Submitted:
07 May 2026
Posted:
08 May 2026
You are already at the latest version
Abstract
A function-based definition of a living individual as a central element and specially distinguished level of the phenomenon of life is proposed. According to this definition a living individual is a causally-closed network of five types of subordinate functions: p-processes (Ps), signals (Ss), feedbacks (Fs), re-writing (R(s)) and decoding (D(s)), directed recursively on the superior function: survival and proliferation (copying of its own identity in a possibly large number of copies possibly similar to the original), being self-dependent and self-sufficient in the realization of this purpose in its environment/ecological niche. This definition separates univocally living phenomena from inanimate processes in the case of the contemporary terrestrial life, origin of life on our planet, spontaneously-originated alien life in the universe, artificial and virtual life. According to this definition, life originated in the moment of formation of the first genetic code, including the “key” to this code coupling autocatalytic cycles of proteins (with random amino-acid sequences and divers catalytic abilities) and RNA (R), supported by and supporting proto-metabolism (P) and embedded in coacervate-like bubbles, into hypercycles of proteins and RNA (D) (“everything first” hypothesis). It clearly distinguishes the living individual from other levels of the organization of biological systems, such as living cells and organs, populations, ecosystems and the whole biosphere. Finally, it clearly decides whether such phenomena of doubtful status as viruses and viroids, plasmids, retrotransposons, chromosomes B, prions, cancer cells, mutualistic symbionts, mitochondria, colonial coelenterates, slime molds and eusocial insect(s) (colonies) are living individuals or not, and to what degree.
Keywords:
life
; living individual
; biological function
; purposefulness
; origin of life
; extraterrestrial life
; artificial life
; virtual life
1. Introduction – Definitions of Life
Several definitions of life have been proposed that frequently overlap to a large extent and concern various physical, chemical and biological aspects of life (see, e.g., Muller 1935, 1966; Gánti 1975; Prigogine 1980; McShea 1991; Schrödinger 1992; Maynard Smith and Szathmáry 1995; Bedau 1998; Koshland 2002; Tsokolov 2009; Trifonov 2011). According to the official definition of NASA, life is: “A self-sustaining chemical system capable of Darwinian evolution.” These definitions mostly concentrate on how living organisms reproduce, inherit genetic information and evolve or quote necessary physical characteristics of life. Living organisms are presented as low-entropy systems significantly displaced from thermodynamic equilibrium, dissipative structures, hierarchically-organized open systems, quantum-level-based systems, systems built of organic compounds, complexes of redox reactions, nonlinear computing systems and/or systems capable of supple adaptation (Prigogine 1980; McShea 1991; Schrödinger 1992; Bedau 1998; Adami 2002; Barrack et al. 2007; Tsokolov 2009; AlKhalili and McFadden 2014).
In order to quote a possible minimum definition, a cybernetic definition of life (a living individual) was proposed by the author (Korzeniewski 2001, 2005). It states: “The living individual is a network of inferior, meaning by connotation negative feedbacks, being a functional representation of various aspects of the world, intentionally (in the operational sense) directed on the realization of the superior positive feedback: copying of itself, that is reproduction”. While this definition still “holds true”, after 25 years a need appeared for its significant re-organization and complementation in order to make it more comprehensible for biologists and more compatible with the huge body of biological knowledge and conceptual paradigms (see discussion below).
Our brains have been shaped by biological evolution in such a way that they perceive and conceive primarily spatial objects, their substance and structure, and only secondarily: processes, dynamic phenomena and changes occurring in time. However, just temporal dynamic aspects are central for the essence of biological systems. Some of the existing definitions of life are focused on substance/structure in space, while other – on dynamic processes involving changes over time. However, in the author’s opinion, they are not sufficiently concentrated on the purposefulness of living beings and the distinction between process and function. Also the inter-relation between various functions, their mutual co-conditioning and directing on the superior function: proliferation of the identity of the whole complex of functions in as many copies as possible is not emphasized strongly enough. Finally, the importance of the absolutely central constituent of the phenomenon of life, that is the living individual, is not sufficiently exposed. An analysis of the vocabulary of over 100 different definitions of life allowed to extract the minimal common definition: “Life is self-reproduction with variations” (Trifonov 2011). There is nothing about functions, purposefulness, inter-relations, co-determination and causal closure. Therefore, the author has decided to propose a new, causally-closed functional-network-based (or function-based in short) definition of life/a living individual.
2. Function-Based Definition of A Living Individual
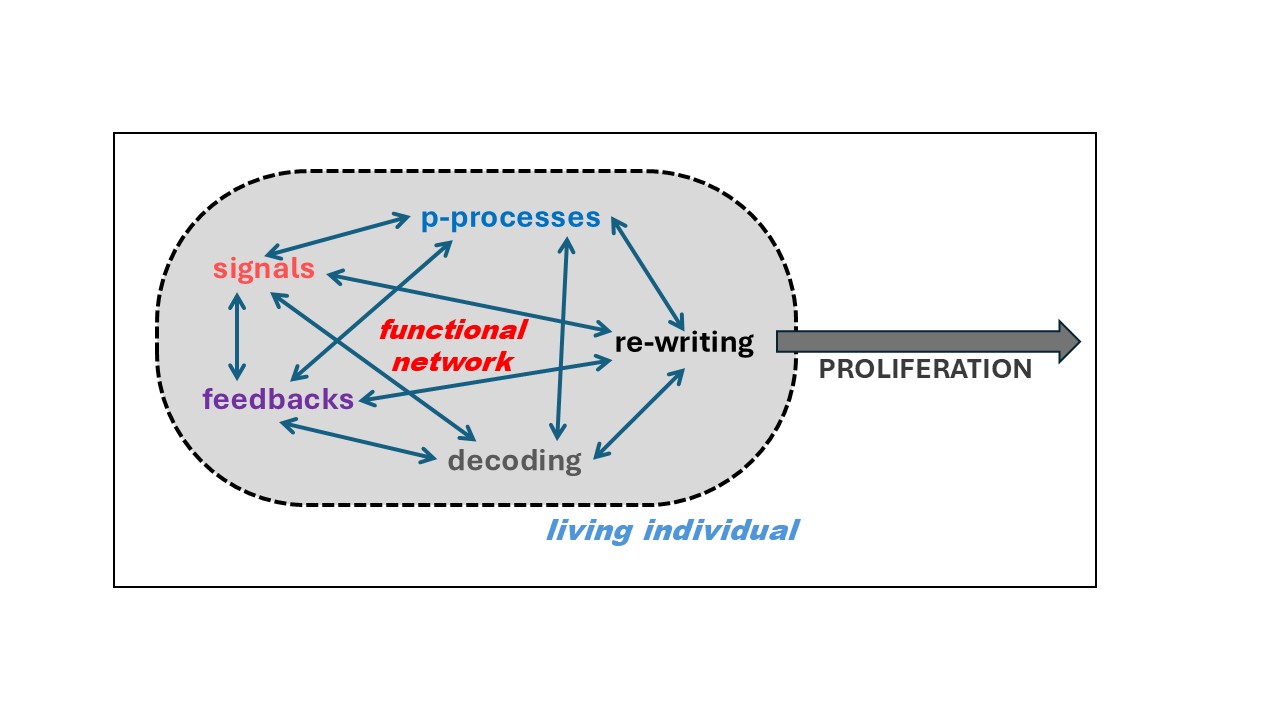
According to the functional-network-based definition of life (living individual), the living individual is a causally-closed network of five distinguished types of subordinate functions: p-processes (Ps), signals (Ss), feedbacks (Fs), re-writing (R(s)) and decoding (D(s)), directed recursively on the realization of the superior function: survival and self-copying (proliferation of its identity in a possibly large number of copies possibly similar to the original), being self-dependent and self-sufficient in the realization of this purpose in its environment/ecological niche.
The function types in question are:
purposeful passive processes: p-processes;
information transduction pathways: signals;
regulatory homeostatic loops: (negative) feedbacks;.
mechanical (physicochemical-fitting-based) information copying: re-writing processes;
conventional (arbitrary-code-based) information transformation: decoding processes;
First of all, purposeful biological functions should be clearly distinguished from purposeless purely physicochemical processes. While physicochemical “indifferent” processes just occur according to laws of physics and chemistry, biological “intentional” (in the operational sense) functions shaped in the course of biological evolution serve some purpose determined by the context of other functions and ultimately by the superior goal of the entire (functional network of the) living individual: survival and reproduction. It may seem an antimony, a sort of vicious circle, that the living individual is defined by its functions, while functions are defined in the context of the living individual. However, there is no escape from this apparent paradox: from the purely physical point of view living systems are nothing but mere physical phenomena, like stones falling down in the gravitational field or crystals growing in a saturating solution of salt. Therefore, while biological organisms are characterized by univocal internal purposefulness and intentionality, from the external point of view they have no goal or target – they simply occur. The essence of biological phenomena cannot by expressed in purely physical terms using purely physical concepts and paradigms.
All biological functions purposefully evolved to carry out their tasks. These tasks only can be defined, determined, “mean” in the context of other tasks and ultimately the whole functional network. All functions serve, co-condition the realization of, directly or indirectly, all other functions and all other functions serve, directly or indirectly, this particular function (the property of causal closure). The subordinate tasks are directed on the realization of the superior purpose. No purely physicochemical system exhibits this kind of purposefulness, reciprocal co-conditioning, causal closure and superior goal. Of course, all, even most complex, “intentional” biological functions are also “indifferent” physicochemical processes, but the inverse relation does not hold.
To be sure, several purely physicochemical processes, purposeless in themselves, occur within living individuals. They can be neutral, useful or harmful for the individual, but nevertheless they “must be taken into account” by its network of functions. For example, diffusion is an elementary physical process, but the entire metabolism could not work without the diffusion of metabolites and enzymes. The enzymatic catalysis is only possible thanks to the electrical (quantum) properties of the enzyme reaction center. The thermodynamic properties of nutrient oxidation or light quanta absorption that enable to gain energy are exploited by living systems. Organisms must cope with gravitation during locomotion, growth, water, nutrients and oxygen transport. Mutations and free radicals (e.g., reactive oxygen species) are generally harmful and must be counter-reacted. However, in the long run, mutations are a necessary factor of biological evolution. Nevertheless, none of this processes evolved during biological evolution as a purposeful function, realizing a determined task within the whole living individual – they equally well occur in the inanimate world, where they fulfil no function.
Of course, in the real physical world any function must be based on structure; material base, spatially-organized carrier. However, within the formal abstract function-based definition of a living individual structure is secondary in relation to function and does not have to be explicitly referred to.
