Submitted:
26 May 2026
Posted:
26 May 2026
You are already at the latest version
Abstract
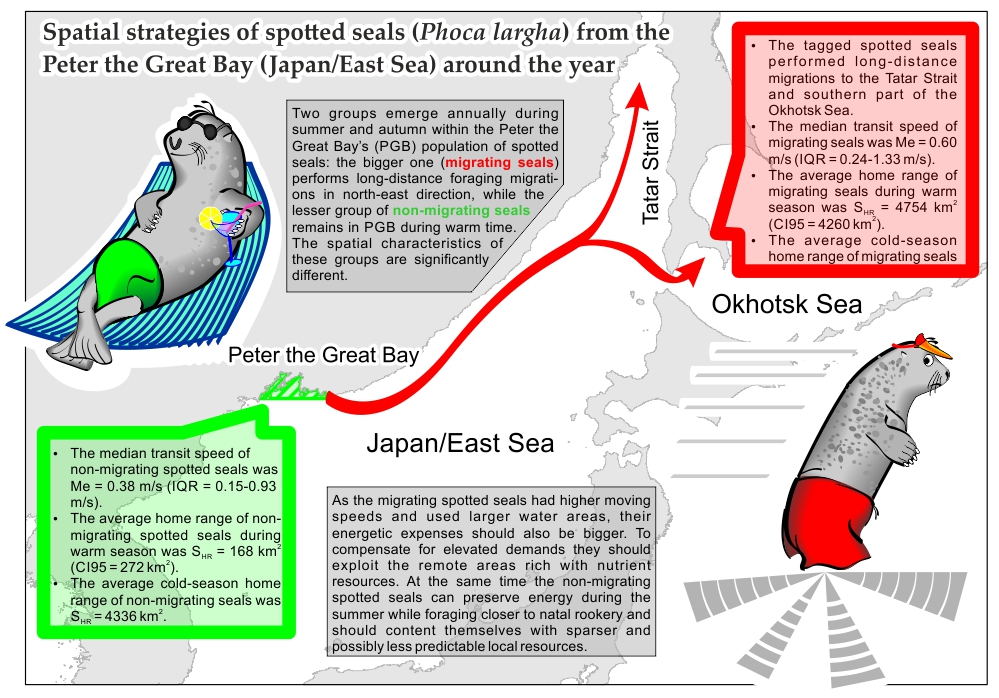
Spotted seals demonstrate unusual ecological feature within population of the Peter the Great Bay (PGB). After the breeding and molting seasons, one their group performs long-distance foraging migrations; another group remains in the bay year-round. To investigate how utilization of space differs between these two groups in 2017-2022 we deployed nine satellite tags on juvenile spotted seals within PGB: of these four seals were tagged at rookery, located at Rimsky-Korsakov Archipelago (RKA), while the remaining were tagged at haul-out of Verkhovskogo Islands (VI). Spotted seals tagged at RKA undertook long summer migrations towards north-east of the Japan/East Sea (JES) and then to south of the Okhotsk Sea. Four spotted seals tagged at VI remained in PGB through warm season. Seals were allocated into two clusters on basis of median speed during transit traveling. A relatively slow group was formed by spotted seals that remained in PBG. Also the spotted seals from this group conducted fewer transit passages and used less extent water areas. Thus non-migrating seals conserved energy, while seals that left PGB showed stronger energetic spending. Presumably, the migrating seals can compensate high energetic demands through foraging in remote feeding grounds with abundant nutrient resources.
Keywords:
spotted seal (Phoca largha)
; summer migrations
; foraging strategies
; the Japan/East Sea (JES)
1. Introduction
The spotted seal (Phoca largha Pallas, 1811) is an abundant species of Phocid seals (Phocidae Gray, 1825) that inhabits near-shore waters in the Northern part of Pacific Ocean and in pacific sector of Arctic [1]. There are eight isolated reproductive centers (populations) of spotted seal within its range with three populations recognized within the Bering Sea, two in the Okhotsk Sea, two in the Japan/East Sea (hereinafter: JES) and one in the Yellow Sea [2]. Through most of its range the spotted seal leads a pagophilic way of life, breeding and nursing their offspring on floating ice. Nevertheless, the population that inhabits the Peter the Great Bay (north-west of the JES; hereinafter: PGB) routinely experiences a limited and short-living ice covering in the bay which does not favor its reproduction. Thereby the PGB population of spotted seals is forced to breed at costal rookery of Rimsky-Korsakov Archipelago (hereinafter: RKA) [3]. Currently the archipelago is protected by conservation area of Far Eastern Marine Reserve. The PGB spotted seals can be distinguished from spotted seals of other populations by larger body size, peculiar coloration of new-born pelt and a number of meristic characteristics [3,4]. The spotted seals’ population of PGB is the most southern and less-abundant in Russia. In 1970th it was assessed in just several hundreds of individuals [4], but during last half of century it grew in numbers up to 4 thousands individuals [5]. The observed grows possibly came due to protective measures applied by Far Eastern Marine Reserve upon the rookery of local spotted seals. The breading season of spotted seal in PGB does not depend on ice conditions and is very prolonged, spanning from January to April [3,6]. At the end of spring and in early summer, after the breeding and molting is finished, the spotted seals leave PGB en-masse and travel to their summer-autumn feeding grounds at north of the JES and at south of the Okhotsk Sea [3,7]. Some spotted seas also travel in south direction along the Korean Peninsula. During the summer-autumn period only 20% of population remains in PGB [8]. This remaining part of population is suggested to be called “resident” in contrast to the “migrating” seals that leave PGB [8]. There is a speculation about possible genetic isolation running between these groups [9]. The adult spotted seals tend to return to same islands within RKA and a number of individually recognizable (marked) seals remained at the rookery through consecutive summer seasons [8]. The migratory routes and spatial characteristics of migrating spotted seals from PGB are strongly understudied. Recently only three juvenile seals from PGB were tagged by satellite tags, and all of them undertook long foraging migrations [7]. Information about spatial characteristics of seals remaining in PGB during the summer-autumn season lacks completely.
