Submitted:
19 May 2026
Posted:
20 May 2026
You are already at the latest version
Abstract
Maize is a major global crop; however, its production is affected by Fusarium verticillioides (Fv), which causes stalk, ear, and root rot (SERR). Bacillus cereus B25 is a maize bacterium that antagonizes Fv, likely through antifungal compounds and possibly by inducing of maize chitinase genes. Fv effectively infects maize by producing a chitinase-modifying protein (Cmp) that disrupts maize chitinases, preventing fungal cell wall degradation and evasion of plant immune responses triggered by PAMPs. The aim of this work was to analyze maize, B25, and Fv gene expression during bipartite and tripartite interactions at early stages (5, 7, 10, and 14 days post-inoculation). Physiological results showed increased root and shoot growth in maize seedlings under the tripartite interactions compared to Fv alone. B25 was demonstrated to grow endophytically and coexist with Fv in maize roots. Maize extracellular chitinase genes were induced, possibly due to chitin fragments released from the fungal cell wall, while Fv genes were also upregulated in response. Furthermore, the chitinase gene Zm00001eb317090 (bk4) may contribute to cell wall strengthening, as suggested by in silico co-expression analyses. Overall, these results support a coordinated interaction between maize and B25 that contributes to controlling Fv infection.
Keywords:
chitinase
; expression
; maize
; fungus
; bacteria
1. Introduction
Fusarium verticillioides (Fv) is one of the main pathogenic fungi that affects maize crops. This fungus belongs to the Nectriaceae family, and it belongs to the Fusarium fujikuroi species complex (FFSC) [1]. Fv is a facultative endophytic fungus which can be found inside the maize plant without presenting any symptoms of infection [2]. In addition, Fv can switch to a necrotrophic stage where the fungus biomass increases resulting in necrosis of infected tissues, Fv infection in grain produces high amounts of mycotoxins (FB1, FB2, and FB3) [3]. FB1 mycotoxin is the most abundant toxin of Fv affecting both animal and human health [4]. FB1 inhibits ceramide synthesis and sphingolipid metabolism, causing the accumulation of reactive oxygen species (ROS) and subsequently cell death [5].
In the systemic infection stage of Fv, conidia produce which can be accumulated in the cells of the root vascular tissue and reaches the different plant tissues through the stem causing infection of the entire plant [6]. Since Fv is a fungus that can live inside the maize tissues and conidia can form inside the vascular vessels [7], its growth control becomes challenging. Chemical applications of fungicide only limit its infection progress, but do not prevent the fungus from invading the plant or producing mycotoxins. However, biological control has been implemented as an alternative to the use of chemical compounds, which can be accumulated in soils and have indirect or direct effects on the environment [8]. Biological control acts using beneficial living microorganisms to prevent or control any fungal infection and this alternative also minimizes pollution by chemical compounds [9]. Bacillus cereus sensu lato B25 was isolated from the maize rhizosphere; it was previously shown that B25 is a plant growth-promoting bacteria and controls the Fv infection [10,11]. B25 has several antagonistic mechanisms against Fv, which include genes for production of biofilm, siderophore, antibiotics and lytic enzymes [12]. Both B25 chitinase genes (A and B) are induced by chitin or Fv lysates [13], or when B25 is confronted directly to Fv [14] and, also, both B25 chitinase recombinant proteins inhibit Fv conidia germination when applied directly [15].
Maize plant chitinases play an important role as a surveillance mechanism for the presence of fungi and can monitor Fv infection, when a fungus is attacking a plant, chitinases degrade chitin on their cell walls producing chito-oligomers which act as elicitors [16]. These elicitors are available for detection by receptor proteins on plant cells, also called pattern recognition receptors (PRRs) [17]. Nevertheless, it has been discovered that some fungi can avoid the hydrolysis of their cell wall by plant chitinases by inducing effectors that can detect and modify chitinases, such as fungalysin (Fvcmp) from Fv [18]. Effectors recognize specific domains of certain extracellular chitinases (ZmChitA and ZmChitB) causing the separation of the chitin binding domain from the catalytic domain and making maize chitinases unable to bind chitin in the fungal cell wall, and therefore, avoiding the release of chitin elicitors (chito-oligosaccharides - COSs) which work as Pathogen Associated Molecular Patterns (PAMPs) that help the plant to induce its own defense system which allows Fv infection to proceed. On the other hand, several fungal effectors capable of suppressing programmed cell death and the plant hypersensitive response have been reported, including CFEM (Common in Fungal Extracellular Membrane) domain-containing proteins, such as CfEC12 and CgCFEM from Colletotrichum graminicola [19,20]. These effectors can also suppress chitin-triggered reactive oxygen species (ROS) accumulation and alter the expression patterns of defense-related genes, for example, CgCFEM1 from Colletotrichum gloeosporioides [21]. However, despite these advances, the biological functions of CFEM proteins in F. verticillioides remain poorly understood.
In a recent study [22], it was demonstrated that ten chitinase genes are induced in 7-day old maize seedlings when infected with Fv, including four genes encoding extracellular chitinases which could be targeted by the fungal fungalysin Fvcmp. This study aims to elucidate the role of B25 during the expression of maize chitinase genes in root samples infected with B25, Fv and with B25/Fv, as well as to analyze the expression of other genes such as the putative maize chito-oligosaccharide (PRR) receptor (ZmCEBiP) and the fungal Fvcmp, CFEM and Fvsep through a time course analysis. We also present evidence that B25 is an endophytic bacterium and can be detected in the xylem vessels, the same niche that Fv occupies when infection of this fungus advances in the plant. These findings suggest a direct interaction between the plant, the fungus and its bacterial control agent inside the maize root and opens new avenues to explain the mechanisms underlying multiple plant-microorganism interactions.
2. Materials and Methods
2.1. Bacterial and Fungal Growth
The bacterial strain, Bacillus cereus sensu lato B25, was isolated from the maize rhizosphere and maintained at -80 °C as a glycerol stock [10]. The bacterium was re-animated in Luria Bertani agar at 28 °C for 16 hours. Fusarium verticillioides strain P03 [6] was grown in potato dextrose agar (PDA) at 24 °C for fourteen days to induce conidia production. Conidia were collected using sterilized water and a Drigalski loop and adjusted to 1 x 106 conidia/mL. B25 was grown in 5 mL of LB liquid media for 16 h at 28 °C. A 250 mL baffled flask containing 100 mL of LB liquid media was inoculated with 1 mL of B25 pre-inoculum and grown at 28 °C for 24 h. B25 spores powder formulation was obtained [23].
2.2. B25 and Fv Colonization Assay in Maize Roots
To visualize the endophytic location of B25 and Fv inside the maize roots a bipartite interaction assay was performed for each of the plant-microorganism interactions. 200 grams of sterile sand was used as substrate containing 1x105 Fv conidia per gram for the maize/Fv interaction. White maize seeds (Asgrow hybrid “Hipopótamo”) were previously disinfected [6]. The maize seeds were treated with 0.2 mL of 1% (w/v) carboxy methyl cellulose (CMC) solution used as an adhesive agent. Seeds were coated with 1 g of B25 powder formulation spores (1 x 109 spores g-1) obtaining coating of ~1 x106 UFC/seed [23]. Control maize plants and maize/Fv plants were inoculated with the base powder formulation without B25. The experiment consisted of three inoculated seeds per magenta box. The boxes were incubated at 25 °C for 16 hours light and 8 hours darkness at 20 °C. After two weeks, the plants were harvested, and roots were taken for further analysis. For confocal laser microscopy, images were taken from B25 or Fv inoculated plant roots. The wheat germ agglutinin (WGA) conjugated to the fluorophore Alexa Fluor 488™ was used to stain the cell walls of both microorganisms. Root samples were red fluorescence counter-stained with rhodamine B (plant cell wall staining). Leica TCS SP5 confocal laser microscope with a white laser was used for capturing the green fluorescence emitted at 511-541 nm by WGA-488 after excitation at 499 nm, and the red counterstain with rhodamine B by exciting it at 573 nm and registering emission at 593-623 nm. In addition, 100 mg of fresh root tissue was superficially disinfected to quantify endophytic B25 colony-forming units (CFU). Roots were washed with tap water and surface-sterilized by sequential immersion in 70% ethanol for 1 min and 1% hypochlorite for 10 min. Samples were then immersed in 10% (v/v) Tween-20 solution for 1 min and rinsed three times with sterile distilled water. The last wash was used directly to plate as a control to demonstrate the efficiency of the surface sterilization procedure. Subsequently, root tissues were ground in a mortar containing 1 mL of sterile distilled water. Serial dilutions were prepared and plated on LBA medium for bacterial quantification.
2.3. Rolled Paper Assay for Tripartite Interaction
Surface-disinfested white maize seeds (Asgrow hybrid “Hipopótamo”) were coated with the B25 powder formulation as described in the previous section. For the rolled paper assay [24] four treatments were established for each time-course point: 1) control maize seeds (inoculated with base powder formulation without B25 spores); 2) maize seeds inoculated with a B. cereus B25 spores powder formulation; 3) maize seeds inoculated with 10 μL of Fv conidia suspension (1x103 conidia/seed) and base powder formulation without B25 spores; and 4) maize seeds inoculated with both microorganisms. The treatments were maintained in a growth chamber with controlled conditions at 25 + 2 °C with a light-dark cycle of 14-h light and 10-h darkness. Maize roots were collected at 5, 7, 10 and 14 days post inoculation (dpi). Each treatment had two biological replicates with four plants per replicate for every time point collected. A disease rating system [23] was employed in which 0 = 0% root rot; 1 = 1-25% root rot; 2 = 26-50% root rot; 3 = 51-75% root rot; 4 = 76-100% root rot; 5 = root system death; and 6 = whole seeding death. Length and fresh weight measurement of plant roots and shoots were also taken. Roots were ground in liquid nitrogen for RNA isolation.
