
Submitted:
04 May 2023
Posted:
05 May 2023
You are already at the latest version
Abstract
The dependence of longleaf pine (Pinus palustris) ecosystems on fire is well-understood, and anthropogenic alteration of fire cycles within its natural range has contributed to its decline. This is increasingly exacerbated in areas of urban interfaces, wherein use of prescribed fire can be problematic. The purpose of this study—the University of West Florida Campus Ecosystem Study—was to examine effects of fire exclusion on longleaf pine in the unique urban interface of a university campus. This was conceived as an interconnected series of investigations on the main campus and three associated natural areas that comprise remnant longleaf stands following cessation of widespread longleaf pine harvesting ~120 years ago. This period of chronic fire exclusion has allowed for a distinct shift in stand structure and composition. The open, savanna-like structure of fire-maintained longleaf stands has transitioned into closed-canopy forests with the increased prevalence of southern evergreen oaks (especially live oak—Quercus virginiana) and Magnolia spp., resulting in complete absence of longleaf regeneration. Fire exclusion has also appeared to have decreased soil fertility. Significant variation in mean age of longleaf pine stems of the main campus and natural areas suggest that the natural areas were likely under separate ownership with contrasting land-use history prior to purchase by the State of Florida for campus construction in 1963.

Keywords:
longleaf pine
; urban interface
; fire exclusion
1. Introduction
Longleaf pine (Pinus palustris) is one of the better-known disturbance-maintained ecosystems, and its dependence on fire is both well studied and understood. Because its natural range—widespread as it is throughout the southeastern United States—coincides with the region of North America that experiences highest frequencies of landfall tropical cyclones, however, another disturbance regime of great significance is that of tropical storms and hurricanes [1,2,3,4]. A phenomenological model has been developed that integrates both fire and tropical cyclones in influencing the physiognomy of longleaf pine stands [5]. This model, developed as an alternative strategy toward creating endpoints for longleaf restoration, views these two disturbance regimes as having contrasting impacts on vegetation strata based on innate differences in disturbance frequency and intensity. Tropical cyclones are more sporadic and intense than fires, which are more frequent with less intensity. Tropical cyclones exert a direct impact on the overstory through wind damage and mortality, with indirect effects on the herb layer, primarily via changes in spatial patterns of accumulation of detritus and shading. By contrast, fire affects the herb layer directly, including the juvenile stages of longleaf regeneration.
Forests in general have been referred to as comprising a paradox of biodiversity, with the most apparent vegetation stratum—the overstory—typically composed of the fewest number of species, with the diminutive herbaceous layer harboring 80-90% of forest plant species. An extreme example of this would be longleaf pine ecosystems. Under conditions of frequent fire, the savanna-like physiognomy of longleaf stands is simple—a single species in the overstory and quite numerous species in the herb layer [6]. Synthesizing data from forest ecosystems throughout the United States, Gilliam [7] reported that there was a significant linear relationship between the species richness of the overstory and that of the herb layer, further noting that conifer forests more commonly occupy the low range of both strata in terms of species richness. A notable outlier in this summary were data from longleaf pine savannas. Indeed, working in old-growth longleaf pine in southeastern Louisiana, Platt et al. [6] found, as expected, only longleaf pine in the overstory, but 251 species in the herb layer. More specifically, they found more herb layer species—at least 20% more—in both recent and persistent canopy openings. They also found notable variation among plant functional groups in response to these openings. For example, plants responding most sensitively to canopy openings were either short-lived, monocarpic perennial forbs or grasses arising from soil seed banks. There was a significant linear increase in total numbers of all species with measured levels of light. Clearly, fire is important in maintaining high light levels that support observed species richness of longleaf savannas.
Among numerous threats to the structure and function of longleaf pine ecosystem is the chronic, widespread alteration of fire cycles that once were both part of the natural landscape and a common land-use practice among indigenous human populations. All of these factors intersect intimately with current human populations, with associated land-use practices and resource demands. The relative percentage of the population living in the southeastern United States, along with the absolute increases in the population of the region, contribute greatly to the decline of the longleaf pine ecosystem. Currently, the nearly 100 million people living in the southeast represent nearly 1/3 the U.S. population. This pattern has contributed to declines in available longleaf habitat via conversion to urban areas, agricultural lands, and pine plantation forestry [8].
Also related to human population growth in the southeastern United States is the increase in the degree of urban interface with longleaf pine sites [9]. Urban interfaces represent a unique juxtaposition of human populations and the natural systems that sustain them [10]. More specifically, college and university campuses comprise a distinctive urban interface, with their spatial patterns of permanent construction—dormitories, administrative buildings, classrooms, academic departments—combined with parking lots and green spaces, such as quadrangles, gardens, and arboreta [11,12]. Ultimately, all urban interfaces, especially those of campuses, typically necessitate a high degree of fire exclusion.
Several features of the campus of the University of West Florida in Pensacola have combined to provide an opportunity to study the ecology of longleaf pine—including the effects of fire exclusion—in a unique setting. Campus construction began in 1963 from 405 ha of longleaf pine stands recovering from extensive logging in the region [13]. The designer of the campus, John E. Jarvis, Jr., used a ‘design with nature’ approach that called for minimal tree removal during construction of buildings and establishment of paved areas [14]. Remnant campus trees were predominantly species native to the Florida Panhandle, including species of pine, oak, and magnolia [15,16]. In addition to his main campus directive, Jarvis called for the establishment of natural areas with an expansive network of low-impact, non-paved trails. The vision of Jarvis has been borne out to be particularly fruitful, as there are numerous habitat types observable on the UWF campus, including a wetland and associated hammock, a cypress pond, and a freshwater stream with riparian plant species. Anecdotally, the campus of UWF has the highest number of vascular plant species (nearly 900) of any university campus in the eastern United States (specimens of each species held in the UWF Michael I. Cousens Herbarium).
In the summer of 2019, these distinctive features of the UWF campus were recognized for their innate value in investigating the ecology of longleaf pine in the urban interface of a university campus [17,18]. Thus, the UWF Campus Ecosystem Study (CES) was established as an interconnected series of research projects. To date, there have been five investigations, with four as part of the original focus on the ecology of longleaf pine [19,20,21,22] and one study [23] monitoring the freshwater stream that flows through the campus and joining the Escambia River (Figure 1).
Most forest stands along the UWF campus nature trails are typical of second-growth longleaf pine forests experiencing chronic fire exclusion [6,20,24,25,26,27,28]. These comprise stands of widely-space longleaf stems primarily 50-100 yr in age [21] with the otherwise fire-maintained open matrix [2,29,30] filled with hardwood species, especially numerous southern oaks, e.g., live oak (Quercus virginiana), water oak (Q. nigra), blackjack oak (Q. marilandica), and laurel oak (Q. laurifolia).
The purpose of this paper is to provide a synthesis review of all work carried out as part of the UWF CES. Also, further novel analysis of soil fertility and forest stand composition and structure will be added to further understand current conditions in the context of past land-use history, with a focus on the effects of chronic fire exclusion in longleaf pine systems.