The main goal of our work is to describe possible summer foraging strategies of spotted seals from population of PGB. The objective was to study spatial and speed characteristics of spotted seals around the year, to reveal possible basic discrepancies among juvenile seals concerning the ways they use water area and to describe parameters in which they differ between each other.
2. Materials and Methods
To assess specific spatial characteristics of spotted seals from PGB population in 2017-2022 we deployed satellite tags on 9 spotted seals of which 5 were weaned under-yearling seals and 4 were juvenile seals aged between 1 to 2 years (Table 1). The tagging was carried out after the breeding season was complete. Four seals were tagged at the rookery of RKA, while the remaining 5 were tagged at the nearby haul-outs (Verkhovskogo Isls.; hereinafter: VI) (Figure 1). We used satellite tags SPOT-293A (ARGOS SPOT, Wildlife Computers, Inc., USA). Tags were attached to the crown area of a head of each seal using Loctite™ Type 422 glue (Henkel Corp., Germany), or using two-component epoxy glue. After the tagging was completed seals were released immediately.
All received location data were pre-processed by means of ARGOS’ Kalman filtering algorithm and underwent additional pre-processing by using the SDA-filtration algorithm [10] which is implemented in “argosfilter” R package [11]. Threshold values for SDA parameters to remove records from dataset were accepted as follows: speed above 3.8 m/s at distances above 5 km [12]; angles between locations more than15o at distances above 2.5 km and more than 25o at distances above 5 km. Additionally we removed records that had a speed above 10 m/s (irrelative to distances passed) or were found 1 km away from shoreline within the dry land.
The installed satellite tags fall away in process of annual molting of seals but sometime it can be lost earlier due to different reasons. In this case there is a possibility that tags lost in a tidal zone will continue to send data on its location because its wet/dry sensor will continue to switch tag on and off following the diurnal tidal cycles. To remove such records from the dataset we tested each tag by keeping it switched on for a day in open space at the platform with precisely measured coordinates. Coordinates were measured using GPS/GLONASS handheld navigation gear Garmin eTrex 30x. For each received record we calculated the Euclidean distances (deviations) between observed (i.e., provided by ARGOS) and true (GPS/GLONASS-measured) coordinates. The deviations were collected into a control set of bias (Table 1). After that for each received field record we measured a deviation of its observed coordinates from “moving” center of masses, which was calculated as a 5-day moving average on X and Y coordinates. These deviations were combined into a sequences of 5-day sets calculated in a way analogous to “moving” procedure and then were compared against control set of bias using t-test (α = 0.05). Whenever the difference between 5-day sets of observed deviations and the control set of bias showed no statistically meaningful difference for 30 consecutive days the tag was considered being lost in tidal zone.
Statistically meaningful areas for spotted seals were delineated using method of hot-spots [13]. Polygons with a side length of 5 km were considered significant when value of assigned statistic Gi* conformed to Z(Gi*)≥2.58 (hereinafter: key areas). Within designated key areas spotted seals spend significantly (α = 0.05) more time than in other parts of a range. Same method was used to describe locations with statistically significant changes in horizontal speed of spotted seals. Locations were considered “cold” (Z(Gi*)≤-2.58) or “hot” (Z(Gi*)≥2.58) which corresponded to significant (α = 0.05) decrease (in “cold” spots) or rise (in “hot” spots) in horizontal speed of a seal. A repeated decrease in horizontal speeds was assumed as a marker for foraging (when seal was spending more time submerged) or resting behavior. The key areas with clusters of “cold” spots within it were recognized as main (foraging) grounds for individual spotted seals. All other key areas were considered secondary.
To study the intra-seasonal changes in spatial distribution of spotted seals during different seasons we delineated their home ranges (the areas within which individual spotted seals can be found with 95% probability; hereinafter: HR) [14]). The “warm” season was assumed to span from May to October (or after accomplishment of summer migration in migrating spotted seals); the “chill” season started in November (or after accomplishment of reverse autumnal migration in migrating spotted seals). The Kernel Density procedure (cell size of raster was set to 1500 m) was applied to smooth the density of probability.
A direct comparison of speeds between spotted seals would be misleading due to a complex nature of their behavior. Thereby we compared only speeds observed during transit passages. To distinguish the transit passages we divided tracks of individual spotted seals into sequences of 8-hour periods, upon which applied the Rayleigh Tests for Uniformity [15]. The Rayleigh Tests highlights the deviation from uniform distribution in circular statistics and thus allows to judge about existence of average travel direction within each 8-hour traveling period. In case that average travel direction did exist all records within a tested 8-hour period were considered belonging to transit passage. To assess the probability P associated with Rayleigh R we used its exponential approximation. Then all spotted seals were assigned to one of two k-clusters using the k-means method [16]. Clustering was made based on spotted seals’ median speed during transit passages. The Euclidean distance was used for proximity function dE.
