Submitted:
29 October 2025
Posted:
31 October 2025
Read the latest preprint version here
Abstract
What mechanisms drive and shape the stepwise evolution from simple carbon-based materials (CBMs) into complex organisms and societies? This fundamental question spans life's origin, biological diversification, and societal development, yet remains unresolved because it has been often studied in isolation. Here, through cross-disciplinary integrative reasoning, empirical validation, and mathematical modeling, we extend Darwinian theory to formulate the Carbon-Based Evolutionary Theory (CBET)—the first explicit and holistic framework that unifies chemical, biological, and social evolution. CBET highlights energy flow, functions of CBMs, environmental influence, and mathematical rules. CBET derives three axioms and two key mechanisms—spirodynamic feedback and natural selection—which respectively drive and shape the stepwise evolution from carbon atoms to societies. CBET explains how CBMs diversify, elevate hierarchies, increase fitness, and gain orderliness and complexity. Furthermore, CBET refines fitness calculation and clarifies widespread misunderstandings about evolution. It elucidates the natural balance between competition and inclusiveness, selfishness and altruism, and individual versus collective interests, thereby offering a scientific foundation for fostering the balanced evolution of human societies.
Keywords:
carbon-based material
; mechanism
; chemical evolution
; biological evolution
; social evolution
; natural selection
; theory
Why have simple carbon-based materials (CBMs)—such as carbon atoms, methane, and carbon dioxide—evolved stepwise into the complex, diverse, and ordered organisms and societies we observe today? This fundamental question has fascinated scholars for millennia but remains unresolved [1,2,3,4,5,6,7,8]. It involves the chemical evolution of organic molecules [5], the biological evolution of organisms, and the social evolution of animal and human societies [7,8]. Traditionally, these three domains have been studied in isolation [1,2,3,4,5,6,7,8], and thus theories developed in one domain often fail to apply to others. For instance, while Darwinian theory provides a robust account for biological evolution, it does not address chemical or social evolution. Consequently, it offers neither an explanation for life’s origin nor rational suggestions for social development [1,2,3,4]. To bridge these gaps, through cross-disciplinary integrative reasoning and robust empirical validation, we propose the Carbon-Based Evolutionary Theory (CBET)—an extension of Darwinian theory designed to unveil the mechanisms underlying chemical, biological, and social evolution within a coherent framework.
1. Results
1.1. Hierarchies and Functions of CBMs
The central concept of ‘organisms’ in Darwinian theory is extended into the more inclusive ‘CBMs’ in CBET. As shown in Figure 1, CBET classifies nine hierarchies of CBMs (H1- to H9-CBMs), according to the criterion that lower-hierarchy CBMs can be building blocks for higher-hierarchy ones. H1 refers to carbon atoms, which likely originated in the interiors of giant or supergiant stars9. H2 and H3 represent small carbon-containing molecules—i.e., CCMs (e.g., CH₄, CO₂) and medium CCMs (e.g., glucose, lysine), respectively. Polymerization of some H2 and H3 molecules can form H4—large CCMs such as proteins and nucleic acids—which can assemble into H5, namely multi-molecule aggregates (MMAs; e.g., phospholipid bilayers). The transition of CBMs to life occurs at H6 (cells) and culminates in H7 (multicellular organisms). Some H7 individuals form social organizations, with H8 representing single-level societies (e.g., ant colonies, small companies) and H9 comprising multi-level societies (e.g., universities, armies, nations). Owing to their extensive internal collaboration, H8 and H9 entities are often regarded as superorganisms [7]. Boundaries between hierarchies are not rigid: transitional forms such as peptides (between H3 and H4) and viruses (between H4 and H5) can act as bridges facilitating the hierarchy elevation of CBMs.
CBET clarifies the following functions of CBMs crucial to CBM evolution.
(1) Macromolecular backbone function. Carbon is unique in its ability to form long, stable chains and complex rings through relatively strong, versatile C-C bonds, which can link to various atoms (such as hydrogen, oxygen, nitrogen, sulfur, and halogens) and functional groups (such as hydroxyl, carbonyl, amino, and carboxyl groups), giving rise to immense molecular diversity. In contrast, other elements lack the ability to form extended chains or networks through self-bonding, except that silicon and boron can form much weaker and shorter covalent chains (e.g., Si-Si, B-B) that are prone to cleavage. Therefore, only carbon atoms can serve as the backbones of myriad stable macromolecules [10]. This fundamental disparity explains why CBMs, not silicon or boron-based ones, have naturally evolved into life.
(2) Hierarchy elevation function. CBMs across H1–H9 can form higher-hierarchy CBMs through physical, chemical, biological, and/or social interactions. For example, CO₂ can participate in amino-acid formation; amino acids can form proteins; individual ants can form ant colonies; and tribes can form alliances.
(3) Relative stability function. Many CBMs are relatively stable. For instance, various H3-CBMs could persist for billions of years in meteorites [11]; some proteins and nucleic acids were preserved in fossils for millions of years [12]; humans typically live for decades; and some companies have existed for years.
(4) Changeability function. The repeated formation and degradation of higher-hierarchy CBMs drive Earth’s carbon cycles [13], and many CBMs produced in these cycles exhibit variations relative to their progenitors.
(5) Information function. H6-CBMs and H7-CBMs can encode, read, translate, change, store, and transmit genetic information through nucleic-acid–protein interactions [2]. They also use diverse signaling pathways to respond to internal and external changes. Some H8-CBMs and H9-CBMs harness additional information forms—such as scents, sounds, behaviors, scripts, and/or electromagnetic waves.
(6) Other functions. The energy-storage and -release functions of H3- and H4-CBMs, the catalytic functions of many H4-CBMs, the reproduction of H6-CBMs, and the knowledge accumulation function of humans (H7) are all crucial to CBM evolution.
1.2. Three Axioms
CBET is founded on three explicit axioms that have long guided research in physics, chemistry, biology, and social sciences. These axioms are derived from principles across diverse disciplines by abstracting away their quantitative parameters to render them applicable to all molecules, organisms, and societies.
(1) The Environment–Influence Axiom. The environment supplies energy, constituent materials, and conditions that influence structural changes in a target material. These structural changes, in turn, can alter the environment. For example, Earth’s habitable environment fosters CBM evolution [14,15], which in turn profoundly transforms Earth’s environment.
(2) The Structure–Function Axiom. A material’s structure partly determines its functions, and structural changes can generate new functions. Concurrently, functions are the internal causes for maintaining or modifying structures. For example, a cell’s structure dictates its metabolic and reproductive abilities, which in turn are essential for sustaining and replicating that structure. This axiom suggests that a complex function—if defined as one requiring multiple ordered and distinct steps (e.g., a car’s operation and a cell’s reproduction)—typically necessitates a complex structure, as each step requires a specialized structural component, and these components must be coordinated in space and time to ensure orderly execution of the steps, collectively constituting a complex structure.
(3) The Driving–Force Axiom. Energy flow drives structural changes (e.g., protein synthesis) and locational changes (e.g., bird flying) under the modulation of functions of the target material (internal causes) and the environment (external causes). This axiom highlights energy flow as the universal driver of structural and locational changes across all CBMs, from carbon atoms to societies. It is derived from and supported by the two axioms above and principles from Newtonian mechanics and organic chemistry, as well as evidence from animal locomotion and social revolutions.
1.3. Reasoning Steps of CBET
CBET was derived through eight steps of integrative reasoning (Figure 2).
Step1. Reasoning the three axioms
This is given in Section 1.2 above.
Steps 2−5. Reasoning spirodynamic feedback from the three axioms
Step 2. According to the Driving–Force Axiom, energy flow under the modulation of functions of CBMs and the environment drives structural or locational changes in CBMs.
Step 3. Structural changes in CBMs (potentially combined with other factors such as environmental shifts) can generate novel functions. These new functions can feed back to Step 2 and thus initiate further structural changes in CBMs. An example is the formation of nucleotides, which enables their polymerization into nucleic acids—a novel function that emerges when nucleotides are formed.
Step 4. Structural or locational changes from Step 2 can cause environmental changes. Other factors like asteroid impacts can also cause environmental changes. Environmental changes can feed back to Step 2 and thus initiate further structural or locational changes in CBMs. For instance, the atmospheric oxygenation by photosynthetic bacteria ~2.5 billion years ago selected against anaerobes in favor of aerobes [2,16].
Step 5. The iterative feedback loops between the changes in structures or locations (Step 2), functions (Step 3), and environments (Step 4) constitute the spirodynamic feedback mechanism. This mechanism—integrating the foundational concept of feedback loops from systems theory and cybernetics with the three axioms given in Section 1.2—can amplify CBM diversity and elevate CBM hierarchies within Earth's habitable environment.
Step 6. Reasoning natural selection
Natural selection shapes the CBM changes generated through spirodynamic feedback (Steps 2−5) governed by the following two self-evident mathematical rules.
(1) The Quantity-Existence Rule. A material exists if its cumulative formation exceeds its cumulative degradation. This applies to the synthesis/breakdown of molecules, birth/death of organisms, and establishment/dissolution of social organizations.
(2) The Quantity-Change Rule. The change in a material's existing quantity over a time interval [a,b] is determined by its initial quantity and the ratio of its total formation () to its total degradation () within that period. This ratio denotes a material’s overall performance in formation and sustenance (OPFS), which equals its fitness during that interval (Equation 1).
In modern biology, fitness is defined as the number of offspring an individual leaves in the next generation [2]. This measure fails to capture the fitness of molecules, social organizations, or worker bees, and it neglects generation time. The universal fitness calculation in Equation 1 overcomes these limitations.
According to the two rules above, a change that increases fitness facilitates its accumulation, and a change that decreases fitness facilitates its elimination. This differential retention—based solely on the effect of changes on fitness—constitutes natural selection in CBET, which non-randomly shapes CBM evolution and increases CBM’s fitness across all hierarchies. In accordance with the Structure–Function Axiom, fitness improvements are usually accompanied by greater structural orderliness and internal collaboration.
Step 7. Complexity enhancement and its feedback
The hierarchical elevation in Step 5 and internal collaboration enhancement in Step 6 collectively lead to an increase in the structural complexity of CBMs. This heightened structural complexity itself becomes an input into the spirodynamic feedback and natural selection mechanisms. This can sometimes intensify the pressure of natural selection—a phenomenon vividly illustrated by the co-evolution between predators and prey, parasites and hosts, alongside plants and herbivores [17]. For instance, structural adaptations that increase the speed of gazelles exert stronger selection pressure for corresponding enhancements in the speed of their predators (e.g., cheetahs). The subsequent evolution of faster predators, in turn, exerts even greater selective pressure on the gazelle population.
Step 8. Synergy of the reasoning steps
The seven reasoning steps above demonstrate how CBET integrates three axioms, two mathematical rules, CBM functions, and environmental influences to formulate two key mechanisms that collectively drive and shape the stepwise evolution of CBMs from simple atoms into the complex, diverse, and ordered organisms and social organizations.
Comparison with Darwinian theory
Darwinian theory aligns primarily with Step 6 of CBET and derives natural selection from over-reproduction, environmental limitations, struggle for survival, and inheritable changes (Figure 2B). It clarifies neither the mathematical rules of natural selection nor the mechanism underlying the hierarchy elevation and diversification of CBMs.
1.4. Explanation of Life's Origin
Life's origin is the most enigmatic event in CBM evolution, and CBET offers a coherent framework to explain its plausibility, as detailed below.
(1) Driving role of spirodynamic feedback. Life's origin was driven by the mechanism of spirodynamic feedback through the stepwise hierarchy elevation of CBMs from H1 to H6 on Earth. This mechanism involves energy flow (which is modulated by functions of CBMs and the environment) and feedback loops among changes in structures, functions, and environments of CBMs (Figure 2). In chemistry and thermodynamics, certain H2-CBMs can absorb energy spontaneously from energy flow sources on Earth, such as sunlight, radiation, or geotherm, and thus react to form diverse H3-CBMs through organic chemical reactions under specific environmental conditions [3,5,6,15]. Likewise, some of the resulting H3-CBMs can further absorb energy spontaneously from energy flow sources and thus react to form H4-CBMs through organic chemical reactions under specific environmental conditions (e.g., presence of catalysts) [5,6,15]. This stepwise process establishes a natural energy-driven trend toward the gradual accumulation and increasing complexity of organic molecules. In astronomy and geology, this process could have persisted on Earth for hundreds of millions of years before life's origin [5,6,15], and thus would have accumulated myriad macromolecules (H4-CBMs). In physics, the energy flows—such as those from wind, water flow, and wet–dry cycles—can drive those macromolecules into innumerable MMAs (H5-CBMs) and MMA clusters. Concurrently, functions of relevant CBMs, such as their interaction through hydrogen bonds, also influence the formation of MMA clusters. Due to their diverse complex structures, some MMA clusters could have acquired various catalytic abilities, facilitating the synthesis or degradation of amino acids, nucleotides, glucose, proteins, nucleic acids, and other organic molecules—accelerating the formation-degradation cycles of H3-CBMs, H4-CBMs, H5-CBMs and MMA clusters. Through stochastic processes, an exceedingly rare fraction of these clusters eventually achieved the complex function of reproduction, becoming the first life structures (H6-CBMs), namely those MMA structures with the reproduction function.
(2) Shaping role of natural selection before life's origin. Prior to life's origin, natural selection preferentially accumulated those H3-CBMs, H4-CBMs, H5-CBMs, and MMA clusters with a higher formation-to-degradation ratio (i.e., higher fitness) during their formation-degradation cycles. Generally, the fitness ranks as follows: unstable nucleic acid A < stable nucleic acid B formed without catalysis < stable nucleic acid C formed through catalysis < stable nucleic acid D replicated through catalysis by certain proteins (e.g., the RNA polymerases of coronaviruses and other RNA viruses). Nucleic acid D was thus favored by natural selection for life's origin. Likewise, stable proteins replicated through catalysis by certain MMA clusters—such as ribosomes18—are also favored by natural selection for life's origin. Some of these stable nucleic acids and proteins, in turn, could be the enzymes or constitute the MMA clusters to catalyze the replicative formation of these stable nucleic acids and proteins. Thus, natural selection can thus shape the evolution or the formation-degradation cycles of MMA clusters towards life's origin.
