
Submitted:
04 April 2023
Posted:
05 April 2023
You are already at the latest version
Abstract
The Asian weaver ant (Oecophylla smaragdina) is a natural enemy, generalist predator of diverse major pest species (i.e. the highly destructive oriental fruit fly Bactrocera dorsalis) in economically strategic agricultural landscapes in Australia and Southeast Asia countries. For effective implementation of the weaver ant for biological control of the invasive bagworms Metisa plana, the dominant pest in the oil palm plantation, its distribution was investigated in Malaysia. From 2018-2022, censuses were first carried out to record Oecophylla colonies presence. Oecophylla colonies’ occupation patterns were monitored on 8 selected plantations. To distinguish between brood and barracks nests, 26 captured nests were evaluated to define the main predictor variable for its practical visual identification. More than 11000 palms with 10821 nests for over 500 colonies were sampled in this study. Results showed that O. smaragdina colonies distribution pattern exhibited three-dimensional ubiquitous occupation dominance. We also found that by observing the colony’s dense population spatial arrangements, it demonstrates them as spreading by waves featuring interconnected irregular geometrical patches shaped for each colony. The height to the ground nest location in palm canopies was the significant visual discriminant factor between Brood and barrack nests. Moreover, polydomous arboreal nesting behavior occupation occurred irrespective of palms or associated host plants sizes (≥ 3 m), throughout the year. However, colony nesting behaviors were polydomous and monodomous on shorter plants (≥ 2 m), while being strictly monodomous on small plants (≤ 1.5 m). Colonies occupied an average 3 to 4 palms yearly beginning from the newly founding phase up to the stable mature phase (3 to 4 years). Furthermore, we found that matured stable colonies (3 ≥ age ≥13) occupied 10-12 palms on average, on 800 m2 to 2500 m2 area delimited by a minimum average 20 m2 of “no ants land” zone between each colony. Our results suggest that a sustained agriculture system in this area would be possible with the presence of matured colonies nests exploitation which could provide an efficient biological control ecosystem service.
Keywords:
Asian weaver ants
; arboreal dominance
; Elaeis guineensis
; spatial distribution
; ecosystem service
; biological control agent
; novel nesting behavior
1. Introduction
The Asian weaver ants (Oecophylla smaragdina) occur from India to the Pacific and from South China to North Australia [1]. Oecophylla smaragdina spatial and geographical distribution throughout Asia has been documented with detailed coordinates [2]. However, the current distributional limit of O. smaragdina in oil palm (Elaeis guineensis) plantations in Peninsular and Borneo Malaysia had not been properly examined. The Asian weaver ants is a proven biological control agent (BCA) of the red-banded thrips, Selenothrips rubrocinctus, an important global pest of economic importance in Australian mango (Mangifera indica) [3]. O. smaragdina it is also a component of integrated pest management (IPM) program to control the fruit fly, Bactrocera jarvisi, a major pest of mango in Australia [4]. Considering the potential application to keep the invasive bagworms, Metisa plana, below the economic threshold level (ETL) in oil palm plantations [5,6], O. smaragdina distribution need to be verified to ensure easy access and the availability of its nests in the oil palms [7] for population manipulations [8].
Weaver ant is well-known for the leaf nests it builds. The nests were constructed by pulling and rolling living leaves in place and binding them together with larval silk [1]. Oecophylla spp. colony expansion is polydomous [9,10], i.e., it consists of multiple nests for each occupied palm tree with the possibility of having several nests in a single palm frond [11]. The nests of a single colony are frequently distributed in the crowns of several trees expanding on variable land surfaces depending on the colony population density [1,12].
Before elaborating an exploitation of the Asian weaver ant colonies as a potential natural enemy that could contribute to biological control in the study area it is compulsory to differentiate which stage between incipient-immature and mature colonies have more predatory potential over the main pest in oil palm crop. Emergent colonies occupy only one palm tree with one nest [1]. Matured colonies are determined by the first emergence of sexual forms and able to reach such a development growth stage upon 18 to 24 months is occupying a higher host tree number [13]. On this note, the measurement method of queen abdomen [9], and workers size [14] to estimate colony age is expensive and time consuming, hence not suitable. Ref. [15] discovered that the lifespan of both queen and workers are dependent on the colony size. The population size of Asian weaver ants is related to the average of total number of occupied trees and nest’s abundance, hence to the type of colonies as incipient or matured [1]. Both colonies can be found within the occupied zone, with clear boundaries of unoccupied trees or artificial plantations borders between palm tree blocks (waterways or roads that provide clear separation) or isolated in a newly explored habitat [11]. Weaver ant average lifespan estimation is helpful for rapid matured colonies detection. This prevents the translocation of vulnerable incipient, immature or pre-mature colonies that could act as biological control agents in commercial crops [16]. Further, a visual discrimination between brood and barrack nests is proposed, hence encouraging only for the capture of barrack nests having only major workers [1], with a balanced mixture of brood nests. Ref. [17] defined brood nest as being colonies main nests consisting of all type of weaver brood ants (eggs, larvae, pupae, reproductive adult forms and workers), while barracks (“pavilions”) represented only satellite nests covering the periphery of territory [18] inhabited only by major veterans workers [19]. Three types of nests were clearly defined as queen right, brood and barracks by [19].
An important aspect concerns the pleometrosis (multiple queens) or haplometrosis (single queen) and territorial status of Oecophylla colonies [20]. In pleometrotic colonies, there is higher brood production to shorten the period of claustral initial growth phase development, hence increase survival [21]. The fecundity of queens in pleometrotic colonies, is higher than haplometrotic colonies [20,21], is inviting to raise the point that colonies having several queens will attain maturity faster than single colonies queens. As a territorial species, with the high densities of founding queens, intraspecific elimination between incipient colonies occurs, increasing mortality and inducing low survival impact between incipient colonies [22,23]. Adding to this, neighboring colonies risk raiding [24] as a natural selection factor, dominant colonies’ population size and territorial distribution expand [25]. This condition needs to be considered during sample collection to evaluate diverse Oecophylla colonies in different geospatial zones.
To our knowledge, there is no information on the spatial distribution of Asian weaver ants’ colonies in major agricultural industries for at least large monoculture. This is necessary to convince the industry entrepreneurs of the worth if the Asian weaver ants could provide an important ecosystem service, by biological control of the recurrent outbreaks of the invasive dominant bagworms in the oil palm crop [26,27]. This may help planters to enhance their pest management program implementation for different agricultural products to attain higher quality with lower cost of management minimizing the use of phytosanitary applications [28]. This study proposes to gather information to estimate the average age of O. smaragdina colonies in oil palm plantations. For sustained nest manipulation in future biological control treatments, we hypothesized that the total nests, occupied palms numbers per colony is correlated to its lifespan up to reaching stable maturity stage growth development by an average of 36 to 42 months. Given that, by direct visual observation we count nests, being faster, and avoid destructive sampling.
The objectives of this study are; (1) to document the distribution of O. smaragdina colonies and boundary estimation of colonized palms, (2) to investigate the colonies occupancy patterns in relation to the nests network existence in canopies, (3) to discriminate between barrack and brood nests visually, by comparing their location heights from the ground and, (4) to estimate colonies age in relation to the average total number of nests and total number of occupied trees per colony.
2. Materials & Methods
2.1. Study sites
The study was conducted in Malaysia (supplementary material Figure S1a,b). Field experiments were approved by the Malaysian Palm Oil Board MPOB and the Federal Land Development Authority FELDA agreement with “Universiti of Malaya” (UM). The study was conducted in both Peninsular and Borneo Malaysia (Supplementary material, Figure S1). Such immense planted area (Sarawak: 1.5 million ha – Sabah: 1.54 million ha and 2.77 million ha for Peninsular) [29] obliged to subdivides the surveys into five geographical zones.
Oecophylla colonies occurrences were recorded as follows: (i) Northern Peninsular (NP), (ii) Eastern Peninsular (EP) (iii) Central Peninsular (CP) (iv), Southern Peninsular (SP) and (v) Malaysia Borneo (MB). The geographical coordinates covered were from “ladang” (plantations in Malay) are presented as supplementary information (Supplementary material, Table S1).
2.2. Surveying the presence-absence of Asian weaver ants in oil palm plantations
An extensive census was conducted in each geographical zone from January 2018 to December 2022 (Figure 1a-h.). Intensive censuses were performed, by establishing five plots (0.5 ha each) covering the center and four extremities of the plantation area (Figure 2). The surveys recorded the existence of canopies-dwelling, ground-dwelling, or any nests on oil palm trees. Binoculars and monocular field glasses were used to help in the detection. During an earlier period of sampling (2018-2019), 12 hours each day of observations was performed to lessen to 6 hours from 2020 onward. Following the approach of [30] and [2] we surveyed all major estates. To locate the occupied areas, the geographical coordinates were extracted using Google Landsat Maxar maps technology CNES/Airbus and GPS Essentials software with Samsung Galaxy Note20 model from September 2020 to examine the colonies spatial distribution in each territory to identify its specific patterns. Weaver ants were observed and monitored during daylight from early morning on random sampled palm trees canopies, trunks and ground vicinity by two hours intervals once during 10 minutes for each observation (0700 hours to 1900 hours, N > 1000 palms).
The host plants associated with colonies of O. smaragdina (promoting expansion) inside and nearby (1 km radius) oil palm plantations were recorded systematically by taking photos of the whole tree and zoom on leaves, flowers/fruits whenever available. Associated favorite plants are beneficial as a conservation buffer zone enhancing Asian weaver ant population stability [19]. Further identification was carried out by a botanist (Supplementary material, Table S2). The plant species IDs were confirmed by the Singapore government portal website “NParks Flora & Fauna Web” plant database (https://www.nparks.gov.sg/florafaunaweb, accessed on 23 February 2023). The presence of antagonist ants such as the invasive Solenopsis geminata and black ant Dolichoderus thoracicus was also recorded by performing censuses along a 1000 m path in all occupied sampled weaver ant in the plantation (See result related figures and discussion). Targeted species were hand-picked and placed in 80 % ethanol vial solution for further identification. Antagonistic ant species are reported to harm the population strength of both Oecophylla species [9,31].
2.3. Nests occupancy patterns in selected plantations
To obtain a more representative size of the variety of colonies, the sampling collection was performed on selected plantations, representing diverse geographical locations (Supplementary material, Table S2). To determine sampling size, we carried out a preliminary study to estimate the average number of weaver ant colonies occupying palm trees per hectare. An average of four colonies fall within a 500 m wide transect set up along a path in each plantation block. The sampling was performed according to [7]. Figure 2 shows the random sample point collection used. Five focal scan sampling points were set up in oil palm blocks covering an average 0.1 ha/ 1000 m2 surface per plot. The data collection began from any of the four palm blocks extremities towards the central territory location or vice-versa following the methodology of [32]. Each colony was identified and recorded for the total number of nests, occupied palms, average growth period (i.e. incipient-immature, pre-matured and mature) and average occupied area (m2). The ground dwelling major workers’ average distance from base trunk was recorded for both individuals on fixed position or active trails away from palm base trunks. Quadrants measuring 1sq. m were used on the ground as a grid marker; however, using a long open reel agricultural measuring tape (100 m) was more appropriate. The estimation of the average occupied land area of each plantation was measured as follows: highest value of occupied land surface in the sample (total number of occupied palm trees x 10-meter distance approximates between each palms) x total number of sampled colonies (Supplementary material, Table S3).