2.4. Primer Design for Chitinase Detection
The design of the primers for the real-time PCR assays was carried out using the PRIMER3Plus software website [25]. The primers were designed using the coding sequence (CDS) of the genes and the 3’UTR region to obtain gene target specificity for each chitinase gene [22]. The housekeeping genes of maize, the cyclin-dependent kinase (Zm00001eb350890) and Fv β-tubulin, were used as internal control expression genes (Table 1).
2.5. RNA Extraction and cDNA Synthesis
Total RNA was isolated from roots of four plants from each biological replicate at each time point. The root samples were ground in liquid nitrogen using a mortar and pestle until a fine powder was obtained. TRIzol was used to obtain total RNA from all root samples from 100 mg of tissue for maize gene expression analysis. For Fv gene expression analysis, total RNA was obtained from root samples with Fv infection using RNeasy Plant Mini Kit (Cat. No. 74904, QIAGEN). RNA was quantitated using a Nanodrop 2000c (Thermo Fisher Scientific, Wilmington, DE, USA) and RNA integrity determined by agarose gel electrophoresis. 1 μg of RNA was treated with 1 unit of RQ1 DNase (PROMEGA, Cat. No. M6101, Fitchburg, WI, USA) to avoid DNA contamination. First-strand cDNA was synthesized from total RNA using oligo(dT) and SuperScript III reverse transcriptase (Thermo Fisher Scientific, Cat. No. 18080-044, Waltham, MA, USA) following the manufacturer’s instructions. Finally, cDNA was adjusted to a concentration of 10 ng/μL and a 1 μL aliquot was used for real-time PCR (qPCR) analyses.
2.6. Quantitative Real Time-PCR (qRT-PCR)
qRT-PCR reactions were performed using QuantiNova™ SYBR PCR kit (Qiagen, Cat. No. 208052, Hilden, Germany) in a reaction that included 5 μL of SYBR Green, 0.2 mM of each primer, 10 ng of cDNA and RNase-free water for a final volume of 10 μL. Each reaction was carried out in triplicate for each biological replicate. The reaction was carried out in a Rotor Gene-Q Real Time PCR System (Qiagen, Cat. No. 9001550, Hilden, Germany) with a thermocycler program included a preheating step at 95 °C for 2 min, followed by 40 cycles of denaturation at 95 °C for 30 s, an annealing step at 60 °C for 30 s, and an extension step at 72 °C for 30 s. Relative quantification of each gene was normalized using the maize cyclin-dependent kinase (CKD) gene, and the comparative threshold cycle method 2-∆∆Ct [26] was used to calculate the fold change (FC) values. For Fv genes, the relative quantification was normalized using Fv β-tubulin gene, and the comparative threshold cycle method 2-∆Ct was used, and for the comparative analyses of maize-Fv vs maize-B25-Fv treatments, the threshold cycle method 2-∆∆Ct was used. All interactions graphics were design using Biorender (https://www.biorender.com).
2.7. Protein-Protein Interaction Network
STRING database (http://string-db.org) was used to generate a co-expression network [27]. Zm00001eb317090 protein sequence was submitted to the STRING database. The minimum required score was set to medium confidence (0.400). The maximum number of interactors showed no more than 20 on the first shell, and no more than 10 on the second shell.
2.8. Statistical Analysis
Rolled paper assays were analyzed by one-way ANOVA using IBM SPSS Statistics program. Differences among treatments were determined using the Tukey test at a significance level of P < 0.05.
3. Results
3.1. Biocontrol of Fv in Maize Plants by B25 Powder Formulation Spores
B25 controlled Fv infection in maize plants in different treatments in the time-course infection days (Table 2). Fv infection reduced shoot length at 5 and 10 dpi compared to the other treatments and reduced shoot fresh weight at 14 dpi compared to Zm and Zm-B25-Fv treatment. Fv also caused a significant reduction of root length at 5 dpi, and in root fresh weight at 5, 10, and 14 dpi compared to the Zm control. In maize plants inoculated with B25 and Fv (Zm-B25-Fv) there was a significant difference in root length compared to the Zm-Fv treatment at 5, 10, and 14 dpi, and the control (Zm) treatment at 10 and 14 dpi. Likewise, no significant differences were detected at 7 dpi. Maize seed treated with Fv only showed a disease index starting at 7 dpi. At 14 dpi, the disease severity index was significantly higher in the Zm-Fv than in the Zm-B25-Fv treatment (Figure 1).
3.2. B25 Colonizes Endophytically the Maize Root
B25 formed biofilm in the root apex (Figure 2A) and was present in the root epidermis (Figure 2 B) and in the root vessels (Figure 2C). We observed that both B25 and Fv (Figure 2D) grew endophytically in the root vessels of Fv and B25 independently colonized plants at 14 dpi sharing the same niche inside the plant. We were able to observe B25 moving actively in the xylem vessels (Video S1). Likewise, endophytic bacterial abundance after two weeks reached 4.7 x 104 CFU/100 mg, while last wash controls showed no bacterial growth, confirming the efficacy of the surface sterilization procedure.
We also tested the endophytic presence of the microorganisms inside root samples by placing surface sterilized root segments in LB and PDA plates finding typical bacterial or fungal growth (Figure 3; Figures S1-S3). B25 and Fv were recovered from root samples at all sampling times. More abundant B25 colonies were observed in the Zm-B25 interaction, whereas reduced Fv mycelia growth was observed in the tripartite (Zm-B25-Fv) compared to the Zm-Fv bipartite interaction.
3.3. Differential Gene Expression and protein-Protein Interaction Analysis
All chitinase genes were upregulated at some Fv infection time point (7, 10 and 14 dpi) (Table 3; Table S1 and S2) in the bi and tripartite interaction. However, the highest upregulated gene was bk4 – brittle stalk (Zm00001eb317090), followed by CEBiP (Zm00001eb002690) and chitinase 29 (Zm00001eb168350). bk4 – brittle stalk was highly expressed in Zm-B25 and Zm-B25-Fv compared to its expression in Zm-Fv. Chitinase 29, a cytoplasmic chitinase of unknown function, was induced at 5 dpi in all interactions and at 7 dpi it was only induced in the Zm-B25 and Zm-B25-Fv interactions, at 10 dpi this gene was upregulated in the Zm-Fv treatment and at 14 dpi it showed an induction only in the tripartite interaction. Chitinase 27 (Zm00001eb167340), an extracellular chitinase which does not contain the protein domain recognized by the chitinase modifying proteins, was induced at 5 dpi in the bipartite treatments, at 7 dpi in the Zm-B25 and Zm-B25-Fv interactions, at 10 dpi in the bipartite interactions, and in all conditions at 14 dpi with the highest induction in the Zm-B25 treatment.
Four fungalysin (Fvcmp)-targeted maize chitinases were analyzed: 1) ChitA (Zm00001eb078730) was upregulated at 10 dpi, being mostly induced by Fv, and at 14 dpi, the Zm-Fv treatment was the only one induced. 2) ChitB (Zm00001eb425600) was induced by day 7 in the tripartite treatment, at 10 dpi in the Zm-B25 treatment, and at 14 dpi in all treatments. 3) chitinase 21 – class I (Zm00001eb346860), which also has an Fvcmp site, was highly expressed at 7 dpi and remained induced at 10 dpi in the Zm-Fv and Zm-B25-Fv interaction, and at 14 dpi it was slightly induced only in the Zm-B25 interaction. 4) EPR4 (Zm00001eb246640) was induced at 7 and 14 dpi in all interactions and induced only in the bipartite interactions at 10 dpi.
Class I chitinases are proposed to be targeted not only by fungalysin but also by subtilisin-like proteins [28]. Therefore, a search was conducted to find a gene ortholog of F. oxysporum (FOX_09801) in the Fv genome. An orthologous gene was found with 95.98% of nucleic acid identity (FVEG_08679) to the one from F. oxysporum and it was denominated Fvsep (serine protease). Fvsep (FVEG_08679) was upregulated at 7 dpi in the Zm-B25-Fv with a fold change of 2.50. Fv CFEM gene, coding for a putative effector possibly involved in suppressing chitin-triggered reactive oxygen species (ROS) accumulation, was induced in all interaction’s days in Zm-Fv with a high FC value at 14 dpi. However, this gene was also induced in the tripartite interactions at 5, 10 and 14 dpi. Fvcmp always showed low levels of expression.
In addition, the putative maize chito-oligosaccharide (PRR) receptor CEBiP was induced since 5 dpi but the induction was highest in Zm-B25 and Zm-B25-Fv compared to Zm-Fv treatment and continued being induced at 7 dpi in the Zm-B25 and Zm-B25-Fv treatments and only in the Zm-B25 interaction at 10 dpi.
Due to the high induction of bk4 brittle stalk (Zm00001eb317090), a protein interaction analysis was performed to find which are the genes that interact with this protein (Figure 4). We found nine possible protein interactors: bk2 brittle stalk, B4FVH0, A0A1D6JR13, A0A1D6F557 and five cellulose-synthase (CesA10, 11, 12, 13 and 14-A0A1D6GBL0) proteins. Most of these proteins are involved in cell wall biosynthesis. B4FVH0 (Zm00001eb397000) is proposed to encode an O-acetyltransferase involved in cell wall formation; A0A1D6JR13 (Zm00001eb138390) and A0A1D6F557 (Zm00001eb112280) are plant-specific domain TIGR01627 family protein members involved in xylan biosynthesis.
