
Submitted:
05 April 2023
Posted:
05 April 2023
You are already at the latest version
Abstract
The Asian weaver ant Oecophylla smaragdina is a natural enemy (predator) used as a biological control agent in Australia and several Southeast Asian nations against the most destructive and economically important oriental fruit fly Bactrocera dorsalis. For biological control of the invasive bagworm species Metisa plana in oil palm plantations, the composition of the colony's social structure was investigated to ensure its efficacy. On two colonies, all castes were examined stereo microscopically after nest collection and dissection. The head's width, head, thorax, abdomen, and body lengths of workers were evaluated during the morphology analysis. All colonies with more than a thousand eggs comprise of a founding egg-laying queen, reproductive winged green and newly emerged yellow queens, as well as adult drone males and workers. Future queens, males, and workers' pupae and larvae comprised immature individuals. It is the first study to characterise one caste of minor workers, two previously unidentified castes of intermediate workers, and two castes of major workers. The lengths of the body and abdomen are proposed as differentiating factors for workers. The discovery of a polymorphic size frequency distribution model contrasts with the consistently reported existence of archetypical bimodal systems in ants. Intermediate workers foraging outside the nest revealed reconnaissance and aggressive behaviours that aided major workers in their daily tasks. Due to their short lifespan and segregated nests, it is difficult to successfully sample adult males in wide plantations. An entire year's worth of reproductive individuals are produced by stable, three-year-old colonies, with a decrease in population size correlated with a significant decline in rainfall interception.
Keywords:
Weaver ant abundance
; Intermediate workers caste
; Major workers
; Behaviours
; Reproductive
; Polymorphic workers
1. Introduction
The Asian weaver ant, Oecophylla smaragdina, is a dominant arboreal ant species inhabiting forests (Lim, 2007) and oil palm plantation canopies in Southeast Asia Malaysia (Pfeiffer et al., 2007; 2008; Exélis & Idris, 2013; Exélis, 2015). Matured Oecophylla smaragdina colonies are polydomous (more than one nest per occupied host tree) and occupy multiple trees (Holldobler & Wilson, 1990; Debout et al., 2007). The debate regarding the existence of strict monogyny (single eggs-laying founding colony queen), polygyny (multiple eggs-laying reproducing queens per colony), and/or haplometrosis or pleometrosis (multiple queens contributing to incipient colony foundation) has not been geographically resolved (Azuma et al., 2002, 2006), with the exception of Australia northern territory species (Peng et al., 1998). Nevertheless, resolving the polygynous-monogynous status of O. smaragdina using a microsatellite DNA marker revealed that polygyny-polyandry (multiple males mating with a single female) occurs in Java, Indonesia (Schlüns et al., 2009).
The colony structure composition of Oecophylla smaragdina is not completely documented (Schlüns et al., 2009) and further investigation may unveil more information on the workers caste (Exélis et al., 2023). A colony is defined by the presence of a founding depositing eggs dealate queen (shed wings), winged reproductive individuals composed of virgin queens and drone males. A matured colony that survived the founding stage develops into an average of half a million or more individuals inhabiting several tree canopies with an extensive nesting network; these individuals include major and minor workers, pupae, larvae, and eggs (Holldobler and Wilson, 1990). Sadly, only one study provides a comprehensive description of all castes of individuals collected from nests of the mango tree (Mangifera indica) and the pongame oil tree (Millettia pinnata) in Malaysia (Pimid et al., 2012). The social structure of the colony consisted of the same individual castes, with the exception of the embryos, which were not well-defined in the study by Pimid et al. (2012). Another report highlighted the extant third intermediate worker, whose average size is double that of minors and one-half that of majors (Vidhu & Evans, 2011). The colony composition, as defined by a distinct caste system between reproductive and sterile individuals, requires additional consideration.
A brood nest is characterised by the presence of all individual castes (winged green queens, recently emerged callow yellow winged reproductive future queens, drone males, major and minor workers, immature caste i.e. pupae, larvae, and eggs), typically in a high position in tree canopies. A barrack nest contains only major workers in the lowest positions (height from the ground) at the tree canopy's periphery (Crozier et al., 2010). We speculated that a thorough investigation of the weaver ant progeny and barrack nest contents could reveal new information about the unreported worker caste. We also predicted that such an evaluation would provide additional insight into how these novel individual worker caste contributions influence colony-level activities that aid the territorial defence and survival of significant worker colonies. The objectives of the study were as follows: (1) to estimate the number of eggs laid per collected colony; (2) to examine the colony social structure composition by describing the average population size; (3) to describe the worker castes and identify the existence of distinct unreported major and intermediate workers; (4) to determine the yearly emergence pattern of the reproductive individual in relation to rainfall interception (RI) and relative humidity (RH); and (5) to briefly describe the population dynamics of the studied species.
2. Methodology and Materials
2.1. Study Sites
The main study was conducted at the oil palm plantation Felda Gunung Besout in Perak, Peninsular Malaysia (03°50'04"N 101°17'48"E). Mango (Mangifera indica), key lime (Limau nipis, Citrus aurantiifolia), lemon (Citrus limon), Calamansi- Philippines lime (Limau kasturi; Citrus microcarpa), and durian (Durio zibethinus) are among Oecophylla ants' favourite host plants (Lim, 2007). The area experiences significant monthly precipitation fluctuations ranging from an average of 400 mm/month to more than 1000 mm/month. Additional sampling sites comprised oil palm plantations located in: Teluk Intan, Perak (Peninsular Malaysia) (3°49'07"N 100°58'59"E) and Felcra-Universiti Malaya plantations, Kota Tinggi Johor (Peninsular Malaysia) (2°02'03"N 103°51'50"E), Lahad Datu, Sabah (Borneo, East Malaysia) (5°01'52.4"N 118°25'33.2"E), and Saratok, Sarawak (Borneo, East Malaysia) (1°55'25.4"N 111°13'33.0"E) under the Malaysian Palm Oil Board (MPOB).
Figure 1.
A-C. Sampling site at Felda Gunung Besout Perak. A. The habitat of the study site within oil palm plantations. B. Brood nest collected from a palm frond in an elevated position. C. A large brood nest was sampled and captured from a Calamansi tree orchard within a palm plantation.
Figure 1.
A-C. Sampling site at Felda Gunung Besout Perak. A. The habitat of the study site within oil palm plantations. B. Brood nest collected from a palm frond in an elevated position. C. A large brood nest was sampled and captured from a Calamansi tree orchard within a palm plantation.

2.2. Nests Collection and Caste Structure Laboratory Assessment
Due to the difficulty of capturing a colony's egg-laying queen and the absence of male alates during sampling, other collection records from sampling sites are included in this evaluation. This is due to the possibility that successful capture of dealate queens is only feasible through the use of invasive, destructive methods, resulting in the extinction of the colony (Holldobler & Wilson, 1990). Exélis and Idris (2013; 2015) conducted a five-kilometer-radius survey to detect the presence of matured O. smaragdina colonies. Upon confirming the presence of large colonies exhibiting polydomous nesting behaviours in seven palm trees within the plantations, a 1000-meter-wide transect was established along a path within the plantation. A minimum distance of 200 metres was maintained between palm tree canopies sampled to prevent the capture of nests from the same O. smaragdina colony. The antagonism test was conducted to further enhance the detection of distinct colonies. Between released and resident ants, ants from the same colony will pass and forage and spontaneously communicate. Different colonies of ants will result in a conflict (Kamhi et al., 2015). Three distinct palm tree-dwelling colonies, Felda I, Felda II, and UM III, were identified and tagged during the day. After sustained daytime rainfalls, three brood nests were collected from each colony during the late night to midnight period 2230 hours to 0030 hours, which corresponds to periods of low main worker activity determined in a separate study (Exélis, 2015). Felda I colony represented a large colony established on Calamansi (Limau kasturi; Citrus microcarpa) short trees of 2 m in height, forming an orchard with an average 100 m x 200 m land area within palm tree plantations. It was only feasible to collect nests during the daytime on short palms (3 m) using pyrethroids. Only samples from Felda I and UM III were reported (samples from Felda II suffered from misplacement during the COVID-19 pandemic and many samples were dehydrated). Brood nests were identified based on a visual discrimination height parameter established in another study, which corresponded to an average position in palm tree canopies between 6 and 10 metres above the ground (Tables 3 & 4). A long hook was used to bring down the tall fronds slowly in order to cut the brood nests with a sharp cutter from the base of leaflets affixed to the rachis of each palm frond, and all weaver ants were collected in a rigid plastic bag (1 m x 60 cm). Pyrethrin knockdown was used to sample weaver ants in order to avoid ferocious stings and minimise individual escape and loss. Nests of O. smaragdina ants were dissected and classified according to the following castes after 24 hours of exposure to open air: dealate queens, egg clusters, larvae, pupae, female alate virgin queens, male alate, major, intermediate, and minor workers (Pimid et al., 2012). For each brood, the number of individuals of each caste was determined and recorded. The average position of barrack nests dissected with a keen surgical blade was determined to be between 3 and 4 metres below the lowest palm fronds, and their caste composition was recorded. Brood and barrack nests (two and one per colony, respectively) were used to evaluate the composition of the individual populations of three colonies Felda I, Felda II, and UM III, which was helpful in determining the expected worker castes (major, intermediate size and minor workers). In the field, all varieties of caste specimens that had not been exposed to the sun were separated, preserved in 95% ethanol, and transferred in 70% ethyl alcohol. The samples were transported to the Entomology Laboratory, Universiti Malaya, and the Centre for Insect Systematics, Universiti Kebangsaan Malaysia, for additional research on the distinct individual caste population and the average total number of eggs per colony.
2.3. Estimation of Eggs Number per Nest
In the first experiment, a total of 20 nests representing ten colonies (2 nests per colony) were selected, with two primary brood nests per colony containing egg clusters encased in white silken chambers that were separated from other individual castes within the nest. Egg clusters must be collected with care, as they are fragile (breaking or exploding under pressure) and attach to the interior nest wall. After filtration and drying of all specimens, the total number of eggs per nest was estimated by clusters (Figure 3 A-B) using dissecting surgical needles and narrow forceps to separate eggs under a solution of ethyl alcohol 80% in a petri dish. Using thin needles and a 70% alcohol solution, the eggs poured into a tiny petri dish glass could be easily separated. The separated embryos were then sucked directly into a microscope using a pipette. When it was challenging to separate eggs from each cluster due to their fragility, they were placed on white filter paper and counted visually with the naked eye before being examined under a Nikon e200 compound microscope. The eggs were then tallied using a stereomicroscope Nikon SMZ800N on a PC HP Compaq running Windows 7 Professional connected to a Nikon digital sight DS-fi2 camera and imaging software NIS Elements version 4.0. A ZEISS Stereo Discovery V20 microscope equipped with AxioCam MRc software Image Analyzer was also utilised.
