Submitted:
05 May 2026
Posted:
07 May 2026
You are already at the latest version
Abstract
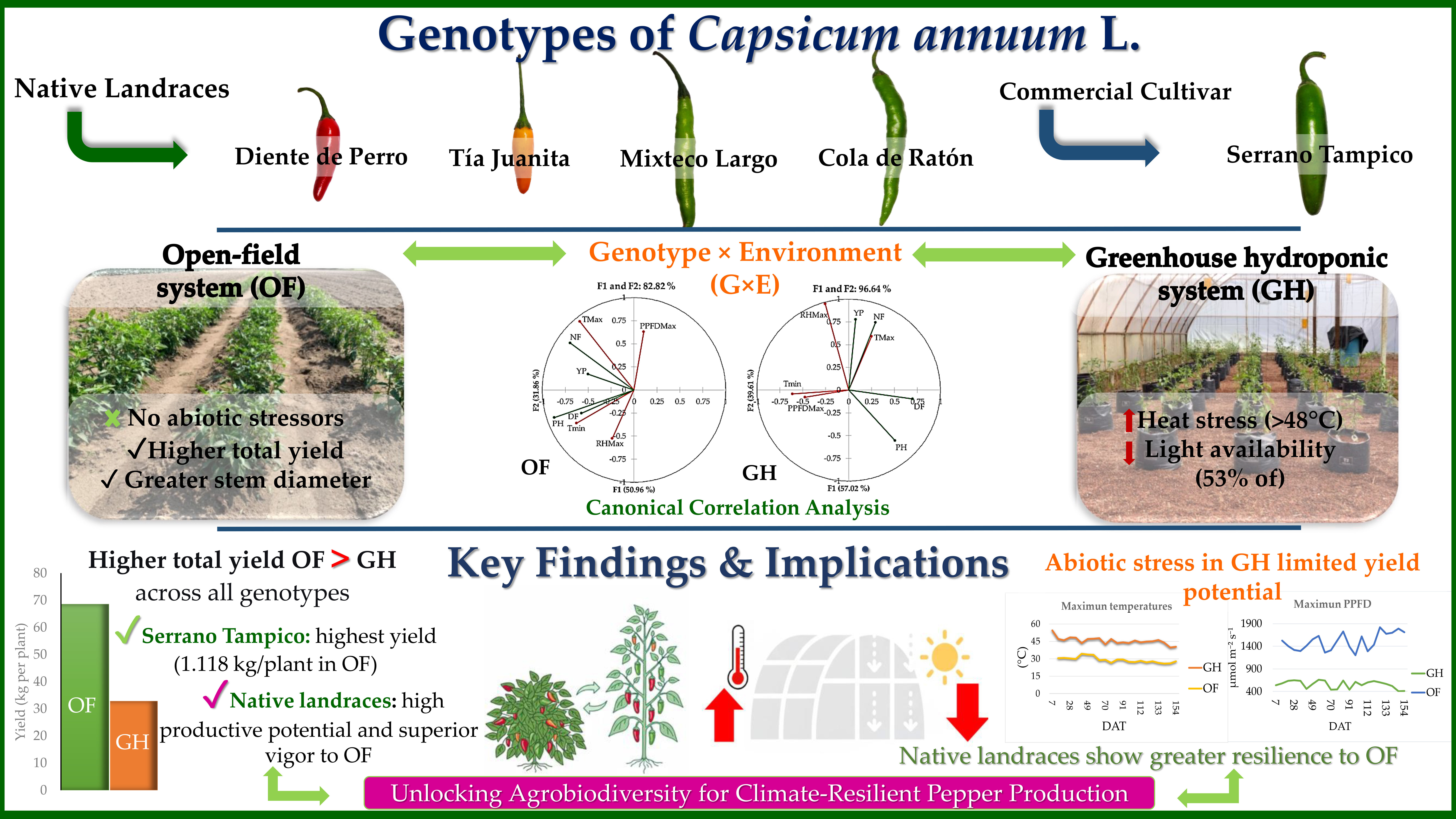
The performance of native landraces of Capsicum annuum L. under contrasting production systems remains poorly understood, limiting their integration into optimized production schemes. This study evaluated the phenological and productive responses of five genotypes (four native landraces and one commercial cultivar) under two sys-tems representing locally relevant production conditions: open-field (OF) and a sub-strate-based hydroponic system under low-technology, passively ventilated tunnel-type greenhouse conditions (GH), in order to identify differences in genotype performance across environments during the 2023 growing season in Puebla, Mexico. Agroclimatic and agronomic variables were analyzed using independent ANOVA by system and canonical correlation analysis (CCA). The GH system exhibited restrictive microclimatic conditions, with maximum temperatures exceeding 48 °C and photo-synthetically active radiation reduced by approximately 53% compared to OF conditions. Environmental conditions were not standardized between systems; therefore, the results reflect the contrasting microclimates of locally relevant production systems and provide a context-specific assessment of genotype performance. These conditions were associated with reduced yield across genotypes under GH, whereas higher productivity levels were observed under OF conditions. The commercial cultivar Serrano Tampico achieved the highest yield (1.118 kg per plant under OF), while Mixteco Largo and Cola de Ratón produced the highest number of fruits. The CCA revealed a strong genotype × environment (G×E) interaction, with native landraces showing greater adaptation to open-field conditions. Overall, the results provide evidence of the sensitivity of these materials to the production environment and underscore the need for system-specific selection and management strategies.
Keywords:
Capsicum annuum
; native landraces
; phenology
; genotype × environment interaction
; agroclimatic variables
; abiotic stress
1. Introduction
Chili pepper (Capsicum annuum L.) is a crop of global strategic importance, with significant economic, nutritional, and cultural value [1,2]. This species, belonging to the Solanaceae family, exhibits extensive genetic diversity, which confers high phenotypic plasticity and enables adaptation to a wide range of environmental conditions [3]. As a result, its global production exceeds 44 million tons annually, consolidating its role as a key crop in both traditional and intensive production systems [4]. This genetic diversity not only supports its broad geographic distribution but also provides a basis for the selection of genotypes with specific agronomic traits, particularly under varying environmental conditions [5].
Beyond its dietary importance, C. annuum is characterized by a complex biochemical composition that determines its commercial and functional value [6,7,8,9]. From an agronomic perspective, however, the accumulation of these compounds is closely linked to the physiological status of the plant and its response to the growing environment [10]. Therefore, current research focuses on understanding how environmental conditions influence phenotypic expression, particularly in terms of phenology, growth, and yield. In this context, evaluating crop performance under different production systems is essential to optimize crop stability and ensure the expression of genetic potential, as productivity is determined by the interaction between genotype and environment [11].
Under open-field conditions, factors such as climatic variability, temperature fluctuations, water availability, and solar radiation introduce high production uncertainty and limit crop efficiency [12,13,14]. These factors directly affect key physiological processes, including photosynthesis, transpiration, and assimilate partitioning, leading to variation in growth and yield. In particular, high temperatures can reduce photosynthetic efficiency [15], while water deficit limits cell expansion and reproductive development [15,16].
As an alternative, protected agriculture has enabled partial control of the growing environment through the use of greenhouses, thereby improving growing conditions and reducing environmental variability [17]. In these systems, soilless cultivation, including hydroponic systems based on inert substrates, allows more precise management of water and nutrients, promoting the expression of productive potential [18,19]. In addition, these systems can improve resource-use efficiency, reduce the incidence of soil-borne pathogens, and facilitate more uniform crop development [20,21]. However, the performance of protected systems critically depends on environmental control [22]. Under conditions of limited ventilation and high external solar radiation, greenhouses may experience excessive heat accumulation, while plastic covers can reduce the availability of photosynthetically active radiation, jointly inducing abiotic stress [23,24]. In particular, stress derived from these conditions affects key physiological processes such as photosynthesis, cellular integrity, metabolic activity, and pollen viability [25,26], ultimately compromising reproductive development and yield.
In this context, the genotype × environment (G×E) interaction provides a fundamental framework for understanding variability in agronomic performance [27]. This interaction describes the differential response of genotypes under contrasting environmental conditions, which is reflected in changes in key agronomic traits [28]. The magnitude of this interaction depends on both genetic variability and environmental heterogeneity. In particular, phenological stages such as flowering and fruiting are highly sensitive to environmental factors including temperature, radiation, and humidity [29,30], making them useful indicators for identifying adaptation patterns and resource-use strategies among genotypes. These insights are particularly relevant for breeding programs and the conservation of native landraces, as they support the identification of genotypes with enhanced adaptation to specific environmental conditions.
Mexico, as one of the main centers of origin and diversification of C. annuum, harbors extensive genetic diversity represented by native landraces that have been selected under local conditions over generations [31,32]. These materials constitute a valuable genetic resource, particularly in terms of adaptation to variable environments and resilience to abiotic stress [33,34]. However, the adoption of commercial cultivars in intensive systems has contributed to a decline in the use of these native materials [35,36], which may lead to genetic erosion and loss of agricultural diversity, creating a knowledge gap regarding their performance under different production systems.
Under this framework, genotype performance may vary significantly across production systems, reflecting the influence of environmental conditions and management practices on phenological and productive responses. This variability highlights the importance of evaluating genotype behavior under contrasting growing environments to identify adaptation patterns and differences in agronomic performance associated with genotype × environment interaction. Therefore, the objective of this study was to characterize the phenological and productive responses of five C. annuum genotypes (four native landraces and one commercial cultivar) under two contrasting production systems: open-field and a substrate-based hydroponic system under low-technology greenhouse conditions, in order to identify differences in genotype performance across environments. It was hypothesized that contrasting production environments would differentially influence phenological and productive performance, revealing a significant genotype × environment interaction, with the greenhouse system expected to enhance productivity.
