
Submitted:
02 May 2023
Posted:
03 May 2023
You are already at the latest version
Abstract
Soil salinity is one of the most serious problems for achieving better crop growth and yield. The study was conducted to isolate, characterize and screen salt tolerant microbes from spinach grown in saline soils. The efficient salt tolerant microbes were characterized for beneficial traits such as nitrogen fixation, phosphate solubilization, indoleacetic acid (IAA), biofilm production, catalase and antagonistic activities, salt tolerance and optimization of bacterial fermentation. Molecular identification of potential microbes was done by 16S rRNA gene sequencing. The collected soil samples were moderately to strongly saline, alkaline in nature, low in N and P while adequate in K and Ca concentration. A total of 17 salt tolerant bacteria differing in colony and cell morphology were characterized. All isolated rhizobacterial microbes had capability to biosynthesize phytohormone IAA, 14 were able to fix atmospheric N2, 11 microbes solubilized inorganic P, 10 produced biofilm and 15 were positive in catalase activity. The isolated rhizobacteria showed salt tolerance up to 10 % induced salinity, only six microbes tolerated salt concentration of 12.5% however, none of any rhizobacteria remained viable at 15% NaCl amended media plates. The two potential salt tolerant microbes (NIA SP-6 & NIA SP-11) were screened and found for optimum growth curve and fermentation at pH 7.0 after 48 hrs of incubation period. Both potential microbes were identified as Enterobacter cloacae (NIA SP-06) and Bacillus safensis (NIA SP-11). Hence, it can be concluded that isolated salt tolerant rhizobacterial microbes have potential for beneficial traits and have ability for salt tolerance under saline conditions.
Keywords:
Activity
; beneficial traits
; bacterial count
; inoculation
; rhizosphere
; salinity
; salt tolerance
1. Introduction
Soil salinity is a global problem that affects the overall quality of the environment and the health of the soil. Salts have contaminated more than 900 million hectares of land. Salinity also reduces crop yields and threatens food security by compacting the soil and negatively impacting soil fertility [1,2]. In general, saline soils have high pH (>8.5) and high electrical conductivity (<4 dS m-1). In the world 76.6 Mha are salt affected soils among them more than 30% of the world’s saline soils are in China and 4.5 Mha are affected soils in Pakistan [3]. This is the factor that aggravates most of the problem. Although external and internal influences play a significant role the plant enhancement and crop production [4], in saline condition growth stress remains one of the most important stresses affecting plant growth and microbial survival [5]. Salinity stress reduces crop productivity by inhibiting plant development and reducing land utilization. As the global population increases at an alarming rate, industrialization and/or habitat use are reducing the amount of land available for agriculture. Therefore, salt-affected land must be utilized to meet the food demand. The salt concentration when increase from the normal levels it will inhibit plant development and crop productivity. Approximately twenty percent of cultivated land is compromised by soil salinity, which reduces crop production. Salt stress has two major effects on plants: osmotic stress and ionic toxicity. These types of pressures influence all main plant functions, such as photosynthesis, cellular metabolism and plant feeding [6]. Environmental elements that are both biotic and abiotic have a substantial impact on agricultural production. One of the main reasons restricting agricultural growth and production worldwide is the prevalence of saline-alkaline soils [7], with the application of chemical fertilizers to increase crop yields being the biggest obstacle. The essential nutrients play a vital role in the growth of plants in the soil. All living things need the element nitrogen to survive, which significantly lowers the productivity of terrestrial ecosystems. Another necessary nutrient that encourages the growth of plant roots and boosts crop production is phosphorus. Synthetic fertilizers are being used to meet these basic food requirements and their access use give negative affect to the ecosystem, soil fertility, and salinity [8]. Contrarily, utilizing phosphate fertilizer may have unfavorable outcomes such as runoff, leaching, denitrification, and volatilization [9]. The microbial population in the soil controls many ecosystem processes, such as the purification of contaminants, the cycling of nutrients, and the preservation of soil structure [10]. Salinity is widely acknowledged to play a crucial role in forming bacterial communities in various settings [11]. High soil salinity prevents some bacteria from growing, which in turn causes the selection to favour a community that can withstand salt [12,13]. However, through root colonization and the production of antimicrobial compounds, many beneficial bacteria can defend plant root surfaces against pathogenic microbes and plant stresses (both biotic and abiotic) [14]. In addition, the plant’s microbes drive growth despite challenging conditions like water shortage or high salt content. Salt-resistant plant microorganisms are favourable in challenging situations and can potentially increase crop productivity [15]. Several bacteria including Pseudomonas, Bacillus, Enterobacter, Agrobacterium, Streptomyces, Klebsiella, and Ochromobacter can increase crop productivity under saline circumstances and is well recognized [16,17,18]. The ability to perform various soil activities and maintain healthy ecosystems and the survival of plants and animals on earth depends on the presence of microorganisms in the soil. Soil health depends on many ecosystems [19] and microbial communities in healthy soil are more diversified in terms of the number of different species. One gram of soil is thought to include thousands of different soil types of microbes, including bacteria (108 to 109), actinomycetes (107 to 108) and fungi (105 to 106) [20]. Agriculture crops must be produced in a way that is both sustainable and takes use of plant-microbe interactions in the rhizosphere [21]. Spinacia oleracea L., also known as spinach, is a green leafy vegetable grown all over the world and a member of the Chenopodiaceae family [22]. Spinach is an excellent dietary fibre source since it is a highly favoured leaf vegetable with a high nutritional value, has several health benefits, and contains many significant vitamins and minerals [23,24]. Cultivation of valuable crops such as spinach is becoming more and more popular to reduce the negative impact of the salinization of water resources and water scarcity caused by drought in many areas [25,26]. In addition, insufficient nutrient supply negatively affects the growth and yield of spinach [27]. A decrease in leaf area is the main morphological change in spinach plants in response to biotic and abiotic stresses, which leads to lessen in productivity [28].
The salinity lessens the quantity and richness of bacterial communities substantially [29]. Soil microorganisms have the potential to be used as a biological indicator to assess and restore the quality of salinization. It has been demonstrated that the expression of bacteria that can endure in salty settings significantly increases plant growth and soil improvement. High-salt conditions can affect the growth of plants but due to presence of salt tolerant microbes which can to plants for growth under salt concentrations. Many strains of salt-tolerant bacteria have been found in saline crop soils [30], and it has been shown that some strains can fix N2 and solubilize phosphorus in the soil with other beneficial traits like IAA and biofim production, antagonistic effect, catalase activity and salt tolerance [31]. Furthermore, compared to simple chemical fertilizers, including salt-tolerant microbes in the inoculation process can increase crop yields in saline soil conditions. The use of these bacteria as an alternative source of plant nutrition saves the use of mineral fertilizers and protects the environment from damage [31]. Hence, study was carried out to isolate, characterize and screen salt-tolerant microbes from spinach grown in various saline concentrations.
2. Materials and methods
2.1. Study site and samples collection
Soil and plant samples were randomly collected from various spinach grown on saline soils of District Tando Allahyar, Sindh, Pakistan. The samples from various locations were labelled, kept in ice box and brought to the microbiology laboratory for further analysis.
2.2. Physico-chemical properties of soil
The soil samples collected from various spinach grown areas were analyzed for the various physico-chemical analysis followed by various methods (Table 1). A total of 11 saline locations were selected from where the soil and spinach plants samples were collected for this study.
2.3. Isolation, characterization and screening of isolated rhizobacteria
The soil and plant rhizosphere fresh samples were collected from the spinach grown areas and kept in icebox. The samples were transported to Soil Microbiology Laboratory for further rhizobacteria isolation, characterization and screening for improving spinach growth yield.
2.3.1. Total bacterial Count
The total bacterial count was done using a standard serial dilution plating technique [37]. Nutrient agar media was used for enumerating the total bacterial population following colony forming unit (CFU) per gram of soil. A sequence of serial dilution was organized and 0.01 mL aliquot was dropped onto the nutrient agar media plates for incubation (28 °Ċ) in incubator for 48-72 h. The bacterial quantity (CFU) was counted on the colony counter, and the population was enumerated.
2.3.2. Colony and cell morphology of isolated rhizobacteria
The isolated rhizobacteria microbes were purified on the media plates and colony morphology was recorded. A loop of the pure culture was kept on glass slide. The emulsion oil was dropped on the pure culture and bacterial cell morphology was observed under the microscope.
2.3.3. Gram reaction
A 48-hour-old pure bacterial culture underwent a Gram staining test. Gram-stained slides were handled using a heat-fixed rhizobacteria smear that had been saturated with two drops of crystal violet for one minute. The crystal violet was gently cleaned with sterile distilled water, then sealed with an iodine smear for a minute before being washed with water. The stain was carefully removed by gently dropping an alcohol solution using a pipette. Smears were counter-stained with safranine for 30 seconds after being rinsed with water. After being cleaned with sterile distilled water, the slides were allowed to air dry and observed under microscope. Gram negative strains were reddish or pink in color, while gram positive strains were bluish or purple in color.
2.3.4. Atmospheric Nitrogen (N2) fixing ability
Atmospheric Nitrogen (N2) fixing ability of isolated microbes was determined using by inserting one loop of bacterial culture in the N2 fixing bacteria (Nfb) semisolid media [38]. The ability was determined by the pellicle formation in the inoculated medium. The pellicle forming strain was recorded positive for nitrogen fixing trait.
