion in Drosophila melanogaster. Scientific Reports. 2017, 7, 15617. [Google Scholar] [CrossRef]
- Pandey, A.; et al. Cellular internalization and stress response of ingested amorphous silica nanoparticles in the midgut of Drosophila melanogaster. Biochimica et Biophysica Acta. 2013, 1830, 2256–2266. [Google Scholar] [CrossRef]
Figure 1.
The top-down technique, in which the bulk substance shrinks in size to generate NPs, and the bottom-up method, in which atoms and molecules self-assemble to form nanoparticles, are the two methods for producing nanoparticles. The top-down strategy includes physical methods such as arc discharge, evaporation-condensation, and laser ablation, whereas the bottom-up approach includes chemical methods such as chemical reduction, micro-emulsion techniques, and electrochemical synthetic techniques. Physical and chemical techniques, despite their popularity, are limited in their application due to limitations such as high costs, toxicity, complexity, and age. NPs are created by a variety of biological processes, including fungi (microbes), bacteria, and plants. The biological process is inexpensive, produces a bulk amount, and is eco-friendly.
Figure 1.
The top-down technique, in which the bulk substance shrinks in size to generate NPs, and the bottom-up method, in which atoms and molecules self-assemble to form nanoparticles, are the two methods for producing nanoparticles. The top-down strategy includes physical methods such as arc discharge, evaporation-condensation, and laser ablation, whereas the bottom-up approach includes chemical methods such as chemical reduction, micro-emulsion techniques, and electrochemical synthetic techniques. Physical and chemical techniques, despite their popularity, are limited in their application due to limitations such as high costs, toxicity, complexity, and age. NPs are created by a variety of biological processes, including fungi (microbes), bacteria, and plants. The biological process is inexpensive, produces a bulk amount, and is eco-friendly.
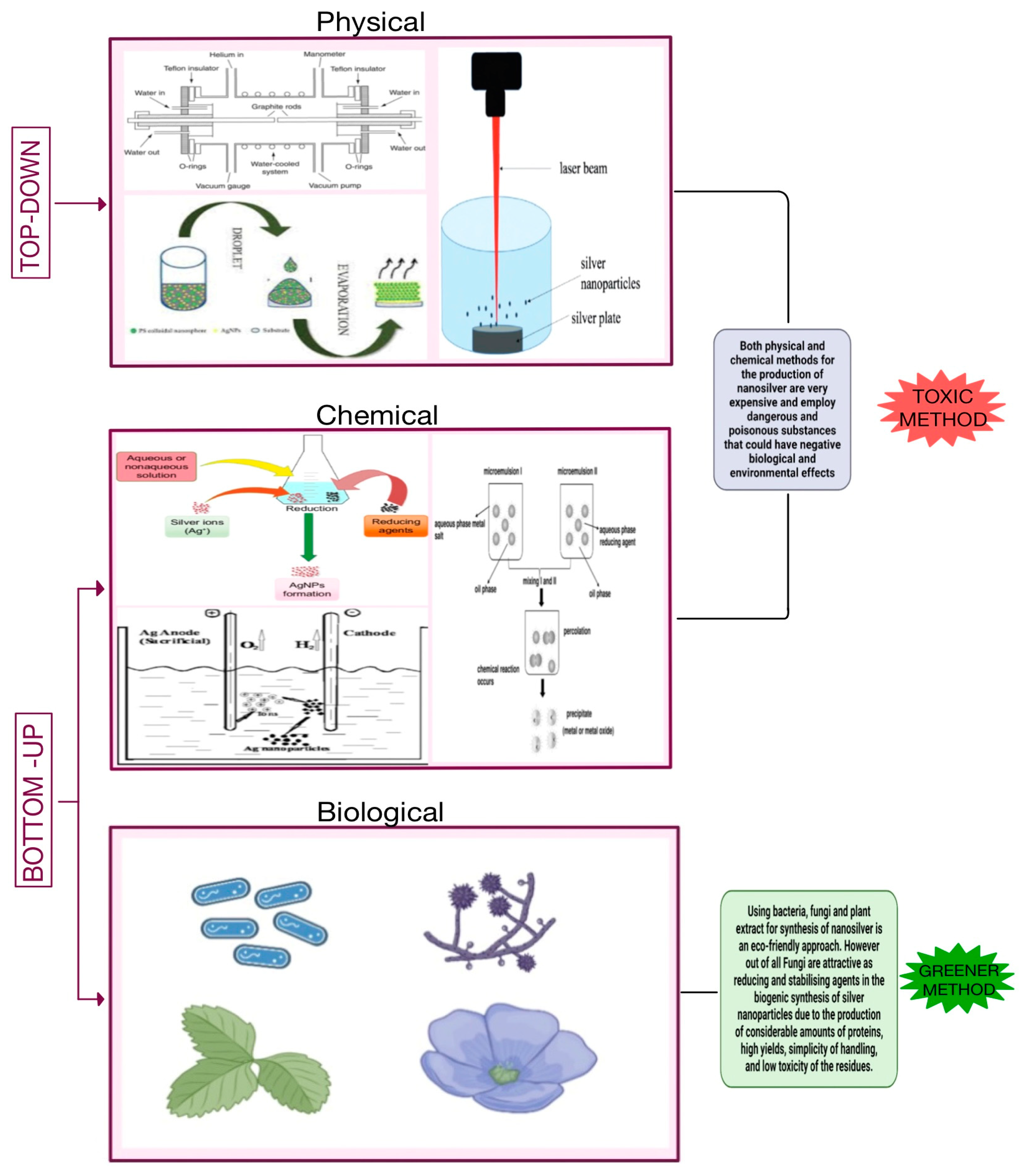
Figure 2.