STRING analysis suggested bk2 brittle stalk as a gene hub involved in cell wall synthesis/strengthening (Figure 4). Thus, we analyzed its expression by qRT-PCR and found out that this gene is up-regulated in the Zm-B25 association at 7 dpi compared to the other treatments being Zm-Fv the one with the lowest gene expression (Table 4, Table S1).
4. Discussion
Fusarium verticillioides is considered an endemic fungus of maize fields and it can prevail in soils as conidia or spores for many years [4]. In addition, Fv can infect other important agricultural crops not only maize, but also tomato [29], potato [30], sorghum [31], and sugarcane [32]. Over the years, great advances have been found regarding the mechanisms of tolerance of maize to Fv infection through cultivation techniques of tolerant genotypes, which have allowed to elucidate the molecular mechanisms underlying the maize-Fv interaction and how Fv can infect maize tissues. For example, ZmWAX gene is involved in maize resistance to Fv seed and stalk rot and cuticular wax deposition [33].
Likewise, two inbred maize lines, IL09 (susceptible, SUS) and IL02 (resistant, RES), showed different defense responses against Fv DA42 infection [34]; an increase in lignification was observed in cells within the sclerenchymatous hypodermal zone in the RES line, in addition to exhibiting a greater number of cell layers and increased cell wall thickness compared to the SUS line. In contrast, the SUS line showed pronounced degradation of root hairs, along with small discontinuities at cell-cell junctions in epidermal cells.
Synthetic fungicides may cause environmental damage due to their ability to accumulate in soils and affect the microbiota rhizosphere and can also affect aquatic ecosystems through soil lixiviation [8]. Therefore, an alternative tool could be the use of antagonistic microorganisms. There can be more than 1010 organisms in one gram of soil, so it is feasible to find one that can control fungal or viral infections, especially organisms isolated from the plant rhizosphere. There is a wide diversity of microbial communities around plant roots, which in turn participate in direct and indirect interactions [35]. However, the rhizosphere microbiota is composed mostly of proteobacteria, Ascomycota and Basidiomycota fungi [36]; the interaction between plant roots and the rhizosphere microbiota is influenced by the physical and chemical soil conditions such as pH, salinity, carbon and oxygen composition [37], and, above all, the production of root exudates.
A control agent for SERR caused by Fv, a bacterium from the maize rhizosphere, Bacillus cereus B25, was identified and demonstrated in vitro that can control Fv disease [10]. B25 also controlled SERR symptoms caused by Fv in field trials where it showed the ability to reduce fumonisins and increase grain yield [11]. B25 genome was sequenced [12] and some of B25 antagonistic mechanisms against Fv were suggested by identifying genes involved in the production of siderophores (bacillibactin and petrobactin), antibiotics (surfactin), lytic enzymes (endoglucanase and chitinases), and biofilm formation which were later corroborated by gene expression, physiological and biochemical studies [14]. B25 chitinases have been found induced in the presence of fungal lysates [13] in addition, recombinant B25 chitinases were shown to have a negative effect on Fv conidia germination [15]. Therefore, to understand which the interaction mechanisms of B25 within the maize root cells are, and the presence of Fv, we investigated if this bacterium could induce maize chitinase genes.
In the bipartite and tripartite assays, we observed the effect of B25 and Fv on maize seedling growth, the greatest root growth in length was obtained when Fv was combined with B25. Maize chitinases can participate in several plant biological processes such as seed germination, growth and fungal control [38]. The increase in plant chitinase gene expression has been observed in the presence of several phytopathogens, for example, the BjChI1 chitinase of Brassica juncea in response to Aspergillus niger [39], the chitinase (CHI) from Vitis vinifera L. in response to Aspergillus carbonarius [40] and the induction of chitinases in soybean by adding oligochitosan complexes in response to Colletotrichum truncatum [41]. The expression of chitinases in plants can be influenced by some symbiotic organisms, such as Medicago truncatula in the presence of Glomus intraradices [42] and soybean chitinases induced in response to different rhizobacteria inoculation [43], this could lead to early detection and rapid response against some pathogens.
Regarding maize chitinase genes, bk4 brittle stalk presented the highest fold change value in all treatments in our study, especially in the bacterial treatments. Notably, this gene has been reported to be involved in stalk tensile strength and could be involved in cell wall regulatory formation [44]. Likewise, this gene is also related at the co-expression level with bk2 brittle stalk, which encodes for a COBRA-LIKE4 protein, this protein can function in a lignin-cellulosic interaction [45]. Therefore, both genes could be participating during the biosynthesis of cell wall compounds.
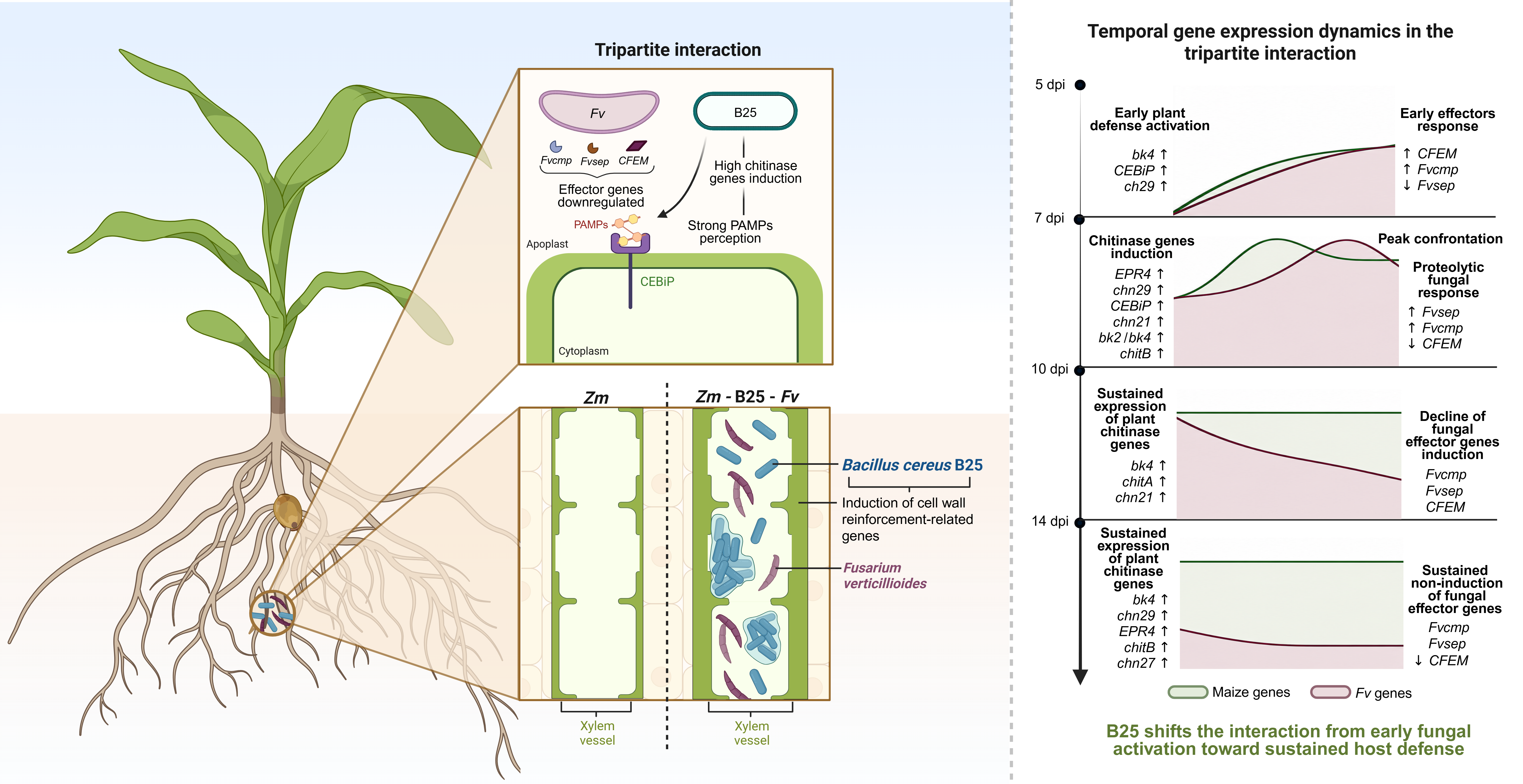
Chitinase-27 (Zm00001eb167340) has been found to be induced in maize silk samples infected with Fv [46]. Chitinase-29 (Zm00001eb168350) is downregulated in leaf samples in maize susceptible plants infected with Fv [47]; interestingly, in this study chitinase-29 was induced mostly in the bacterial treatments, suggesting that B25 may be regulating its expression, and since its induction was higher in the tripartite interaction, it may be being induced in response to Fv infection while interacting with B25. In addition, chitinase-21 (Zm00001eb346860), one of the Fvcmp target chitinases, has been found induced in the maize line (RIL165) which is susceptible to Cercospora Zeina [48]. Taking all together, we proposed an interaction model for chitinase-related gene expression between maize, B25 and Fv for each day of interaction (Figure 5).