2.3.5. Phosphate solubilization
According to Pikovskaya’s method, the phosphate-solubilizing activity of microbes was assessed [39] the rhizobacteria culture was dropped in the center of agar medium, after one week of incubation at (28-32 ºC) the media plates were checked for the formation of clear halo zone surrounding the bacterial colony and calculated using by following formula [40].
2.3.6. Biofilm production
The mechanical shaker device was used to incubate the bacterial broth cultures and biofilm production was noticed after 72 hours. The tissue plate approach was used to find biofilm development. As a result of positive biofilm-producing bacteria developing a film on it [41].
2.3.7. Antagonistic activity
The antagonistic effect of the potential isolated microbes was tested against fungal (Fusarium sp.) by dual culture method [42]. Fungal strain was grown in the center of potato dextrose agar (PDA) medium plate. Bacterial culture was grown on the both sides of growing fungi with a distance of two centimeter. The culture was incubated for seven days on ambient temperature. Inhibition activity was calculated on the basis of radius of normal growing hyphae compared to bacterial culture and their inhibition.
2.3.8. Catalase production
The production of catalase was measured by applying bacterial colony on the surface of a dry glass slide with a sterile wooden stick loop. The bacterial colony was given a drop of hydrogen peroxide (3% H2O2) and was thoroughly mixed. The quick evolution of oxygen (within 5 to10 sec.) and the creation of bubbles were evidence of the successful outcome. However, a drawback was that colonies did not form any bubbles after hydrogen peroxide was applied and mixed with them [43].
2.3.9. Indole-3-acetic acid (IAA) production activity
The IAA production of selected rhizobacterial microbes was estimated by inoculating the bacterial culture and incubated on the mechanical shaker at 120 rpm (28 °C for 48 hour). The culture was supplemented with 200μg/ml L-tryptophan and incubated for further 72 hrs ambient temperature of 28-32 °C. After 4-7 days, the cells were centrifuged at 8000 rpm for 10 min and 2ml of supernatant was taken and 2ml of Salkowski’s reagent (0.5 M FeCl3, 35% HClO4, 2:100) was added. The mixture was left for 30 min at room temperature for development of pink colour [44]. The absorbance of sample was detected at 535 nm on spectrophotometer and the concentration was projected by the curve of IAA standards (0-35 ppm).
2.4. Screening and identification of salt tolerance rhizobacteria isolated from spinach growing areas
2.4.1. Screening the of salt resistant rhizobacteria
The selected microbes isolated from spinach vegetable were evaluated for their efficacy for salt tolerance. Different salinity levels (NaCl: 0, 0.5, 1.0, 1.5, 2.0, 2.5, 3.0, 3.5, 4.0, 5.0, 7.5, 10.0, 12.5 and 15.0 %) were developed in nutrient agar media plates. The isolated rhizobacteria were grown on respective media plates. The efficiency of isolated rhizobacteria was determined by presence/existence colony formation units on respective media plates. The bacterial survival and count was estimated after 48-72 hrs. The most tolerant microbes were selected for further studies.
2.4.2. Potential salt resistant rhizobacterial growth Curve and biomass measurement
Two potential salt tolerant bacterial strains (NIA SP-6 & NIA SP-11) were on the basis of their salt tolerance under various salinity levels selected for growth curve measurement. The bacterial strains were inoculated in broth culture and incubated at 37°C with rotation rate 120 rpm for 72 hours on mechanical shaker. The growth curve was measured by monitoring optical density (OD600) every 24 hrs using by plate count method [45]. Finally the bacterial culture was centrifuged and the supernatant was removed and pellet was collected for the determination of biomass weight.
2.4.3. Optimization of bacterial fermentation condition under various pH levels
The selected potential microbes (NIA SP-6 & NIA SP-11) were inoculated in broth culture modified with various pH levels (6.0, 7.0, 8.0 & 9.0). The fermentation was carried out from 0 to 72 hrs and the production rate was measured after 24 hrs intervals. The most suitable pH of the fermentation bacterial media was determined.
2.4.4. Optimization of carbon and nitrogen (C:N) ratio
The potential microbes (NIA SP-6 & NIA SP-11) were inoculated in Lysogeny broth. The bacterial culture was incubated at 37 oC on at mechanical shaker for 72 hours at pH 7.0. The optimal proportion of carbon and nitrogen sources was measured by varying the added proportions to 1:9, 1:4, 2:3, 1:1, 3:2, 4:1 and 9:1.
2.4.5. Molecular identification (16S rRNA Gene Sequencing) of potential microbes
Extraction of total genomic DNA
Two potential microbes (NIA SP-06 and NIA SP-11) were selected for 16S rRNA partial gene sequencing analysis. Alkaline lysis method was employed to extract the total genomic DNA from the selected bacterial isolates with slight modifications [46]. A 1.5 mL aliquot from five mL of fully grown bacterial culture was centrifuged at 13000 × g for 2 min Cells were re-suspended in 567 µL TE buffer (10 mM tris HCl and 1 mM EDTA) by pipetting. The mixture was incubated at 37 °C for 1 h, or till the time they became transparent after adding 30 µL of 10% (w/v) SDS and 3 µL of 20 mg mL-1 proteinase K (Fermentas). The mixture was added with 100 µL of 5 M NaC1, mixed thoroughly and then 80 µL cetyle trimethyle ammonium bromide/ sodium chloride (CTAB/NaCl) solution (10% (w/v) CTAB and 0.7 M NaC1, 50 mM tris-HCl pH 8, 25 mM EDTA) were added. The mixture was mixed properly again and incubated at 65 °C for 10 minutes. An equal volume (780 µL) of phenol: chloroform: isoamyle alcohol (25: 24: 1) was added and gently mixed by inverted shaking. The supernatant was collected at 13000 × g for 10 min in 1.5 mL tubes. The supernatant was added with 0.6 volume (360 µL) isopropanol and incubated at 20 oC for 30 min. The tubes were again centrifuged at 13000 × g for 10 min and supernatant was discarded. The DNA pellet was then washed with 70% (v/v) ethanol and dissolved in appropriate amount of (70-100 µL) TE buffer.
Amplification and sequencing of 16S rRNA
Amplification and sequencing of 16S rRNA gene was carried out using primers fD1 (5--AGAGTTTGATCCTGCTCAG-3,) and rD1 (5--AAGGAGGTGATCCAGCC-3,) as described earlier [47] with modification. Fifty µL reaction was prepared in purified water having: 5µL of Taq buffer (Fermentas), 1.5 µL of 50 mM MgCL2 (Fermentas), 5 µL of 2 mM dNTPs (Fermentas), 0.5 µL of 100% DMSO, 1.5 µL each of forward and reverse primer, 0.75 µL of 5 U µL-1 Taq DNA polymerase (Fermentas) and 50 ng of template DNA. Polymerase chian reaction was carried out in PCR machine (PeQLab, advanced Primus 96) and temperature conditions were also modified as: 30 cycles of 95 oC for 2 min, 55 °C for 30 sec, and 72 °C for 5 min. The initial denaturation and final extension steps was carried out at 95 centigrade for 5 min and 72 centigrade for 10 min, respectively. Amplified PCR products were purified using QIAquick PCR purification kit (Qiagen, USA). The purified amplicons were sequenced directly by Eurofins, Germany. Both ends were joined by Caps 3 assembly software and compared with others in the GenBank database using the NCBI BLASTn technique.
2.5. Statistical Analysis
The collected data and biochemical analyses exposed with 5 replicates. Significance at the 5% level was tested followed by Tukey’s studentized range using SAS statistical program version 9.1 (SAS Institute, Cary, NC, USA, 2008).
3. Results
3.1. Sample collection and, bacterial isolation and characterization from the spinach grown on saline soil
The soil and plant samples of spinach were randomly collected from the spinach grown in the salt affected areas from District Tando Allahyar, Sindh, Pakistan at various crop ages for further analysis.
3.1.1. Physio-chemical properties of soil samples
A total of eleven (11) saline locations were selected from where the soil and spinach plants samples were collected (Table 2). The physico-chemical properties of the samples were carried out.
3.1.2. Isolation characterization of salt tolerant rhizobacteria from spinach grown under saline soil
Generally soil of the various spinach grown areas was alkaline (pH >8.0) and moderately to strongly saline (>4.0dS m-1) soil in nature. The soil organic matter contents (<0.86%) were very low except a few samples. The soil of sample collection site was deficient in nitrogen, AB-DTPA extractable P and was sufficient in K. The soil texture was medium to heavy in all locations. The Ca and Na concentrations were higher while Mg+ concentration was low in the collected soil samples (Table 3).
3.1.3. Total bacterial count in rhizosphere
Total bacterial count was in collected from spinach rhizosphere samples. Significant variations in bacterial count were reported in the rhizosphere of spinach growing locations. Relatively, the higher bacterial abundance was observed among all the locations however, significantly (P<0.05) the highest bacterial population (42×107 CFU g-1 soil) was found in Muhammad Afzal Khawaja, Khawaja stop, Tando Allahyar followed by (23×107cfu g-1 soil) Rustam Khawaja, near civil hospital, Sultanabad locations (Table 4).
3.1.4. Biochemical characterization
The isolated microbes were observed under light microscope for their morphological characteristics. The isolated microbes were then selected on the basis of their colony morphology for further biochemical characterization. A total of seventeen (17) rhizobacterial strains were selected. Thirteen (13) were gram positive while four were Gram negative in reaction (Table 5). Mostly all isolates have shown N2 fixing ability except SP-7, SP-12 & SP-13 similarly twelve (12) strains possess their P-solubilization and antagonistic effect against Fusarium sp. A total of twelve (12) isolates were able for mineral biofilm production on sold surface and fifteen were positive in catalase test (Table 5).