Mechanism for the synthesis of fungi mediated silver nanoparticles [24].
Figure 2.
Mechanism for the synthesis of fungi mediated silver nanoparticles [24].

Figure 3.
Nanoparticles are very small-sized particles that can easily enter exposed spiracles on the insect body and travel through the trachea and tracheoles into various tissues and cells of the insect, causing cell toxicity and eventually leading to apoptosis and tissue damage. Direct penetration, endocytosis, and cation channels are all ways for nanoparticles to enter the cell membrane. When nanoparticles travel through the endocytic pathway, they are engulfed by lysosomes, causing membrane disruption and exposing lysosomal enzymes containing NPs to other cell organelles such as mitochondria, nuclei, and even neighboring cells. NPs in the nucleus cause irreversible DNA damage. In Mitochondria NPs cause the generation of ROS. Release of excessive ROS induces the Apoptosis Pathway by increasing expression of cytochrome c, increasing MDA levels, disruption of bax/bcl-2 expression, and cleavage of caspase 3. As a result, cell and tissue death occurs, impairing all physiological and biological processes.
Figure 3.
Nanoparticles are very small-sized particles that can easily enter exposed spiracles on the insect body and travel through the trachea and tracheoles into various tissues and cells of the insect, causing cell toxicity and eventually leading to apoptosis and tissue damage. Direct penetration, endocytosis, and cation channels are all ways for nanoparticles to enter the cell membrane. When nanoparticles travel through the endocytic pathway, they are engulfed by lysosomes, causing membrane disruption and exposing lysosomal enzymes containing NPs to other cell organelles such as mitochondria, nuclei, and even neighboring cells. NPs in the nucleus cause irreversible DNA damage. In Mitochondria NPs cause the generation of ROS. Release of excessive ROS induces the Apoptosis Pathway by increasing expression of cytochrome c, increasing MDA levels, disruption of bax/bcl-2 expression, and cleavage of caspase 3. As a result, cell and tissue death occurs, impairing all physiological and biological processes.