(3) Shaping role of natural selection after life's origin. The reproduction function provided a decisive advantage of life structures to increase their prevalence, as the function could facilitate the formation of life structures to consistently exceed their degradation. In contrast, no such sustaining mechanism exists for non-replicating MMA clusters to increase their quantities. Moreover, life structures have the changeability function, and consequently, myriad variants of life structures could have emerged on Earth over billions of years, and those variants with mutations that enhance their fitness are favored by natural selection, making life structures increasingly advantageous over non-reproductive MMA clusters. This could have led to the prosperity of life structures and mass extinction of non-reproductive MMA clusters on Earth, along with their constituent H2-CBMs, H3-CBMs, and H4-CBMs.
(4) Unique capacity of CBMs and Earth's habitability. While the three axioms and two mechanisms in CBET could be extended from CBMs to many other materials and allow their hierarchy elevation and fitness enhancement, only CBMs possess the function to reach and surpass the hierarchy of diverse and stable macromolecules, as explained in Section 1.1. Moreover, according to CBET, CBMs can form such complex structures only in a suitable environment like Earth, which provides abundant energy, ample H2-CBMs, plentiful liquid water, and other necessary conditions [14,15]. Previously, it was hypothesized that life originated in hydrothermal vents, or volcanic lakes [2,4]. Section 3.4.2.1.2 below will explain why it was much more likely for life to emerge in Earth’s early atmosphere than other spaces.
(5) Probabilistic nature and intermediate steps. Due to the extraordinary complexity of life structures—particularly exemplified by systems like the ribosome, which produces proteins according to the sequences of nucleic acids [18]—CBET posits that the probability of an inanimate MMA cluster transitioning into a life structure, while greater than zero, is exceedingly low. Consequently, some intermediate steps were likely essential for life's origin. A plausible sequence of these intermediate steps might involve transitions such as H4-CBMs → [small H4-CBMs → middle H4-CBMs → large H4-CBMs] → MMAs → MMA clusters → metabolic MMA clusters → life structures capable of reproduction. This postulated pathway, while consistent with CBET, is not a component of CBET and should be validated by novel studies.
Together, CBET provides a coherent framework rather than details to explain the plausibility of life's origin, and meanwhile, the CBET framework can integrate knowledge across diverse disciplines to explain life's origin in more detail. Based on this framework, Section 3.4.2 below will employ mathematical modeling, along with relevant assumptions and publications [19,20,21,22,23,24], to further analyze the plausibility of life's origin.
1.5. Clarification of Misunderstandings
The CBET framework can be employed to clarify the following five common misunderstandings about natural selection. These misunderstandings are not flaws in Darwinian theory but rather in the common interpretations.
(1) Misunderstanding about 'Survival of the fittest'. Darwin adopted this phrase as shorthand for natural selection, but it is often misread literally to mean that only the fittest can survive1,4. According to Equation 1 in CBET, a bacterial mutant with a detrimental mutation can increase in population size, even if its growth rate (e.g., 0.30% per day) is lower than that of its wild-type counterpart (e.g., 0.05% per day). This illustrates that 'survival of the fit enough' is more literally accurate than 'survival of the fittest.' Likewise, major evolutionary transitions—such as multicellularity, endothermy, and eusociality—could have arisen from pioneers that were not fitter than their progenitors or competitors but were fit enough to establish new lineages. Additionally, a CBM’s OPFS (fitness) reflects the combined contribution of all its traits and hierarchies, while specific key traits (e.g., antelope speed, wolf cooperation, human intelligence) can disproportionately influence OPFS (fitness). Thus, natural selection shapes both all-around and specialized adaptation.
(2) Misunderstanding that all traits are advantageous. The example above shows that neutral or even detrimental traits can persist, provided the relevant CBM is fit enough. This is also evidenced by human infants’ vulnerability, the slow running speed of African buffalo, and the prevalence of neutral mutations in genomes [2]. Furthermore, traits may be advantageous in some contexts but detrimental in others. For instance, giraffes’ considerable height aids browsing and combat but hinders rising from the ground [25].
(3) Misunderstanding that natural selection is purely competitive. As elucidated above, natural selection is also inclusive as it can accommodate advantageous, neutral, and even detrimental traits, provided the relevant CBM is fit enough. This inclusiveness is also evidenced by the long-term increase in biodiversity throughout Earth's history [2], though catastrophes such as asteroid impacts occasionally trigger sharp declines in biodiversity [2].
(4) Misunderstanding that natural selection equals selfishness. It is true that selfish behavior is critical for acquiring energy, resources, favorable environments, and mating opportunities [1,26]. Yet CBET shows that selfishness and altruism coexist and balance throughout evolution. Diverse molecules often serve as catalysts, energy sources, or building blocks for other molecules [4]; diverse macromolecules in cells enable genetic transmission of genomic DNA; somatic cells in multicellular organisms support the passage of reproductive cells; ant workers sacrifice for colony reproduction [7]; and humans in armies or police may die for society. According to the Driving-Force Axiom, these altruistic behaviors above are all driven by energy flow modulated by the functions of the relevant CBMs and the environment.
(5) Misunderstanding that only heritable changes matter. In contrast to the classical view that natural selection acts solely on heritable genetic changes [1,2], CBET recognizes the biological fact that non-heritable factors (e.g., vaccination, education) and epigenetic changes [27], whether inherited or not, influence survival and reproduction. Thus, these factors are directly under natural selection and play crucial roles in shaping evolutionary outcomes.
1.6. Validation of CBET
As detailed in Section 3.1 below, the spirodynamic feedback mechanism was tested against 27 representative facts randomly selected from diverse hierarchical levels of CBMs. These facts—ranging from charcoal combustion and DNA synthesis to the formation of basketball teams and international conflicts—systematically support the mechanism.
A survey of 35 undergraduate students from Foshan University unanimously affirmed the foundational premises of the three axioms underlying spirodynamic feedback. Furthermore, three AI tools (DeepSeek, Doubao, ChatGLM) and five professors from physics, biology, chemistry, and geology were invited to falsify the axioms and reasoning of CBET; none identified robust counterevidence.
The natural selection mechanism was validated through its consistency with established biological phenomena [2], organic chemical reactions where molecules with higher formation-to-degradation ratios accumulate preferentially, and social dynamics, as seen in the competition among companies.
1.7. Mathematical Formalization and Modeling
Mathematical formulation and modeling of CBET were conducted to support the CBET framework in formal logic (see Methods for details). The mathematical formulation translated the three axioms and core mechanisms into a set of universal equations, serving to formally demonstrate the internal consistency of the theory. In parallel, the mathematical modeling employed certain quantitative assumptions to visually demonstrate the dynamics of natural selection across diverse scenarios and to explicitly evaluate the plausibility of life’s origin through abiogenesis or chemical evolution.
2. Discussion
2.1. Significance in Natural Sciences
In natural sciences, CBET offers a novel, explicit, and holistic explanation for the origins and evolution of life and societies. It potentially stands as the first theory to unify chemical, biological, and social evolution within a single coherent framework, which has been desired for centuries in evolutionary studies [2,3,4,8,28]. CBET effectively clarifies five common misunderstandings and the intricate relationships among six fundamental concepts, including energy, structures, functions, information, orderliness, and complexity [29]. Furthermore, CBET clarifies the mathematical rules governing natural selection, refines fitness calculation, and integrates multiple facts (e.g., the prevalence of neutral mutations) that have not been integrated by Darwinian theory in biology (Table 1). CBET elucidates the prevalence of altruistic behavior and the importance of epigenetic and non-heritable changes in CBM evolution. It also elucidates that organisms can obtain some complex functions, such as nonrandom mutations and epigenetic changes, which stem from their complex inner structures [27,30].
2.2. Significance in Social Sciences
In social sciences, CBET offers an innovative explanation for the evolution of societies from molecules and individual organisms. It reveals that changes in CBMs—from organic molecules to societies—are governed by the same axioms, mathematical rules, and mechanisms of spirodynamic feedback and natural selection. Furthermore, CBET illuminates the persistent natural balance between seemingly opposing forces, such as self-functioning and environmental influence, competition and inclusiveness, and individual versus collective interests. It thus helps resolve long-standing debates in sociology, ethics, and political philosophy while providing a foundation for fostering balanced and harmonious societal development. Additionally, the spirodynamic feedback mechanism underscores the importance of considering the systemic and long-term effects of policies. Collectively, CBET establishes an ontological foundation for social sciences that is grounded in natural sciences.
Notably, within the CBET framework, the core concepts of energy, function, structure, and environment exhibit substantially increasing complexity as hierarchies rise. Consequently, CBET seamlessly incorporates non-material social factors—such as culture, laws, and institutions—because they are emergent functions of societies, which can influence and be influenced by social changes. Thus, CBET does not simplistically reduce complex social phenomena to mere biological or chemical terms.
2.3. Limitations of CBET
Both Darwinian theory and CBET only provide a schematic (not detailed) account for the plausibility (not inevitability) of evolution mostly in a qualitative (not quantitative) manner. These three limitations have lowered the threshold for establishing the two theories while enhancing their credibility in accounting for the evolution of all organisms or all CBMs. For example, CBET would be uncredible if it described protein synthesis and the emergence dynamics of companies using the same qualitative equation.
2.4. Outlook
The CBET framework opens new avenues for cross-disciplinary reflection. It suggests that in physics, energy acts as both the 'architect' and 'disruptor' of structures, while structures serve as reservoirs, stabilizers, converters, and regulators of energy, with structural complexity dictating the pathways and efficiency of energy storage and transformation. In chemistry, CBET predicts—based on the Structure–Function Axiom—that self-replicating macromolecules, long sought in prebiotic studies [31], may be exceedingly rare or even non-existent, as such a complex function typically requires a structural system far more sophisticated than a single molecule, and the function of biological reproduction is a novel function resulting from molecular interactions rather than the function of self-replication of certain macromolecules. In biology, CBET anticipates the persistent prevalence of deleterious mutations in humans and other organisms, provided their carriers remain 'fit enough' overall. CBET also suggests that some organisms (e.g., rabbits in Australia) can consume more energy than their ancestors, also provided they remain 'fit enough' overall. In social sciences, CBET predicts overarching trends of global integration and balanced governance, driven by spirodynamic feedback and shaped by natural selection.
3. Methods
3.1. Reasoning, Validation, and Refinement of CBET
3.1.1. Methodology of CBET
Darwinian theory was initially deduced by Darwin and Wallace through the integration of the insights from diverse fields such as Malthusian demography (the struggle for existence), Lyellian geology (deep time), artificial selection (the power of selective breeding), biogeography (patterns of species distribution)2. Later, this theory has been empirically validated by more observations and scientific advancements (e.g., those in genetics and developmental biology) and refined thereby (Figure 3). CBET extends this methodology through the integration of modern knowledge from physics, chemistry, biology, and social sciences to derive its core mechanisms(namely spirodynamic feedback and natural selection) (Figure 3). This is followed by empirical validation and iterative refinements. CBET is further reinforced with mathematical formalization and modeling (See Section 3.4 below).
3.1.2. Integrative Reasoning of CBET
– Selection of CBMs as CBET’s core concept
- ✧
- Like the core concept of ‘organism’ in biological evolution, carbon-based material (CBM)’ is an explicit and precise core concept for the study of the entire CBM evolution.
- ✧
- Other concepts, such as ‘molecule’, ‘system’, ‘material’, ‘gene’, or ‘entropy’, are less suited to unify carbon atoms, organic molecules, organisms, and societies. They are therefore less suited to be the core concept for the study of the entire CBM evolution.
– Integration of various key factors
- ✧
- CBET integrates various key factors, such as hierarchy, energy, structures, functions, environmental factors, feedback, natural selection, competition, collaboration, water, genes, epigenetics, neutral mutations, Earth’s features, and carbon atomic structure.
- ✧
- The factors above are concrete and known to be crucial for CBM evolution and directly associated with CBM evolution. Thus, the selection of these key factors is important for CBET to provide a direct and explicit explanation for CBM evolution.
– Eight integrative reasoning steps
CBET was derived through eight reasoning steps of integrative reasoning, as detailed in the main text and outlined in Figure 2.
– Circumvention of circular reasoning
CBET has derived the spirodynamic feedback mechanism, which is grounded in the three universal axioms derived from extensive empirical facts, to address the availability of the options of natural selection (i.e., structural changes of CBMs). CBET has also clarified the mathematical rules of natural selection. Thus, CBET circumvents circular reasoning (i.e., tautology).
In contrast, Darwinian theory has not clarified the availability of the options of natural selection (i.e., heritable changes), a gap that has long drawn criticism for its potential to lapse into tautology [32].
3.1.3. Empirical Validation of Spirodynamic Feedback
– Empirical validation against 27 representative facts
The validity of the spirodynamic feedback mechanism was tested against 27 representative randomly selected facts spanning molecular to social phenomena (Table 2). These 27 facts collectively and systematically validate the spirodynamic feedback mechanism, as the changes they embody consistently exhibit the following characteristics:
- They require energy flow, which may originate from the environment, the target material itself, or their interactions;
- They are influenced by environmental factors;
- They rely on the functions of the relevant material, which are in turn partially determined by its structure;
- They demonstrate that structural changes can generate new functions not present in the original forms. These emergent functions can initiate further structural changes, thereby establishing a spirodynamic feedback loop that, in some cases, leads to serial hierarchical elevation—as illustrated by Facts 1, 4, 7, 10, 13, 16, 19, 22, and 25;
- They show that structural changes can influence or alter the environment, which may also be modified by other factors. Such environmental changes can, in turn, contribute to subsequent rounds of structural changes or further engage in the spirodynamic feedback process—as evidenced by Facts 1, 3, 5, 18, 20, and 27.
– Survey of undergraduate students
Undergraduate students (n=35) randomly selected from Foshan University were invited to answer the following four questions.
- ✧
- Question 1: Do you think all structural changes in molecules, organisms, or social organizations are driven by or rely on energy flow?
- ✧
- Question 2: Do you think many structural changes in molecules, organisms, or social organizations are affected by the environment?
- ✧
- Question 3: Do you think some functions of molecules, organisms, or social organizations are important to maintain or change the structures of themselves?
- ✧
- Question 4: Do you think some functions of molecules, organisms, or social organizations are partially determined by their structures?
All the 35 students answered the four questions above with ‘Yes’.
– Falsification by AI tools and scholars
The three AI tools of DeepSeek (https://chat.deepseek.com/), Doubao (https://www.doubao.com/), and ChatGLM (https://chatglm.cn) and five professors (1 in physics, 2 in biology, 1 in chemistry, and 1 in geology in Foshan University) were invited to answer these two questions.