3.1.5. Determination of IAA production
The isolated indigenous rhizobacteria microbes were able to produce indoleacetic acid (IAA) after supplementing L-tryptophan. Among isolated strains, SP-4 (22.18 mg L-1), followed by SP-6 (20.76 mg L-1) significantly (P<0.05) produced the highest IAA in comparison to other isolates while the lowest (7.33 mg L-1) was produced by SP-7 (Figure 1).
3.1.6. Phosphate solubilizing activities
A total of eleven (11) microbes were able to solubilize P from insoluble P source. Among all, isolates SP-11 significantly (P<0.05) solubilized the highest P (70%) followed by SP-6 (50%), while six isolates (SP-1, SP-7, SP-12, SP-13, SP -15 and SP-17) were not able to solubilize P on the P insoluble modified media plates (Figure 2).
3.2. Screening and identification of salt tolerance rhizobacteria isolated from spinach growing areas
3.2.1. Salt tolerance efficiency of rhizobacteria
The isolated microbes were tested for salt tolerance using various salt concentrations (NaCl 0 to 15%) during the incubation period at (37 °C) in in vitro conditions. Most of the isolated microbial culture showed salt tolerance ranges from 10 to 7.5% salt concentrations. At 12.5% salt concentration SP-5, SP-6, SP-8, SP-9, SP-11and SP-13 were able to grow while rest of isolated microbes were not able to show their tolerance. When the salt concentration was 15% none of the rhizobacteria was able to grow/survive (Table 6).
3.2.2. Rhizobacteria growth in response to increasing salt (NaCl) concentration
The relative bacterial growth was observed at various salt concentrations (NaCl 0%, 1%, 2.5%, 5%, 7.5%, 10%, 12.5% and 15 %) mediated media culture during the incubation period in in vitro conditions. There was growth response depending on the isolated strains and varied among the microbes. Mostly all the isolated strains were able to grow on salt concentration their response up to 7.5% salt concentration. Most of the different bacterial strains were able to show their growth on salt concentration up to 12.5%. However, none of the isolated grown on salt concentration of 15% (Figure 3).
3.2.3. Potential salt resistant rhizobacterial growth curve measurement and bacterial biomass
The bacterial inoculum was cultured in media at optimal temperature (37 °C) and growth curve was measured at various incubation interval periods. The maximum growth activity of both potential rhizobacterial strains was observed after 48 hrs then activity was decreased significantly (Figure 4a). The maximum bacterial biomass was recorded in 40.80 g L-1 followed by 28.0 g L-1 in NIA SP-11: Bacillus safensis and NIA SP-6: Enterobacter cloacae during the incubation period respectively (Figure 4b).
3.2.4. Optimization of bacterial fermentation condition under various pH levels
The bacterial fermentation activity was measured at different pH levels (6.0, 7.0, 8.0 and 9.0). The optimization of bacterial fermentation condition was varied under various pH levels. The optimum pH value of the fermentation media of both bacterial strains NIA SP-6: Enterobacter cloacae and NIA SP-11: Bacillus safensis was at pH 7.0 after 48 hrs. However, the activity was showing decline with the increase of pH during incubation period. The minimum activity was observed at pH 9.0 after 72 hrs of incubation period (Figure 5).
3.2.5. Optimization of carbon and nitrogen ratio
The incubation temperature was set at 37 °C for incubation period from 0 to 72 hrs at pH 6.0, with 75 ml of bacterial culture and carbon and nitrogen ratio (1:9, 1:4, 2:3, 1:1, 3:2, 4:1 and 9:1) was kept for optimization. The results showed that the both NIA SP-06: Enterobacter cloacae and NIA SP-11: Bacillus safensis strains showed better growth and higher enzymatic activity in the fermentation medium with carbon and nitrogen ratio of 1:1. However, the maximum activity was observed by NIA SP-11: Bacillus safensis strain (Figure 6).
3.2.6. Bacterial identification
The isolated microbes from the spinach grown in saline soil were identified as NIA SP-06: Enterobacter cloacae and NIA SP-11: Bacillus safensis using by 16S rRNA Gene Sequencing technique. In this study, we used gene sequences where different species of Enterobacter and Bacillus species were determine the phylogenic relationship among the tested strains. The Neighbor-Joining tree was subjected to the numerical re-sampling by bootstrapping, and the resulting bootstrap values are shown at the tree branch nodes. Each value represents the number of times (out of 1000 replicates) that the represented groupings occurred in the re-samplings. The consensus tree showed 08-100%% confidence levels whereas NIA SP-06 Enterobacter cloacae strain had a 99% confidence level and NIA SP-11 also showed the 99% confidence level with Bacillus safensis strain VS-16 with accession numbers NR MT507092.1 and NR MK785115.1 respectively (Figure 7).
4. Discussion
The saline soils are common and one of the significant issues for crop production, due to salinity, plants suffer and hindering crop yield in these areas. Salinity-induced abiotic stresses provide severe problems to crop growth on a global scale. It affects plant development and agricultural productivity when it is subjected to extreme salinity level. It occurs directly in areas of high osmotic stress, leading to a rise in ionic toxicity and a fundamental transformation of the physicochemical properties of the soil. Therefore, the efforts to alleviate salinize stress got much attention, including the isolation and application of salt-tolerant microbes in this study. Eleven (11) saline locations were selected from where the soil and spinach plants, along with adhering soil samples, were collected. The soil analysis indicated that the sampled area was saline and the spinach plant had salt tolerance ability. Most of the studied soils were deficient in main nutrients N and P except K and high in Ca, Na and Mg. Rhizobacteria were present in all the sampled locations. Seventeen (17) rhizobacterial microbes with the differing colony and cell morphology were isolated from the collected samples. The rhizobacterial microbes were available in saline soils and can survive under various salinity levels [48]. In addition, soil salinity is a worldwide problem, and finding salt-tolerant microbes to alleviate the salinization problem along with other management practices is a sustainable and economical solution to this problem [49].
The screened salt-tolerant rhizobacterial microbes have the potential to fix atmospheric nitrogen, solubilize mineral P from soil, and antagonistic against plant pathogens. The soil microorganisms carried out numerous processes necessary for the development of plant growth, including supplementing fixed nitrogen under various soil stress situations [48]. Previous report showed several types of beneficial microbes such as Pseudomonas, Azotobacter, and other microbes which solubilize insoluble P, release exopolysaccharides, produced biofilm and indole-3-acetic acid (IAA), siderophores as well as enzymes to tolerate under saline conditions [48]. These attributes might be more supportive for plants directly for plant growth and root initiation. The antagonist effect of the isolated bacterial strains could protect plants from fungal diseases such as Fusarium sp.. Furthermore, isolated microbes had the ability for biofilm and enzyme production. The bacterial strains produced a higher amount of indoleacetic acid (IAA) which may lead to the growth promotion of crop plants by altering root architecture. Naher et al. [50] noticed that IAA production by the bacterial strains improve plant growth enhancement of several crops. The microbes isolated from the salt affected soils are reported to possess phosphate solubilization activity determined through halo zones performances on Pikovaskaya media plates [39] and it may solubilize fixed Ca-P in saline soil and improve bioavailable P to the plant grown in saline stress.
The isolated bacteria existed and survived at various salt concentrations of 7.5%, whereas, at the higher salt concentrations (NaCl from 7.5 to 12.5%), a few tolerant microbes could survive. However, none of inoculated microbes could survive beyond 15% of NaCl induced salinity level. Similar findings were reported in the earlier study by Qurashi et al. [51] who found that the salt tolerant microbes have the capability to survive under saline stress conditions and among them have variability in the survival and their bacterial population at various salinity levels in the NaCl amended media culture. Further, it was investigated in vitro effects of NaCl concentrations on microbes from a variety of characteristics which is expected to assist the considerate the physical changes of bacterial strains under salt stress conditions that might help to clarify the association between bacterial durability and virulence outside hosts [52]. It was found that in saline conditions, microbes can bind with the sodium ion via secretion of EPS which, therefore, lessens its toxicity in the soil [53] In the present study salt tolerant gummy microbes exposed their existence at various salinity levels due to their tolerant efficiency as these microbes might have the ability for EPS-production and that could chelate various cations including sodium. Bacterial biofilm production is closely related to EPS production. Several studies proved that microbial biofilm attached to the roots improve soil fertility and reduce salinity stress [54].
Furthermore, it is essential to ensure high microbial activity at the vicinity of plant rhizosphere to get benefit of these novel strains. To ensure survival of the bacteria, cell lysis, C:N and pH of the growth media checked. In this study, it has been observed during the incubation period microbes showed their exponential growth and that was the optimized period for increase in cell concentration in in vitro condition at various salt concentrations. It was known that the various strains have different incubation period and pH for their growth and optimization fermentation that mostly depends on their cell lysis and other activities [52].