2. Materials and Methods
2.1. Experimental Site
The study was conducted in San Agustín Calvario, San Pedro Cholula, Puebla, Mexico, during the 2023 growing season (January–October), under two contrasting production systems: open field (OF) and a substrate-based hydroponic system under low-technology greenhouse conditions (hereafter referred to as the GH) (Figure 1).
Open-field system (OF): The experiment was established in a cooperating farmer’s field located at 19°03′18″ N, 98°20′08″ W, at 2164 m a.s.l. The soil had a pH of 7.30, electrical conductivity (EC) of 0.75 dS·m−1, and organic matter content of 0.74%.
Substrate-based hydroponic system under low-technology greenhouse conditions (GH): The experiment was conducted in a low-technology tunnel-type greenhouse (100 m2), covered with polyethylene (caliber 800; 200 μm) and protected with anti-aphid mesh, located at 19°02′59″ N, 98°20′03″ W, at 2158 m a.s.l. The plastic cover reduced light transmission by approximately 15% and filtered 76% of ultraviolet radiation.
2.2. Plant Material
Five genotypes of C. annuum were evaluated: four native landraces (Diente de Perro, Tía Juanita, Mixteco Largo, and Cola de Ratón) and one commercial cultivar (Serrano Tampico). Taxonomic identity was confirmed by the Botanical Garden of BUAP (herbarium vouchers 88725–88729). Plant morphology is shown in Figure 2.
2.3. Seedling Production
For each genotype, 200 seeds were sown in 200-cavity polystyrene trays filled with commercial substrate (Sunshine® Mix 3). One seed per cavity was used; no thinning was required. Seedlings were grown under nursery conditions (25 ± 2 °C, 12 h photoperiod) for 80-89 days, and only the most vigorous and uniform seedlings were selected for transplanting. To synchronize plant establishment under each system, sowing dates were adjusted: January 29, 2023 for OF and February 19, 2023 for GH. This offset compensated for differences in early growth rates and ensured uniform vigor at transplanting. Germination percentage was below 100% and is detailed in Section 3.2.1.
2.4. Crop Management
2.4.1. Open-Field System (OF)
Experimental design and transplanting: Transplanting was performed on April 19, 2023 (80 days after sowing, das) under a randomized complete block design (RCBD) with four replicates. Plants were arranged in 6.0 m long rows spaced 0.80 m apart, with 0.50 m between plants, corresponding to a density of 25,000 plants·ha−1. Each experimental unit consisted of one row per genotype within each block (13 plants). The five central plants were evaluated to minimize border effects (20 plants per genotype).
Fertilization: A fertilization rate of 180–100–30 (N–P2O5–K2O) was applied using urea (46% N), diammonium phosphate (DAP; 18% N and 46% P2O5), and potassium chloride (60% K2O). One-third of the nitrogen and all phosphorus and potassium were applied at transplanting; the remaining nitrogen was split at 45 and 90 days after transplanting (dat).
Irrigation: Irrigation followed the farmer’s schedule. Furrow irrigation was initially applied and later replaced by sprinkler irrigation from 50 dat. Soil moisture sensors were not used.
Phytosanitary management: Preventive and corrective strategies were implemented.
Whitefly (Bemisia tabaci) was controlled using imidacloprid (350 g·L−1) applied at 0.75 L·ha−1. Bacterial diseases (Xanthomonas spp.) were managed using copper sulfate pentahydrate (≈25% Cu) at 1 L·ha−1. Soil-borne pathogens were controlled by alternating applications of Trichoderma harzianum (11.5 g·kg−1, 1.15%) at 15 mL·plant−1 with systemic fungicides, including propamocarb (530 g·L−1) + fosetyl-Al (310 g·L−1) applied at 6 L·ha−1, azoxystrobin (46 g·L−1) + chlorothalonil (460 g·L−1) at 3 L·ha−1, and carbendazim (500.76 g·L−1) at 2.5 L·ha−1.
Cultural practices: Periodic manual weeding and one tillage operation at 45 dat were performed to improve root development and weed control.
2.4.2. Substrate-Based Hydroponic System Under Low-Technology Greenhouse Conditions (GH)
This system corresponds to a substrate-based hydroponic scheme using an inert volcanic substrate (tezontle).
Experimental design and transplanting: Transplanting was carried out on May 19, 2023 (89 DAS). The experiment was conducted under a completely randomized design (CRD) with 12 replications. Each experimental unit was a black polyethylene bag (40 × 40 cm) filled with 11 kg of disinfected red tezontle (volcanic substrate), containing two plants, resulting in a density of 6.67 plants·m−2. One plant per unit was selected for measurements (12 plants per genotype).
Nutrient solution: The Steiner universal nutrient solution (1984) was used, with ionic concentrations (meq·L−1) of 12 NO3−, 1 H2PO4−, 7 SO42−, 7 K+, 9 Ca2+, and 4 Mg2+. The solution was prepared using KNO3, Ca(NO3)2, KH2PO4, MgSO4, K2SO4, and a micronutrient mix (Fe, Mn, B, Zn, Cu, Mo).
Irrigation and nutrient management: Solution concentration was adjusted according to phenological stage: 25% after transplanting, 50% during early vegetative growth, and 100% from 15 dat onward. Irrigation volume ranged from 250 mL to 1.5 L per plant. Electrical conductivity was maintained at 2.5 dS·m−1 and pH at 6.0 using phosphoric acid. Weekly leaching irrigation with water only was applied to prevent salt accumulation.
Phytosanitary management: Systemic insecticides included imidacloprid (350 g·L−1) applied at 0.75 L·ha−1 and a mixture of imidacloprid (210 g·L−1) + bifenthrin (72 g·L−1), applied in separate applications at 0.75 L·ha−1 during the flowering stage to control whitefly (B. tabaci), aphids (Aphididae), and thrips (Thysanoptera). Additionally, elemental sulfur (800 g·kg−1, 80% p/p) was applied at 2 g·L−1 as a preventive measure against fungal pathogens.
Cultural practices: Plants were supported using trellising with agricultural raffia.
2.5. Harvest Management
Fruits were harvested at physiological maturity, defined by size and characteristic color for each genotype. Eight harvests were performed under OF conditions and six under GH during the experimental period. In OF, harvests occurred between July and September, while in GH they were conducted from August to October 2023. This schedule allowed estimation of cumulative yield per plant.
2.6. Variables Evaluated
Phenological variables. Emergence time was evaluated, defined as the number of days elapsed from sowing until 50% of the seeds of each genotype had emerged. Emergence percentage was calculated as the proportion of seedlings that successfully emerged relative to the total number of seeds sown per genotype. In addition, days to flowering (DF) were recorded, defined as the time elapsed from transplanting to the anthesis of the first flower, and days to fruiting (DFR), corresponding to the time until the appearance of the first fruit. Both variables were determined when 50% of the plants of each genotype reached each stage.
Growth variables. Plant height (PH) and stem diameter (SD) were recorded weekly from transplanting. Plant height was measured as the vertical distance (cm) from the base of the stem to the apex, and stem diameter (mm) was measured using a digital caliper at 2 cm above the substrate level.
Yield variables. The number of fruits per plant (NF) was evaluated, defined as the total number of fruits developed and harvested at physiological maturity. Total yield per plant (YP) was calculated as the total fresh weight of fruits harvested per plant (kg per plant) at the end of the experimental period.
Agroclimatic variables. Agroclimatic variables were recorded continuously and simultaneously, including air temperature (°C), relative humidity (%), and photosynthetically active radiation, expressed as photosynthetic photon flux density (PPFD; μmol·m−2·s−1). Measurements were obtained using HOBO® dataloggers (models MX2301A, MX2202, and MX1104), configured to record data at 60-minute intervals throughout the experimental period.
2.7. Statistical Analysis
Phenological, growth, and yield data were analyzed separately for each production system. In OF, a randomized complete block design (RCBD) was used, while in GH a completely randomized design (CRD) was applied. In both cases, varietal effects were evaluated using analysis of variance (ANOVA), and means were compared using Tukey’s HSD test (α ≤ 0.05) with SAS Studio (SAS® OnDemand for Academics, version 3.1.0).
Additionally, canonical correlation analysis (CCA) was performed to evaluate relationships between agronomic variables (plant height, days to flowering, number of fruits, and yield) and agroclimatic variables (temperature, relative humidity, and PPFD), grouped by growth stage. Analyses were conducted using XLSTAT (version 2019.2.2) and SAS Studio.
3. Results
Significant genotype effects were detected for phenological, growth, and yield variables in both production systems, allowing characterization of genotype-specific performance within each cropping environment. Detailed results of the analysis of variance (ANOVA), including mean squares, coefficients of variation, and coefficients of determination, are presented in Table S1 (Supplementary Material).
3.1. Agroclimatic Conditions
Agroclimatic variables exhibited distinct patterns within each production system, reflecting substantial differences in the crop microenvironment (Figure 3). Due to differences in experimental design, plant density, and agronomic management, comparisons between systems were addressed descriptively, prioritizing the independent characterization of each environment.