- ✧
- Question 1: Can you find any evidence able to falsify any viewpoints of the three axioms or the two mathematical rules given in this article?
- ✧
- Question 2: Can you find any evidence able to falsify the reasoning or viewpoints of CBET given in this article?
- ✧
- All three AI tools and five professors reported that they could not find robust evidence to falsify the viewpoints.
3.1.4. Empirical Validation of Natural Selection
- ✧
- ✧
- In chemical evolution, molecular populations undergo natural selection in those organic chemical reactions with multiple possible products, where the molecules that are easily formed and difficult to degrade (i.e., with a high formation-to-degradation ratio) tend to accumulate in greater quantities than those that are difficult to form and easily degraded.
- ✧
- In social evolution, companies or schools undergo competition and selection where those with the formation-to-dissolution ratio >1 demonstrate progressive accumulation, consistent with the natural selection dynamics.
- ✧
- The two mathematical rules of natural selection are explicit and self-evident. They were deduced by fundamental mathematics (See Equation 1 in the main text). They are universally applicable to all molecules, organisms, and social organizations.
- ✧
- When an asteroid impact causes extinction, the cumulative formation of the affected species throughout its evolutionary history becomes equal to its cumulative degradation at the extinction moment. This satisfies the Quantity-Existence rule.
- ✧
- For a company, Equation 1 does not represent the relationship between the company’s economic income and expenditure and its survival. Instead, it indicates the relationship between the number of establishments and closures of a specific type of company and the existing quantity of that type of company.
- ✧
- For a country, Equation 1 does not represent the relationship between its net economic income and its survival. Instead, it indicates the relationship between the number of new formations and dissolutions of a specific type of country (e.g., slave-holding countries) and the existing number of that type of country.
3.1.5. Refinement of CBET
- ✧
- The CBET was iteratively refined based on validation results, leading to improvements across multiple dimensions to strengthen the theory’s coherence, rigor, clarity, and explanatory power. These included redefining the hierarchies of CBMs, reformulating the core axioms and mechanisms, streamlining the reasoning steps, updating the mathematical formulations, and redesigning the explanatory figures.
- ✧
- The core components of CBET—including its nine hierarchies of CBMs, eight reasoning steps, three axioms, and two mechanisms—have been iteratively refined for clarity and accessibility, and all elusive concepts are excluded from the theory. An online test conducted by the authors showed that 33 out of 35 undergraduates from Foshan University successfully grasped the core components of CBET within a one-hour session.
- ✧
- Additionally, as shown in the main text, multiple sentences in the definitions of the three axioms and two mechanisms in CBET use the words like ‘some, many, partially, or can’, rather than ‘all’ or ‘must’, so that they allow exceptions.
3.2. Comparison of CBET to other theories
3.2.1. Compared to Theories in Chemistry
– Oparin's chemical evolution theory developed in the 1920s5
- ✧
- Similarity:
Both Oparin's theory and CBET claim that chemical evolution led to life’s origin and natural selection existed in chemical evolution.
- ✧
- Distinction:
- •
- Oparin's theory is limited to chemical evolution and the description of the possibility rather than revealing the underlying mechanisms and mathematical rules.
- •
- CBET applies to chemical, biological, and social evolution, and reveals the underlying mechanisms and mathematical rules of natural selection as well as chemical, biological, and social evolution.
- ✧
- Similarity:
Both theories aim to explain the origin of life from a material-based perspective.
Both acknowledge the role of information-carrying molecules in early life.
Both recognize that natural selection can operate at the molecular level.
- ✧
- Distinction:
- •
- RWH posits that self-replicating RNA molecules were the crucial, foundational step for life’s origin, acting as both the primary information carrier and catalyst. In contrast, CBET argues that such self-replicating macromolecules are exceedingly rare in nature. Instead, CBET proposes that life originated from the collaborative interactions of diverse molecules, where no single type of molecule is granted primacy. To date, though some RNA molecules can catalyze their degradation, no RNA molecules that can catalyze their replication have been identified.
- •
- RWH is primarily a theory of early biological evolution, focusing on the transition from prebiotic chemistry to the first living systems. In contrast, CBET presents itself as a universal framework, aiming to unify the mechanisms behind not only chemical and biological evolution but also social evolution, tracing a continuous path from carbon atoms to human civilizations.
- •
- RWH is grounded in the functional capabilities of RNA itself—specifically, its ability to replicate and catalyze reactions. In contrast, CBET introduces two overarching mechanisms: "spirodynamic feedback," which drives the hierarchical elevation and diversification of CBMs, and "natural selection," which shapes these changes based on fitness defined as a formation-to-degradation ratio.
3.2.2. Compared to Theories in Physics
– Ilya Prigogine's dissipative structure theory [36]
- ✧
- Similarity:
Both CBET and Prigogine’s theory, which was awarded with Nobel Prize, explain complexity emergence and orderliness enhancement in open systems.
- ✧
- Distinction:
- •
- Prigogine assumed that the still crystal-like orderliness in thermodynamics results from the same mechanism with biological and social orderliness, while CBET recognizes that the orderliness in thermodynamics is different from biological orderliness and they result from different mechanisms.
- •
- Prigogine interpreted orderliness and complexity with only a few abstract and elusive concepts (e.g., entropy), while CBET explains the mechanisms behind the natural increases in the orderliness and complexity of molecules, organisms, and social organizations with diverse concrete and explicit concepts across multiple disciplines (e.g., energy, structures, functions, feedback, and natural selection).
- •
- Prigogine highlighted self-organization and neglected the role of natural selection, while CBET highlights natural selection and highlights both functions of the target materials and the environmental influence.
- •
- Prigogine did not highlight the unique functions of carbon atoms and other CBMs, while CBET highlights the unique functions of CBMs.
- ✧
- Similarity:
Both FEP and CBET highlight feedback, energy, non-heritable traits, and the interplay of organisms and the environment.
- ✧
- Distinction:
- •
- FEP uses abstract concepts (e.g., entropy, surprise) and thus does not explicitly explain the evolution of CBMs, while CBET provides explicit explanation of the mechanisms behind the evolution of CBMs using concrete concepts (e.g., energy, structures, functions, feedback, and natural selection).
- •
- FEP does not highlight the unique functions of carbon atoms and other CBMs, while CBET highlights the unique functions of CBMs.
- •
- FEP focuses on interpreting how a complex system achieves and maintains its complex functions, rather than addressing the origins of the complex system itself, while CBET addresses the origins of the complex system.
3.2.3. Compared to Darwinian Theory in Biology
- ✧
- Similarity:
- •
- Both Darwinian theory and CBET highlight the mechanism of natural selection. In this sense, CBET is an extension rather than a replacement of Darwinian theory.
- •
- Both Darwinian theory and CBET only provide a schematic rather than detailed account for the plausibility rather than inevitability of evolution mostly in a qualitative rather than quantitative manner.
- ✧
- Distinction:
- •
- Darwinian theory is confined to biological evolution and devoid of a coherent framework to integrate various evolutionary phenomena, while CBET applies to chemical, biological, and social evolution and integrates various evolutionary phenomena in a coherent framework.
- •
- Darwinian theory has not accounted for the origins of life and societies, while CBET provides a holistic explanation for the entire evolution of CBMs, which covers the origins of life and societies.
- •
- Darwinian theory cannot calculate the fitness of molecules, worker bees, and societies, while CBET can rationally calculate their fitness.
- •
- Darwinian theory does not highlight the unique functions of carbon atoms, while CBET highlights the unique functions of carbon atoms.
- •
- Darwinian theory reveals only the natural selection mechanism of evolution, while CBET reveals the natural selection mechanism that shapes the evolution and the spirodynamic feedback mechanism that drives the evolution. Thus, CBET is more powerful in accounting for evolutionary facts. Consequently, neutral mutations, epigenetic changes, and non-heritable traits, which cannot be integrated into Darwinian theory [2,4], can be well integrated into CBET. This makes CBET more consistent with modern biological findings.
- •
- CBET clarifies the mathematical rules of natural selection and refines ‘survival of the fittest' to ‘survival of the fit enough' to prevent literal misunderstanding. It denotes that a neutral or detrimental trait can exist provided that its carrier maintains net positive growth to persist. This provides a rigorous theoretical foundation for the widespread existence of neutral evolution and detrimental traits, aligning better with the reality revealed by genomics.
3.2.4. Compared to Relevant Social Theories
- ✧
- Similarity:
Both Spencer and CBET extend evolution to society and highlight competition and elimination.
- ✧
- Distinction:
- •
- Spencer emphasized ‘survival of the fittest' in social competition and justified elimination, selfishness, and colonialism.
- •
- CBET accurately clarifies the mathematical rule of ‘survival of the fit enough', the natural balance between competition versus inclusiveness, and the natural balance between selfishness versus altruism, in natural selection.
- ✧
- Similarity:
Both MLS and CBET highlight hierarchies, altruism, and selection at multiple levels.
- ✧
- Distinction:
- •
- MLS lists populations and species without close internal collaboration as hierarchies, while CBET defines that higher-hierarchy CBMs are formed through close collaboration of lower-hierarchy CBMs.
- •
- MLS focuses on altruistic behavior in biological evolution and explains them using the claim that natural selection exists at multiple levels. Furthermore, MLS has not revealed the mechanisms underlying natural selection and chemical, biological, and social evolution. In contrast, CBET develops a coherent theory interpreting the mechanisms behind natural selection and chemical, biological, and social evolution, and consequently explains the pervasive altruistic behavior and other important phenomena in CBM evolution.
- •
- MLS does not highlight the unique functions of carbon atoms and other CBMs. CBET highlights the unique functions of CBMs.
– Compare to John Holland's Complex System Theory (CST) [42]
- ✧
- Similarity:
Both CST and CBET emphasize the emergence of new functions, adaptation to the environment, and feedback of complex systems.
- ✧
- Distinction:
- •
- CST uses abstract models and focuses on the emergence of new functions, active adaptation to the environment, and modeling the current complex systems, while CBET interprets complexity with concrete and explicit concepts and focuses on revealing the mechanisms behind the evolution.
- •
- CST has not revealed the underlying mechanisms of chemical, biological, and social evolution with concrete and explicit concepts.
- •
- CST does not highlight the unique functions of carbon atoms and other CBMs, while CBET highlights the unique functions of CBMs.
3.2.5. Conclusions and Discussion
- ✧
- While some prior theories share partial similarities with CBET, none articulates the underlying mechanisms and mathematical rules of natural selection, as well as chemical, biological, and social evolution, using concrete, empirical concepts under a coherent theoretical framework. CBET distinguishes itself by integrating concrete concepts, mathematical rules, and interdisciplinary empirical evidence to illuminate the mechanisms driving the stepwise chemical, biological, and social evolution. This systematic approach bridges gaps left by prior theories, offering a novel and holistic perspective on the CBM world and on us.
- ✧
- Previous evolutionary theories were mainly based on a few factors, such as natural selection, entropy, free energy, self-organization, or dissipative structure, some of which are incomprehensible to many common readers. Additionally, few evolutionary theories have mentioned that functions of carbon atoms and other CBMs are crucial to evolution. In contrast, CBET integrates numerous concrete comprehensible factors, such as energy, structures, functions, information pathways, genetics, epigenetics, natural selection, carbon atoms, organic molecules, cells, organisms, and social organizations, into a unified framework.
- ✧
- Previous evolutionary theories often neglected or denied the role of natural selection [3,28,36,37,38,43] and explained CBM evolution using a single principle, such as Schrödinger's negative entropy notion of dissipating entropy into the environment [44], self-organization in Prigogine's dissipative structure theory [36], the constructal law [45], the maximum entropy production hypothesis [46], or the free energy principle [38]. In contrast, CBET uses three axioms along with two mechanisms (namely spirodynamic feedback and natural selection). This contrasts with previous studies, which
- ✧
- Some other evolutionary studies highlight the hierarchies of CBMs [41]. They have categorized molecules, cells, organs, and ecosystems into different hierarchies, and these studies have not deduced the spirodynamic feedback mechanism that drives stepwise evolution from carbon atoms to societies. In contrast, in CBET, organs are defined as components of individual organisms, whereas ecosystems are regarded as a kind of environment, and CBET has derived the spirodynamic feedback mechanism.
3.3. Mathematical Formulation of CBET
3.3.1. Equations of the Three Axioms
The three axioms in CBET, which have been neglected in previous studies of evolution theories, are core axioms that have underpinned extensive empirical studies in physics, chemistry, biology, and social sciences. They can be expressed with the following equations.
Ψ: physical, chemical, biological, and social functions
m: the target material, which is typically an open system exchanging energy and matter with its environment
S: physical, chemical, biological, and social structures
: the deterministic relationship between Ψ and S
∂: change e.g., ∂Sm denotes structural changes in the target material
e: the environment (e.g., ∂S e denotes structural changes in the environment)
Φ: energy flow
𝒜: the synergistic effect of Ψm and Ψe that modulates structural changes
: the generative relationship between structural changes and the combined effects of energy flow, material's functions, and environmental functions
Equation S1 states that a material's functions are partially determined by its structure, which represents the first core tenet of the Structure-Function Axiom.
Equation S2 states that structural modifications in the target material can alter its functions, potentially yielding new functions?another core tenet of the Structure-Function Axiom.
Equations S3 and S4 extend the relationships embodied in Equations S1 and S2 to the environment, which can itself be considered as a material system.
Equation S5 reveals that energy flow drives structural changes in the target material under the modulation of the material and the environment. This equation embodies:
- ✧
- The Driving-Force Axiom (i.e., energy flow drives structural or locational changes in the target material under the modulation of the material and the environment)
- ✧
- The first core tenet of the Environment-Influence Axiom (i.e., environmental factors, which can act as the external cause, influence structural changes of the target material)
- ✧
- The third core tenet of the Structure-Function Axiom (i.e., material's functions, which can act as the internal cause, influence structural changes of the target material)
Equation S6 shows that energy flow, which is under the modulation of the material and the environment to drive structural or locational changes in the target material, simultaneously drives structural changes in the environment. This equation is grounded in fundamental physical laws, namely the Law of Conservation of Energy, the Law of Conservation of Mass, and the Principles of Newtonian Mechanics.
For example, when a company, acting as an environmental factor, donates one billion dollars to a university, its property decreases. This equation represents the second core tenet of the Environment-Influence Axiom (namely, changes of the target material can influence the environment).