In the study two potential strains based on their biochemical properties selected and were identified using as 16S rRNA partial gene sequencing as NIA SP-06: Enterobacter cloacae and NIA SP-11: Bacillus safensis. Both of the selected potential rhizobacterial microbes showed maximum growth and optimization fermentation after 48 hrs at pH 7.0 with the increase of pH and incubation period the activity was minimized. The suitable carbon and nitrogen ratio for enzymatic activity was observed for both rhizobacterial microbes was found 1:1 as the carbon and nitrogen are essential for the bacterial growth and their ratio is very much important for the production of cellulose [55]. The beneficial traits, enzymatic and physicochemical properties that can help to identify high enzymes ability and stability can be used for the development of bio-fertilizer for applications in crops [52]. Bio-chemical characters of the identified microbes from the spinach grown in saline soil proved and found both of the microbes have beneficial traits, enzymatic and biochemical properties that can help for nutrient fixation and their solubility in soil and production of growth hormones so that they can be used for the development of bio-fertilizer.
5. Conclusions
The soil and plant samples were collected from spinach grown under moderately to strongly saline (>4.0 dS m-1) conditions. A number of microbes (11 locations) were isolated from the spinach rhizosphere and a total of seventeen (17) salt tolerant microbes that differ in colony morphology having multiple beneficial traits were documented. The majority (13) of microbes were Gram positive, 14 were N2 fixers, 15 were positive in the catalase test, 12 were P-solubilizers, IAA and 10 biofilm producers. The isolated microbes were able to tolerate salinity levels up to 10% (NaCl); however, six (6) microbes showed their tolerance at 12.5% (NaCl) amended media culture. Two potential bacteria were identified using as 16S rRNA partial gene sequencing as NIA SP-06: Enterobacter cloacae and NIA SP-11: Bacillus safensi. The optimum pH value of the fermentation media of Enterobacter cloacae and Bacillus safensi was at pH 7.0 after 48 hrs with better growth and enzymatic activity at 1:1 carbon and nitrogen ratio. Biochemical characterization proved both strains have potential to reduce stress of Spinach grown in saline soil.
Author Contributions
Conceptualization, S.P.M., Q.A.P, X.W. and Q.A.P., S.P.M; methodology, S.P.M., S.R.M., and Q.A.P; formal analysis, S.P.M. and M.Q.; data curation, X.W., Q.A.P. and S.P.M. and M.U.Y.; writing—original draft preparation, S.P.M. and U.A.N.; writing—review and editing, X.W. and Q.A.P.; visualization, Q.A.P.; supervision, X.W.; funding acquisition, X.W. All authors have read and agreed to the published version of the manuscript.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to confidentiality.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pau, D. Osmotic stress adaptations in rhizobacteria.” J. basic Microbiol. 2013, 53(2), 101-110.
- Nagler, K.;Moeller, R. Systematic investigation of germination responses of Bacillus subtilis spores in different high-salinity environments.” FEMS Microbiol. Ecol. 2015, 91(5), 23. [CrossRef]
- Li, X.; Wan, W.; Luo, X.; Zheng, L.; He, G.; Huang, D.; Chen, W. Huang, Q. “High salinity inhibits soil bacterial community mediating nitrogen cycling.” Appl. Environ. Microbiol. 2021, 87(21), e1366-21. [CrossRef]
- Mao, X.;Yang, Y.; Guan,P.; Geng, L.; Ma,L.; Hongjie Di; Wenju Liu, Bowen Li. Remediation of organic amendments on soil salinization: Focusing on the relationship between soil salts and microbial communities.” Ecotoxicol. Environ. Safety. 2022, 239, 113616. [CrossRef]
- Nadeem, S.M.; Ahmad, M.; Naveed, M., Imran, M., Zahir, Z.A.; Crowley, D.E. Relationship between in vitro characterization and comparative efficacy of plant growth-promoting rhizobacteria for improving cucumber salt tolerance. Archives Microbiol. 2016, 198, 379–387. [CrossRef]
- Havlin, J.L.; Soltanpour. A. nitric acid plant tissue digest method for use with inductively coupled plasma spectrometry. Commun. Soil Sci. Plant Anal. 1980, 11(10), 969-980. [CrossRef]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: physiological, biochemical, and molecular characterization. 2014, 70, 1596. [CrossRef]
- Li, Y.; Sun, Y.; Zhang, H.; Wang, L.; Zhang, W., Niu, L.; Wang, P.; Wang, C. The responses of bacterial community and N2O emission to nitrogen input in lake sediment: estrogen as a co-pollutant.” Environ. Res. 2019, 179, 108769. [CrossRef]
- Zhang, K.; Yu Shi, Xiaoqing Cui, Ping Yue, Kaihui Li, Xuejun Liu, Binu M. Tripathi, Haiyan Chu. Salinity is a key determinant for soil microbial communities in a desert ecosystem. Msystems. 2019, 4(1), e00225-18. [CrossRef]
- Chaparro, J.M.; Dayakar V. B.; Jorge M. Rhizosphere microbiome assemblage is affected by plant development. The ISME J. 2014, 8(4), 790-803.
- Gavrichkova, O.; Brykova, R.A., Brugnoli, E., Calfapietra, C., Cheng, Z., Kuzyakov, Y., Liberati, D., Moscatelli, M.C. Secondary soil salinization in urban lawns: Microbial functioning, vegetation state, and implications for carbon balance. Land Degrad. Dev. 2020, 31(17), 2591-2604. [CrossRef]
- Rath, K.M.; Rousk J. Salt effects on the soil microbial decomposer community and their role in organic carbon cycling: a review. Soil Biol. Biochem. 2015, 81, 108–123. [CrossRef]
- Zhang, J.; Xiao, Q.; Guo, T.; Wang, P. Effect of sodium chloride on the expression of genes involved in the salt tolerance of Bacillus sp. strain “SX4” isolated from salinized greenhouse soil. Open Chem., 2021, 19(1), 9-22.
- Jiao, S.; Lu, Y.H. Abundant fungi adapt to broader environmental gradients than rare fungi in agricultural fields. Global Change Biol. 2020, 26, 8, 4506-4520. [CrossRef]
- Niu, X.; Song, L.; Yinong X.; Weide G. Drought-tolerant plant growth-promoting rhizobacteria associated with foxtail millet in a semi-arid agroecosystem and their potential in alleviating drought stress. Frontiers Microbiol. 2018, 8, 2580.
- Singh, R.P.; Jha, P.N. The multifarious PGPR Serratia marcescens CDP-13 augments induced systemic resistance and enhanced salinity tolerance of wheat (Triticum aestivum L.).” PLos one. 2016, 11, 6. e0155026. [CrossRef]
- Sharma, S.; Kulkarni, J.; Jha, B. Halotolerant rhizobacteria promote growth and enhance salinity tolerance in peanut.” Frontiers Microbiol. 2016, 7, 1600. [CrossRef]
- Sarkar, A.; Ghosh, P.K.; Pramanik, K.; Mitra, S., Soren, T., Pandey, S., Mondal, M.H.; Maiti, T.K. A halotolerant Enterobacter sp. displaying ACC deaminase activity promotes rice seedling growth under salt stress.” Res. Microbiol. 2018, 169(1), 20-32. [CrossRef]
- Prakash, O.; Sharma, R.; Rahi, P.; Karthikeyan, N. Role of microorganisms in plant nutrition and health. Nutrient use efficiency: from basics to advances. 2015, pp. 125–161.
- Microbes, S.; Understanding soil microbes and nutrient recycling. Actinomycetes. 2020, 107, 40–50.
- Glick, B.R.; Gamalero, E. Recent developments in the study of plant microbiomes. Microorganisms. 2021, 9(7), 15-33. [CrossRef]
- Caparrotta, S.; Masi, E; Atzori G.; Diamanti I.; Azzarello E.; Mancuso S.; Pandolfi, C. Growing spinach (Spinacia oleracea) with different seawater concentrations: Effects on fresh, boiled and steamed leaves. Scientia Horticulturae. 2019, 256, 108540. [CrossRef]
- Shannon, M.C.; Grieve C.M. Shannon, M. C., and C. M. Grieve. Tolerance of vegetable crops to salinity. Scientia Horticulturae, 1998, 78(1-4), 5-38.
- Thomas, R.M.; Verma, A.K.; Prakash, C., Krishna, H.; Prakash, S.; Kumar, A. Utilization of Inland saline underground water for bio-integration of Nile tilapia (Oreochromis niloticus) and spinach (Spinacia oleracea). Agri. Water Manage. 2019, 222,154-160. [CrossRef]
- Zheng, P.; Bai, X.; Long, J., Li, K.; Xu, H. Nitric oxide enhances the nitrate stress tolerance of spinach by scavenging ROS and RNS. Scientia Horticulturae. 2016, 213, 24–33. [CrossRef]
- Ors, S.; Suarez, D.L. Spinach biomass yield and physiological response to interactive salinity and water stress.” Agri. Water Manage. 2017, 190, 31–41. [CrossRef]
- Ibekwe, A.M.; Ors, S.; Ferreira, J.F.; Liu, X.; Suarez, D.L. Seasonal induced changes in spinach rhizosphere microbial community structure with varying salinity and drought.” Sci. Total Environ. 2017, 579, 1485–1495.