2.4. Estimation of Individual Population Size from Matured Colonies
In the second experiment, six nests (3 for each colony) were brought back to the laboratory from two sizable colonies inhabiting 8 and 12 palm trees, representing Felda I and UM III, respectively. The Felda I and UM III polydomous colonies consisted of 33 and 74 geographically interconnected nests, respectively. The contents of every nest were once spread out on a white laboratory table. Each nest was meticulously dissected to obtain an accurate count of its inhabitants by directly counting all individual castes: adults (major, intermediate, and minor workers with winged reproductive of each sex), pupae (worker, male, and queen pupae), and larvae. By multiplying the mean of the total worker population for each colony by the total number of nests, the colony population size (PS) can be estimated on average: PS= ∑x (Total nest number)*(TW), where TW represents the total worker values.
2.5. Individual Worker Caste Measurements
In the third experiment, the body and abdomen lengths (BL-AL) of the collected adult and pupae workers revealed distinct differences in body size. To determine the size distribution of workers within a colony, hundreds of individuals were chosen at random to have their body measurements recorded. After mounting an individual with a pin, images and measurements are recorded. The sizes of the maximum head width (HW, transversal black arrow in figure 2A), head length (HL, longitudinal black arrow in figure 1C), thorax (TL-figure 2B), abdomen length (AL-figure 2B), and body length (BL, transversal red arrow in figure 2C) (hypothesized different sizes for five distinct variables) were recorded. The examination of all variable measurements was performed using a stereomicroscope Nikon SMZ800N on a PC HP Compaq running Windows 7 Professional attached to an integrated Nikon digital sight DS-fi2 camera and imaging software NIS Elements 4.0. The morphological characteristics were compared to images of an O. smaragdina specimen from Chonburi Pattaya, Thailand, on AntWeb (CASENT0173647). Using the stereomicroscope Nikon SMZ800N and aided by the Zeiss stereo discovery V20, a dissection was performed to identify any imperfect sightings or concealed body parts. The dataset of body size records was pre-processed and filtered to reorganise and categorise them into distinct castes based on the similarity of the values of the five variables in order to visualise the size distribution. It is anticipated that the distribution will be either normal, bimodal, or multimodal.
Figure 2.
A-C. Comparison between specimens.

Major worker from Thailand (CASENT0173647, April Nobile) and specimens seen in Felda Gunung Besout plantations during this research. Three views are shown: a frontal view (HW, red line), a side view (TL, yellow line, AL, red line, and BL, black line), and a dorsal view (C). (HL, red line – BL, black line). Images are accessible at www.antweb.org.
2.6. Monitoring Reproductive Caste Emergence
This fourth experiment's goal was to evaluate the productive caste output throughout the entire year from established colonies. The plantation management confirmed that the occupied areas were chosen based on their known presence and that the average colony age was 3 years and older. From 18 to 24 months, these colonies start to generate adult sexual. In order to track the appearance of productive individuals, such as winged virgin queens and male drones, from 2011 to 2022, two different methods were used: destructive sampling and direct surgical nest dissection. At the selected plantations described in section 2.1, the sampling of brood nests was typically done overnight between the hours of 2200 and 0100. From 2011 to 2016 on a three-month basis, alternating to various months each year to cover the entire year. Nest sampling frequency decreased from 2018 to 2022 (November 2018, August 2019, January-March and November-December 2020, January 2021, April-May-June to October 2022). We can acquire nests for every month of the year. An overall dataset displaying which month of the year had positive or negative emergence instances was created by recording and pooling the values. The department of irrigation and drainage (Ministry of Environment and water - https://publicinfobanjir.water.gov.my/hujan/?lang=en) provides the rainfall interception (RI) means (mm) dataset log.
2.7. Intermediate Workers Evaluation
The fifth experiment consisted of monitoring and evaluating the specific function of the two identified intermediate worker castes using three mature, distinct colonies selected from separate palm tree blocks and designated colony I, II, and III. The intermediate size workers (an average of 10 per colony) were observed in the field on fronds, leaflets, trunks, and the ground in order to observe their foraging activity and distinguish their behaviour from that of the major workers. Following this, a comparison was made with minor workers who are typically restricted to the nest compound alone. The surveillance was conducted daily from 30 November to 6 December 2020 and 6-13 July 2022 during peak hours (1100-1300 hours & 1600-1800 hours). Each observation lasted five minutes and three observations were performed every hour (1100-1130-1155 hours). Photographs were captured to document the activities of workers outside nests.
3. Statistical Analysis
A One-way ANOVA post-hoc Tukey HSD, Scheffé test supported by Bonferroni and Holm was conducted to compare the ten egg-producing colonies. For further analysis, all random head width (HW), head length (HL), thorax length (TL), abdomen length (AL), and body length (BL) measurements were tabulated in Excel tables. The mean number, standard deviation (SD), and standard error of the mean (SEM) for each caste were calculated using SPSS version 26 (Pallant, 2010).Analysis of Variance of the five sub-castes of workers was performed to determine if there are statistically significant differences in each factor/variable (AL, BL, HL, HW, and TL; see Table 4). Since there are distinctions between these five sub-castes, it became necessary to conduct additional analyses (Figure 8). Prior to analysis, the mean number of each variable size was log-transformed to achieve normal dataset distribution. To determine whether there are statistically significant differences between the five workers castes, a one-way between-groups analysis was conducted. On HW, HL, TL, AL, and BL, we conducted a one-way repeated measures ANOVA with Tukey HSD test (Table 5). For each caste, hundreds of individuals were observed in total. Upon processing and categorization into five groups, hundreds of workers examined in section 2.4 were reduced to twenty individuals per category under five recorded parameters, for a total of 500 dataset values. Using the following online statistical analysis tool, the dataset was processed: (https://www.socscistatistics.com/tests/anovarepeated/default.aspx). Further analysis was conducted by considering a comparative study for each size parameter (HW, HL, TL, AL, and BL) across all five distinct workers (major V, IV, intermediates III, II, and minor I) in order to highlight the differences and detect the hypothesised potential predictor factors (AL and BL). Using box plot profiling, the MATLAB software R2021b utility (2021) was utilised to determine whether or not there are statistically significant differences between the five worker castes.
4. Results
4.1. Oecophylla Smaragdina Colony Social Structure Description
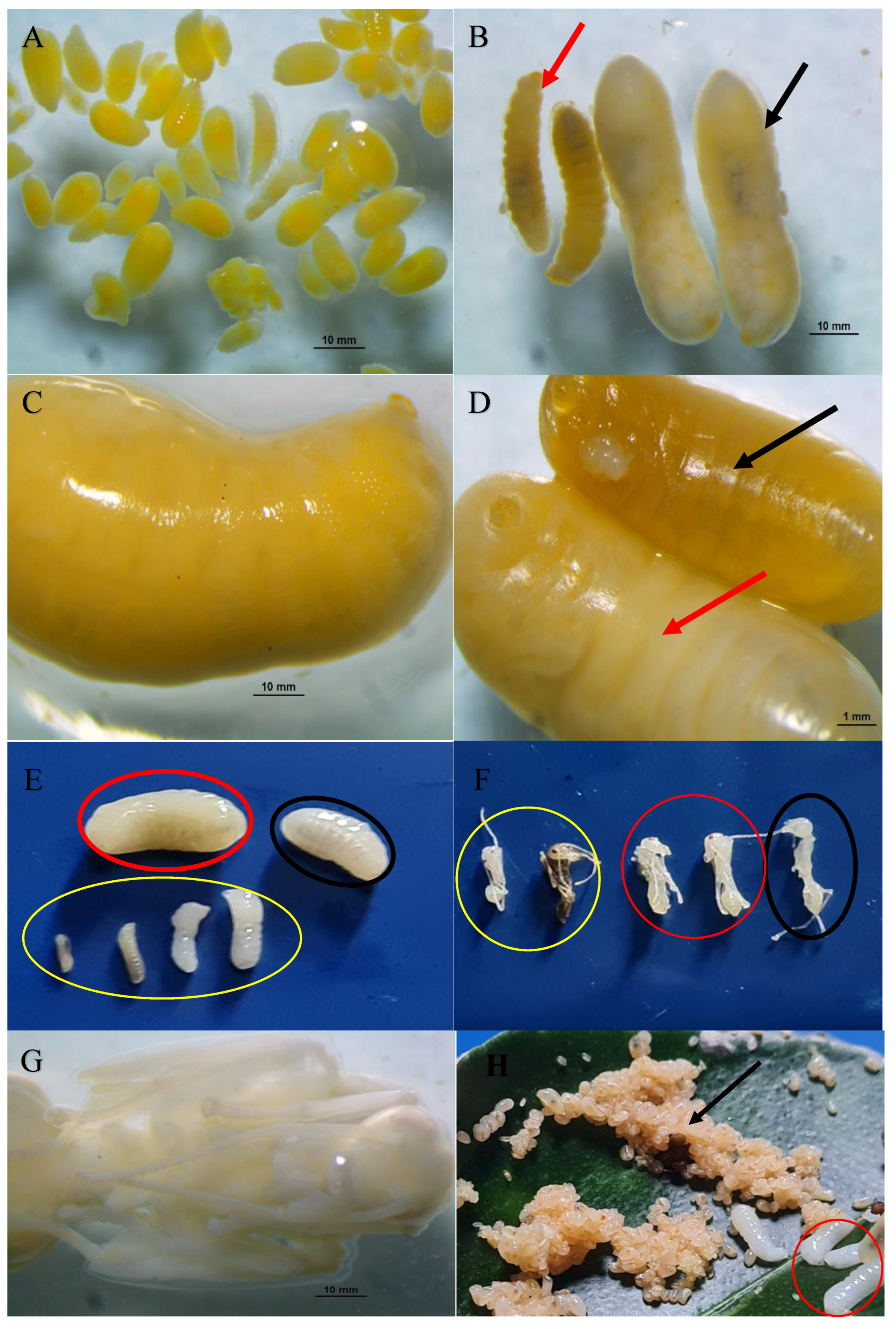
Figures 3 A-E, 4 A-B, 5 A-G, and 6 A-E show how diverse individuals make up a colony in terms of caste. Within each colony structure, members of the Oecophylla smaragdina caste represented unique individuals. Figure 3A–E show the adult stage of each individual caste, including adult workers and the gravid dealate queen, drone males, winged green, and newly emerged yellow queen. In comparison to green alate virgin queens, the captured dealate founding queen (Lahad Datu, Sabah MPOB plantations) had a larger abdomen. When alate female sexuality first emerged, it had a yellow colour and a slightly smaller abdomen. Within a week, it changed to green hue. In the MPOB plantations in Saratok, Sarawak, the mature alate males, which are smaller than the queens are black in colour (like house flies), were discovered living in several nests by themselves in large numbers (more than 200 for two nests). All other locations' nest collections revealed a lack of adult male presence. It was discovered that there were five different size individuals for the workers caste, comprised of individuals with barely discernible colour contrasts and basically noticeable differences in their abdominal and body sizes. The individuals were the major workers V, who were larger and darker reddish-brown in colour; the major workers IV, who were intermediate workers III and II; and the minor workers I (Figure 3 A-E). Among workers, the two-novel caste was recognised (Figure 3 E, red and yellow circles). The considerable variation in body length between each worker is depicted in Figure 3E. The large major workers abundance is noticeably greater than for minor workers in all collected nests from various colonies and geographical sampling locations, but more balanced with the two intermediate workers III and II.