To compare the difference between datasets we used permutation test [17]. The critical value α for achieved significance level (ASL) was set at 0.05.
All calculations were done for Asia North Equidistant Conic projection; illustrative materials were drawn in WGS-84.
3. Results
3.1. Migration of the Seals from Verkhovskogo Islands
Five spotted seals aged between weaned under-yearlings and 2-years old were tagged between 24 May and 7 June at the haul-out of VI. The installed tags remained functional during 21 to 276 days (Table 1). Spotted seals #36694, #36695 and #55009 generally remained within boundaries of PGB and rarely crossed 200 m isobaths while moving at sea (Figure 2a) (hereinafter: “non-migrating”). Using of water area by female #36694 was followed until next spring. In June-October female inhabited a limited water area adjusted to VI from south. In November spotted seal began to undertake passages into seaward part of PGB, towards the depth drop-off (Figure 2b). The HR of this female was SHR = 105 km2 during warm season and SHR = 4336 km2 during the chill season (Table 2).
Spatial characteristics of male #36695 and female #55009 were studied only through warm season. Both spotted seals spent summer time in compact areas. The male #36695 generally inhabited the water area spanned between VI and Rikorda I. (SHR = 88 km2; Figure 2c). After its tag ceased to send signals (on 14 July, 2017) the male #36695 was re-sighted by opportunistic divers in 29 September, 2019 at VI [20]. Registration was photo-confirmed and seal was identified from photo using a pattern of spots on its pelt. The tag remained attached to a head of the seal during the contact. By the end of May, 2019, the female #55009 moved to Rifovaya Bay and lived there until the end of August (SHR = 312 km2; Figure 2d). The female regularly visited nearby Vostok Bay and south-eastern end of Putyatin I., presumably to attend the haul-out situated at Pyat Paltsev Rock and Iretskogo I. The tag of #55009 worked smoothly until 30 August, 2019, but then it ceased to send signals and turned on again on 10 October, 2019. The tag continued to function until March, 2020, but during the remaining phase of its work the deviations of tag’s locations from 5-day “moving” center of mass did not surpass a control set of measured bias (p < 0.05). Data from the last phase of the tag’s service were removed from the dataset.
The remaining spotted seals from the group tagged at VI (#55010 and #55014) left PGB, moving within the shelf area (Figure 3). Female #55010 left PGB on 19 June, 2019, and went eastward along the coast of Primorye Region. Spatial characteristic of this spotted seal were not assessed due to insufficiency of data. Last registration of #55010 was received in the end of June, which assumes that the female could start its foraging migration, but it was not tracked due to a short service time of the tag.
The yearling female #55014 left PGB in June, 2019, and went more than 270 km in north-east direction. Then she suddenly ceased her movement and returned to PGB. Until the January, 2020, she remained within main key area adjacent to Povorotny Cape (most eastern point of PGB). During the warm season HR of female #55014 was SHR = 2986 km2 (Table 2). Main feeding ground of the seal was situated around Povorotny Cape and to the west of Putyatin I. (Figure 3; see Key areas and Low-velocity spots). After November HR of that spotted seal began to grow in size and by January, 2020 it increased up to SHR = 4460 km2 (Table 2). In the beginning of 2020 female left PGB again and moved in south-west direction until it reached Yongdae-gap, 340 km away from tagging spot (Figure 3). While moving along the coast of Korea female #55014 formed several secondary key areas within which she did not slow her pace in statistically meaningful way. The tag ceased to send signals in 24 February, 2020.
3.2. Migration of the Seals from Rimskiy-Korsakov Archipelago
Information on spatial and speed characteristics of three juvenile spotted seals that were tagged in 2017 (Table 1) was partially used in this analysis and was published earlier [7]. All these seals undertook long foraging migrations to north of the JES and to south of the Okhotsk Sea during the warm season (Figure 1).
The under-yearling female #36693 was tagged at RKA on 25 May, 2022 (Table 1). Female left PGB three days later and went alongside the JES cost towards Tatar Strait. On 19-22 June female crossed Tatar Str. between Peschany Cape and Crillon Cape and on 23 June she entered its main foraging area that spread in waters of Japan/East and Okhotsk seas, both sides from La Perouse Str. (see Figure 1, Key areas). Female #36693 spent most of its foraging time at Rayatomari Bank and to south from Crillon Cape (Figure 1) where she repeatedly moved from Sakhalin I. to Hokkaido I., and in reverse, along the local belt of cold waters that emerges annually in summer along the front of Soya Warm Current [21,22,23]. During the warm season HR of spotted seal was SHR = 4447 km2 (Table 2). The seal died within fishing net in Rishiri Suidō in 5 October, 2022 [24].