- Çakmakçı, R.; Erat, M.; Erdoğan, Ü.; Dönmez, M.F. The influence of plant growth–promoting rhizobacteria on growth and enzyme activities in wheat and spinach plants. J. Plant Nutr. Soil Sci. 2007, 170, 288–95. [CrossRef]
- Shi, S.; Tian, L.; Nasir, F.; Bahadur, A., Batool, A., Luo, S., Yang, F., Wang, Z. and Tian, C. Response of microbial communities and enzyme activities to amendments in saline-alkaline soils. Appl. Soil Ecol. 2019, 135, 16–24. [CrossRef]
- Zhang, X.; Qi, L.; Li, W.; Hu, B.X. Dai, Z. Bacterial community variations with salinity in the saltwater-intruded estuarine aquifer. Sci. Total Environ. 2021, 755,1424. [CrossRef]
- Othman, N.M.I.; Othman, R.; Zuan, A.T.K.; Shamsuddin, A.S.; Zaman, N.B.K.; Sari, N.A.; Panhwar, Q.A. 2022. Isolation, characterization, and identification of zinc-solubilizing bacteria (ZSB) from wetland rice fields in Peninsular Malaysia. Agriculture, 2022, 12, 1823.
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37(1), 29-38. [CrossRef]
- Jones, J.B. Laboratory guide for conducting soil tests and plant analysis. Chemical Rubber Company (CRC) Press, 2001.
- Bouyoucos, G.J. Hydrometer method improved for making particle size analyses of soils 1.” Agron. J. 1962, 54(5), 464-465. [CrossRef]
- Bremner, J.M.; Mulvaney, C.S. Nitrogen–total. In ‘Methods of Soil Analysis, Part 2. Chemical and microbiological properties’, 2nd edn. (Eds A.L. Page; R.H. Miller; D.R. Keeney) pp. 595–624. Soil Science Society of America, Inc. and American Society of Agronomy, Inc.: Madison, WI 1982.
- Havlin, J.L.; Soltanpour. A. nitric acid plant tissue digest method for use with inductively coupled plasma spectrometry. Commun. Soil Sci. Plant Anal. 1980, 11(10), 969-980. [CrossRef]
- Somasegaran, P.; Hoben, H.J. Methods in legume-Rhizobium technology. Paia, Maui: University of Hawaii NifTAL Project and MIRCEN, Department of Agronomy and Soil Science, Hawaii Institute of Tropical Agriculture and Human Resources, College of Tropical Agriculture and Human Resources, 1985, 1–52.
- Gyaneshwar, P.; James, E.K.; Mathan, N.; Reddy, P.M., Reinhold-Hurek, B.; Ladha, J.K. Endophytic colonization of rice by a diazotrophic strain of Serratia marcescens. J. Bacteriol. 2001, 183(8), 2634–2645.
- Pikovskaya, R.I.; Mobilization of phosphorus in soil in connection with vital activity of some microbial species. Mikrobiologiya, 1948, 17, 362–370.
- Nguyen, C.; W. Yan, W.; F. Le Tacon Lapeyrie, F. Genetic variability of phosphate solubilizing activity by monocaryotic and dicaryotic mycelia of the ectomyccorhizal fungus Laccaria bicolor (Maire) P.D. Orton. Plant Soil, 1992, 143, 193–199.
- Watnick, K.; Koltre. Biofilm, City of Microbes. J. Bacteriol. 2000, 182, 2675–79.
- Sariah, M. Potential of Bacillus spp. as biocontrol agent for anthracnose fruit rot of chilli. Malaysian Appl. Biol. 1994, 23, 53–60.
- Serra, B.; Zhang, J., Morales, M.D., de Prada, A.G.V., Reviejo, A.J.; Pingarron, J.M. A rapid method for detection of catalase-positive and catalase-negative bacteria based on monitoring of hydrogen peroxide evolution at a composite peroxidase biosensor. Talanta. 2008, 75(4), 1134–1139. [CrossRef]
- Gordon, A.S.; Weber, R.P. Colorimetric estimation of indole acetic acid. Plant physiol. 1951, 26(1), 192.
- Morales-Borrell, D.; González-Fernández, N.; Silva, R.S.; Gómez, E.S. Optimization of fermentation conditions of bacterium Pseudoxanthomonas indica H32.” Biotechnol. Reports. 2021, 32, e00676. [CrossRef]
- Xue, Z.; Kable, M.E.; Marco, M.L. Impact of DNA sequencing and analysis methods on 16S rRNA gene bacterial community analysis of dairy products.” Msphere. 2018, 3(5), e00410-18. [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173(2), 697-703.
- Sadiq, H. M.; Jahangir, G. Z.; Nasir, I. A.; Iqtidar, M.; Iqbal, M. Isolation and Characterization of Phosphate-Solubilizing Bacteria from Rhizosphere Soil. Biotechnol. Biotechnol. Equipment. 2014, 27(6), 4248-4255. [CrossRef]
- Mao, X.; Yang, Y.; Guan, P.; Geng, L.; Ma, L.; Di, H.; Liu, W,; Li, B,; Remediation of organic amendments on soil salinization: Focusing on the relationship between soil salts and microbial communities. Ecotoxicol. Environ. Safety. 2022, 239, 113616. [CrossRef]
- Naher, U.A.; Biswas, J.C.; Maniruzzaman, M.; Khan, F.H.; Sarker, M.I.U.; Jahan, A.; Hera, M.H.R.; Hossain, M.B.; Islam, A.; Islam, M.R.; Kabir, M.S. Bio-Organic Fertilizer: A Green Technology to Reduce Synthetic N and P Fertilizer for Rice Production. Frontiers Plant Sci. 2021, 12, 152. [Google Scholar]
- Qurashi, A.W.; Sabri, A.N. Osmoadaptation and plant growth promotion by salt tolerant bacteria under salt stress. Afr. J. Microbiol. Res. 2011, 5(21), 3546–3554. [Google Scholar]
- Li, F.; Xiong, X.S.; Yang, Y.Y., Wang, J.J., Wang, M.M., Tang, J.W., Liu, Q.H., Wang, L.; Gu, B. Effects of NaCl concentrations on growth patterns, phenotypes associated with virulence, and energy metabolism in Escherichia coli BW25113. Frontiers Microbiol. 2021, 12, 705326. [CrossRef]
- Abuhena, Md.; Al-Rashid, J.; Azim, M.F.; Morshed, M. N.; Golam Kabidr, M.K.; Barman, N.C.; Rasul, N.M.; Akter, S.; Huq, M.A. Optimization of industrial (3000 L) production of Bacillus subtilis CW-S and its novel application for minituber and industrial-grade potato cultivation. Sci. Rep. 2022, 12, 11153. [Google Scholar]
- Shultana, R.; Zuan, A.T.K.; Naher, U.A.; Islam, A.K.M.M.; Rana, M.M.; Rashid, M.H.; Irin, I.J.; Islam, S.S.; Rim, A.A.; Hasan, A.K. The PGPR Mechanisms of Salt Stress Adaptation and Plant Growth Promotion. Agronomy 2022, 12, 2266. [Google Scholar] [CrossRef]
- Sharma, K.M.; Kumar, R.; Panwar, S.; Kumar, A. Microbial alkaline proteases: Optimization of production parameters and their properties. J. Genet. Eng. Biotechnol. 2017, 15(1), 115–126. [Google Scholar] [CrossRef]
Figure 1.
IAA production by potential rhizobacteria isolated from spinach. Means within the same column followed by the same letters are not significantly different at P<0.05.
Figure 1.
IAA production by potential rhizobacteria isolated from spinach. Means within the same column followed by the same letters are not significantly different at P<0.05.
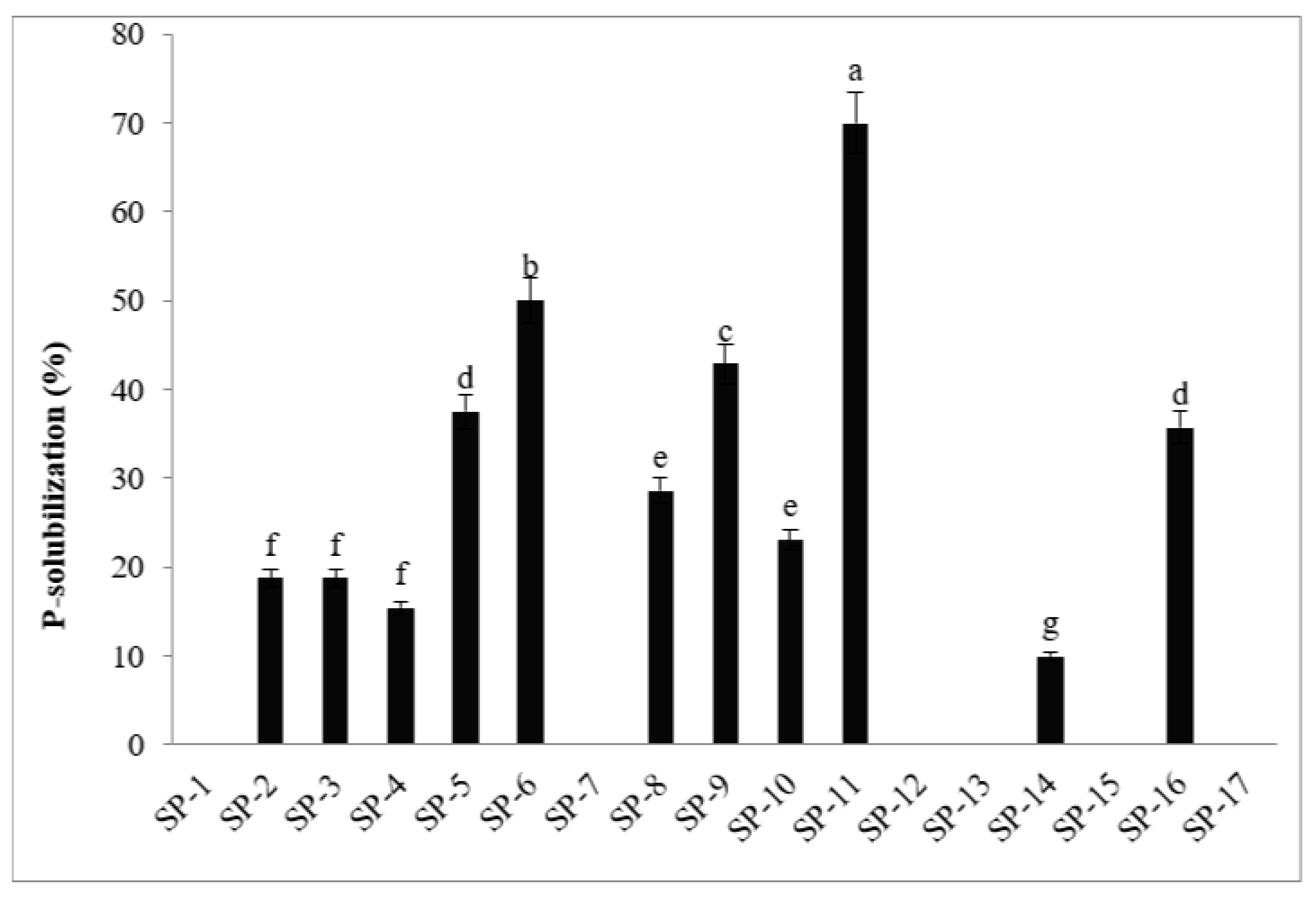
Figure 2.