3.1.1. Air Temperature
In the open field (OF), air temperature exhibited a defined seasonal pattern throughout the crop cycle. During the initial period (14–35 dat), the average temperature was 16.2 °C, with a minimum of 9.36 °C. The maximum value of the cycle (32.80 °C) occurred during the vegetative stage, followed by a progressive decrease toward the maturation stage, where an average of 17.3 °C was recorded (Figure 3A).
In the greenhouse hydroponic system (GH), temperature was characterized by higher values throughout the cycle (Figure 3B). During the initial and vegetative stages, maximum temperatures exceeded 30 °C, reaching a peak of 48.78 °C, above the optimal ranges reported for the crop. Toward the final stage of the experimental period (119–154 dat), temperature stabilized, with an average of 22.3 °C, consistent with the seasonal transition toward autumn.
3.1.2. Relative Humidity
Relative humidity (RH) exhibited contrasting patterns between environments (Figure 3C, D). In OF, a marked diurnal variation was observed throughout the cycle, with maximum values during early morning (87–96%) and minimum values at midday (23–35%). During the initial period, the widest range of RH was recorded (23.85–90.88%), with an average of 72.4%. During the vegetative stage, RH decreased to 58.2%, whereas during the maturation stage it increased to 78.3%, with less pronounced daytime minima.
In GH, RH remained relatively high and more stable throughout the cycle (Figure 3D), with average values ranging from 78.2% to 83.7%. During the initial period, the widest range was observed (8.93–92.4%). During the vegetative stage, variation became more restricted, with an average of 77.8% and maximum values close to 94.8%. During reproductive and maturation stages, RH variability decreased further, with minimum values above 30% and maximum values close to 95%, indicating a more humid and stable environment.
3.1.3. Photosynthetic Photon Flux Density (PPFD)
The availability of photosynthetic photon flux density (PPFD) showed marked differences between environments (Figure 3E, F). In OF, the average PPFD during the cycle was 523.4 μmol m−2 s−1. During the initial period, the average was 458.2 μmol m−2 s−1, whereas the vegetative stage recorded the highest values (712.8 μmol m−2 s−1) and the maximum peak of the cycle (1818.8 μmol m−2 s−1). During the maturation stage, PPFD decreased to 543.6 μmol m−2 s−1, consistent with seasonal progression.
In GH, the average PPFD was 247.3 μmol m−2 s−1, representing an approximate 53% reduction relative to OF (Figure 3F). During the initial period, the average was 278.5 μmol m−2 s−1, with maximum values of 643.2 μmol m−2 s−1. During the vegetative stage, the highest average (312.8 μmol m−2 s−1) and the maximum value of the system (657.0 μmol m−2 s−1) were recorded. Toward the maturation stage, PPFD decreased to 183.6 μmol m−2 s−1. Radiation attenuation showed seasonal variation, with greater reductions during summer (56–64%) compared to autumn (40–45%), indicating consistent light limitation in GH.
3.2. Phenological Development
3.2.1. Emergence Time and Percentage
Emergence parameters were evaluated under controlled nursery conditions (identical for both production systems) prior to transplanting. Therefore, the data in Table 1 represent baseline seed quality and vigor for each genotype and are not specific to either the OF or GH system.
Significant differences (p ≤ 0.05) were observed in emergence time and percentage among genotypes (Table 1). Serrano Tampico recorded the shortest emergence time (15.25 ± 1.26 days), followed by Diente de Perro (16.25 ± 1.26 days), whereas Tía Juanita and Cola de Ratón showed the longest emergence times (24.75 ± 0.96 and 22.75 ± 2.22 days, respectively).
Emergence percentage was highest in Serrano Tampico (98.50 ± 1.08%), Tía Juanita (97.75 ± 1.55%), and Diente de Perro (93.50 ± 5.15%), whereas Mixteco Largo and Cola de Ratón showed significantly lower values (78.63 ± 6.69% and 75.13 ± 2.66%, respectively). Detailed ANOVA results are presented in Table S1 (Supplementary Material).
3.2.2. Growth Dynamics
Significant differences (p ≤ 0.05) in plant height (PH) and stem diameter (SD) were observed among genotypes within each production system (Figure 4). Detailed ANOVA results are provided in Tables S2 and S3 (Supplementary Material).
In OF, final plant height ranged from 65.45 to 105.44 cm. Tía Juanita showed the highest value (105.44 cm), followed by Cola de Ratón (99.31 cm) and Mixteco Largo (94.53 cm), whereas Serrano Tampico showed the lowest value (65.45 cm). Plant height stabilized from 63 dat onward. In GH, final plant height ranged from 108.33 to 155.04 cm. Tía Juanita showed the highest value (155.04 cm), followed by Mixteco Largo (132.00 cm) and Cola de Ratón (123.57 cm), whereas Serrano Tampico and Diente de Perro showed the lowest values (109.42 and 108.33 cm, respectively).
In OF, stem diameter ranged from 19.77 to 26.41 mm. Tía Juanita showed the highest value (26.41 mm), followed by Mixteco Largo (23.85 mm) and Cola de Ratón (20.89 mm), whereas Serrano Tampico showed the lowest value (19.77 mm). Stem diameter stabilized from 63 dat onward. In GH, stem diameter ranged from 10.90 to 12.86 mm, showing a consistent reduction relative to OF. Tía Juanita and Serrano Tampico showed the highest values (12.86 and 12.25 mm, respectively), whereas Mixteco Largo showed the lowest value (10.90 mm). Stabilization occurred from 119 dat onward.
3.2.3. Days to Flowering and Fruiting
Significant differences (p ≤ 0.05) in reproductive timing were observed among genotypes within each production system (Table 2). Detailed ANOVA results are presented in Tables S2 and S3 (Supplementary Material).
In OF, days to flowering ranged from 51 to 82 dat. Diente de Perro showed the lowest value (51 dat), followed by Serrano Tampico (57 dat), whereas Mixteco Largo (59 dat) and Cola de Ratón (62 dat) showed intermediate values. Tía Juanita showed the highest value (82 dat). In GH, days to flowering ranged from 45 to 77 dat. Diente de Perro (45 dat) and Serrano Tampico (50 dat) showed the lowest values, whereas Mixteco Largo (52 dat) and Cola de Ratón (57 dat) showed intermediate values. Tía Juanita showed the highest value (77 dat).
For fruiting, in OF values ranged from 67 to 92 dat, whereas in GH values ranged from 54 to 85 dat. Diente de Perro and Serrano Tampico showed the lowest values, while Tía Juanita showed the highest values in both systems.
3.3. Yield Components
Significant differences (p ≤ 0.05) were observed in number of fruits per plant and yield among genotypes within each production system (Table 3; Tables S2 and S3 in Supplementary Material).
In OF, fruit number ranged from 289.65 to 642.40 fruits per plant. Mixteco Largo (642.40) and Cola de Ratón (628.60) showed the highest values, whereas Diente de Perro showed the lowest (289.65). In GH, fruit number ranged from 130.00 to 288.33 fruits per plant. Tía Juanita (288.33) and Cola de Ratón (279.92) showed the highest values, whereas Diente de Perro showed the lowest (130.00).
Yield in OF ranged from 0.294 to 1.118 kg per plant. Serrano Tampico showed the highest value (1.118 kg), followed by Cola de Ratón (0.872 kg) and Mixteco Largo (0.759 kg), whereas Tía Juanita (0.294 kg) and Diente de Perro (0.387 kg) showed the lowest values. In GH, yield ranged from 0.181 to 0.603 kg per plant. Serrano Tampico showed the highest value (0.603 kg), followed by Cola de Ratón (0.332 kg), whereas Tía Juanita showed the lowest value (0.181 kg).
Descriptively, yield values in GH were consistently lower than those recorded in OF for all genotypes. Yield reductions relative to the open-field system ranged from 34.1% to 64.0% across genotypes. The largest reductions were observed in Mixteco Largo (64.0%) and Cola de Ratón (61.9%), followed by Serrano Tampico (46.1%) and Tía Juanita (38.4%), whereas Diente de Perro showed the smallest reduction (34.1%).
3.4. Canonical Correlation Analysis
Canonical correlation analysis (CCA) showed genotype-specific associations between environmental and agronomic variables within each production system (Table 4). Canonical correlation coefficients ranged from 0.6002 to 0.9607, indicating moderate to strong relationships.
Environmental variables, particularly temperature and radiation, were associated with growth and yield variables. The magnitude and direction of these associations varied among genotypes and production systems. In OF, associations tended to be positive between temperature and productive variables, whereas in GH more variable patterns were observed, including negative associations with yield. These patterns indicate the presence of genotype × environment interaction in phenological and productive responses. Mean values and direction of associations are presented in Tables S4 and S5 (Supplementary Material).
3.4.1. Interaction Patterns by Genotype
Diente de Perro showed canonical correlation coefficients of 0.8789 in OF and 0.9607 in GH. In OF, maximum (r = 0.52) and minimum temperature (r = 0.55) were associated with plant height (r = 0.76) and number of fruits (r = 0.61). In GH, minimum temperature (r = 0.59) and maximum PPFD (r = 0.46) were negatively associated with plant height (r = −0.49) and days to flowering (r = −0.67).
Tía Juanita showed coefficients of 0.9034 in OF and 0.8760 in GH. In OF, maximum PPFD (r = 0.41) was associated with number of fruits (r = 0.79) and yield (r = 0.51). In GH, maximum PPFD (r = 0.57) and maximum RH (r = 0.43) were negatively associated with yield (r = −0.29).