4.2. Colonies Eggs Count Estimation
Table 1 shows the average number of eggs inside 8.34 mm x 5.02 mm standardized size clusters from Felda Gunung Besout Perak oil palm plantations.
Table 2's p-value for the F-statistic of a one-way ANOVA is greater than 0.05, indicating that the treatments do not differ significantly at this level of significance. The Tukey HSD test and other multiple comparison tests, such as Scheffe and Bonferroni, yielded similar values; therefore, the colonies did not reveal differences in egg production.
aBonferroni and Holm tests gave same results, significant level, < 0.05.
4.3. Colony Caste Composition-Description
Eggs recently laid by queens were white to slightly yellowish and formed into cubic clusters surrounded by an unidentified yellow gelatinous substance. The sharp, thin tip of an egg corresponds to the head (Figure 4 A-B; Figure 5 A). All captured brood nests contained these egg clusters in distinct whitish silken compartments on the nest's underside. At a later time, the embryos were white and had a distinct miniature oval shape. Individually, eggs are smaller than the characteristic white, elongated worm shape of the larvae, and there is no growth in body parts. The width varies between 0.37 mm and 0.50 mm, while the length varies between 0.96 mm and 1.15 mm (N = 50). Eggs are always found as compact, clustered, shiny, adhesive entities (Figure 4 A-B), whereas larvae and pupae are found separately as dry, solitary, non-sticky entities. (Figure 5 A-B).
Figure 4.
A-B. O. smaragdina yellowish eggs clusters. (A) Separated and tallied eggs from brood nests with a glossy appearance. (B) Compact egg clusters with a micron-sized standard. Examining using Nikon SMZ800N stereomicroscope. Photo credit:Exelis Moïse Pierre.
Figure 4.
A-B. O. smaragdina yellowish eggs clusters. (A) Separated and tallied eggs from brood nests with a glossy appearance. (B) Compact egg clusters with a micron-sized standard. Examining using Nikon SMZ800N stereomicroscope. Photo credit:Exelis Moïse Pierre.
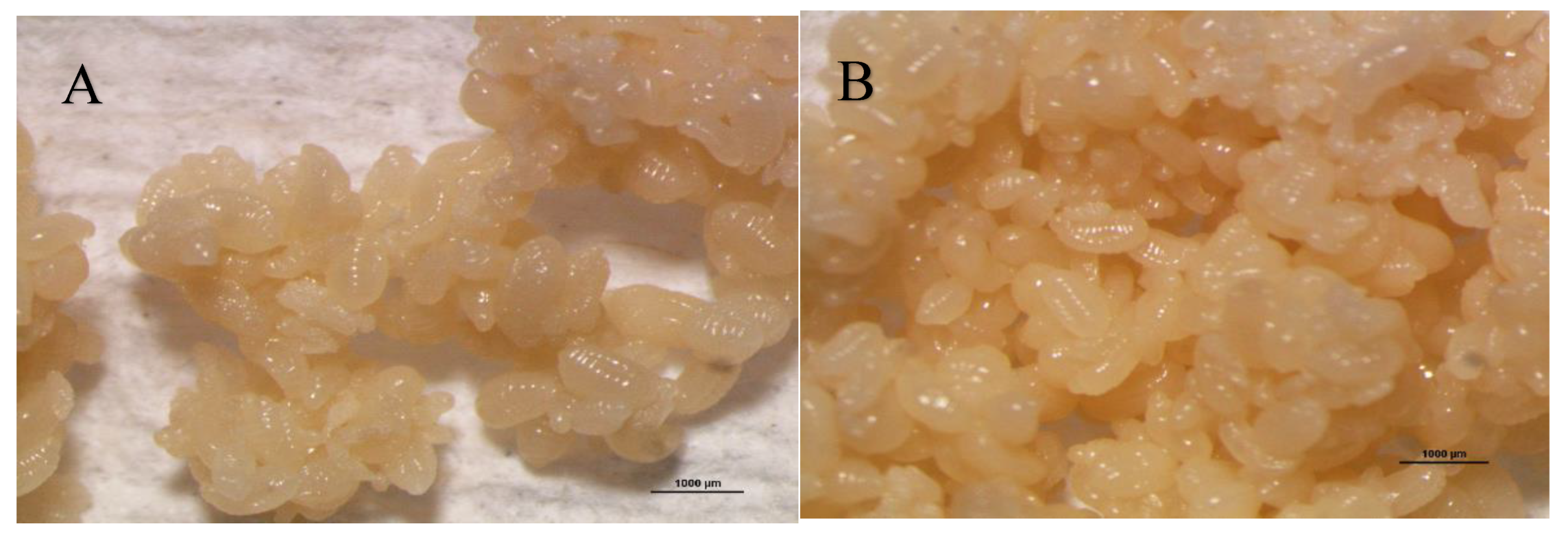
Figure 5 A-H depict the immature stages of the various castes that make up the social structure of the O. smaragdina colony, which consists of eggs, workers, the queen, male larvae, and queen and worker pupae. The eggs are clustered, encased in an unidentified protective substance, and much smaller than larvae (Figure 5 A, separated under 80% ethyl alcohol). Larvae are whitish, more elongated than eggs, and have a thinner, sharper extremity that corresponds to the head and resembles the shape of worms (Figure 5 B-E-H). Pupae (Figure 5 F) are distinguished from larvae (Figure 5 E) by a foetal position displaying the appearance of eyes, mouth, legs, and wing growth that accurately predicts the shape of the adult weaver ant. Queen and male larvae are distinguished by their interior curve's lighter and darker hues, respectively (Fig. 5 D).
Figure 5.
A-H. Immature O. smaragdina individuals.O. smaragdina distinguished yellow eggs (A). Major (black arrow) and subordinate worker larvae (red arrow) (B). Big queen larvae (C). Male drone larva (black circle) and female queen larva (red arrow) (D). Queen (red circle) and male larvae (black circle). Major worker, intermediate III, intermediate II, and minor worker I (yellow circle), from right to left. (E). Workers' pupae with their distinctive sizes, from right to left: a major worker (black circle), an intermediate worker (red circle), and a minor worker (yellow circle). (F). Queen pupae (G). Identifies the distinction between eggs (black arrow) and larvae (red circle). (H). All figures scale are 1 mm. Examination via Nikon SMZ800N stereomicroscope (Photo credit: Exelis Moïse Pierre).
Figure 5.
A-H. Immature O. smaragdina individuals.O. smaragdina distinguished yellow eggs (A). Major (black arrow) and subordinate worker larvae (red arrow) (B). Big queen larvae (C). Male drone larva (black circle) and female queen larva (red arrow) (D). Queen (red circle) and male larvae (black circle). Major worker, intermediate III, intermediate II, and minor worker I (yellow circle), from right to left. (E). Workers' pupae with their distinctive sizes, from right to left: a major worker (black circle), an intermediate worker (red circle), and a minor worker (yellow circle). (F). Queen pupae (G). Identifies the distinction between eggs (black arrow) and larvae (red circle). (H). All figures scale are 1 mm. Examination via Nikon SMZ800N stereomicroscope (Photo credit: Exelis Moïse Pierre).
4.4. Colonies Dissected Nest Content – Reproductive Population Size Estimation
Due to the impossibility of distinguishing between queen and brood colonies, only two categories are considered. The height of the brood and the barrack nest made it simple to distinguish between the two. Depending on the extent of the palm tree, brood nests were located at heights between 6 m and 10 m, whereas barrack nests were typically located between 2 m and 5 m (Tables 3 & 4). This factor facilitated the sampling of several types of brood nests from diverse colonies, allowing the avoidance of barracks nests. The majority of the dissected barrack nests (N=50) in the field were inhabited by larger-sized dark brown to reddish large major workers V and IV sub-caste (Figure 6 A-B). In Felda I and UM III colony nests, the absence of adult male drones was compensated by the presence of queens and male larvae. As described in Section 4.3, Table 3 and Table 4 demonstrate that all caste forms are represented (see Figure 5 A-H). An estimate of the reproductive winged queen population size ranged from 9 to 11 per nest in early matured colonies (24-36 months of age; N= 10 colonies), yielding a range of 63 to 156 queens per colony on average. In older matured colony (48 months and above), the number of alate queens range from 19 to 21 per nest and 475 to 630 queens per colony (T). In brood nests with extensive egg clusters, yellow larvae of an unidentified species of large homopterans were discovered (collected first in May 2012 at MPOB plantations in Teluk Intan, Perak). The yellow larvae exuded a transparent nectar-like substance from their midline. These homopterans were cared for by major workers observed gathering these ingredients (under laboratory condition). The contents of O. smaragdina's nests in the Felda I colony are analysed descriptively in Table 3. Table 4 displays a sample result for a UM III large size colony with a large total worker population of 64,761 individuals. The presence of reproductive adult male was not detected in the samples. At the exception of a large number of adult male, collected from guava trees (Psidium guajava) nearby to others occupied palms, secluded in two separate nests in Borneo Sarawak. Significantly more major workers exist than intermediate or minor workers. The average number of workers in this colony was estimated to be 4.8 million. Another large population of Asian weaver ants was discovered in MPOB plantations in Saratok, Sarawak, with 149 interconnected polydomous nests occupying 12 palms.