Mineral Phosphate solubilization by potential rhizobacteria strains isolated from spinach. Means within the same column followed by the same letters are not significantly different at P<0.05.
Figure 2.
Mineral Phosphate solubilization by potential rhizobacteria strains isolated from spinach. Means within the same column followed by the same letters are not significantly different at P<0.05.
Figure 3.
Correlation of salt (NaCl) concentration and rhizobacterial growth.
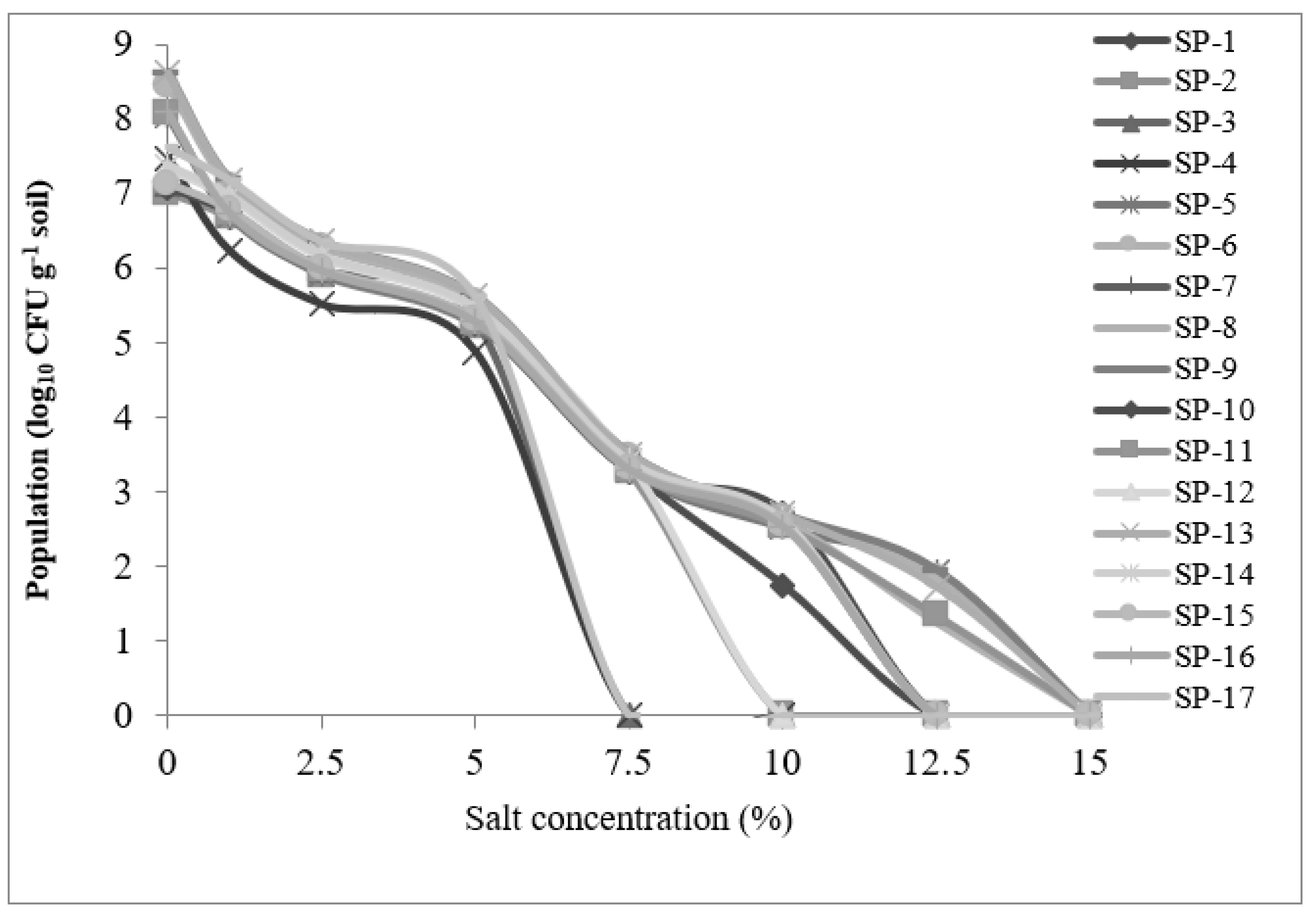
Figure 4.
(a) Growth curve and (b) bacterial biomass of salt tolerant rhizobacteria in enriched medium.
Figure 4.
(a) Growth curve and (b) bacterial biomass of salt tolerant rhizobacteria in enriched medium.
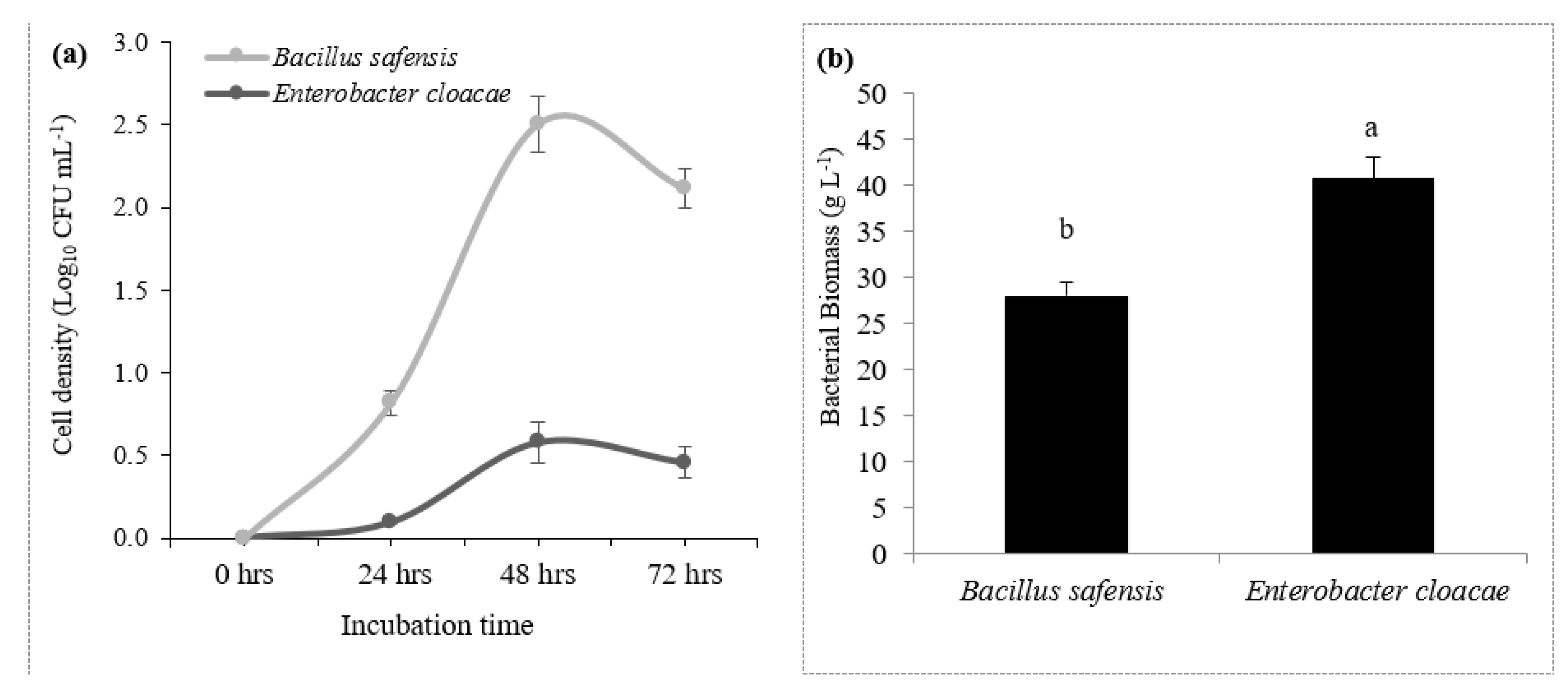
Figure 5.