The five purposeful function types enumerated above, entering into the composition of the function-based definition of the living individual, can be characterized as follows:
► P-processes (P) – passive purposeful processes characterized by their qualitative aspects and specificity; they simply do something for a strictly determined purpose; Pn means that the nth process occurs/proceeds; p-processes occur on different levels of hierarchy and are frequently composed of many sub-processes; examples: biochemical reaction – conversion of substance A to substance B catalyzed by an enzyme; energy supply in the cell in the form of ATP; the whole metabolic pathway composed of several enzymatic reactions directed on production of a given chemical compound; pumping of blood by the heart; carrying oxygen by hemoglobin in blood; skeletal muscle contraction; locomotion, manual operations and speech generation realized through a coordinated contractions/relaxations of appropriate muscle groups; multi-stage digesting of different organic compounds in food; complex hierarchically-organized process of morphogenesis during embryonic/individual development based on the morphogenetic cascade;
► Signals (S) – processes determining (activating or inhibiting) the intensity (quantitative commanding aspect) of other functions: p-processes, other signals, re-writing and decoding; they determine whether or not and how fast they proceed; Sa causes an increase in Pn (in the case of p-processes) intensity with a force fna = ΔPn/ΔSa (f > 0 for activating signals, f < 0 for inhibitory signals and f > 1 for signal enhancement); Ss can act in sequential signaling cascades, forming Ss on a higher level of hierarchy; signals are also p-processes (like p-processes are also physicochemical processes); examples: NO-induced blood vessels widening; hormone action (e.g., acceleration of the blood pumping by the heart by adrenaline); hormone-induced signaling pathways within the cell regulating its metabolism (see below); protein phosphorylation/dephosphorylation increasing/decreasing enzyme activity and thus enzymatic reaction rate; neural signal transduction; neural stimulation of muscle contraction; release of Ca2+ within muscle cells from sarcoplasmic reticulum (SR) cisterns to cytosol causing myosin head movement along actin filaments (muscle cell contraction); blood coagulation cascade leading to blood coagulation; activation or repression of gene expression: regulation of DNA transcription to mRNA and mRNA translation to proteins; pheromone action affecting behavior of other individuals; gene switching on and off during cell differentiation (a reach network/cascade of signals is involved); control of embryonic and ontogenetic development based on morphogenetic centers, morphogen gradients, positional information and whole hierarchically-organized morphogenetic cascades (a reach network of signals is involved) (Wolpert 1981; Maynard Smith and Szathmáry 1995; Gilbert 2010); complex system of sensory, motor and cognitive processes conducted by the brain in order to control behavior (Banich and Compton 2018);
► Feedbacks (negative) (F) – homeostatic processes (signals) aiming at maintaining of an optimal (under given conditions) intensity (quantitative stabilizing aspect) of other functions: p-processes, signals, other feedbacks, re-writing and decoding; they steer them to proceed with an optimal intensity; Fx causes an increase or decrease in Pn (in the case of p-processes) intensity with a strength snx = ΔPn/ΔFx; feedbacks are also signals and p-processes (like signals are also p-processes and p-processes are also physicochemical processes); examples: acceleration/deceleration of amino-acid X synthesis through a decreased/increased X level (Umbarger 1978); acceleration/deceleration of ATP synthesis by oxidative phosphorylation through an increased/decreased level of ADP and Pi (products of ATP hydrolysis) (Chance and Williams 1956); regulation of lactose synthesis by the lactose operon (Jacob and Monod 1961); regulation of the glucose level in blood by glucagon and insulin (see below for details) (Cryer 1991); coordination of muscle contraction: an increase/decrease in the strength and/or velocity of particular muscle contractions through a change in the temporal pattern of neural signals from motor cortex activating muscles in response to feedback signals received from muscles by the cerebellum and motor cortex; complex control of sensory, cognitive and motor functions responsible for the organism behavior; supervising of cell differentiation (involving switching on and off gene expression); maintaining of the correct course of embryonic and ontogenetic development based on morphogenetic centers, morphogen gradients, positional information and morphogenetic cascades (Wolpert 1981; Gilbert 2010);
► Re-writing (R) – processes of sequential (at least potentially bi-directional) information copying based on necessary physicochemical correspondence (fitting) between the matrix and the copy; synthesis of a descendant linear hetero-polymer strand with a sequence of monomers (information record) corresponding to (e.g., complementary with) such a sequence of monomers (information record) in the matrix linear hetero-polymer strand; the matrix and the copy have identical or similar physical nature; the correspondence between monomers (information units) in the new and old strand is based on a purely mechanistic physicochemical fitting: the shape and distribution of electrical charges in monomers laying opposite to one another; theoretically, different patterns of the monomer correspondence can be imagined in the case of natural, artificial and virtual life (see below and Korzeniewski 2023); in the present terrestrial life the correspondence is of course C-G and A-U or A-T (pairs of nucleotides/nitrogenous bases in RNA and DNA) and the processes of re-writing involve DNA replication, RNA replication, transcription of DNA to RNA and reverse transcription of RNA to DNA; re-writing processes are also p-processes (like p-processes are also physicochemical processes); in the case of artificial or virtual life one can speak about information elements/units (e.g., 0 and 1) and information records (binary information) instead of monomers and their sequence in hetero-polymers (see below);
► Decoding (D) – processes of sequential one-directional information transformation based on arbitrary accidental (conventional, established “by a chance”) correspondence; synthesis of a linear derivative hetero-polymer chain with a sequence of monomers corresponding to the sequence of monomers in the matrix linear hetero-polymer strand with a different chemical nature and likely the number of monomers than the synthesized chain; the correspondence between the monomers is not physicochemical (based on the shape, electrical charge distribution or other) and is based on an arbitrary, accidental, resulting from the system history of the system correspondence (the code) between the sequence of monomers in the matrix strand and derived chain; this requires a key to this code establishing the correspondence between the monomer sequences; theoretically, various forms of such a code can be imagined (see below and Korzeniewski 2023); in the present terrestrial life, the translation of information from RNA to proteins represents the decoding process and the code is based on four nucleotides (nitrogenous bases): C, G, A and U/T in RNA/DNA and twenty amino-acids in proteins and is triadic (in fact: dyadic-triadic): the sequence (triplet, codon) of three nucleotides corresponds to one amino acid; there are two keys to this code: tRNA and aminoacyl-synthase (in fact, families of different types of these compounds corresponding to different codons and amino-acids); at the beginning of the life history on the Earth this code could be simpler: dyadic (based on two coding nucleotides) or monadic (involving only one coding nucleotide, see below), involve a less number of amino-acids and contain only one key: the ancestor of the contemporary tRNA (see below); after a poly-peptide (a linear chain of amino-acids) synthesis, its folding into a functional protein takes place – a spontaneous (with some help of other proteins) transformation of one-dimensional information record (sequence) to three-dimensional structure and function (followed by possible post-transcriptional modification, attaching of polysaccharides or lipids, cofactor binding and/or subunit assembling into protein complexes).
As the plural forms “re-writings” and “decodings” would look rather odd, the formulations “re-writing processes (Rs)” and “decoding processes (Ds)” will be frequently used.
3. Examples of Functional Networks
A very simple example of a partly causally-closed functional network (truly causally-closed is only the whole functional network constituting the identity of the function-based individual) within a single skeletal muscle cell (myocyte) (a part of it occurs outside the cell) comprising only a few p-processes (Ps), signals (Ss) and feedbacks (Fs), with only a symbolic presentation of the involvement of re-writing R(s) and decoding D(s) is presented in Figure 1. Each of the three shown Ps supports and enables the realization of the next P in a (causally) closed sequence, and generally each of these Ps conditions, directly or indirectly, each other. The ATP supply process delivers energy in the form of ATP for muscle contraction (caused on the molecular level by movement of myosin filaments along actin filaments). Coordinated contraction of appropriate skeletal muscles groups enables searching for and acquiring food (e.g., browsing, hunting, chewing). Most of food supply occurs outside the myocyte, on the hierarchical level of physiology and behavior of the whole organism, but this process involves also substrate transport within the cell through the cellular and mitochondrial membranes. The food contains organic compounds that, after digestion (another function involved, not shown explicitly in Figure 1) supplies substrates for ATP synthesis and the loop closes. The neural stimulation of skeletal muscle coming from the brain (a function occurring mostly outside the myocyte) (S) causes Ca2+ ions release from sarcoplasmic reticulum (SR) cisterns to cytosol (S) and initiates some still unknown factor/mechanism X (perhaps related to protein phosphorylation/dephosphorylation). Ca2+ ions stimulate (S) muscle contraction (mutual myosin and actin movement) (P) and co-operate with X in a direct activation (S) of various elements of the ATP-producing system (the each-step activation, ESA, mechanism: Korzeniewski 1998, 2017). This is responsible for a rough adjustment (S) of the rate of ATP synthesis (P) to the current (highly increased during muscle contraction) rate of ATP hydrolysis (P). A “fine-tunning” of the rate of ATP supply takes place by a negative feedback (F) through an increase of ADP and Pi (inorganic phosphate) concentrations that are products of ATP hydrolysis. The three explicitly demonstrated Ps deliver, directly or indirectly, matter (organic compounds) and energy (ATP) to transcription (R) and translation (D) processes, which in turn produce (on the matrix of appropriate genes) proteins underlying and carrying out all the enumerated Ps, Ss and Fs (in the last case, appropriate enzymes/carriers increase their reaction rates in the result of binding of ADP and Pi). In turn, all these functions participate in providing the whole genetic apparatus carrying out transcription (R) and translation (D) with matter (organic compounds, for example nucleotides and amino-acids) and energy (ATP). “In return”, these processes produce proteins that underlie all these functions (PSFs), and also recursively themselves (RDs).
Of course, this scheme is extremely simplified. All the functions presented are composed of numerous sub-functions, all condition and are conditioned by many other functions of different types (PSFRD) on different levels of hierarchy. For instance, ATP is synthesized by several processes in the cell, including oxidative phosphorylation in mitochondria and anaerobic glycolysis and creatine-kinase-catalyzed reaction in the cytosol. The muscle contraction is a complex process on the molecular level and is subject of multiply control. Food supply occurs on different hierarchy levels: behavioral, including locomotion and search for food, browsing or hunting, chewing; physiological, involving differentiated digestion of complex organic molecules to simple molecules and absorption of the latter in subsequent segments of the digestive tract, their transport to particular organs and cells of the individual through the circulatory system (carried out by blood flow driven by heart beating) and uptake of nutrients by the cell. The signal transduction by neurons and neural networks is itself a complex process. In order to acquire food on the behavioral level of function organization hierarchy, the organism must appropriately coordinate particular muscles contractions on the physiological level, in which the motor parts of the brain are involved, especially pre-motor and motor cortex and cerebellum (Banich and Compton 2018). This complex task involves both neural Ss and Fs, where proprioreceptors (receptors of “internal sensing”) and afferent neural connections coming from them check the current speed and tension of particular muscles, the brain (cerebellum) compares them with the “assigned” speed and tension, and in the case when any deviation of the current speed/tension from the “target” speed/tension is detected, correcting neural signals are sent through efferent neural connections to particular muscles, modifying their speed/tension. On the behavioral level, the sensory and cognitive parts of the brain supervise search for food and acquiring it (the entire action is initiated by the hypothalamus (S), which is responsible for feeling hunger through sensing the glucose level in blood and mechanical press in the stomach walls (F)). Of course, all these networks and sub-networks of functions need energy (ATP), matter (organic substances) and proteins (enzymes, structural proteins and molecular machines like actomyosin) provided within appropriate (neural, muscle) cells, which involves many other processes necessary for their functioning. The functional network of the skeletal muscle cell is only a small sub-network, which is a part of, and is interconnected through numerous functions with, the entire rest of the whole complex functional network of the organism. And the set of all the elements and interactions enumerated above still constitutes only a dramatic simplification of the complexity of the network of functions entering into the composition of the function-based living individual. All these functions condition each other and form a causally-closed functional network. Of course, the ultimate goal of this network, constituting the functional identity of the living individual, is the sustaining of its own existence, survival and proliferation.
It should be mentioned that in many cases “the same” functions can be considered as belonging at the same time to different classes. For instance, neural signal transduction by single neurons or groups of neurons frequently constitutes at the same time a p-process (P), control signal (S) and homeostatic negative feedback (F).
Hundreds and thousands of similar examples could be quoted. However, the general idea of the causally-closed functional network of the function-based living individual should be clear now. Therefore, an abstract scheme of the function-based living individual is presented in Figure 2. Here, a causally-closed network of inter-connected abstract Ps, Ss, Fs, R(s) and D(s) is ultimately directed on its own maintenance and proliferation, multiplication of its own identity in as many copies as possible.
4. Characteristics of the Function-Based Individual
The formal functional-network-based definition of the living individual can be further disambiguated, pinpointed and specified by quoting additional characteristics of the function-based living individual. They involve:
The functional identity of the living individual is composed of such-and-not-another unique network of functions and is univocally determined by this network;
Each function within the living individual is causally linked, directly or indirectly, to other functions and conditions their existence, determines operational meaning and supports action; reciprocally, all other functions are causally linked to a given function, condition its existence, co-determine its operational meaning and support action (the causal-closure property);
All these and only these subordinate functions that are ultimately directed, through co-operation with other functions, on the realization of the superior purpose enter into the composition of the functional identity of the living individual;
All subordinate (inferior) functions cooperate with each other for the good of the whole individual (its superior goal) and do not compete or fall into antagonisms;
The (degree and kind of the) complexity of the network of subordinate functions reflects the (degree and kind of the) complexity of various aspects of the external (and internal) environment (ecological niche) (physicochemical, biological and social) it must cope with, its variability in space and time and various disturbances coming from it; the relation is reciprocal: the complexity of the features of the surroundings important for functioning and proliferation imposes the complexity to the functional network, while the already-existing complexity of the functional network determines recursively to a significant extent the complexity of the ecological niche (the set of aspects of the external and internal environment that are crucial for the individual’s fitness);
The living individual comprises all five distinguished types of functions enumerated above and further discussed below;
The (identity of the) living individual comprises its whole life cycle, including the course of embryonic and individual development, and not only the “adult” stage;
The living individual is a central element of the essence of life and a very distinguished level in the hierarchy of the organization of living systems (see below);
The living individual constitutes the unit of natural selection and subject to Darwinian evolution (Darwin 1859).
5. Functional Vs. Cybernetic Definition of Life
The functional-network-based definition of a living individual is to some extent complementary to the cybernetic definition proposed previously (Korzeniewski 2001). It is both its continuation and extension. The advantage of the cybernetic definition is its parsimony and formality (it is expressed in terms of formal sciences). Its shortest formulation says that the cybernetic living individual is a homeostatic replicator. The advantage of the function-based definition is its intuitive accessibility and completeness. It is not as concise as the cybernetic definition, but intuitively it is easier to conceive as less abstract and closer to the real life. The cybernetic definition is contained within the functional definition, as the negative feedback is one of the five function types contained in the functional definition. The former is a possibly minimal definition that reaches to the very heart of the essence of life (living individual). However, it may be easier to conceive and conceptually encompass for mathematicians and physicists rather than biologists. On the other hand, the latter is decidedly closer to the real biological life we encounter presently on the Earth. Generally, the cybernetic definition may be more appropriate for the analytical style of thinking, while the functional definition is closer to the synthetic style of thinking. Therefore, both definitions rather supplement than exclude each other.