2. Materials and Methods
2.1. Study Site
The UWF CES is an ongoing study that comprises the main campus and forested stands off of low impact, unpaved sandy trails in several natural areas in Pensacola, Florida (30° 33’ N, 87° 11’ W, Figure 1). The natural areas include the Edward Ball Nature Trails (hereafter, ‘Ball Trails’), the Baars-Firestone Wildlife Sanctuary (hereafter, ‘Sanctuary’), and the Campus Side Trails (hereafter, ‘Side Trails’). Soils of these sites are dominated by the Troup series, which consist of deep, well-drained soils that arose from marine sediments of a sandy, loamy nature [31]. Troup soils are loamy, kaolinitic, thermic Grossarenic Kandiudults with a chronically-high seasonal water table below 2 m in depth [32]. Previous work has shown these soils to be acidic and infertile [22].
2.2. Field Sampling
The initial investigation of the UWF CES began in summer 2019. Although that study examined two complementary aspects of the longleaf pine ecosystem associated with the campus, i.e., (1) the effects of gopher tortoise burrowing on herbaceous plant communities and soil fertility, (2) size structure of longleaf pines on the main campus, only the data from the longleaf pine sampling will be covered here. These data were obtained by measuring all longleaf stems ≥2.5 cm diameter at breast height (DBH) on the main campus for DBH.
In summer 2020, longleaf stands of the Ball Trails and Sanctuary sites were studied to determine the response of longleaf pine stands to chronic fire exclusion. For this part of the study, 15 400-m2 circular plots were established in each site to characterize composition and structure and sample mineral soil. Living stems that were ≥ 2.5 cm DBH were identified in each sample plot to species and measured for DBH. These data were transcribed to yield basal area and density for each species per sample site for the construction of importance value tables (data not shown), as well as serving is input for canonical correspondence analysis (CCA—see 2.3 Data analysis). Following removal of organic material, mineral soil was sampled to a depth of 5 cm and analyzed for pH, organic matter (OM), cation exchange capacity (CEC), extractable macro- and micronutrients, and Al.
On 16 September 2020, Hurricane Sally—a Category 2 tropical cyclone—impacted the region around and including the UWF campus with a 12-hr eyewall and winds reaching 49 m/sec. This resulted in numerous trees, but especially longleaf pines, being windfallen, including the main campus and the stands along the nature trails. In summer 2021, the UWF CES was able to take advantage of longleaf pine windthrows by Hurricane Sally to obtain cross-sections for age determinations, sampled primarily in the Ball Trails and Sanctuary areas. A total of 50 sections were taken for this study, with 40 being windthrows and 10 being live-tree harvests to provide for an adequate low-end range of diameters, as small, young longleaf pines tend to be unsusceptible to windthrow. For each stem section sampled, DBH and number and width of annual rings (to the nearest 0.01 mm) were recorded.
Sampling was expanded in summer 2022 to include the Side Trails area composed of hardwood stands to compare with adjacent longleaf pine stands to assess variation in stand composition/structure, soil fertility, and soil microbiome. For this study, 12 circular sample plots (400 m2 ha) were established per stand type. Stand and soil sampling were carried out as in previous UWF CES investigations. Longleaf pine and soil data from only the longleaf pine stands are included in this analysis.
2.3. Data analysis
Size structure of longleaf pine was determined by constructing size-class frequency distributions based on DBH. To assess possible effects of fire exclusion on soil fertility, t-tests were used to compare mean soil data between pre- versus post-fire exclusion sites [33]. Linear regression was used to compare age of longleaf stems to measured DBH for 50 stems to yield a model allowing estimation of stem age based on DBH. Age of each measured longleaf pine stem was estimated from DBH with the following model based on this analysis [22]:
where Age is stem age in years; DBH is in cm. Mean age of longleaf pine was compared among main campus, Ball Trails, Sanctuary, and Side Trails sites via analysis of variance and Least Significant Difference tests [33]. Age-class frequency distributions were also constructed for longleaf pine populations of these sites.
Age = 1.76*DBH + 0.83
Stand composition was compared among natural area sample sites using CCA with Canoco 5.11, Windows release [34]. As previously stated, input data for tree species were basal areas for each species in each sample plot. Soil data were used as environmental variables. Among numerous ordination techniques, CCA has the advantage of providing a direct gradient analysis of environmental (i.e., soil) variables and species composition [35,36].
3. Results and Discussion
The inaugural effort of the UWF CES in Summer 2019 determined DBH of all main campus longleaf pine stems, a tally which totaled 2,165 stems. When this number is adjusted for estimated sample area, stem density was ~64 stems/ha, a value clearly lower than that of natural stands (e.g., 120–140 stems/ha) [6,24]. On the other hand, it is well within range of tree densities in other urban settings, i.e., 40 to 100 stems/ha [11]. Given the chronically unburned state of urban interfaces [8], it is also unsurprising that the distribution of stem size resembled fire-excluded longleaf pine stands and was in sharp contrast to frequently burned stands [18].
The two natural areas—Ball Trails and Sanctuary—sampled in Summer 2020 displayed interesting patterns of both similarities and contrasts, ultimately suggesting that the two areas had been tracts/parcels of land under separate ownership prior to the initiation of construction of the UWF campus in 1963. Regarding similarities, both sites had live oak as the species of highest importance value as a result of high density, with longleaf pine being of secondary importance, even with its much higher basal area, because of low density. These sites were of essentially identical stand structure, with total basal area of 30.3 and 29.6 m2/ha for Ball Trails and Sanctuary, respectively, and total density of 1225 and 1197 stems/ha, respectively [20].
There were also, however, notable contrasts between sites regarding size structure of longleaf pine. In the Sanctuary site, more than 30% of longleaf stems were found in classes > 30–40 cm, but only ~10% were > 30–40 cm for the Ball Trails. Furthermore, the Sanctuary site was more similar to the main campus than the Ball Trails site. Median DBH was 35.0 and 37.1 cm for the main campus and Sanctuary, respectively, and 31.2 cm for the Ball Trails (Figure 2). Mean DBH of longleaf pine stems was not significantly different between the main campus and Sanctuary (34.4 ± 0.2 and 35.9 ± 1.1 cm, respectively), but means for both were significantly higher (P < 0.05) than the Ball Trails (30.9 ± 0.8 cm).
Two conclusions arise from these observed patterns. The overall similarity confirms that the longleaf pines of the UWF campus represent remaining stems from a time when extensive logging in northwest Florida ceased >100 years ago [13]. Sharp contrasts between the Sanctuary and Ball Trail sites, however, suggest that the two sites likely had different ownership and contrasting land-use history prior to the construction of the UWF campus in 1963. Indeed, these are ~2 km apart and occur on opposite sides of the main campus.
As already discussed, the work of summer 2021 was to sample windthrown longleaf stems resulting from Hurricane Sally in September 2022. This resulted in the linear model presented in the Methods (see Data analysis) which allowed for estimation of age of all measured longleaf, including the main campus and all natural areas, based on DBH.
The scope of the UWF CES was expanded in summer 2022 to include two new facets: (1) comparative analysis of an additional longleaf-dominated stand with an adjacent hardwood-dominated stand, and (2) determination of the soil microbiome of these stands. Sampling took place in the Side Trails site (see Methods). The component of that work that is relevant for this review comprises the DBH data of longleaf pine stems in the pine-dominated Side Trails stand, with a specific focus on stem age and age-class distribution of all sampled areas, including the main campus, Sanctuary, Ball Trails, and Side Trails. Data from the Side Trails place that site more in line with the Ball Trails site, as mean age of the pine-dominated stand was also significantly lower than the main campus and Sanctuary (Figure 3).