In the presence of B25 bacteria, the highest induction of CEBiP receptor and the bk4 gene at 5 dpi suggests the plant prepares to repel pathogen attack by plant cell wall reinforcement and preparedness for PAMPs signal detection. The chn27 gene, an extracellular chitinase, gets induced at that time which could degrade chitin from fungal pathogens in case they try to colonize the plant. The same scenario persists at days 7 and 10 with bk2 being induced by B25 which could assist plant cell reinforcement even more. CEBiP remains induced and chn27 and different cmp-containing chitinases get induced at those times which should allow PAMPs generation in case of fungal invasion. Extracellular chitinases persist induced at day 14, as well as bk4. Only CEBiP is not induced but it remains possible PAMPs detection by remaining CEBiP protein sensing chito-oligosaccharides. In summary, B25 allows plants to get ready to fight fungal pathogens through plant cell wall reinforcement, PAMPs generation and PAMPs signal detection.
Zm-Fv interaction by 5 dpi induces slightly CEBiP, chn27 and chn29. This raises the question: is the extracellular chitinase chn27 able to degrade chitin from the fungal cell wall and induce PAMPs detection by the plant? At this time, also bk4 is not induced with the lack of reinforcement of the plant cell wall. The plant prepares to respond to Fv by inducing cmp-containing chitinases by 7 dpi, however, since CEBiP induction is also lost currently, it can be possible that Fv can affect PAMPs detection by an unknow mechanism, possibly mediated by CFEM proteins. To respond to fungal attack the plant induces slightly bk4 expression, but this is possibly not enough to cause a reinforcement of the plant cell wall. Fv would partially be avoiding gene induction of bk2 to lower levels than the ones induced in the B25 treatments and avoiding plant cell wall biosynthesis and reinforcement. Root biomass is affected by Fv since 10 dpi, at this time point the plant attempts to defend from fungal invasion increasing the expression of three extracellular cmp-containing site chitinases, chn27, and the cytosolic chitinase chn29, suggesting the plant tries to rescue its mechanism of plant defense mediated by plant chitinases in order to fight against Fv. A similar gene expression scenario is encountered at 14 dpi where Fv also causes a noticeable decrease in root biomass and shoot biomass. In summary, maize can response to Fv infection by inducing several chitinases genes and cell-wall related chitinases, however, this response is not enough to prevent fungal colonization.
In the Zm-B25-Fv interaction at 5 dpi only bk4, CEBiP and the cytosolic chn29 are induced. These genes remained induced at 7 dpi, but at this time three cmp-containing site chitinases and chn27 are also induced possibly causing PAMP generation and detection, bk2 is also induced enhancing plant cell wall reinforcement. At this point the fungus tries to protect itself from maize chitinases by inducing fvsep. As in the Zm-Fv interaction at 10 dpi CEBiP is not induced, but in the Zm-B25-Fv interaction bk4 induction is higher and chitA and chn21 remained induced. A different set of cmp-containing site chitinases were induced at 14 dpi: chitB and EPR4. Chn27 was also induced, as well as chn29 which was highly induced. A noticeable increase in gene expression at this time point was observed for bk4 with respect to 10 dpi (120.33 vs. 25.33-fold change). The symptoms and disease-severity index of the plants associated with both organisms are lower than in the maize associated to Fv only. It is interesting to note that the Fv CFEM gene was induced at 5 dpi with a high FC value in Zm-B25-Fv interaction compared to the bipartite Zm-Fv interaction suggesting that Fv tries counter attacking B25 antagonistic effect.
In order to understand the effect of B25 on Fv expression genes, a 2-∆∆Ct analysis was performed comparing the expression levels of Zm-B25-Fv against Zm-Fv (Figure 6, Table S3).
In summary, when the tripartite interaction occurs in roots B25 seems to slow down the advancement of Fv in the first 7 dpi by plant cell wall reinforcement and PAMP detection by inducing CEBiP, at 10 and 14 dpi plant cell wall reinforcement mediated by bk4 may continue in the plant. It is plausible to suggest that B25 could be producing chitinases that could directly affect the fungal cell wall at days 10 and 14 dpi, since fvcmp and fvsep were highly induced at 7 dpi followed by a lack of induction at days 10 and 14 post inoculation. However, B25 chitinase gene expression has remained elusive to be detectable in our maize root system. The Fvsep subtilisin-like protein may be acting together in the tripartite interaction with the fungalysin Fvcmp to modify class I and IV chitinases [28] counteracting the plant response of induction of the extracellular chitinases which could degrade the fungal cell wall. In addition, Fv CFEM proteins have been described as a dual-function effector that suppress plant immunity while contributing to fungal cell wall integrity during early stages of infection [49], this could be associated with the high-level expression of CFEM gene at 5 dpi. The induction of two B25 extracellular chitinases in response to chitin or fungal lysates [13], along with evidence that bacterial chitinases can decrease conidia germination [15] and mycelial branching of Fv [14] and that B25 and Fv appear to share the same niche in the root vascular tissue (Figure 2), suggests that when in contact inside the plant root, direct antagonistic mechanisms of B25 against Fv may help to debilitate the fungal defenses. Even when the fungus can respond by mounting an avoidance mechanism of PAMP signaling, B25 can help the plant in a coordinated dual defense. While the plant reinforces its cell wall, B25 uses all its weaponry against the fungus, including lytic fungal cell wall enzymes such as chitinases, chitosanase and endoglucanase, biofilm production, antibiotics (surfactin) and siderophores (petrobactin and bacillibactin) to delay fungal infection.
5. Conclusions
In summary, maize chitinase genes were analyzed to learn about their expression patterns in a time-course interaction assay involving the biocontrol bacterium B25, the fungal phytopathogen Fv or both together. Plant physiological analyses suggest that B25 plays a major role in controlling Fv infection. Furthermore, we demonstrate that B25 can grow endophytically inside the vasculature of maize roots, just like Fv. Additionally, maize chitinases may regulate the MAP kinase cascade through the production of PAMPs and the activation or repression of the CEBiP chito-oligosaccharides receptor. Chitinases gene expression changes were dynamic, as well as the expression of the CEBiP receptor; however, B25 may induce this PAMPs receptor as a mechanism to induce in the plant not only fungal cell-wall degrading chitinases but also those associated with plant cell wall biosynthesis to counteract the fungal infection through a still unknown mechanism that may involve plant cell-wall reinforcement. Meanwhile, Fv may protect its cell wall from degradation by releasing three proteins, CFEM, Fvcmp and Fvsep, which the last two could act in a coordinated fashion to modify Class I and IV extracellular maize chitinases. These findings highlight the maize chitinase gene expression variability during bi- and tripartite interactions in the presence of a beneficial bacterium and/or a phytopathogenic fungus.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/doi/s1. Figure S1: Growth of maize seedlings and microorganisms at 5 dpi; Figure S2: Growth of maize seedlings and microorganisms at 7 dpi; Figure S3: Growth of maize seedlings and microorganisms at 10 dpi; Table S1: Quantitative real time PCR expression (2-∆∆Ct ) of maize chitinase genes; Table S2: Quantitative real time PCR expression (2-∆Ct ) of Fusarium verticillioides genes title; Table S3: Quantitative real time PCR expression (2-∆∆Ct ) of Fusarium verticillioides genes; Video S1: endophytic growth of B25 inside xylem vessel cells.