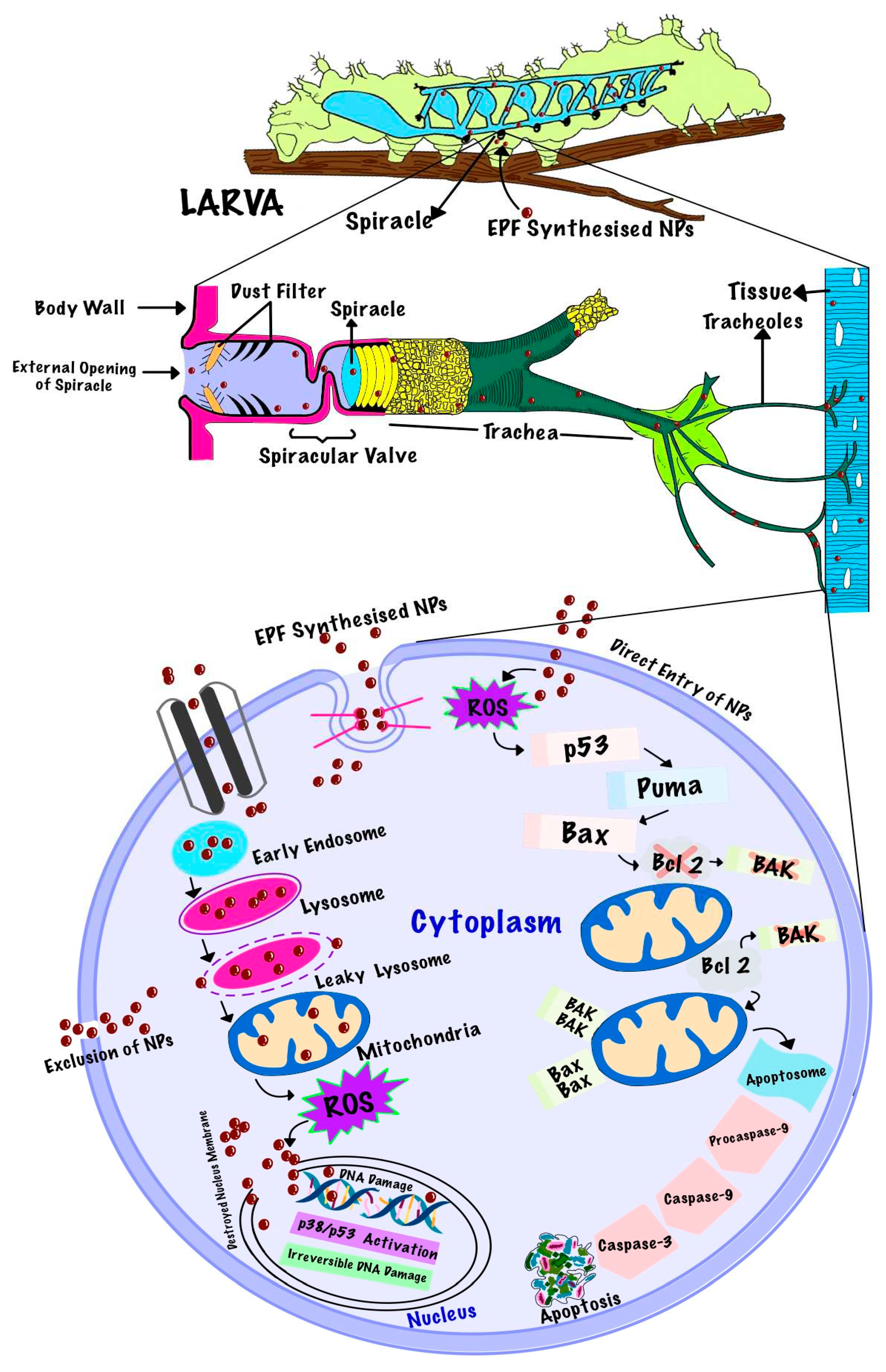

Table 1.
Different entomopathogenic fungi used for the Biosynthesis of silver nanoparticles.
| Entomopathogenic fungi used | Particle size | Time is taken for synthesis | Reference |
|---|---|---|---|
| Aspergillus tubingensis | 35nm | 96hrs | [101] |
| Penicillium citrinum | 109nm | 24hrs | [102] |
| Aspergillus foetidus | 20-40 nm | 24hrs | [103] |
| Isariafumosorosea | 51.31 -111.02 nm | 72hrs | [104] |
| Metarhizium anisopliae | 28–38 nm | 72hrs | [105] |
| Beauveria bassaina | 20–30 nm | 5 days | [106] |
| Trichoderma longibrachiatum | 24.43 nm | 72hrs | [107] |
| Penicillium oxalicum | 150nm | 96hs | [108] |
Table 2.
Mortality value of entomopathogenic based nanoparticles against their targeted pest.
| Entomopathogenic based Nanoparticles | Targeted Pest | Mortality value | References |
|---|---|---|---|
| Penecillium verucosum based silver nanoparticles | Culex quinquefasciatus | LC50-4.91, 5.16, 5.95 7.83; LC90-8.13, 8.44,7.76,12.63 | [120] |
| Cochliobolus lunatus based silver nanoparticles | Aedes aegypti | LC50- 1.29, 1.48, 1.58 ppm ; LC90 3.08, 3.33, 3.41 ppm. | [121] |
| Cochliobo luslunatus based silver nanoparticles | Anopheles stephensiListon | LC50- 1.17, 1.30, 1.41 ppm; LC90 2.99, 3.13, 3.29 ppm | [122] |
| Beauveria bassiana TS11 based zinc oxide nanoparticles | Trialeurodesvaporariorum | LC50-7.35 ppm | [123] |
| Isaria fumosorosea based iron nanoparticles | Bemisiatabaci | LC50- 19.17, 26.10,37.71 ppm | [124] |
|
Beauveria brongniartii based iron nanoparticles |
Spodoptera litura | LC50-59 ppm | [125] |
| Trichoderma based selenium nanoparticles | Spodoptera litura | LC50-39.739; LC90-142.839 ppm | [52] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