Age class distributions reveal further contrasts in age structure among campus longleaf populations, confirming earlier conclusions that these areas likely were of different ownership under contrasting land use at the time of purchase by the State of Florida for construction of the UWF campus [19,20]. Most longleaf stems were between 60 and 70 yr for the main campus and Trails, but between 70 and 80 yr for the Sanctuary (Figure 4). Of even sharper contrast was the longleaf-dominated stand of the Side Trails site. That is, not only were most stems of this stand 40-50 yr, but stems <50 yr were virtually absent (Figure 4). Median stem age was 64, 69, 58, and 55.3 yr for main campus, Sanctuary, Ball Trails, and Side Trails, respectively.
Despite the sharp contrasts in stand age and longleaf age structure among the three natural areas, they were generally quite similar with respect to species composition, as evidenced by the results of canonical correspondence analysis (CCA) (Figure 5). The distribution of sample plots in CCA ordination space reveals no patterns consistent with site. These data support previous findings of close similarities between Ball Trails and Sanctuary stands [20], wherein live oak and longleaf pine were co-dominant species at both sites (to emphasize this, these two species are highlighted in bold, green font). Appropriately, they are found at the center of ordination space, owing to their ubiquitous nature, as they were generally found in all plots.
It merits restating that this high diversity of hardwood species is the direct result of chronic fire exclusion. Although these are second-growth stands, this phenomenon has also been described for old-growth longleaf stands [6,8,25]. Of particular importance is the success of live oak with its importance in all stands, regardless of stand age of longleaf pine. More specifically, the age structure for longleaf pine at UWF (e.g., Figure 4) closely approximates the region’s land-use history, wherein the wide-spread logging extended from 1870 to 1930 [13].
To assess the potential changes in soil fertility in response to chronic fire exclusion in the UWF sample sites, Gilliam et al. [20] compared soil data from the Ball Trails and Sanctuary sites to the open matrix used for the gopher tortoise study on UWF campus [19]. This is admittedly based on an assumption that cannot be tested empirically, i.e., that soils from these open areas represent pre-exclusion conditions. Despite this, two observations support the veracity the assumption. First, the open matrix displays the patterns of longleaf regeneration typical of the period prior to fire exclusion. Second, soils of all sites (open matrix and three longleaf pine sites) are of the same series—the Troup series (see Methods).
Direct studies of the effects of chronic fire exclusion on soils is underrepresented in the literature. Comparisons presented here suggest that fire exclusion results in an overall decline in soil fertility, much of which is related to increases in acidity of already acidic soils, as the pH of post-exclusion soil was ~0.7 unit lower that pre-exclusion soil (Table 1). This likely resulted from two acidifying processes occurring interactively and simultaneously. First, litter from longleaf pine and live oak is acidic, and as acidic litter builds up on the forest floor, organic acids are leached and accumulate in the A horizon. Furthermore, Moore et al. [37] demonstrated that throughfall from canopies of live oak is acidified, with precipitation pH decreasing by as much as 0.5 as it passes through live oak canopies. Also, trees have a far higher nutrient demand than do herbaceous plants [38], and uptake of cations by plant roots acidifies the rhizosphere from the exchange of H+ for nutrient cations from exchange sites [39]. Because chronic fire exclusion has allowed extensive hardwood establishment, it is likely that nutrient cations, such as Ca, Mg, and K, have been taken up in great amounts during fire exclusion. Also, hardwoods, which represent up to 60% of the basal area of the longleaf sites, have a higher demand for base cations, particularly Ca and Mg, than do conifers [40]. Patterns of significantly lower Ca, Mg, and K in post- versus pre-exclusion soil (Table 1) support this second mechanism.
Adding to fire exclusion-mediated declines in soil fertility is the contrast in soil organic matter (OM). Soils of all sampled areas are composed of deep, coarse-textured sands that had low OM (~2%) and CEC (~1.5 meq/100 g), typical of this region, with OM being lower than soils observed in old-growth longleaf pine (~4%) [25]. Organic matter in post-exclusion soil was significantly lower than that in pre-exclusion soil (Table 1). Although this may seem counterintuitive (i.e., it may have been expected that fire exclusion may have been expected to increase in soil OM from increased litterfall from trees), OM is probably higher in pre-exclusion soils because in these coarse-textured soils derive OM from fine-root turnover from dominant grass species, such as broomsedge (Andropogon virginicus), wiregrass (Aristida stricta), and bluegrass (Poa spp.) [41]. These grasses are predominant in open areas, but they exhibit rapid decline because of insufficient light during canopy closure.
4. Conclusions
The University of West Florida Campus Ecosystem Study, initiated in Summer 2019, was conceived as interconnected investigations on the main campus and associated natural areas that comprise remnant longleaf stands following cessation of widespread logging ~120 years ago. This has allowed assessment of effects of chronic fire exclusion on longleaf pine in the unique urban interface of a university campus. Data presented here offer a compelling picture that fire exclusion has resulted in profound changes in stand structure and composition. The savanna physiognomy of fire-maintained longleaf stands has become closed-canopy forests with numerous hardwood species. Live oak has been particularly ubiquitous in these stands which inhibits longleaf regeneration. Indeed, it is notable that it is found in all stands, regardless of the presence of longleaf pine [22]. Data also suggest that soil fertility declines significantly with chronic fire exclusion. Significant variation in the mean age of longleaf pine stems of the main campus and the three natural areas suggest that these natural areas were likely under separate private ownership prior to the construction of the UWF campus in 1963.
A further implication for the research reviewed here is the essential value of campuses of higher education as sites for ecological study. As already established, college and university campuses comprise a unique type of urban interface with their various buildings, paved areas, and lawns [11,12,18]. Indeed, Gilliam et al. [20] identified several institutions within the natural range of longleaf pine. In Georgia, these were Berry College and Valdosta State University. In Florida, these were Stetson University, Florida A&M University, Florida State University, University of Central Florida, and Rollins College. There are numerous others beyond these, all with wide landcover of longleaf pine. In many, restoration and research projects are either planned or underway [17,42,43].
More specifically regarding the investigations of the UWF CES, such efforts have involved the work of undergraduate students, providing them with the invaluable and unique opportunity to extend their education and learning through research. To date, results of the research carried out by these students have provided deeper insights into longleaf pine ecology, especially regarding the effects of chronic fire exclusion on the structure and function of longleaf pine ecosystems, including species composition and soil resources. Consistent with the aims of the UWF CES, future investigations will build further on the results presented herein, including measurements of photosynthetically-active radiation and characterization of the soil seed bank among hardwood-dominated versus longleaf pine-dominated stands.
Author Contributions
F.S.G. wrote the manuscript, and has read/agreed to the published version.
Funding
This research received no external funding.
Data Availability Statement
Not applicable.
Acknowledgments
I gratefully acknowledge the financial assistance of the UWF Summer Undergraduate Research Program, Dr. Allison Schwartz, Director. I further acknowledge the invaluable contributions of the following individuals who were undergraduates at the University of West Florida when they carried their respective research as part of the UWF Campus Ecosystems Study: Emily Harmon, Skylar Boyles, Selina Detzel, Katie Bray, Emily Major, Heather Patten, Sarah Rabinowitz, Elizabeth Hargis, Leo Sweet, Brandon Davis. The writing of this manuscript was supported with support from the Hal Marcus College of Science and Engineering, Dr. Jaromy Kuhl, Dean.