An estimation of the population size of Felda I is: PSFelda I = 33 nests x 11,962.00 = 394,746.00 ± 159,647and UM III, PSUMIII = 74 x 64,761.00 = 4,792,314.00 = 4.8 106 ± 525,136 representing an average of major-intermediate-minor total workers.
4.5. Workers Caste Description
Figure 6 A to E show the size variations between the main body parts, including HW, HL, TL, AL, and BL, with the abdomen and body length showing the most notable variation. There are a total of 12 antennal segments for all the workers (Figure 7, black circle).
Figure 6.
A-E. Distinct workers caste I to V. A: Senior major worker V. B: Large major worker IV. C: Intermediate size worker III. D: Intermediate size worker II. E: Minor worker I. Examination by stereomicroscope Nikon SMZ800N (A-B) and ZEISS Stereo Discovery V20 microscope (C-E). The lines are indicating the sizes of variables HW, HL, TL, AL and BL. Photo Credit (Exélis Moïse Pierre).
Figure 6.
A-E. Distinct workers caste I to V. A: Senior major worker V. B: Large major worker IV. C: Intermediate size worker III. D: Intermediate size worker II. E: Minor worker I. Examination by stereomicroscope Nikon SMZ800N (A-B) and ZEISS Stereo Discovery V20 microscope (C-E). The lines are indicating the sizes of variables HW, HL, TL, AL and BL. Photo Credit (Exélis Moïse Pierre).

Figure 7.
O. smaragdina worker Antennal segments. Examination by stereomicroscope Nikon SMZ800N. Photo Credit (Exélis Moïse Pierre). .
Figure 7.
O. smaragdina worker Antennal segments. Examination by stereomicroscope Nikon SMZ800N. Photo Credit (Exélis Moïse Pierre). .

4.6. Worker Castes Body Size Analysis
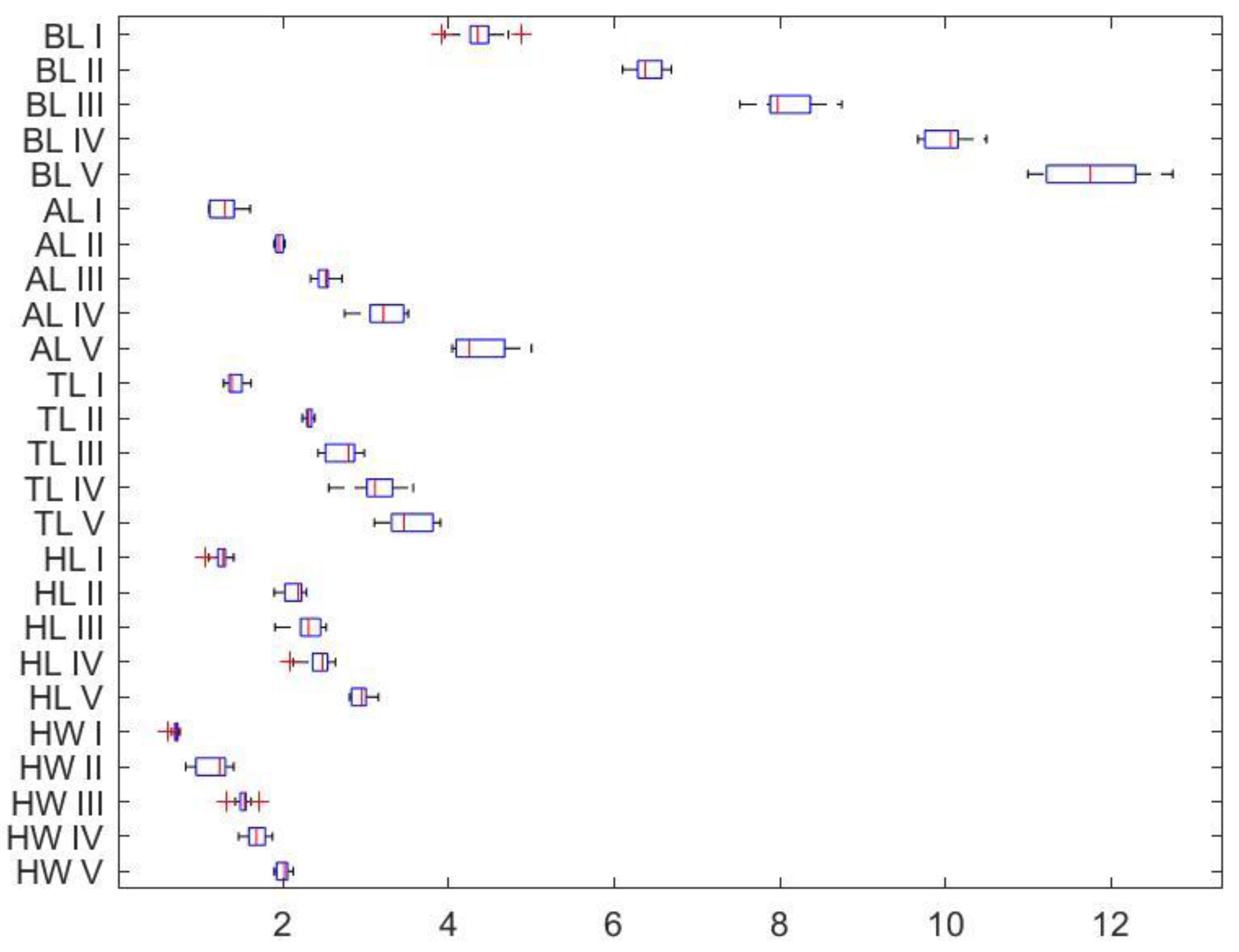
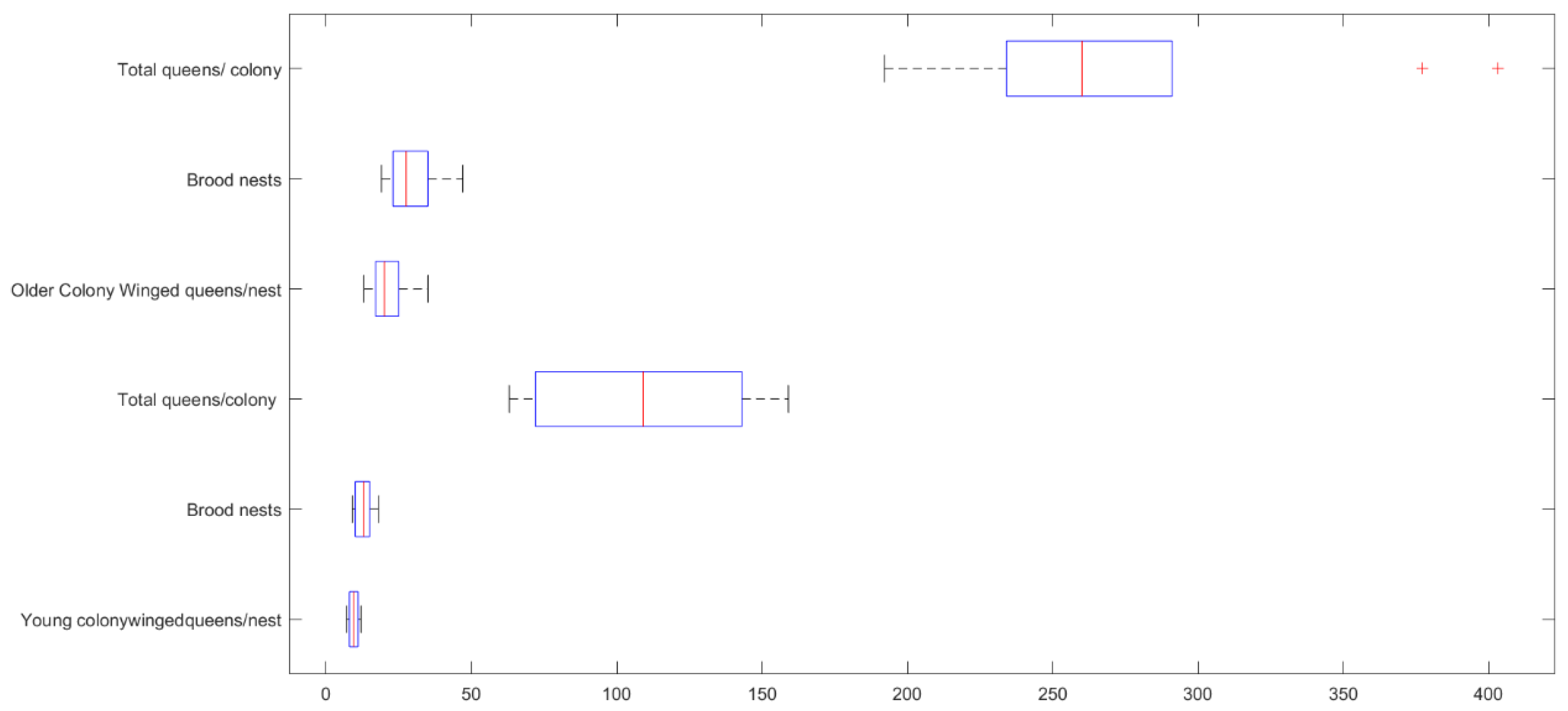
Figure 8 demonstrates that the ratings of the abdomen and body length (AL, BL) have greater significant variability than the measurements of the head width, head length, and thorax length (HW, HL, TL), which include all five different worker groups. Figure 8 shows how each variable differs for each of the five distinct workers. Table 5 and Table 6 show the statistically significant differences between all 5 worker classes, which are primarily dependent on the size of the abdomen and body.
The overall means for each variable for each worker identified as belonging to a subcaste are shown in Table 5 along with the corresponding standard deviations (SD) and standard error means (SEM). One-way ANOVA is used to clarify the significance of the found differences between each worker category.
The combined means of each variable for the five workers are shown in Table 6 as major V, IV, intermediate III, II, and minor I, respectively, based on the one-way ANOVA findings. The edge of significance is to AL and BL as the primary possible predictor factors with higher sum of squares (SS), mean sum of squares (MS), F and p values, and significantly different from all other three variables at p .00001.

Table 5.
Variables means with SD and SEM.
| HW mm | HL mm | TL mm | AL mm | BL mm | Worker Castes |
|---|---|---|---|---|---|
| 1.99 | 2.93 | 3.53 | 4.38 | 11.8 | V |
| 1.68 | 2.43 | 3.14 | 3.20 | 9.99 | IV |
| 1.51 | 2.28 | 2.73 | 2.50 | 8.10 | III |
| 1.15 | 2.12 | 2.30 | 1.95 | 6.41 | II |
| 0.70 | 1.25 | 1.411 | 1.27 | 4.36 | I |
| ±0.49 | ±0.61 | ±0.81 | ±1.19 | ±2.92 | ±SD |
| 0.223 | 0.27 | 0.365 | 0.53 | 1.30 | SEM |
Table 6.