Optimization of bacterial fermentation condition under various pH levels.
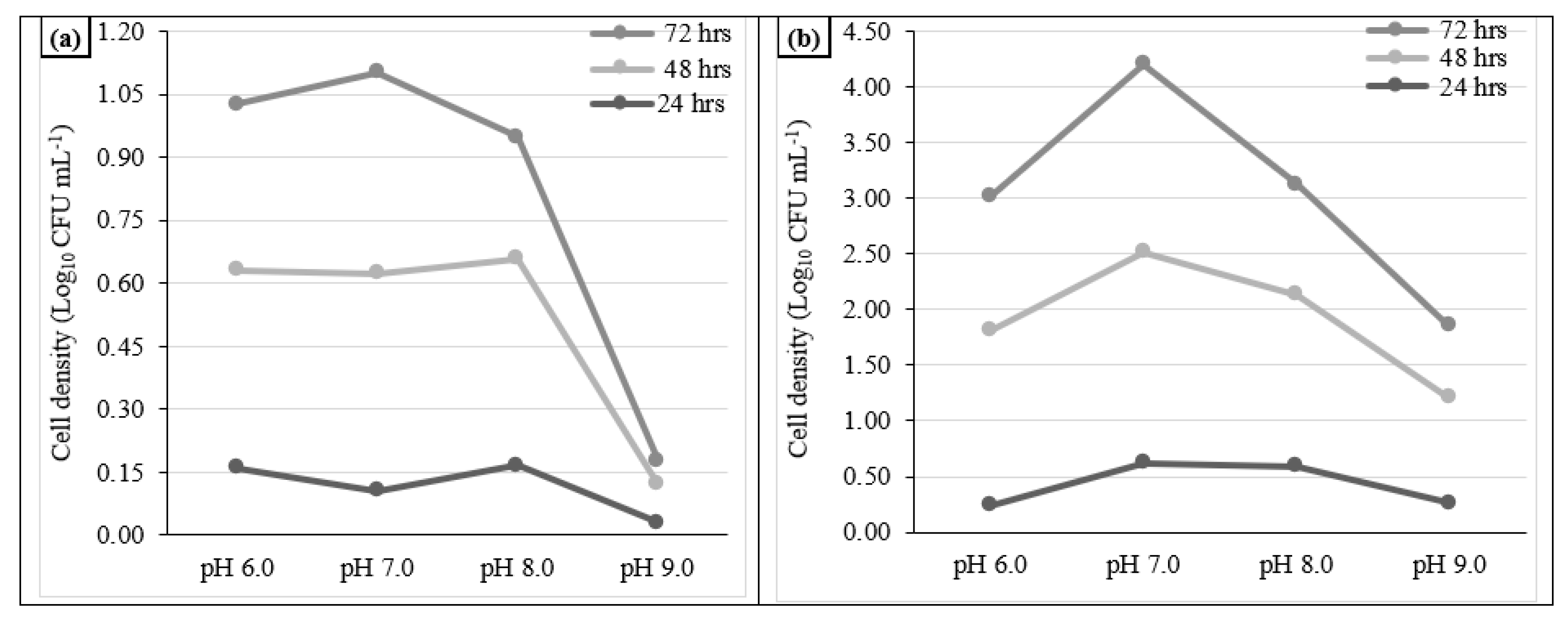
Figure 6.
Optimization of fermentation conditions secreted by NIA SP-06: Enterobacter cloacae and NIA SP-11: Bacillus safensis on enzyme production.
Figure 6.
Optimization of fermentation conditions secreted by NIA SP-06: Enterobacter cloacae and NIA SP-11: Bacillus safensis on enzyme production.
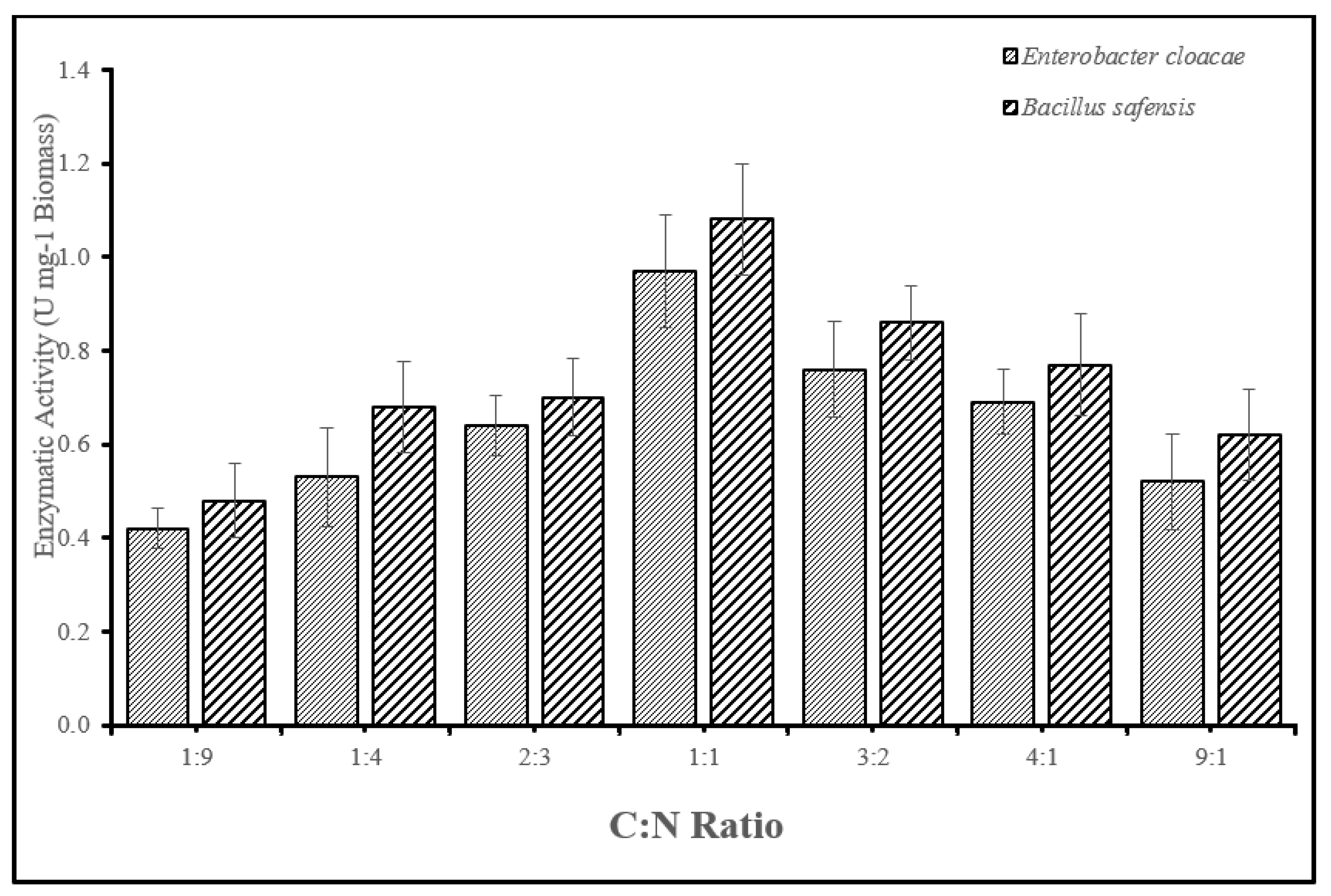
Figure 7.
Phylogenetic tree with bootstrap values. Tree constructed using Neighbor-Joining (NJ) method. NIA SP-06: accession NR MT507092.1 and NIA SP-11 accession NR MK785115.1.
Figure 7.
Phylogenetic tree with bootstrap values. Tree constructed using Neighbor-Joining (NJ) method. NIA SP-06: accession NR MT507092.1 and NIA SP-11 accession NR MK785115.1.


Table 1.
Analytical methods used for analysis.
| No. | Parameter | Analytical Procedure |
|---|---|---|
| 1. | Organic matter | Walkley and Black (1934) method [32] |
| 2. | Soil pH and EC | Soil: water (1:2.5) extract using PHM210 standard pH meter and EC meter [33] |
| 3. | Soil texture | Bouyoucos hydrometer method [34] |
| 4. | Total N | Kjeldahl method [35] |
| 5. | Exchangeable P | Wet digestion [36] |
| 6. | Exchangeable cations (Ca, Mg, Na and K) | Wet digestion [36] |
Table 2.
Collection of spinach and soil plant samples from different locations of district Tando Allahyar.
Table 2.