3.3.2. Equations of Spirodynamic Feedback
Example 1. Certain forms of energy (e.g., fires)?when modulated by the properties of charcoal (which consists primarily of carbon atoms) and the environment (e.g., oxygen availability) ?drive the structural transformation of charcoal into CO2. The resulting CO2 exhibits novel functions that are absent in charcoal, such as participating in amino acid synthesis under appropriate energy flows and environmental conditions. This process simultaneously alters the environmental composition (e.g., increasing atmospheric CO2 concentration). The environment can also be altered by other factors. The synergistic actions of energy flows, environmental changes, and the new function of CO2 (which is absent in charcoal) can drive the transformation of CO2 into amino acids or other carbon-containing molecules.
Example 2. Certain forms of energy (e.g., the energy in food, thermal energy, electric energy, and mechanic energy) ?when modulated by the functions of a group of people (who possess financial resources, technologies, management capabilities, and team spirit) and the environment (e.g., a favorable natural, political, and economic environment) ?drive the establishment of a company. The resulting company acquires novel functions that that its individual members lack, such as manufacturing and selling a new brand of mobile phones. This process simultaneously alters the environment (e.g., the prices of mobile phones on the market have declined due to the emergence of this new brand). The environment can also be altered by other factors (e.g., changes in tariffs or social security). The synergistic actions of energy flows, environmental changes, and the new function (which emerges in the newly established company and is absent in individual humans) can drive the transformation of the company (e.g., expanding production scale, transitioning to robot manufacturing, or going out of business).
The two examples above exemplify the mechanism of spirodynamic feedback, which is outlined in Figure 2 in the main text and can be deduced from the six equations (Equations S1−S6) above:
Consequently,
The letters in Equations S7−S9 share the same meanings in Equations S1−S6.
∂Sm represents material structural changes, and ∂(∂Sm) denotes subsequent rounds of structural changes.
Because certain structural changes in CBMs on habitable Earth constitute hierarchical changes of CBMs, Equation S9 suggests the possibility of serially hierarchical changes, which can be shown by the following equation:
Equation S10 demonstrates the theoretical possibility of serial hierarchical elevation of CBMs if supported by energy flow, the functions of the target material (e.g., CBMs), and the functions of the environment.
In reality, as stated in the main text, unlike other materials, CBMs possess the functions to support the serial hierarchical elevation to life structures and social organizations, and Earth is a rare habitable planet that supports such serial elevation of CBMs, with abundant liquid water, CBMs, and diverse energy flows ?including sunlight, radiation, chemical energy release, and geothermal energy. Consequently, the origins of life and social organizations on Earth become both theoretically and empirically plausible.
The ten equations above can only serve as qualitative and universal formulations applicable to molecules, biological systems, and social organizations. They can be used to deduce the plausibility of serial hierarchical elevation of CBMs. Logically, they cannot simultaneously be quantitative and universal in that way. For example, those equations quantitatively modeling the dynamic process of carbon combustion into carbon dioxide would most likely prove inapplicable to modeling the development of social organizations.
3.3.3. Equations of Natural Selection
The following equation is self-evident in fundamental mathematics.
Nexisting: The existing quantity
Γcumulative: The cumulative formation quantity
Λcumulative: The cumulative degradation quantity
Equation S11 applies to the synthesis/breakdown of molecules, birth/death of organisms, and establishment/dissolution of social organizations, and thereby can be employed to describe the existing quantity of a molecule, organism, or social organization during a time interval.
Equation S11 suggests that a material exists if its cumulative formation exceeds its cumulative degradation, regardless of whether it is fitter than its progenitors or other materials. This validates the Quantity-Existence Rule.
According to Equation S11, the existing quantity of a material increases if its formation-to-degradation ratio is >1, and the existing quantity of a material decreases if its formation-to-degradation ratio is <1, regardless of whether it is fitter than its progenitors or other materials. This validates the Quantity-Change Rule of natural selection?The change in a material's existing quantity over a time interval is determined by the ratio of its total formation to its total degradation within that period.
The Quantity-Change Rule supports the formation/degradation ratio can be directly employed to calculate the OPFS and fitness of a CBM using Equation 1 in the main text. The formation/degradation ratio equals the ratio of formation rate versus degradation rate, as shown below.
Γab: The formation quantity of the CBM during the period [a,b]
Λab: The degradation quantity of the CBM during the period [a,b]
Fitnessab: Fitness of a CBM during the period [a,b]
tab: The length of the period [a,b]
Γrate: The formation rate during the period [a,b]
Λrate: The degradation rate during the period [a,b]
To minimize sampling biases, the observation period should be long enough to ensure that Γab and Λab reach sufficiently large values.
From Equation S11 and Equation 1, the quantity of a CBM at time a can be calculated from the quantity of a CBM at time b with the following equation:
3.4. Mathematical Modeling of CBET
3.4.1. Modeling of the Effect of Natural Selection
This section provides the mathematical formulation to model the effect of natural selection on the quantities of six mock CBMs. The model simulates how the population of each CBM changes over discrete time steps based on its fitness.
3.4.1.1. Fitness Function Definitions
The fitness of a CBM during a time interval , denoted as , is defined as its formation-to-degradation ratio (see Equation 1 in the main text). For modeling purposes, the for each CBM at time step is defined by the following piecewise functions:
- CBM_a: Constant baseline fitness,
- CBM_b: Constant exponential growth,
- CBM_c: Stepwise decrease in fitness.
- CBM_d: Parabolic growth and stabilization.
- CBM_e: Constant fitness leading to population decline,
- CBM_f: Complex pattern simulating a catastrophic crash followed by recovery.
3.4.1.2. Core Evolution Equation and Simulation Setup
The quantity dynamics of each mock CBM are governed by a recursive equation derived from the natural selection rules.
(1) Core Evolution Equation
The quantity of CBM at time step is calculated based on its previous quantity and its current fitness:
where:
- is the quantity of CBM at time step .
- is the fitness value of CBM at time step .
(2) Extinction Condition
If the quantity of a CBM falls to 1 or below, it is considered extinct, and its quantity remains at 1 for all subsequent time steps:
(3) Initial Quantities
The simulation begins with the following initial quantities ():
- CBM_a, CBM_b, CBM_c:
- CBM_d, CBM_e, CBM_f:
3.4.1.3. Modeling Results
The modeling results are given in Table 3 and Figure 4. Briefly, the modeling results demonstrate how fitness values dictate distinct population trajectories under natural selection. CBM_a, with a constant fitness of 1.0, maintains a perfectly stable population, establishing a critical baseline of neutral change. In contrast, CBM_b with sustained fitness >1 expands exponentially, while CBM_e with fitness <1 inevitably face extinction. CBM_d illustrates a more complex, parabolic pattern of growth and stabilization, showing that fitness can vary non-monotonically over time. Furthermore, the crash and recovery observed in CBM_f underscore the non-linear impact of natural selection on population resilience, revealing that populations can not only rebound from severe declines but even surpass their original size given a period of high fitness.
3.4.2. Modeling of the Possibility of Life’s Origin
3.4.2.1. Background
This mathematical modeling aims to clarify the plausibility of life's origin via chemical evolution on Earth under the CBET framework, even though the probability of inanimate molecular clusters becoming life structures is extremely low. This modeling is not intended to retrospectively fit the precise history of Earth.
CBET provides a framework (namely the eight reasoning steps involving the three axioms and two mechanisms) accounting for the plausibility of life’s origin, as described Section 1.4.
Since CBET only provides a schematic (not detailed) account for the plausibility of evolution mostly in a qualitative manner, some facts, assumptions, and hypotheses about life’s origin are needed for the quantitative modeling. These facts, assumptions, and hypotheses are not intrinsic components of CBET but can be integrated into its framework, much like walls, windows, and doors are integrated into a building's structure. They must be validated independently through future research.
3.4.2.1.1. Roles of Spirodynamic Feedback
Life's origin was driven by the spirodynamic feedback mechanism through the stepwise hierarchy elevation of CBMs from H1 to H6, as given in Section 1.4 and detailed below.
- ✧
- H2-CBMs → H3-CBMs. Chemically, some H2-CBMs can spontaneously absorb energy (e.g., heat) from certain energy flow sources (e.g., sunlight) on Earth. The absorbed energy and some functions of H2-CBMs—such as the reactivity of CO2 with H2O and the reactivity of CH4 with NH3—collectively transform H2-CBMs into diverse H3-CBMs through organic chemical reactions [47]. The prebiotic chemical synthesis of various H3-CBMs, such as amino acids, nucleotides, and monosaccharaides, has been validated in laboratories [4,5,6,20,21,23], and supported by the fact that myriad distinct H3-CBMs have been identified from meteorites [48].
- ✧
- H3-CBMs → small H4-CBMs. Likewise, chemically, many H3-CBMs can spontaneously absorb energy from certain energy flow sources on Earth. The absorbed energy and some functions of H3-CBMs—such as the one that some H3-CBMs have two or more active function groups—collectively transform H3-CBMs into diverse small H4-CBMs through organic chemical reactions. Studies have shown that high pressure, certain inorganic molecules like boric acids, and certain H3-CBMs—such as N-phosphoryl amino acids—can facilitate the abiotic synthesis of some small H4-CBMs, like short proteins, nucleic acids, lipids, and myriad heteropolymers—namely those hybrid macromolecules incorporating residues of multiple types, such as macromolecules bearing both amino acids and nucleotide residues [2,5,6,23,49].
- ✧
- Small H3-CBMs → large H4-CBMs. Likewise, chemically, small peptides can form larger peptides in the same chemical way. Thus, through intermediate steps, amino acids can indirectly form medium or large proteins, although amino acids inefficiently form large proteins directly [49]. Likewise, chemically, other medium or large H4-CBMs, including various heteropolymers, can also be formed through various intermediate steps.
- ✧
- H4-CBMs → H5-CBMs (MMAs) and MMA clusters. In physics and chemistry, some H4-CBMs have the function of transforming into H5-CBMs (multiple molecule aggregates, MMAs), such as virus-like particles or water-in-oil emulsion, through physical and chemical reactions [50]. MMAs can further form MMA clusters through physical and chemical reactions. The energy provided by the wind, rain, lightning, and solar evaporation and some functions of H4-CBMs, such as the interaction among certain proteins through hydrogen bonds, can drive the formation of myriad MMAs and MMA clusters.
- ✧
- MMA clusters → Life structures. Chemically, many proteins generated through abiogenesis are enzymes and have certain catalysis activities. They, along with diverse H3-CBMs (e.g., glucose, nucleotides, amino acids), can thus form diverse MMAs and MMA clusters with certain metabolic functions, such as catalyzing the random formation of various H4-CBMs without fixed sequences, catalyzing the formation of certain H4-CBMs (e.g., RNA or DNA) according to certain sequences (which represent certain information), and catalyzing the degradation of certain H3-CBMs or H4- CBMs, etc. The formation of these metabolic MMAs and MMA clusters, in turn, could facilitate the formation of MMAs and MMA clusters through their catalysis activities or metabolic functions. Various metabolic MMAs, such as polymerase-chain-reaction (PCR) systems for DNA synthesis according to certain sequences, have been widely applied in laboratories. Through stochastic processes, an exceedingly rare fraction of these clusters eventually achieved the complex function of reproduction, becoming the first life structures (H6-CBMs).
3.4.2.1.2. Roles of Natural Selection
- ✧
- Before life's origin, as given in Section 1.4, natural selection shaped the formation-degradation cycles of MMA clusters towards life's origin.
- ✧
- After life's origin, as given in Section 1.4, life structures are advantageous over non-replicating MMA clusters in natural selection, and life structures can be increasingly advantageous over non-reproductive MMA clusters in natural selection.
3.4.2.1.3. The Atmosphere-Origin Hypothesis
Previously, it was hypothesized that life originated in a warm pond (by Darwin), hydrothermal vents, or volcanic lakes [2,4]. Here the Atmosphere-Origin Hypothesis is proposed to explain why it was much more likely for life to emerge in Earth’s early atmosphere than in hydrothermal vents or volcanic lakes. Notably, this hypothesis is not an integral component of CBET, and it remains a hypothesis rather than established knowledge.
- ✧
- ✧
- The Atmosphere-Origin Hypothesis is grounded in the established understanding that the early Earth was too hot—with its surface in the form of liquid magma—to retain surface water. Consequently, the early (Hadean) atmosphere differed markedly from today’s, characterized by high temperatures, pressure, and water concentration [15,22]. Due to frequent volcanic activity on the early Earth, the early atmosphere contained high concentrations of volcanic ash, along with abundant reactive gases such as CO2, H2, and NH3, as well as variable winds and frequent heavy rains [6,15]. As a result, the early atmosphere, which is usually termed ‘magma-ocean atmosphere’ or ‘steam atmosphere’, resembled a floating ocean, with darkness prevailing near its base.
- ✧
- The early atmosphere—shared advantages with hydrothermal vents and volcanic lakes: abundant energy flow, water, and mineral catalysis (via suspended volcanic ash that enhances reaction yields)—was a more conducive environment for life’s origin than hydrothermal vents and volcanic lakes for the following five reasons, which could greatly enhance the plausibility of life’s origin on Earth.
- •
- Time. The early atmosphere sustained high pressure and abundant water for about 0.2 billion years before hydrothermal vents and volcanic lakes emerged.
- •
- Materials. The early atmosphere contained more abundant reactive gases such as CO2, H2, and NH3 than hydrothermal vents and volcanic lakes.
- •
- Energy flows. The early atmosphere held more diverse energy flows, such as solar radiation, lightning, and various forms of radiation, which led to vast planetary-scale spaces for the synthesis of H3-CBMs and H4-CBMs.
- •
- Spatial differences. The thermal and pressure gradients of the early atmosphere across heights and altitudes facilitated the formation, storage, and accumulation of organic molecules.
- •
- Formation of MMA clusters. The early atmosphere could naturally generate vast numbers of dynamic aerosol particles and droplets, driven by wind, dust, and evaporation-condensation cycles. These aerosol particles and droplets effectively constituted innumerable MMA clusters, providing endless trial-and-error opportunities for the emergence of life structures.
- •
- Global mixing. Efficient global mixing of MMAs and MMA clusters via wind facilitated the renewal of MMAs and MMA clusters in the early atmosphere.