Mixteco Largo showed coefficients of 0.7095 in OF and 0.8421 in GH. In OF, maximum temperature (r = 0.55) was associated with yield (r = 0.36). In GH, minimum temperature (r = 0.78) was associated with plant height (r = 0.52), while negative associations were observed with yield (r = −0.53) and number of fruits (r = −0.47).
Serrano Tampico showed coefficients of 0.7699 in OF and 0.7582 in GH. In OF, maximum temperature (r = 0.64) and maximum PPFD (r = 0.50) were associated with days to flowering (r = 0.73). In GH, minimum temperature (r = 0.69) was associated with yield (r = 0.32) and number of fruits (r = 0.40), while plant height showed a negative association (r = −0.59).
Cola de Ratón showed coefficients of 0.6002 in OF and 0.8690 in GH. In OF, minimum temperature (r = 0.43) and maximum RH (r = 0.47) were negatively associated with yield (r = −0.46) and days to flowering (r = −0.42). In GH, maximum PPFD (r = 0.69) was associated with plant height (r = 0.34) and days to flowering (r = 0.69).
The structural correlation coefficients of the first three canonical factors for all the variables and genotypes are represented graphically in Figures S1 to S5 of the Supplementary Material, providing an integral visualization of these multivariate interrelations.
4. Discussion
The results obtained showed consistent patterns in the phenological and productive performance of C. annuum across the evaluated environments, with differentiated responses among genotypes depending on the growing conditions. The magnitude of these differences, including yield reductions of up to 64% and an approximate 53% decrease in photosynthetic photon flux density in the GH system, highlights the likely role of genotype × environment interaction (G×E) in the expression of productive potential. In this context, variations in temperature and radiation availability were associated with changes in phenology, plant architecture, and productive efficiency, suggesting that genotypes may differ in their ability to adjust the allocation of resources under contrasting environmental conditions.
It is important to note that the evaluated system corresponded to a low-technology tunnel-type GH with limited passive ventilation, a condition commonly found in production systems in developing regions. Therefore, the results should be interpreted within this specific context and not directly extrapolated to high-technology hydroponic systems with active climate control. Likewise, due to differences in planting density, substrate type, and water management between OF and GH, the observed differences between systems reflect the interaction of multiple factors, including both environmental and agronomic management conditions. Consequently, it is not possible to attribute the observed effects to a single variable in isolation. Under this context, although greenhouse microclimatic conditions were associated with a reduction in yield, this effect cannot be attributed exclusively to those factors, as other uncontrolled variables may also have influenced the observed response.
4.1. Microclimatic Conditions as a Limiting Factor in the Protected System
The microclimatic conditions recorded in the GH, characterized by maximum temperatures of up to 48.78 °C and an approximate 53% reduction in photosynthetically active radiation compared to OF, suggest a potentially restrictive environment for crop development. These conditions suggest that the observed microclimatic conditions may exceed commonly reported ranges favorable for C. annuum growth (18–30 °C) [25,37,38], suggesting a likely simultaneous occurrence of thermal stress and light limitation. The occurrence of these temperatures is consistent with the structural characteristics of the greenhouse, particularly its low height (3 m) and the absence of roof ventilation, conditions that may have limited heat dissipation and air renewal in the upper canopy.
From a physiological perspective, temperatures above 32 °C have been associated with alterations in physiological and reproductive processes in C. annuum, potentially affecting pollen viability and fruit development [39,40,41]. In this study, although direct physiological measurements were not performed, these mechanisms may partially explain the yield reduction observed in the GH. Additionally, the recorded radiation levels (247.3 μmol m−2 s−1) fall below those commonly associated with high photosynthetic activity in C. annuum, which, according to previous studies, may be linked to reduced net photosynthetic rates and consequently lower availability of photoassimilates for fruit filling [42,43]. This is consistent with studies reporting changes in photosynthesis and yield under reduced radiation conditions [44]. Moreover, the wide variability in relative humidity (8.93–94.78%), outside the optimal range (65–85%) [45], indicates an unstable environment, consistent with potential alterations in plant water balance and transpiration processes, with possible implications for physiological performance [46]. Collectively, these microclimatic conditions suggest potential physiological limitations that may affect key processes such as photosynthesis, photoassimilate partitioning, and reproductive development. In this regard, studies in C. annuum have documented reductions in growth and yield under abiotic stress conditions, particularly when water balance and resource availability are compromised [16].
In contrast, OF presented a less extreme thermal regime and greater radiation availability, conditions that favored a more balanced physiological development and higher productivity levels. In this context, the results contrast with the general assumption that protected systems automatically improve crop performance [47], showing that under low-technology conditions and limited environmental control, the greenhouse may represent a more restrictive environment for crop development. Such conditions have been widely documented in protected production systems [48,49], in some cases being more limiting than open-field conditions. These findings provide empirical evidence that, in low-technology systems, greenhouse environments do not necessarily represent an automatic improvement over open-field conditions, but rather their performance depends critically on the microclimatic conditions generated.
Since the production system was not evaluated as a replicated experimental factor, these differences should be interpreted in terms of the specific environments observed. From this perspective, although the system corresponded to a substrate-based hydroponic scheme, the results suggest that crop performance was mainly associated with the recorded microclimatic conditions rather than with the production technique itself. Additionally, although hydroponic systems typically offer advantages related to controlled water and nutrient supply, allowing greater precision in resource availability [50], these advantages may not have been fully expressed under conditions of thermal stress and light limitation.
4.2. Phenological and Growth Responses: Genotypic Plasticity and Adaptive Strategies
Differences observed in emergence percentage (75.13–98.50%) reflect variability in seed vigor and suggest the presence of intrinsic characteristics associated with the genotypes, particularly in native materials. The greater precocity observed in Serrano Tampico and Diente de Perro indicates faster establishment, whereas lower percentages in Mixteco Largo and Cola de Ratón suggest lower germination uniformity. In this context, the implementation of seed priming strategies could contribute to improving germination, seedling vigor, and establishment uniformity [51,52,53].
At the phenological level, clear differences among genotypes were observed within each production system. In OF, flowering and fruiting times were longer, suggesting a more gradual development compared to protected systems, where these phases tend to be accelerated [54]. This pattern is consistent with reports documenting variations in agronomic performance of C. annuum between open-field and protected conditions [11]. In contrast, the acceleration of the crop cycle observed in GH (4–13 days) is consistent with the higher temperatures recorded in this environment. However, this reduction in cycle duration was not associated with increased yield, indicating that greater precocity does not necessarily translate into higher productive efficiency.
Vegetative growth showed contrasting patterns among genotypes within each production system. In OF, plants exhibited lower height but greater stem diameter, suggesting a more balanced structural development. In GH, greater plant height combined with reduced stem diameter indicates a potential asymmetry in growth. This pattern is consistent with low radiation conditions, which have been associated with modifications in growth and yield in C. annuum [55], and may induce elongation processes as an adaptive response to maximize light interception [56].
Under these conditions, the combination of high temperature and low radiation may have promoted morphophysiological responses associated with a shade avoidance syndrome, where plants prioritize vertical growth over structural thickening [57,58]. These responses may compromise mechanical stability and the ability to support reproductive structures, with direct implications for yield.
Overall, the evaluated genotypes exhibited considerable diversity in their phenological and morphological behavior across both production systems, suggesting different adaptive strategies in response to environmental conditions. The precocity of Serrano Tampico and Diente de Perro may represent an adaptive trait favorable for short-cycle environments, whereas the longer cycle and greater vegetative development of Tía Juanita suggest a strategy oriented toward biomass accumulation, characteristic of genotypes that may perform better under less restrictive conditions.
4.3. Yield Response and Genotype × Environment Interaction
Differences observed in fruit number and yield reflect variations in the efficiency of biomass conversion into marketable yield. In OF, genotypes such as Mixteco Largo and Cola de Ratón produced the highest number of fruits, whereas Serrano Tampico achieved the highest yield (1.118 kg per plant), indicating that a greater number of fruits does not necessarily translate into higher productivity. This decoupling suggests differences in fruit size or individual fruit weight and in the efficiency of photoassimilate allocation, consistent with biomass partitioning patterns described in plants [59,60,61]. In this sense, Serrano Tampico may exhibit greater efficiency in the allocation of photoassimilates toward fruits, allowing it to reach higher yield levels despite not producing the highest number of fruits.
In GH, a generalized reduction in yield (34–64% depending on genotype) was observed, suggesting a consistent limitation in productive efficiency under this system. Although some genotypes such as Tía Juanita and Cola de Ratón maintained relatively high fruit numbers under GH conditions, their yield remained lower, suggesting possible constraints in resource supply for the full development of reproductive structures. This behavior is consistent with the previously described microclimatic conditions, particularly the combination of high temperatures and low radiation availability, factors that have been associated with reductions in fruit set due to alterations in pollen viability, as well as limitations in biomass accumulation during fruit filling under reduced radiation conditions in C. annuum [55,62].
Conversely, Serrano Tampico showed a smaller relative reduction in yield between systems, suggesting greater productive stability and a better capacity to adjust to contrasting environmental conditions. This may indicate higher physiological efficiency in resource use or lower sensitivity to stress conditions present in the protected system.
Overall, these results indicate that yield in C. annuum does not depend exclusively on the number of fruits produced, but rather on the interaction between reproductive structure formation and the availability of resources for their development, highlighting the importance of evaluating productive efficiency under different environmental conditions.