To avoid overlapping on distinctive colonies, the antagonism test was performed (hostile behaviors): (i) the mandibles open toward the intruder, with or without elevated abdomen, (ii) chasing acts, (iii) any part of the trespasser’s body seized in the maxillae of one or more resident ants in act of biting, and finally one or more resident ants with its body curled around the invader while biting to the death as in wrestling. A small number of ants were captured from one tree and were released on a neighboring tree. Ants from the same colony will pass and forage, communicating with each other spontaneously between released and resident ants. To monitor distinctive colonies of O. smaragdina within a plantation, the data was collected at different blocks either from a far distance or separated by human made obstacles.
The experimental plan is presenting a portion of palm blocks used for the study of nesting occupation patterns.
2.4. Brood and barrack nests discrimination
The nests position was investigated by visual discrimination (Figure 3 (a-d). To discriminate between brood and barrack nests, observations of the distinct type of nests was carried out in selected plantations by random palm trees sampling. Upon noticing systematically different nests located at high and low position above the ground, we captured 42 brood and 10 barracks nests from identified matured Oecophylla colonies (See supplementary material, Tables S5 and S6). The sampling location were at UM, Felcra from 8-14 August 2019, Felda Gunung Besout (28/11-15/12/2020; 15/6-27/6/2022), MPOB Teluk Intan (16/12 – 19/12/2020 and 10/01 – 15/01/2021) and Felda Keratong (3/04 -14/05/2022). First, we sought for nests in the daytime and tagged the tree fronds where those nests are selected for further collection. Upon finding a nest, we recorded the height from the nest's ground to the nearest meter unit. We collected those nests at night (2300 - 0400 hours) as the genus O. smaragdina is known to be diurnal [33]. The tree fronds where the nests rested were slowly bent using a long scythe. By using scissors, the exact nest-portions of the frond were cut and immediately put into a sealed thick plastic bag. The ants were neutralised by pyrethroid knock down and brought back to the laboratory. Careful collection permitted to keep nests integrity. Each nest's volume was measured by fitting the nests to an ellipsoid form and measuring the principal axes to the nearest centimetre. The number of leaflets that consist of the nest architecture, i.e., leaflets attached by larval silk, was also counted (Supplementary material, Tables S4 and S5).
Incipient colony with first brood nests under construction (a.) at highest position (red circles) newly occupying territory of invasive bagworms M. plana defoliating palm trees (red circle). New large brood nest located at high frond position from ground (b.). New barrack type of nests (3) at lower palm frond level (red arrows) (c.) showing the ground level reach is at close distance. Below is the enlarge photo Figure 3 a named as Figure 3 d showing a new brood nest located at higher position from ground at the top of palm frond canopies (red arrow), occupying that infested palm (red right bracket).
In the laboratory, each nest was carefully disentangled and number of ants based on its age [i.e. adults (major workers, minor workers, and alates of each sex), pupae (worker, male, and queen pupae), and larvae] was directly counted. Since direct counting of eggs was not feasible without a stereomicroscope, egg clusters and their approximated size (to the nearest 0.5 cm in diameter) were determined.
2.5. Nests distribution patterns in relation to maturity status
Preliminary studies to estimate age were conducted from January 2014 to end in 2016 by monitoring nine incipient colonies in UM plantations. By visual check it is possible to estimate the average prime of life status of a colony. The nest distribution pattern of O. smaragdina was investigated in relation to their age status; i.e. Matured (ant richness level 4), pre-mature (ant richness level 3), immature (ant richness level 2), and incipient colonies (ant richness level 1) are arbitrary index designing the four growth stage of colony life development. The following description given in Supplementary material, Tables S3 and S4 identify the status of incipient, immature, pre-mature and mature colonies.
These definitions are the prelude to describe the growth stage of development of a colony. It is necessary to verify if each stage corresponding to an age calculated in months, is associated to the number of both occupied tree and nest abundance fluctuations. In order to verify the species average age parameters in relation to the total nest and occupied palm trees per colony, the above sampling dataset was again repeated for validation by 2018. From early January 2018 to the end of 2022, the nests and occupied palms dynamic fluctuation of nine tagged incipient colonies were monitored on every three months frequency basis with their age noted in months (plantations in Sungai Buloh, Selangor state). From earlier colony foundation (incipient) up to maturity level, every six months during 1.5 year and bimonthly up to subsequent 2 years, the first emergence of sexual forms was monitored in several selected plantations (using a sharp surgical blade to slightly dissect brood nests on short palm trees of 3 m). To avoid bites nuisance, brood nests were dissected from 1 m high occupied plants nearby (few meters) each sampled colonies from the same palm blocks. Difficulties of maintaining records arise once the Oecophylla ants shift to new nests abandoning earlier broken ones due to leaves deterioration or fronds pruning. Once any highly positioned occupied fronds is bent down for checking brood nests content, it does not come back to the initial higher position which may provoke the nests to be vacated. To compensate for this shortcoming, monitoring on shorter various occupied plants helped resolve this. The upper first third of frond occupied nests should be used, by holding the tips of leaflets, for such sampling to avoid ant bites.
3. Statistical analysis
3.1. Discriminant analysis
Ref. [34] method was applied to identify the discriminant factor between brood and barrack nests. Data was preprocessed and subjected to outlier removal by using the box and whisker plot method. The data normality was tested using the Shapiro-Wilk test (See Supplementary material, Table S4), following [35]. Then the dataset was transformed and standardized. To test for differences between nest types, an ANOVA test and Tukey's test were fitted for the individual variable.
Discriminant analysis (DA) was employed to differentiate the barrack and brood nests. The establishment of the DA model was executed at α = 0.05. The DA was performed on the training and validation datasets to identify the significant variables that contribute to the discrimination of the nests. The DA model was validated by partial least square discriminant analysis (PLS-DA). The correct classification by the PLS-DA model was assessed using the training and validation datasets. Then, a list of variable importance in the projection (VIPs) was compared to the significant variables determined by the DA model (Supplementary material, Table S7).
3.2. Colonies age estimation
Outline of the colonies age estimate applied using statistical analysis based on [36]: Obtain data on the age structure of the colony: This information was collected through census data from incipient stage growth development (1 occupied palm with new egg laying founding queen having a complete sealed nest means already 3 months old at least): 9 colonies total. The experimented was conducted in Ladang Sungai Buloh. In the meantime, other known colonies’ age were sampled (18) by recording the total nest and occupied palm tree per each colony. The plantation management had recorded the time of first occupation history (only error by a few months). So the data is strong. The mathematical models is used to estimate the age of the colony: the Poisson Probability model as explained in methodology. Validate the model: The model was validated by comparing the estimated age structure with the observed recorded and known precise age structure of the colony.
For age estimation, data was pre-processed and subjected to outlier removal by using the Tukey Fence method (K=1.5). Then data normality was tested using the Shapiro-Wilk test. We computed the Pearson product moment correlation coefficients between the total numbers of nests per colony versus the total number of occupied palm-trees and age estimated in months for each colony. Subsequently, we analyzed the correlation of the total number of occupied palm-trees per colony versus the estimated age in months. The Pearson correlation range strength was validated according to [37] and [38]. We verify the probability of predicting the average colonies’ age in months in function of its nest abundance and number of occupied palms.
For this evaluation, we sampled 18 colonies from the selected plantations in which it was known the age in months of nest and occupied palms, this information was given by plantation managers. We added the 9 monitored incipient colonies to have a total of 27 colonies for this analysis (See raw dataset Excel tables’ attachment). Based on the study of [39], we check which lambda (λ) setting of the Poisson Probability Density Function produced the best range to detect the most acceptable prediction of the maximum total number of nests per colony in relation to their age in months.
Frequency distribution of occupied tree-palms per colony was fitted to the negative binomial and Poisson distribution in order to determine whether the temporal distribution of matured and incipient nests followed an aggregated or random pattern. Then the number of nests occupying palms per sampling unit was summarized. The K parameter was used to estimate the temporal factor associated to the nests and palms distribution level (i.e., it gives an indication of colonies approximate age in months), for this we obtain as the best estimate for K [40]. The parameter is estimated with successive iterations and the fits of the negative binomial and Poisson models were tested with the Pearson χ2 statistic [40]. Finally, the Negative binomial and Poisson distributions were fitted to the observed frequency of nests in the 1st and 49th month and the distribution patterns at the two times were compared. The correlations between the total abundance of old and incipient nests occupying palms-trees per colony, as well as nests from different areas with different ages, were analysed using Spearman’s correlation.
In addition, the colony age estimation takes into consideration the study of [41], which suggested that new nests’ construction duration is 14 days. For this purpose, six matured colonies per each three plantations (identified by rapid visual nesting method) were randomly selected. The colonies were examined under the Poisson model for the probability of predicting their average lifespan expressed in months in relation to each colony total nests occupying palms (Supplementary material, raw dataset Excel tables). The Asian weaver ant colonies have an average temporal maximum period to reach the optimal growth development stage of full maturity: there is no more extra occupied palm tree and nest construction considering that the population dynamic attained its peak. The MATLAB software R2021b tool [42] was applied to estimate the maximum age of a colony expressed in months and corresponding to the optimum growth development stage period corresponding to an average total number of nests per colony that is reliable, acceptable to determine a colony average age.
4. Results
4.1. Spatial distribution of Asian weaver ant colonies
The surveys recorded that the Asian weaver ants are widely distributed throughout Malaysia. The results show that sites record of their areas of occurrences, existing throughout the year, Oecophylla colonies prevalence. Colonies of O. smaragdina develop a systematic procedure of occupation. The weaver ants established absolute control of palm trees by colonizing under a three-dimensional pattern of colony occupation (canopies, trunk-palm fronds and ground, central, secondary and tertiary territories respectively). A majority of studied areas/estates confirmed the presence of Oecophylla colonies. It is important to notice that the large palm trees and imposing canopies type of phenology appear to be a preferred habitat of Oecophylla ants. Some plantations management is practicing a combination of mix cropping with a variety of the Oecophylla colonies host plants (diverse plants species) to provide refuge. Moreover, the common host plants harboring weaver ant colonies are presented (Supplementary material, Table S5). Weaver ants demonstrated polydomous nesting behaviors when occupying associated fruit trees with a height ≥ 3 m, and polydomous/monodomous pattern for shorter plants ≤ 2 m high. However, colonies were strictly monodomous when occupying shorter plants inferior or equal to an average ≤ 1 m (Supplementary material, Table S8). However, a survey conducted in Felda Keratong Pahang exposed a negative presence of the Asian weaver ant colonies from large plantation blocs (Supplementary material, Table S1). Only few isolated palms were occupied with matured colonies ant richness level 4 in one spot closer to residential areas. Some plantations had a poor O. smaragdina colonies presence (Felda Gunung Besout Perak). Some plantations management is practicing a combination of mix cropping with a variety of host plants (diverse plants species) to provide refuge within and beside the plantations. Moreover, the common associated plants harboring weaver ant colonies are presented (Supplementary material, Table S5). Odontoponera denticulata ant species presence was consequent with many nest holes recorded in the vicinity of occupied O. smaragdina territories (Figure 4 A-B). Such species are potentially strongly antagonistic to weave ant, since the collapse of O. smaragdina colonies was high in each area whenever colonies of O. denticulata and Solenopsis geminata (Figure 4 C) presence was positive (N = 50).