6. Confrontation of the Functional Definition of Life with the Real Physical World
The causally-closed functional-network based definition of life (a living individual) is still rather abstract, as it is intended to reach to the core of the essence of life and refer not only to the contemporary natural terrestrial life, but also to the origin of life on the Earth, life existing on other planets/moons in the universe, artificial and virtual life. However, many properties of the life we know on the Earth, and probably of any natural spontaneously-originated life have been imposed by the nature and properties of the physicochemical environment it exists in. In the real physical world a function-based living individual is characterized, among others, by the following properties:
all its functions must of course be based on some material/structural carrier;
it must be separated by some structural/functional barrier from its environment, including other individuals, in order to preserve its identity, autonomy and, nomen omen, individuality;
it must be a process significantly displaced from thermodynamic equilibrium, in particular a dissipative structure or, alternatively, a complex set of dissipative structures, driven by matter flow, energy dissipation and entropy increase;
it must be built of sufficiently complex and diverse chemical molecules, in particular to a large extent reduced organic compounds of carbon;
it must be exposed to a significant redox gradient, that is the presence of both donors and acceptors of electrons in the immediate neighborhood;
it must have a one-dimensional record of its identity that is transferred from parents to progeny and is somehow transformed in every generation into a three-dimensional structure and function, as only a linear record of information can be effectively copied in the three-dimensional world;
its record of identity must undergo accidental changes (mutations), and as a result its identity also must undergo modifications, because of the physical nature of the carrier of this record fulfilling elementary thermodynamic laws;
its record of identity must be copied with the same or greater effective rate (involving loss of information during accidental errors in copying) than the rate with which it decays (is destroyed) in order for the identity to endure and possibly evolve;
its record of identity, and thus the identity itself, must evolve from generation to generation as a consequence of its copying, modifications and the limited capacity of the environment (ecological niche) (according to the “logical proof of evolution”, Korzeniewski 2023);
it must self-assemble itself in its environment using its identity record and “make-yourself-by-yourself” tool kit inherited from its parents from some simple germ (e.g., fertilized egg cell), with a possible help of the mother, father or both parents (Korzeniewski 2023);
there exists some minimal evolutionarily-stable complexity of its functional network reflecting (representing) the complexity of the physical reality it must cope with, possibly corresponding to the level of organization of prokaryotes that probably have not changed significantly during over 3 billions of years of biological evolution on the Earth;
7. Degree of Individuality
A free-living function-based individual is fully autonomous, self-dependent and self-sufficient in carrying out its superior task – self-maintenance and reproduction in its environment/ecological niche. However, there are cases like parasites, mutualistic symbionts and colonial organisms that do not entirely fulfil this condition – they depend on and are involved in the networks of functions of other individuals. Are they individuals and, if so, to what extent? The degree of individuality can be graded – it is not just a digital, zero-or-one distinction, a Hamletic problem “to be or not to be” an individual. It is possible to be an individual only partly, to some extent. The degree of individuality can be very roughly and generally defined as follows:
Of course, a major objection that can be addressed to the formulated degree of individuality is that it would be very difficult in practice to determine the actual number of functions in a considered system, as there are frequently no sharp limits between functions and many functions are nested within other functions, constitute their sub-functions (the functional network has a strongly-expresses hierarchical character). However, this measure of individuality is not intended to give a strict particular value of IA, but rather to offer a general conceptual frame of reference for the discussion of the “evolutionary selfishness” of the “candidates” for function-based living individuals. Anyway, the degree of individuality should not be confused with the degree of autonomy (see below).
8. Which Phenomena Are Function-Based Living Individuals?
The functional-network-based definition of a living individual encompasses the cybernetic definition proposed previously (Korzeniewski 2001, 2005, 2023). Therefore, phenomena that were counted as cybernetic individuals also belong to function-based individuals. However, the justification of their classification is frequently significantly different and the functional definition delivers significantly stronger conceptual tools to deal with this problem than the cybernetic definition.
First, all processes traditionally regarded as inanimate phenomena, such as stones falling in the gravitational field, crystals growing in a saturated solution of salt or “flowers” “painted” by frost on window glasses certainly do not constitute (elements of) biological systems in general, and function-based individuals in particular. They are not purposeful, shaped by biological evolution, directed on the realization of the superior goal of self-sustaining and proliferation. Some processes, like convective currents belonging to dissipative structures, are dynamic processes organized in space and time, sustained by a ceaseless circular flow of matter, dissipation of energy and production of entropy. However, they lack any operational intentionality as they are not directed on any ultimate task. They are not (composed of) Ps, Ss, Fs, R(s) and D(s).
Some devices produced artificially by humans, for example robots equipped with computers, exhibit a set of functions and realize concrete tasks. Several functions occur within and are carried out by them. They comprise passive purposeful processes (e.g., movements), signal transduction (e.g., electrical), negative feedbacks (e.g., responsible for precise control of operations or navigation in the surroundings), re-writing processes (e.g., mechanistic information copying by computers) and decoding (arbitrary transformation of information from one form into another using available hardware and software by computers, for instance during constructing of complex devices on the basis of sets of instructions). However, these functions and tasks of human-produced devices are not “their own”. On the contrary, they are imposed to them by their human creators to whom these devices serve. They do not reproduce by themselves (copy themselves), possess any mechanisms directed on this purpose, and can be only produced in human factories in a certain, frequently large number of copies. Certainly, they are not self-dependent, self-sufficient in and self-directed on the production of themselves. Therefore, they are by no means artificial function-based living individuals.
However, there exist numerous biological phenomena that are “alive” in this sense that they are complex sub-systems of living systems or are composed of living individuals (of the same or different species), but they are not function-based living individuals themselves. There also exist systems that have only a limited autonomy as individuals and therefore are only individuals to a certain degree and not fully-fledged function-based living individuals (their IA < 1). Let us consider some doubtful cases in order.
Viruses are frequently regarded as situated on the border of life and non-life. Their infectious particles outside the host cell, virions, have the form of a nucleic acid molecule (one- or two-stranded, linear or circular RNA or DNA) encapsulated in a protein capsid composed of single protein molecules: capsomers (Harris and Hill 2021). They exhibit no “vital functions”: metabolism, movement, growth or replication. They execute no Ps, Ss, Fs, R(s) and D(s). Are they therefore “dead”? Not at all. They resemble e.g., bacteria spores produced in unfavorable (hostile) conditions for the purpose of survival that are completely dehydrated (devoid of water) and carry out no metabolism and other functions. However, as discussed above, it is not just one stage, but the whole life cycle of a given organism that constitutes the function-based living individual. When a virion enters the host cell, the situation changes dramatically. It takes over the control of the cell functioning, re-directs its metabolism and genetic apparatus (all its Ps, Ss, Fs, R(s) and D(s)) and forces it, instead to perform normal functions and maintain homeostasis, to produce maximal possible amount of copies of the virus particles (virons), which dramatically disturbs the cell functioning by depriving it of building substances and energy, leading eventually to its death (not all viruses are lethal for the infected cell). The virus is very efficient in the realization of this task. These functions of the host cell captured by the virus and imposed by itself involve p-processes (e.g., synthesis of organic compounds being substrates for new virions production), signals and feedbacks (taking over and controlling the cell functioning, but also many of the existing regulatory mechanism of the cell metabolism), re-writing processes (i.e. copying the virus nucleic acid) and decoding processes (e.g., synthesis of proteins encoded in this nucleic acid such as capsomers or reverse transcriptase in retroviruses) (anyway, viruses also use numerous proteins already synthesized by the host cell). Of course, all these functions are executed by the host cell components (especially enzymes) or by a few proteins synthesized by these enzymes on the matrix of the virus nucleic acid. However, the essence of being a parasite is to get rid of as many vital functions of its own as possible and to “cede” these functions to the host organism. Viruses realize this strategy to an extreme extent. While isolate virions outside host cells do not seem “alive”, viruses “dipped” in the functional network of the host cell switched by the virus to its own proliferation certainly behave as fully-fledged living individuals. Additionally, as the virus RNA or DNA undergoes mutations, viruses are subject to normal natural selection and Darwinian evolution. Therefore, viruses are certainly function-based living individuals, although with a very low value of IA, much more close to 0 than 1.
Viroids are even simpler than viruses, as their naked circular single (but containing many intra-molecular nitrogenous base-pairing regions) RNA strand contains only a few hundreds of nucleotides and they are devoid of the protein capsid (Navarro et al. 2021). However, essentially all the reasoning concerning the status of viruses as function-based living individuals refers also to viroids. When they enter into the host angiosperm plant cell, its functioning is re-directed and it executes all the functions necessary for the viroid RNA to replicate: Ps, Ss, Fs, R(s) and D(s) (of course, most of them are functions that normally occur in non-infected cells). To be sure, viroid RNA does not code for any protein. However, this does not mean that decoding is not involved in the viroid proliferation. All the necessary proteins are, before infection or under the viroid “command”, synthesized by the cell’s genetic apparatus. For example, the viroid RNA is replicated by enzyme RNA polymerase II, which in a “normal” (not infected) cell catalyzes transcription of DNA to mRNA. Therefore, viroids are also, like viruses, function-based living individuals, although characterized by an extremely low, even lower than in viruses, value of IA.
Depending on the particular case and conditions, plasmids (Kado 1998; Brown 2012) can be regarded as either mutualistic symbionts or parasites of bacteria and some simple eukaryotes. They are small extrachromosomal circular DNA molecules present in one or several copies in the host cell. The proliferate independently (they are replicons) and code for they own system of replication. They also code for proteins controlling their separation to descendent cells during the host cell division. They can contain the addiction system that kills the host cells that have not inherited a copy of the plasmid during cell division. They can transfer themselves between host cells during the process of conjunction. They are not essential for the host functioning and survival under normal conditions. However, they can be very helpful in special circumstances, as they can condition the resistance to antibiotics, ability to decompose poisons, induce virulence or code for a system being a substitute of sexual reproduction: when transferring themselves (their copies) from a cell to a cell, they can carry with them fragments of the host genomes and thus participate in the horizontal gene transfer. Generally, they are equipped with a rich network of functions, including Ps, Ss, Fs, R(s) and D(s), both their own and that of the host cell. Therefore, they certainly should be counted among function-based living individuals with a relatively high IA value, undoubtedly higher than in viruses and viroids.
Transposons (transposable elements (TE), jumping genes) (Pray 2008; Brown 2012; Futuyma 2013), counted among parasitic DNA (Doolittle and Sapienza 1980; Orgel and Crick 1980), are mobile genetic elements – segments of DNA of eukaryotic chromosomes that can change their position within the genome. There are distinguished two apparently quite similar classes of transposons. Class I transposons (retrotransposons) pass through the RNA stage during their “life cycle”. They encode a few proteins (D), including reverse transcriptase, which catalyzes synthesis of a DNA strand on the matrix of an RNA strand (R). The regular cell DNA-dependent RNA polymerase carries out the transcription of the retrotransposon DNA to RNA (R). Next reverse transcriptase rewrites this RNA to a single DNA strand (R), that is next doubled by DNA-dependent DNA polymerase (R). The newly-synthesized double DNA segment of the retrotransposon is then inbuilt in a new place in the “host” genome. In this way, the number of the transposon copies can be substantially increased. Class II transposons (DNA transposons) do not pass through the RNA stage. They code for enzyme transposase (D)responsible for insertion and excision of DNA transposon to/from the “host” DNA. The whole process of transposition consists in just relocation of the transposon from one place to another. The number of transposons does not increase – they do not proliferate, but just relocate themselves. Both classes of transposons deprive the host cell of building substances and energy and unnecessarily (to our knowledge) occupy space in chromosomal DNA. Of course the whole process of transposition of Class I and II transposons is involved the functional network (Ps, Ss, Fs, R(s) and D(s)) of the “host” cell with a minor contribution of the transposons own mechanisms. However, retrotransposons do proliferate and they are purposefully directed on this task, albeit only within the genome of the “host”, while DNA transposons just travel from one place in the genome to another, although they are evidently purposefully directed on doing so. Therefore, while retrotransposons are function-based living individuals, albeit with a very low IA, lower even than in viruses, then DNA transposons are not. Therefore, the purposeful transposition of the latter remains a puzzle unless it is somehow supported by the “host”. Anyway, the “environment” of the former in which they can spread remains very limited: it is just the genome of their “host” (or “carrier”), and they can “infect” only the haploid genome coming from the other parent (its gamete) in the fertilized egg cell (zygote). The genetic record of retrotransposons can mutate and evolve, as various transposons can compete for the limited “environment capacity” in the genome, which is a hallmark of a living individual.
Supernumerary chromosomes B (Jones 1975; Bougourd and Jones 1997), another type of parasitic DNA, are unfunctional degenerated chromosomes that nevertheless are present in genomes of many eukaryotic species. The individuals devoid of chromosomes B do equally well, if not better, than their “carriers”. It seems that organisms try to get rid of chromosomes B and that the latter have purposeful mechanisms counter-acting this (Nur et al. 1988). They most probably evolved from “normal” functional chromosomes. Of course, chromosomes B are sustained and replicated with the aid of the whole metabolism and genetic apparatus (all Ps, Ss, Fs, R(s) and D(s)) of the cell containing them. However, they cannot proliferate within the cell. In this aspect they resemble Class II (DNA) transposons. In such a case, are they not living individuals at all? The situation is not so simple, as they are replicated by the cell genetic apparatus during cell division, including meiosis. If they are able to get preferably to gametes, especially the egg cell, they can “infect” the fertilized egg cell containing half of the genome coming from another parent that might not to contain B chromosomes of a given type. In such a way, chromosomes B could propagate and thus deserve classifying as function-based living individuals, although only to a very rudimentary extent with a very low IA. Furthermore, if a given type of DNA transposon (Class II) can also be preferably transferred to gametes, it should regain the status of a function-based (barely) living individual. As it seems, the fact of being or not a function-based individual is not such a clear-cut distinction as could be expected.
Prions (Prusiner 1998) are pathogenic misfolded protein molecules that cause neurodegenerative diseases of the brain in mammals (including humans). The normal (correctly folded) molecules of the prion protein (PrP), possessing α-helix structure, are encoded in the animal genome, synthesized in the brain and other organs and fulfil useful, still not fully understood, function in the organism. The misfolding or transition to the pathological form of a PrP molecule, resulting in the β-sheet structure, can be spontaneous (rarely) or induced by an already misfolded PrP molecule. Such molecules can come from consumed brain tissue of another animal (or human in the case of New Guinea cannibals). Each pathogenic molecule can induce misfolding of several normal molecules, which can in turn induce misfolding of subsequent molecules. In the result of this, the number of misfolded molecules increases exponentially and a positive feedback appears, analogous e.g. to a stony avalanche or bacteria proliferation on laboratory medium. Like viroids, prions are single molecules of an organic linear hetero-polymer, this time protein. Like all proteins synthesized by a living cell, its production is involved in a rich network of functions (Ps, Ss, Fs, R(s) and D(s)). They even exhibit a rudimentary form of replication: pathological molecules proliferate at the cost of normal molecules. Therefore, are they function-based living individuals? Of course not. They constitute a case of a spontaneous living system misfunctioning, something resembling to some extent auto-allergy. They are equipped with no functions/mechanisms purposefully supporting (directed on) their reproduction (IA = 0). They cannot evolve in order to increase their “virulence”, as their identity is encoded in the cell DNA. They just constitute a kind on an “evolutionary trap”.