Conflicts of Interest
The author declares no conflict of interest.
References
- Platt, W.J. Southeastern pine savannas. In The Savanna, Barren, and Rock Outcrop Communities of North America. Anderson, R.C.; Fralish, J.S.; Baskin, J., Eds.; Cambridge University Press, Cambridge, England, 1999; pp. 23–51.
- Zampieri, N.E. , Pau, S., and Okamoto, D.K. The impact of Hurricane Michael on longleaf pine habitats in Florida. Sci. Rep.-UK, 2020, 10, 8483. [Google Scholar] [CrossRef] [PubMed]
- Gilliam, F.S. Impacts of tropical cyclones on longleaf pine ecosystems of Florida: tropical cyclogenesis, landfall frequencies, and climate change. Front. Ecol. Evol., 2021, 9, 595791. [Google Scholar] [CrossRef]
- Zampieri, N.E.; Pau, S. The effects of fire, climate, and species composition on longleaf pine stand structure and growth rates across diverse natural communities in Florida. Forest Ecology and Management, 2022, 526, 120568. [Google Scholar] [CrossRef]
- Gilliam, F.S.; Platt, W.J. , Peet, R.K. Natural disturbances and the physiognomy of pine savannas: a phenomenological model. Appl. Veg. Sci., 2006, 9, 83–96. [Google Scholar] [CrossRef]
- Platt, W.J.; Carr, S.M.; Reilly, M.; Fahr, J. Pine savanna overstorey influences on ground-cover biodiversity. Appl. Veg. Sci., 2006, 9, 37–50. [Google Scholar] [CrossRef]
- Gilliam, F.S. Introduction: the herbaceous layer—the forest between the trees. In The Herbaceous Layer in Forests of North America. Gilliam, F.S., Ed.; Oxford University Press, Oxford, England, 2014.; pp. 1-9.
- Gilliam, F.S.; Platt, W.J. Conservation and restoration of the longleaf pine ecosystem. Appl. Veg. Sci., 2006, 9, 7–10. [Google Scholar] [CrossRef]
- Francos, M.; Stefanuto, E.B.; Úbeda, X.; Pereira, P. Long-term impact of prescribed fire on soil chemical properties in a wildland-urban interface. Northeastern Iberian Peninsula. Sci. Tot. Environ., 2019, 689, 305–311. [Google Scholar] [CrossRef]
- Barrington-Leigh, C.; Millard-Ball, A. A century of sprawl in the United States. PNAS, 2015, 112, 8244–8249. [Google Scholar] [CrossRef]
- Turner, P.V. Campus: an American planning tradition. MIT Press, Cambridge, Massachusetts, 1984.
- Roman, L.A.; Fristensky, J.P.; Eisenman, T.S.; Greenfield, E.J.; Lundgren, R.E.; Cerwinka, C.E.; Hewitt, D.A.; Welsh, C.C. Growing canopy on a College campus: understanding urban forest change through archival records and aerial photography. Environ. Manage., 2017, 60, 1042–1061. [Google Scholar] [CrossRef]
- Knight, G.R.; Oetting, J.B.; Cross, L. Atlas of Florida’s natural heritage—biodiversity, landscapes, stewardship, and opportunities. Institute of Science and Public Affairs, Florida State University, Tallahassee, FL, USA, 2011.
- Jarvis, J.E. The Next Larger Picture: an Anecdotal History of the Master Planning of the Campus of the University of West Florida and the Design and Construction of its Buildings for the Initial Thirty-Something Years; a Memoir, of Sorts. Pioneer Series/University of West Florida Foundation, Inc, Pensacola, FL, USA, 2008.
- Daubenmire, R. The Magnolia grandiflora-Quercus virginiana forest of Florida. Am. Midl. Nat., 1990, 123, 331–347. [Google Scholar] [CrossRef]
- Marse, C. Rising Fiery Constellation: a Portrait of the University of West Florida from Birth to Forty. Pioneer Series, Pensacola, FL, USA, 2007.
- Cole, K.; Bennington, C. From the ground up: natural history education in an urban campus restoration. Southeast. Nat., 2017, 16, 132–145. [Google Scholar] [CrossRef]
- Copenheaver, C.A.; Seiler, J.R.; Peterson, J.A.; Evans, A.M.; McVay, J.L.; White, J.H. Stadium woods: a dendroecological analysis of an old-growth forest fragment on a university campus. Dendrochronologia, 2014, 32, 62–70. [Google Scholar] [CrossRef]
- Gilliam, F.S.; Harmon, E.E.; Boyles, S.C. The University of West Florida campus ecosystem study: gopher tortoise and longleaf pine populations in an urban interface. Urban Ecosyst., 2020, 23, 355–362. [Google Scholar] [CrossRef]
- Gilliam, F.S.; Detzel, S.J.; Bray, K.D.; Major, E.A. The University of West Florida campus ecosystem study: the college/university campus as a unit for study of the ecology of longleaf pine. Urban Ecosyst., 2021, 24, 1073–1082. [Google Scholar] [CrossRef]
- Gilliam, F.S.; Patten, H.N.; Rabinowitz, S.K. The University of West Florida campus ecosystem study: age-diameter and growth relationships of longleaf pine using hurricane-induced windthrows. Urban Ecosyst., 2022, 25, 839–848. [Google Scholar] [CrossRef]
- Gilliam, F.S.; Hargis, E.A.; Rabinowitz, S.K.; Davis, B.C.; Sweet, L.L.; Moss, J.A. Soil microbiomes of hardwood- versus pine-dominated stands: linkage with overstory species. Ecosphere, 2023; in press. [Google Scholar]
- Gilliam, F.S.; Hardin, J.W.; Williams, J.A; Lackaye, R.L. The University of West Florida Campus Ecosystem Study: spatial and temporal variation in water quality of Thompson Bayou. Water, 2022, 14, 2916. [Google Scholar] [CrossRef]
- Addington, R.N.; Knapp, B.O.; Sorrell, G.G.; Elmore, M.L.; Wang, G.G.; Walker, J.L. Factors affecting broadleaf woody vegetation in upland pine forests managed for longleaf pine restoration. For. Ecol. Manag., 2015, 354, 130–138. [Google Scholar] [CrossRef]
- Gilliam, F.S.; Platt, W.J. Effects of long-term fire exclusion on tree species composition and stand structure in an old-growth longleaf pine forest. Plant Ecol., 1999, 140, 15–26. [Google Scholar] [CrossRef]
- Hiers, J.K.; Walters, J.R.; Mitchel, R.J.; Varner, J.M.; Conner, L.M.; Blanc, L.A.; Stowe, J. Ecological value of retaining pyrophytic oaks in longleaf pine ecosystems. J. Wildlife. Manage., 2014, 78, 383–393. [Google Scholar] [CrossRef]
- Provencher, L.; Litt, A.R.; Gordon, D.R. Predictors of species richness in Northwest Florida longleaf pine sandhills. Conserv. Biol., 2003, 17, 1660–1671. [Google Scholar] [CrossRef]
- Varner, J.M.; Gordon, D.R.; Putz, F.E.; Hiers, J.K. Restoring fire to long-unburned Pinus palustris ecosystems: novel fire effects and consequences for long-unburned ecosystems. Restor. Ecol., 2005, 13, 536–544. [Google Scholar] [CrossRef]
- Platt, W.J.; Evans, G.W.; Rathbun, S.L. The population dynamics of a long-lived conifer (Pinus palustris). Am. Nat., 1988, 131, 491–525. [Google Scholar] [CrossRef]
- Noel, J.M.; Platt, W.J.; Moser, E.B. Characteristics of old and second growth stands of longleaf pine (Pinus palustris) in the Gulf coastal region of the U.S.A. Cons. Biol., 1998, 12, 533–548. [Google Scholar]
- Hine, A.C. Geologic History of Florida: Major Events That Formed the Sunshine State. University Press of Florida, Gainesville, FL, USA, 2013.