The following modeling is grounded in the CBET framework and the Atmosphere-Origin Hypothesis. A key merit of the Atmosphere-Origin Hypothesis is its proposition that Earth's early atmosphere functioned as a vast, planet-scale, and fully integrated organic reactor, providing a far more extensive and dynamic environment for prebiotic synthesis of organic molecules than localized settings like hydrothermal vents.
3.4.2.2. Initial Setup and Time Segmentation
- Initial amount of H1-CBMs on Earth: Changes are not considered (due to the chemical instability of uncombined H1-CBMs under high temperatures on early Earth and their significantly smaller quantity compared to H2-CBMs).
- Initial amount of H2-CBMs on Earth: NH2⁰ = 10²⁴ mol
This value represents a substantial but plausible reservoir of reactive carbon available for prebiotic chemistry. While the bulk carbon content of the early Earth is uncertain, this estimate is consistent with the planet having accreted from carbon-bearing building blocks and subsequently outgassing a significant fraction of this carbon into the early atmosphere and hydrosphere as simple molecules (H2-CBMs like CO₂ and CH₄). The high temperatures on early Earth would have favored the formation of these stable molecular compounds over uncombined carbon atoms, justifying the focus on H2-CBMs as the primary starting reservoir.
- Initial amount of H3-CBMs and higher-hierarchy CBMs on Earth: 0
Although H3-CBMs and higher-hierarchy CBMs could have accumulated vastly by or during Earth's formation11, this would not affect the conclusion of this modeling.
-
Time Segments:
- ○
- 1st 10 million years: 0 – 10 Myr
- ○
- 2nd 10 million years: 10 – 20 Myr
- ○
- 3rd 10 million years: 20 – 30 Myr
3.4.2.3. Consumption of H2-CBMs
- According to the CBET framework, H2-CBMs were consumed to form H3-CBMs, which can further to form H4-CBMs, and the formation was driven by the synergetic action of energy flow, functions of H2-CBMs and H3-CBMs, and influence of the environment.
- The formation of H3-CBMs and H4-CBMs could be accelerated due to the catalysis function of some H3-CBMs and higher-hierarchy CBMs (an effect of the spirodynamic feedback mechanism), and thus the consumption of H2-CBMs could be accelerated during the first thirty million years (Myr).
- H2-CBMs can be generated through the degradation of H3-CBMs and higher-hierarchy CBMs, and thus the quantity of H2-CBMs could be relatively stable after the first thirty Myr.
Assumed H2-CBMs Remaining Ratio
- 0–10 Myr: 99.8% → Remaining 0.998
- 10–20 Myr: 99.0% → Remaining 0.990
- 20–30 Myr: 90.0% → Remaining 0.900
- H2-CBMs are assumed to remain relatively constant thereafter (steady state).
3.4.2.4. Growth Calculation of H3-CBMs
-
During the first 10 Myr
- ○
- Reduction of H2-CBMs: ΔNH2 = 10²⁴ × (1 - 0.998) = 2 × 10²¹
- ○
- Converted to H3-CBMs (each H3-CBM is assumed to contain 10 carbon atoms, and 5% of H3-CBMs further formed H4-CBMs):
ΔNH3⁽¹⁾ = (2 × 10²¹ / 10) × 0.95 = 1.9 × 10²⁰ mol
-
During the second 10 Myr
- ○
- ΔNH2= 10²⁴ × (0.998 - 0.990) = 8 × 10²¹
- ○
- Converted to H3-CBMs:
ΔNH3⁽²⁾ = (8 × 10²¹ / 10) × 0.95 = 7.6 × 10²⁰ mol
-
During the third 10 Myr
- ○
- ΔNH2 = 10²⁴ × (0.990 - 0.900) = 9 × 10²²
- ○
- Converted to H3-CBMs:
ΔNH3⁽³⁾ = (9 × 10²² / 10) × 0.95 = 8.55 × 10²¹ mol
- Cumulative H3-CBMs (at 30 Myr)
NH3= 1.9 × 10²⁰ + 7.6 × 10²⁰ + 8.55 × 10²¹ = 9.5 × 10²¹ mol
- H3-CBMs are assumed to remain relatively constant thereafter (steady state).
3.4.2.5. Growth Calculation of H4-CBMs
Small H4-CBMs
- Each small H4-CBM is assumed to contain 15 H3-CBM residues.
- 5% of H3-CBMs are consumed to form small H4-CBMs, with 97% retained (3% of small H4-CBMs are consumed to form middle or large H4-CBMs), and thus at the end of the third 10 Myr, the quantity of small H4-CBMs:
Nsmall H4 = (NH3 × 0.05 / 15) × 0.97 = (9.5 × 10²¹ × 0.05 / 15) × 0.97 = 3.069 × 10¹⁹ mol
Middle H4-CBMs
- Each middle H4-CBM is assumed to contain 160 H3-CBM residues.
- 3% of small H4-CBMs are consumed to form middle H4-CBMs, with 98% retained (2% of middle H4-CBMs are consumed to form large H4-CBMs), and thus at the end of the third 10 Myr, the quantity of middle H4-CBMs:
Nmiddle H4= (NH3 × 0.05 × 0.03 / 160) × 0.98 = (9.5 × 10²¹ × 1.5 × 10⁻³ / 160) × 0.98 = 8.728 × 10¹⁶ mol
Large H4-CBMs
- Each large H4-CBM is assumed to contain 600 H3 residues.
- 2% of middle H4-CBMs are consumed to form large H4-CBMs, and thus at the end of the third 10 Myr, the quantity of large H4-CBMs
Nlarge H4 = (NH3 × 0.05 × 0.03 × 0.02 / 600) = (9.5 × 10²¹ × 3 × 10⁻⁵ / 600) = 4.75 × 10¹⁴ mol
- Although the total numbers of small, middle, and large H4-CBMs after the end of the third 10 Myr are assumed to be constant, they were formed and degraded simultaneously at largely equal rates after that time.
3.4.2.6. Growth Calculation of MMA Clusters
Number oflarge H4-CBMsmolecules at the end of the third 10 Myr
Number of molecules = 4.75 × 10¹⁴ × 6.022 × 10²³ = 2.860 × 10³⁸ molecules
Large H4moleculesavailable for MMAclusterformation
- Participation probability: 10% (10 in 100)
Participating Nlarge H4 = 2.860 × 10³⁸ × 0.1 = 2.860 × 10³⁷ molecules
Total MMA Clusters
- Each MMA cluster is assumed to contain 1,000 large H4-CBMs molecules (Other materials such as water are assumed to be abundant for the formation of MMA clusters)
- Total MMA clusters:
Nmc= 2.860 × 10³⁷ / 1000 = 2.860 × 10³⁴ clusters
Annual Newly Generated MMA Clusters
- MMA clusters are assumed to be renewed (e.g., addition, removal, or alteration of a macromolecule within a cluster) on average every second.
- Annual renewal cycles: 365 × 24 × 60 × 60 = 15,156,000
- Annual production:
Rmc, gen = 15,156,000 × 2.860 × 10³⁴ = 4.335 × 10⁴¹ clusters/year
3.4.2.7. Calculation of Life Origin Events
Probability of a new MMA cluster becoming a life structure is assumed to be:
Plife = 10⁻⁴⁵
If "becoming a life structure" were defined as the instantaneous abiogenesis of a modern cell with its super-sophisticated internal machinery, such as the ribosome, this probability (10⁻⁴⁵) could be a significant overestimate. However, this model refers to the emergence of a primordial life prototype. Such an initial structure could be vastly simpler than any modern cell—akin to the difference between a primitive telegraph and a modern smartphone. Furthermore, the formation and accumulation of MMA clusters on early Earth were not random but shaped by natural selection towards the direction beneficial to life’s origin, as exemplified in Section 3.4.2.1.2. Therefore, for the spontaneous formation of a minimal, self-replicating molecular system, the estimated probability of could be a plausible and appropriate value for modeling purposes. Nevertheless, this extremely low value aligns with the universal expectation that the transition from an inanimate molecular cluster to a living structure is an event of exceptionally low probability.
Annual life formation events
Lyear = 4.335 × 10⁴¹ × 10⁻⁴⁵ = 4.335 × 10⁻⁴ events/year
Total life formation events during the first 800 million years
770 Myr = 7.7 × 10⁸ years
Ltotal = 4.335 × 10⁻⁴ × 7.7 × 10⁸ = 333,800 events
3.4.2.8. Conclusion
This modeling shows that over the first 800 million years after Earth's formation, there could be 33,380 times that MMA clusters accidentally became self-replicating life structures, even though the probability of an inanimate molecular cluster becoming a life structure is assumed to be as low as 10⁻⁴⁵. The possibility above for life’s origin through abiogenesis is reinforced by the fact that experiments on the abiotic synthesis of viruses and cells have been successful (Fact 12 and Fact 13 listed in Table 2) [34,35].
3.5. Questions and Answers
3.5.1. Is CBET Falsifiable
Answer:
Yes, it is. CBET's viewpoints pertain to molecules, organisms, and social organizations—all of which are directly observable and testable. Furthermore, its viewpoints—such as the one that structural changes are driven by energy flow that is modulated by functions of the target material and the environment—are expressed without elusive concepts. Therefore, its viewpoints are explicit and testable. This distinguishes CBET from indirect inferences, like the composition of the Earth's core, grounding it firmly in empirical science.
3.5.2. What Is the Most Revolutionary Aspect of CBET
Answer:
The most revolutionary aspect of CBET is its establishment of a unifying theoretical framework—one that explains the fundamental mechanisms underlying chemical, biological, and social evolution.
3.5.3. Why Is CBET Not Derived from Novel Experiments
Answer:
- ✧
- Experiments, which are suitable to address specific issues, are unsuitable to derive the broad-scoped theory of CBET, which addresses the changes of countless molecules, organisms, and social organizations.
- ✧
- Instead, CBET can be established through integrative reasoning from vast established knowledge and empirical validation via falsification against countless molecules, organisms, and social organizations. This process effectively validated CBET against a myriad of facts across diverse disciplines. Likewise, Darwinian theory was established through the same methodology rather than by a few experiments.
- ✧
- Since Darwin’s era, researchers have accumulated extensive knowledge on molecules, organisms, and social systems. Over more than three decades, the authors have synthesized this vast body of established knowledge to derive, validate, and refine CBET to explain the mechanisms underlying chemical, biological, and social evolution, and this synthetic work became promising when they adopted carbon-based materials as the core concept in 2018.
3.5.4. Why Is CBET Easy to Understand
Answer:
- ✧
- As shown in Figure 2, while CBET is significantly more complex and harder to understand than Darwinian theory, it is more accessible than quantum mechanics and astronomy.
- ✧
-
CBET itself can be easy to understand for two reasons:
- •
- Carbon-based materials, ranging from carbon atoms to social organizations, are substances that humans can directly observe with naked eyes or instruments. They follow relatively simple and understandable laws, as they are neither obscure substances below the atomic level nor esoteric ones above the galactic level.
- •
- The evolutionary mechanisms unveiled by CBET are shared by molecules, organisms, and social organizations. Therefore, these mechanisms and their underlying mathematical rules apply to social organizations—which are familiar to the general public—making them easy to comprehend.
- ✧
-
CBET has been derived towards an explicit theory, according to the principle that scientific explanations should be as explicit as possible.
- •
- The authors have spent over three decades employing concrete and understandable terms (e.g., structure, function, change, energy, carbon atoms, and reproduction), rather than elusive concepts, to derive and describe CBET.
- •
- Again, CBET is like the first aerial photograph of a maze, which clearly shows the correct path from its entrance to its exit.
3.5.5. Does CBET Oversimplify the Behavior of Countries
Answer:
- ✧
- CBET only describes certain not all aspects of countries.
- ✧
- CBET is inclusive in its statement. For example, its statement that structures partially determine functions allows the influence of the history, culture, and the environment on the functions of countries.
- ✧
- CBET is applicable to CBMs at diverse hierarchical levels because its key concepts of CBMs, energy, changes, structures, functions, and environments—apply universally to diverse hierarchies and can become increasingly complex as hierarchies are elevated.
- ✧
- A country can emerge through an evolutionary trajectory where carbon atoms → small carbon-containing molecules → medium carbon-containing molecules → large carbon-containing molecules → multiple molecular aggregates → cells → multicellular organisms → humans → tribes → tribal alliances → countries.
- ✧
- A country's behavior and evolution are shaped by both natural and social environments, relying on energy flow as well as its physical, chemical, biological, and social functions. Its social functions include political, cultural, legislative, religious, and military functions.
- ✧
- Reforming the structure of a country can generate new functions, enabling more or less adaptation to environmental changes.
- ✧
- The facts above collectively demonstrate that CBET neither oversimplifies country's behavior nor ignores political or cultural factors.
3.5.6. Can CBET Describe CBM Evolution Quantitatively
Answer:
- ✧
- No, and such quantification is unnecessary and theoretically unjustified.
- ✧
- CBET is in part derived from principles across diverse disciplines by abstracting away their quantitative parameters to render them applicable to all molecules, organisms, and societies. Consequently, CBET provides a qualitative description of the relationships among environmental factors, energy flow, structures, and functions. These relationships vary profoundly among different hierarchies, materials, and environments, making the unified quantitative framework impractical and theoretically unjustified. For example, a quantitative description of the dynamic process of carbon combustion into carbon dioxide would most likely prove inapplicable to describe the development of social organizations.
3.5.7. Can CBET Explain the Inevitability of Life’s Origin
Answer:
No, It cannot. CBET can explain the plausibility of life’s origin, which could be a milestone in science. Since it took hundreds of millions of years for the origin of life2, the origin of life could be exceedingly rare rather than inevitable events. Thus, it is rational for CBET not to account for the inevitability of life’s origin.
3.5.8. Can CBET Reveal the Details of CBM Evolution
Answer:
No, It cannot. CBET is a framework theory that can unify chemical, biological, and social evolution, which could be a milestone in science. However, because CBM evolution involves countless distinct changes in molecules, organisms, and social organizations, and many changes occurred millions or even billions of years ago, without robust records, it is rational for CBET not to provide a detailed explanation for CBM evolution.
3.5.9. Can CBET Predict Anything
Answer:
This question can be addressed from two distinct perspectives:
(1) When predicting concrete changes in a specific CBM—such as the growth rate of a tree or a cell's metabolic pathway—theories from relevant specialized fields (e.g., plant physiology, molecular biology) are often more precise and powerful than CBET. As a qualitative framework, CBET does not aim to replace these domain-specific theories in their own areas of expertise.