Cancer cells (Papaccio et al. 2017) originate in the result of tumorous transformation caused by somatic mutations in normal cells. They proliferate in the place where the transformation took place and possibly migrate to other organs and there also proliferate forming metastases. Eventually, they die together with the death of the host organism, frequently caused by them. The divisions of normal cells are strictly controlled by the organism. Mutations cause cancer cells to escape this control and increase their efficiency in proliferation and migration. Traditionally, cancer cells are not regarded as living individuals, perhaps partly because they are still almost identical genetically with their “carrier”. However, the tumorous transformation of a normal cell constitutes breaking out from the superior purpose of the whole organism and establishing its own superior purpose, that is its own survival, reproduction and spreading in the “environment” of the host organism. Cancer cells are subject to normal Darwinian selection and evolution. Only those cells survive and proliferate that develop (due to mutations) numerous adaptations involving purposeful functions of their own (comprising Ps, Ss, Fs, R(s) and D(s)) that support and are directed on their survival, replication and spreading within the host organism. These functions include:
insensitivity to the organism’s cell proliferation control;
resistance to the immunological system and other defensive mechanisms of the host organism;
adaptation of metabolism to lowered oxygen levels, for instance anaerobic (glycolytic) ATP production; the neoplastic tumor has limited vascularization and thus blood flow;
stimulation of angiogenesis (formation of blood vessels) in the neoplastic tumor;
resistance to the induction of apoptosis (programed cell death);
maximization of the rate of cancer cells divisions;
maximization of the rate of cancer cells migration and metastases formation;
competition with other cancer cells, with somewhat different genetic identity, for the limited capacity of the “living space” within the host organism.
Therefore, in the “environment” of the host organism cancer cells behave as regular fully-fledged living individuals. From normal parasites, they differ mainly by the genetic near-identity with the host and impossibility to infect other hosts. However, there are known examples of cancers that can be transferred from one host individual to another, such as dog venereal sarcoma that is transferred through sexual contact in dogs (Barski and Corneferd-Jensen 1966) or facial tumor in Tasmanian devils (Woods et al. 2015), where mutated tetraploid lymphocytes spread to other devils by bites during fights for sexual mate or territory. Generally, cancer cells decidedly constitute function-based living individuals with quite a high value of IA.
In some cases two initially completely independent function-based individuals (IA = 1) start to cooperate and support each other through offering to the partner some functions the partner does not possess and using some functions of the partner that the individual in question is not equipped with. Both partners benefit from such cooperation provided that the advantages prevail over the costs. Mutualistic symbionts are individuals of (very) different species that entered into such a relation called mutualism. It can be facultative, when the partners can, but do not have to cooperate (they can survive and reproduce, although less successfully, in separation), or obligatory, when the partners are absolutely necessary for each other in order to survive and/or reproduce. Examples of facultative symbiosis are cleaner fish and their “clients” – bigger fish (e.g., sharks), whose teeth and skin they clean. The cleaners get food and the cleaned – more healthy teeth and skin, but they can manage alone. In this case, IA of both cleaner fish and their clients is decidedly close to 1. Ants of many species protect and in fact “culture” aphids that in return feed them secreting honeydew. However, both ants and aphids can survive without their partners (ants would have to look for another source of food and aphids would be less protected). Therefore, for both of them 0 < IA < 1. In most cases of lichens, conglomerates of the alga and fungus components (fungi supply water and mineral substances from the substrate, while algae – organic products of photosynthesis), the partners cannot live alone under natural conditions. However, in some cases they can, and they can be cultivated separately in laboratory conditions. Therefore, IA of algae and fungi is closer to 0 than in the previous case, or simply reaches 0. There are many examples of obligatory mutualism, partnership “for life and death”. They include mycorrhiza, that is cooperation between a fungus that fulfill the role of root hair and takes up water and minerals from the soil, and a vascular plant that provides the fungus with organic products of photosynthesis. Flowering plants are fertilized by various insects, birds or bats (fulfilling the role of pollinators) and in return offer sweet nectar and pollen as food, using their colorful flowers as a lure. Termites cannot digest by themselves the dead wood they feed on and they culture for this purpose protozoans or bacteria in their guts: termites get easily assimilable organic compounds, while protozoans/bacteria – raw food and place to live. As the close relationship between obligatory mutualistic symbionts tightens, their value of IA approaches 0. Although the partners in symbiosis retain much of its autonomy, they cannot survive and reproduce independently. If there are still some genetic/evolutionary conflicts between the partners, as they still have their own reproductive systems, IA is slightly greater than zero. If not, it reaches zero. The degree of individuality is not the same what autonomy. For example, the living cell or liver in a complex animal organism possess quite a great degree of autonomy, but they have no individuality (see below).
Mitochondria, called the power plants of eukaryotic cells that deliver most of energy in the form of ATP through oxidizing organic compounds (respiratory substrates) using oxygen, are descendants of once free-living α-proteobacteria (IA = 1) (Reece et al. 2011). They entered into endo-symbiosis (living within) with ancestors of eukaryotic cells: Asgard archaea. Apart from other functions, mitochondria deliver ATP to the cytosol of the eukaryotic cell, receiving from it respiratory substrates as well as most of proteins, carbohydrates and lipids they are built of. Most of their genes have been transferred to the nucleus or lost and presently they are significantly simplified in relation to their ancestors. Nevertheless, they have still preserved some of their reproductive autonomy, although their number is strictly controlled by the cell. On the other hand, mitochondria never originate de novo but only through the division of maternal mitochondria. They contain several copies of their own circular DNA, code for some proteins, for example some subunits of the oxidative phosphorylation complexes, for their own tRNA and rRNA, possess small ribosomes much more resembling bacterial ribosomes than that situated just outside them in the cytosol, are encompassed by two lipid-protein membranes. The mitochondrial DNA still can mutate, and therefore particular mitochondria can possess their own identity and potentially care about their own (evolutionary) interests. For example, in plants, there can occur a genetic conflict between mitochondria and the “cytosolic rest” of the plant. Mitochondria are transferred only through female gametes (egg cells) and therefore a mitochondrial gene cms prevents production of male sex organs in the flower: the stamens. However, the plant activates its own nuclear gene r that suppresses the action of gene cms (Futuyma 2005). On the other hand, the mitochondrial DNA in animals, especially mammals, is much smaller than in plants and contains no such “conflicting” genes. It is not clear whether mutated and thus difunctional mitochondrial can be somehow eliminated by host cells – certainly such mutations cause mitochondrial myopathies (MMs). Taking into account all these facts, mitochondria can be treated either as rudimentary individuals (IA very close to 0), descendants of fully-fledged free-living individuals, or as objects that have already completely got rid of their individuality (IA = 0).
Hydrogenosomes (Yarlett and Hackstein 2005) evolved from mitochondria and are organelles that produce ATP in the process of fermentation in certain primitive protists living in strictly anaerobic (devoid of oxygen) conditions. They are encompassed by a double protein-lipid membrane, but devoid of their own DNA and genetic apparatus. Therefore, in this case the fusion of initially two separate individuals into one has ultimately proceeded to the very end. Hydrogenosomal IA = 0.
Chloroplasts (Reece et al. 2011) are to some extent analogous to mitochondria. They are organelles carrying out photosynthesis in plants and are (descendants of) bacterial endosymbionts, this time cyanobacteria that settled in the ancestors of plants. They have a significantly greater DNA encoding more proteins and more complex structure than mitochondria, their own genetic apparatus and lipid-protein membranes. Therefore, they are rudimentary function-based individuals with IA close to 0, but probably slightly greater, than in mitochondria. However, it cannot be excluded that their IA = 0.
Colonial coelenterates, bryozoans and tunicates (Reece 2011) are composed of “quasi-individuals” or polyps/zooids of the same or different types (e.g., feeding, defensive, reproductive, locomotory). Genetically, they are colonies of clones budding from one larva or ancestral zooid developed from the zygote. The polyps/zooids have connected digestive systems – in fact the share one common gastrovascular cavity/gut – and they exchange food. They are to some extent physiologically (functionally) integrated and exhibit cooperation. The whole colony controls the budding and growth of particular (types of) polyps/zooids, depending on conditions. Therefore, in the case of functional (and, of course, structural) differentiation of polyps/zooids (into sexual and fulfilling other functions), the whole colony constitutes a function-based living individual, as only the whole colony can survive and reproduce. However, even in the case of one type of polyps/zooids that can reproduce, they are all genetically identical and can be regarded as a single function-based individual. On the other, if some mutations occur during the colony growth by budding, different zooids start to differ genetically and therefore to possess different identities they tend to proliferate. Therefore, in this case such mutated zooids become to a tiny extent separate function-based individuals. Anyway, this concerns in fact all organisms that do not possess separated generative (germ) cell lines and thus are in fact well-integrated colonies of somatic groups of cells, for instance trees, where “mutated” branches (somatic groups of cells) with “mutated” flowers produce gametes with a modified identity. Summing up, this is the whole colony of coelenterates or bryozoans that constitutes the function-based individual (IA close to 1), while individual zooids have zero or very low level of individuality (IA equal or very close to 0).
Another interesting case is slime molds (mycetozoans) (Maynard Smith and Szathmary 1995), distant relatives of fungi. During most of their life cycle they have the form of unicellular amoeboids (amoeba-like cells) spread over some area and feeding independently. However, in the case of nutrients deficit, the amoeboids approach each other attracted by chemical signals (chemotaxis) and congregate into multi-cellular plasmodium (slug). After some time the plasmodium forms a sporangium (fruiting body). The chamber of the sporangium is situated at the top of a stalk and contains spores. When the wall of the sporangium breaks, the spores are disseminated. In hospitable conditions (plentiful food) they develop into amoeboids that feed and proliferate and the cycle closes. It is uncertain whether particular amoeboids or the whole plasmodium constitute the function-based living individual. This depends on the genetic identicalness of amoeboids in the plasmodium. If the whole local population of amoeboids comes from spores of one plasmodium, the whole population of cloned amoeboids simply constitutes an “individual in dispersion”, which undergoes consolidation into plasmodium to produce and disseminate spores. If, however, a few genetically different amoeboid groups enter into the plasmodium composition, either because they come from spores produced by different parental plasmodia or they originated in the result of sexual reproduction that facultatively occurs in slime molds, an interesting case of an intra-species reproductive symbiosis (mutualism) takes place. However, this automatically creates a possibility of “reproductive parasitism”, where one of the genetically different groups of amoeboids possesses a mutation that would cause all amoeboids from this group to transform themselves selfishly into spores instead of honestly co-creating the stalk and sporangium walls with other groups of amoeboids. Of course, the mutated amoeboids will transfer more efficiently their identity (genes) to the progeny. Therefore, perhaps appropriate mechanisms preventing such a situation and suppressing the selfish strategy develop. Generally, it is likely that the plasmodium constitutes a kind if intra-species mutualistic symbiont composed of partial individuals (genetically identical groups of amoeboids) cooperating for the common good, with IA between 0 and 1, varying significantly depending on the stage of their life cycle, as a given amoeboid can potentially enter into co-operation with different other amoeboids.
Eusocial insects (wasps, bees, ants and termites) (Wilson 1971) and one rodent species (the naked mole) form colonies containing sexual forms and sterile (unable to reproduce or with suppressed reproduction) forms. The former can be one or more queens (fertile females) in hymenopterans (the males die after copulation) or a queen and a king (a fertile male) in termites and hairless moles. The latter may be workers (females in hymenopterans, females and/or males in termites and naked moles), frequently of a few kinds of different sizes and morphology (and, of course, fulfilling different functions) or soldiers (females in ants, females and/or males in termites). Insects of one type form casts. The role of queens and males is in principle limited to reproduction. The sterile casts realize all other tasks: taking care of queens (and kings), eggs and larvae, feeding them, acquiring food, building the nest, controlling its internal conditions, cultivating “fungi gardens”, defending the colony and so on.
Individual insects, both reproducing and sterile, although each equipped with a reach network of functions (almost all Ps, Ss, Fs, R(s) and D(s) present in solitary insects constituting living individuals), are not self-sufficient in the realization of the superior goal of the function-based individual: self-copying (propagation of own identity), requiring self-dependent sustaining of own existence (at least to the reproduction period) and reproduction. Obviously, sterile insects cannot reproduce and sexual insects (and their progeny) cannot sustain alone their functioning and survive without the help of sterile insects. Sexual and sterile forms must cooperate in order to realize the superior goal of a certain greater whole – the entire colony, its survival and reproduction (propagation of its identity). Particular insects in a colony are interconnected by a reach network of functions involving coordinated cooperation, communication and division of labor. These functions include:
► exchange of food between fed workers and hungry sexual forms, larvae, workers and soldiers (Ps, Fs);
► cooperation in building and repair of the nest involving communication and coordinated actions (Ps, Ss, Fs);
► maintaining homeostasis, involving regulation of the temperature and humidity in the nest, removing dead insects, scraps and debris (mostly Fs);
► cultivation of fungi gardens (Ps, Ss, Fs);
► marking odor traces or carrying out waggle dances indicating newly discovered sources of food (Ss);
► releasing chemical alarm substances in situations of danger (Ss);
► coordinated defense of the nest or column of insects outside the nest (Ps, Ss, Fs);
► common expeditions for food, forming floating islands or hanging bridges from coupled bodies of ants (Ps, Ss, Fs);
► regulation of the numerical strength of particular casts by pheromones or appropriate feeding of larvae (Fs).