- USDA. 2004. Soil survey of Escambia County, Florida. United States Department of Agriculture, Natural Resources Conservation Service, Washington, DC, USA.
- Zar, J.H. Biostatistical Analysis, 5th ed.; Prentice-Hall: Upper Saddle River, NJ, USA, 2009. [Google Scholar]
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data Using CANOCO 5, 2nd ed. Cambridge University Press: Cambridge, England, 2014.
- Barbour, M.G.; Burk, J.H; Pitts, W.D.; Gilliam, F.S.; Schwartz, M.W. Terrestrial plant ecology, 3rd Edition. Benjamin/Cummings Publishing Company, Inc.: Menlo Park, CA, USA, 1999.
- Gilliam, F.S.; Saunders, N.E. Making more sense of the order: A review of Canoco for Windows 4.5, PC-ORD version 4 and SYN-TAX 2000. J. Veg. Sci., 2003, 14, 297–304. [Google Scholar] [CrossRef]
- Moore, L.D.; Van Stan II, J.T.; Gay, T.E.; Rosier, C.; Wu, T. Alteration of soil chitinolytic bacterial and ammonia oxidizing archaeal community diversity by rainwater redistribution in an epiphyte-laden Quercus virginiana canopy. Soil Biol. Biochem., 2016, 100, 33–41. [Google Scholar] [CrossRef]
- Muller, R.N. Nutrient relations of the herbaceous layer in deciduous forest ecosystems. In The Herbaceous Layer in Forests of North America. Gilliam, F.S., Ed.; Oxford University Press, Oxford, England, 2014.; pp. 13-34.
- Marschner, H . Mineral Nutrition of Higher Plants, 3rd edn. Academic Press, London, England, 2012.
- Schroth, A.W.; Friedland, A.J.; Bostick, B.C. Macronutrient depletion and redistribution in soils under conifer and northern hardwood forests. Can. J. For. Res., 2007, 71, 457–468. [Google Scholar] [CrossRef]
- West, J.B.; Espeleta, J.F.; Donovan, L.A. Fine root production and turnover across a complex edaphic gradient of a Pinus palustris—Aristida stricta savanna ecosystem. For. Ecol. Manag., 2004, 189, 397–405. [Google Scholar] [CrossRef]
- Cipollini, M.; Felch, P.; Dingley, N.R.; Maddox, C. Changes in tree canopy, understory vegetation, and fuel composition after 10 years of restoration management in an old-growth mountain longleaf pine forest. Nat. Areas J., 2019, 39, 197–211. [Google Scholar] [CrossRef]
- Cipollini, M.L.; Culberson, J.; Stripplehoff, C.; Baldvins, T.; Miller, K. Herbaceous plants and grasses in a mountain longleaf pine forest undergoing restoration: a survey and comparative study. Southeast. Nat., 2012, 11, 637–668. [Google Scholar] [CrossRef]
Figure 1.
The campus of the University of West Florida, Pensacola, Florida (30.5102° N, 87.2125° W). The main campus is composed of all roads, parking lots, and permanent structures; university property extends beyond this to wooded areas. In the summer of 2019, all longleaf pines of the main campus ≥2.5 cm diameter at breast height (DBH) were measured for DBH—a total of 2,165 stems [19]. In the summer of 2020, sample plots were established and sampled in two of the UWF campus natural areas: Ball Trails (T) and Baars-Firestone Wildlife Sanctuary (S) [20]. The most recent study [22] is indicated by the red circle.
Figure 1.
The campus of the University of West Florida, Pensacola, Florida (30.5102° N, 87.2125° W). The main campus is composed of all roads, parking lots, and permanent structures; university property extends beyond this to wooded areas. In the summer of 2019, all longleaf pines of the main campus ≥2.5 cm diameter at breast height (DBH) were measured for DBH—a total of 2,165 stems [19]. In the summer of 2020, sample plots were established and sampled in two of the UWF campus natural areas: Ball Trails (T) and Baars-Firestone Wildlife Sanctuary (S) [20]. The most recent study [22] is indicated by the red circle.
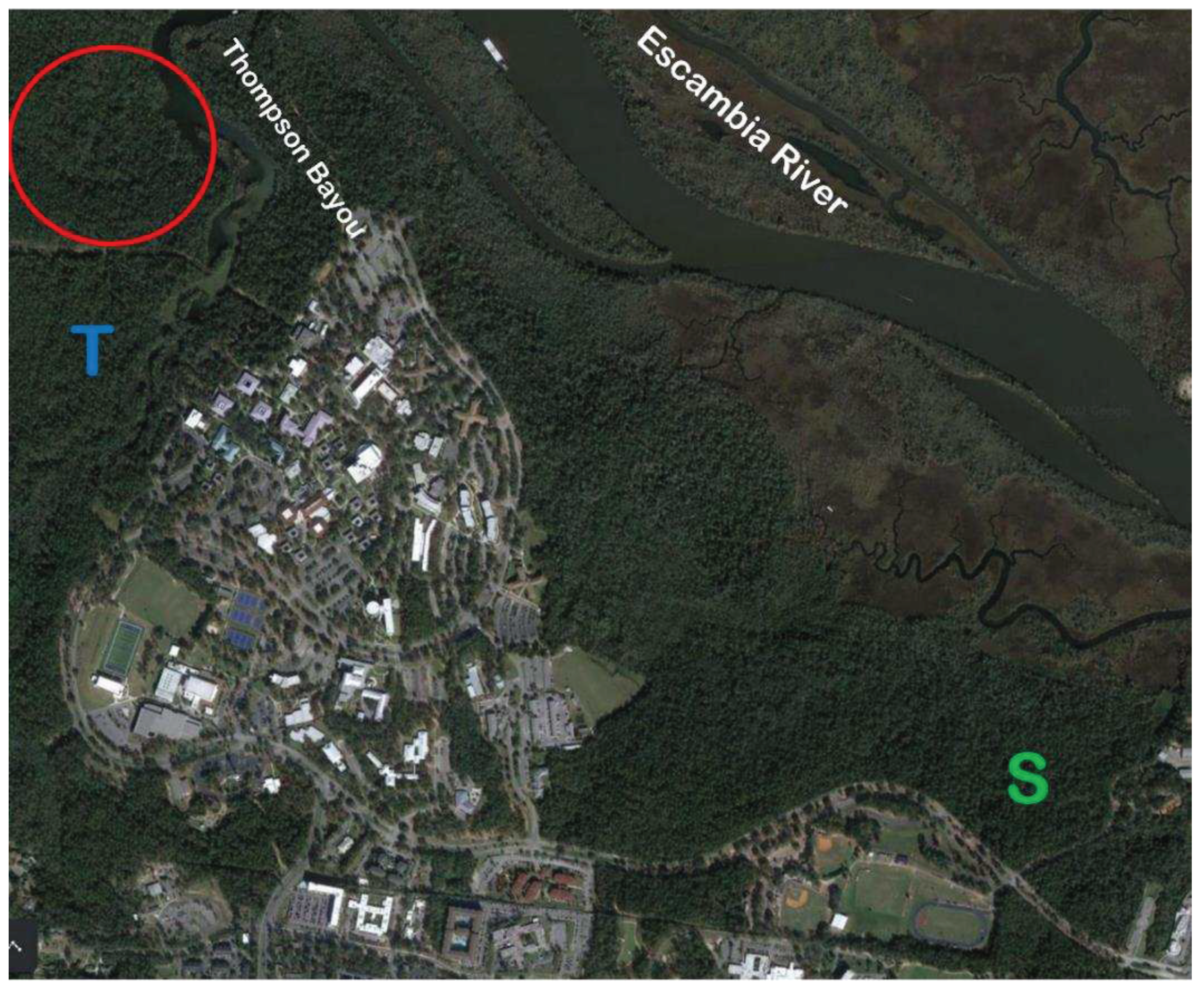
Figure 2.