(2) When predicting systemic or cross-hierarchical issues, CBET's predictive power can be more powerful than others, as demonstrated by the following examples:
- CBET predicts that many organisms (e.g., humans) and social organizations (e.g., companies) can sustain numerous detrimental traits, as CBET clarifies that these organisms or social organizations can exist on Earth as long as their cumulative formation exceeds their cumulative degradation.
- CBET predicts that sustainable social organizations typically exhibit a natural balance between competition and inclusiveness, as well as between selfishness and altruism, as these balances are inherent outcomes of the spirodynamic feedback and natural selection mechanisms underlying the evolution of social organizations.
- CBET predicts that self-replicating macromolecules—long sought in studies on the origin of life29—are likely rare in nature and were not a crucial prerequisite for life's emergence. This is because the complex function of self-replication, involving information encoding, reading, and multi-step synthesis, typically requires a structural complexity far beyond that of a single macromolecule, and the reproduction function of life emerged on Earth due to collaboration of various molecules rather than the self-replication function of certain molecules.
3.5.10. Is CBET a Theory of Philosophy
Answer:
No, CBET is a scientific theory, not a theory of philosophy, as explained below.
- ✧
- CBET is grounded in empirical evidence and scientific principles, not philosophical reasoning.
- ✧
- Its three axioms are derived from well-established knowledge in physics, chemistry, biology, and social sciences (e.g., principles from Newtonian mechanics, thermodynamics, heredity, and social change).
- ✧
- Its core mechanisms (spirodynamic feedback and natural selection) are reasoned from the three axioms, mathematical rules, and the observable functions of carbon-based materials (CBMs) and environments.
- ✧
- CBET is testable and falsifiable, which are hallmarks of scientific theories.
- ✧
- CBET's validity was explicitly tested against numerous facts spanning molecular, biological, and social phenomena.
- ✧
- The authors actively sought counterevidence through self-scrutiny, peer review, and AI-assisted analysis, adhering to scientific practice.
- ✧
- CBET does not rely on ‘a priori’ reasoning, metaphysical assumptions, or any specific school of philosophical thought.
- ✧
- Whereas philosophy also engages with broad concepts like 'hierarchy' and 'complexity', CBET distinguishes itself by explaining them through concrete, scientific concepts—such as energy flow and structural changes—within a coherent empirical framework.
- ✧
- Since CBET shares a broad scope with certain philosophical studies, it can act as a bridge between philosophy and science. This is particularly relevant in the mesocosmic world—the realm between atoms and the solar system, in contrast with the picocosmic world (below the atomic level) and the megacosmic world (beyond the solar system).
3.5.11. How Can AI Tools Support This Work
Answer:
- ✧
- AI tools have significantly assisted in refining the expression of this article. For example, they suggested revising ‘total formation' to the more precise ‘cumulative formation' in the statement of the Quantity-Existence Rule.
- ✧
- AI tools have significantly assisted in the validation of CBET through their efficient seeking of counterevidence to challenge CBET.
- ✧
- The authors find themselves in a race against rapidly advancing AI tools in the development of this theory. It is highly plausible that AI could independently formulate a similar theory in the near future—even without referencing this article.
3.5.12. What Energy Flows Drive the Transitions of CBMs Across Hierarchies
Answer:
As elucidated in Section 3.4.2.1, changes from H2-CBMs to H3-CBMs and from H3-CBMs to H4-CBMs are driven by chemical energy flows, while changes from H4-CBMs to H5-CBMs and from H5-CBMs to inanimate MMA clusters are driven by physical energy flows such as wind or wet-dry cycles. In contrast, changes from H5-CBMs to H6-CBMs and from H6-CBMs to H7-CBMs through biological approaches are driven by chemical and physical energy flows under the modulation of biological functions of organisms, and changes from H7-CBMs to H8-CBMs and from H8-CBMs to H9-CBMs are driven by chemical, physical, and/or biological energy flows under the modulation of social functions of relevant individuals or societies.
3.6. Glossary
Altruism —Selfless behaviors that benefit others.
Carbon —A chemical element with the symbol C and atomic number 6. It is a non-metallic, tetravalent element—meaning it can form four covalent chemical bonds with other atoms, a property that enables it to construct an extraordinarily diverse range of complex molecules. Naturally occurring in three primary allotropes (diamond, graphite, and fullerenes), carbon is ubiquitous across the universe and essential to all known forms of life on Earth, serving as the foundational ”backbone” of organic compounds like carbohydrates, lipids, proteins, and nucleic acids (DNA and RNA) through the chains of C-C bonds. Beyond biology, it has widespread industrial applications: for example, diamond is used in cutting tools due to its extreme hardness, graphite in pencils and lubricants for its layered structure, and carbon fibers in high-strength materials for aerospace and automotive industries. Carbon also plays a critical role in Earth’s carbon cycle, regulating atmospheric carbon dioxide levels and influencing global climate.
Carbon cycle —A complex, naturally occurring cycle of the formation and degradation of carbon- based materials. It describes the continuous movement and transformation of carbon-based materials between the Earth’s four main reservoirs: the atmosphere, oceans, terrestrial biosphere (including plants, animals, and soil), and geosphere (rocks, fossil fuels, and sediment).
CBET (the Carbon-Based Evolutionary Theory) —A theory that explains the stepwise evolution of carbon-based materials from carbon atoms to social organizations in a unified framework. It states that, on habitable Earth, three axioms associated with energy flow, structures, functions, and changes of carbon-based materials and the environment, together with two universal mathematical rules, collectively lead to stepwise chemical, biological, and social evolution from carbon atoms to social organizations via spirodynamic feedback and natural selection. Because CBET highlights the importance of natural selection in evolution, it is an extension rather than replacement of Darwinian theory.
CBMs (carbon-based materials) —Carbon-centered substances ranging from carbon atoms to complex social organizations, classified into nine hierarchies (H1-CBMs to H9-CBMs). The authors propose to create a new word, such as ‘carbonal’, as its synonym.
- ✧
- H1-CBMs: carbon atoms.
- ✧
- H2-CBMs: small carbon-containing molecules (e.g., CH4, CO2, HCN).
- ✧
- H3-CBMs: medium carbon-containing molecules (e.g., lysine, glucose).
- ✧
- H4-CBMs: large carbon-containing molecules (e.g., lipids, proteins, nucleic acids).
- ✧
- H5-CBMs: multi-molecule aggregates (e.g., phospholipid bilayers, cellulose microfibrils).
- ✧
- H6-CBMs: individual cells (e.g., bacteria, cells in animals).
- ✧
- H7-CBMs: multicellular organisms (e.g., pines, rabbits).
- ✧
- H8-CBMs: single-level social organizations (e.g., colonies, small companies).
- ✧
- H9-CBMs: multi-level social organizations (e.g., universities, armies, countries) - Higher-hierarchy CBMs and lower-hierarchy CBMs: they are defined by hierarchy comparison. For example, H6-CBMs are at a higher hierarchy relative to H5-CBMs but at a lower hierarchy relative to H7-CBMs. Higher-hierarchy are formed by lower-hierarchy CBMs and other materials through physical, chemical, biological, or social interactions.
CBM Evolution —The evolution of carbon-based materials, which includes chemical, biological, and social evolution.
- ✧
- Chemical Evolution The evolution from small to large carbon-containing molecules, and further to multi-molecule aggregates.
- ✧
- Biological Evolution The origin and evolution of life.
- ✧
- Social Evolution The origin and evolution of animal and human social organizations (e.g., colonies, tribes, countries).
CCMs (carbon-containing molecules) —molecules with carbon as a key component, which are fundamental building blocks of CBMs. They are categorized into small (H2-CBMs, e.g., CH4, CO2), medium (H3-CBMs, e.g., glucose), and large (H4-CBMs, e.g., proteins, nucleic acids) based on size and complexity.
Darwinian theory —A developing evolutionary theory that is grounded in Charles Darwin’s original ideas (e.g., natural selection, descent with heritable modifications) and refined by subsequent scientific advancements (e.g., those in genetics and developmental biology). It focuses on biological evolution and explains biological evolution using natural selection.
Driving Force —The force driving structural or locational changes in materials. In CBET, the driving force of changes are derived from energy flow, functions of target materials (as internal causes), and functions of the environment (as external causes)
Energy —A fundamental and concrete concept across physics, chemistry, biology, and social sciences that offers the capacity of a system to perform work, generate heat, or produce structural changes. It exists in various forms (e.g., kinetic, potential, and thermal energies) and can be transformed from one type to another, but it cannot be created or destroyed (Law of Conservation of Energy). It originates from the target materials, the environment, or the interaction between the target materials and the environment. The effects of energy on making changes are influenced by the functions of both the target materials and the environment.
Fitness (in CBET) —A measure of the overall performance of a carbon-based material (CBM) in its formation and sustenance, calculated as the ratio of formation to degradation over a time interval. It determines how fast the quantity of a CBM increases or decreases.
Function —The capability of a material partially determined by its structure.
Heteropolymers—Those macromolecules incorporating residues of multiple types, such as macromolecules bearing both amino acids and nucleotide residues.
Hierarchy —A level or rank in the classification of carbon-based materials (CBMs), ranging from H1 (carbon atoms) to H9 (multi-level social organizations).
Information —Data or signals that enable a material to make changes not randomly.
Life (life structures) —Those clusters of multi-molecule aggregates having the reproduction function.
Mathematical Rules of Natural Selection —Quantitative rules describing natural selection in CBET:
- ✧
- The Quantity-Existence Rule: A material exists if its cumulative formation exceeds its cumulative degradation (‘survival of the fit enough’).
- ✧
- The Quantity-Change Rule: The quantity of a material increases if its formation-to-degradation ratio >1 and decreases if <1.
MMA (multiple-molecule aggregate) —An aggregate of multiple large carbon-containing molecules. Examples include phospholipid bilayers and cellulose microfibrils, which serve as intermediate structures in the evolution from large CCMs to cells.
MMA clusters —Clusters formed by multiple molecule aggregates (MMAs) driven by intermolecular forces and environmental factors (e.g., wind, water flow).
Natural selection (in CBET) —the natural process that a carbon-based material (CBM) carrying a change increasing its fitness is prone to be more prevalent and a CBM carrying a change decreasing its fitness is prone to be less prevalent. This natural process stems from mathematical rules and its selection options, namely the changes of CBMs that result from energy flow under the modulation of the CBMs and the environment.
Orderliness —A characteristic of a material referring to the degree of organization, regularity, and non-randomness in its structure and functions. Notably, biological orderliness and social orderliness are sometimes different from thermodynamical orderliness (e.g., cold, still, and crystal-like orderliness) [3,27]. In biology, orderliness does not immediately result from one-step of thermodynamic process (e.g., entropy decline or dissipation), but results from multiple processes, including encoding, reading, translating, mutating, storing, and/or transmitting genetic information primarily through the interaction of nucleic acids and proteins, together with long-term natural selection.
OPFS (Overall Performance in Formation and Sustenance) —the collective contribution of a material’s constituents, traits, and hierarchies to its formation and sustenance, determining the fate of the material in natural selection.
Spirodynamic feedback —Energy flow under the modulation of the functions of CBMs and the environment drives structural and/or locational changes in CBMs; the changes, in turn, can create new functions of CBMs and change the environment; the emergence of new functions of CBMs and new changes in the environment, along with energy flow, can lead to a new round of structural changes of CBMs. These processes constitute the spirodynamic feedback mechanism. It leads to endless structural changes of CBMs, diversification of CBMs, and hierarchy elevation of CBMs.
Structure —The arrangement, organization, or configuration of components (such as atoms, molecules, cells, or larger entities) that is associated with specific spatial relationships, connections, or patterns between the parts of the entities.
The three axioms in CBET —The three axioms in CBET have been derived from empirical evidence across disciplines and have guided many empirical studies in physics, chemistry, biology, and social sciences.
- ✧
- The Environment-Influence Axiom: The environment has the function to provide energy, constituent materials, and other conditions to influence structural changes of the target material, and the structural changes of the target material can in turn influence the environment.
- ✧
- The Structure-Function Axiom: For a material, its structure partially determines its functions, and its structural changes can generate new functions, and in turn, its functions (acting as the internal causes) are crucial for maintaining or altering its structure.
- ✧
- The Driving-Force Axiom: Qualitatively, energy flow drives structural or locational changes of the target material under the modulation of the functions of the material (as internal causes) and the environment (as external causes).
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
J.M.C. conceived, designed, performed, and financially supported this study; Y.Q.C. conducted the mathematical formulation and drew figures; J.W.C. performed and financially supported this study; J.M.C. wrote the manuscript; all authors revised and approved the manuscript.
Acknowledgements
This study was supported by the High-Level Talent Fund of Foshan University (No. 20210036). The authors thank Meng Yang, Feng Tang, Zhen Liu, Hai Xiang, Ying Li, and many other people for their comments on this study.
Conflicts of Interest
The authors declare no competing interests.
References
- Darwin, C. On the Origin of Species 1st edn (John Murray, 1859).
- Futuyma, D. J. & Kirkpatrick, M. Evolution 4th edn (Sinauer Associates, 2017).
- Chen, J. A new evolutionary theory deduced mathematically from entropy amplification. Chin. Sci. Bull. 2000, 45, 91–96. [CrossRef]
- Xie, P. The Aufhebung and Breakthrough of the Theories on the Origin and Evolution of Life (Science Press, 2014).
- Oparin, A. I. Chemistry and the origin of life. R. Inst. Chem. Rev. 2, 1–12 (1969).
- Fiore, M. Prebiotic Chemistry and Life's Origin (Royal Society of Chemistry, 2022).
- Nowak, M. A., Tarnita, C. E. & Wilson, E. O. The evolution of eusociality. Nature 466, 1057–1062 (2010).
- Birch, J. The Philosophy of Social Evolution (Oxford Univ. Press, 2017).
- Cowan, J. J. One of the first of the second stars. Nature 488, 288–289 (2012).
- Razeghi, M. The Mystery of Carbon: An Introduction to Carbon Materials (IOP Publishing, 2019).
- Heck, P.R.; Greer, J.; Kööp, L.; Trappitsch, R.; Gyngard, F.; Busemann, H.; Maden, C.; Ávila, J.N.; Davis, A.M.; Wieler, R. Lifetimes of interstellar dust from cosmic ray exposure ages of presolar silicon carbide. Proc. Natl. Acad. Sci. 2020, 117, 1884–1889. [CrossRef]
- Li, Z. H. et al. Exceptional preservation of an extinct ostrich from the Late Miocene Linxia Basin of China. Vertebrata PalAsiatica 59, 229–248 (2021).