3.3. Spatial and Speed Characteristics of the Peter the Great Bay Spotted Seals
A proportion of transit traveling records within activity budget of all nine spotted seals partially overlapped (2-25%% transit records in spotted seals tagged at VI, against 15-21%% tagged at RKA (Table 3)). Its medians did not differ statistically between tagging spots (permutation test, ASL > 0.05). The median speeds of transit traveling were 0.50-0.88 m/s and 0.93-1.03 m/s, correspondingly. Median speeds of seals tagged at VI and PGB were significantly different (ASL < 0.05), which was a reason after allocation of spotted seals in two different groups (clusters). When all spotted seals were assigned into clusters on basis of their median speeds the intra-group centroids were 0.59 m/s for the first k-cluster and 0.94 m/s – for the second; the proximity function was dE = 0.20 m/s. After the pooling of all locations by k-clusters the median transit speed of seals within first k-cluster was Me = 0.38 m/s (IQR = 0.15-0.93 m/s), and Me = 0.60 m/s (IQR = 0.24-1.33 m/s) within second k-cluster. The difference between median transit speeds in k-clusters was statistically significant (ASL < 0.05). First k-cluster was composed by spotted seals that did not leave PGB (non-migrating spotted seals #55009, #36694 and #36695), while the second k-cluster included all other spotted seals (hereinafter: migrating spotted seals) (Table 3).
The spotted seals tagged on VI haul-out were distributed between k-clusters, which mean the factor of place of tagging did not play role in seals’ tendencies towards using high or low traveling speeds. Likewise no effect had the age of spotted seals, since grouping by the age (“under-yearlings” and “older than 1 year”) was not statistically meaningful between k-clusters (Fisher’s Exact Test, P = 0.36). Migrating spotted seals shoved generally high level of activity: besides the high speeds of traveling they had a higher proportion of transit records (in average 19% against 7% in non-migrating spotted seals (Table 3)). They also used wider water areas during warm season: in average non-migrating spotted seals’ HR were only SHR = 168 km2 (CI95 = 272 km2) while migrating spotted seals used tens time bigger HR (SHR = 4754 km2, CI95 = 4260 km2) (Table 2). Warm season water areas were significantly different between seals in k-clusters (ASL < 0.05).
Movements of one non-migrating (#36694) and three migrating (#94842, #148661 and #55014) spotted seals were tracked through chill season (Table 2). During chill time the sizes of water areas used by spotted seals were comparable between k-clusters (SHR = 4336 km2 and SHR = 7120 km2, correspondingly) and did not differ statistically (ASL > 0.05). During the late autumn water areas that were used by non-migrating spotted seals grew up significantly – from several hundreds to thousands of square kilometers (for example, see Figure 2b). Unlike the non-migrating spotted seals, the migrating seals tend to use bigger water areas regardless of a season (Table 2).
4. Discussion
4.1. Migrating and Non-Migrating Spotted Seals of the Peter the Great Bay
Some discrepancies in exploitation of water area by spotted seals inhabiting PGB were disclosed long ago. The assumption that in summer and autumn spotted seals can leave PGB dates back to the 1980th [25]. Summer-time re-distribution of spotted seals along the continental shelf of the JES was suggested as an evidence for foraging migrations of spotted seals to north-east along the coast of the Primorsky Kray [26]. Nesterenko and Katin [8] assessed number of spotted seals remaining in PGB in summer as 450-500 individuals, which at that moment accounted for 20% of population. The re-sighting of branded and tagged individuals indicated that in summer spotted seals leave PGB for prolonged migrations [5]. The most recent research of migratory behavior of PGB spotted seals used satellite tagging allowed to assert the existence of stable route of foraging migrations towards Tatar Strait and to south of the Okhotsk Sea [7].
The observed difference between median speeds of spotted seals from two k-clusters of seals has a high biological importance. The migrating spotted seals showed much higher level of activity during their transit passages (58% higher than spotted seals that spent warm season in PGB) which should translate into significantly higher energetic spending if compared to non-migrating seals. The proportion of transit records in activity budget and the size of water areas used during warm season were also higher in migrating spotted seals (Table 2). It should be noted that exploitation of small water areas by juvenile spotted seals can be typical for non-migrating individuals as the same was observed in other population of spotted seal [27].
The reasons of dissimilarity in spatial and speed characteristics of migrating and non-migrating spotted seals are not completely clear. Recent research of genetics of PGB spotted seals revealed at least two mitochondrial lines, which allowed authors to suggest genetic isolation of spotted seals in PGB [9]. Such isolation could have occurred historically as a result of sea level changes due to glaciations in Pleistocene or later. The reports on repetitive observations of the same individually recognizable spotted seals in PGB year-by-year in summer [8] also suggest a certain inter-annual stability of non-migrating spotted seals. The most plausible explanation for supposed existence of migrating and non-migrating ecological groups within PGB population of spotted seals would be found in history of the JES region. The Quaternary period was characterized by significant changes in sea levels as a consequence of regular regressions which by some accounts could lead to a complete or near-complete isolation of the JES from the Pacific Ocean. The result of such isolation was emergence of a closed water body with strongly altered oceanographic parameters [28,29,30,31,32]. If spotted seals did inhabit this isolated water body – which is a probable assumption though lacks corroboration by solid evidences, – they would be forced to adapt to specific conditions of Quaternary JES, possibly gaining some inclination to a settled way of life in process. Later, when the straits of the JES opened, the newly obtained trait could be preserved as a feature characteristic to a group of spotted seals that inhabited the JES through glaciations. In other words, the arguable non-migrating group can be a relic of Quaternary period in the JES. The other group of ancestors of PGB population of spotted seals could go through glaciations in adjacent water areas and was less constrained spatially. This group could preserve its ability to commit long migrations. This hypothesis demands a through-out research of genetics of both ecological groups of PGB spotted seals and would require sampling of individual spotted seals whose affiliation to one or another ecological group is undisputable.