Diameter size-class frequency distributions for longleaf pine for two UWF natural areas (Ball Trails and Wildlife Sanctuary) and the UWF main campus. Figure used with permission from [20].
Figure 2.
Diameter size-class frequency distributions for longleaf pine for two UWF natural areas (Ball Trails and Wildlife Sanctuary) and the UWF main campus. Figure used with permission from [20].
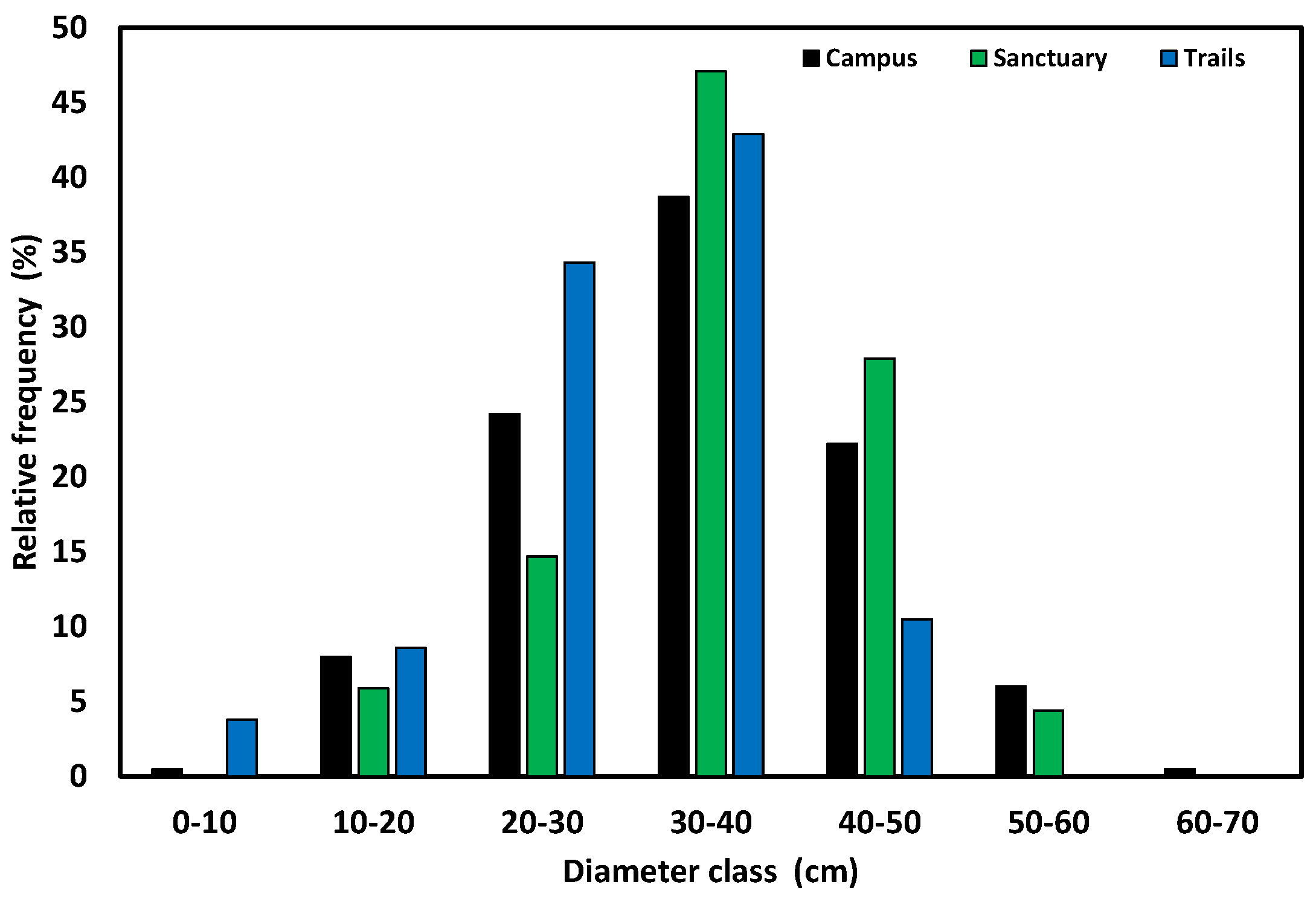
Figure 3.
Mean age of longleaf pine for four areas at UWF: main campus, Baars-Firestone Wildlife Sanctuary, Edward Ball Trails, and pine-dominated stands of the current study (see Figure 1 for locations). Values shown are means ±1SE. Means with the same superscript are not significantly different at P<0.05. (B) Frequency age class distributions for these groups.
Figure 3.
Mean age of longleaf pine for four areas at UWF: main campus, Baars-Firestone Wildlife Sanctuary, Edward Ball Trails, and pine-dominated stands of the current study (see Figure 1 for locations). Values shown are means ±1SE. Means with the same superscript are not significantly different at P<0.05. (B) Frequency age class distributions for these groups.