4.4. Environmental Drivers and Functional Relationships of Genotype × Environment Interaction (CCA)
Canonical correlation analysis (CCA) allowed the identification of differentiated associations between environmental and agronomic variables, showing that genotype responses varied depending on the evaluated environmental conditions. The high canonical correlation values observed in several genotypes suggest a strong association between environmental conditions and agronomic performance, supporting the importance of genotype × environment interaction (G×E) [63].
In OF, environmental variables, particularly temperature and radiation, showed predominantly positive associations with productive variables such as fruit number and yield in some genotypes, which is consistent with an environment that favored the expression of productive potential through greater energy availability for photosynthesis and biomass accumulation [64]. In contrast, in GH, more variable patterns were observed, including negative associations between environmental and agronomic variables, consistent with the presence of stress conditions or physiological imbalance.
At the genotypic level, differentiated responses were observed. Diente de Perro showed negative associations between environmental variables and development in GH, indicating greater sensitivity to conditions in this system. In contrast, Serrano Tampico exhibited positive associations with productive variables in both systems, indicating greater stability in its performance. In genotypes such as Mixteco Largo and Cola de Ratón, the observed associations suggest that, under greenhouse conditions, increases in certain environmental variables may not translate into improvements in productivity, indicating possible limitations in the ability of these genotypes to respond favorably to environmental changes.
Overall, the patterns identified through CCA suggest that the evaluated genotypes exhibit different strategies in response to environmental conditions, where some show greater productive stability and others greater sensitivity to specific conditions. These patterns reinforce the importance of considering G×E interaction in the selection of genetic materials and in the design of production systems adapted to specific environmental conditions.
4.5. Agronomic Implications and Perspectives
Varietal selection under contrasting systems: The results indicate that no single genotype was consistently superior across the evaluated environments, highlighting the importance of genotype × environment interaction in yield expression. Under open-field conditions, genotypes such as Mixteco Largo, Cola de Ratón, and Serrano Tampico showed favorable productive performance, suggesting their potential for this system. However, their behavior under protected environments with limited climate control may differ; therefore, their evaluation should consider the specific microclimatic conditions.
Greenhouse management in warm climates: The conditions recorded in the GH, with temperatures exceeding 48 °C and an approximate 53% reduction in photosynthetic photon flux density, suggest that passive systems may generate suboptimal environments. In this context, strategies such as improved ventilation, shading, or evaporative cooling could contribute to moderating the microclimate and improving crop performance [65]. Additionally, the management of light transmission emerges as a key factor for maintaining adequate PPFD levels [66].
Agronomic value of native germplasm: The performance observed in native landraces under open-field conditions suggests processes of local adaptation, highlighting their value as genetic resources in breeding programs aimed at abiotic stress tolerance and yield stability [67].
Future research perspectives: The results identify relevant research directions, including evaluation under systems with active climate control to separate environmental effects, analysis of fruit quality under genotype × environment interaction, and physiological studies focusing on photosynthesis and water-use efficiency. Likewise, strategies to improve germination and early establishment could optimize crop performance.
4.6. Study Limitations and Contextualization of Results
The present study has inherent limitations associated with its experimental design that should be considered when interpreting the results. In particular, the evaluation was conducted over a single growing season and at specific sites; therefore, the findings should be interpreted within the context of the analyzed environments rather than as universal generalizations.
Comparisons between production systems were influenced not only by microclimatic conditions but also by differences in agronomic management, including planting density, substrate type, and irrigation regime, which are integral components of each system. Therefore, the observed differences cannot be attributed exclusively to microclimatic conditions and should be interpreted as the combined effect of these interacting factors.
Additionally, the lack of replication of the production system factor across multiple environments or growing cycles limits the possibility of making statistically generalizable inferences regarding this factor. However, the adopted approach, based on the independent analysis of each system and the use of canonical correlation analysis (CCA), allows the identification of consistent associations between environmental and agronomic variables, providing relevant evidence of genotype × environment interaction.
It is important to emphasize that the evaluated system corresponded to a substrate-based hydroponic system under low-technology greenhouse conditions, with passive ventilation and without active climate control, which conditioned the recorded microclimatic conditions. Therefore, the results should be interpreted within this specific context and should not be directly extrapolated to hydroponic systems with active climate control, where the environment is more precisely regulated.
Overall, this study does not aim to establish absolute superiority between production systems, but rather to provide evidence on the differential performance of genotypes under contrasting environmental conditions. Within this framework, the results provide a solid basis for the formulation of new hypotheses and the design of future studies incorporating environmental replication, multiple evaluation cycles, and systems with greater microclimatic control.
5. Conclusions
The results of this study suggest that the phenological and productive performance of Capsicum annuum was associated with genotype × environment interaction, as reflected in differences in cycle duration, growth, and productive efficiency across growing environments.
In the greenhouse hydroponic system (GH), the recorded microclimatic conditions were associated with a reduction in yield, indicating that, under limited ventilation, the environment may restrict the expression of productive potential. This behavior is particularly relevant in production systems used by smallholder farmers and in developing regions, where environmental control is limited.
In contrast, under the evaluated conditions, the open-field system (OF) showed higher productive performance, highlighting the importance of matching the production system to local environmental conditions.
The patterns identified through canonical correlation analysis confirm a strong genotype × environment interaction, revealing differential responses among genotypes. In this context, Serrano Tampico showed greater productive stability, whereas native genotypes such as Mixteco Largo and Cola de Ratón exhibited better performance under open-field conditions, suggesting specific adaptation to this environment.
Overall, these results demonstrate that yield in Capsicum annuum depends critically on the interaction between genotype and environmental conditions, rather than solely on the production system employed, underscoring the need for management and selection strategies adapted to the specific microclimatic context.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, A.D.-A.; Investigation, B.N.H.-H.; methodology, A.D.-A., B.N.H.-H and M.A.T-C.; data curation and statistical analysis, B.N.H.-H. and B.E.H.-C.; resources, A.D.-A.; funding acquisition, A.D.-A. and B.N.H.-H.; writing—original draft preparation, B.N.H.-H.; supervision, M.A.T.-C., writing-review and editing, B.E.H.-C., M.A.T.-C., L.C.L.-E. and J.L.J.-V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was primarily supported by Colegio de Postgraduados, Campus Puebla, through the Student Activity Support Program (AAE, Apoyo a la Actividad de Estudiantes). Additional funding was provided by Colegio de Postgraduados under Grant CONV_RGAA_2024_68 (Research and Incidence Projects for the Conservation and Sustainable Use of Genetic Resources for Food and Agriculture).
Data Availability Statement
The original contributions presented in this study are included in the article/Supplementary Materials. Further inquiries can be directed to the corresponding author.
Acknowledgments
This study is product of the doctoral dissertation of the first, who wishes to thank the Ministry of Science, Humanities, Technology and Innovation (Secretaría de Ciencia, Humanidades, Tecnología e Innovación, SECIHTI), for scholarship number 784576 in support of her doctoral studies. The authors express their sincere gratitude to the local farmers for providing the native variety seeds used in this study. Special thanks are extended to Mr. Tomás Cuautle for providing the farmland and logistical support for the field experiment. The authors also acknowledge Colegio de Postgraduados, Campus Puebla, for providing greenhouse facilities and technical support.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Bijalwan, P.; Shukla, Y.R. Bell pepper (Capsicum annuum) productivity and economics as influenced by different weed management strategies. Indian J. Weed Sci. 2024, 56, 91–94. [Google Scholar] [CrossRef]
- Hossain, M.M.; Azad, M.A.K.; Soren, E.B.; Alam, M.N.; Ahmed, M.S.; Islam, M.S.; Monira, S. Enhancing growth, yield, and nutritional value of Capsicum annuum: Evaluating micronutrient efficiency and varietal performance. J. Agric. Food Res. 2025, 101945. [Google Scholar] [CrossRef]
- Organización de las Naciones Unidas para la Alimentación y la Agricultura (FAO). Estadísticas sobre alimentación y agricultura. FAOSTAT. 2023. Available online: https://www.fao.org/faostat/es/#data (accessed on 15 July 2023).