4.2. Colonies nests distribution patterns
From the overall occupancy, it is revealed that weaver ants do not have any particular preference for palm trees based on age and height. Sight occupation depends largely on colonies’ safety and survival factor coupled with their association of arboreal fruit host tree species (Supplementary material, Table S5). While O. smaragdina predilection for dwelling in taller palms is unmatched, their absence is still rare in shorter palms. In one case only circumstantially due to the black ants D. thoracicus presence [7], then weaver ants manage to take dominance status the following years (Teluk Intan, Perak plantations) [11]. Conversely, in a southern plantation in Johor, their abundance was higher in a younger plantation with the first fresh fruit bunches yet to emerge soon.
Of the 1599 recorded palms ranging from 2.5 to 15 years and 3 m to 15 m high, Oecophylla colonies occupied the trees irrespectively of their age and height. It was observed that incipient colonies were established within close distance from each other. All newly constructed nests comprise only young leaflets while old ones are being abandoned gradually. A nest occupation can last up to more than 6 months, depending on the quality of freshness of palm fronds leaflets.
Between matured and pre-matured colonies, there is a threefold superiority difference in mean nest numbers. The distribution of nests consisting of a dominant sequence facilitates the rapid detection of matured colonies in the field (Supplementary material, Table S4). All observed matured colonies in this sampling fulfilled the definition (Table 1).
There was a strong positive correlation for both nest and palm numbers in association with the age of each colony as both Pearson correlation coefficients gave significant large positive relationship with r(7)= 0.987, p <0.001, covariance=165.5 (outliers: 101, 63); r(7)= 0.846, p = 0.004, covariance = 11.4; for total nests and occupied palms respectively (Figure 5).
4.3. Discrimination of brood and barrack nests
All collected nests from selected plantations presented close similarity among the main factor differentiating between nest categories. There were significant differences between brood and barracks nests (Table 2; Supplementary material, Table S6; supplementary material Figure S2). The discriminant analysis model had identified seven variables that significantly contributed to the discrimination of the barrack and brood nests (Supplementary material, Table S6). Based on the F-statistics, the ranking of significant variables was as follows: Number of minor workers > height from the ground > number of worker pupae > number of larvae > number of total workers > number of leaflets > number of winged queen (Supplementary material, Table S6).
Wilks' lambda for the DA model (p < 0.0001) showed that the brood and barrack nests were significantly different from each other. This finding is confirmed by Fisher distance (p < 0.0001). The training and validation datasets had a 100% correct classification of the nest type. The DA's ability to differentiate the brood and barrack nests was evident with the separation of both groups (See Figure 6 and supplementary material, Figure S3). The PLS-DA model corroborated the ability of the DA model to discriminate between brood and barrack nests (Table 3; Supplementary material, Table S6). Based on variable importance in the projection (VIP) value, the ranking of significant variables was as follows: Number of minor workers > height from the ground > number of worker pupae > number of larvae > number of total workers > number of major workers > number of leaflets. The DA results supported the ANOVA finding (Table 2) since these variables showed a significant difference in value between the barrack and brood nests. Moreover, the DA had successfully identified these variables' ranking, which the ANOVA test cannot provide. The DA model also found that the variables associated with nest characteristics played insignificant roles in discriminating the barrack and brood nests except for the height from the ground. Since the ground's height is the most practical approach to determine the nest type compared to the six significant variables, the nest which height is 8.0 ± 1.3 m is identified as a brood nest, as shown in supplementary material Table S6.
Contrary to the DA model, the PLSDA model did not identify the number of winged queens as significant variables. The PLS-DA had 96.15% correct classification of brood and barrack nests when applying the training and validation datasets, which was lower than the DA model. Nevertheless, the Wilks' lambda for the PLS-DA model (p < 0.0001) showed that the brood and barrack nests were significantly different from each other. The Fisher distance (p < 0.0001) confirmed this result. Hence, the DA model was able to differentiate the brood and barrack nests (Table 3).
The DA model also found that the variables associated with nest characteristics played insignificant roles in discriminating the brood and barrack nests except for the height from the ground. Since the ground's height is the most practical approach to determine the nest type compared to the six significant variables, the nest which height is 8.0 ± 1.3 m is identified as a brood nest, as shown in Table 2 and supplementary Table S6.
4.4. Age estimation validation by Pearson correlation coefficient analysis
All residual distributions followed a normal distribution. Results of the Pearson correlation indicated that there is a significant large positive relationship between total nests (two outliers, 85, 77 representing 11.11% of the observations) and total palms (no outliers) (r (16) = 0.831, p < .001) (Supplementary material, Table S4). There is a significant large positive relationship between number of nests and the occupied palm trees (no outliers), and strong positive correlation (r (16) = .991, p < .001), (r (16) = .845, p < .001) with the estimated colonies age (2 outliers, 53, 49), respectively. In the second analysis, there was one potential outlier in the pair, which is 5.56% of the observations ([34,48]), while in the third the pair did not contain outliers (Supplementary material, Table S8). All Pearson correlation coefficients had a strong correlation between number of nests and occupied palms with each recorded colony age in months (14 colonies) (Supplementary material, Table S8 and Figure S4 a-c).
4.5. Colonies age estimation
The results for λ = 25, λ = 40 and λ = 50 are not informative enough to detect any association between the average nest abundance and colony age (Figure 7 a-c). The analysis for λ = 60 is expressing a relation between the total number of nests per colony and the expected age in months for each colony. This corresponds to a Poisson ratio of 0.05. The Poisson distribution detected a prediction probability for each colony average age in function of the total number of nests per colony up to the age of 4-5 years old. It is better to include Lambda (x-axis) in an interval [50,60] to obtain the maximum Poisson probability density function (PPDF) (this is the y-axis, 0.05, 0.055) because the error is a very small value (5/1000) (Figure 7 a-d). Hence the PPDF is valid to predict the association between the total numbers of nests per colony in function of the age in months up to a maximum of 5 years (See Figure 7 d; supplementary material, Figure S4 a-c).
Figure 7 a-d show the Poisson probability density function at lambda 25 (Figure 7 a), lambda 45 (Figure 7 b), lambda 50 (Figure 7 c) and lambda 60 (Figure 7 d). The results for lambda 60 is expressing a relation between the total number of nest per colony and the expected age in months for each colony. This corresponds to a Poisson ratio of 0.05. The Poisson distribution detected a prediction probability for each colony average age in function of the total number of nests per colony up to the age of 4-5 years old.
5. Discussion
5.1. Distribution of Oecophylla smaragdina colonies
This paper engaged in this investigation to widen the latitude of information and knowledge contributing to understand better the versatile functions of this generalist predator. Previous studies mentioned the omnipresence of Oecophylla colonies in oil palm plantations but without defining the extension of their distribution nationwide [28]. O. smaragdina established prevalence by its abundance and rapid spatial expansion and colonization in the Australian, Brunei and Sulawesi forest canopies [43,44] and oil palm trees of Southeast Asia, as it rarely was challenged by other ant species [45]. This study managed to offer a detailed mapping of the Asian weaver ants in Malaysia, establishing the first investigation ever. The study findings demonstrate Oecophylla colonies wide accessibility. However, there was no record from the forest mangroves or oil palm plantation in Rembau, Negeri Sembilan [46]. The Asian weaver ants have not been well documented in this major Southeast Asia industry. Our findings corroborate well with other studies in terms of the prevalence and colonies abundance of Oecophylla genus in the tropics [2,18]. Palm trees age, location and the nature of the cultivated soil are not influential factors on the Asian weaver ant habitat choices. It is rather the density of palm trees population, offering a good sunny exposure angle to colonies that would become determinant under thicker and large canopies [47]. Ref. [48] suggested that such phenology factor of host trees influence Oecophylla colonies preference choices. Ref. [9] expressed the concept of a new matured colony with few trees occupation along some dozen of nests. This is showing much less population abundance (based on the higher nests number per colony) than Oecophylla colonies found in oil palms. The conditions of colonization of weaver ants in large monoculture plantations are different from Australian orange, cashew nuts and citrus orchard tree phenology. This idea of an association between tree crops phenology and colony of O. smaragdina was expressed by several authors, which provided better shelters with larger canopies [47] and easier folding softer large leaves, a mixture of a suitable ambient microclimate and sunny exposition to nests [19,49]. By analogy of this concept fruiting trees producing extra floral nectar and honeydew items, such as rambutan, mangosteen and mango are reported to promote Oecophylla ant colonies distribution and expansion which is similar with present study findings [50,51,52]. In addition, weaver ants are also known to inhabit wild or domesticated fruit trees such as citrus [45] with an extensive list of plants’ family species provided by [19]. These plants are promoting the expansion of Oecophylla colonies in tropical agrosystems as they establish a buffer zone to support Oecophylla colonies expansion, hence enhancing ant long-term conservation [19,31].