- Reichle, D. E. The Global Carbon Cycle and Climate Change 2nd edn (Elsevier, 2023).
- Seager, S. Exoplanet Habitability. Science 2013, 340, 577–581. [CrossRef]
- Rodriguez, L.E.; Altair, T.; Hermis, N.Y.; Jia, T.Z.; Roche, T.P.; Steller, L.H.; Weber, J.M. Chapter 4: A Geological and Chemical Context for the Origins of Life on Early Earth. Astrobiology 2024, 24, S76–76. [CrossRef]
- Olejarz, J.; Iwasa, Y.; Knoll, A.H.; Nowak, M.A. The Great Oxygenation Event as a consequence of ecological dynamics modulated by planetary change. Nat. Commun. 2021, 12, 1–9. [CrossRef]
- Benton, M.J. The Red Queen and the Court Jester: Species Diversity and the Role of Biotic and Abiotic Factors Through Time. Science 2009, 323, 728–732. [CrossRef]
- De Capitani, J., Mutschler, H. The long road to a synthetic self-replicating central dogma. Biochemistry 62(7), 1221–1232 (2023).
- Marchi, S.; Korenaga, J. The shaping of terrestrial planets by late accretions. Nature 2025, 641, 1111–1120. [CrossRef]
- Guo, X.; Fu, S.; Ying, J.; Zhao, Y. Prebiotic chemistry: a review of nucleoside phosphorylation and polymerization. Open Biol. 2023, 13, 220234. [CrossRef]
- Nam, I., Lee, J. K., Nam, H. G. & Zare, R. N. Abiotic production of sugar phosphates and uridine ribonucleoside in aqueous microdroplets. Proc. Natl Acad. Sci. USA 114, 12396–12400 (2017).
- Dobson, C.M.; Ellison, G.B.; Tuck, A.F.; Vaida, V. Atmospheric aerosols as prebiotic chemical reactors. Proc. Natl. Acad. Sci. 2000, 97, 11864–11868. [CrossRef]
- Griffith, E.C.; Vaida, V. In situ observation of peptide bond formation at the water–air interface. Proc. Natl. Acad. Sci. 2012, 109, 15697–15701. [CrossRef]
- Trainer, M. Atmospheric Prebiotic Chemistry and Organic Hazes. Curr. Org. Chem. 2013, 17, 1710–1723. [CrossRef]
- Wang, S.-Q.; Ye, J.; Meng, J.; Li, C.; Costeur, L.; Mennecart, B.; Zhang, C.; Zhang, J.; Aiglstorfer, M.; Wang, Y.; et al. Sexual selection promotes giraffoid head-neck evolution and ecological adaptation. Science 2022, 376, 1067–+. [CrossRef]
- Dawkins, R. The Selfish Gene (Oxford Univ. Press, 1976).
- Low, D. Advantage of an epigenetic switch in response to alternate environments. Proc. Natl. Acad. Sci. 2024, 121. [CrossRef]
- Veigl, S.J.; Vasilyeva, Z.; Müller, R. Scientific-intellectual movements in the post-truth age: The case of the Extended Evolutionary Synthesis. Soc. Stud. Sci. 2025. [CrossRef]
- Chen, J. M., Chen, J. W. & Zivieri, R. Systematically challenging three prevailing notions about entropy and life. Qeios. 2024. [CrossRef]
- Fitzgerald, D.M.; Rosenberg, S.M. What is mutation? A chapter in the series: How microbes “jeopardize” the modern synthesis. PLOS Genet. 2019, 15, e1007995. [CrossRef]
- Fine, J.L.; Moses, A.M. An RNA Condensate Model for the Origin of Life. J. Mol. Biol. 2025, 437, 169124. [CrossRef]
- Hunt, T. Reconsidering the logical structure of the theory of natural selection. Commun. Integr. Biol. 2014, 7, e972848–e972848. [CrossRef]
- Zhu, S.-C.; Chen, G.-W.; Yuan, X.-H.; Cheng, Y.; Xu, B.; Wang, M.-S.; Tang, H.; Hou, Y. Key for Hexagonal Diamond Formation: Theoretical and Experimental Study. J. Am. Chem. Soc. 2025, 147, 2158–2167. [CrossRef]
- Stobart, C.C.; Moore, M.L. RNA Virus Reverse Genetics and Vaccine Design. Viruses 2014, 6, 2531–2550. [CrossRef]
- Hutchison, C.A., 3rd; Chuang, R.-Y.; Noskov, V.N.; Assad-Garcia, N.; Deerinck, T.J.; Ellisman, M.H.; Gill, J.; Kannan, K.; Karas, B.J.; Ma, L.; et al. Design and synthesis of a minimal bacterial genome. Science 2016, 351, aad6253–aad6253. [CrossRef]
- Prigogine, I. Time, structure and fluctuation. Science 201, 777–785 (1978).
- Ramstead, M. J. D., Badcock, P. B. & Friston, K. J. Answering Schrödinger’s question: A free-energy formulation. Phys. Life Rev. 24, 1–16 (2018).
- Friston, K. The free-energy principle: a unified brain theory?. Nat. Rev. Neurosci. 2010, 11, 127–138. [CrossRef]
- Rudman, L.A.; Saud, L.H. Justifying Social Inequalities: The Role of Social Darwinism. Pers. Soc. Psychol. Bull. 2020, 46, 1139–1155. [CrossRef]
- Stanford, P.K.; Sober, E.; Wilson, D.S. Unto Others: The Evolution and Psychology of Unselfish Behavior. J. Philos. 2001, 98, 43. [CrossRef]
- Eldredge, N., Pievani, T., Serrelli, E. & Tëmkin, I. Evolutionary Theory: A Hierarchical Perspective (Univ. Chicago Press, 2016).
- Miller, J.H.; Page, S.E. Complex Adaptive Systems: An Introduction to Computational Models of Social Life; Walter de Gruyter GmbH: Berlin, Germany, 2009; ISBN: .
- Sabater, B. Entropy Perspectives of Molecular and Evolutionary Biology. Int. J. Mol. Sci. 2022, 23, 4098. [CrossRef]
- Schrödinger, E. What is Life (Cambridge Univ. Press, 2012).
- Bejan, A. The principle underlying all evolution, biological, geophysical, social and technological. Philos. Trans. R. Soc. A: Math. Phys. Eng. Sci. 2023, 381, 20220288. [CrossRef]
- Dewar, R.C. Maximum entropy production and plant optimization theories. Philos. Trans. R. Soc. B: Biol. Sci. 2010, 365, 1429–1435. [CrossRef]
- Borgnakke, C. & Sonntag, R. E. Fundamentals of Thermodynamics (Wiley, 2022).
- Schmitt-Kopplin, P.; Gabelica, Z.; Gougeon, R.D.; Fekete, A.; Kanawati, B.; Harir, M.; Gebefuegi, I.; Eckel, G.; Hertkorn, N. High molecular diversity of extraterrestrial organic matter in Murchison meteorite revealed 40 years after its fall. Proc. Natl. Acad. Sci. 2010, 107, 2763–2768. [CrossRef]
- Yang, Y.; Wang, Z.; Bai, J.; Qiao, H. Prebiotic Peptide Synthesis: How Did Longest Peptide Appear?. J. Mol. Evol. 2025, 93, 193–211. [CrossRef]
- Yang, B.; Liu, A.; Li, X.; Miao, Z. Research progress on mRNA drug delivery strategies in the nervous system. Nanomedicine 2025, 1–16. [CrossRef]
Figure 1.
Schematic evolutionary pathway of carbon-based materials (CBMs) from carbon atoms and carbon-containing molecules (CCMs) to social organizations. Arrows indicate the stepwise or cross-hierarchy formation or degradation of higher-hierarchy CBMs.
Figure 1.
Schematic evolutionary pathway of carbon-based materials (CBMs) from carbon atoms and carbon-containing molecules (CCMs) to social organizations. Arrows indicate the stepwise or cross-hierarchy formation or degradation of higher-hierarchy CBMs.
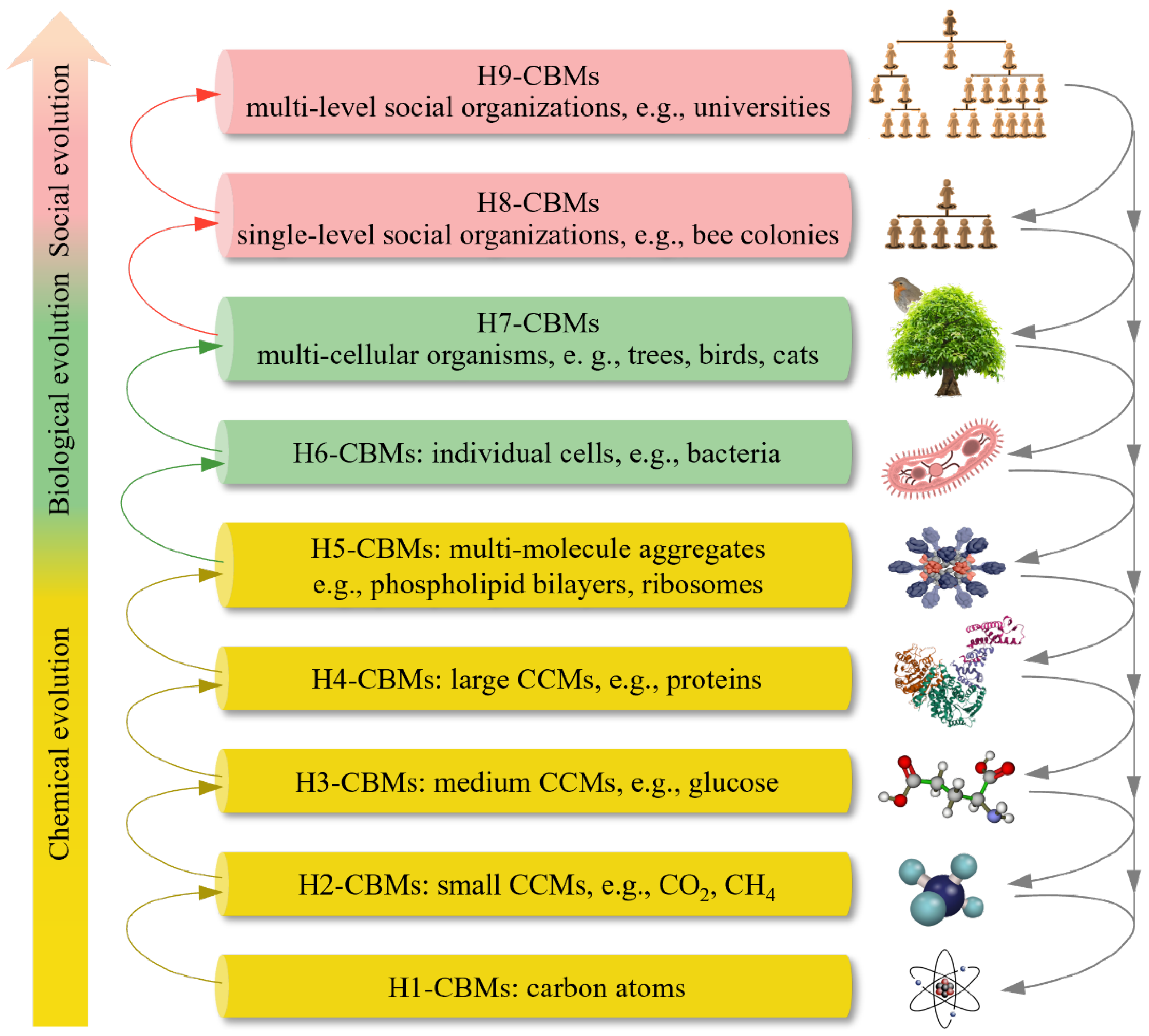
Figure 2.
The reasoning steps of CBET and Darwinian theory. *: Exemplified by the functions of catalysis, information coding, and reproduction of carbon-based materials (CBMs).
Figure 2.
The reasoning steps of CBET and Darwinian theory. *: Exemplified by the functions of catalysis, information coding, and reproduction of carbon-based materials (CBMs).
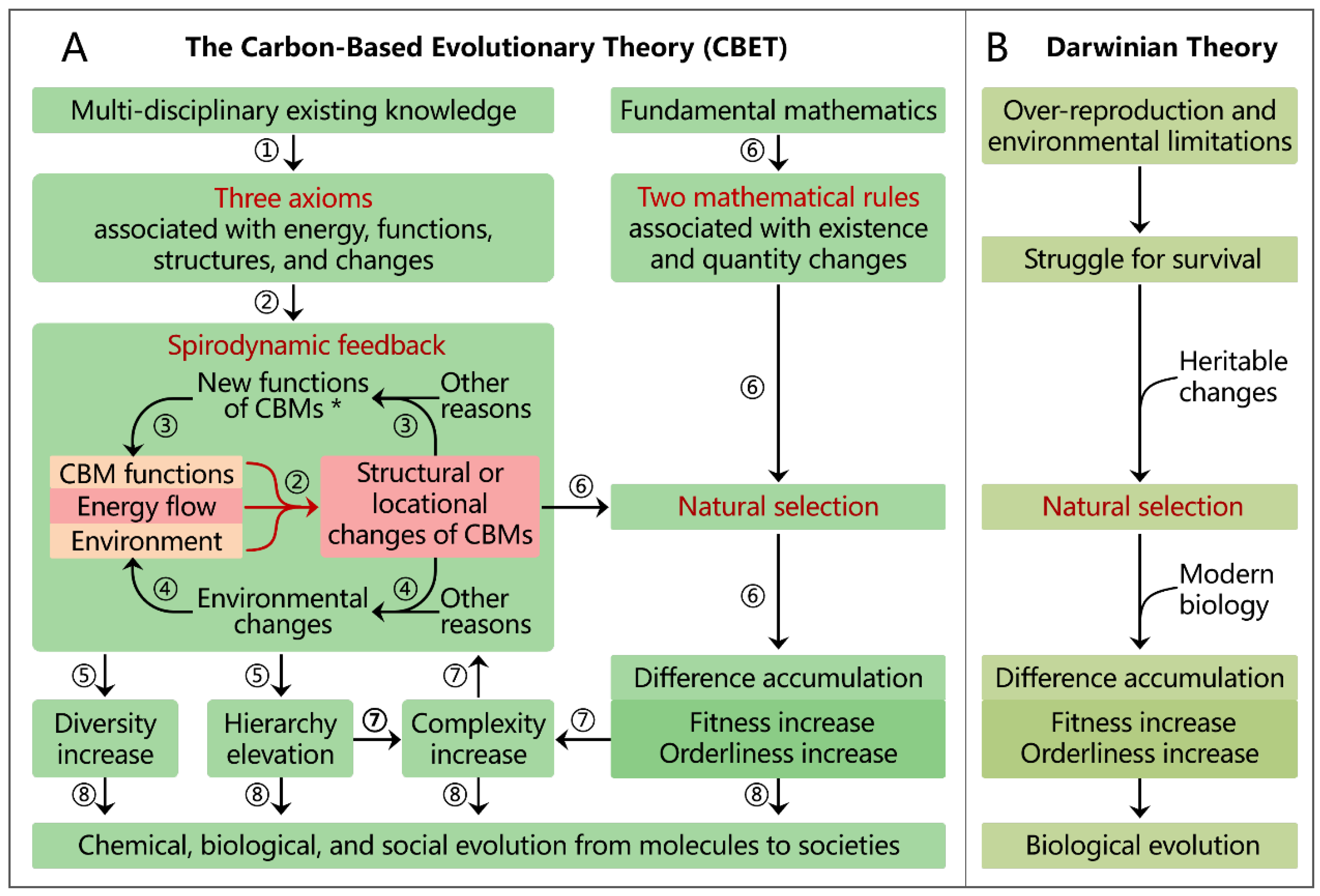
Figure 3.