- Penella, C.; Calatayud, A. Pepper crop under climate change: Grafting as an environmental friendly strategy. In Pepper Crops: Production and Management; Calatayud, A., Barreto, M.G., Eds.; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef]
- Toxqui-Tapia, R.; Peñaloza-Ramírez, J.M.; Pacheco-Olvera, A.; Albarrán-Lara, A.L.; Oyama, K. Genetic Diversity and Landscape Genetic Structure of Capsicum annuum L. from Wild, Backyard, and Cultivated Populations in a Heterogeneous Environment in Oaxaca, Mexico. Polibotánica 2022, 53, 87–103. [Google Scholar] [CrossRef]
- Hudáková, T.; Šuleková, M.; Tauchen, J.; Šemeláková, M.; Várady, M.; Popelka, P. Bioactive compounds and antioxidant activities of selected types of chilli peppers. Czech. J. Food Sci. 2023, 41, 3. [Google Scholar] [CrossRef]
- Mandal, S.K.; Rath, S.K.; Logesh, R.; Mishra, S.K.; Devkota, H.P.; Das, N. Capsicum annuum L. and its bioactive constituents: A critical review of a traditional culinary spice in terms of its modern pharmacological potentials with toxicological issues. Phytother. Res. 2023, 37, 965–1002. [Google Scholar] [CrossRef]
- Akhter, M.J.; Akhter, S.; Islam, S.; Sarker, M.S.H.; Hasan, S.K. Varietal influence on bioactive compounds and antioxidant activity in chilies during development stages. Heliyon 2024, 10, e17441. [Google Scholar] [CrossRef]
- Baenas, N.; Belović, M.; Ilic, N.; Moreno, D.A.; García-Viguera, C. Industrial use of pepper (Capsicum annuum L.) derived products: Technological benefits and biological advantages. Food Chem. 2019, 274, 872–885. [Google Scholar] [CrossRef]
- Bulle, M.; Rahman, M.M.; Islam, M.R.; Abbagani, S. Strategies to develop climate-resilient chili peppers: transcription factor optimization through genome editing. Planta 2025, 262, 30. [Google Scholar] [CrossRef]
- Lozano-Fernández, J.; Orozco-Orozco, L.F.; Grisales-Vásquez, N.Y. Comportamiento agronómico de cultivares de pimentón (Capsicum annuum L.) cultivados en campo abierto y en condiciones protegidas. Terra Latinoam. 2022, 40. [Google Scholar] [CrossRef]
- Arya, M.; Kumar, G.; Giridhar, P. Serotonin-salt-stress model-induced cell growth via promoting an antioxidant system and secondary metabolites in Capsicum annuum cell suspension culture. ACS Omega 2024, 9, 37330–37342. [Google Scholar] [CrossRef]
- Thuy, T.L.; Kenji, M. Effect of high temperature on fruit productivity and seed-set of sweet pepper (Capsicum annuum L.) in the field condition. J. Agric. Sci. Technol. A B Hue Univ. J. Sci. 2015, 5, 515–520. [Google Scholar] [CrossRef]
- Park, B.; Kim, S.; Kim, S. Simulation of the impacts of high temperature stress on pepper (Capsicum annum L.) yields. Front. Plant Sci. 2025, 16, 1590193. [Google Scholar] [CrossRef]
- Rajametov, S.N.; Yang, E.Y.; Cho, M.C.; Chae, S.Y.; Pae, D.H.; Lee, M.H.; Baek, H.J.; Kang, W.H. Heat-tolerant hot pepper exhibits constant photosynthesis via increased transpiration rate, high proline content and fast recovery in heat stress condition. Sci. Rep. 2021, 11, 14328. [Google Scholar] [CrossRef]
- Ricardez-Miranda, L.E.; Lagunes-Espinoza, L.C.; Hernández-Nataren, E.; Palma-López, D.J.; Conde-Martínez, F.V. Water Restriction during the Vegetative and Reproductive Stages of Capsicum annuum var. glabriusculum and Its Effect on Growth, Secondary Metabolites and Fruit Yield. Sci. Hortic. 2021, 287, 110129. [Google Scholar] [CrossRef]
- Abdelhafez, A.A.; Al Dhumri, S.A.; Shaban, M.S.; Elgeheny, O.K.; Saleh, A.S.; Metwally, A.S. Exploring future cultivation strategies in greenhouses. In EntechOpen; 2025. [Google Scholar] [CrossRef]
- Velazquez-Gonzalez, R.S.; Garcia-Garcia, A.L.; Ventura-Zapata, E.; Barceinas-Sanchez, J.D.O.; Sosa-Savedra, J.C. A review on hydroponics and the technologies associated for medium- and small-scale operations. Agriculture 2022, 12, 646. [Google Scholar] [CrossRef]
- Rajaseger, G.; Chan, K.L.; Tan, K.Y.; Ramasamy, S.; Khin, M.C.; Amaladoss, A.; Kadamb Haribhai, P. Hydroponics: current trends in sustainable crop production. Bioinformation 2023, 19, 925–938. [Google Scholar] [CrossRef]
- Naresh, R.; Jadav, S.K.; Singh, M.; Patel, A.; Singh, B.; Beese, S.; Pandey, S.K. Role of hydroponics in improving water-use efficiency and food security. Int. J. Environ. Clim. Change 2024, 14, 608–633. [Google Scholar] [CrossRef]
- Sela Saldinger, S.; Rodov, V.; Kenigsbuch, D.; Bar-Tal, A. Hydroponic Agriculture and Microbial Safety of Vegetables: Promises, Challenges, and Solutions. Horticulturae 2023, 9, 51. [Google Scholar] [CrossRef]
- Sylvain, W.; Lalonde, T.; Monfet, D.; Haillot, D. Standardised Framework for Analysis of Greenhouse Performance Using Key Performance Indicators. Biosyst. Eng. 2019, 181, 59–72. [Google Scholar] [CrossRef]
- Shamshiri, R.R.; Jones, J.W.; Thorp, K.R.; Ahmad, D.; Man, H.C.; Taheri, S. Review of optimum temperature, humidity, and vapour pressure deficit for microclimate evaluation and control in greenhouse cultivation of tomato: a review. Int. Agrophys. 2018, 32, 287–302. [Google Scholar] [CrossRef]
- Gruda, N.; Tanny, J. Protected crops—recent advances, innovative technologies and future challenges. Horticulturae 2022, 8, 1087. [Google Scholar] [CrossRef]
- Oh, S.Y.; Koh, S.C. Fruit development and quality of hot pepper (Capsicum annuum L.) under various temperature regimes. Hortic. Sci. Technol. 2019, 37, 313–321. [Google Scholar] [CrossRef]
- Jang, Y.; Schafleitner, R.; Barchenger, D.W.; Lin, Y.P.; Lee, J. Evaluation of heat stress response in pepper (Capsicum annuum L.) seedlings under controlled environmental conditions using a high-throughput 3d multispectral phenotyping. Sci. Hortic. 2025, 345, 114136. [Google Scholar] [CrossRef]
- Parra-Londono, S.; López-Hernández, F.; Montoya, G.; Henao-Rojas, J.C.; Ossa-Ossa, G.A.; Cortés, A.J. Merging Phenotypic Stability Analysis and Genomic Prediction for Multi-Environment Breeding in Capsicum spp. Agronomy 2025, 15, 2690. [Google Scholar] [CrossRef]
- Bustos-Korts, D.; Romagosa, I.; Borràs-Gelonch, G.; Casas, A.M.; Slafer, G.A.; van Eeuwijk, F. Genotype by Environment Interaction and Adaptation. In Encyclopedia of Sustainability Science and Technology; Meyers, R., Ed.; Springer: New York, NY, USA, 2018. [Google Scholar] [CrossRef]
- Ntanasi, T.; Karavidas, I.; Savvas, D.; Spyrou, G.P.; Giannothanasis, E.; Cosentino, B.B.; Papasotiropoulos, V.; Sabatino, L.; Ntatsi, G. Physiological and Yield Responses of Pepper (Capsicum annuum L.) Genotypes to Drought Stress. Plants 2025, 14, 1934. [Google Scholar] [CrossRef]
- Kim, M.K.; Jeong, H.B.; Yu, N.; Park, B.M.; Chae, W.B.; Lee, O.J.; Lee, H.E.; Kim, S. Comparative Heat Stress Responses of Three Hot Pepper (Capsicum annuum L.) Genotypes Differing in Temperature Sensitivity. Sci. Rep. 2023, 13, 14203. [Google Scholar] [CrossRef] [PubMed]
- García-Vásquez, R.; Vera-Guzmán, A.M.; Carrillo-Rodríguez, J.C.; Pérez-Ochoa, M.L.; Aquino-Bolaños, E.N.; Alba-Jiménez, J.E.; Chávez-Servia, J.L. Bioactive and nutritional compounds in fruits of pepper (Capsicum annuum L.) landraces conserved among indigenous communities from Mexico. AIMS Agric. Food 2023, 8, 3. [Google Scholar] [CrossRef]
- Taitano, N.; Bernau, V.; Jardón-Barbolla, L.; Leckie, B.; Mazourek, M.; Mercer, K.; McHale, L.; Michel, A.; Baumler, D.; Kantar, M.; van der Knaap, E. Genome-wide genotyping of a novel Mexican Chile Pepper collection illuminates the history of landrace differentiation after Capsicum annuum L. domestication. Evol. Appl. 2018, 12, 78–92. [Google Scholar] [CrossRef]
- Castillo-Zavala, J.L.; Martínez, J.L.; Mostacero-León, J.; De la Cruz-Castillo, A.J.; Vásquez-Boyer, C.A.; Palacios-Díaz, M.; Escobar, J.; Villacorta-Vásquez, J.A.; López-Medina, S.E.; Gil-Rivero, A.E.; León-Torres, C.A.; Charcape-Ravelo, J.M. Estudio de la sensibilidad de Capsicum annuum al aumento de la salinidad debido al cambio climático. Bonplandia 2025, 34, 85–94. Available online: https://www.jstor.org/stable/48812407. [CrossRef]
- Sanjuán-Martínez, J.; Ortiz-Hernández, Y.D.; Aquino-Bolaños, T.; Cruz-Izquierdo, S.; Pérez-Pacheco, R. Response of Chile Huacle (Capsicum annuum L.) to Water Stress under Greenhouse Conditions. Agric. Soc. Y Desarro. 2023, 19, 421–435. [Google Scholar] [CrossRef]