5.2. Colonies nests distribution patterns
The distribution of nests was similar to other studies in various fruit plants [53] such as in mango [54] in coconut [55] in Citrus spp [31]. They are polydomous in nature [12] and a single tree may harbour many nests [7,11]. Ref. [13], demonstrated that in mango orchards, a single colony of O. smaragdina managed to occupy 2 to 30 trees with 25 to 153 nests. A colony by its nests network expansion could occupy an average surface of 1600 m2, comprising 17 major trees [56] with over hundreds of nests [1] which is in accordance with the findings of this study. Nest distribution may be influenced by the proximity of palm trees and interconnected palm canopies provide bridges for colony expansion. Nevertheless, the weaver ants exhibited the obvious trend of ground foraging activities [31,57], suggesting that the existence of unoccupied palm trees is not depressing their expansion potential. Further in already occupied palm trees blocks, the nests’ absence in these empty trees are representing the border zone between these absolute territorial Asian weaver ants’ colonies [1]. Each colony of the weaver ants featured a deliberate control with systematic no “ants land” as separation zones. Chemical cues are left by ants as a demarcation between colonies to strongly mark their boundaries [1]. Their territorial behavior towards trespassers which comprises all other ant species and arthropods. Any established O. smaragdina colony in the oil palm plantations represents a vast community of dozens of thousands of workers [58]. The Asian weaver ants demonstrated a three-dimensional pattern of enlargement resulting in a mosaic of colonies through the oil palm landscape with its surroundings [45]. The ants are able to expand faster and occupy a larger portion of the oil palm including all the surrounding landscape (various trees, plants within the plantations and available nearby forestry corridors) taking their presence with a far distance in depth. Weaver ants are establishing their new habitats with a high level of cooperation between nest mates. In general, it was easy to obtain the exact number of nests per palm tree and per colony. Each colony establishes a large security perimeter around their territory with various layers of successive columns used as filtration defensive levels to expel any strangers, intruders into their residency areas [19,59]. On the other hand, Asian weaver ants were seen multiplying their nests with individuals spreading further from each palm tree occupied in both ground and canopies as reported by [32]. There was no obvious competition from any other ant ground nesting species or arboreal species within occupied oil palm blocks which coincide with [45] study on O. smaragdina dominance in oil palm plantations. However, any nearby insect or neighboring Oecophylla ants passing by their territory was fiercely and promptly eliminated with the help of colony mates’ cooperation. Occupancy patterns are specifically territorial with compact groups and individuals spreading from nests in palm canopies, trunk trails to the ground with an average radius of 5 m around each tree with few layers of weaver ant defensive rows (n > 1000 palm trees) [19]. The presence and intense foraging activities of the ants on large soil areas is conspicuous and their repeated attacks once anyone passed their residence zones is systematic. On the ground of base trunks of each occupied palm tree, the sight of layers of ants’ successive columns was striking. Since many palm trees in a particular block and plantations areas were not occupied by extensive nesting networks, the control measures of the antagonistic presence of D. thoracicus (and invasive ants S. geminata) is compulsory. The black ants are reported to impair significantly or stop the promotion of Oecophylla colonies distribution and expansion for unknown reasons [9,60], hence it is recommended to destroy any antagonistic ant species if any control treatment for the invasive bagworms M. plana is to be applied. Ref. [61] made the same recommendations for control measures against antagonistic ants Pheidole megacephala and Anoplolepis longipes in T. cacao plantations. On this note, Odontoponera denticulata and O. smaragdina can be co-dominant [62] and further investigation to verify any adverse antagonistic effect is needed. It is necessary to conduct further study on the seasonal colonies nest territorial dynamic [32] and abundance [63], in particular its interaction with rainfall interception [64].
5.3. Difference between brood and barrack nests
O. smaragdina preference to position their brood nests at the top of the canopies under direct sunlight, has demonstrated a predilection to solar radiation exposure [65]. These findings concur with those found by [45,66]. Functionally, Oecophylla colonies brood nests location at top of palm canopies having higher insolation is verified in the literature with a tendency towards sunshiny exposure, because they are being unable to capitalise on the heat radiance of the soil [18]. Beside that aspect, another important point touches to the territorial defensive strategy of O. smaragdina [28]. The characteristic arrangement observed between brood and barrack nests in the palm trees is seen as a pure safety and survival strategy [1]. By putting brood nests at higher height from the ground, it is to prevent the colony overall individuals exposure to danger related to the maintenance activity during fronds pruning. The barrack nests are positioned at much lower ground level with only bigger size major workers securing the canopies territory and representing the first line of defence [7]. The barrack nests are functioning as a different level of defence line for each occupied palm tree to secure the colony from any intruders [18,67,68]. There might be a possible relation between the height impacts of palm trees preferential occupancy by O. smaragdina determined by the mutualistic relation with honey dew tended homopterans predilection for shaded larger canopy found in taller trees as reported for O. longinoda [69].
5.4. Average colonies age status of Oecophylla smaragdina
Between each stage of colony development (incipient-immature, pre-mature and mature), there is an average evolution period which determines their population stability. There is a uniform replication of the task from an incipient colony to become an immature, pre-matured colony. It takes above two years and half to establish a matured colony fully autonomous under the leadership of their gravid mother queens [13] with three years being more reasonable and a minimum to define the maturity period and age of established stable colonies more suitable for manipulations (BC, IPM treatments) [16]. This shift phase is crucial during the ending of pre-matured growth as newly matured colonies (aged 24 months to 30 months) need one more year (36 to 42 months of age) to strengthen the colony size to optimum level [16]. The temporal factor consideration could be variable in function of the number of dealate queens initially taking part in colony foundation [20]. The cooperation of more queens to develop a colony is predictive of a significant fluctuation in the average period taken to establish a colony from embryonic stage towards a final stable juncture [70,71]. Incipient polygynous colonies reach maturity much earlier than monogynous colonies since the former may produce a far more abundant work force and broods [13,16,20]. Once the nest population increases, Oecophylla ant reaction is to shift the overcrowding of individuals to a new satellite nest to fulfil the balance threshold of an optimum acceptable repartition, thus such abundance contributing to the colonies wider distribution and expansion [72]. This might explain one of the factors of the rapid distribution expansion of the species in oil palm plantations and tropical forests. In the present study the labelling of pre-matured colonies up to the first emergence of sexual forms signifies that such colonies are still in a vulnerable stage, hence not yet suitable for usage as pest control manipulations as suggested by [16] and [73]. This is the first time that such adaptive period evolution is detected in the growth development of Oecophylla colonies. This is inferred that a newly matured colony is not yet stable until a further year of its existence. Oecophylla population is expected to reach a satisfactory level of size strength upon that extended period [16].
The founding queen body size fluctuation over years has been evaluated and shows a correlation with the average age of the colony [9]. Identifying the location of the mother queens’ colonies [13,74] is logistically unpractical with high improbability to replicate successfully the similar experiment carried out in situ [75,76,77]. Destructive sampling was avoided by visual check only, consumes less time, and is easier, hence sustainable. Incipient colonies ages one to two months correspond to the emergence of the first adults in Oecophylla spp [78]. It is rare that incipient colony’s lifespan is monitored for ten years. Only barrack nests were affected by pruning during harvesting of fresh fruit bunches (FFB) due to its position at lower height (statistically strongly significant).
Agricultural intensification simplifies and modifies the agricultural landscape, disturbing the ecosystem services that biodiversity provides to agroecosystems, particularly the biological control [79,80]. This is the case in neighbouring Philippines by a recent study proposing the adoption of the Asian weaver ant colonies as an efficient substitute of the harmful highly toxic chemicals in use inside cocoa plantations [81]. Biological control is offering ecosystem services free from nature to humans and a vast monoculture like the oil palm plantations in South- East Asia could have a high impact that could be affecting such ecosystem service (conservation biological control) [82]. Many of the associated host plants are flower nectar producers, like the Melastoma malabathricum whom attractant by recruiting the Asian weaver ant, induce the repellence of poorly efficient pollinators to promote more effective Xylocopa bees [83]. The Asian weaver ants in South-East Asia represents a strategic potential influential of the ecological and evolutionary paths of plant–pollinator interactions [84]. The future introduction of the Asian weaver ant might ease management burden costs, terminate highly toxic pesticides applications harmful to non-target taxa and accordingly provide environmental preservation [28].
6. Conclusion
This is the first study highlighting Oecophylla ant geographical distribution in Malaysia in the mega oil palm plantations major industry. Despite their absolute territorial behaviour, the wide availability of weaver ants, does give clues on the low cost of this species management if included for the control of invasive nuisance in areas where O. smaragdina is present. The colony’s dense population, three-dimensional occupation pattern, exhibited spreading waves featuring interconnected irregular geometrical patches shaped for each. However, some plantations were not occupied, hence the planting of additional favourite host plants mixed crops is recommended to enhance weaver ant conservation and three-dimensional distribution expansion. While O. smaragdina is a characteristic polydomous species, it is the first case reporting monodomous nesting behaviors on short plants. Odontoponera denticulata was identified, for the first time ever, as a possible antagonistic species disrupting the smooth Asian weaver ant colonies’ territorial dynamic. Brood and barrack nest visual distinction is possible by the height to the ground factor, corresponding to higher and lower location respectively in the palm canopies. It is also the first report establishing a strong correlation between the numbers of nests, occupied palm trees with the estimated colonies age up to 60 months by Poisson probability density function. Matured stable colonies (average 3 years) of O. smaragdina demonstrated occupation stability above 10 years period by resilience and altruism. This is the first study estimating the optimal population superiority average reaching a peak by five years of age in relation to the nest abundance. The combined method of visual fast identification of the colonies maturity status with the brood and barrack nests distinction is practical, innovative and sustainable for future potential biological control treatments.
The possibility of having the introduction of multiple queens in O. smaragdina could be advantageous by establishing a biological control efficient ecosystem services. This could provide a complementary method from the ongoing excessive toxic chemicals applications prevailing in the oil palm industry pest management. The Asian weaver ant survival might be vulnerable in the long term from such practices. Constituting sub-colonies for relocation might maintain the integrity of the original colony. An evaluation on the polygyny status of O. smaragdina species in South East-Asia is necessary. It might lend more clues on the factors enabling colonies to enhance their abundance, developing their expansion by colonising more territories and stabilising their population dynamic faster. Further studies about the effect of O. smaragdina over natural enemies in oil palm crop should be investigated.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Exélis Moïse Pierre proposed the hypothesis and the approach to test it experimentally, conceptualized the project, conceived and designed the experiments, performed all experiments, analyzed the data, prepared tables, and figures, wrote original draft, edited, authored or reviewed drafts of the paper. Rosli Ramli supervised the project, edited, authored or reviewed drafts of the paper. Azarae Idris supervised the project, performed experiments, authored or reviewed drafts of the paper. Rabha W. Ibrahim performed experiments, analyzed the data, prepared figures, authored or reviewed drafts of the paper. Gemma Clemente-Orta edited, authored or reviewed drafts of the paper. All authors approved the final version of the paper.
Funding
This research was funded by Universiti Malaya through the project no. PS320-2010B and PV078-2011B (awarded to EMP), and (Sustainability & Biodiversity) UMRG project no. RG187/12SUS, AFR (Frontier Science) project no. RP016A-14-AFR (to EMP & AHI). Another major funding was also secured under a French-European Union complete scholarship-grant through the local government authority of “Collectivité Territoriale de la Martinique-CTM” (all to EMP). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Data Availability Statement
All data generated or analysed during this study are included in this published article [and its supplementary information files under Excel format named as Supplementary dataset].
Acknowledgments
Special thanks to the Malaysian Palm Oil Board (MPOB), Felda Malaysia and Universiti Malaya Sdn. Bhd for granting access permission to its facilities, plantations in Malaysia. Prof Emeritus Dr. Seiki Yamane identified the ant species.
Conflicts of Interest
The authors declare there are no competing interests.
References
- Hölldobler B, Wilson EO. The ants. Harvard University Press; 1990.
- Wetterer, JK. Geographic distribution of the weaver ant Oecophylla smaragdina. Asian myrmecology. 2017 Jan 1; 9:1-2.