One-Way Repeated Measures ANOVA with Tukey HSD test.

Figure 8.
Box plot showing the overall distribution of the ratings variables of all five distinct workers.
Figure 8.
Box plot showing the overall distribution of the ratings variables of all five distinct workers.
4.7. Emergence-Abundance of Reproductive Individuals
Every month of the year, samples of reproductive individuals were taken, but their dynamic was not assessed. It was made up of adult yellow- and green-winged queens. In MPOB plantation in Saratok, we only tested drone males once (Tab 3 and 4). In oil palm plantations, it has been rare to see adult males. Table 7 and figure 9 demonstrate that O. smaragdina sexual occurs throughout the year, with an increase correlating with greater relative humidity levels. May through August saw fewer mature or immature queens, which was correlated with lower monthly rainfall interception and drier days. October through February saw greater monthly rainfall interception, which led to a rise in reproductive emergence. In one brood nest that was 10 metres high, there was a massive concentration of mature green winged queens that was exceptionally much higher than other months. The abundance of reproducing queens in both young and older colonies, per nest and per colony, is depicted in Figure 10 and Table 8. Figure 10 shows a higher population density in favour of older colony, while the presence-production of adult males might be decreasing in much older matured colony.
Table 7.
Yearly means of O .smaragdina reproductive per nest.
|
Months |
Jan |
Feb |
Ma |
Ap |
May |
Jun |
Jul |
Aug |
Sep |
Oct |
Nov |
Dec |
Means ±SD |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Yellow winged queens |
33 |
21 |
11 |
13 |
11 |
0 |
7 |
9 |
15 |
25 |
41 |
58 |
20.3 ±16.5 |
| Green winged queens |
51 |
48 |
21 |
19 |
13 |
0 |
15 |
17 |
33 |
35 |
55 |
138 |
37 ±36 |
| Gravid queen | 0 | 0 | 0 | 0 | 3 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | NA |
| Drone males | 0 | 0 | 0 | 0 | 0 | 0 | 250 | 0 | 0 | 0 | 0 | 0 | NA |
| *RI mm | 460 | 420 | 310 | 290 | 280 | 215 | 210 | 280 | 310 | 380 | 480 | 550 | 348.75 ±108.3 |
|
1RH % Locations |
95 MPOB TIP |
95 ** |
93 Felda PK |
92 ** |
93 ** |
89 Felda GBP |
90 MPOB SB |
89 UM |
91 Felcra Johor |
93 Felcra Johor |
95 Felda GBP |
97 UM |
92.6 ±2.5 |
Nov to Feb: higher rainy days frequency; May to Oct feature drier day frequency; 1Relative humidity, NA: not applicable. *Rainfall interception. **MPOB TIP: Teluk Intan, Perak. FGBP: Felda Gunung Besout, Perak. UM: Universiti Malaya plantations in Johor. Felda PK: Keratong, Pahang. MPOB SB: Sabah/Sarawak Borneo.
Figure 9.
Monthly Distribution of reproductive emergence.
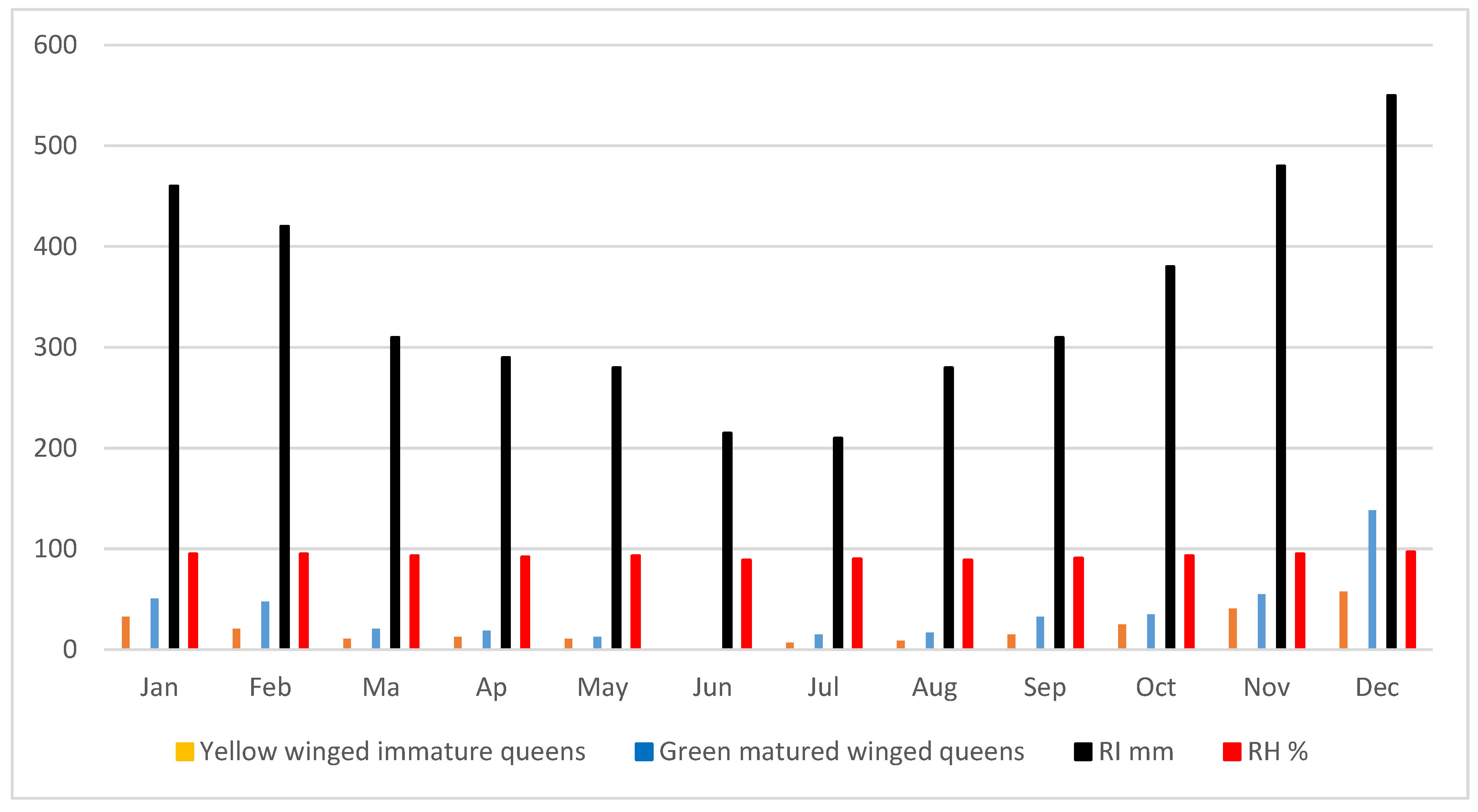
Table 8.
Reproductive population size estimation: young-older colony.
| Colony No. |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
Means ±SD |
||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Young colony winged queens/nest Brood nests Total queens/colony Older Colony Winged queens/nest |
9 11 99 19 |
8 9 72 21 |
11 14 143 17 |
10 13 110 21 |
12 16 156 31 |
8 10 64 25 |
7 9 63 17 |
9 13 108 15 |
11 15 132 13 |
12 18 159 35 |
9. 7 ±1.76 12.9 ±2.9 111.8 ±36.5 20.2 ±7.04 |
||
|
Brood nests Total queens/ colony *Drone males |
25 251 |
30 269 |
23 234 |
34 283 |
41 377 |
35 291 |
24 245 |
21 216 |
19 192 |
47 403 |
28.32 ±9.2 257.7 ±67.2 N/A |
*Only 1 sample found in two separate nests with more than 250 adults in MPOB plantation in Saratok, Sarawak. N/A: Not available.
Figure 10.
Comparative abundance of reproductive virgin queens in young-older colonies.
4.8. Comparative Novel Intermediate Workers Behaviours
The two previously undescribed castes of intermediate workers do not function as nurses, as do minor workers, but rather exhibit the same behaviours as large major workers (i.e. defensive guard duty, aggressive, exploration of territories). They have been observed patrolling extensively with major workers along the weaver ant trails at the primary trunks, fronds, and rachis of palm trees (Figure 10). Figure 10 depicts an intermediate worker II (red circle) foraging alongside an intermediate worker III (yellow circle) on a palm tree frond beside large major workers IV (black circle) in Felda Gunung Besout plantations. These individuals were observed foraging on the ground and monitoring the trunk bases of palm trees. They also react aggressively by opening their mandibles widely and raising their gaster to a defiant 90-degree angle before assisting in the attack of intruders along larger major workers. However, it is unclear how precisely they are positioned in the first line of repulsion against alien elements in the vicinity of their territorial interaction zone. Their occurrence outside of colonies appears to be less frequent than that of larger-sized major workers and is correlated with varying daily weather parameters such as air temperature and atmospheric pressure in the absence of precipitation. However, the intermediates have been spotted acting on solitary foraging or by small group of two-three individuals, exploring far from their territorial boundary. Similarly, during the daily hot hours (average: 1200 - 1700 hours), major workers V and IV were frequently observed acting as supervisors to limit the activity of intermediate workers III and II far from nest canopies by bringing them back to nearby nests by holding them at the thorax baseline with their mandibles (Figure 11). It appears that during the hottest daily period (1100-1600 hours), the number of foragers of intermediate size that were permitted to be active was severely restricted, as was their ability to travel beyond nest perimeters. More study and observation are required to corroborate and comprehend their actual colony-level function and activity.
Various workers in daily activity on palm frond: Major worker IV (black circle); intermediate III (yellow circle); intermediate II (red circle). Photo Credit (Exélis Moïse Pierre).
The measurements shown by the lines were taken using a ZEISS Stereo Discovery V20 microscope. Photo Credit (Exélis Moïse Pierre).
Figure 10.
Major and intermediate workers collective daily foraging activity.

Figure 11.
Major worker caste V transporting intermediate worker caste II back to nest.