Therefore, of course this is the whole colony and not particular sexual or sterile insects that constitute the function-based living individual. In most cases (apart from e.g., primitive colonial wasps) the value of IA for both reproductive and sterile insects is zero. It is frequently argued that sterile workers sacrifice themselves for the queen in the frame of the so called “kin altruism”, as they are very closely related to her and between them. However, in most cases this reasoning is wrong. Leaving apart the cases where a queen is fertilized by many males (which dramatically lowers the statistical kinship of particular workers) or many not-related queens co-exist within a single nest/colony and are supported by the same workers, the sterile insects do not choose their fate, but it is determined genetically, by larvae feeding or pheromones – they “have no choice”. While it is likely that the kin altruism played an important role in eusocial insect evolution, in developed insect societies it can be neglected (see Korzeniewski 2023 for discussion). From the formal point of view, eusocial insect colonies resemble multicellular organisms. Both systems are composed of sexual/reproductive and sterile/somatic elements: insects/cells and casts/organs, interconnected by a rich causally-closed network of functions directed on the proliferation of the identity of the whole system. Multicellular organisms evolved from unicellular ones, while eusocial insect colonies from solitary insects. In both cases the elements ceded their independence and autonomy to the entire system. In both cases, the “selfish” proliferation of the non-reproducing component elements of the whole: sterile insects/somatic cells is strongly suppressed by the whole (cancer cells or functional workers trying to lay eggs are rare exceptions from this rule). The main difference between them is that cells in multicellular organisms are integrated both structurally and functionally, while insects in insect colonies – mostly functionally (Korzeniewski 2023). Therefore, eusocial insect colonies even more deserve the name “functional individuals”.
9. Other Levels of Biological Systems
There can be distinguished, to some extent arbitrarily, several levels of the hierarchy of the structural and functional organization of living systems. They involve atoms, inorganic molecules, simple (monomeric) organic molecules, complex (polymeric) organic molecules, assemblies of complex organic molecules of different kinds (multi-unit proteins, ribosomes, chromosomes, cytoskeleton elements, protein-lipid membranes, flagella), organelles (nucleus, mitochondria, chloroplasts, endoplasmic apparatus, Golgi apparatus, lysosomes), whole cells of many varieties, tissues, organs (liver, brain, heart, lungs), organ systems, whole complex organisms, populations, ecosystems and the entire biosphere. On the other hand, the living individual, as discussed above, is decidedly a very distinguished level of organization in the vital hierarchy. As contemporary free-living individuals (neglecting all the discussed above cases of individuals dependent on other individuals) represent different levels of organization, from the prokaryotic cell (losing in the way the level of organelles) to advanced complex organisms like humans, it is worth to analyze which of these levels are “alive” and why they are or, in most cases, are not living function-based individuals.
A system can be regarded as “alive” when it is complex and autonomous enough and possess a sufficiently reach network of interconnected functions (as opposed to purely physicochemical processes). Therefore, atoms, molecules and their complexes should be automatically excluded from the circle of living systems (or organization hierarchy levels). Organelles can be disputable – compare the above discussion concerning mitochondria. Therefore, let us start from the cell – the most fundamental level of structural and functional organization of the contemporary terrestrial life. Of course the cells of unicellular organisms, both prokaryotic and eukaryotic, are living function-based individuals. Therefore, let us consider cells of complex multicellular organisms. They carry out multiply functions, in fact all the five distinguished types of subordinate functions of the function-based living individual. Are they therefore living individuals? Of course not. Their functioning is not directed on their own proliferation (apart from cancer cells, which however escaped the control of the whole organism and have become living individuals, see above), but on the reproductive success of the whole organism (the actual living individual). They sustain their own functioning, but they are not self-dependent and self-sufficient in doing this – they would not survive without the support of other cells (as well as organs, systems of organs and so on). Also reproductive cells that directly participate in reproduction would not be able to do this without other cells, organs and the whole organism (and they transfer to the progeny the functional identity of the whole organism, and not just of themselves). The cells do not “contain” all functions needed for survival and reproduction in natural conditions. Somatic cells can be cultured and even proliferated in special conditions in laboratories. However, their artificial environment, in which they are provided with all resources they need and protected from any dangers and disturbances, is very poor and simple. Nevertheless, in such a poor environment they can be regarded as living individuals (if they are stimulated to proliferate), like cancer cells in the environment of the host cell or virtual individuals in the environment of the specially programmed operational memory of a computer (see below).
The same reasoning as for cells can be applied to organs, organ systems and other complex units within the body. While they can be regarded as “alive”, they are certainly not leaving individuals, as they do not reproduce by themselves, they are not self-dependent and self-sufficient, and they are directed on the functioning, survival and reproduction of the (identity of) the entire organism they are an element of, and not (only) their own.
To be sure, particular cells and organs have a large degree of autonomy, which can be defined as a partially causally-closed sub-network of internal functions. However, they have no individuality. Both a living cell and an organ, e.g., liver, possesses a complex network of internal functions of all the required types (P, S, F, R, D). However, these functions are not ultimately directed on the “selfish” evolutionary interests of the cell or organ, but serve the superior purpose of the whole individual. Therefore, the degree of individuality IA of the liver or liver cell equals exactly 0. Individuality is something significantly different from (although to some extent related to) autonomy.
Populations of individuals of a given species are not at all purposefully directed on the survival and reproduction of themselves. The individuals within a population take care exclusively of the own selfish evolutionary interest, and not of the good of the whole population or species. Mostly they compete with other individuals (possessing other genetic and functional identities) for the limited capacity of the environment. There are no or few (e.g., in animals forming herds) direct functional interactions between them including rivalry for mating or some cooperation. For example, they can warn other individuals in the whole herd against predators, but this concerns also individuals of different species living in mixed herds. This can be regarded as a case of moderate facultative intra- or inter-species mutualism (the benefit of the warning individual is that the attention of predators is focused on the whole escaping herd, and not only on itself). There is nothing like altruistic acting for the good of the species or population – every individual egoistically cares only about the propagation of its own identity (Williams 1997). Populations can spread by occupying new places in a given area or populating other areas. However, these are particular individuals that are purposefully directed on their numerical and spatial spreading. Therefore, populations are composed of function-based individuals, but are not individuals themselves.
Ecosystems (biocenoses) are composed of individuals of different species, between which various types of relations occur: competition, feeding of herbivores on plants, predation, parasitism or cooperation (mutualism). However, again, ecosystems as a whole do not possess any functions that are purposefully directed on their survival and proliferation. They emerge as a result of interactions: competition, exploitation or (rather rarely) purely egoistic co-operation of individuals of the same or different species. Ecosystems are mostly built of antagonistic, and not co-operative relations. Certainly, they are not function-based living individuals.
Finally, the whole biosphere is composed of particular ecosystems that are only loosely connected by some global factors like oxygen or carbon dioxide in the atmosphere. It is in no way directed on the sustaining of its existence, not to mention reproduction. It results from selfish, mostly antagonistic relations between particular living individuals. Some very naïve ideas like the “Gaia hypothesis” (somewhat analogous to the “good of the species” concept), according to which all organisms on the earth collaborate for the good of the entire terrestrial biosphere (Lovelock 1979), contradict the whole contemporary biological-evolutionary knowledge. The biosphere is not a function-based (or any other) living individual.
10. Sexual Reproduction and the Function-Based Individual Identity
In the result of sexual reproduction, each parent transfers to progeny about a half of its functional identity (such-and-not-another network of functions) and genetic identity underlying it. Like the degree of individuality discussed above is not a zero-or-one feature, the (functional) identity is not indivisible and can be graded. Therefore, it should be determined how sexual reproduction relates to the superior goal of the function-based individual, that is as efficient as possible spreading of its own (functional) identity.
There is no consensus concerning the reasons of the existence of sexual reproduction, although a few propositions have been formulated (Futuyma 2013). Apparently, it seems to do only harms and lower the individual (its identity) fitness (efficiency of proliferation). First, sexual reproduction is a waste of resources (energy and building material) for the production of males. The population/species composed exclusively of parthenogenic females could produce twice as much progeny as a population/species in which females constitute only 50% (sex proportion 1:1), as the number of females, for purely energetic reasons, is limiting for the amount of offspring. Second, and more important from the point of view of the undergoing proliferation identity of the function-based individual, in sexual reproduction each of the parents transfers to progeny only 50% of its identity. Parthenogenically reproducing females transfer entire 100%. An additional disadvantage is waste of time and energy for courtship, mating rituals, bright coloration, exhaustive singing, fighting for females, searching for a partner, copulation, and so on. This time and energy could be invested in, for example, additional feeding or protection against predators, which would enable to produce more better-equipped progeny, and thus propagate own identity more efficiently. From this point of view, sexual reproduction seems decidedly harmful for the proliferation of the identity of function-based individuals. Therefore, as a decisive majority of more complex biological organisms reproduce this way, the immediate (here and now) advantages of sexual reproduction must prevail over its disadvantages.
It has been proposed (Korzeniewski 2023) that the sense and rationale of sexual reproduction and its advantage over asexual reproduction is production, because of random mixing and synergistic action of the elements of parental identities, of a great diversity of genetic and functional identities of the progeny, including “good”, “mean” and “bad” descendants. On the other hand, asexually-reproducing organisms produce only mostly “mean” progeny with the identity (almost) the same as of their parents, although twice as numerous as the progeny of “sexual” organisms. As only a small fraction of the descendant individuals survive and reproduce (in given environmental conditions), this strategy decidedly pays, as statistically only the few most successful “good” descendants of “sexual” individuals will pass their functional identity further, while the numerous “mean” and “bad” progeny of both “sexual” and “asexual” organisms will die without achieving the reproductive success (Korzeniewski 2023). Generally, it is better to spread (pass) a half of someone’s own identity in (to) many descendants than not to spread the whole intact identity at all.
Living organisms being a product of sexual reproduction have a much greater variability of the functional identity among progeny than the organisms produced through asexual reproduction. Both haploids and diploids have statistically a half of the genetic record coming from each parent (leaving aside the inheriting of mitochondria only from the mother in oogamy and the fact that X chromosome contains more genes than Y chromosome), and thus a half of their functional identity. This is caused by crossing-over (intra-chromosomal gene shuffling), random chromosome distribution to gametes (extra-chromosomal gene shuffling) and random gametes pairing during fertilization (both). As the genetic identity determines to a large extent the functional identity, the progeny of “sexual” individuals will be functionally much more diverse, than the twice more numerous progeny of “asexual” individuals. The mean fitness will be equal in both cases. However, “sexual” individuals will produce better-, moderately- and worse-fitted descendants, while “asexual” individuals – only moderately-fitted descendants. It does not matter that some of the descendants of “sexual” individuals are “bad” and will inevitably die without leaving progeny, together with the “mean” descendants of both “sexual” and “asexual” individuals. It matters that few “good” individuals will survive and reproduce (Korzeniewski 2023). They will inherit and pass further on a part of genetic and functional identity of both parents. As stated above, the identity is divisible and it is better when a part of it is propagated than when its entirety dies.
11. Origin of Terrestrial Life
The phenomenon of life emerged from inanimate matter in the moment when first systems appeared that fulfilled the functional-network-based definition of the living individual.
Life appeared on our planet at least 3.8 billion years ago, less than one billion years after its formation (Maynard Smith and Szathmáry 1995; Macdougall 1996; Lane 2010, 2015; Chambers and Mitton 2014; Baggott 2015; Deamer 2019, 2020; Knoll 2021). It could originate in shallow warm ponds (Darwin 1859; Haldane 1929; Oparin 1938), ancient hydrothermal vents (Lane 2010, 2015; Chambers and Mitton 2014; Baggott 2015; Deamer 2019, 2020) or small water reservoirs around places with volcanic/geyser activity (Deamer 2019, 2020; Korzeniewski 2023) There are three most popular general scenarios of the origin of life on the Earth: “metabolism-first”, “genes-first” (or “control-first”) and, perhaps most seldom enumerated “cell-first” hypotheses. According to the first view (metabolism-first), somewhere, perhaps in small empty calcareous cells of alkaline hydrothermal vents (white smokers), appropriate conditions (first of all redox and concentration gradients of appropriate ions and chemicals) prevailed favoring spontaneous synthesis of organic compounds through CO2 assimilation with an aid of electron donors, for example H2 molecules. These chemicals and reactions organized themselves into primeval proto-metabolic pathways or cycles, for example the reverse tricarboxylic acid (TCA) cycle, still without the aid of proteins (enzymes). They constituted self-sustaining, but not self-replicating networks of chemical reactions. Then, such a simple pre-metabolic system acquired in some way a primitive genetic apparatus coding for and producing, among others, biocatalysts (enzymes or ribozymes) sustaining, directing and driving metabolism, and envelope composed of amphiphilic molecules surrounding a bubble containing the metabolism and genetic apparatus (Lane 2010, 2015; Baggott 2013; Kee and Monnard 2016; Smith and Morowitz 2016). From the point of view of the functional definition of a living individuals, this scenario can be called “P-first” or “p-processes first”). Currently, the most popular (if not to say fashionable) scenario is the genes-first hypothesis, which starts from the genetic aspect of living individuals and says that at the very beginning short self-replicating RNA molecules fulfilled both the function of information (genetic) record and catalysts (ribozymes). This conception is called the “RNA world” (Maynard Smith and Szathmary 1995; Copley et al. 2007; Ricardo and Szostak 2009; Cech 2012; Baggott 2013; Lane 2015). Ribozymes started to organize and conduct the first proto-metabolism. Then proteins were co-opted to the cooperation as efficient molecular machines and structural elements (enzymes, carriers, cytoskeleton) and the structural (first proto-cellular) aspect of life (living individuals) was acquired, when the RNA-(and later also protein-)based self-replicating system was located into a kind of a bubble surrounded by a semi-permeable membrane composed of some kind of amphiphilic molecules. The exact sequence of the latter events is disputable and their detailed relative chronology can be changed. Finally, DNA was included to the system as a tough, resistant to damages genetic information carrier. Within the conceptual frame of the functional definition of life, this scenario can be called “R-first or “re-writing first”. According to the third scenario (“cell-first” hypothesis) dating back to Oparin’s coacervates (Oparin 1938; Smokers et al. 2022) and Fox’s microspheres (Nakashima 1987), life started from some purely physicochemical bubbles built of organic compounds and capable of self-dependent growth and division (proto-proliferation), perhaps encompassed by a semi-permeable proto-membrane. These proto-cells then acquired both a proto-genetic apparatus and proto-metabolism (Nakashima 1987; Lane 2010; Cavalier-Smith 2001).