- Peng RK, Christian K. The weaver ant, Oecophylla smaragdina (Hymenoptera: Formicidae), an effective biological control agent of the red-banded thrips, Selenothrips rubrocinctus (Thysanoptera: Thripidae) in mango crops in the Northern Territory of Australia. International journal of pest management. 2004 Apr 1; 50(2):107-14. [CrossRef]
- Peng, R.K.; Christian, K. Effective control of Jarvis's fruit fly,Bactrocera jarvisi(Diptera: Tephritidae), by the weaver ant,Oecophylla smaragdina(Hymenoptera: Formicidae), in mango orchards in the Northern Territory of Australia. Int. J. Pest Manag. 2006, 52, 275–282. [Google Scholar] [CrossRef]
- Salim, H.; Rawi, C.S.M.; Ahmad, A.H.; Al-Shami, S.A. Efficacy of Insecticide and Bioinsecticide Ground Sprays to Control Metisa plana Walker (Lepidoptera: Psychidae) in Oil Palm Plantations, Malaysia. . 2015, 26, 73–83. [Google Scholar] [PubMed]
- Kamarudin N, Ali SR, Masri MM, Ahmad MN, Manan CA, Kamarudin N. Controlling Metisa plana Walker (Lepidoptera: Psychidae) outbreak using Bacillus thuringiensis at an oil palm plantation in Slim River, Perak, Malaysia. Journal of Oil Palm Research. 2017 Mar 1; 29(1):47-54. [CrossRef]
- Exélis MP, Idris AH. Studies on the predatory activities of Oecophylla smaragdina (Hymenoptera: Formicidae) on Pteroma pendula (Lepidoptera: Psychidae) in oil palm plantations in Teluk Intan, Perak (Malaysia). Asian myrmecology. 2013 Jan 1; 5(1):163-76.
- Peng, R.; Christian, K.; Gibb, K. The best time of day to monitor and manipulate weaver ant colonies in biological control. J. Appl. Èntomol. 2011, 136, 155–160. [Google Scholar] [CrossRef]
- Peng RK, Christian K, Gibb K. Implementing Ant Technology in Commercial Cashew Plantations and Continuation of Transplanted Green Ant Colony Monitoring: A Report for the Rural Industries Research and Development Corporation. 2004 May; 1-37.
- Binh, NT. (2010). Assessment of an integrated cashew improvement program, using weaver ants as a major component, in cashew production in Vietnam. Charles Darwin University and Institute of Agricultural Science for South Vietnam. 2010 March; (Doctoral Dissertation, Charles Darwin University), 17-135. https://ris.cdu.edu.au/ws/portalfiles/portal/44469117/Thesis_CDU_34457_Nguyen_TB. [CrossRef]
- Exélis, MP. An ecological study of Pteroma pendula (Lepidoptera: Psychidae) in oil palm plantations with emphasis on the predatory activities of Oecophylla smaragdina (Hymenoptera: Formicidae) on the bagworm/Exelis Moise Pierre. 2015 March; (Msc Dissertation, University of Malaya).
- Dejean A, Corbara B, Orivel J, Leponce M. Rainforest canopy ants: the implications of territoriality and predatory behavior. Functional Ecosystems and Communities. 2007; 1(2):105-20.
- Peng, R.; Christian, K.; Gibb, K. How many queens are there in mature colonies of the green ant, Oecophylla smaragdina (Fabricius)? Aust. J. Èntomol. 1998, 37, 249–253. [Google Scholar] [CrossRef]
- Chapuisat, M.; Keller, L. Division of labour influences the rate of ageing in weaver ant workers. 269. [CrossRef]
- Kramer, B.H.; Schaible, R. Life Span Evolution in Eusocial Workers—A Theoretical Approach to Understanding the Effects of Extrinsic Mortality in a Hierarchical System. PLOS ONE 2013, 8, e61813. [Google Scholar] [CrossRef] [PubMed]
- Peng RK, Nielsen MG, Offenberg J, Birkmose D. Utilisation of multiple queens and pupae transplantation to boost early colony growth of weaver ants Oecophylla smaragdina. Asian Myrmecology. 2013; 5(1):177-84.
- Blüthgen N, Fiedler K. Interactions between weaver ants Oecophylla smaragdina, homopterans, trees and lianas in an Australian rain forest canopy. Journal of animal ecology. 2002 Sep; 71(5):793-801. [CrossRef]
- Crozier RH, Newey PS, Schluens EA, Robson SK. A masterpiece of evolution–Oecophylla weaver ants (Hymenoptera: Formicidae). Myrmecological News. 2010 May; 13(5):57-71.
- Lim, GT. Enhancing the weaver ant, Oecophylla smaragdina (Hymenoptera: Formicidae), for biological control of a shoot borer, Hypsipyla robusta (Lepidoptera: Pyralidae), in Malaysian mahogany plantations. 2007. (Doctoral dissertation, Virginia Tech).
- Offenberg, J.; Peng, R.; Nielsen, M.G. Development rate and brood production in haplo- and pleometrotic colonies of Oecophylla smaragdina. Insectes Sociaux 2012, 59, 307–311. [Google Scholar] [CrossRef]
- Peeters, C.; Andersen, A.N. Cooperation Between Dealate Queens During Colony Foundation in the Green Tree Ant, Oecophylla Smaragdina. Psyche: A J. Èntomol. 1989, 96, 39–44. [Google Scholar] [CrossRef]
- Rissing, S.W.; Pollock, G.B. An experimental analysis of pleometrotic advantage in the desert seed-harvester antMessor pergandei (Hymenoptera; Formicidae). Insectes Sociaux 1991, 38, 205–211. [Google Scholar] [CrossRef]
- Sommer, K.; Hölldobler, B. Colony founding by queen association and determinants of reduction in queen number in the antLasius niger. Anim. Behav. 1995, 50, 287–294. [Google Scholar] [CrossRef]
- Kronauer, D.J.C.; Miller, D.J.; Hölldobler, B. Genetic evidence for intra– and interspecific slavery in honey ants (genusMyrmecocystus). 270. [CrossRef]
- Tschinkel, W.R. Brood raiding and the population dynamics of founding and incipient colonies of the fire ant, Solenopsis invicta. Ecol. Èntomol. 1992, 17, 179–188. [Google Scholar] [CrossRef]
- Wood BJ, Kamarudin N. A review of developments in integrated pest management (IPM) of bagworm (Lepidoptera: Psychidae) infestation in oil palms in Malaysia. Journal of Oil Palm Research. 2019 Dec 10; 31(4):529-39.
- Wood BJ, Kamarudin N. Bagworm (Lepidoptera: Psychidae) Infestation in the centennial of the Malaysian oil palm industry—A review of causes and control. Journal of Oil Palm Research. 2019 Sep 1; 31(3):364-80. [CrossRef]
- Exélis, M.P.; Ramli, R.; Ibrahim, R.W.; Idris, A.H. Foraging Behaviour and Population Dynamics of Asian Weaver Ants: Assessing Its Potential as Biological Control Agent of the Invasive Bagworms Metisa plana (Lepidoptera: Psychidae) in Oil Palm Plantations. Sustainability 2022, 15, 780. [Google Scholar] [CrossRef]
- Abubakar, A.; Ishak, M.Y.; Makmom, A.A. Nexus between climate change and oil palm production in Malaysia: a review. Environ. Monit. Assess. 2022, 194, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Lokkers, C. Colony dynamics of the green tree ant (Oecophylla smaragdina Fab.) in a seasonal tropical climate. 1990. (Doctoral dissertation, James Cook University).
- Van Mele P, Cuc NT. Evolution and status of Oecophylla smaragdina (Fabricius) as a pest control agent in citrus in the Mekong Delta, Vietnam. International Journal of Pest Management. 2000 Jan 1; 46(4):295-301. [CrossRef]
- Rastogi, N. Seasonal pattern in the territorial dynamics of the arboreal ant Oecophylla smaragdina (Hymenoptera: Formicidae). Journal-Bombay Natural History Society. 2007 Jan; 104(1):30.
- Jander, R.; Jander, U. The Light and Magnetic Compass of the Weaver Ant, Oecophylla smaragdina (Hymenoptera: Formicidae). Ethology 1998, 104, 743–758. [Google Scholar] [CrossRef]
- Samsudin, M.S.; Azid, A.; Khalit, S.I.; Sani, M.S.A.; Lananan, F. Comparison of prediction model using spatial discriminant analysis for marine water quality index in mangrove estuarine zones. Mar. Pollut. Bull. 2019, 141, 472–481. [Google Scholar] [CrossRef]
- Samsudin, M.S.; Azid, A.; Khalit, S.I.; Sani, M.S.A.; Lananan, F. Comparison of prediction model using spatial discriminant analysis for marine water quality index in mangrove estuarine zones. Mar. Pollut. Bull. 2019, 141, 472–481. [Google Scholar] [CrossRef] [PubMed]
- McCrea RS, Morgan BJ. Analysis of capture-recapture data. CRC Press; 2014 Aug 1.
- Mukaka, M.M. Statistics corner: A guide to appropriate use of correlation coefficient in medical research. . 2012, 24, 69–71. [Google Scholar]
- Akoglu, H. User's guide to correlation coefficients. Turkish journal of emergency medicine. 2018 Sep 1; 18(3):91-3. [CrossRef]
- Ludwig JA, Reynolds JF. Statistical ecology: a primer in methods and computing. John Wiley & Sons; 1988. 18 May.
- Gowda, D.M. PROBABILITY MODELS TO STUDY THE SPATIAL PATTERN, ABUNDANCE AND DIVERSITY OF TREE SPECIES. 2011. [Google Scholar] [CrossRef]
- Pimid M, Hassan A, Tahir NA, Thevan K. Colony structure of the weaver ant, Oecophylla smaragdina (Fabricius)(Hymenoptera: Formicidae). Sociobiology. 2012; 59(1):1-0. [CrossRef]
- MATLAB and Statistics Toolbox. The Mathworks Inc. Natick, Massachusetts, USA. 2021.
- Blüthgen, N.; Stork, N.E. Ant mosaics in a tropical rainforest in Australia and elsewhere: A critical review. Austral Ecol. 2007, 32, 93–104. [Google Scholar] [CrossRef]
- Davidson, D.W.; Lessard, J.; Bernau, C.R.; Cook, S.C. The Tropical Ant Mosaic in a Primary Bornean Rain Forest. Biotropica 2007, 39, 468–475. [Google Scholar] [CrossRef]
- Pfeiffer, M.; Tuck, H.C.; Lay, T.C. Exploring arboreal ant community composition and co-occurrence patterns in plantations of oil palm Elaeis guineensis in Borneo and Peninsular Malaysia. Ecography 2008, 31, 21–32. [Google Scholar] [CrossRef]
- Hashim NR, Wan Jusoh WFA, Mohd Nasir MNS. Short Communication. Ant diversity in a Peninsular Malaysian mangrove forest and oil palm plantation. Asian Myrmecology. 2010; Volume 3, 5–8.