The methodology of Darwinian theory and the Carbon-Based Evolutionary Theory (CBET) starting from integrative reasoning. CBMs: carbon-based materials.
Figure 3.
The methodology of Darwinian theory and the Carbon-Based Evolutionary Theory (CBET) starting from integrative reasoning. CBMs: carbon-based materials.
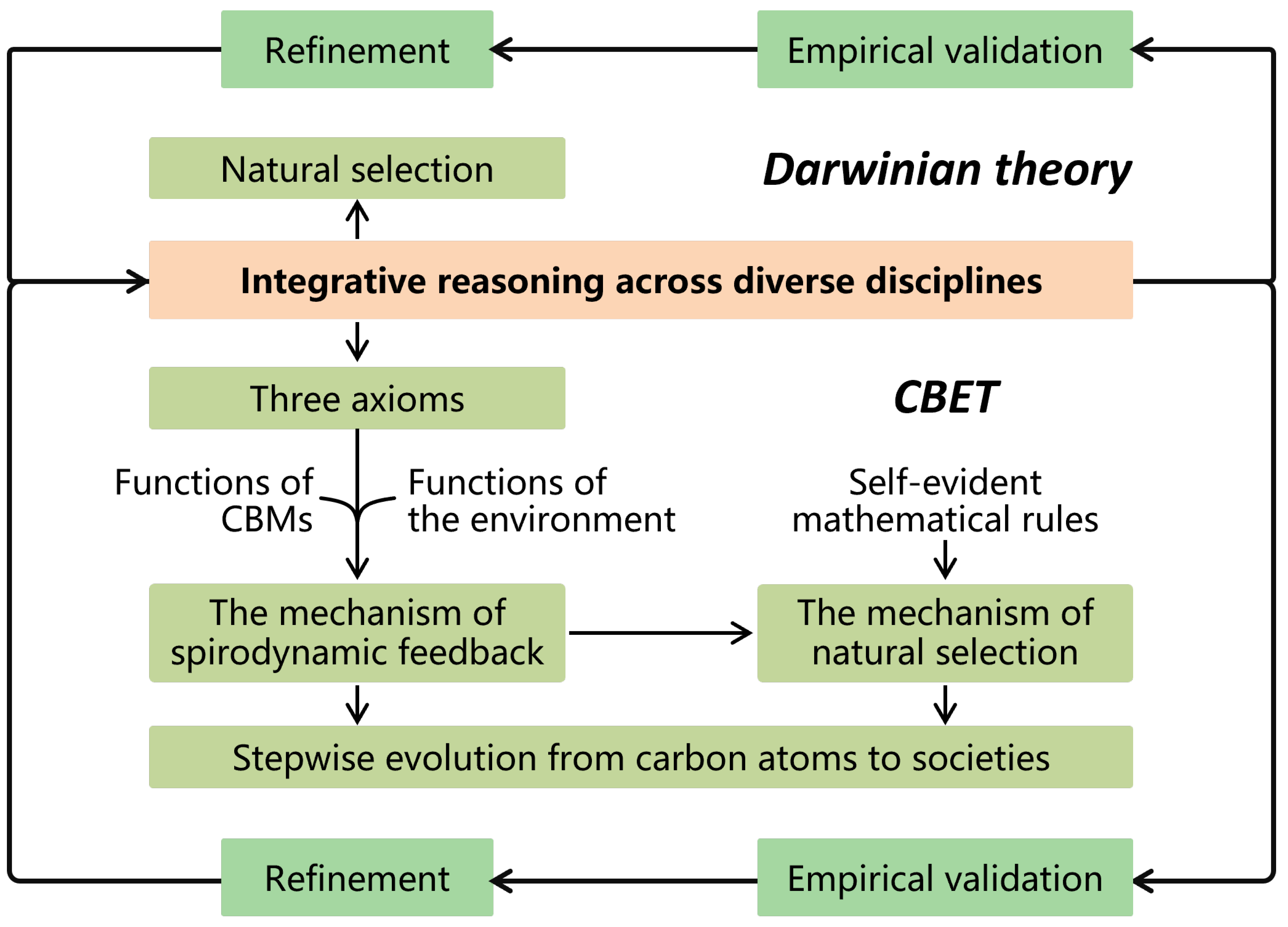
Figure 4.
Changes in the quantities of six mock carbon-based materials (CBMs) according to their fitness values under natural selection. The fitness and quantity changes of these CBMs are described in Table 3.
Figure 4.
Changes in the quantities of six mock carbon-based materials (CBMs) according to their fitness values under natural selection. The fitness and quantity changes of these CBMs are described in Table 3.
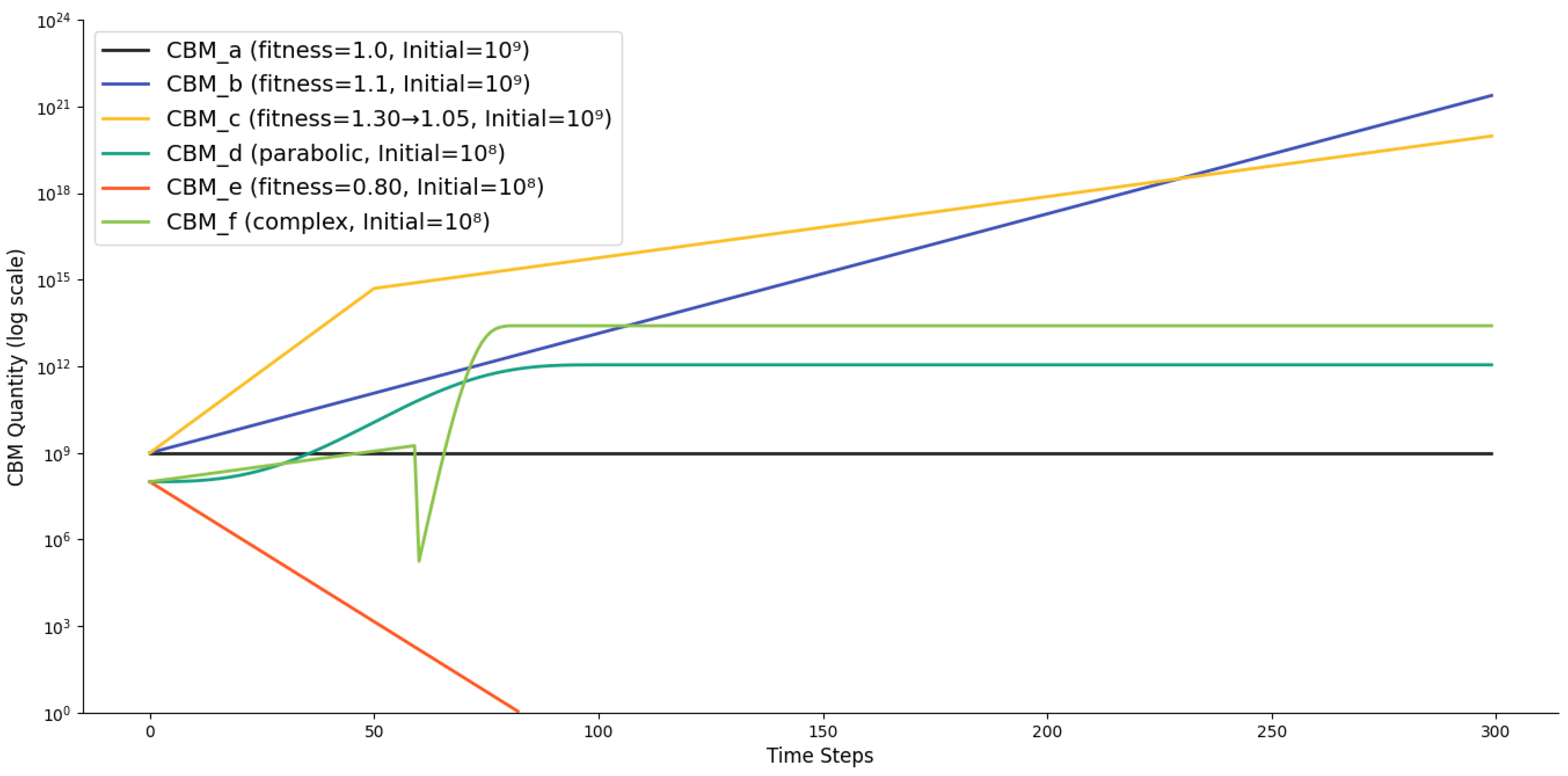

Table 1.
Differences between two theories of evolution in biology.
| Darwinian theory | The Carbon-Based Evolutionary Theory | |
| Significance | Challenged creationism and inheritance of acquired traits | Challenged creationism and inheritance of acquired traits |
| Core concept | Organisms | Carbon-based materials (CBMs) |
| Driving force | Natural selection | Energy flow under the modulation of functions of CBMs and the environment |
| Mechanisms | Natural selection | Spirodynamic feedback and natural selection |
| Fitness calculation | The number of an individual's offspring in the next generation | The formation to degradation ratio of a CBM over a time interval |
| Life’s origin | Lacks an account for life’s origin | Schematically accounts for life’s origin |
| Inclusiveness | Lacks an explicit account for neutral or detrimental mutations | Neutral or detrimental mutations can persist if relevant CBMs are fit enough |
| Non-heritable changes | Focuses on heritable genetic changes | Considers heritable, non-heritable, and epigenetic changes |
Table 2.
The 27 representative facts for the primary validation of CBET.
| Order | Description of the fact |
| 1 | Charcoal combustion which consumes oxygen and forms carbon dioxide |
| 2 | Diamond synthesis from graphite under certain conditions [33] |
| 3 | Water adsorption of activated carbon |
| 4 | CO2, NH3, and other molecules form amino acids via chemical reactions [2,5] |
| 5 | Methane explosion in coal mines |
| 6 | Evaporation of ethanol from an open bottle |
| 7 | The dehydration condensation of amino acids into peptides in tubes |
| 8 | The binding of gentamicin to the 30S ribosomal subunit of some bacteria |
| 9 | The degradation of glucose into carbon dioxide and water in cells |
| 10 | Some recombinant proteins self-assembling into virus-like particles |
| 11 | DNA synthesis catalyzed by a protein |
| 12 | Starch degradation in the stomach |
| 13 | Inputting certain DNA molecules into certain cells can create new viruses [34] |
| 14 | Inputting certain DNA molecules into certain cells can create new cells [35] |
| 15 | The formation of bilayer vesicles by amphiphilic lipid molecules |
| 16 | The development of a fertilized egg into a tadpole |
| 17 | Escherichia coli absorbs glucose via its membrane carrier proteins |
| 18 | The reproduction of bacteria within chicken intestines |
| 19 | A group of ant individuals form a eusocial ant colony |
| 20 | Some trees undergo spring budding and autumn leaf shedding |
| 21 | A car collision causes human casualties |
| 22 | The formation of a basketball team by a group of young people |
| 23 | Leafcutter ants harvest foliage to cultivate symbiotic fungi in their nests |
| 24 | The destruction of a beehive by a forest fire |
| 25 | Several institutions merged into Paris-Saclay University in 2018 |
| 26 | University of the Arts Philadelphia was closed in 2024 |
| 27 | The ongoing war between Russia and Ukraine |
Table 3.
The quantity changes in six mock CBMs with different fitness under the effect of natural selection.
Table 3.
The quantity changes in six mock CBMs with different fitness under the effect of natural selection.
| Representative significance of the CBM | |
| CBM_a | The baseline scenario where the formation/degradation rates are perfectly balanced (fitness=1.0 for all time steps). CBM_a can be a species maintaining constant population size in a stable environment. |
| CBM_b | The continuous exponential growth by 10% every time step (fitness=1.10 for all time steps). CBM_b can be a bacterium beginning to grow in a broth with abundant materials and energy. |
| CBM_c | The fitness drops from 1.30 to 1.05, which can be a sign of a species adapting to environmental changes like climate shift slowing the rapid population growth of the species. |
| CBM_d | The smooth parabola shows accelerating growth followed by decelerating growth to stabilization in the population of the CBM, which can be a bacterium that has grown in a broth, with its population reaching a peak. |
| CBM_e | The curve demonstrates the inevitable fate of the CBM (fitness=0.80 for all time steps) with sustained negative growth in its population. CBM_e can be endangered species with consistently low reproductive success. |
| CBM_f | This CBM can represent those species (e.g., humans) whose fitness oscillates around 1.0 for a certain period. This phase is followed by a sharp decline in fitness to 0.05 due to a catastrophe, which causes the population to drop by approximately 95%. Subsequently, the fitness recovers rapidly—reaching a high of 3.0—before gradually returning to the earlier oscillating pattern. During this recovery stage, the population shows rapid exponential growth and eventually surpasses its original size. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