- Khoury, C.K.; Brush, S.; Costich, D.E.; Curry, H.A.; de Haan, S.; Engels, J.M.M.; Guarino, L.; Hoban, S.; Mercer, K.L.; Miller, A.J.; Nabhan, G.P.; Perales, H.R.; Richards, C.; Riggins, C.; Thormann, I. Crop Genetic Erosion: Understanding and Responding to Loss of Crop Diversity. New Phytol. 2022, 233, 84–118. [Google Scholar] [CrossRef]
- Lazaridi, E.; Kapazoglou, A.; Gerakari, M.; Kleftogianni, K.; Passa, K.; Sarri, E.; Papasotiropoulos, V.; Tani, E.; Bebeli, P.J. Crop Landraces and Indigenous Varieties: A Valuable Source of Genes for Plant Breeding. Plants 2024, 13, 758. [Google Scholar] [CrossRef]
- Hawraa, I.; Khan, M.A.; Akram, M.T.; Rana, R.M.; Tipu, F.A.; Ali, I.; Nawaz, H.; Khan, M.H. Heat Stress in Chillies: Integrating Physiological Responses and Heterosis Breeding Approaches for Enhanced Resilience. Biol. Life Sci. Forum 2025, 51, 12. [Google Scholar] [CrossRef]
- Lopez, R.G.; Runkle, E.S.; Heins, R.D.; Blanchard, M.G. Quantifying temperature effects on plant development rates and quality of compact container-grown pepper. HortScience 2024, 59, 1293–1298. [Google Scholar] [CrossRef]
- Gisbert-Mullor, R.; Padilla, Y.G.; Calatayud, Á.; López-Galarza, S. Rootstock-mediated physiological and fruit set responses in pepper under heat stress. Sci. Hortic. 2022, 305, 111699. [Google Scholar] [CrossRef]
- Lin, S.; Lin, T.-H.; Yee, C.K.M.; Chen, J.; Wang, Y.-W.; Nalla, M.K.; Barchenger, D.W. Impedance Flow Cytometry for Selection of Pollen Traits Under High Temperature Stress in Pepper (Capsicum annuum L.). HortScience 2022, 57, 181–190. [Google Scholar] [CrossRef]
- Manoharan, S.; Srivastava, A.; Mangal, M.; et al. Temperature stress induced anatomical and histological alterations in reproductive organs of Capsicum annuum L. Protoplasma 2026. [Google Scholar] [CrossRef]
- Goo, H.; Roh, Y.; Lee, J.; Park, K.S. Analysis of bell pepper (Capsicum annuum L.) leaf spectral properties and photosynthesis according to growth period. Horticulturae 2024, 10, 646. [Google Scholar] [CrossRef]
- Wittmann, S.; Wittmann, I.; Mewis, I.; Förster, N.; Asseng, S.; Mempel, H. Capsicum annuum in vertical indoor farming: yield and capsaicinoid responses to reduced light and additional UV-A. Sci. Hortic. 2025, 350, 114364. [Google Scholar] [CrossRef]
- Kabir, M.Y.; Díaz-Pérez, J.C.; Nambeesan, S.U. Effect of shade levels on plant growth, physiology, and fruit yield in bell pepper (Capsicum annuum L.). Acta Hortic. 2020, 1268, 311–318. [Google Scholar] [CrossRef]
- López-Bravo, E.; Placeres-Remior, A.; Carbonell-Saavedra, E.; Martínez-Rodríguez, A.; González Cueto, O. Variability of Agroclimatic Factors and Irrigation Rate in Protected Cultivation of Pepper (Capsicum annuum). Rev. Cienc. Téc. Agropecu. 2023, 32, 4. [Google Scholar]
- Erwin, J.; Hussein, T.; Baumler, D.J. Pepper photosynthesis, stomatal conductance, transpiration, and water use efficiency differ with variety, indigenous habitat, and species of origin. HortScience 2019, 54, 1662–1666. [Google Scholar] [CrossRef]
- Nagarsheth, S.; Agbossou, K.; Henao, N.; Bendouma, M. The Advancements in Agricultural Greenhouse Technologies: An Energy Management Perspective. Sustainability 2025, 17, 3407. [Google Scholar] [CrossRef]
- Jung, W.; Kim, H. Evaluation of heat stress levels inside greenhouses during summer in Korea. Int. J. Environ. Res. Public Health 2022, 19, 12497. [Google Scholar] [CrossRef]
- Akpenpuun, T.D.; Ogunlowo, Q.; Na, W.-H.; Rabiu, A. Review of temperature management strategies and techniques in the greenhouse microenvironment. Adeleke Univ. J. Eng. Technol. 2023, 6, 126–147. [Google Scholar]
- Pomoni, D.I.; Koukou, M.K.; Vrachopoulos, M.G.; Vasiliadis, L. A Review of Hydroponics and Conventional Agriculture Based on Energy and Water Consumption, Environmental Impact, and Land Use. Energies 2023, 16, 1690. [Google Scholar] [CrossRef]
- Garruña-Hernández, R.; Latournerie-Moreno, L.; Ayala-Garay, O.; Santamaría, J.M.; Pinzón-López, L. Acondicionamiento pre-siembra: una opción para incrementar la germinación de semillas de chile habanero (Capsicum chinense Jacq.). Agrociencia 2014, 48, 413–423. [Google Scholar]
- Brondo-Ricárdez, R.; Domínguez-Angulo, S.; Pérez-Hernández, I.; D’Artola-Barceló, A.L. Tratamientos pregerminativos a semillas y desarrollo inicial de plántulas de chile amashito (Capsicum annuum L. var. glabriusculum). Agro Product. 2020, 13, 53–59. [Google Scholar] [CrossRef]
- Antúnez-Ocampo, O.M.; Croseños-Palazin, M.I.; Espinosa-Rodríguez, M.; Vázquez-Villamar, M.; Rojas-García, A.R.; Sabino-López, J.E. Germinación de la semilla y vigor de plántula de chiles apaxtlecos en respuesta a la radiación gamma. Bio Cienc. 2024, 11, e1591. [Google Scholar] [CrossRef]
- Vega-Alfaro, A.; Ramírez-Vargas, C.; Chávez, G.; Lacayo, F.; Bethke, P.C.; Nienhuis, J. Flowering time and productivity of interspecific grafts between pepper species in contrasting high tunnel-sheltered and open-field production environments in Costa Rica. HortTechnology 2021, 31, 828–837. [Google Scholar] [CrossRef]
- Maier, C.R.; Chavan, S.G.; Klause, N.; Liang, W.; Cazzonelli, C.I.; Ghannoum, O.; Chen, Z.-H.; Tissue, D.T. Light blocking film in a glasshouse impacts Capsicum annuum L. yield differentially across planting season. Front. Plant Sci. 2023, 14, 1277037. [Google Scholar] [CrossRef]
- Mao, L.; Dai, Y.; Huang, Y.; Sun, H.; Li, Z.; Yang, B.; Zhang, Z.; Chen, W.; Ou, L.; Yang, S. Effect of light intensity on gene expression in hypocotyl during elongation in a leaf-yellowing mutant of pepper (Capsicum annuum L.). Agronomy 2022, 12, 2762. [Google Scholar] [CrossRef]
- Bashyal, M.; Coolong, T.W.; Díaz-Pérez, J.C. Shade nets increase plant growth but not fruit yield in organic jalapeño pepper (Capsicum annuum L.). Agriculture 2025, 15, 1757. [Google Scholar] [CrossRef]
- Formisano, L.; Miras-Moreno, B.; Ciriello, M.; Zhang, L.; De Pascale, S.; Lucini, L.; Rouphael, Y. Between light and shading: Morphological, biochemical, and metabolomics insights into the influence of blue photoselective shading on vegetable seedlings. Front. Plant Sci. 2022, 13, 890830. [Google Scholar] [CrossRef]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Cockshull, K.E. The effect of flowering on the growth of the tomato. J. Exp. Bot. 1989, 40, 863–870. [Google Scholar]
- Rosado-Souza, L.; Yokoyama, R.; Sonnewald, U.; Fernie, A.R. Understanding source–sink interactions: Progress in model plants and translational research to crops. Mol. Plant 2023, 16, 96–121. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Jansma, S.Y.; Wolters-Arts, M.; de Groot, P.F.M.; Jansen, M.J.; Rieu, I. Long-term mild heat causes post-mitotic pollen abortion through a local effect on flowers. Front. Plant Sci. 2022, 13, 925754. [Google Scholar] [CrossRef]
- Sahmat, S.S.; Rafii, M.Y.; Oladosu, Y.; et al. Unravelling the dynamics of genotype and environment interactions on chilli (Capsicum annuum L.) yield-related attributes in soilless planting systems. Sci. Rep. 2024, 14, 1698. [Google Scholar] [CrossRef]
- Marcelis, L.F.M.; Heuvelink, E.; Goudriaan, J. Modelling biomass production and yield of horticultural crops: A review. Sci. Hortic. 1998, 74, 83–111. [Google Scholar] [CrossRef]
- Allali, F.E.; Fatnassi, H.; Demrati, H.; Errais, R.; Wifaya, A.; Aharoune, A. Greenhouse cooling systems: A systematic review of research trends, challenges, and recommendations for improving sustainability. Clean. Eng. Technol. 2025, 26, 100973. [Google Scholar] [CrossRef]
- Bantis, F.; Koukounaras, A. Impact of Light on Horticultural Crops. Agriculture 2023, 13, 828. [Google Scholar] [CrossRef]
- Ix-Nahuat, J.G.; Latournerie-Moreno, L.; Pech-May, A.M.; Pérez-Gutiérrez, A.; Tun-Suárez, J.M.; Ayora-Ricalde, G.; Mijangos-Cortes, J.O.; Castañón-Nájera, G.; López-Vázquez, J.S.; Montes-Hernández, S. Valor agronómico de germoplasma de chile dulce (Capsicum annuum L.) en Yucatán, México. Univ. Y Cienc. 2013, 29, 175–185. [Google Scholar]
Figure 1.