- Van Wijngaarden PM, Van Kessel M, Van Huis A. Oecophylla longinoda (Hymenoptera: Formicidae) as a biological control agent for cocoa capsids (Hemiptera: Miridae). In Proceedings of the Netherlands Entomological Society Meeting. 2007; (Vol. 18, 21-30).
- Nene, WA. Aspects of ecology of weaver ants (Oecophylla longinoda Latreille) (Hymenoptera: Formicidae) in Tanzania. 2016, (. Doctoral Dissertation, Sokoine University of Agriculture, Morogoro.
- Wargui R, Offenberg J, Sinzogan A, Adandonon A, Kossou D, Vayssières JF. Comparing different methods to assess weaver ant abundance in plantation trees. Asian Myrmecology. 2015; 7, 159-170.
- Dejean, A.; Djieto-Lordon, C.; Durand, J.L. Ant Mosaic in Oil Palm Plantations of the Southwest Province of Cameroon: Impact on Leaf Miner Beetle (Coleoptera: Chrysomelidae). J. Econ. Èntomol. 1997, 90, 1092–1096. [Google Scholar] [CrossRef]
- Noor Izwan A, Amiruddin BA. Diversity of ants (Hymenoptera: Formicidae) at Kuala Lompat, Krau Wildlife Reserve, Pahang, Malaysia. The Journal of Wildlife and Parks. 2014; 28, 31-39.
- Vayssières, J.; Grechi, I.; Sinzogan, A.; Ouagoussounon, I.; Todjihoundé, R.; Modjibou, S.; Tossou, J.; Adandonon, A.; Kikissagbé, C.; Tamò, M.; et al. Host plants and associated trophobionts of the weaver ant Oecophylla longinoda Latreille (Hymenoptera Formicidae) in Benin. Agric. For. Èntomol. 2021, 24, 137–151. [Google Scholar] [CrossRef]
- Fiedler, K.; Maschwitz, U. The symbiosis between the weaver ant,Oecophylla smaragdina, andAnthene emolus, an obligate myrmecophilous lycaenid butterfly. J. Nat. Hist. 1989, 23, 833–846. [Google Scholar] [CrossRef]
- Peng, R.; Christian, K. Integrated pest management in mango orchards in the Northern Territory Australia, using the weaver ant,Oecophylla smaragdina, (Hymenoptera: Formicidae) as a key element. Int. J. Pest Manag. 2005, 51, 149–155. [Google Scholar] [CrossRef]
- Way, M.; Bolton, B. Competition between ants for coconut palm nesting sites. J. Nat. Hist. 1997, 31, 439–455. [Google Scholar] [CrossRef]
- Hölldobler, B. Territories of the African Weaver Ant (Oecophylla longinoda [Latreille]); A Field Study. 51. [CrossRef]
- Offenberg, J. The use of artificial nests by weaver ants: A preliminary field observation. Asian Myrmecology. 2014 Jan 1; 6(1):6.
- Schlüns EA, Wegener BJ, Schlüns H, Azuma N, Robson SK, Crozier RH. Breeding system, colony and population structure in the weaver ant Oecophylla smaragdina. Molecular Ecology. 2009 Jan; 18(1):156-67.
- Exélis, MP, Ramli R, Idris AH, Latif SA, Yaakop S, Ibrahim RW. Relationships between weather parameters and foraging daily activity of Oecophylla smaragdina (Hymenoptera: Formicidae) in oil palm plantations: Case Study. 14 November. [CrossRef]
- Van Mele, P.; Cuc, N.T.T.; VanHuis, A. Direct and indirect influences of the weaver ant Oecophylla smaragdina on citrus farmers' pest perceptions and management practices in the Mekong Delta, Vietnam. Int. J. Pest Manag. 2002, 48, 225–232. [Google Scholar] [CrossRef]
- Way, M.J.; Khoo, K.C. Role of Ants in Pest Management. Annu. Rev. Èntomol. 1992, 37, 479–503. [Google Scholar] [CrossRef]
- Sitthicharoencha D, Chantarasawat N. Ant species diversity in the establishing area for advanced technology institute at Lai-Nan sub-district, Wiang Sa district, Nan province, Thailand. Tropical Natural History. 2006 Oct 1; 6(2):67-74.
- Olotu, MI. Effect of seasonality on abundance of African weaver ant Oecophylla longinoda (Hymenoptera: Formicidae) in cashew agro-ecosystems in Tanzania. African Journal of Agricultural Research. 2016 Apr 21; 11(16):1439-44. [CrossRef]
- Rwegasira RG, Mwatawala MW, Rwegasira GM. Seasonal population structure of African weaver ants Oecophylla longinoda (Latreille) (Hymenoptera: Formicidae) under bimodal rainfall pattern in Tanzania. Methods for Enhancing Weaver Ant (Oecophylla Longinoda latreille) (Hymenoptera: Formicidae) Populations for Sustainable Control of Insect Pests of Crops. Doctoral Dissertation, Sokoine University of Agriculture. MOROGORO, TANZANIA. 2016; 74-88.
- Ribeiro S, Espirito Santo N, Delabie J, Majer J. Competition, resources and the ant (Hymenoptera: Formicidae) mosaic: a comparison of upper and lower canopy. Mycological Progress. 2013; 18:113-20.
- Pfeiffer, M.; Tuck, H.C.; Lay, T.C. Exploring arboreal ant community composition and co-occurrence patterns in plantations of oil palm Elaeis guineensis in Borneo and Peninsular Malaysia. Ecography 2008, 31, 21–32. [Google Scholar] [CrossRef]
- Holldobler, B. Territorial Behavior in the Green Tree Ant (Oecophylla smaragdina). Biotropica 1983, 15, 241. [Google Scholar] [CrossRef]
- Exélis, MP, Ramli R. A Review on Distribution Pattern, Nesting Style, Mating Behavior, Colony Organization of Asian weaver ant Oecophylla smaragdina (Hymenoptera: Formicidae) Occupying Strategic Biome-agroforestry Systems of Asia. 2022. [CrossRef]
- Bigger, M. Observations on the insect fauna of shaded and unshaded Amelonado cocoa. Bull. Èntomol. Res. 1981, 71, 107–119. [Google Scholar] [CrossRef]
- Hakala, S.M.; Ittonen, M.; Seppä, P.; Helanterä, H. Limited dispersal and an unexpected aggression pattern in a native supercolonial ant. Ecol. Evol. 2020, 10, 3671–3685. [Google Scholar] [CrossRef]
- Ostwald, M.M.; Guo, X.; Wong, T.; Malaekeh, A.; Harrison, J.F.; Fewell, J.H. Cooperation among unrelated ant queens provides persistent growth and survival benefits during colony ontogeny. Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sangma JS, Prasad SB. Population and Nesting Behaviour of Weaver Ants, Oecophylla smaragdina from Meghalaya, India. Sociobiology. 2021 Dec 23; 68(4):e7204-. [CrossRef]
- Offenberg J, Cuc NT, Wiwatwitaya D. The effectiveness of weaver ant (Oecophylla smaragdina) biocontrol in Southeast Asian citrus and mango. Asian Myrmecology. 2013 Jan 1; 5(1):139-49.
- Peng, R.; Christian, K.; Gibb, K. Locating queen ant nests in the green ant, Oecophylla smaragdina (Hymenoptera, Formicidae). Insectes Sociaux 1998, 45, 477–480. [Google Scholar] [CrossRef]
- Van Itterbeeck, J.; Sivongxay, N.; Praxaysombath, B.; Van Huis, A. Indigenous Knowledge of the Edible Weaver Ant Oecophylla smaragdina Fabricius Hymenoptera: Formicidae from the Vientiane Plain, Lao PDR. Ethnobiol. Lett. 2014, 5, 4–12. [Google Scholar] [CrossRef]
- Van Itterbeeck, J.; Sivongxay, N.; Praxaysombath, B.; van Huis, A. Location and external characteristics of the Oecophylla smaragdina queen nest. Insectes Sociaux 2015, 62, 351–356. [Google Scholar] [CrossRef]
- Rwegasira, R.G.; Mwatawala, M.; Rwegasira, G.M.; Mogens, G.N.; Offenberg, J. Comparing different methods for trapping mated queens of weaver ants (Oecophylla longinoda; Hymenoptera: Formicidae). Biocontrol Sci. Technol. 2015, 25, 503–512. [Google Scholar] [CrossRef]
- Vanderplank, F.L. The Bionomics and Ecology of the Red Tree Ant, Oecophylla sp., and its Relationship to the Coconut Bug Pseudotheraptus wayi Brown (Coreidae). J. Anim. Ecol. 1960, 29, 15. [Google Scholar] [CrossRef]
- Fayle, T.M.; Turner, E.C.; Snaddon, J.L.; Chey, V.K.; Chung, A.Y.; Eggleton, P.; Foster, W.A. Oil palm expansion into rain forest greatly reduces ant biodiversity in canopy, epiphytes and leaf-litter. Basic Appl. Ecol. 2010, 11, 337–345. [Google Scholar] [CrossRef]
- Landis, D.A. Designing agricultural landscapes for biodiversity-based ecosystem services. Basic Appl. Ecol. 2017, 18, 1–12. [Google Scholar] [CrossRef]
- Pag-Ong, AI. Taxonomy and diversity of ants associated with cacao and evaluation of Oecophylla smaragdina Fabricius (Hymenoptera: Formicidae) for biological control of Helopeltis bakeri Poppius (Hemiptera: Miridae) in Quezon and Laguna, Philippines. 2021. Doctorate dissertation, De La Salle University, Manila Philippines.
- Eilenberg, J. Concepts and visions of biological control. In: Eilenberg J, Hokkanen H. (eds). An ecological and societal approach to biological control. Springer, Dordrecht. 2006:1-1.
- Gonzálvez, F.G.; Santamaría, L.; Corlett, R.T.; Rodríguez-Gironés, M.A. Flowers attract weaver ants that deter less effective pollinators. J. Ecol. 2012, 101, 78–85. [Google Scholar] [CrossRef]
- Rodríguez-Gironés MA, Gonzálvez FG, Llandres AL, Corlett RT, Santamaría L. Possible role of weaver ants, Oecophylla smaragdina, in shaping plant–pollinator interactions in South-East Asia. Journal of Ecology. 2013 Jul; 101(4):1000-6. [CrossRef]
Figure 1.
Selected sampling plantations sites in Peninsular and Borneo Malaysia. MPOB Sabah Lahad Datu (a.), MPOB Saratok, Sarawak (b.), Sg. Buloh, Selangor (c.), UM plantations Johor (d.), Felcra, Johor (e.), MPOB Teluk Intan, Perak (f.), Felda Keratong Pahang (g.) and, Felda Gunung Besout, Perak (h.).
Figure 1.