However, perhaps it is not necessary to choose between these different possibilities, as life did not have to start from something very simple. In the author’s opinion, the abilities of purely inanimate matter to self-organize is frequently underestimated. In fact, it was proposed (Korzeniewski 2023) that life originated in coacervate-like bubbles containing, because of purely physicochemical reasons (e.g., better solubility in their interior or semi-permeable surface/envelope), auto-catalytic cycles of proteins (Kauffmann 1986) and RNA (R), the latter supported non-intentionally (“by a chance”) by the former, and some related proto-metabolism (P), also supported by polypeptide chains with random amino-acid sequences and diverse, including catalytic, spectrum of properties. Of course, this proto-metabolism supported reciprocally the autocatalytic cycles of proteins (and RNA). These systems constituted relative complex proto-individuals or “ur-individuals” (Korzeniewski 2023). The first living individual appeared in the moment of the of the joining of the protein and RNA autocatalytic cycles into protein-RNA hypercycles1, that is the origin of the first primitive genetic proto-code (Korzeniewski 2023) (D). The sole “key” to this code was proto-tRNA (a family of its different types) able to recognize by its one functional end (the medium RNA loop of the “clover leaf” of the modern tRNA) the codon in proto-mRNA and by its other (second) functional end (the “footstalk” of the “clover leaf” of the modern tRNA) – an appropriate amino-acid by a purely physicochemical spatio-electrical correspondence (Korzeniewski 2023) (today a family of amino-acyl synthases is responsible for this; thus, the code involves two “keys”: tRNA and amino-acyl synthase). The genetic code was not triadic (or in fact: dyadic-triadic) as today, but most probably dyadic (Smith and Horowitz 2016) with two meaningful (coding) nucleotides separated by one non-coding (meaningless) nucleotide (in fact even in the contemporary code in most cases the third nucleotide in the codon triplet is meaningless, which is called the degeneration of the genetic code), or even monadic (Korzeniewski 2023) with one coding nucleotide separated by two non-coding nucleotides. Of course, the number of the coded amino-acids would be smaller: not 20 as today, but maximally 16 in the dyadic code and 4 in the monadic code. Perhaps not single amino-acids, but their groups of similar properties, for example hydrophobic, polar, acidic and alkalic were encoded.
Such first function-based living individual equipped with at least the genetic (R(s) and D(s)), metabolic (Ps) and structural (cellular) aspect could take care of its functioning, survive, reproduce and evolve. The origination of life most probably involved a parallel development of several different, but mutually inter-related, supporting each other and co-conditioning aspects: genetic, structural-individual, energetic-thermodynamic, metabolic, enzymatic and redox aspect (Korzeniewski 2023). Signal transduction (Ss) and negative feedbacks (Fs) joined the emerging living organisms very early, if not from the very beginning (they could e.g., support the efficient functioning of proto-metabolism or the stability of coacervate-like proto-cells). According to the causal-closure paradigm, on which the functional definition of the living individual is based, all aspects of the living individual functioning co-condition all other aspects, and this was the case already during the process of the origination of life (Korzeniewski 2023).
Therefore, the “everything first” scenario or the origin of life is promoted here and supported by the function-based definition of a living individual. According to this hypothesis, first living organisms contained at least P, R and D functions and very quickly acquired (through evolving appropriate proteins) S and F functions (if they were not present from the very beginning). It must be emphasized that the variety of protein molecules with random amino-acids sequences in autocatalytic cycles were able to perform (especially catalyze), although inefficiently, a wide range of different functions, including new protein molecules synthesis, supporting RNA strands replication and channeling metabolism (Korzeniewski 2023). The “RNA world” never existed, as proteins from the very beginning supported RNA replication. No self-replicating RNA molecules, that is synthesizing new complementary strands on the matrix of old strands from single nucleotides present in the solution, have been discovered in the nature or synthesized in laboratories, despite long-lasting strenuous search and trials. This does not change the fact that at the beginning, before advanced proteins encoded by nucleic acids evolved, the executive role of RNA was much greater, than today, to what such relics as ribozymes testify. Until today, rRNA is the key element of the translation process, many introns self-excise themselves from RNA strands, reverse transcriptase and RNA-dependent RNA polymerase (de Farais et al. 2017) still participate in the reproduction of many viruses and small RNA molecules fulfil many control functions (Barrack et al. 2007). At the beginning, as proposed above, proto-tRNA was perhaps the only “key” to the genetic code. Nevertheless, in the author’s opinion life started from the “extended RNA-protein world” containing both RNA and proteins, but also many other components/elements of a living individual fulfilling various functions. Subsequently, it passed into the “extended DNA-RNA-protein world”, in which DNA took over the function of genetic record, but it was still RNA that underwent replication, while DNA was synthesized through reverse transcription. This stage was represented by Last Universal Common Ancestor (LUCA, Weiss et al. 2016) (Korzeniewski 2023). Then, bacteria and archaea invented independently DNA replication. Some Asgard archaea incorporated into their cells α-proteobacteria and next cyanobacteria that, as endosymbionts, evolved into mitochondria and chloroplasts, and developed the cellular nucleus and complex system of intracellular protein-lipid membranes: eukaryotes originated.
Summing up, life has not begun from some very simple “germ” or “object”, such as a single self-replicating RNA molecule (or a set of such molecules), proto-metabolism or growing-and-dividing bubble. It started from a much more complex system created in the result of self-organization of inanimate nature, for example something resembling hypercycles of nucleic acids (RNA) and proteins “embedded” in a sort of “bubbles” or “droplets” surrounded by a semi-permeable membrane and filled with a mixture of organic compounds reacting with other compounds. The very event of the transformation from an inanimate system to a living function-based individual took placed in the moment of the appearance of the first genetic code (and the key to it), coupling autocatalytic cycles of proteins and RNA into hypercycles, and thus appearance of the first hypercycle of proteins and RNA (Korzeniewski 2023). This was equivalent to an origin of the decoding function (D) in a system already containing multiply p-processes (Ps) and re-writing processes (R(s)), and possibly signal (S) and feedback (F) functions. If not, first individuals very quickly evolved these (S and F) types of functions. This scenario constitutes the “everything first” hypothesis of the origin of life.
12. Extraterrestrial Natural Life
We know little about the possible spontaneously-originated life on other planets or moons in the universe. We can speculate that individuals forming such life would have to be based on molecules of organic compounds as the only complex and diverse enough to act as molecular machines or information record carriers, although not necessarily of the exactly the same kind as those underlying the contemporary terrestrial life. We can also hypothesize that they would possess something analogous to cellular structure, metabolism and genetic apparatus. However, they could most probably develop different body plans, genetic codes and ecologies. Some potential possibilities are presented elsewhere (Korzeniewski 2023). However, most probably the whole spectrum of all the available structural and functional solutions is out of our imagination.
The causally-closed networks of functions constituting the identity of extraterrestrial function-based individuals would be most probably significantly different from those of the terrestrial individuals, although the same types of functions (Ps, Ss, Fs, R(s) and D(s)) would be involved. The degree and kind of complexity of such networks would reflect the degree and kind of the complexity of various aspects and variability in space and time of the environments (ecological niches), in which the individuals constituted by these networks would live. Different physicochemical environments, different basic components, material fundaments of structure and function, different other individuals with which they would have to interact with (compete with, feed on, being fed on by, cooperate with) would force the extraterrestrial natural evolution to shape the functional aspects of living individuals in a completely different way. Also chance would play a crucial role here, as in the case of life on the Earth. Therefore, markedly different (types of) P, S and F functions could be expected in them. Depending on different building materials and just chance, alien life could also generate completely different R processes. For example, they could be based on two monomers: X-X and Y-Y2 (symmetrical pairs) or X-Y (asymmetrical pairs); three monomers: e.g., X-X, Y-Y and Z-Z or X-Y and Z-Z; four monomers: e.g., A-B and C-D (as in the contemporary terrestrial life); five monomers and so on (compare Korzeniewski 2023). Therefore, their D functions would also have to be based on different principles. The number of the coding and coded monomers could be different. Even in the case of life on Earth, it is easy to imagine a genetic code that involves more than 20 amino-acids, which is allowed because of the degeneration of the known genetic code. As discussed above, at the very beginning of life on the Earth, the code could contain two coding nucleotides and up to 16 coded amino-acids, or even one encoding nucleotide and 4 coded (groups of) amino-acids (with similar properties). In the last case, the number of coding monomers (nucleotides) is identical with the number of the coded monomers (although one coding nucleotide is separated by two non-coding nucleotides in the proto-mRNA strand). The same could happen in life on other planets, although the number of both types (coding and coded) of monomers could be different, for example (significantly) greater than four. Presently, the genetic code on our planet involves two “keys” to it: tRNA and amino-acyl synthase. Presumably, when life was originating, it contained only one “key”: proto-tRNA (see above; Korzeniewski 2023). It is imaginable that extraterrestrial life “employs” three or more such keys, although this does not seem very practical.
It should be mentioned that, in order to drive all the functions (PSFRD) underlying the identity of the living individual, it must be exposed to significant thermodynamic, especially redox gradient. This is important in the case of possible life on some moons in the Solar System possessing oceans of liquid water under their frozen ice crust, having contact with the rocky core heated up by tidal forces, chiefly Europa, Ganimedes and Kallisto (Jupiter’s moons), Titan and Enceladus (Saturn’s moon) and Triton (Neptune’s moon). It is not enough to possess liquid water and mineral substances delivered from the core. It is also necessary to be exposed to an appropriate redox gradient in order to sustain life. Such a gradient on the Earth’s surface occurs between the more reduced Earth’s mantle and more oxidized hydrosphere and atmosphere, and is maintained by a continuous flow of hydrogen from the mantle, released in tectonic processes (e.g., volcanism or hydrothermal vents), through hydrosphere and atmosphere to the cosmic space, escaping there after water photolysis to oxygen and hydrogen in atmosphere (Smith and Morowitz 2016; Korzeniewski 2023). While the broken ice surface and water outflows of Europa and water geysers of Enceladus can ensure such continuous hydrogen flow, it seems that the water oceans of Ganimedes, Titan and Triton are firmly sealed by the thick ice shell, while in Kallisto its water ocean (if it exists at all) is additionally separated from the rocky core by a layer of water ice.
Summing up, extraterrestrial function-based living individuals would have to be causally-closed networks of PSFRD functions directed on sustaining of their existence and reproduction in their environment. However, their detailed P, S, F, R and D functions would be most probably slightly or dramatically different (in kind) from the set of functions terrestrial individuals are equipped with (composed of). We do not know how different, as we do not conceive and encompass the whole spectrum of potential possibilities offered by the nature.
13. Real Artificial Life
Artificial life is life that has not originated spontaneously but has been constructed by intelligent beings (Adami 1998). It was argued that artificial real life, the functioning of which is based on the supra-molecular level, can be only composed of dispersed individuals, comprised of structurally-separated but functionally-integrated sub-individuals, such as insects in an eusocial insect colony (Korzeniewski 2011, 2023). The original reasoning concerned the cybernetic definition of life/ling individual, but it generally applies also the functional definition, and in fact any reasonable definition.
The natural spontaneously-originated living individuals on the Earth function on the molecular level – they are composed of organic compounds, to our knowledge the only class of chemical molecules that are variable and complex enough to constitute the basis of life/living individuals, the function of which is based on the molecular level, including molecular machines – proteins and molecular information record carriers – nucleic acids. Therefore, most probably also artificial life, created by humas or other intelligent beings, functioning on the molecular level would have to be based on organic compounds, albeit not necessarily of exactly the same types the life on the Earth is based on.