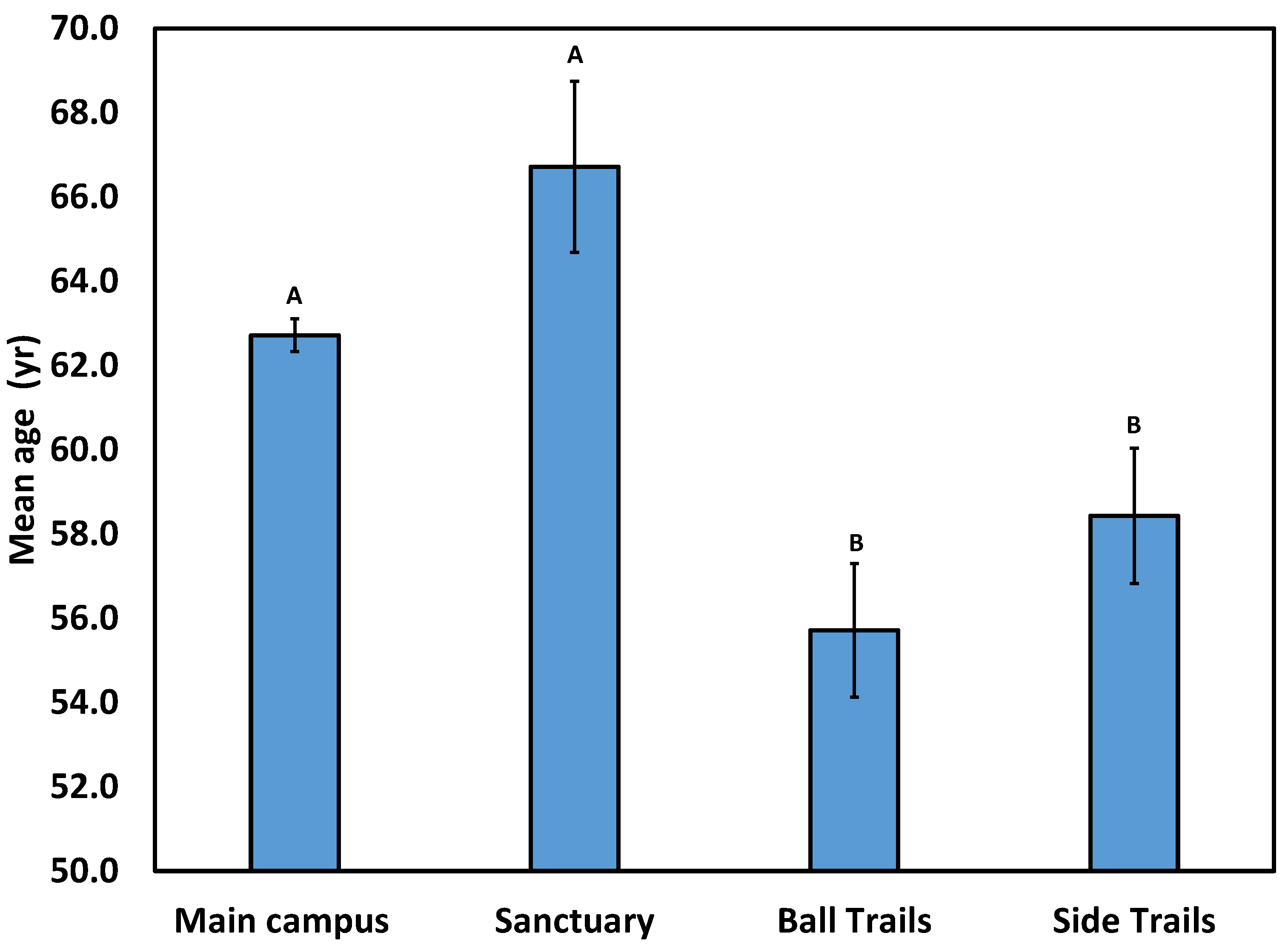
Figure 4.
Frequency age class distributions for four areas at UWF: main campus, Baars-Firestone Wildlife Sanctuary, Edward Ball Trails, and pine-dominated stands of the current study (see Figure 1 for locations).
Figure 4.
Frequency age class distributions for four areas at UWF: main campus, Baars-Firestone Wildlife Sanctuary, Edward Ball Trails, and pine-dominated stands of the current study (see Figure 1 for locations).
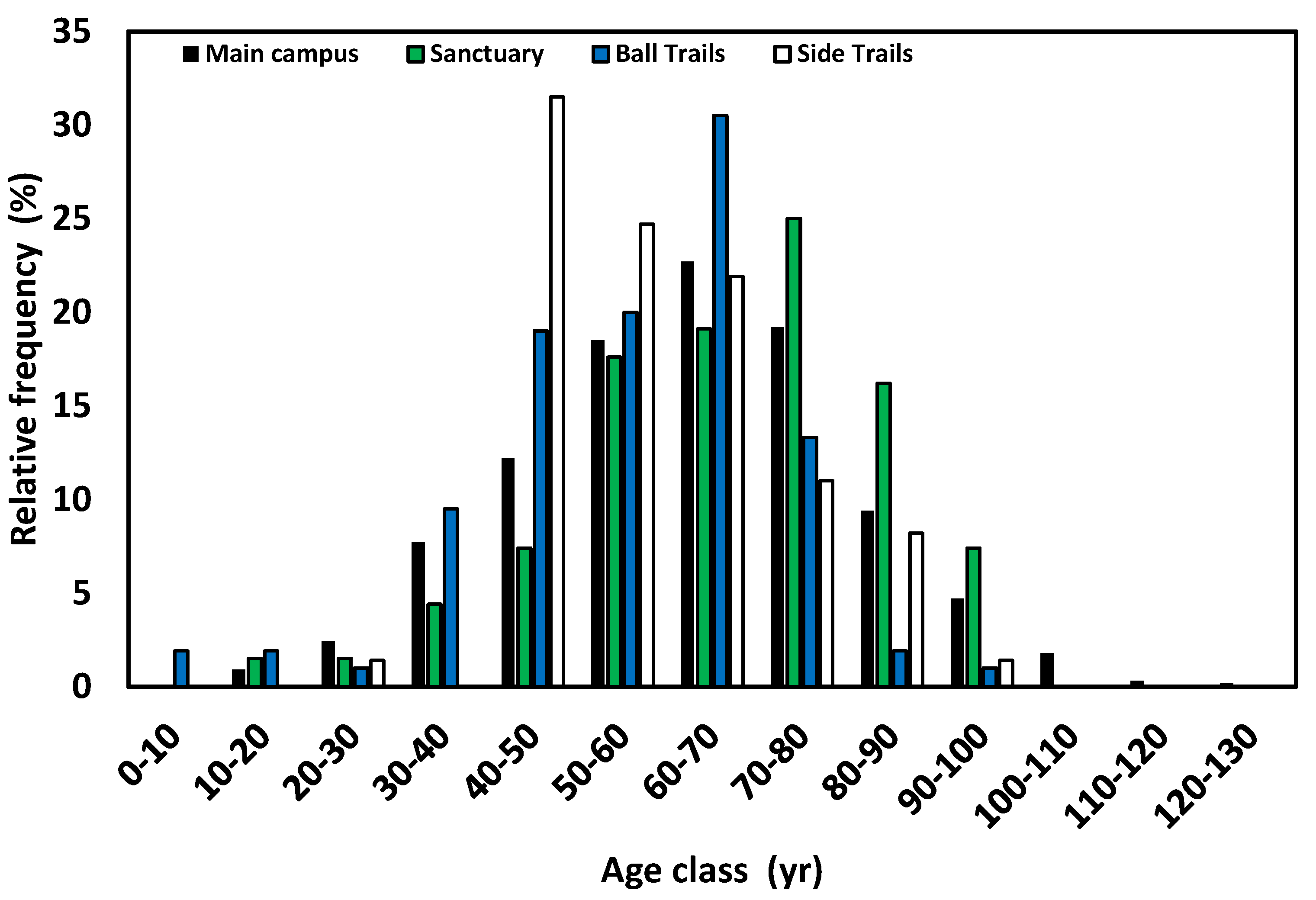
Figure 5.