Selected sampling plantations sites in Peninsular and Borneo Malaysia. MPOB Sabah Lahad Datu (a.), MPOB Saratok, Sarawak (b.), Sg. Buloh, Selangor (c.), UM plantations Johor (d.), Felcra, Johor (e.), MPOB Teluk Intan, Perak (f.), Felda Keratong Pahang (g.) and, Felda Gunung Besout, Perak (h.).

Figure 2.
Experimental design demarcation plan.
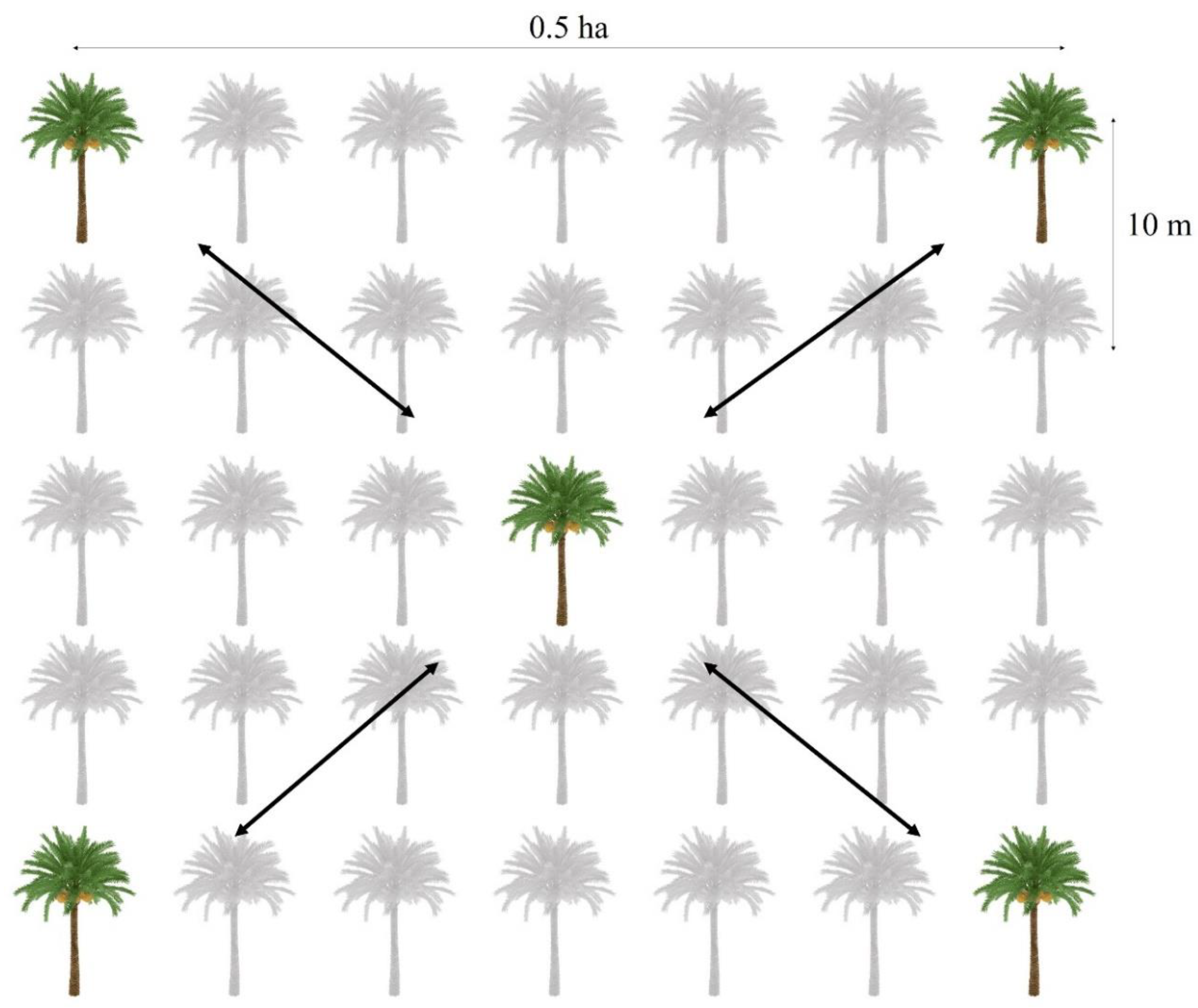
Figure 3.
a-d. Identification of brood and barrack nests. .

Figure 4.
(A-C). Antagonistic ant species presence interference. Photos credit: Exélis Moïse Pierre.
Figure 4.
(A-C). Antagonistic ant species presence interference. Photos credit: Exélis Moïse Pierre.

Figure 5.
Box plot of mature, pre-mature and Immature/incipient O. smaragdina colonies.
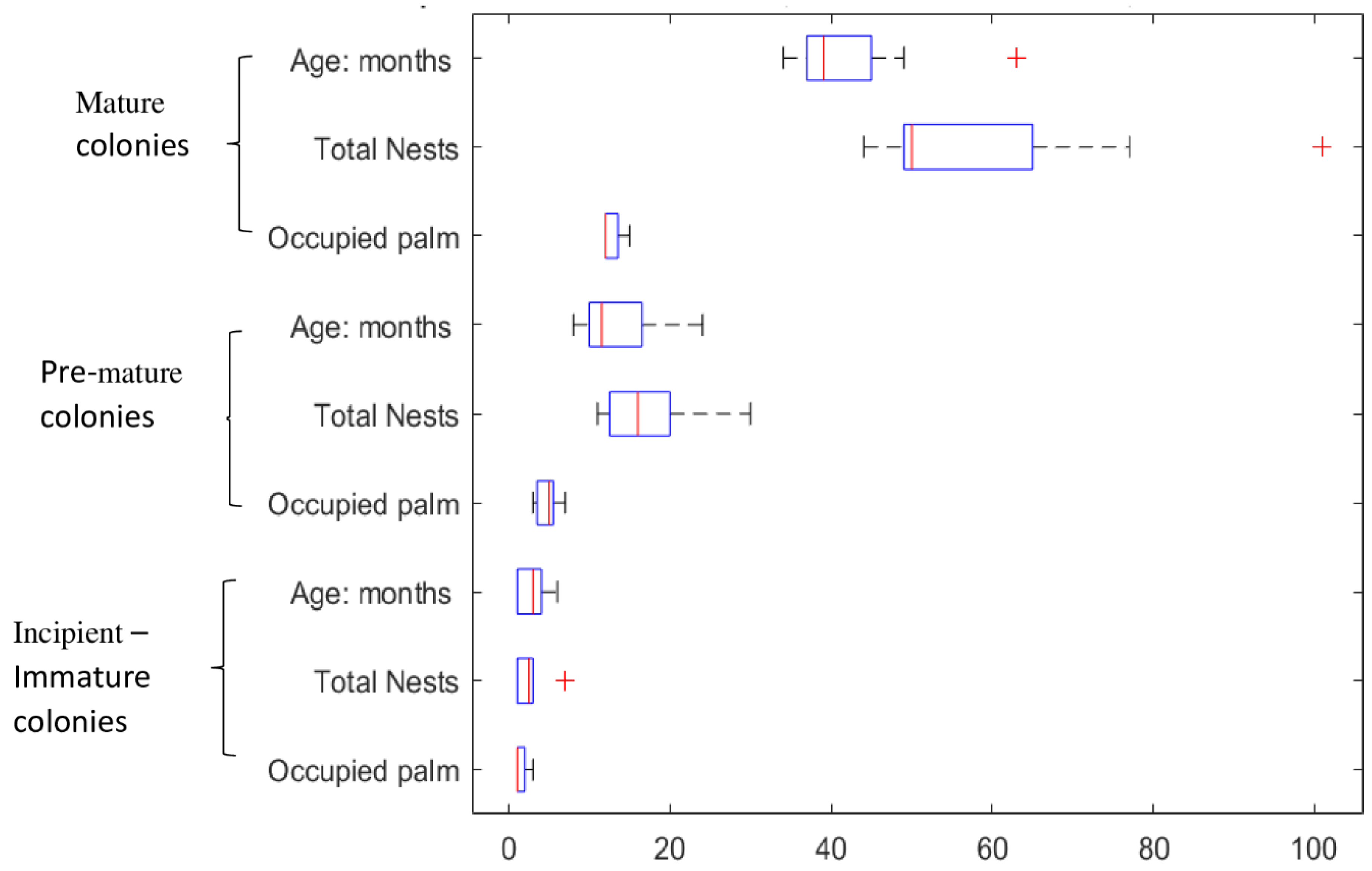
Figure 6.
Discriminant analysis centroids graph. .
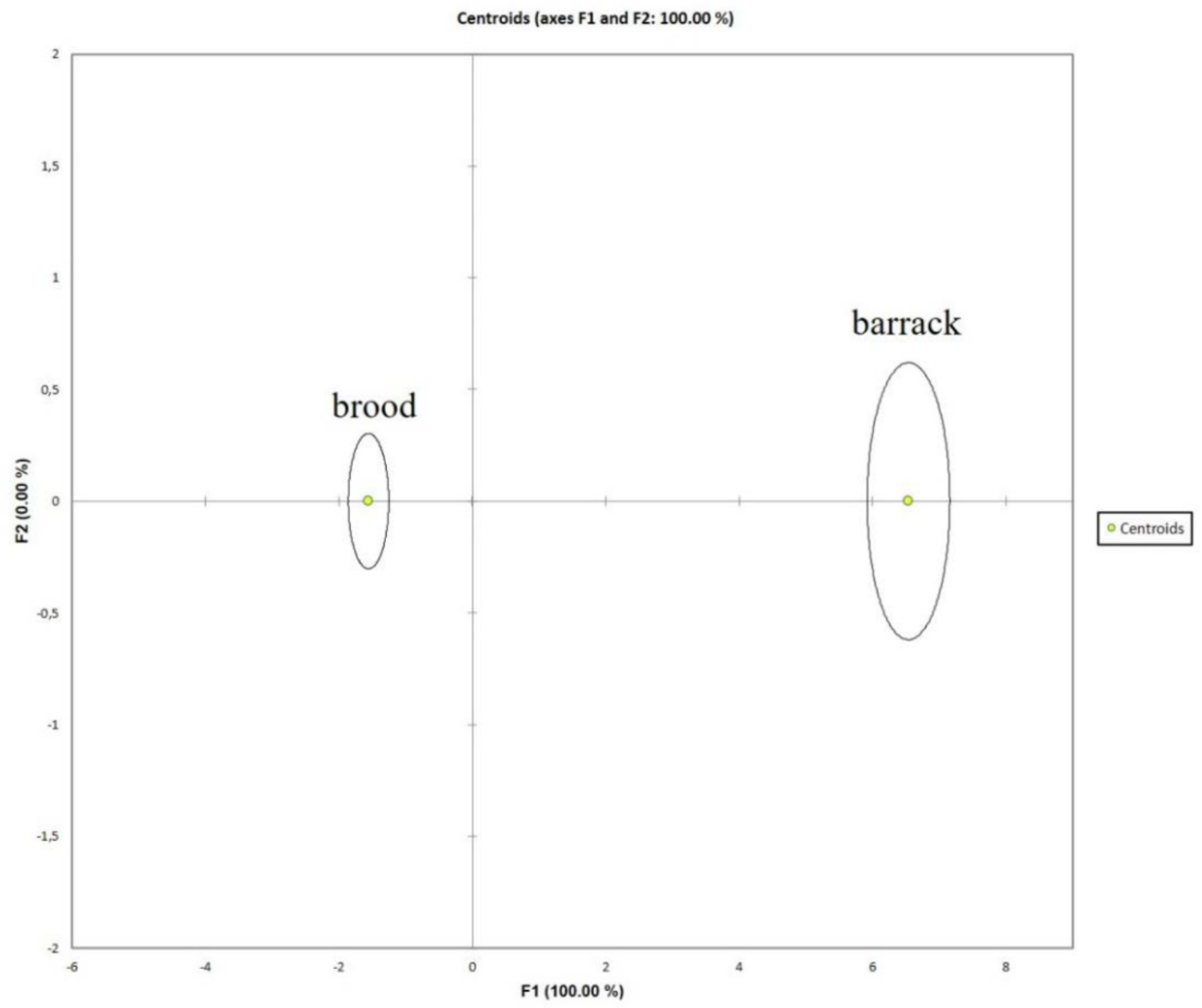
Figure 7.