Author Contributions
Conceptualization, J.E.C.-Á and I.E.M.-M.; methodology, J.E.C.-Á., and F.R.Q-F.; software, J.E.C.-Á, and F.R.Q-F.; validation, J.E.C.-Á; formal analysis, J.E.C.-Á, A.M.F.-L., and K.M.F.-B.; investigation, J.E.C.-Á, A.M.F.-L., and K.M.F.-B.; resources, I.E.M.-M.; data curation, J.E.C.-Á, F.R.Q-F., and I.E.M.-M.; writing—original draft preparation, J.E.C.-Á., and I.E.M.-M.; writing—review and editing, J.E.C.-Á., A.M.F.-L., and K.M.F.-B.; visualization, J.E.C.-Á., A.M.F.-L., F.R.Q-F., and K.M.F.-B.; supervision, I.E.M.-M. and K.M.F.-B.; project administration, I.E.M.-M.; funding acquisition, I.E.M.-M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by SECIHTI FOINS Fronteras de la Ciencia Grant No. 2016-01-2510, and projects SIP-IPN 20241338, 20232027, 20221357, and 20211507. J.E.C.A. was granted a PhD fellowship (No. 934735) from SECIHTI and K.M.F.B. received a Master fellowship (No. 1360647). K.M.F.B. was granted complementary fellowship support from IPN (BEIFI fellowship program).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in article/Supplementary Material.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Summerell, B. A. Resolving Fusarium: Current Status of the Genus. Annual Review of Phytopathology 2019, 57, 323–339. [CrossRef]
- Oren, L., Ezrati, S., Cohen, D., & Sharon, A. Early events in the Fusarium verticillioides-maize interaction characterized by using a green fluorescent protein-expressing transgenic isolate. Applied and Environmental Microbiology 2003, 69, 1695–1701. [CrossRef]
- Torre-Hernández, M. E. De, Sánchez-Rangel, D., Galeana-Sánchez, E., & Plasencia-de, J. Fumonisinas-síntesis y función en la interacción Fusarium verticillioides-Maíz. TIP Revista Especializada En Ciencias Químico-Biológico 2014, 17, 77–91. [CrossRef]
- Omotayo, O. P., & Babalola, O. O. (2023). Fusarium verticillioides of maize plant: Potentials of propitious phytomicrobiome as biocontrol agents. Frontiers in Fungal Biology 2023, 4, 1095765. [CrossRef]
- Zeng, H. Y., Li, C. Y., & Yao, N. Fumonisin B1: a tool for exploring the multiple functions of sphingolipids in plants. Frontiers in Plant Science 2020, 11, 1–16. [CrossRef]
- Leyva-Madrigal, K. Y., Larralde-Corona, C. P., Apodaca-Sánchez, M. A., Quiroz-Figueroa, F. R., Mexia-Bolaños, P. A., Portillo-Valenzuela, S., Ordaz-Ochoa, J., & Maldonado-Mendoza, I. E. Fusarium species from the Fusarium fujikuroi species complex involved in mixed infections of maize in Northern Sinaloa, Mexico. Journal of Phytopathology 2015, 163, 486–497. [CrossRef]
- Terna, T. P., Mohamed Nor, N. M. I., & Zakaria, L. Histopathology of corn plants infected by endophytic fungi. Biology 2022, 11. [CrossRef]
- Zubrod, J. P., Bundschuh, M., Arts, G., Brühl, C. A., Imfeld, G., Knäbel, A., Payraudeau, S., Rasmussen, J. J., Rohr, J., Scharmüller, A., Smalling, K., Stehle, S., Schulz, R., & Schäfer, R. B. Fungicides: An Overlooked Pesticide Class?. Environmental Science and Technology 2019, 53, 3347–3365. [CrossRef]
- Lahlali, R., Ezrari, S., Radouane, N., Kenfaoui, J., & Esmaeel, Q. Biological Control of Plant Pathogens: A Global Perspective. Microorganisms 2022, 10, 596. [CrossRef]
- Figueroa-López, A. M., Cordero-Ramírez, J. D., Martínez-Álvarez, J. C., López-Meyer, M., Lizárraga-Sánchez, G. J., Félix-Gastélum, R., Castro-Martínez, C., & Maldonado-Mendoza, I. E. Rhizospheric bacteria of maize with potential for biocontrol of Fusarium verticillioides. SpringerPlus 2016, 5. [CrossRef]
- Lizárraga-Sánchez, G. J., Leyva-Madrigal, K. Y., Sánchez-Peña, P., Quiroz-Figueroa, F. R., & Maldonado-Mendoza, I. E. Bacillus cereus sensu lato strain B25 controls maize stalk and ear rot in Sinaloa, Mexico. Field Crops Research 2015, 176, 11–21. [CrossRef]
- Douriet-Gámez, N. R., Maldonado-Mendoza, I. E., Ibarra-Laclette, E., Blom, J., & Calderón-Vázquez, C. L. Genomic analysis of Bacillus sp. Strain B25, a biocontrol agent of maize pathogen Fusarium verticillioides. Current Microbiology 2018, 75, 247–255. [CrossRef]
- Figueroa-López, A. M., Leyva-Madrigal, K. Y., Cervantes-Gámez, R. G., Beltran-Arredondo, L. I., Douriet-Gámez, N. R., Castro-Martínez, C., & Maldonado-Mendoza, I. E. Induction of Bacillus cereus chitinases as a response to lysates of Fusarium verticillioides. Romanian Biotechnological Letters 2017, 22, 12722–12731.
- Báez-Astorga, P. A., Cazares-Álvarez, J. E., Cruz-Mendívil, A., Quiroz-Figueroa, F. R., Sánchez-Valle, V. I., & Maldonado-Mendoza, I. E. Molecular and biochemical characterization of antagonistic mechanisms of the biocontrol agent Bacillus cereus B25 inhibiting the growth of the phytopathogen Fusarium verticillioides P03 during their direct interaction in vitro. Biocontrol Science and Technology 2022, 1074-1094. [CrossRef]
- Morales-Ruiz, E., Priego-Rivera, R., Figueroa-López, A. M., Cazares-Álvarez, J. E., & Maldonado-Mendoza, I. E. Biochemical characterization of two chitinases from Bacillus cereus sensu lato B25 with antifungal activity against Fusarium verticillioides P03. FEMS Microbiology Letters 2021, 368, fnaa218. [CrossRef]
- Vaghela, B., Vashi, R., Rajput, K., & Joshi, R. Plant chitinases and their role in plant defense: A comprehensive review. Enzyme and Microbial Technology 2022, 159, 110055. [CrossRef]
- Huang, C., Yan, Y., Zhao, H., Ye, Y., & Cao, Y. Arabidopsis CPK5 phosphorylates the chitin receptor LYK5 to regulate plant innate immunity. Frontiers in Plant Science 2020, 11, 702. [CrossRef]
- Naumann, T. A., Wicklow, D. T., & Price, N. P. J. Identification of a chitinase-modifying protein from Fusarium verticillioides: Truncation of a host resistance protein by a fungalysin metalloprotease. Journal of Biological Chemistry 2011, 286, 35358–35366. [CrossRef]
- Gong, A., Jing, Z., Zhang, K., Tan, Q., Wang, G., & Liu, W. Bioinformatic analysis and functional characterization of the CFEM proteins in maize anthracnose fungus Colletotrichum graminicola. Journal of Integrative Agriculture 2020, 19, 541–550. [CrossRef]
- Shang, S., Liu, G., Zhang, S., Liang, X., Zhang, R., & Sun, G. A fungal CFEM-containing effector targets NPR1 regulator NIMIN2 to suppress plant immunity. Plant Biotechnology Journal 2024, 22, 82–97. [CrossRef]
- Feng, L., Dong, M., Huang, Z., Wang, Q., An, B., He, C., Wang, Q., & Luo, H. CgCFEM1 Is Required for the Full Virulence of Colletotrichum gloeosporioides. International Journal of Molecular Sciences 2024, 25, 2937. [CrossRef]
- Cazares-Álvarez, J.E.; Báez-Astorga, P.A.; Arroyo-Becerra, A.; Maldonado-Mendoza, I.E. Genome-Wide Identification of a Maize Chitinase Gene Family and the Induction of Its Expression by Fusarium verticillioides (Sacc.) Nirenberg (1976) Infection. Genes 2024, 15, 1087. [CrossRef]
- Martínez-Álvarez, J. C., Castro-Martínez, C., Sánchez-Peña, P., Gutiérrez-Dorado, R., & Maldonado-Mendoza, I. E. Development of a powder formulation based on Bacillus cereus sensu lato strain B25 spores for biological control of Fusarium verticillioides in maize plants. World Journal of Microbiology and Biotechnology 2016, 32. [CrossRef]
- Warham, E.J.; Butler, L.D.; Sutton, B.C. Ensayos para la semilla de maíz y de trigo: Manual de laboratorio; Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT): México, 1997.
- Untergasser, A., Nijveen, H., Rao, X., Bisseling, T., Geurts, R., & Leunissen, J. A. M. Primer3Plus, an enhanced web interface to Primer3. Nucleic Acids Research 2007, 35, W71-4. [CrossRef]
- Livak, K. J., & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [CrossRef]
- Szklarczyk, D., Franceschini, A., Wyder, S., Forslund, K., Heller, D., Huerta-Cepas, J., Simonovic, M., Roth, A., Santos, A., Tsafou, K. P., Kuhn, M., Bork, P., Jensen, L. J., & von Mering, C. STRING v10: protein-protein interaction networks, integrated over the tree of life. Nucleic acids research 2015, 43, D447–D452. [CrossRef]
- Jashni, M. K., Dols, I. H., Iida, Y., Boeren, S., Beenen, H. G., Mehrabi, R., Collemare, J., & de Wit, P. J. Synergistic Action of a Metalloprotease and a Serine Protease from Fusarium oxysporum f. sp. lycopersici Cleaves Chitin-Binding Tomato Chitinases, Reduces Their Antifungal Activity, and Enhances Fungal Virulence. Molecular plant-microbe interaction 2015, 28, 996–1008. [CrossRef]
- Mwangi, M. W., Muiru, W. M., & Kimenju, J. W. Characterization of Fusarium species infecting tomato in Mwea West Sub-county, Kirinyaga County, Kenya. Canadian Journal of Plant Pathology 2021, 43, 56–61. [CrossRef]