5. Discussion
5.1. O. smaragdina Colony Social Structure – Novel Intermediate Workers Caste
The establishment of a colony of Asian weaver ants is contingent on the successful mating of drone males and winged virgin females (potential future queens) during a nuptial flight swarm (Lokkers, 1990; Nielsen et al., 2015). Once inseminated, mated alate queens seek for a suitable curved leaf to lay the first egg batch in a "claustral" state and then shed their wings. This does correspond to a state of self-sufficiency by confinement, with the queen relying on her reserves to care for the brood until their emergence (Holldobler & Wilson, 1990; Nielsen et al., 2015; Nene et al., 2015). The first generation consists solely of major workers tasked with foraging and taking over all activities at the colony level, including care for the brood (Crozier et al., 2010). When the second egg group produces a mixture of major and minor workers, colony expansion commences with the completion of the nest. At this point, main workers use late instar larvae as a shuttle to seal the nest with silk (Holldobler & Wilson, 1990; Lim, 2007). Workers of the Asian weaver ant exhibit typical dimorphism, or a bimodal size-incidence distribution characterised by the major and minor workers (Lim, 2007; Schlüns et al., 2009; Crozier et al., 2010).
This study suggests the existence of a polymorphic reality by reporting for the first time the occurrence of a "polymodal size" distribution among five distinct worker castes comprised of two major, two intermediate, and minor individuals. Masram & Basargade (2021) did mention structural workers size differentiation and a more complex condition, but failed to provide unambiguous data on the existence of additional castes of Asian weaver ants. The study mentioned the existence of O. smaragdina polymorphs without distinguishing in depth distinct worker castes based on leg structure and sensilla size (Masram & Basargade, 2021) and instead provided a description of sensilla among all worker castes (Basargade et al., 2020).
This study presented the fundamental social colony structure composition of the Asian weaver ant in Malaysia, which is similar to previous studies (Holldobler & Wilson, 1990; Schlüns et al., 2009; Pimid et al., 2012) with the exception of two novel intermediate workers and larger-sized major workers that have never been reported. Pimid et al. (2012) reported O. smaragdina basic colony structure of throws the chain of mutation from eggs to immature stages (larvae, pupae) to adulthood emergence. It consisted of reproductive phase individuals (alate virgin queens, dealate shed wings founding queens and alate males) with non-reproductive ones (workers). However, Pimid et al. (2012) referred to the larvae as eggs, despite the fact that both characteristics are distinguishable (the photograph depicted only larvae, not eggs as stated in the manuscript), as shown in figure 5H and described in the results section. In accordance with Rwegasira et al. (2015) and Van Itterbeeck et al. (2014), the reproductive individuals are available throughout the year, particularly on rainy days when relative humidity is higher. It may be difficult to acquire adult male alate samples due to nest occupation segregation between males and the rest of the colony's members. Vanderplank (1960) described O. longinoda male inhabited different nests than the remainder of the colony members, which is consistent with our findings. However, it is not clear if this occurs just prior to the nuptial flight or if it is just their common natural behaviour. In spite of the difficulty of capturing adult male drones, we are able to capture their larvae. In this study, male weaver ants were only discovered in a peat soil oil palm plantation in Saratok, Sarawak. Due to the limited sample size of colonies (n=10) and the existence of split sex ratios in many ant species (Meunier et al., 2008; Kümmerli & Keller, 2009), this may be a coincidence. After sustained rainfalls, alate male lifespan exterminated immediately following the post-massive swarming nuptial flights during the mating period is suggested to be another potential factor (Nielsen et al., 2016a; Nene et al., 2015). It's conceivable that the sampled reproductive male nests with the abundance of green-winged queens represented an aggregation preceding the nuptial flights (Nielsen et al., 2015; Nene et al., 2016a; Nene et al., 2016b). Pimid et al. (2012) performed brood collection on O. smaragdina colonies occupying a smaller area (mango and pongame oil trees) on an urban university campus compare to a random sampling conducted in vast oil palm plantation zones. Compared to common palm tree plantations, the probability of sampling adult male nests in restricted occupied spaces from much shorter trees is greater. As suggested for other arboreal ant species by Powell et al. (2011) and for the Asian weaver ants by Rahim & Okahwara (2019), nest spatial distribution and configuration may also vary according to topography and tree phenology. In extensive oil palm plantations, therefore, it is very difficult to sample the nests of winged males. Our findings concur with those of Holldobler & Wilson (1990) and Hasegwa (2013), who reported that male production is scarcer in mature, larger colonies than in smaller colonies (Table 4). In a previous study, Vanderplank (1960) conducted dietary experiments on the African species O. longinoda, noting the appearance of large, intermediate, and small-sized workers in colonies fed a mixture of protein (insects) and honey diluted with water. However, there is no evidence that the type of diet correlates with workers' caste differentiation, with the exception of variations in aggressiveness and population density (Advento et al., 2022a). Despite the fact that Vanderplank (1960) did not provide morphometric data on these workers’ distinct intermediate categories, his observations are incorporated into the current study's findings. Consistent with all previous research, there are a greater number of major workers than minor workers (Verghese et al., 2013; Pimid et al., 2014; Pinkalski et al., 2015; Wargui et al., 2015). Foragers have a greater functional spectrum than the rest of the colony. They protect the entire colony perimeter as defensive elements (Holldobler & Wilson, 1990). They provide many veteran workers in permanent close protection of the founding queen, are responsible for brood transport and nest construction-repair, and search for daily food items (Chapuisat & Keller, 2002; Lim, 2007; Kahmi et al., 2015). According to reports, majors also assist drone males during the nuptial flight by escorting them back to the nest in the event of strong winds (Nene et al., 2016a). Minor workers serving as nurses are primarily confined to the vicinity of the nest, where they care for the nest broods (Kahmi et al., 2015). The existence of novel intermediate workers contradicts the concept of a bimodal size distribution in which major and minor worker sizes do not coincide (Holldobler & Wilson, 1990; Chapuisat & Keller, 2002). In incipient colonies, the first yield of gelatinous, sticky eggs and the first generation are susceptible to dehydration from heat (Nene et al., 2016). It is possible that the chambers found within the brood nests (Bochynek & Robson, 2014) are maintaining a cooling effect, thereby preventing the eggs from drying out. The purpose of these silken compartments is unknown, which may serve to separate reproductive forms from each other's castes. Nuptial flights and mating are preceded by days of rainfall and high humidity before, during, and after (Nene et al., 2016), resulting in much cooler air temperatures and a safer climatic environment (Nielsen et al., 2015). It is possible that the presence of chamber layers within the same brood structures contributes to some cooling effects. This is done to avoid the temperature threshold (defined as the highest temperature tolerance level that would inhibit the growth of any ectothermic organism) in order to maintain larval development (Villalta et al., 2020).
5.2. Novel Intermediate Workers Behaviours
It is evident from field observations and monitoring that intermediate workers also serve a purpose within the colony. Our study indicates that their behaviours are consistent with the concept of labour division in Oecophylla weaver ants (Bochynek & Robson, 2014). However, additional experimental analysis is required for a more thorough evaluation of this novel caste. While performing extensive foraging and reconnaissance activity far from nests along palm tree trunks and foliage, intermediate workers displayed territorial behaviours with less intensity in aggressive responses. In accordance with the division of labour among Oecophylla castes, this indicated a striking similarity between the attitude of the major workers and the minor workers. According to Holldobler and Wilson (1990), the significance of labour division determining each caste's specific duty at the colony level through synchronised cooperation among all individuals is a key factor in the colony's sustained survival. Another study analysing the biochemical secretions (i.e. formic acid, free amino acid, and acetylcholine) of all castes in India discovered that O. smaragdina colonies are polymorphic (Vidhu & Evans, 2015). Workers of Oecophylla smaragdina are divided into three distinct castes, namely major, intermediate, and minor, without defining their respective size (Vidhu & Evans, 2015). Beyond age or caste morphological differences, it will be necessary to determine the precise contribution of these novel, specialised catalyst individuals to the group's colony-level benefits (Robson & Traniello, 1999). Another essential element of the colony management system may be the exclusivity and unconventionality of these strategic individuals, who are physically distinct from the rest and who perform task allocation with creativity in contrast to the automatism-repetitive acts of their respective caste (Robson & Traniello, 1999). In ants, a caste is defined as a morphological distinction between individuals with specialised functions within the colony, thus referring to the concept of polymorphism (Wilson, 1984). In our study, however, the novel intermediate workers correspond more closely to a subdivided group or subcaste. Trible and Kronauer (2017) highlighted the exclusive significance of body size measurements for caste determination and distinction. The mechanism underlying the unexplored concept of caste distinction remains poorly explored. Caste is defined as being associated with the effect of tissue growth variations during the pre-pupae and earlier pupae life cycle development stages, establishing the size-frequency range variation of individual ants within a species (Trible & Kronauer, 2017). Observed caste foraging (collective or solitary) and scouting activity in the field by these new workers suggests a degree of autonomy in task allocation decision making. More observation is required to determine the precise contribution of this caste to the colony's overall benefits. In Indonesian oil palm plantations, the observed yellow larvae tended by Oecophylla workers within brood nests have been identified as Liphyra brassolis, an obligate myrmecophilous butterfly larva (Advento et al., 2022b).
6. Conclusion
This study provided a comprehensive evaluation of the social structure composition of O. smaragdina colonies, revealing a population similar to that reported in other studies. With the exception of the worker castes, which are comprised of extremely polymorphic individuals. The high fecundity of queens is supported by the abundance of eggs laid by thousands in brood nests, which is the first report of its kind for the O. smaragdina species. The population size of reproductive individuals can reach a dozen per nest and hundreds per colony. It is the first report to detect the existence of five distinct worker castes (major V-IV, intermediate III-II, and minor I) distinguished by size differences based on the head width, head, thorax, abdomen, and body lengths of the workers. The abdomen and body mass are visually idiosyncratic and are hypothesised to be predictors of this significant difference. The study's findings contradict the conventional exclusive concept of the bimodal size frequency distribution model for workers ant by demonstrating the existence of a clear multimodal distribution. The study shows the difficulty of adult male collection due to a short lifespan and their nests segregation. The results of the study indicated that environmental factors (rainfalls and relative humidity) play a crucial role in the emergence of reproductive individuals. The precise function of the intermediate workers' contribution to the survival of the colony requires additional evaluation. The findings of this study demonstrate that the biological system of the O. smaragdina species serves as a model for more diverse biological systems.
Data Availability Statement
The data that support the findings of this study are available and are included within the submitted documents: http://dx.doi.org/10.13140/RG.2.2.27717.12009
Acknowledgements
Special gratitude to the Malaysian Palm Oil Board (MPOB) and Felda Malaysia for granting access to their facilities and plantations. This research was supported by the Universiti Malaya through project numbers PS320-2010B and PV078-2011B awarded to EMP, and (Sustainability & Biodiversity) UMRG project numbers RG187/12SUS and RP016A-14-AFR awarded to EMP and AHI. EMP also received financing under a French full scholarship-grant from the local government authority "Collectivité Territoriale de la Martinique-CTM."