On the other hand, the supra-molecular function-based life/living individuals would be based on such materials as metals, silicon, carbon (graphene, fullerenes), semi-conductors, plastic materials, inorganic polymers, glass and so on. The structure and function of the elements built of these substances (e.g., chips, microprocessors, cables, capacitors, mechanical parts) are based on the supra-molecular level – they do not result from physicochemical properties of single molecules, but are shaped in the technological process and contain millions of atoms and/or molecules. The supra-molecular artificial function-based individuals would have to cope with numerous tasks, analogous to that executed by biological function-based individuals (comprising Ps, Ss, Fs, R(s) and D(s)), including: 1. finding and exploiting various substances (minerals, metal ores, silicon, carbon, sulfur, oil, gas, radioactive materials and so on) needed for building the components of their bodies and producing energy; 2. transforming these raw materials into secondary substances directly necessary for the production of the supra-molecular components of artificial individuals (smelting metals from ores, carrying out various chemical syntheses, producing plastic or glass, enriching uranium with radioactive isotopes and so on); 3. producing the building components/elements of their bodies (appropriately-shaped metal elements, silicon plates, wires, semi-conductors and so on); 4. producing structural and functional elements/parts of higher order (microprocessors, chips, batteries/accumulators, electrical circuits, engines, information storage and processing media); 5. detecting defects/damages within themselves; 6. exchanging/repairing their incorrectly working parts/elements; 7. constructing/producing their descendants with the same (or very similar) functional identity; 8. preserving, copying and implementing the record of their functional identity i.e., a prescription and set of instructions leading to the creation of appropriate structure underlying and executing such-and-not-another complex of functions in descendant individuals; 9. receiving various stimuli (“sensations”) from the surroundings, perceiving varying in space and time variability of their environment; 10. responding appropriately to these stimuli through implementing suitable behavior and executing this behavior; 11. exploiting available sources of energy (thermal gradients, electromagnetic radiation, nuclear processes, chemical reactions, atmospheric electric discharges) for the construction of their bodies and functioning; 12. maintaining “internal” homeostasis and compensating for various disturbances coming from the surroundings; 13. supervising, regulating, steering and integrating all the above functions; 14. of course, all these functions/goals/tasks would be subordinated to the proliferation of (the functional identity of) such artificial living individuals through production of a possibly large number of copies possibly similar to the original. These functions involve p-processes, signals, feedbacks, re-writing and decoding processes.
Natural spontaneously-originated living individuals cope with tasks analogous to the enumerated tasks without any problems – these are functions for executing of which they originated and evolved. The discussed functions are implemented in them, more or less directly, by molecular machines possessing the character of complex organic compounds.
However, it is not possible (at least to the author) to imagine a structurally-integrated function-based individual based on the supra-molecular level, a kind of a monstrous robot (Miglino et al. 1995; Lipson and Pollack 2000), that would carry out all these sorts of functions. Therefore, supra-molecular artificial individuals, for example introduced and left alone on a distant planet/moon, can only be dispersed individuals, integrated only functionally, but not structurally, composed of several sub-individuals, resembling robots, machines, factories, vehicles, computers, information-processing and control centers, and so on, interconnected between each other by a rich network of interactions, but not firmly attached physically (Korzeniewski 2011, 2023). Such individuals could not originate spontaneously, but only be created by intelligent beings. They would resemble to some extent the units of the human technical civilization, which, however, would not be supervised and controlled by intelligent beings, but left for themselves in a separated environment (e.g., the surface of a distant planet or moon). Such artificial supra-molecular function-based life/living individuals may seem monstrous, but this is the only way how a supra-molecular system can become a function-based living individual.
Such artificial supra-molecular individuals would not develop by themselves from some relatively simple “germ” (like zygote) (through a process analogous to morphogenesis in natural molecular life), but be constructed from elements by parental individuals.
Artificial supra-molecular living individuals would have to perform the same general types of functions as natural molecular individuals: Ps, Ss, Fs, R(s) and D(s). However, these functions in detail would look completely differently. They would resemble various technological processes in human factories, mines and power stations (Ps), supervision, control and steering of technological processes by various computes and homeostatic robots, communication through cables and electromagnetic radiation (Ss and Fs), copying of information records concerning the self-construction, functioning and reproduction using just two “informational monomers”: 0 and 1 (digital devices) (R(s)) and realization of these records through sustaining of the current functioning of the individuals themselves and constructing descendant individuals, also analogous to that carried out by present complex technological systems (complex system of processes involving D(s), for instance transformation of the sequence of 0s and 1s into the sequence of commands in a construction prescription and its realization).
14. Virtual Artificial Life
The presently-known (at least to the author) virtual life is devoid of any sufficiently complex environment varying in space and time, with different disturbances, opportunities and dangers coming from it. Also, as a consequence, the artificial “living” individuals themselves can be very simple. The game “Life” is of course no life at all, it is not “alive”. In order to create “true” virtual life it would be necessary to program complex individuals stiving for survival and reproduction in a complex virtual environment, competing and interacting with other similar individuals, possibly feeding on and being fed on by other, significantly different individuals. The complexity of the function-based living individual is proportional to the number of inter-related functions in its causally-closed functional network. The number of functions reflects the number of various aspects of the variable environment the individual must cope with and respond to. Therefore, the more complex the environment (and the ecological niche occupied in it), the more complex is the function-based individual.
15. General Discussion
One could doubt whether decoding D(s) is a necessary element of a living individual. Without decoding, the same (type of) molecules would have to fulfil both the function of the identity record and executive machinery (and many others). This would very severely limit their efficiency, versatility and the degree of complexity of the systems based on them. It is problematic whether such systems could spontaneously originate at all. In the author’s opinion they could not. For instance, no RNA molecules, either natural or artificially synthesized, are capable of self-replication (synthesis of a complementary strand from free single nucleotides), despite very intense search for such molecules. Additionally, decoding enables specialization: a certain type of molecules (e.g., nucleic acids) can fulfil the function of information record and another type of molecules (e.g., proteins) the function of molecular machines and structural elements. In the author’s opinion, in the real complex physical world (in general: in any sufficiently complex environment, which concerns also virtual life) no individuals that do not exhibit this specialization are possible. Therefore, a full-fledged living individual must contain (be dynamically “built of”) all five enumerated types of functions, including decoding.
There occurs a kind of a recursive loop or vicious circle between the genetic record, on the one hand, and the genetic code and the whole genetic apparatus, on the other hand. The genetic code (the correspondence between the codon triplets of nucleotides in mRNA and amino-acids in proteins) and the whole genetic apparatus is contained in the genetic record (through genes for tRNA, rRNA, AA-tRNA synthases, ribosome proteins, DNA and RNA polymerases and so forth), while the genetic record is copied and realized by the genetic apparatus using the genetic code. All these processes associated with the genetic record copying and realization are controlled and regulated (activated or inhibited) by transcription factors (proteins) and small RNA molecules (e.g., miRNA), which are in turn encoded in the genetic record (DNA and/or RNA), which also possesses binding sequences for these regulatory molecules. This constitutes another manifestation of the recursive causal closure so characteristic for the functional networks of living individuals. And, in the author’s opinion, this recursive loop based on the self-application relation constitutes an inseparable element of the phenomenon of life (the living individual).
The function-based living individual constitutes the unit of natural selection and Darwinian evolution. This is certainly not a “selfish gene” (Dawkins 1976), as only a causally-closed self-copying system self-dependent and self-sufficient in realization of this task in its environment can be regarded as a “true” subject to evolution (see Korzeniewski 2023 for a detailed discussion).
16. Biological Individual vs. Psychical Individual
It was proposed previously how consciousness and self-consciousness emerges in the result of the neuronal network functioning in the human brain (Korzeniewski 2010, 2013, 2015, 2020, 2023). Namely, the special unique kind of the functional organization of the information transfer by neurons and neural networks that underlies the psychical individual endowed with the subjective sphere of mental phenomena was postulated. In other words, a functional definition of the psychical individual was formulated. Its essence can be summarized in four points. First, the neural network constituted by the system of dynamic connections between neurons within the brain, in particular its cognitive part, was shaped (during both biological evolution and individual development) as an operational representation of the external world: the functional “knots” of this network correspond to different, variable in space and time, aspects, objects, processes, properties and regularities of the physical, biological and social environment. Second, the elements of the functional neural network: neurons and their complexes mean by connotation, mutual reference to each other, as particular neurons co-condition each other, constitute the functional context for other neurons, and ultimately only the whole network of neurons, receptors and effectors determines the role and sense of all its components. Third, the neural network is intentionally (in the operational sense) directed on the survival and reproduction of its “carrier”, that is the whole living individual equipped with it (the neural network is only a part of the functional network of the biological individual). Fourth, psychical consciousness and self-consciousness emerge due to the recursive and recurrent relation of self-directing on itself of the “cognitive center” in the human brain. This center, which in beings devoid of self-consciousness receives processed stimuli coming, directly or indirectly, from the sense organs and the sensory part of the brain, in beings endowed with self-consciousness receives and processes stimuli also coming from itself. In other words, this center becomes self-directed on itself, its own self-representation, formation of its own model within itself, which is especially useful in social relations and language development (Korzeniewski 2020). As a result, it is continuously creating a map of itself within itself, awareness of its own existence in time and space. These are in short the most important characteristics of the functional psychical individual.
Of course, evident similarities between the functional biological and psychical individual can be noticed. It was proposed that both types of individualsare analogous and to a large extent formally isomorphic, and therefore a unified definition of an individual was formulated (Korzeniewski 2013, 2023). This definition, after some function-based re-formulation3, states that an individual (biological or psychical) is a system constituted by a dynamic network of functionally-interconnected processes (PSFRD functions or neural signals) that possesses the following common properties: (1) it is intentional (in the operational sense); (2) its elements have meaning (sense) through connotation (mutual relations between them); (3) it contains an instrumental (procedural) representation of the (aspects of the variable in space and time) world; (4) it is self-referent, recurrently and recursively directed on itself (its own self-copying or self-representation). Therefore, the phenomena of life and self-consciousness exhibit deep formal functional similarities. The special kind of complexity characterizing both biological and psychical individuals, that is the self-directing on itself of an operationally-intentional network of elements constituting an instrumental representation of the world, is responsible for the very distinguished status of the phenomenon of life and (self-)consciousness. Generally, this is an exemplification of the relation of self-applicability, underlying famous logico-mathematical paradoxes: the liar’s paradox, Russell’s class antinomy and Gödel’s incompleteness theorem.
In short, the phenomena of life (biological individual) and self-consciousness (psychical individual) originated, one by another, when some instrumentally intentional dynamic network of elements meaning by connotation, being an operational representation of the external world, became recursively directed on itself. It was argued that some degree of “individuality” can also be attributed to various social, economic and political systems (Korzeniewski 2013).
17. Conclusions
The causally-closed functional-network-based definition of life determines the function-based living individual as a causally-closed network of five function types: purposeful processes (Ps), signals (Ss), feedbacks (Fs), re-writing (R(s)) and decoding (D(s)) recursively directed on the superior function: survival (self-sustaining) and reproduction, proliferation of its own identity in its environment/ecological niche, being self-sufficient in the realization of this goal. This definition:
► determines in a clear and univocal way the function-based living individual, which is a fundamental constituent of any possible form of life, and thus allows to distinguish living systems from inanimate phenomena;
► quotes general properties that characterize in an abstract way the functional core of the essence of life and the living individual;
► demonstrates which properties and aspects of the contemporary terrestrial life and how result from the concrete physicochemical nature of the real environment on the Earth;
► emphasizes that the kind and degree of complexity of the functional organization of living individuals reflects the kind and degree of complexity and variability in space and time of the concrete physical, biological and social environment (ecological niche);
► decides which systems and phenomena are living individuals, and to what degree, within the whole vital hierarchy of biological phenomena on the Earth;
► identifies unambiguously living phenomena on other planets/moons in the universe, as well as artificial and virtual life created by intelligent beings;
► distinguishes properties that are immanent for the abstract concept of life and living individual and thus specify the core of its essence form properties of the real terrestrial life resulting from the nature of the physical external world they exist in and the chain of chances underlying the process of the origination and evolution of life.
The life on Earth originated in the moment when a system appeared that contained all the enumerated types of functions. In particular, the first living individual was formed when the genetic code, establishing the correspondence between amino-acid sequences in proteins and nucleotide sequences in nucleic acids (primarily RNA), representing the D function, was formed. This happened in systems resembling coacervate-like bubbles capable of self-dependent growth and divisions (proto-cells), hosting some proto-metabolism (Ps) and possibly equipped with some stabilizing processes (Ss and Fs), that contained auto-catalytic cycles of proteins and RNA (R). The former, constituted by proteins with a random amino-acid sequences and diverse set of properties, “by a chance” supported the latter and proto-metabolism. The first “key” to the genetic code, proto-tRNA, coupled proteins and RNA into hyper-cycles. This is the “everything first” scenario of the origin of life.
There exist significant formal similarities between the essence of the functional-network-based living individual (phenomenon of life) and neuronal-network-based psychical individual (phenomenon of subjective psyche and self-consciousness). Both constitute a result of the relation of recursive self-directing on itself of an operationally-intentional dynamic network of elements meaning by connotation that constitutes an instrumental representation of the external (and internal) world.
Funding
There are no funders to report for this submission.