- Gherbawy, Y., Hussein, M., El-Dawy, E., Abdo, N., & Alamri, S. Identification of Fusarium spp. associated with potato tubers in upper Egypt by morphological and molecular characters. Asian Journal of Biochemistry, Genetics and Molecular Biology 2019, 1–14. [CrossRef]
- Ferrigo, D., Mondin, M., & Raiola, A. Pathogenic and genetic characterization of Fusarium verticillioides strains collected from maize and sorghum kernels. Agriculture 2023, 13. [CrossRef]
- Mohammadi, A., Nejad, F. R., & Mofrad, N. N. Fusarium verticillioides from sugarcane, vegetative compatibility groups and pathogenicity. Plant Protection Science 2012, 48, 80–84. [CrossRef]
- Ma, P., Liu, E., Zhang, Z., Li, T., Zhou, Z., Yao, W., Chen, J., Wu, J., Xu, Y., & Zhang, H. Genetic variation in ZmWAX2 confers maize resistance to Fusarium verticillioides. Plant Biotechnology Journal 2023, 21, 1812–1826. [CrossRef]
- Quiroz-Figueroa, F. R., Cruz-Mendívil, A., Ibarra-Laclette, E., García-Pérez, L. M., Gómez-Peraza, R. L., Hanako-Rosas, G., Ruíz-May, E., Santamaría-Miranda, A., Singh, R. K., Campos-Rivero, G., García-Ramírez, E., & Narváez-Zapata, J. A. Cell wall-related genes and lignin accumulation contribute to the root resistance in different maize (Zea mays L.) genotypes to Fusarium verticillioides (Sacc.) Nirenberg infection. Frontiers in plant science 2023, 14, 1195794. [CrossRef]
- Bai, B., Liu, W., Qiu, X., Zhang, J., Zhang, J., & Bai, Y. The root microbiome: Community assembly and its contributions to plant fitness. Journal of Integrative Plant Biology 2022, 64, 230–243. [CrossRef]
- Lu, D., Jin, H., Yang, X., Zhang, D., Yan, Z., Li, X., Zhao, Y., Han, R., & Qin, B. Characterization of rhizosphere and endophytic fungal communities from roots of Stipa purpurea in alpine steppe around Qinghai Lake. Canadian Journal of Microbiology 2016, 62, 643–656. [CrossRef]
- Ling, N., Wang, T., & Kuzyakov, Y. Rhizosphere bacteriome structure and functions. Nature Communications 2022, 13, 836. [CrossRef]
- Hamid, R., Khan, M. A., Ahmad, M., Ahmad, M. M., Abdin, M. Z., Musarrat, J., & Javed, S. Chitinases: An update. Journal of Pharmacy and Bioallied Sciences 2013, 5, 21–29. [CrossRef]
- Fung, K.-L., Zhao, K.-J., He, Z.-M., & Chye, M.-L. Tobacco-expressed Brassica juncea chitinase BjCHI1 shows antifungal activity in vitro. Plant Molecular Biology 2002, 50, 283–294. [CrossRef]
- Jiang, N., Wang, L., Jiang, D., Wang, M., Yu, H., & Yao, W. Combined metabolome and transcriptome analysis reveal the mechanism of eugenol inhibition of Aspergillus carbonarius growth in table grapes (Vitis vinifera L.). Food Research International 2023, 170(1800), 112934. [CrossRef]
- Dang, P. Van, Tran, H. T., Nguyen, D. N., Le, Q. A., Bui, D. D., Nguyen, H. Q., Tran, C. S., & Bui, H. M. Study on the chitinase-induced efficiency against anthracnose on soybean plant by oligochitosan-Zn2+ complexes. Case Studies in Chemical and Environmental Engineering 2023, 7, 100285. [CrossRef]
- Salzer, P., Bonanomi, A., Beyer, K., Vögeli-Lange, R., Aeschbacher, R. A., Lange, J., Wiemken, A., Kim, D., Cook, D. R., & Boller, T. Differential expression of eight chitinase genes in Medicago truncatula roots during mycorrhiza formation, nodulation, and pathogen infection. Molecular Plant-Microbe Interactions 2000, 13, 763–777. [CrossRef]
- Chen, J.-Y., Sang, H., Chilvers, M. I., Wu, C.-H., & Chang, H.-X. Characterization of soybean chitinase genes induced by rhizobacteria involved in the defense against Fusarium oxysporum. Frontiers in Plant Science 2024, 15. [CrossRef]
- Jiao, S., Hazebroek, J. P., Chamberlin, M. A., Perkins, M., Sandhu, A. S., Gupta, R., Simcox, K. D., Yinghong, L., Prall, A., Heetland, L., Meeley, R. B., & Multani, D. S. Chitinase-like1 plays a role in stalk tensile strength in maize. Plant Physiology 2019, 181, 1127–1147. [CrossRef]
- Sindhu, A., Langewisch, T., Olek, A., Multani, D. S., McCann, M. C., Vermerris, W., Carpita, N. C., & Johal, G. Maize Brittle stalk2 encodes a COBRA-like protein expressed in early organ development but required for tissue flexibility at maturity. Plant Physiology 2007, 145, 1444–1459. [CrossRef]
- Agostini, R. B., Postigo, A., Rius, S. P., Rech, G. E., Campos-Bermudez, V. A., & Vargas, W. A. Long-Lasting primed state in maize plants: salicylic acid and steroid signaling pathways as key players in the early activation of immune responses in silks. Molecular Plant-Microbe Interactions, 2019, 32, 95–106. [CrossRef]
- Lambarey, H., Moola, N., Veenstra, A., Murray, S., & Rafudeen, M. Transcriptomic analysis of a susceptible african maize line to Fusarium verticillioides infection. Plants 2020, 9. [CrossRef]
- Meyer, J., Berger, D. K., Christensen, S. A., & Murray, S. L. RNA-Seq analysis of resistant and susceptible sub-tropical maize lines reveals a role for kauralexins in resistance to grey leaf spot disease, caused by Cercospora zeina. BMC Plant Biology 2017, 17, 197. [CrossRef]
- Li, H., Ishfaq, S., Liang, X., Wang, R., Wei, H., & Guo, W. A Novel CFEM Effector in Fusarium verticillioides Required for Virulence Involved in Plant Immunity Suppression and Fungal Cell Wall Integrity. International Journal of Molecular Sciences 2025, 26, 4369. [CrossRef]
Figure 1.
Effect of Bacillus cereus B25 on the disease severity of maize-Fv infection. Bars represent the disease severity index in percentage (%). Different letters indicate significant differences (P<0.05) according to a T-student test. The statistical analyses were performed using the values for each day separately. Data are presented as mean + standard deviation.
Figure 1.
Effect of Bacillus cereus B25 on the disease severity of maize-Fv infection. Bars represent the disease severity index in percentage (%). Different letters indicate significant differences (P<0.05) according to a T-student test. The statistical analyses were performed using the values for each day separately. Data are presented as mean + standard deviation.

Figure 2.
Confocal laser scanning microscopy showing the endophytic location of B25 and Fv inside vascular vessels at 14 dpi. A) Bacillus cereus B25 colonization in root apex. B) Bacillus cereus B25 colonization in the root cortex, C) Bacillus cereus B25 colonization in root vessels. D) Fusarium verticillioides colonization in the root vessels. Arrows indicate the bacteria cells. Letter “c” indicates cortex cells and “v” indicates vascular cells.
Figure 2.
Confocal laser scanning microscopy showing the endophytic location of B25 and Fv inside vascular vessels at 14 dpi. A) Bacillus cereus B25 colonization in root apex. B) Bacillus cereus B25 colonization in the root cortex, C) Bacillus cereus B25 colonization in root vessels. D) Fusarium verticillioides colonization in the root vessels. Arrows indicate the bacteria cells. Letter “c” indicates cortex cells and “v” indicates vascular cells.

Figure 3.
Growth of maize seedlings and detection of endophytic microorganism at 14 dpi. a) maize seedlings from each treatment; (b) close up of maize control seed; (c-e) close up of inoculated maize seed with B25 (c), Fv (d) and both microorganisms (e); (f) growth of microorganisms from superficially disinfected root samples. Green arrows indicate B25 growth, and red arrows indicate Fv growth. LBA: Luria Bertani Agar. PDA: Potato Dextrose Agar.
Figure 3.
Growth of maize seedlings and detection of endophytic microorganism at 14 dpi. a) maize seedlings from each treatment; (b) close up of maize control seed; (c-e) close up of inoculated maize seed with B25 (c), Fv (d) and both microorganisms (e); (f) growth of microorganisms from superficially disinfected root samples. Green arrows indicate B25 growth, and red arrows indicate Fv growth. LBA: Luria Bertani Agar. PDA: Potato Dextrose Agar.

Figure 4.
Protein interaction network between Zm00001eb317090 and other maize proteins by STRING analysis. Blue lines: protein homology; violet lines: experimentally determined; navy blue lines: gene co-occurrence; yellow lines: textmining; black lines: co-expression and aqua blue: from curated databases. Colored nodes represent each protein’s interactors.
Figure 4.
Protein interaction network between Zm00001eb317090 and other maize proteins by STRING analysis. Blue lines: protein homology; violet lines: experimentally determined; navy blue lines: gene co-occurrence; yellow lines: textmining; black lines: co-expression and aqua blue: from curated databases. Colored nodes represent each protein’s interactors.

Figure 5.