Production systems of C. annuum evaluated in San Agustín Calvario, Puebla: (A) open-field system and (B) substrate-based hydroponic system under low-technology greenhouse conditions.
Figure 1.
Production systems of C. annuum evaluated in San Agustín Calvario, Puebla: (A) open-field system and (B) substrate-based hydroponic system under low-technology greenhouse conditions.

Figure 2.
Plant morphology of five C. annuum genotypes: (A) Diente de Perro, (B) Tía Juanita, (C) Mixteco Largo, (D) Serrano Tampico, and (E) Cola de Ratón.
Figure 2.
Plant morphology of five C. annuum genotypes: (A) Diente de Perro, (B) Tía Juanita, (C) Mixteco Largo, (D) Serrano Tampico, and (E) Cola de Ratón.

Figure 3.
Agroclimatic dynamics in (A, C, E) open-field and (B, D, F) substrate-based hydroponic greenhouse systems: (A, B) air temperature, (C, D) relative humidity, and (E, F) photosynthetic photon flux density (PPFD). Each boxplot represents 24 daily observations. The first and third quartiles indicate the interquartile range, the horizontal line represents the mean, whiskers indicate minimum and maximum values, and points represent outliers.
Figure 3.
Agroclimatic dynamics in (A, C, E) open-field and (B, D, F) substrate-based hydroponic greenhouse systems: (A, B) air temperature, (C, D) relative humidity, and (E, F) photosynthetic photon flux density (PPFD). Each boxplot represents 24 daily observations. The first and third quartiles indicate the interquartile range, the horizontal line represents the mean, whiskers indicate minimum and maximum values, and points represent outliers.

Figure 4.
Growth dynamics of plant height (A, B) and stem diameter (C, D) of five C. annuum genotypes grown in open-field (A, C) and substrate-based hydroponic greenhouse systems (B, D). Data points represent mean values. Different letters at the final point indicate significant differences among genotypes within each production system according to Tukey’s test (p ≤ 0.05).
Figure 4.
Growth dynamics of plant height (A, B) and stem diameter (C, D) of five C. annuum genotypes grown in open-field (A, C) and substrate-based hydroponic greenhouse systems (B, D). Data points represent mean values. Different letters at the final point indicate significant differences among genotypes within each production system according to Tukey’s test (p ≤ 0.05).


Table 1.
Emergence time and percentage of five C. annuum genotypes evaluated during the pre-transplant establishment phase.
Table 1.
Emergence time and percentage of five C. annuum genotypes evaluated during the pre-transplant establishment phase.
| Genotype | Emergence time (days) |
Emergence (%) |
|---|---|---|
| Diente de Perro | 16.25 ab ± 1.26 | 93.50 a ± 5.15 |
| Tía Juanita | 24.75 c ± 0.96 | 97.75 a ± 1.55 |
| Mixteco Largo | 19.25 b ± 1.50 | 78.63 b ± 6.69 |
| Serrano Tampico | 15.25 a ± 1.26 | 98.50 a ± 1.08 |
| Cola de Ratón | 22.75 c ± 2.22 | 75.13 b ± 2.66 |
| Tukey’s HSD | 3.28 | 8.84 |
Values represent mean ± standard deviation (SD). Means within a column followed by the same letter are not significantly different according to Tukey’s HSD test (p ≤ 0.05). HSD: honestly significant difference. For emergence time, the letter “a” indicates earlier emergence.
Table 2.
Days to flowering and fruiting of five C. annuum genotypes evaluated under two production systems.
Table 2.
Days to flowering and fruiting of five C. annuum genotypes evaluated under two production systems.
| System | Genotype | Flowering (dat) | Fruiting (dat) |
|---|---|---|---|
| Open-field (OF) | Diente de Perro | 51 a ± 1.76 | 67 a ± 2.24 |
| Tía Juanita | 82 e ± 1.76 | 92 d ± 1.97 | |
| Mixteco Largo | 59 c ± 1.76 | 69 b ± 1.96 | |
| Serrano Tampico | 57 b ± 2.01 | 68 ab ± 2.16 | |
| Cola de Ratón | 62 d ± 2.01 | 74 c ± 3.50 | |
| Tukey’s HSD | 1.47 | 2.14 | |
|
Hydroponic Greenhouse (GH) |
Diente de Perro | 45 a ± 3.16 | 54 a ± 4.39 |
| Tía Juanita | 77 d ± 3.09 | 85 d ± 4.39 | |
| Mixteco Largo | 52 b ± 3.09 | 61 b ± 4.39 | |
| Serrano Tampico | 50 b ± 2.81 | 59 ab ± 4.39 | |
| Cola de Ratón | 57 c ± 1.88 | 68 c ± 4.39 | |
| Tukey’s HSD | 2.53 | 3.95 |
Values represent mean ± standard deviation (SD). Means within each production system followed by the same letter are not significantly different according to Tukey’s HSD test (p ≤ 0.05). HSD: honestly significant difference. For flowering and fruiting, the letter “a” indicates earlier phenology. In OF, n = 20 plants per genotype; in GH, n = 12 plants per genotype.
Table 3.
Number of fruits per plant and yield of five C. annuum genotypes grown in open-field and substrate-based hydroponic greenhouse systems.
Table 3.
Number of fruits per plant and yield of five C. annuum genotypes grown in open-field and substrate-based hydroponic greenhouse systems.
| System | Genotype | Number of fruits per plant | Total Yield (kg per plant) |
|---|---|---|---|
| Open-field (OF) | Diente de Perro | 289.65 c ± 50.70 | 0.387 c ± 0.09 |
| Tía Juanita | 402.00 b ± 70.61 | 0.294 c ± 0.05 | |
| Mixteco Largo | 642.40 a ± 152.07 | 0.759 b ± 0.14 | |
| Serrano Tampico | 371.25 bc ± 85.71 | 1.118 a ± 0.27 | |
| Cola de Ratón | 628.60 a ± 148.03 | 0.872 b ± 0.19 | |
| Tukey’s HSD | 93.11 | 0.141 | |
|
Hydroponic Greenhouse (GH) |
Diente de Perro | 130.00 c ± 10.79 | 0.255 c ± 0.02 |
| Tía Juanita | 288.33 a ± 39.75 | 0.181 d ± 0.03 | |
| Mixteco Largo | 240.25 b ± 21.68 | 0.273 c ± 0.03 | |
| Serrano Tampico | 152.42 c ± 25.23 | 0.603 a ± 0.09 | |
| Cola de Ratón | 279.92 a ± 46.86 | 0.332 b ± 0.05 | |
| Tukey’s HSD | 35.37 | 0.055 |
Values represent mean ± standard deviation (SD). Means within each production system followed by the same letter are not significantly different according to Tukey’s HSD test (p ≤ 0.05). HSD: honestly significant difference. Statistical analyses were conducted independently for each system. In OF, n = 20 plants per genotype; in GH, n = 12 plants per genotype.
Table 4.
Key environmental factors and agronomic variables associated in canonical correlation analysis for five C. annuum genotypes under two production systems.
Table 4.
Key environmental factors and agronomic variables associated in canonical correlation analysis for five C. annuum genotypes under two production systems.
| Genotype | System | Canonical correlation (r) |
Key environmental factors (correlation) |
Key agronomic variables (correlation) |
|---|---|---|---|---|
| Diente de Perro | OF | 0.8789 | TMax (+0.52), TMin (+0.55) | PH (+0.76), NF (+0.61), DF (+0.50), YP (+0.44) |
| GH | 0.9607 | TMin (+0.59), PPFDMax (+0.46) | PH (-0.49), DF (-0.67) | |
| Tía Juanita | OF | 0.9034 | PPFDMax (+0.41) | NF (+0.79), YP (+0.51) |
| GH | 0.8760 | RHMax (+0.43), PPFDMax (+0.57) | YP (-0.29) | |
| Mixteco Largo | OF | 0.7095 | TMax (+0.55) | YP (+0.36) |
| GH | 0.8421 | TMin (+0.78) | PH (+0.52), YP (-0.53), NF (-0.47) | |
| Serrano Tampico |
OF | 0.7699 | TMax (+0.64), PPFDMax (+0.50) | DF (+0.73) |
| GH | 0.7582 | TMin (+0.69) | PH (-0.59), YP (+0.32), NF (+0.40) | |
| Cola de Ratón | OF | 0.6002 | TMin (+0.43), RHMax (+0.47) | YP (-0.46), DF (-0.42) |
| GH | 0.8690 | PPFDMax (+0.69) | PH (+0.34), DF (+0.69) |
Only structural correlations ≥ 0.29 are presented to highlight the most relevant associations. Canonical correlation analysis was performed independently for each production system. (+) indicates positive correlation and (−) indicates negative correlation. OF: open field; GH: substrate-based hydroponic system under low-technology greenhouse conditions; TMax: maximum temperature; TMin: minimum temperature; RHMax: maximum relative humidity; PPFDMax: photosynthetic photon flux density; PH: plant height; DF: days to flowering; Y: yield; NF: number of fruits.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