At the moment, there are no reliable evidences confirming that non-migrating spotted seals from PGB belong to a separate genetic line. It is possible that their group emerge annually as a response to multiple factors. If assumed that non-migrating spotted seals belong to a stabile demographic unit within PGB population, their group can be expected to gain a competitive superiority in form of lower energetic spending which can result in their numerical dominance over migrating spotted seals within a conceivable time [33]. But as long as only a fifth of population remains in PGB in summer [8] the suggestion about competitive superiority of non-migrating group is questionable. Then a possible explanation for a low number of spotted seals remaining in PGB in summer can be based on relatively lower availability and abundance of local nutrient resources. For example, in summer some generally cold-water prey items that form a base for spotted seals diet (such as walleye pollack (Theragra chalcogramma) and navaga cod (Eleginus gracilis)) tend to migrate beneath 50 m isobaths [34], which can temporarily deplete their abundance in shallow water areas of PGB. Possibly to compensate for a summer-time decrease in availability of fish in their ration the spotted seals of PGB are forced to resort to a trophic specialization at which they base their diet on cephalopods, such as Japanese common squid (Todarodes pacificus) [8]. After early 1990-th Japanese common squid dominate in biomass of nekton in the JES [35], but its annual runs into PGB can be very inconsistent depending on short-term oceanographic conditions [36]. This makes it unreliable source of nutrients for population of spotted seals in PGB.
4.2. The Strategies of Summer Foraging
The observed differences in spatial and speed characteristics of migrating and non-migrating spotted seals may be determined by the difference in strategies of obtaining and spending energy, which, in turn, results from opposite strategies of summer foraging. We suppose that a conservative approach is more typical for non-migrating spotted seals, while a high-cost approach is used mainly by migrating spotted seals. The allocation of seals into one or another group may not be pre-determined at the beginning of warm season and may depend on many variables. Juvenile spotted seals may be dependent on the extended breeding season, typical for local seal population, which leads to the fact that pups born in different time are facing different sets of ecological conditions (temperature, availability of nutrients, anthropogenic pressure, etc.) when they get weaned. For spotted seals that already switched to external sources of feeding the factors of exogenous nature that determine availability of feeding resources may also be relevant. Summer time energetic demands of adult spotted seals are high because in addition to regular vital needs they should use additional energy to build-up their internally stored energetic resource of fat which allow them to participate in reproduction which in Phocid seals align to “capital” breeding strategy [37] whereas the nutrients required for spotted seals’ foraging can be limited during warm season in PGB. That can force spotted seals to leave PGB for the areas rich with nutrient resources. The tendency to nurse offspring at mid-latitude during the spring, and then to forage and regain a lost body mass at zones of increased productivity was observed in different “capital-breeder” marine mammals, such as in northern elephant seal (Mirounga angustirostris) and grey whale (Eschrichtius robustus) [1,37,38]. The migration of spotted seals into waters of Tatar Strait and south of the Okhotsk Sea after completion of their breeding season may be a specific adaptation to seasonably variable conditions of inhabitance in PGB which allows abating intra-population competition for feeding resources and hauling-out grounds, though makes competition with neighboring populations in shared feeding areas more acute.
Individual spotted seals may be affected by factors of endogenous nature. For example, some seals of non-migrating group may be unfit to begin long travels because of physiologic issues, such as health conditions, insufficient amount of internally stored energetic reserves, etc. The endogenous reasons can contribute to differences in seasonal use of water areas by migrating and non-migrating spotted seals: the seals remaining in PGB find themselves in ambient temperature that stand closer to their body temperature, which is less expansive energetically for thermoregulation and maintaining the metabolic rates [37]. On the other hand, the migrating spotted seals spend summer in colder northern areas where they face altered energetic demands due to lower ambient temperature and should show higher foraging activity to be able to replenish a loss of the energy and keep blubber layer thick enough to provide thermal insulation and serve an internal reserve of the energy. This assumption aligns with our observation that the size of water area used by non-migrating spotted seals grow sharply from warm to chill season, while the area used by migrating spotted seals (that spend both summer and winter in cold waters) remains comparable in size inter-seasonally. The areas used by the seals from both groups do not differ statistically during the chill season.
It worth noting that some spotted seals can show peculiar “hybrid” strategies. For example, female #55014 in summer stayed within PGB but kept high movement speed and used big water areas. At the same time she did not perform (possibly ceased) foraging migration and never formed main key areas outside of PGB (Figure 3).
5. Conclusions
Our observations on the spotted seals of the Peter the Great Bay, the Japan/East Sea using satellites tags confirmed that the population during warm season divides in two groups of the seals with different behavior. In total 9 seals were tagged during 2017-2022 in two locations of the bay: Rimsky-Korsakov Archipelago (main rookery) and Verkhovskogo Islands (non-breeding haul-outs). One group of the spotted seals left the bay for forage migration in summer and travelled far to the northeast up to the Tatar Strait and the Okhotsk Sea. The group came back in late autumn. The second group of the spotted seals remained in the bay all year round. The seals of these two groups demonstrated different speed during their transit travelling. A relatively slow group was formed by spotted seals that remained in PBG. In addition to slower movements spotted seals from this group conducted fewer transit passages and used less extent water areas. The migrating spotted seals have elevated energetic demands due to higher level of their activity, which manifest itself in higher traveling speeds and bigger sizes of HRs. They compensate it through exploiting the remote northern areas rich with nutrient resources. At the same time the non-migrating spotted seals can preserve energy during the summer while foraging closer to natal rookery, where they tend to move at slower pace, within less extended areas. During warm season non-migrating spotted seals should content themselves with sparser local resources. Long foraging migrations outside of PGB allow local spotted seals to ease intra-population competition for nutrient resources, though at a cost of increased competition with spotted seals from neighboring populations. Our study is the first to show that seals breeding in the same location can exhibit markedly different utilization of space, resulting in the divergence of their energetic strategies. The underlying causes of this segregation remain unknown and represent a promising avenue for future research.