Proposed interaction model between maize, Fusarium verticillioides and Bacillus cereus B25 in bipartite and tripartite associations. A) 5 dpi: Zm-B25 interaction: B25 induced the highest fold change value of the CEBiP gene involved in PAMPs detection; as well as the highest induction for bk4 brittle stalk, this possibly helps to reinforce plant cell walls since this protein has been predicted to be functionally associated to cellulose synthases suggesting the formation of new cell wall compounds [22]. B25 also induced the extracellular chitinase chn27 (without Fvcmp recognition site) and a cytoplasmic chitinase (chn29) of unknown function. Zm-Fv interaction: Fv presence induced similar levels of these two chitinases, a slight induction of the CEBiP gene may indicate that the presence of the fungus causes the plant to prepare for PAMPs detection, also the lack of induction of bk4 expression suggests that Fv does not allow plant cell wall reinforcement. Fv gene, CFEM, was induced even no maize chitinases (with chitin-binding domain) are present. Zm-B25-Fv interaction: in the tripartite interaction, B25 rescues the induction of bk4 possibly allowing plant cell wall reinforcement, and of CEBiP preparing the plant to sense PAMPs and detect the presence of Fv. While the extracellular chitinase chit27 remains unchanged, the cytoplasmic chn29 gene expression is highly induced in the plant cell. CFEM gene was induced with a high FC value of 22.34 compared with maize-Fv (FC 2.21) possibly as a first defense mechanism. B) 7 dpi: Zm-B25 interaction: B25 induced bk4 and bk2 gene expression, these two putatively interacting proteins may allow plant cell wall reinforcement through their association with cellulose synthase, CEBiP also remained induced at lower levels than at 5 dpi. The two chitinases detected at 5 dpi chn27 and chn29 remained induced, and the Fvcmp containing site chitinases were also induced (EPR4 and chn21). Zm-Fv interaction: in the presence of Fv chn27 and chn29 were uninduced. CFEM was also induced and no fvcmp and fvsep genes were observed, however the Fvcmp targets EPR4 and chn21 were induced, however, chitA and chitB were neither induced, this could be the reason that fvcmp is not induced due to its affinity for these chitinases, while CEBiP was not induced suggesting that CEBiP recognition system can be suppressed by another Fv mechanism such as CFEM proteins. The plant responds to inducing levels of bk4 in an effort of reinforcing plant cell walls in the presence of Fv. Zm-B25-Fv interaction: Fvcmp containing site chitinases EPR4, chn21 and chitB (class IV) were induced in the tripartite interaction while the levels of fvcmp expression were not observed but fvsep was induced in this interaction, possibly due to the maize chitinase activity. Chn27 and chn29 remained induced with this last one raising at higher levels than in the Zm-B25 interaction. Bk4 and bk2 remained induced as in the Zm-B25 interaction possibly allowing plant cell wall reinforcement mediated by B25. Interestingly, the CEBiP gene is also induced at higher levels than in Zm-B25, possibly allowing lower levels of PAMPs to be detected by the plant cell. If this raise in CEBiP expression allows PAMPs detection is something which can only be suggested and remains to be proved. C) 10 dpi: Zm-B25 interaction: in B25 CEBiP, bk4, bk2 and chn27 remain induced compared to 7 dpi and only chn29 was uninduced. EPR4, cmp containing site chitinase, remained induced, but at this time point chitA and chitB were induced instead of chn21 compared to 7 dpi. Under this scenario B25 allows plant cell wall reinforcement as well as good levels of CEBiP for adequate PAMPs detection. Zm-Fv interaction: the presence of Fv causes the induction of cmp site chitinases EPR4, chn21 and chitA; chn27 and chn29 were also induced which were not induced at 7 dpi. The CEBiP PAMPs receptor remained non-induced possibly due to suppression by the Fv CFEM protein, and bk4 showed a slight induction suggesting an attempt of the plant to reinforce its cell wall. Zm-B25-Fv interaction: CEBiP expression was not induced suggesting that blockage of PAMPs signaling may occur under this scenario. Similar levels of bk4 as in the Zm-B25 interaction were observed suggesting that plant cell wall reinforcement is feasible. fvcmp and fvsep were not induced, however, we observed CFEM gene induction. D) 14 dpi: Zm-B25 interaction: cmp containing site chitinases EPR4, chitB and chn21, as well as the extracellular chitinase chn27 were induced in this interaction while CEBiP expression was not induced, it is possible that under this scenario PAMPs signaling detection could still be possible even when CEBiP was not induced. Bk4 gene expression remained induced suggesting plant cell wall reinforcement was still possible. Zm-Fv interaction: cmp containing site chitinases (EPR4, chitB and chitA) and chn27 were induced and CEBiP expression was low. CFEM was induced more than 10 dpi and no fvcmp and fvsep gene were induced even when maize chitinases are present. Zm-B25-Fv interaction: in the tripartite interaction, the induction of fvcmp and fvsep was not observed while EPR4 and chitB remained induced as well as the extracellular chitinase chn27. However, CFEM gene was upregulated, suggesting that Fv can change its infection mechanisms: producing chitinase modifying proteins to protect its cell wall or producing CFEM proteins to suppress plant defense by reducing CEBiP expression.
Figure 5.
Proposed interaction model between maize, Fusarium verticillioides and Bacillus cereus B25 in bipartite and tripartite associations. A) 5 dpi: Zm-B25 interaction: B25 induced the highest fold change value of the CEBiP gene involved in PAMPs detection; as well as the highest induction for bk4 brittle stalk, this possibly helps to reinforce plant cell walls since this protein has been predicted to be functionally associated to cellulose synthases suggesting the formation of new cell wall compounds [22]. B25 also induced the extracellular chitinase chn27 (without Fvcmp recognition site) and a cytoplasmic chitinase (chn29) of unknown function. Zm-Fv interaction: Fv presence induced similar levels of these two chitinases, a slight induction of the CEBiP gene may indicate that the presence of the fungus causes the plant to prepare for PAMPs detection, also the lack of induction of bk4 expression suggests that Fv does not allow plant cell wall reinforcement. Fv gene, CFEM, was induced even no maize chitinases (with chitin-binding domain) are present. Zm-B25-Fv interaction: in the tripartite interaction, B25 rescues the induction of bk4 possibly allowing plant cell wall reinforcement, and of CEBiP preparing the plant to sense PAMPs and detect the presence of Fv. While the extracellular chitinase chit27 remains unchanged, the cytoplasmic chn29 gene expression is highly induced in the plant cell. CFEM gene was induced with a high FC value of 22.34 compared with maize-Fv (FC 2.21) possibly as a first defense mechanism. B) 7 dpi: Zm-B25 interaction: B25 induced bk4 and bk2 gene expression, these two putatively interacting proteins may allow plant cell wall reinforcement through their association with cellulose synthase, CEBiP also remained induced at lower levels than at 5 dpi. The two chitinases detected at 5 dpi chn27 and chn29 remained induced, and the Fvcmp containing site chitinases were also induced (EPR4 and chn21). Zm-Fv interaction: in the presence of Fv chn27 and chn29 were uninduced. CFEM was also induced and no fvcmp and fvsep genes were observed, however the Fvcmp targets EPR4 and chn21 were induced, however, chitA and chitB were neither induced, this could be the reason that fvcmp is not induced due to its affinity for these chitinases, while CEBiP was not induced suggesting that CEBiP recognition system can be suppressed by another Fv mechanism such as CFEM proteins. The plant responds to inducing levels of bk4 in an effort of reinforcing plant cell walls in the presence of Fv. Zm-B25-Fv interaction: Fvcmp containing site chitinases EPR4, chn21 and chitB (class IV) were induced in the tripartite interaction while the levels of fvcmp expression were not observed but fvsep was induced in this interaction, possibly due to the maize chitinase activity. Chn27 and chn29 remained induced with this last one raising at higher levels than in the Zm-B25 interaction. Bk4 and bk2 remained induced as in the Zm-B25 interaction possibly allowing plant cell wall reinforcement mediated by B25. Interestingly, the CEBiP gene is also induced at higher levels than in Zm-B25, possibly allowing lower levels of PAMPs to be detected by the plant cell. If this raise in CEBiP expression allows PAMPs detection is something which can only be suggested and remains to be proved. C) 10 dpi: Zm-B25 interaction: in B25 CEBiP, bk4, bk2 and chn27 remain induced compared to 7 dpi and only chn29 was uninduced. EPR4, cmp containing site chitinase, remained induced, but at this time point chitA and chitB were induced instead of chn21 compared to 7 dpi. Under this scenario B25 allows plant cell wall reinforcement as well as good levels of CEBiP for adequate PAMPs detection. Zm-Fv interaction: the presence of Fv causes the induction of cmp site chitinases EPR4, chn21 and chitA; chn27 and chn29 were also induced which were not induced at 7 dpi. The CEBiP PAMPs receptor remained non-induced possibly due to suppression by the Fv CFEM protein, and bk4 showed a slight induction suggesting an attempt of the plant to reinforce its cell wall. Zm-B25-Fv interaction: CEBiP expression was not induced suggesting that blockage of PAMPs signaling may occur under this scenario. Similar levels of bk4 as in the Zm-B25 interaction were observed suggesting that plant cell wall reinforcement is feasible. fvcmp and fvsep were not induced, however, we observed CFEM gene induction. D) 14 dpi: Zm-B25 interaction: cmp containing site chitinases EPR4, chitB and chn21, as well as the extracellular chitinase chn27 were induced in this interaction while CEBiP expression was not induced, it is possible that under this scenario PAMPs signaling detection could still be possible even when CEBiP was not induced. Bk4 gene expression remained induced suggesting plant cell wall reinforcement was still possible. Zm-Fv interaction: cmp containing site chitinases (EPR4, chitB and chitA) and chn27 were induced and CEBiP expression was low. CFEM was induced more than 10 dpi and no fvcmp and fvsep gene were induced even when maize chitinases are present. Zm-B25-Fv interaction: in the tripartite interaction, the induction of fvcmp and fvsep was not observed while EPR4 and chitB remained induced as well as the extracellular chitinase chn27. However, CFEM gene was upregulated, suggesting that Fv can change its infection mechanisms: producing chitinase modifying proteins to protect its cell wall or producing CFEM proteins to suppress plant defense by reducing CEBiP expression.

Figure 6.
Proposed interaction model between maize-B25-Fv. A) 5 dpi: B25 is associated with an early modulation of Fv response by inducing CFEM and fvcmp genes, suggesting and initial adjustment of fungal virulence and host-interaction responses, consistent with CFEM previously reported functions associated with suppression of plant immunity and fungal cell wall integrity. B) 7 dpi: the tripartite interaction reaches a peak confrontation stage, where fungal expression shifts toward a proteolytic response, as indicated by a high induction of fvsep and fvcmp genes, while CFEM is not induced. This may be related to the increase of maize chitinase induced genes, suggesting that B25 enhanced pathogen perception by increasing PAMPs recognition. C) 10 dpi: fungal gene effectors are not induced. However, maize still induces chitinases and bk4 with a high FC value; B25 may be producing an antagonistic effect towards Fv possibly through direct antagonistic mechanisms. Even without CFEM gene induction, CEBiP receptor remains uninduced; this may be caused by the fungus suppressing PAMPs detection through a still unknown mechanism. D) 14 dpi: fungal genes remain repressed. Like at 10 dpi, B25 could be directly controlling Fv growth, therefore Fv would be unable to produce its effector genes; at the same time, B25 could also be involved in strengthening maize cell wall.