Poisson probability density function (a-d).
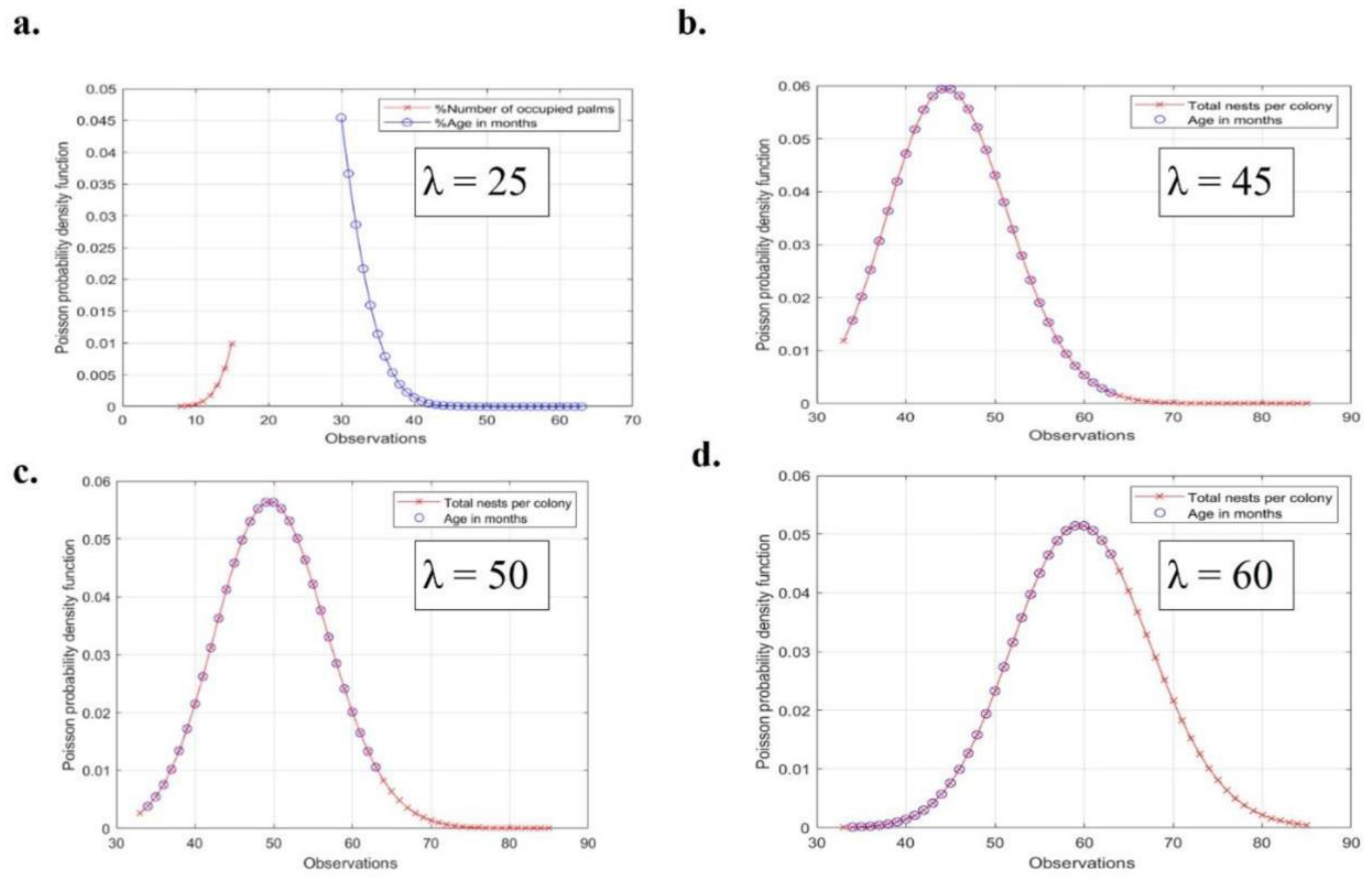

Table 1.
Random sampling of selected oil palm plantations in Malaysia.
| Plantations | Number of colonies | Total Number of nests** | Average Number of nests/ colony | Range of occupied palms/colony | Total Number of occupied palms/total sampling | Average number of palm/ colony | Estimated range of occupied land surface/colony m2 |
|---|---|---|---|---|---|---|---|
| UM Jr* | 27 | 1105 | 40.92 | 8-21 | 320 | 11.85 | 800-2100 |
| Felcra Jr | 20 | 1109 | 55.45 | 10-12 | 200 | 10 | 1000 |
| Boustead Jr | 16 | 832 | 52 | 10-15 | 197 | 12.3 | 1000-1500 |
| MPOB Pr* | 27 | 1216 | 45 | 7-15 | 351 | 13 | 700-1500 |
| MPOB Sb | 20 | 1083 | 54.15 | 12 | 240 | 12 | 1200 |
| MPOB Sr | 16 | 1598 | 99.8 | 12-25 | 219 | 13.6 | 1200-2500 |
| Felda Pr | 10 | 390 | 39 | 7-8 | 72 | 7.2 | 700-800 |
| Total | 136 | 7333 | 386.32 | 7-25 | 1599 | 80 | 4 x 106*** |
| Mean | 19.4 | 1047.5 | 55.2 | 7-25 | 228.42 | 11.42 | 1342.85 |
. *A dozen incipient colonies were detected at UM plantations, Johor; MPOB Teluk Intan, Perak had many immature colonies: data discarded. **Outlier very low occupation surface; Sb = Sabah, Sr = Sarawak, Jr = Johor, Pr = Perak. **Nests formed only by young palm leaflets; old ones are abandoned. *** Total surface = 5,100 ha = 5,100,000 m2; considering 80% of occupation, 4.08 x 106 m2 is the average total occupied surface for all plantations representing an average 816,000 palm trees. 1Age is estimated from close monitoring beginning from 2018 up to 2022: this does not interfere with the method of age maturity estimation incipient stage of this study.
Table 2.
Differences between brood and barrack nests by ANOVA – Tukey (HSD) analysis.
| Contrast | Difference | Standardized difference | Critical value | Pr > Diff | Significant |
|---|---|---|---|---|---|
| brood vs barrack | 3.9524 | 9.4998 | 2.0086 | < 0.0001 | Yes |
| Tukey's d critical value: | 2.8405 | ||||
| Category | LS means | Standard error | Lower bound (95%) | Upper bound (95%) | Groups |
| brood | 7.9524 | 0.1824 | 7.5859 | 8.3188 | A |
| barrack | 4.0000 | 0.3739 | 3.2490 | 4.7510 | B |
| Category* | LS means(Var1) | Groups | |||
| brood | 7.9524 | A | |||
| barrack | 4.0000 | B | |||
* Is the summary of all pairwise comparisons for Q1 (Tukey HSD).
Table 3.
Classification matrix of barrack and brood nests by discriminant analysis and partial least square – discriminating models.
Table 3.
Classification matrix of barrack and brood nests by discriminant analysis and partial least square – discriminating models.
| Model type | Significant variables ranking | Dataset | Correct classification, % | Number of nest and p-values of Fisher distance1,2 | Total nest | ||
|---|---|---|---|---|---|---|---|
| Variable, F-statistics and p-value | Variable and VIP value | Brood | Barrack | ||||
| Discriminant analysis | Number of minor workers (F-statistics: 127.6578, p-value: < 0.0001); Height from the ground (F-statistics: 90.2458, p-value: < 0.0001); Number of worker pupae (F-statistics: 39.5015, p-value: < 0.0001); Number of larvae (F-statistics: 38.3714, p-value: < 0.0001); Number of total workers (F-statistics: 21.0614, p-value: < 0.0001); Number of leaflets (F-statistics: 9.6910, p-value: < 0.0031); and Number of winged queen (F-statistics: 5.1609, p-value: < 0.0274) |
Not related | Training dataset | ||||
| Brood | 100.00 | 18 (1) | 0 (< 0.0001) | 18 | |||
| Barrack | 100.00 | 0 (< 0.0001) | 10 (1) | 8 | |||
| Total | 100.00 | 26 | |||||
| Validation dataset | |||||||
| Brood | 100.00 | 18 (1) | 0 (< 0.0001) | 18 | |||
| Barrack | 100.00 | 0 (< 0.0001) | 10 (1) | 8 | |||
| Total | 100.00 | 26 | |||||
| Partial least square -discriminant analysis | Not related | Number of minor workers (VIP: 1.7272); Height from the ground (VIP: 1.6345); Number of worker pupae (VIP: 1.3537); Number of larvae (VIP: 1.3427); Number of total workers (VIP: 1.1093); Number of major workers (VIP: 0.9928); and Number of leaflets (VIP: 0.8210) |
Training dataset | ||||
| Brood | 95.24 | 16 (1) | 2 (< 0.0001) | 18 | |||
| Barrack | 100.00 | 0 (< 0.0001) | 10 (1) | 8 | |||
| Total | 96.15 | 26 | |||||
| Validation dataset | |||||||
| Brood | 95.24 | 16 (1) | 2 (< 0.0001) | 18 | |||
| Barrack | 100.00 | 0 (< 0.0001) | 10 (1) | 8 | |||
| Total | 96.15 | 26 | |||||
Note: 1Wilks' lambda calculated p-value < 0.0001 at significant level (α) of 0.05. 2Calculated p-value of Fisher distance < 0.05 in parenthesis indicated the brood and barrack nests were significantly different.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