Author Contributions
Conceptualization, A.T., P.P., S.R., V.L., H.K. and S.K.; methodology, A.T., P.P. and S.R.; software, P.P. and S.R.; validation, P.P. and S.R.; formal analysis, P.P.; investigation, A.T., P.P., S.R. and H.K.; resources, H.K. and S.K.; data curation, S.R.; writing—original draft preparation, A.T., P.P. and V.L.; writing—review and editing, A.T., P.P., S.R., V.L., H.K. and S.K.; visualization, P.P.; supervision, A.T.; project administration, A.T.; funding acquisition, V.L. and S.K. All authors have read and agreed to the published version of the manuscript.
Funding
The study was funded jointly by POI FEB RAS (state project No. 124022100077-0) and the National Institute of Fisheries Science, Ministry of Oceans and Fisheries, Republic of Korea (R2026004).
Institutional Review Board Statement
Animal handling and tag attachment procedures were permitted by the Federal Agency for Fisheries of the Ministry of Agriculture of the Russian Federation and approved by the Pacific Fisheries Research Center (order No. 818 of 05.12.2017). Application and exploitation of satellite tags was permitted by Federal Service for Technical and Export Control of Russia (conclusion No. 316 of 12.02.2018). All tags were registered by Federal Service for Supervision of Communications, Information Technology and Mass Media, Russia (certificates 25 18 No. 06279, 25 18 No. 06280, 25 18 No. 06281). Authors declare that they have no conflict of interest.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original data for three spotted seals tagged in 2017 are available at Mendeley Data, https://doi.org/10.17632/b46bjcdtmd.2 was introduced in Trukhin, et al. [7]. The original data for six spotted seals tagged in 2019-2022 are available at Mendeley Data, https://doi.org/10.17632/t9zrchj5hx.2.
Acknowledgments
We are deeply grateful to Dr. Kobayashi M. of Tokyo University of Agriculture and Mr. Lubyanichenko S. of V.I. Il’ichev Pacific Oceanological Institute (FEB RAS) for sharing with us information on re-sighting of tagged spotted seals. We express our sincere gratitude to associates of Far Eastern Marine Reserve (MNRE Russia) Dolganov S., Minov S. and Svintsitskiy E. and to technicians of V.I. Il’ichev Pacific Oceanological Institute (FEB RAS) Pavin L. and Lubyanichenko S. for their assistance in our field work.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| JES | the Japan/East Sea |
| PGB | Peter the Great Bay |
| RKA | Rimsky-Korsakov Archipelago |
| VI | Verkhovskogo Islands |
| HR | home range |
| SHR | area of home range |
| ASL | achieved significance level |
| Me | median |
| IQR | interquartile range |
| CI95 | 95% confidence interval |
References
- Belikov, S.E.; Burkanov, V.N.; Varentsov, M.I.; Varentsova, N.A.; Vladimirov, V.A.; Glazov, D.M.; Danilov, M.B.; Evdokimov, A.A.; Zagretdinova, D.R.; Iliushin, D.G.; Isachenko, A.I.; Kornev, S.I.; Kochi, K.V.; Kuznetsova, D.M.; Logetskaya, M.S.; Svetochev, V.N.; Solovieva, M.A.; Trukhin, A.M.; Udovik, D.A.; Filatova, O.A.; Shpak, O.V.; Shulezhko, T.S. Marine mammals of Russian Arctic and Far East. 2017, Moskow: Arkticheskii Nauchnii Center. 311 p.; in Russian.
- Shaughnessy, P.D.; Fay, F.H. A review of the taxonomy and nomenclature of North Pacific Harbour seals. Journal of Zoology, London 1977, 182, pp. 385-419. [CrossRef]
- Trukhin, A.M. Spotted seal. 2005, Vladivostok: Dalnauka. 246 p.; In Russian.
- Kosygin, G.M.; Tihomirov, E.A. Largha (Phoca largha Pallas) in the Peter the Great Bay. Izvestiya TINRO 1970, 70, pp. 114-137; in Russian.
- Trukhin, A.M. Current status of the spotted seal (Phoca largha) population in the Peter the Great Bay, Sea of Japan. Mammal Study 2024, 49, pp. 257-271. [CrossRef]
- Trukhin, A.M. Changes in the Breeding Range of Spotted Seals, Phoca largha (Pallas, 1811) (Carnivora, Pinnipedia), in the Western Sea of Japan: Causes and Effects. Russ J Mar Biol 2022, 48(2), pp. 101-107. [CrossRef]
- Trukhin, A.M.; Permyakov, P.A.; Ryazanov, S.D.; Lobanov, V.B.; Kim, H.W.; Choi, Y.M.; Sohn, H. Migrations of young spotted seals (Phoca largha) from Peter the Great Bay, Sea of Japan/East Sea, and the pattern of their use of seasonal habitats. PLoS ONE 2021, 16(1), e0244232. [CrossRef]
- Nesterenko, V.A.; Katin, I.O. The spotted seal (Phoca largha) in the Peter the Great Bay. 2014, Vladivostok: Dalnauka. 219 p.; In Russian.