Figure 6.
Proposed interaction model between maize-B25-Fv. A) 5 dpi: B25 is associated with an early modulation of Fv response by inducing CFEM and fvcmp genes, suggesting and initial adjustment of fungal virulence and host-interaction responses, consistent with CFEM previously reported functions associated with suppression of plant immunity and fungal cell wall integrity. B) 7 dpi: the tripartite interaction reaches a peak confrontation stage, where fungal expression shifts toward a proteolytic response, as indicated by a high induction of fvsep and fvcmp genes, while CFEM is not induced. This may be related to the increase of maize chitinase induced genes, suggesting that B25 enhanced pathogen perception by increasing PAMPs recognition. C) 10 dpi: fungal gene effectors are not induced. However, maize still induces chitinases and bk4 with a high FC value; B25 may be producing an antagonistic effect towards Fv possibly through direct antagonistic mechanisms. Even without CFEM gene induction, CEBiP receptor remains uninduced; this may be caused by the fungus suppressing PAMPs detection through a still unknown mechanism. D) 14 dpi: fungal genes remain repressed. Like at 10 dpi, B25 could be directly controlling Fv growth, therefore Fv would be unable to produce its effector genes; at the same time, B25 could also be involved in strengthening maize cell wall.


Table 1.
Primers used in this study
| Gene ID | Gene name | Forward primer (5’—3’) | Reverse primer (5’—3’) |
| Maize chitinases | |||
| Zm00001eb393070 | bk2 | ACTTGGGTTTTCGTCAGAGG | TGCCAATCTTGTAGGAGACG |
| Zm00001eb317090 | bk4 | TAGTTGCCACTTCGCTTTCC | AAGATCTCGCGGTTGTTGAG |
| Zm00001eb346860 | chn21 | CTACAAGCGCTACTGCGATG | CACACACACGTTTTCACTGC |
| Zm00001eb167340 | chn27 | ACGCGTACATGTTCCAGAAG | AGATCATGAGGCCACCGTAG |
| Zm00001eb168350 | chn29 | AAACAATCAGGGGTCCATCC | AGCTAACGAAGGCGTTGATG |
| Zm00001eb078730 | chitA | TCACCTCACACAACAAGCTG | TACTGGGTTCACAGCGAACTAC |
| Zm00001eb425600 | chitB | CAGTATGGCTATGGCAAAGG | ACAGCGCAGAGGAGTGATAG |
| Zm00001eb246640 | EPR4 | ACAACCTCACCTGCTGAATG | GCAATCGCCATCTATCCATC |
| COs receptor | |||
| Zm00001eb002690 | CEBiP | TAGACTGCACTCCGGTGAAAG | GGTGTTGGTATAACCGCTGTAAG |
| Fvgenes | |||
| FVEG_13630 | Fvcmp | GCACCAGCCTTACCA CTAACC | GCATCACTGTTCCCGTGC |
| FVEG_08679 | Fvsep | GGCAGAATCACTGGTACTCTC | TGAACCCTTCGCATTTACG |
| FVEG_07535 | CFEM | ATGGCCCTTGCTCTGTAAAC | AACAATGCCTGTCACCTCAC |
| Housekeeping genes | |||
| Zm00001eb350890 | cdk | CCGTCATCGCCTCACGAAGAG | AGAGCCTGCCTTACGGAATTG G |
| Fvtub | β-tubulin | ACATCCAGACAGCCCTTTGTG | AGTTTCCGATGAAGGTCGAAGA |
Table 2.
Growth of maize plants (Zm) in bi-and tripartite interactions with B25 and Fv in time course experiment.
Table 2.
Growth of maize plants (Zm) in bi-and tripartite interactions with B25 and Fv in time course experiment.
| Treatment | Length (cm) | Fresh weight (g) | ||||||||||||||
| 5 dpi | 7 dpi | 10 dpi | 14 dpi | 5 dpi | 7 dpi | 10 dpi | 14 dpi | |||||||||
| Shoot | Root | Shoot | Root | Shoot | Root | Shoot | Root | Shoot | Root | Shoot | Root | Shoot | Root | Shoot | Root | |
| Zm | 2.70a | 12.60ab | 6.22a | 17.35a | 13.45a | 19.35bc | 22.36ab | 23.45bc | 0.038a | 0.058ab | 0.192a | 0.305a | 0.353a | 0.517a | 0.535a | 0.703a |
| Zm-B25 | 2.41a | 11.55b | 6.50a | 16.46a | 14.06a | 22.20ab | 21.87ab | 26.40ab | 0.051a | 0.080a | 0.227a | 0.347a | 0.337a | 0.462ab | 0.502ab | 0.728a |
| Zm-Fv | 1.60b | 8.47c | 5.72a | 16.82a | 10.85b | 17.65c | 19.78b | 19.84c | 0.030a | 0.048b | 0.192a | 0.287a | 0.233a | 0.377b | 0.425b | 0.505b |
| Zm-B25-Fv | 2.91a | 14.01a | 6.76a | 18.30a | 13.23a | 23.82a | 22.91a | 28.48a | 0.040a | 0.071ab | 0.188a | 0.286a | 0.290a | 0.478a | 0.573a | 0.685ab |
Different letters indicate significant differences (P<0.05) according to Tukey test. Values indicate the average of eight plants per treatment.
Table 3.
Quantitative real time PCR expression analysis of maize chitinases and the maize oligomer chitin-receptor; fungalysin, subtilisin, and CFEM from Fusarium verticillioides.
Table 3.
Quantitative real time PCR expression analysis of maize chitinases and the maize oligomer chitin-receptor; fungalysin, subtilisin, and CFEM from Fusarium verticillioides.
| Genes | |||||||||||
| Treatment |
Zm167340 chn 27 |
Zm168350 chn 29 |
Zm317090 bk4 |
Zm346860 chn 21 |
Zm078730 chitA |
Zm425600 chitB |
Zm246640 EPR4 |
Zm002690 CEBiP |
FVEG_13630 Fvcmp |
FVEG_08679 Fvsep |
FVEG_07535 CFEM |
| 5 dpi | |||||||||||
| Zm-B25 | 3.24 | 6.01 | 145.68 | 1.14 | 0.77 | 0.21 | 0.16 | 23.21 | - | - | - |
| Zm-Fv | 2.29 | 2.59 | 0.52 | 0.86 | 0.67 | 0.01 | 0.5 | 2.09 | 0.12 | 1.35 | 2.21 |
| Zm-B25-Fv | 1.81 | 17.55 | 41.93 | 0.79 | 1.86 | 0.09 | 0.23 | 6.82 | 0.98 | 0.49 | 22.34 |
| 7 dpi | |||||||||||
| Zm-B25 | 2.36 | 3.76 | 38.65 | 2.51 | 1.28 | 1.71 | 17.19 | 3.51 | - | - | - |
| Zm-Fv | 0.81 | 1.11 | 2.45 | 7.45 | 0.47 | 0.54 | 2.35 | 0.79 | 0.12 | 0.22 | 7.03 |
| Zm-B25-Fv | 2.96 | 10.22 | 4.44 | 7.51 | 0.50 | 5.82 | 15.1 | 7.52 | 0.67 | 2.50 | 1.54 |
| 10 dpi | |||||||||||
| Zm-B25 | 2.39 | 0.37 | 27.73 | 1.05 | 5.81 | 2.2 | 6.28 | 4.81 | - | - | - |
| Zm-Fv | 2.10 | 5.63 | 3.47 | 8.51 | 8.22 | 1.32 | 3.96 | 0.8 | 1.02 | 0.65 | 3.54 |
| Zm-B25-Fv | 1.25 | 1.28 | 25.33 | 2.11 | 3.68 | 0.59 | 1.42 | 1.71 | 0.44 | 0.61 | 3.94 |
| 14 dpi | |||||||||||
| Zm-B25 | 6.58 | 0.21 | 10.04 | 2.46 | 0.32 | 2.09 | 5.52 | 0.11 | - | - | - |
| Zm-Fv | 3.36 | 0.09 | 6.77 | 1.10 | 3.86 | 2.68 | 2.16 | 0.1 | 0.17 | 0.80 | 7.67 |
| Zm-B25-Fv | 3.15 | 12.43 | 120.33 | 0.86 | 0.26 | 2.63 | 5.05 | 0.37 | 0.12 | 0.66 | 3.75 |
The cyclin-dependent kinase gene was used for normalization of maize genes and β-tubulin for Fv genes. Fv genes were analyzed using the 2-∆Ct method. (-) refers to undetectable expressions due to the lack of the presence of Fv.
Table 4.
Quantitative real time PCR expression analyses of brittlestalk2 in all treatments.
| Treatment | 5 dpi | 7 dpi | 10 dpi | 14 dpi |
| Zm-B25 | 1.42 | 6.52 | 2.11 | 0.83 |
| Zm-Fv | 0.53 | 0.85 | 1.47 | 0.35 |
| Zm-B25-Fv | 1.41 | 2.37 | 0.70 | 1.40 |
The cyclin-dependent kinase gene was used for gene normalization. The FC value was obtained from 2-∆∆Ct analysis.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



