Submitted:
05 May 2026
Posted:
06 May 2026
You are already at the latest version
Abstract
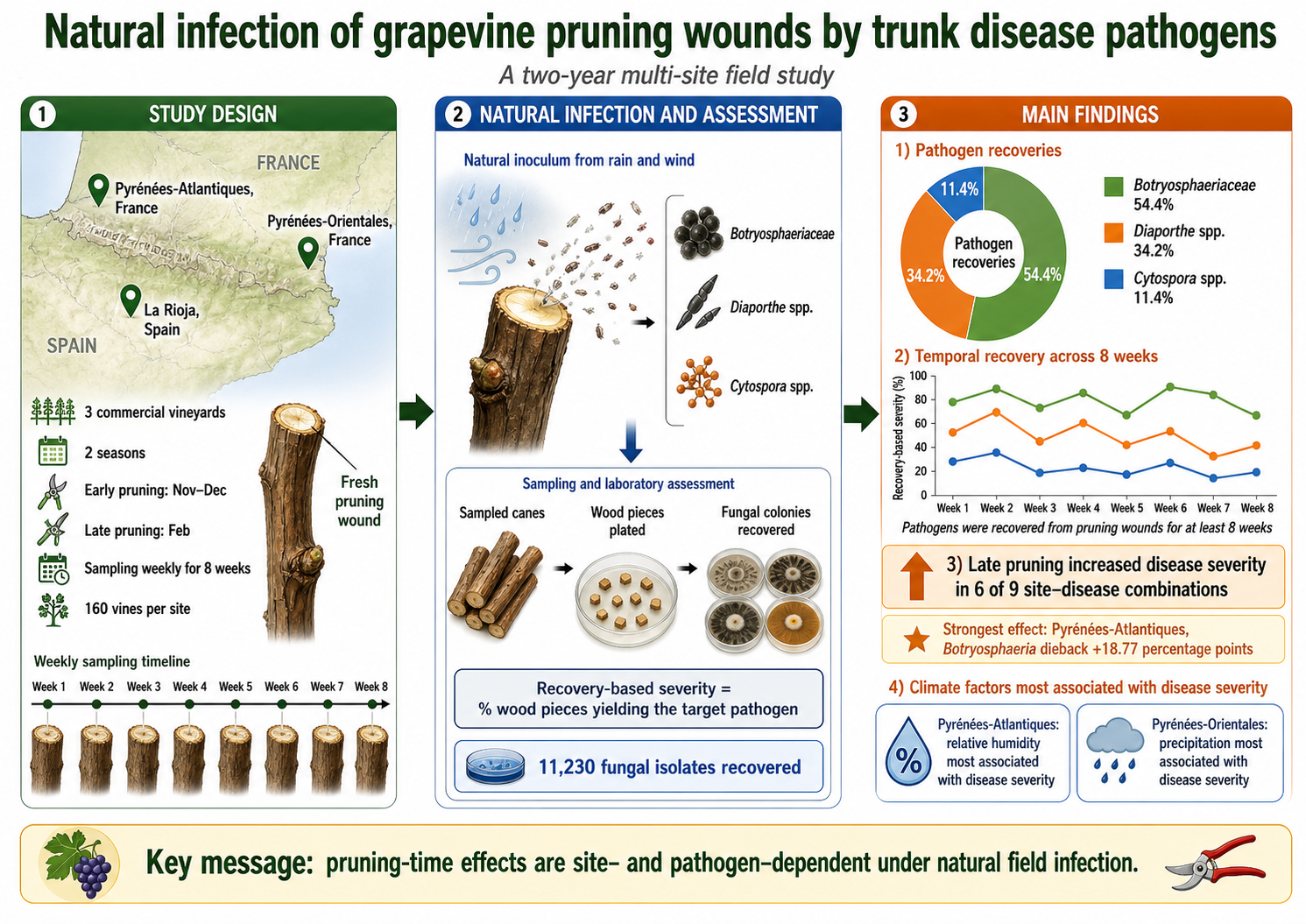
Grapevine trunk diseases (GTDs) are major constraints to vineyard longevity and productivity worldwide, and pruning wounds are recognized as key infection courts for their causal fungi. However, the dynamics of natural infection after pruning under field conditions remain insufficiently defined. This study evaluated natural infection of grapevine pruning wounds by GTD pathogens in three commercial vineyards in Spain and France over two growing seasons. At each site, vines were pruned either early in the dormant season (November-December) or late (February), and wounds were sampled weekly for 8 weeks. Disease severity was quantified as the percentage of wood pieces yielding GTD pathogens after isolation. A total of 11,230 fungal isolates were recovered, of which Botryosphaeriaceae accounted for 54.4%, followed by Diaporthe spp. (34.2%) and Cytospora spp. (11.4%). Disease severity varied significantly over time in all site-disease combinations, and temporal trajectories differed with pruning time and season. Late pruning resulted in significantly greater disease severity than early pruning in 6 of 9 site-disease combinations. The strongest effect was observed in Pyrénées-Atlantiques for Botryosphaeria dieback, where late pruning increased severity by 18.77 percentage points; Cytospora canker at the same site increased by 7.24 percentage points. Climatic analyses revealed site-specific associations, with relative humidity most strongly associated with disease severity in Pyrénées-Atlantiques and precipitation in Pyrénées-Orientales. These results indicate that GTD pathogens can be recovered from pruning wounds for at least 8 weeks after pruning and that the effect of pruning time depends strongly on vineyard and pathogen group.
Keywords:
grapevine trunk diseases
; pruning wounds
; natural infection
; Botryosphaeria dieback
; Phomopsis dieback
; Cytospora canker
; disease epidemiology
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