- Podlesnykh, A.V.; Katin, I.O. Genetic variation in the spotted seal (Phoca largha Pallas, 1811) from the Rimsky-Korsakov Archipelago (Peter the Great Bay, western sea of Japan) as inferred from mitochondrial DNA control region sequences. Zoologischer Anzeiger [A Journal of Comparative Zoology] 2022, 301, pp. 174-178. [CrossRef]
- Freitas, C.; Lydersen, C.; Fedak, M.A.; Kovacs, K.M. A simple new algorithm to filter marine mammal Argos locations. Maine Mammal Science 2008, 24(2), pp. 315-325, 315. [CrossRef]
- R Development Core Team. R: A language and environment for statistical computing.; R Foundation for Statistical Computing: Vienna, Austria, 2007.
- Chugunkov, D.I. Observation on common seal of Utkinsk rookery. Izvestiya TINRO 1970, 70, pp. 164-168; in Russian.
- Getis, A.; Ord, J.K. The Analysis of Spatial Association by Use of Distance Statistics. Geographical Analysis 1992, 24, pp. 189-206. [CrossRef]
- Worton, B.J. Kernel Methods for Estimating the Utilization Distribution in Home-Range Studies. Ecology 1989, 70, pp. 164-168. [CrossRef]
- Zar, J.H. Biostatistical analysis, 5th ed.; Pearson Prentice Hall: New Jersey, 2010; 944 p.
- Mandel, I.D. Cluster analysis. 1988, Moskow: Finansy i Statistika. 176 p.; in Russian.
- Efron, B.; Tibshirani, R.J. An Introduction to the Bootstrap; Chapman & Hall: London, 1993; 456 p.
- OpenStreet. Map Data. 2019 [cited 2022 13 April]; Available from: https://osmdata.openstreetmap.de.
- GEBCO. Maps. 2020 [cited 2022 13 April]; Available from: https://www.gebco.net.
- Lubyanichenko, S.N. (Pacific Oceanological Institute (FEB RAS), Vladivostok, Russia), Personal communication, 2019.
- Ishizu, M.; Kitade, Y.; Matsuyama, M. Characteristics of the cold-water belt formed off Soya Warm Current. J. Geophys. Res. 2008, 113, C12010. [CrossRef]
- Mitnuk, L.M.; Shevchenko, G.V.; Sofienko, Y.A.; Dubina, V.A. On tidal origin of belt of cold waters in area of a Kamen’ Opasnosti Rock within La Perouse Strait. Issledovaniya Zemli iz kosmosa 2006, 5, pp. 86-96; in Russian.
- Supranovich, T.I.; Yurasov, G.I.; Kantakov, G.A. Non-periodical currents and water exchange in La Perouse Strait. Meteorol. Gidrol. 2001, 3, pp. 80-84; in Russian.
- Kobayashi, M. (Tokyo University of Agriculture, Tokyo, Japan), Personal communication, 2022.
- Trukhin, A.M.; Kosygin, G.M. Materials about distribution and population of largha seals in the Peter the Great Bay. Nauchno-issledovatelskie raboti po morskim mlekopitayushchim severnoi chasti Tihogo okeana v 1986/87 1988, pp. 97-103; In Russian.
- Trukhin, A.M.; Mizuno, A.W. Distribution and abundance of the largha seal (Phoca largha Pall.) on the coast of Primorye Region (Russia): a literature review and survey report. Mammal Study 2002, 27(1), pp. 1-14. [CrossRef]
- Kim, H.W.; Uh, N.; Lee, S. First satellite tracking of a free-ranging spotted seal (Phoca largha) from the Baengnyeongdo Island. Fisheries and Aquatic Sciences 2023, 26(11), pp. 669-677. [CrossRef]
- Gorbarenko, S.A.; Shi, X.; Bosin, A.A.; Liu, Y.; Artemova, A.V.; Zou, J.; Yanchenko, E.A.; Vasilenko, Y.P.; Wu, Y.; Vladimirov, A.S. Relative sea level changes during the Last Glacial Maximum and deglaciation (33-15 ka) inferred from the δ18O records of planktic foraminifera from the Sea of Japan. Quat Sci Rev 2022, 279, 107386. [CrossRef]
- Ikehara, K. Late Quaternary Seasonal Sea-Ice History of the Northeastern Japan Sea. J Oceanogr 2003, 59, pp. 585-593.
- Ikehara, K.; Fujine, K. Fluctuations in the Late Quaternary East Asian winter monsoon recorded in sediment records of surface water cooling in the northern Japan Sea. J Quat Sci 2012, 27(9), pp. 866-872. [CrossRef]
- Oba, T.; Irino, T. Sea level at the Last Glacial Maximum, constrained by oxygen isotopic curves of planktonic foraminifera in the Japan Sea. J Quat Sci 2012, 27(9), pp. 941-947. [CrossRef]
- Park, S.-C.; Yoo, D.-G.; Lee, C.-K.; Lee, E. Last glacial sea-level changes and paleogeography of the Korea (Tsushima) Strait. Geo-Mar Lett 2000, 20(2), pp. 64-71. [CrossRef]
- Solbrig, O.T.; Solbrig, D.J. Introduction to population biology and evolution; A.D. Bazikin, ed. 1982, Moskow: Mir. 488 p.; in Russian.