Canonical correspondence analysis of overstory species in Ball Trails (T), Sanctuary (S), and Side Trails (P) sites. For vectors, element symbols are extractable concentrations of stated elements, ‘CEC’ is cation exchange capacity, ‘OM’ is organic matter, ‘pH’ is H2O-extractable soil pH, and ‘C’ and ‘N’ are total C and N, respectively. Species are indicated by four-letter codes: Fagus grandifolia (FAGR), Ilex opaca (ILOP), Ilex vomitoria (ILVO), Lagerstroemia indica (LAIN), Liquidambar styraciflua (LIST), Magnolia grandifolia (MAGR), Magnolia virginiana (MAVI), Nyssa sylvatica (NYSY), Oxydendrum arboreum (OXAR), Pinus palustris (PIPA), Prunus caroliniana (PRCA), Quercus alba (QUAL), Quercus falcata (QUFA), Quercus laurifolia (QULA), Quercus marilandica (QUMA), Quercus nigra (QUNI), Quercus virginiana (QUVI), Symplocos tinctoria (SYTI), Vaccinium arboretum (VAAR).
Figure 5.
Canonical correspondence analysis of overstory species in Ball Trails (T), Sanctuary (S), and Side Trails (P) sites. For vectors, element symbols are extractable concentrations of stated elements, ‘CEC’ is cation exchange capacity, ‘OM’ is organic matter, ‘pH’ is H2O-extractable soil pH, and ‘C’ and ‘N’ are total C and N, respectively. Species are indicated by four-letter codes: Fagus grandifolia (FAGR), Ilex opaca (ILOP), Ilex vomitoria (ILVO), Lagerstroemia indica (LAIN), Liquidambar styraciflua (LIST), Magnolia grandifolia (MAGR), Magnolia virginiana (MAVI), Nyssa sylvatica (NYSY), Oxydendrum arboreum (OXAR), Pinus palustris (PIPA), Prunus caroliniana (PRCA), Quercus alba (QUAL), Quercus falcata (QUFA), Quercus laurifolia (QULA), Quercus marilandica (QUMA), Quercus nigra (QUNI), Quercus virginiana (QUVI), Symplocos tinctoria (SYTI), Vaccinium arboretum (VAAR).
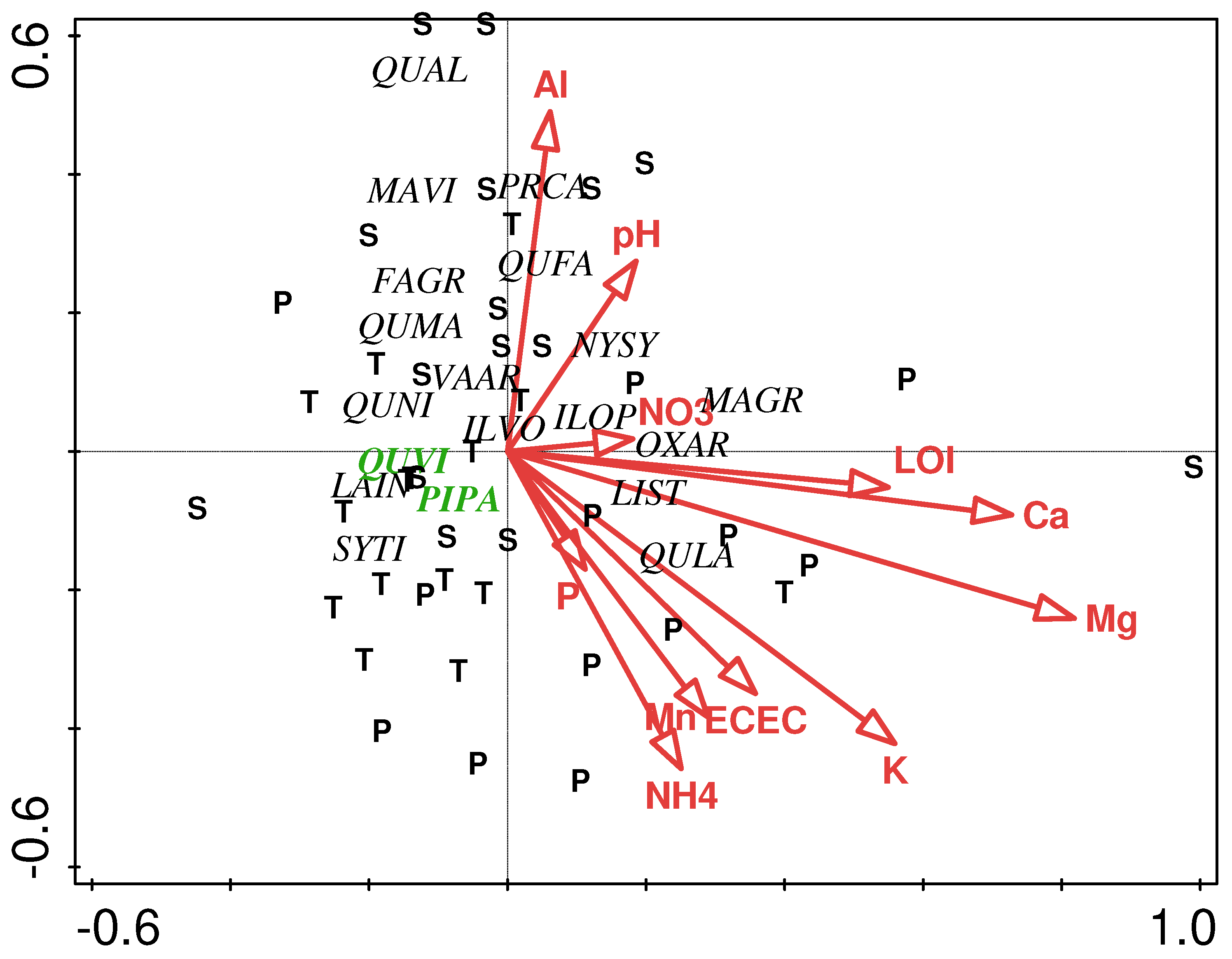

Table 1.
T-test comparisons of pre- vs post-fire exclusion soil variables. All nutrients/Al are extractable (see Methods). OM is organic matter; CEC is cation exchange capacity.
Table 1.
T-test comparisons of pre- vs post-fire exclusion soil variables. All nutrients/Al are extractable (see Methods). OM is organic matter; CEC is cation exchange capacity.
| Status | pH | OM | NO3-N | NH4-N | Ca | K | Mg | CEC | P | Al |
| % | µg/g | µg/g | µg/g | µg/g | µg/g | meq/100 g | µg/g | µg/g | ||
| Pre (n=32) | 5.37±0.08 | 2.09±0.13 | 0.23±0.05 | 2.60±0.43 | 175.2±36.1 | 16.8±1.9 | 22.0±2.5 | 1.51±0.17 | 0.37±0.03 | 52.0±3.8 |
| **** | * | NS | * | **** | **** | **** | NS | ** | **** | |
| Post (n=42) | 4.67±0.03 | 1.66±0.10 | 0.30±0.02 | 3.77±0.25 | 26.9±4.1 | 8.2±0.5 | 6.5±0.5 | 1.47±0.10 | 0.49±0.01 | 113.3±3.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