References
- Adami, C. Introduction to artificial life; Springer-Verlag: Santa Clara, 1998. [Google Scholar]
- Adami, C. What is Complexity? BioEssays 2002, 24, 1085–1094. [Google Scholar] [CrossRef]
- Al-Khalili, J.; McFadden, J. Life on the edge. The coming of age of quantum biology; Bantam Press, 2014. [Google Scholar]
- Baggott, J. Origins: The scientific story of creation; Oxford University Press: Oxford, 2015. [Google Scholar]
- Banich, M.T.; Compton, R.J. Cognitive Neuroscience; Cambridge University Press: Cambridge, 2018. [Google Scholar]
- Barrack, J.E.; Breaker, R.R. The Power of Riboswitches. Sci. Am. 2007, 296(1), 50–57. [Google Scholar] [CrossRef]
- Barski, G.; Cornefert-Jensen, F. Cytogenetic study of Sticker venereal sarcoma in European dogs. J. Natl. Canc Inst. 1966, 37(6), 787–797. [Google Scholar] [CrossRef]
- Bedau, M.A. Four puzzles about life. Artif. Life 1998, 4(2), 125–140. [Google Scholar] [CrossRef] [PubMed]
- Bougourd, S.M.; Jones, R.N. B chromosomes: a physiological enigma. New Phytol. 1997, 137, 43–54. [Google Scholar] [CrossRef]
- Brooks, D.; Wiley, E.O. Evolution as entropy. Toward a unified theory of biology; University of Chicago Press: Chicago & London, 1986. [Google Scholar]
- Brown, T. Introduction to genetics: a molecular approach; Garland Science: New York & London, 2012. [Google Scholar]
- Carroll, S.B.; Prud’homme, B.; Gompel, N. Regulating evolution: how gene switches make life. Sci. Am. 2008, 298(5), 60–67. [Google Scholar] [CrossRef]
- Cech, T.R. The RNA worlds in context. Cold Spring Harb. Perspect. Biol. 2012, 4(7), a006742. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. Obcells as proto-organisms: membrane heredity, lithophosphorylation, and the origins of the genetic code, the first cells, and photosynthesis. J. Mol. Evol. 2001, 53, 555–595. [Google Scholar] [CrossRef]
- Chambers, J.; Mitton, J. From dust to life. The origin and evolution of our solar system; Princetin University Press: Princeton and Oxford, 2014. [Google Scholar]
- Chance, B.; Williams, G.R. The respiratory chain and oxidative phosphorylation. Adv. Enzymol. 1956, 17, 65–134. [Google Scholar] [CrossRef]
- Copley, S.D.; Smith, E.; Morowitz, H.J. The origin of the RNA world: co-evolution of genes and metabolism. Bioorg Chem. 2007, 35(66), 430–43. [Google Scholar] [CrossRef] [PubMed]
- Cryer, P.E. Regulation of glucose metabolism in man. J. Int. Med. Suppl. 1991, 735, 31–39. [Google Scholar]
- Darwin, C. The origin of species by means of natural selection, or the preservation of favored races in the struggle for life; Modern Library: New York, 1859. [Google Scholar]
- Dawkins, R. The selfish gene; Oxford University Press: Oxford, 1976. [Google Scholar]
- Doolittle, W.F.; Sapienza, C. Selfish genes, the phenotype paradigm and genome evolution. Nature 1980, 284, 601–603. [Google Scholar] [CrossRef]
- Deamer, D.W. Assembling life: How can life begin on earth and other habitable planets; Oxford University Press: Oxford, 2019. [Google Scholar]
- Deamer, D.W. Origin of life. What everyone needs to know; Oxford University Press: Oxford, 2020. [Google Scholar]
- de Farias, S.T.; dos Santos, AP Jun; Rêgo, T.G.; José, M.V. Origin and evolution of RNA-dependent RNA polymerase. Front Genet 2017, 8, 125. [Google Scholar] [CrossRef]
- Eigen, M.; Schuster, P. The hypercycle: a principle of natural self-organization; Springer: Berlin, 1979. [Google Scholar]
- Futuyma, D.J. Evolution; Sinauer Associates, 2013. [Google Scholar]
- Gánti, T. Organisation of chemical reactions into dividing and metabolizing units: the chemotons. BioSystems 1975, 7(1), 189–195. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, S.F. Developmental biology; Sinauer Associates: Sunderland, 2010. [Google Scholar]
- Haldane, J.B.S. The origin of life. Ration. Annu. 1929, 148, 3–10. [Google Scholar]
- Harris, H.M.B.; Hill, C. A place for viruses on the tree of life. Front Microbiol. 2021, 11, 604048. [Google Scholar] [CrossRef] [PubMed]
- Jacob, F.; Monod, J. On the regulation of gene activity. Cold Spring Harb. Symp. Quant. Biol. 1961, 26, 193–211. [Google Scholar] [CrossRef]
- Jones, R.N. B-Chromosome system in flowering plants and animal species. Int. Rev. Cytol. 1975, 40, 1–100. [Google Scholar] [CrossRef]
- Kauffman, S.A. Autocatalytic sets of proteins. J. Theor. Biol. 1986, 119(1), 1–24. [Google Scholar] [CrossRef]
- Kee, T.P.; Monnard, P.-A. On the emergence of a proto-metabolism and the assembly of early protocells. Elements 2016, 12(6), 419–424. [Google Scholar] [CrossRef]
- Knoll, A.H. A brief history of earth: four billion years in eight chapters; Harper Collins, 2021. [Google Scholar]
- Korzeniewski, B. Regulation of ATP supply during muscle contraction: theoretical studies. Biochem. J. 1998, 330, 1189–1195. [Google Scholar] [CrossRef]
- Korzeniewski, B. Cybernetic formulation of the definition of life. J. Theor. Biol. 2001, 209(3), 275–286. [Google Scholar] [CrossRef]
- Korzeniewski, B. Confrontation of the cybernetic definition of a living individual with the real world. Acta Biotheor. 2005, 53(1), 1–28. [Google Scholar] [CrossRef] [PubMed]
- Korzeniewski, B. From neurons to self-consciousness. How the brain generates the mind; Humanity Books: Amherst, 2010. [Google Scholar]
- Korzeniewski, B. Artificial cybernetic living individuals based on supramolecular-level organization as dispersed individuals. Artif. Life 2011, 17(1), 51–67. [Google Scholar] [CrossRef]
- Korzeniewski, B. Formal similarities between cybernetic definition of life and cybernetic model of self-consciousness: universal definition/model of individual. Open J. Philos. 2013, 3(2), 314–328. [Google Scholar] [CrossRef]
- Korzeniewski, B. Regulation of oxidative phosphorylation through each-step activation: evidence from computer modeling. Prog. Biophys. Mol. Biol. 2017, 125, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Korzeniewski, B. Self-consciousness as product of biological evolution. J. Conscious. Stud. 2020, 27(7-8), 50–76. [Google Scholar]
- Korzeniewski, B. How life emerges from inanimate matter: From matter to a living individual; Cambridge Scholars Publishing: Newcastle upon Tyne, UK, 2023. [Google Scholar]
- Koshland, D.E., Jr. The seven pillars of life. Science 2002, 295(5563), 2215–2216. [Google Scholar] [CrossRef]
- Lane, N. Life ascending. The ten greatest inventions of evolution; W. W. Norton & Company, 2010. [Google Scholar]
- Lane, N. The vital question. Why is life the way it is? Profile Books, 2015. [Google Scholar]
- Lipson, H.; Pollack, J. Automatic design and manufacture of robotic life forms. Nature 2000, 406(6799), 974–978. [Google Scholar] [CrossRef] [PubMed]
- Macdougall, J.D. A short history of planet earth. In Mountains, mammals, fire and ice; John Wiley & Sons: New York, 1996. [Google Scholar]
- Maynard Smith, J.; Szathmáry, E. The major transitions in evolution; W.H. Freeman: Oxford, 1995. [Google Scholar]
- McShea, D.W. Complexity and evolution: what everybody knows. Biol. Philos. 1991, 6, 303–324. [Google Scholar] [CrossRef]
- Miglino, O.; Lund, H.H.; Nolfi, S. Evolving mobile robots in simulated and real environments. Artif. Life 1995, 2(4), 417–434. [Google Scholar] [CrossRef] [PubMed]
- Muller, H.J. (1935) Life. Science 121(3132), 1–9. [CrossRef]
- Muller, H.J. The gene material as the initiator and organizing basis of life. Am. Nat. 1966, 100(915), 493–517. [Google Scholar] [CrossRef]
- Nair, A.; Chauhan, P.; Saha, B.; Kubatzky, K.F. Conceptual Evolution of Cell Signaling. Int. J. Mol. Sci. 2019, 20(13), 3292. [Google Scholar] [CrossRef]
- Nakashima, T. Metabolism of proteinoid microspheres. Top. Curr. Chem. 1987, 139, 58–81. [Google Scholar]
- Navarro, B.; Flores, R.; Di Serio, F. Advances in Viroid-Host Interactions. Annu. Rev. Virol. 2021, 8(1), 305–325. [Google Scholar] [CrossRef] [PubMed]
- Nur, U.; Werren, J.H.; Eickbush, D.G.; Burke, W.D.; Eickbush, T.H. A “selfish” B chromosome that enhances its transmission by elimination of the paternal genome. Science 1988, 240(4851), 512–514. [Google Scholar] [CrossRef] [PubMed]
- Oparin, A.I. The Origin of Life; Dover Publications: New York, 1938. [Google Scholar]
- Orgel, L.E.; Crick, F.H.C. Selfish DNA: the ultimate parasite. Nature 1980, 284(5757), 604–607. [Google Scholar] [CrossRef]
- Papaccio, F.; Paino, F.; Regad, T.; Papaccio, G.; Desiderio, V.; Tirino, V. Concise Review: Cancer Cells, Cancer Stem Cells, and Mesenchymal Stem Cells: Influence in Cancer Development. Stem Cells Transl. Med. 2017, 6(12), 2115–2125. [Google Scholar] [CrossRef]
- Pray, L.A. Transposons: The jumping genes. Nat. Educ. 2008, 1(1), 204. [Google Scholar]
- Prigogine, I. From being to becoming: Time and complexity in physical sciences; W.H. Freeman: San Francisco, 1980. [Google Scholar]
- Prigogine, I.; Stengers, I. Order out of chaos. Man’s new dialogue with nature; Heinemann: London, 1984. [Google Scholar]
- Prusiner, S.B. Prions. Proc. Natl. Acad. Sci. USA 1998, 95(23), 3363–13383. [Google Scholar] [CrossRef] [PubMed]
- Reece, J.B.; Urry, L.A.; Cain, M.L.; Wasserman, S.A.; Minorsky, P.V.; Jacksomn, R.B. Campbell Biology; Pearson: Boston, 2011. [Google Scholar]
- Ricardo, A.; Szostak, J.W. The Origin of Life on Earth. Sci. Am. 2009, 301(3), 54–61. [Google Scholar] [CrossRef] [PubMed]
- Schrödinger, E. What is life? Press Syndicate of the University of Cambridge: Cambridge, 1992. [Google Scholar]
- Smith, E.; Morowitz, H.J. The origin and nature of life on earth. The emergence of the fourth geosphere; Cambridge University Press: Cambridge, 2016. [Google Scholar]
- Smokers, I.B.A.; van Haren, M.H.I.; Lu, T.; Evan Spruijt, E. Complex coacervation and compartmentalized conversion of prebiotically relevant metabolites. ChemSystemsChem 2022, 4, e202200004. [Google Scholar] [CrossRef]
- Trifonov, E.N. Vocabulary of definitions of life suggests a definition. J. Biomolec Struct. Dynam 2011, 29(2), 259–266. [Google Scholar] [CrossRef] [PubMed]
- Tsokolov, S.A. Why is the definition of life so elusive? Epistemological Considerations. Astrobiology 2009, 9(4), 401–412. [Google Scholar] [CrossRef]
- Umbarger, H.E. Amino acid biosynthesis and its regulation. Annu Rev. Biochem 1978, 47, 533–606. [Google Scholar] [CrossRef]
- Weiss, M.C.; Sousa, F.L.; Mrnjavac, N.; Neukirchen, S.; Roettger, M.; Nelson-Sathi, S.; Martin, W.F. The phylogeny and habitat of the last universal common ancestor. Nat. Microbiol. 2016, 1, 16116. [Google Scholar] [CrossRef]
- Williams, G.C. The pony fish’s glow and other clues to plan and purpose in nature; Basic Books, 1997. [Google Scholar]
- Wilson, E.O. The insect societies; Harvard University Press: Cambridge, MA, 1971. [Google Scholar]
- Wolpert, L. Positional information and pattern formation. Curr. Top. Dev. Biol. 2016, 117, 597–608. [Google Scholar] [CrossRef]
- Woods, G.M.; Howson, L.J.; Brown, G.K.; Tovar, C.; Kreiss, A.; Corcoran, L.M.; Lyons, A.B. Immunology of a transmissible cancer spreading among tasmanian devils. J. Immunol. 2015, 195(1), 23–29. [Google Scholar] [CrossRef]
- Yarlett, N.; Hackstein, J.H.P. Hydrogenosomes: one organelle, multiple origins. BioScience 2005, 55(8), 657–668. [Google Scholar] [CrossRef]
Figure 1.
A very simple example of a functional network, involving p-processes (P), signals (S), feedbacks (F), re-writing (R(s)) and decoding (D(s)) occurring within and outside the skeletal muscle cell (myocyte). The details are described in the text.
Figure 1.
A very simple example of a functional network, involving p-processes (P), signals (S), feedbacks (F), re-writing (R(s)) and decoding (D(s)) occurring within and outside the skeletal muscle cell (myocyte). The details are described in the text.

Figure 2.
An abstract presentation of the function-based living individual. Various numerous p-processes (Ps), signals (Ss), feedbacks (Fs), re-writing processes (R(s)) and decoding processes (D(s)) form a causally-closed network of functions directed only and exclusively on its own proliferation.
Figure 2.
An abstract presentation of the function-based living individual. Various numerous p-processes (Ps), signals (Ss), feedbacks (Fs), re-writing processes (R(s)) and decoding processes (D(s)) form a causally-closed network of functions directed only and exclusively on its own proliferation.

| 1 | The term introduced by Eigen and Schuster (1979) in a slightly different context. |
| 2 | It is possible that identical monomers in strands of opposite polarity (like complementary strands in DNA and RNA) will thus fit to each other like two identical right-angle triangles turned in relation to each other by 180°. |
| 3 | Strictly speaking, the cybernetic living individual was considered there, as the functional definition of the living individual is only proposed in the present study. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