Competing Interests
The authors declare that they have no competing interests.
References
- Advento, A. D., Yusah, K. M., Naim, M., Caliman, J. P., & Fayle, T. M. (2022a). Which Protein Source is Best for Mass-Rearing of Asian Weaver Ants?. Journal of Tropical Biology & Conservation (JTBC), 19, 93-107. [CrossRef]
- Advento, A. D., Yusah, K., Salim, H., Naim, M., Caliman, J. P., & Fayle, T. (2022b). The first record of the parasitic myrmecophilous caterpillar Liphyra brassolis (Lepidoptera, Lycaenidae) inside Asian weaver ant (Oecophylla smaragdina) nests in oil palm plantations. Biodiversity Data Journal, 10, e83842. [CrossRef]
- AntWeb. Version 8.64.2. California Academy of Science, online at https://www.antweb.org. Accessed 22 September 2021.
- Azuma, N., Kikuchi, T., Ogata, K., & Higashi, S. (2002). Molecular phylogeny among local populations of weaver ant Oecophylla smaragdina. Zoological Science, 19(11), 1321-1328. [CrossRef]
- Azuma, N., Ogata, K., Kikuchi, T., & Higashi, S. (2006). Phylogeography of Asian weaver ants, Oecophylla smaragdina. Ecological Research, 21(1), 126-136. [CrossRef]
- Barsagade, D. D., Masram, P. P., & Nagarkar, D. A. (2020). Surface ultra-structural studies on antennae in polymorphs of weaver ant Oecophylla smaragdina; Fabricius (Hymenoptera: Formicidae) with reference to sensilla. Indian Journal of Agricultural Research, 54(5), 555-562. [CrossRef]
- Bochynek, T., & Robson, S. K. A. (2014). Physical and Biological Determinants of Collective Behavioural Dynamics in Complex Systems: Pulling Chain Formation in the Nest-Weaving Ant Oecophylla smaragdina. PLoS ONE, 9(4). [CrossRef]
- Chapuisat, M., & Keller, L. (2002). Division of labour influences the rate of ageing in weaver ant workers. Proceedings of Biological Sciences, 269(1494), 909–913. [CrossRef]
- Crozier, R. H., Schlüns, E. A., Robson, S. K. A., & Newey, P. S. (2010). A masterpiece of evolution Oecophylla weaver ants (Hymenoptera: Formicidae). Myrmecological News, 13, 57–71.
- Dejean, A., Corbara, B., Orivel, J., & Leponce, M. (2007). Rainforest canopy ants: the implications of territoriality and predatory behavior. Functional Ecosystems and Communities, 1(2), 105-120.
- Dejean, A., Ryder, S., Bolton, B., Compin, A., Leponce, M., Azémar, F.,... & Corbara, B. (2015). How territoriality and host-tree taxa determine the structure of ant mosaics. The Science of Nature, 102(5-6), 1-9. [CrossRef]
- Exélis, M. P & Azarae, H. I. (2013). Studies on the predatory activities of Oecophylla smaragdina (Hymenoptera: Formicidae) on Pteroma pendula (Lepidoptera: Psychidae) in oil palm plantations in Teluk Intan, Perak (Malaysia). Asian Myrmecology, 5, 163–176.
- Exélis, M. P. (2015). An ecological study of Pteroma pendula (Lepidoptera: Psychidae): in oil palm plantation with emphasis on the predatory activities of Oecophylla smaragdina (Hymenoptera: Formicidae) on the bagworm. 2013. Masters. Dissertation, Universiti of Malaya, Kuala Lumpur Malaysia.
- Exélis, M. P., Ramli, R., Ibrahim, R. W., & Idris, A. H. (2023). Foraging Behaviour and Population Dynamics of Asian Weaver Ants: Assessing Its Potential as Biological Control Agent of the Invasive Bagworms Metisa plana (Lepidoptera: Psychidae) in Oil Palm Plantations. Sustainability, 15(1), 780. [CrossRef]
- Greenslade, P. J. M. 1972. Comparative ecology of four tropical ant species. Insectes Sociaux, 19, 195–212. [CrossRef]
- Hölldobler, B. (1983). Territorial behavior in the green tree ant (Oecophylla smaragdina). Biotropica, 15(4), 241-250. [CrossRef]
- Hölldobler, B. (1998). Cooperation and Communication of Weaver Ants (Oecophylla). Jiussi News, 40.
- Hölldobler, B., & Wilson, E. O. (1990). The Ants. Belknap Press, Cambridge.
- Kamhi, J. F., Nunn, K., Robson, S. K. A., & Traniello, J. F. A. (2015). Polymorphism and division of labour in a socially complex ant: neuromodulation of aggression in the Australian weaver ant, Oecophylla smaragdina. Proceedings of the Royal Society B, 282(1811). [CrossRef]
- Kümmerli, R., & Keller, L. (2009). Patterns of split sex ratio in ants have multiple evolutionary causes based on different within-colony conflicts. Biology Letters 5, 713–716. [CrossRef]
- Lim, G. T. (2007). Enhancing the weaver ant, Oecophylla smaragdina (Hymenoptera: Formicidae), for biological control of a shoot borer, Hypsipyla robusta (Lepidoptera: Pyralidae), in Malaysian mahogany plantations. Virginia Tech University electronic Doctoral Dissertation. https://vtechworks.lib.vt.edu/handle/10919/26850.
- Lokkers, C. (1990). Colony dynamics of the green tree ant (Oecophylla smaragdina FAB.) in a seasonal tropical climate. – PhD thesis, James Cook University of North Queensland, Townsville, Queensland, 301.
- Mahima, K. V., Anand, P. P., Seena, S., Shameema, K & Manogem, E. M. (2021). Caste-specific phenotypic plasticity of Asian weaver ants: Revealing the allometric and non-allometric component of female caste system of Oecophylla smaragdina (Hymenoptera: Formicidae) by using geometric morphometrics. Sociobiology, 68(2), 5941. http://periodicos.uefs.br/index.php/sociobiology/article/view/5941/5904 . [CrossRef]
- Masram, P. P., & Barsagade D. D. (2021). Morphological and surface ultra-structural studies of legs in polymorphs of weaver ant Oecophylla smaragdina Fabricius (Hymenoptera: Formicidae) with reference to sensilla. International Journal of Zoological Investigations, 7, 26-39. [CrossRef]
- MATLAB and Statistics Toolbox. (2021). The Mathworks Inc. Natick, Massachusetts, USA.
- Meunier, J., West, S. A., & Chapuisat, M. (2008). Split sex ratios in the social Hymenoptera: a meta-analysis. Behavioral Ecology, 19(2), 382–390. [CrossRef]
- Nene, W., Rwegasira, G. M., Mwatawala, M., Nielsen, M. G., & Offenberg, J. (2016a). Foraging behavior and Preferences for Alternative Supplementary Feeds by the African Weaver Ant, Oecophylla longinoda Latreille (Hymenoptera, Formicidae). Journal of Hymenoptera Research, 50, 117-128. [CrossRef]
- Nene, W. A., Rwegasira, G. M., Nielsen, M. G., Mwatawala, M., & Offenberg, J. (2016b). Nuptial flights behavior of the African weaver ant, Oecophylla longinoda Latreille (Hymenoptera: Formicidae) and weather factors triggering flights. Insecte Sociaux, 63, 243–248. [CrossRef]
- Newey, P. S., Robson, S. K. A & Crozier, R. H. (2010). Weaver ants Oecophylla smaragdina encounter nasty neighbours rather than dear enemies. Ecology, 91(8), 2366-2372.
- Nielsen, M. G., Peng, R., Offenberg, J., & Birkmose, D. (2015). Mating strategy of Oecophylla smaragdina (Hymenoptera: Formicidae) in northern Australia. Austral Entomology, 55(3), 261–267. [CrossRef]
- Offenberg, J. (2015). Ants as tools in sustainable agriculture. Journal of Applied Ecology, 52(5), 1197–1205. [CrossRef]
- Peng, R., Christian, K., & Gibb, K. (1998). How many queens are there in mature colonies of the green ant, Oecophylla smaragdina (Fabricius)? Australian Journal of Entomology, 37(3), 249-253.
- Pfeiffer, M., Ho, C.T., & Lay, T. C. (2007). Exploring arboreal ant community and co-occurrence patterns in plantations of oil palms (Elaeis guineensis) in Borneo and Peninsular Malaysia. In structuring of animal communities: Interspecific interactions and habitat selection among ants and small mammals. Habilitationsschrift zur Erlangung der Venia Legendi an der Universität Ulm. Fakultät für Naturwissenschaften. http://www.antbase.net/pdf/pfeiffer-habil.pdf. [CrossRef]
- Pfeiffer, M., Ho, C. T., & Chong Lay, T. (2008). Exploring arboreal ant community composition and co-occurrence patterns in plantations of oil palm Elaeis guineensis in Borneo and Peninsular Malaysia. Ecography, 31(1), 21-32. [CrossRef]
- Pimid, M., Hassan, A., Tahir, N. A., & Thevan, K. (2012). Colony Structure of the Weaver Ant, Oecophylla smaragdina (Fabricius) (Hymenoptera: Formicidae). Sociobiology, 59(1), 1–10. [CrossRef]
- Pinkalski, C., Damgaard, C., Jensen, K. V, Gislum, R., Peng, R., & Offenberg, J. (2015). Non-destructive biomass estimation of Oecophylla smaragdina colonies: a model species for the ecological impact of ants. Insect Conservation and Diversity, 8, 464–473. [CrossRef]
- Powell, S., Costa, A. N., Lopes, C. T., & Vasconcelos, H. L. (2011). Canopy connectivity and the availability of diverse nesting resources affect species coexistence in arboreal ants. Journal of Animal Ecology, 80, 352-360. [CrossRef]
- Rahim, A & Ohkawara, K. (2019). Invasive Ants Affect Spatial Distribution Pattern and Diversity of Arboreal Ant Communities in Fruit Plantations, in Tarakan Island, Borneo. Sociobiology, 66(4), 527-535. [CrossRef]
- Robson, S. K., & Traniello, J. F. A. (1999). Key individuals and the organisation of labor in ants. In: Detrain C., Deneubourg J.L., Pasteels J.M. (eds) Information Processing in Social Insects. Birkhäuser, Basel, 239-259.