Collection of spinach and soil plant samples from different locations of district Tando Allahyar.
| No. | Farmer’s Nam | Location | GPS Location | Variety | Method of sowing | Crop Age |
|---|---|---|---|---|---|---|
| 1. | Muhammad Afzal Khawaja | Khawaja stop | 25°29’32.1”N68°51’15.9”E, | local | Ridge sowing | 30 days |
| 2. | Muhammad Afzal Khawaja | Tando Allahyar | 25°29’51.8”N68°53’04.0”E | local | Ridge sowing | 40 days |
| 3. | Ghulam Mustafa | Awami Markaz road, Nawazabad | 25°30’02.9”N 68°51’14.7”E | local | Ridge sowing | 45 days |
| 4. | Rustam Khawaja | Nawazabad | 25°32’04.4”N 68°52’27.7”E | Hybrid | Ridge sowing | 65 days |
| 5. | Rustam Khawaja | Near Civil Hospital, Sultanabad | 25°28’09.8”N 68°43’15.4”E | local | Ridge sowing | 60 days |
| 6. | Nawaz Khawaja | Village Kamal Khan Mastoi | 25°34’14.0”N 68°49’32.4”E | local | Ridge sowing | 45 days |
| 7. | Shah Nawaz | Village Shah Nawaz | 25°33’19.5”N 68°48’02.2”E | local | Ridge sowing | 50 days |
| 8. | Muhammad Asif | Village DatoKarloo | 25°30’11.1”N 68°47’30.3”E | local | Ridge sowing | 55 days |
| 9. | Muhammad Asif | Village Ghulam Hussain Lund | 25°30’14.6”N 68°45’10.5”E | Hybrid | Ridge sowing | 45 days |
| 10. | Shah Nawaz | Village Kaamaro Sharif | 25°29’09.3”N 68°47’59.7”E | local | Ridge sowing | 50 days |
| 11. | Mubeen Laghari | Village JadoLaghari | 25°27’03.7”N 68°47’31.3”E | local | Ridge sowing | 45 days |
Table 3.
Physio-chemical properties of soil samples collected from spinach fields of Tando Allahyar.
Table 3.
Physio-chemical properties of soil samples collected from spinach fields of Tando Allahyar.
| No. | Farmer’s Name | Location | OM (%) |
EC (dS/m) |
pH | Texture | N (%) |
P (ppm) |
K (ppm) | Ca (mqL-1) |
Na (ppm) |
Mg (mqL-1) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1. | Muhammad Afzal Khawaja | Khawaja stop | 1.27 | 4.52 | 8.5 | Silty loam | 0.04 | 2.9 | 177 | 40.6 | 0.48 | 60.65 |
| 2. | Muhammad Afzal Khawaja | Tando Allahyar | 0.78 | 8.80 | 8.7 | Clay loam | 0.03 | 3.0 | 158 | 36.0 | 0.47 | 52.03 |
| 3. | Ghulam Mustafa | Awami Markaz road, Nawazabad | 0.92 | 7.36 | 8.8 | Silty clay loam | 0.03 | 2.9 | 105 | 33.24 | 0.43 | 42.61 |
| 4. | RustamKhawaja | Nawazabad | 0.63 | 10.12 | 8.6 | Clay | 0.03 | 2.6 | 120 | 41.23 | 0.49 | 31.23 |
| 5. | Rustam Khawaja | Near civil hospital, Sultanabad | 0.87 | 6.77 | 8.4 | Clay loam | 0.04 | 2.3 | 167 | 34.28 | 0.31 | 45.67 |
| 6. | Nawaz Khawaja | Goth Kamal Khan Mastoi | 0.69 | 7.84 | 8.4 | Silty loam | 0.04 | 3.2 | 156 | 37.71 | 0.37 | 49.15 |
| 7. | Shah Nawaz | Village Shah Nawaz | 1.02 | 3.92 | 8.7 | Silty loam | 0.06 | 3.0 | 177 | 39.76 | 0.34 | 36.74 |
| 8. | Muhammad Asif | Village DatoKarloo | 0.94 | 6.26 | 8.5 | Silty loam | 0.04 | 2.9 | 143 | 40.01 | 0.32 | 14.78 |
| 9. | Muhammad Asif | Village Ghulam Hussain lund | 0.85 | 7.90 | 8.4 | Clay loam | 0.03 | 2.9 | 171 | 35.51 | 0.36 | 38.10 |
| 10. | Shah Nawaz | Village Kaamaro Sharif | 1.20 | 4.52 | 8.6 | Silty clay loam | 0.03 | 2.5 | 124 | 37.32 | 0.37 | 36.54 |
| 11. | Mubeen Laghari | Village Jado Laghari | 1.05 | 5.49 | 8.0 | Silty loam | 0.04 | 2.6 | 161 | 38.13 | 0.35 | 40.14 |
Table 4.
Total bacterial count of isolated strains from various locations of spinach grown areas.
| No. | Farmer’s Name | Location | Total bacteria count (CFU g-1 soil) |
|---|---|---|---|
| 1. | Muhammad Afzal Khawaja | Khawaja stop,Tando Allahyar | 12×107c |
| 2. | Muhammad Afzal Khawaja | Khawaja stop,Tando Allahyar | 42×107a |
| 3. | Ghulam Mustafa | Awami Markaz road, Nawazabad | 14×107c |
| 4. | RustamKhawaja | Nawazabad | 2×107e |
| 5. | Rustam Khawaja | Near civil hospital, Sultanabad | 23×107b |
| 6. | Nawaz Khawaja | Goth Kamal Khan Mastoi | 21×107b |
| 7. | Shah Nawaz | Village Shah Nawaz | 11×107c |
| 8. | Muhammad Asif | Village DatoKarloo | 6×107d |
| 9. | Muhammad Asif | Village Ghulam Hussain Lund | 5×107d |
| 10. | Shah Nawaz | Village Kaamaro Sharif | 14×107c |
| 11. | Mubeen Laghari | Village Jado Laghari | 13×107c |
Means within the same column followed by the same letters are not significantly different at P<0.05.
Table 5.
Biochemical characteristics of potential rhizobacteria isolated from spinach fields.
| No. | Isolates | Colony Morphology | Gram Staining | Nitrogen fixing ability | Phosphate- solubilizing ability | Biofilm production | Antagonistic effect (%) | Catalase test |
|---|---|---|---|---|---|---|---|---|
| 1 | SP-1 | Long rod | -ve | + | - | - | - | + |
| 2 | SP-2 | Rod shape | -ve | + | + | + | - | + |
| 3 | SP-3 | Short rod | +ve | + | + | + | - | +++ |
| 4 | SP-4 | Short rod | +ve | + | + | + | + | - |
| 5 | SP-5 | Short rod | +ve | + | + | + | - | + |
| 6 | SP-6 | Long rod shape | +ve | + | + | - | + | +++ |
| 7 | SP-7 | Short rod shape | +ve | - | - | ++ | - | +++ |
| 8 | SP-8 | Medium rod shape | -ve | + | + | - | +++ | + |
| 9 | SP-9 | Short rod | +ve | + | + | +++ | +++ | ++ |
| 10 | SP-10 | Short rod | +ve | + | + | - | +++ | + |
| 11 | SP-11 | Round | -ve | + | + | + | +++ | + |
| 12 | SP-12 | Round shape | +ve | - | - | + | ++ | +++ |
| 13 | SP-13 | Long rod shape | +ve | - | - | + | + | + |
| 14 | SP-14 | Round shape | +ve | + | + | - | + | - |
| 15 | SP-15 | Long rod shape | +ve | + | + | + | +++ | + |
| 16 | SP-16 | Short rod | +ve | + | + | - | +++ | + |
| 17 | SP-17 | Short rod | +ve | + | - | - | +++ | +++ |
| Note: +ve = positive, -ve = negative | ||||||||
Table 6.
Salt tolerance efficiency of bacterial strains isolated from various locations of spinach grown areas
Table 6.
Salt tolerance efficiency of bacterial strains isolated from various locations of spinach grown areas
| No. | Strain | Salt concentration (%) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 0.5 | 1 | 1.5 | 2.0 | 2.5 | 3.0 | 3.5 | 4 | 5 | 7.5 | 10 | 12.5 | 15 | ||
| 1 | SP-1 | +++ | +++ | +++ | + | + | + | + | + | + | + | + | + | - | - |
| 2 | SP-2 | +++ | +++ | + | + | + | + | + | + | + | + | + | - | - | - |
| 3 | SP-3 | +++ | +++ | + | + | + | + | + | + | + | + | - | - | - | - |
| 4 | SP-4 | +++ | +++ | + | + | + | + | + | + | + | + | - | - | - | - |
| 5 | SP-5 | +++ | +++ | +++ | +++ | +++ | ++ | ++ | ++ | ++ | + | + | + | + | - |
| 6 | SP-6 | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | ++ | ++ | + | - |
| 7 | SP-7 | +++ | +++ | +++ | +++ | ++ | + | + | + | + | + | + | + | - | - |
| 8 | SP-8 | +++ | +++ | ++ | ++ | ++ | ++ | + | + | + | + | + | + | + | - |
| 9 | SP-9 | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | ++ | + | + | - |
| 10 | SP-10 | +++ | +++ | +++ | +++ | ++ | ++ | ++ | ++ | ++ | + | + | + | - | - |
| 11 | SP-11 | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | +++ | ++ | ++ | + | - |
| 12 | SP-12 | +++ | +++ | ++ | ++ | + | + | + | + | + | + | + | - | - | - |
| 13 | SP-13 | +++ | +++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | + | + | + | + | - |
| 14 | SP-14 | +++ | +++ | ++ | ++ | ++ | ++ | + | + | + | + | + | + | - | - |
| 15 | SP-15 | +++ | +++ | ++ | ++ | + | + | + | + | + | + | + | + | - | - |
| 16 | SP-16 | +++ | +++ | ++ | ++ | ++ | ++ | + | + | + | + | + | + | - | - |
| 17 | SP-17 | +++ | +++ | +++ | ++ | + | + | + | + | + | + | - | - | - | - |
| Note: - Sensitive, + Slightly tolerant, ++ Moderately Tolerant, +++ Tolerant | |||||||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