- Shuntov, V.P.; Volvenko, I.V.; Kulik, V.V.; Bocharov, L.N. Bentic macrofauna of Peter the Great Bay (Japan/East Sea): occurance, abundance, and biomass. 1978-2009; V.P. Shuntov and L.N. Bocharov, eds. 2014, Vladivostok: TINRO-Centre. 307 p.; in Russian.
- Zuenko, Y.I. Fisheries oceanography of the Japan Sea. 2008, Vladivostok: TINRO-Center. 227 p.; in Russian.
- Bulygin, V.V.; Ribnikova, I.G. Distribution and catches of pacific flying squid in Peter the Great Bay (Sea of Japan) during the summer-autumn period. Nauchnie trudi Dalribvtuza 2016, 39, pp. 3-8; in Russian.
- Davis, R.W. Marine Mammals: Adaptations for an Aquatic Life; Springer Nature: Cham (Switzerland), 2019; 302 p. [CrossRef]
- Holser, R.R.; Favilla, A.B.; Crocker, D.E.; Costa, D.P. Tracking interannual variation of a large-scale ocean front influences foraging in a mesopelagic predator. Ecosphere 2026, 17(4), e70599. [CrossRef]
Figure 1.
Spatial distribution of spotted seals that committed long foraging migrations from the Peter the Great Bay (PGB). #94842, #148660 and #148661 adopted from Trukhin, et al. [7]; #36693 – our data.
Figure 1.
Spatial distribution of spotted seals that committed long foraging migrations from the Peter the Great Bay (PGB). #94842, #148660 and #148661 adopted from Trukhin, et al. [7]; #36693 – our data.

Figure 2.
Spatial distribution of locations of spotted seals tagged at Verkhovskogo Isls. (VI) that remained in PGB: for all “resident” spotted seals (a), and for individual seals #36694 (b), #36695 (c) and #55009 (d).
Figure 2.
Spatial distribution of locations of spotted seals tagged at Verkhovskogo Isls. (VI) that remained in PGB: for all “resident” spotted seals (a), and for individual seals #36694 (b), #36695 (c) and #55009 (d).

Figure 3.
Spatial distribution of spotted seals tagged at VI that left PGB.


Table 1.
Information on the tagged spotted seals and on working time of each tag.
| Releasing date | PTT | Working time a tag, days |
Tagging area 1 | Sex of seal | Age of seal, month |
Number of datums accepted for research |
| 18.05.2017 | #94842 2 | 316 | RKA | F | 2.5-3 | 2780 |
| 18.05.2017 | #148660 2 | 207 | RKA | M | >12 | 2154 |
| 19.05.2017 | #148661 2 | 351 | RKA | M | 2.5-3 | 3146 |
| 24.05.2019 | #55009 | 98 | VI | F | 3-4 | 1944 |
| 01.06.2019 | #36695 | 43 | VI | M | >24 | 761 |
| 03.06.2019 | #36694 | 276 | VI | F | >12 | 4602 |
| 04.06.2019 | #55014 | 265 | VI | F | >12 | 4827 |
| 07.06.2019 | #55010 | 21 | VI | F | 3-4 | 440 |
| 25.05.2022 | #36693 | 133 | RKA | F | 2.5-3 | 2928 |
1 RKA – Rimsky-Korsakov Archipelago; VI – Verkhovskogo Isls. 2 adopted from Trukhin, et al. [7], except the Number of datums accepted for research.
Table 2.
Areas of home ranges for spotted seals from different k-clusters.
| PTT | Season | k-cluster | Area of HR, km2 |
Average area of HR 1, km2 |
| #36694 | warm | 1 | 105 | 168 |
| #36695 | warm | 1 | 88 | |
| #55009 | warm | 1 | 312 | |
| #148660 | warm | 2 | 5472 | 4 754 |
| #148661 | warm | 2 | 9638 | |
| #36693 | warm | 2 | 4447 | |
| #55014 | warm | 2 | 2986 | |
| #94842 | warm | 2 | 1229 | |
| #36694 | chill | 1 | 4336 | 4 336 |
| #148661 | chill | 2 | 5780 | 7 120 |
| #55014 | chill | 2 | 4460 | |
| #94842 | chill | 2 | 11,121 |
1 - grouping was done between seasons and k-clusters.
Table 3.
Grouping of spotted seals into k-clusters on basis of their median speed.
| PTT | Place of tagging | Proportion of transit traveling records, % |
Median transit traveling speed, m/s |
k-cluster |
| #94842 | RKA | 15% | 0.96 | 2 |
| #148660 | RKA | 21% | 0.93 | 2 |
| #148661 | RKA | 19% | 1.01 | 2 |
| #55009 | VI | 2% | 0.65 | 1 |
| #36695 | VI | 9% | 0.5 | 1 |
| #36694 | VI | 10% | 0.62 | 1 |
| #55014 | VI | 16% | 0.88 | 2 |
| #55010 | VI | 25% | 0.82 | 2 |
| #36693 | RKA | 18% | 1.03 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