- Rwegasira, R. G., Mwatawala, M., Rwegasira, G. M., & Offenberg, J. (2015). Occurrence of sexuals of African weaver ant (Oecophylla longinoda Latreille) (Hymenoptera: Formicidae) under a bimodal rainfall pattern in eastern Tanzania. Bulletin of Entomological Research, 105(2), 182-186. [CrossRef]
- Schlüns, E. A., Wegener, B. J., Schlüns, H., Azuma, N., Robson, S. K. A., & Crozier, R. H. (2009). Breeding system, colony and population structure in the weaver ant Oecophylla smaragdina. Molecular Ecology, 18(1), 156-167.
- Trible, W., & Kronauer, D. J. C. (2017). Caste development and evolution in ants: it's all about size. Journal of Experimental Biology, 220(1), 53-62.
- Vanderplank, F. L. (1960). The Bionomics and Ecology of the Red Tree Ant, Oecophylla sp., and its Relationship to the Coconut Bug Pseudotheraptus wayi Brown (Coreidae). Journal of Animal Ecology, 29(1), 15-33. [CrossRef]
- Van Itterbeeck, J., Sivongxay, N., Praxaysombath, B., & van Huis, A. (2014). Indigenous knowledge of the edible weaver ant Oecophylla smaragdina Fabricius Hymenoptera: Formicidae from the Vientiane Plain, Lao PDR. Ethnobiology Letters, 5, 4-12. [CrossRef]
- Verghese, A., Jayanthi, P. D. K., Sreedevi, K., Devi, K. S., & Pinto, V. (2013). A quick and non-destructive population estimate for the weaver ant Oecophylla smaragdina Fab. (Hymenoptera: Formicidae). Current Science, 104, 641–646.
- Vidhu, V. V., & Evans, D. A. (2011). Identification of a third worker caste in the colony of Oecophylla smaragdina (Fabricius) based on morphology and content of total protein, free amino acids, formic acid and related enzymes. Entomon, 36(1/4), 205-212.
- Vidhu, V. V., & Evans, D. A. (2015). Importance of Formic Acid in Various Ethological States of Oecophylla smaragdina (Fabricius). In: A. K Chakravarthy Ed. New Horizons in Insect Science: Towards Sustainable Pest Management. Part II: Insect Physiology. Springer India, II, 53-60.
- Villalta, I., Oms, C. S., Angulo, E., Molinas-González, C. R., Devers, S., Cerdá, X., & Boulay, R. (2020). Does social thermal regulation constrain individual thermal tolerance in an ant species? Journal of Animal Ecology, 89(9), 2063-2076.
- Wargui, R., Offenberg, J., Sinzogan, A., Adandonon, A., Kossou, D., & Vayssières, J. F. (2015). Comparing different methods to assess weaver ant abundance in plantation trees. Asian Myrmecology, 7, 1–12.
- Wilson, E. O. (1984). The relation between caste ratios and division of labor in the ant genus Pheidole (Hymenoptera: Formicidae). Behavioral Ecology and Sociobiology, 16(1), 89-98. [CrossRef]
Figure 3.
A-E. Various castes of O. smaragdina: A- Gravid dealate founding queen with large gaster surrounded by major workers; B- Drone male (red circle) among major workers; C- Winged green virgin queens & yellow callow; D- Dealate gravid queen (red circle) & another wingless queen surrounded by major workers (yellow circle); Multiple winged queens (black circles), major workers-minor workers & larvae-pupae exhibiting a sharp white colour; E- Large major worker IV (Black circle); intermediate worker III (red circle); intermediate worker II (yellow circle) and minor worker I (blue circle). Photo Credit (Moïse Pierre Exélis).
Figure 3.
A-E. Various castes of O. smaragdina: A- Gravid dealate founding queen with large gaster surrounded by major workers; B- Drone male (red circle) among major workers; C- Winged green virgin queens & yellow callow; D- Dealate gravid queen (red circle) & another wingless queen surrounded by major workers (yellow circle); Multiple winged queens (black circles), major workers-minor workers & larvae-pupae exhibiting a sharp white colour; E- Large major worker IV (Black circle); intermediate worker III (red circle); intermediate worker II (yellow circle) and minor worker I (blue circle). Photo Credit (Moïse Pierre Exélis).

Table 1.
O. smaragdina eggs count estimation per nest.
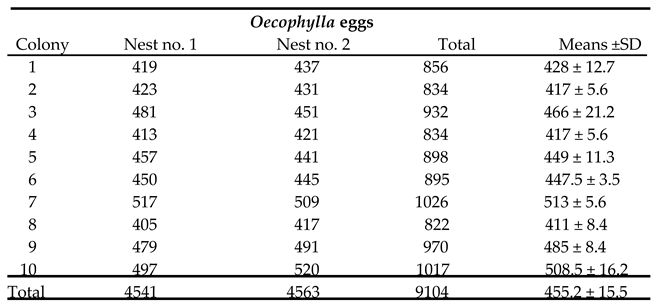
Table 2.
One-way ANOVA post-hoc Tukey HSD, Scheffé T-statistic test.
| Treatments | Nest 1 | Nest 2 | Pooled Total |
|---|---|---|---|
| Observations N | 10 | 10 | 20 |
| Sum | 4,541.0000 | 4,563.0000 | 9,104.0000 |
| Mean | 454.1000 | 456.3000 | 455.2000 |
| Sum of squares SS | 2,075,533.0000 | 2,094,329.0000 | 4,169,862.0000 |
| Sample variance S2 | 1,496.1000 | 1,359.1222 | 1,353.7474 |
| Sample std. dev. S | 38.6795 | 36.8663 | 36.7933 |
| std. dev. of mean SE | 12.2315 | 11.6581 | 8.2272 |
| SS | Df | Mean square MS |
F-statistic P value |
| 24.2000 | 1 | 24.2000 | F: 0.0170; P: 0.8979 |
| Nest 1 vs Nest 2 | aTukey HSD Q statistic | Tukey HSD P-value | Tukey HSD inference |
| N = 20 | 0.1841 | 0.8980410 | > 0.05 |
| Scheffé T-statistic | Scheffé p-value | Scheffé inference | |
| Nest 1 – Nest 2 pair | 0.1302 | 0.8978541 | > 0.05 |
aBonferroni and Holm tests gave same results, significant level, < 0.05.
Table 3.
Descriptive analysis of O. smaragdina Felda I colony.
| Variables | Nest one | Nest two | Nest three | Total nest | ||
|---|---|---|---|---|---|---|
| Min | Max | Mean ±SD | ||||
| Number of leaflets | 8 | 8 | 9 | 8 | 9 | 8.33 ± 0.5 |
| Nest length | 57.0 | 47.0 | 60.0 | 47.0 | 60.0 | 54.6 ± 11.5a |
| Nest width | 25.0 | 11.0 | 27.0 | 11.0 | 27.0 | 21.0 ± 7.0a |
| Nest height | 15.0 | 9.0 | 20.0 | 2.0 | 23.0 | 14.6 ± 8.5a |
| ***Height from the ground | 6.0 | 8.0 | 8.0 | 6.0 | 8.0 | 7.33 ± 1.15 |
| Total workers | 14279 | 4985 | 16623 | 4985 | 16623 | 11962 ±6155 |
| Number of major workers Number of intermediate workers** |
5753 4175 |
2009 1458 |
6379 5101 |
2009 1458 |
5753 5101 |
4713 ±2363 3578 ±1893 |
| Number of minor workers | 4351 | 3721 | 5143 | 3721 | 5143 | 1387 ± 1098a |
| Number of winged green queens Number of newly emerged queens* |
17 5 |
15 9 |
19 11 |
15 5 |
19 11 |
17 ± 2 8.33 ±3 |
| Number of drone males : 0 Number of worker pupae |
1255 | 342 | 357 | 15 | 2038 | 878 ± 792a |
| Number of larvae | 854 | 427 | 1375 | 427 | 1375 | 885.3 ±474.7 |
| Egg volume | 3.1 | 4.5 | 15.2 | 3.1 | 15.2 | 7.6 ± 1.6a |
| Queen larvae | 21 | 15 | 0 | 0 | 21 | 12 ±10.8 |
| Male larvae | 37 | 29 | 0 | 0 | 37 | 22 ±19.4 |
| Eggs count | 1874 | 4890 | 0 | 0 | 4890 | 2254 ±2515 |
Note: aVariable with the different superscript alphabet had significantly different mean value at p < 0.05 *Callow yellow queens becoming green after 7 days - **novel workers caste - ***Barrack nest range 2-4 m height to the ground.
Table 4.
Descriptive analysis of O. smaragdina matured big size colony UM III.
| Variables | Colony (3 nests) | ||
|---|---|---|---|
| Min | Max | Mean ±SD | |
| Number of leaflets | 6 | 19 | 13.33 ± 4a |
| Nest length | 30.0 | 60.0 | 49.0 ± 12.3a |
| Nest width | 8.0 | 30.0 | 17.0 ± 7.0a |
| Nest height | 2.0 | 23.0 | 10.4 ± 6.7a |
| Nest volume | 829.4 | 16618.5 | 5889.1 ± 6347.9a |
| Height from the ground | 9.0 | 10.0 | 9.6 ± 0.5a |
| Total workers | 2448 | 64761 | 16432 ± 20896a |
| Number of major workers Number of intermediate workers |
1205 201 |
40873 19102 |
15045 ± 19828a 6723.3 ±20170 |
| Number of minor workers | 376 | 4786 | 1387 ± 1098a |
| Number of winged queens* Number of newly emerged queens** Number of males1 |
0 10 0 |
138 33 0 |
64.6 ± 63.9b 21.3 ±11.5 0 |
| Number of worker pupae Queen larvae Male larvae |
154 0 0 |
2038 37 49 |
918 ± 991a 22 ±19.4 29.6 ±26 |
| Number of workers larvae | 71 | 6509 | 2821.6 ± 3319.6a |
| Egg volume Eggs count |
0.0 0 |
4.7 3845 |
1.0 ± 1.6a 1505.6 ± 2053.6 |
Note: *Green colour gradually reaching a week period of development; ** Emergence as yellow pale colour changing gradually to green upon high water consumption; aVariables with the different superscript alphabet had significantly different mean value at p < 0.05. 1An average 250 drone reproductive males were found from two big nests in Saratok, Sarawak Borneo MPOB. 1 An estimation of the population size of Felda I is: PSFelda I = 33 nests x 11,962.00 = 394,746.00 ± 159,647and UM III, PSUMIII = 74 x 64,761.00 = 4,792,314.00 = 4.8 106 ± 525,136 representing an average of major-intermediate-minor total workers.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



